
Submitted:
01 February 2024
Posted:
02 February 2024
You are already at the latest version
Abstract
Soil contamination with toxic heavy metals, such as lead (Pb), is a critical environmental issue with adverse effects on ecosystems. Cymbopogon citratus, known as lemongrass, is a medicinal plant with phytoremediation potential for Pb-contaminated soils, besides producing essential oil (EO) and bioactive compounds. Azospirillum brasilense is a plant growth promoting bacteria, and its inoculation into the soil can benefit plants in many ways, such as increased root growth and improved nutrient absorption. This study aimed to investigate the phytochemistry of lemongrass inoculated with A. brasilense and grown in Pb-contaminated soil to assess its responses to inoculation under different Pb levels. The experimental design was completely randomized in a 2 x 5 factorial scheme: two levels of A. brasilense (absence or presence) and five Pb levels (0, 50, 100, 300, and 500 mg Pb kg–1 of soil), totaling 60 experimental plots in ten treatments with six replicates. After four months of treatment, was analyzed total and reducing sugars, total phenolic content, flavonoids, antioxidant activity, antioxidant enzymes, proline, and EO content and composition. Soil Pb levels and A. brasilense inoculation affected phytochemical in lemongrass plants. Azospirillum inoculation reduced total sugars in the roots at all soil Pb levels, while increasing Pb levels favored a rise in sugar contents. There was an increase in flavonoid content in treatments associated with Pb and inoculated with A. brasilense. Antioxidant capacity was lower at lower Pb levels, regardless of bacterial inoculation. Enzymatic response was mainly affected by Pb concentrations between 50 and 100 mg kg–1 soil. EO content was influenced by soil Pb levels, with higher EO production at 500 mg Pb kg–1 soil and without A. brasilense inoculation. Overall, lemongrass cultivation in Pb-contaminated areas can be an alternative for phytoremediation and EO production for the industry. Moreover, A. brasilense inoculation proved to be beneficial in mitigating stress caused by high Pb concentrations in contaminated soils.
Keywords:
Antioxidant enzymes
; beneficial microorganisms
; phytoremediation
; lemongrass
; essential oil.
1. Introduction
Since the beginning of industrialization, a vast amount of organic pollutants (hydrocarbons, volatile organic compounds, and solvents) and inorganic pollutants (heavy metals) have been released into the environment [1]. Mining is the primary source of heavy metal contamination in soil, causing direct or indirect harm to plants and humans [2]. Among these pollutants, Lead (Pb), considered a heavy metal, poses a significant risk due to its high toxic and mutagenic potential [3,4].
In plants, Pb has negative effects on growth and development, impairing and altering production of active compounds [5]. High levels of Pb in plants can impair chloroplast function, inhibiting chlorophyll biosynthesis, CO2 fixation, and assembly of pigment-protein complexes in photosystems [6,7]. Pb-induced stress primarily damages the oxygen-evolving complex located in photosystem II [8].
Pb toxicity leads to root growth inhibition, stunted plant growth resulting in chlorosis, and disruption of various plant activities, including antioxidant systems, respiration, mineral nutrition, membrane structure and properties, and gene expression [9]. As Pb-contaminated areas often become unsuitable for cultivation of food crops, growing plants known as phytoremediators stands out as an environmentally sustainable approach to remove heavy metals from the soil, retaining them in the aboveground or root biomass [10]. This allows for reclamation of contaminated areas for cultivation of medicinal and aromatic plants, potentially generating a marketable end product, such as essential oil - EO [1,11].
Lemongrass, Cymbopogon citratus (D.C.) Stapf, is a plant widely used for phytoremediation due to its resistance to different heavy metals [12]. Its commercial interest in cultivation is primarily related to the cosmetics and perfumery industries, thanks to its EO [13], mainly composed of citral, which has two geometric isomers, geranial and neral, with a characteristic lemon scent [14]. Additionally, C. citratus contains minerals, vitamins, and bioactive compounds (alkaloids, terpenoids, flavonoids, phenols, saponins, and tannins), responsible for its pharmacological properties (antioxidant, antifungal, anticancer, antihypertensive, antidiabetic, and anxiolytic) [15,16]. Moreover, amid the global COVID-19 pandemic caused by the SARS-CoV-2 virus, the need for bioactive food ingredients has increased as they stimulate the immune system, and natural polyphenols are reported as potential inhibitors of the main protease of COVID-19 [17], with the use of C. citratus being studied in the prevention, treatment, and control of the virus [18].
Plant metabolite responses are influenced by various factors, including the availability of heavy metals in the soil [5]. An alternative to establishing stable conditions for better plant development is the use of plant growth promoting bacteria (PGPB), such as Azospirillum brasilense [19]. Bacteria of the Azospirillum genus can associate with the plant’s rhizosphere in external colonization or endophytically (Fukami et al., 2017, 2018). These bacteria promote plant growth through mechanisms such as amino acid biosynthesis and release, indole-acetic acid, cytokinins, gibberellins, and other polyamines, which enhance root growth and, consequently, improve water and nutrient absorption by plants [20,21]. Additionally, these microorganisms are capable of inducing the synthesis of antioxidant enzymes, reducing the deleterious effects of reactive oxygen species (ROS), and promoting greater root elongation, consequently improving the photosynthetic rate [22,23]. Plants under stress conditions have increased EO content since oil production is a plant defense mechanism; A. brasilense reduces oxidative stress, which may result in reduced EO content [24].
Plants have developed mechanisms to alleviate heavy metal toxicity and survive in polluted soils, with one mechanism being the elimination of ROS by increasing the activity of antioxidant enzymes [25]. According to Basu et al. [26], this defense system includes enzymatic antioxidants such as catalase (CAT), superoxide dismutase (SOD), and ascorbate peroxidase (APX), as well as non-enzymatic antioxidants. These enzymes are involved in detoxification of oxygen radicals and can be induced by stress caused by high Pb concentrations in contaminated soils [27,28].
Studies have suggested that induction of antioxidant responses is an adaptive mechanism in plants to counter the oxidative stress of Pb accumulation. Enzymes such as superoxide SOD and peroxidase (POD) demonstrate increased activity, confirming that the antioxidant system can play a crucial role in mitigating Pb toxicity [28,29,30]. The activity of enzymes like SOD, ascorbate peroxidases (APX), and glutathione peroxidase (GPX) can be increased with inoculation of symbiotic microorganisms, enhancing alleviation of Pb toxicity by eliminating reactive oxygen species (ROS) and reducing Pb concentrations in leaves [31,32].
In this context, this study aimed to investigate the growth and phytochemical responses of C. citratus inoculated with A. brasilense under different Pb levels, as well as to evaluate its responses to PGPB inoculation at different soil Pb levels.
2. Results and Discussion
2.1. Primary and Secondary Metabolites
Sugars are molecules that play a crucial role in essential cellular processes, such as seedling development, leaf formation, and growth of adventitious roots [33]. Even small changes in sugar content can affect metabolic status, gene expression, reserve accumulation and mobilization, as well as plant growth and development [33,34]. Moreover, they serve as signaling molecules indicating cellular energy availability in response to environmental fluctuations and cellular metabolism.
Depending on their levels in plants, sugars can also influence expression of various genes, primarily regulating the dynamics between source and sink tissues [34,35]. Sugar-regulated gene expression is not yet fully understood, but it is known that the ability of a specific gene to respond to sugar depends on the availability of particular signals and may also rely on plant developmental stage [36,37].
The shoot total sugar content in T6 (presence of A. brasilense and 0 Pb) was 788.39 mg g–1 of fresh mass, which was approximately 76.60% higher than in T1 (control). Therefore, the bacteria can increase production of total sugars in leaves (Table 2). Such an increase may have occurred because carbohydrate reserves are used to provide the energy required to maintain the association with diazotrophic bacteria [38,39].
As for the roots, total sugar contents were higher in T3 (no bacteria and 100 Pb) at 4714.73 mg g–1 fresh mass. This result was 96.7% higher than the average of treatments inoculated with A. brasilense. Hence, plants inoculated with A. brasilense had lower total sugar contents in the roots at all Pb levels in the soil. This outcome may be due to its effects in alleviating stress caused by heavy metals in plant metabolism. However, plants grown in Pb-contaminated soils without the presence of Azospirillum bacteria produced higher levels of total sugars in the roots, probably due to stress caused by Pb on their metabolism. Reducing sugars were minimally affected by Pb levels in the soil, with higher production of reducing sugars in the roots in T9 (bacteria present and 300 Pb), reaching 2764.81 µg g–1 fresh mass.
The higher contents of total sugars due to Pb may be related to their function in plants. High sugar concentrations are known to promote carbohydrate storage, while low levels stimulate photosynthesis, reserve mobilization, and export, directly influencing the source-sink relationships in plants [40,41]. Sugars can also act as osmoprotectant in regulating osmotic adjustment, providing membrane protection and eliminating toxic reactive oxygen species (ROS) under various stress conditions, including Pb stress [42,43].
Low concentrations of soluble sugars, such as glucose and sucrose, under stress conditions also stimulates the activity of antioxidant enzymes such as peroxidase (APX), catalase (CAT), and superoxide dismutase (SOD), which are essential enzymatic systems responsible for cellular homeostasis and detoxification [44].
Production of flavonoids in plant shoots (Table 3) was increased in T10 (674.06 mg g–1 of fresh mass), decreasing by approximately 80.32% in T6 and 77.43% in T1. The same trend was observed in the roots, where plants produced higher amounts of flavonoids in T10 (767.54 mg g–1 of fresh mass). Higher Pb concentrations in the soil increase production of flavonoids due to the need for protection against oxidizing agents [45,46].
Overall, all treatments associated with Pb and inoculated with A. brasilense showed higher productions of flavonoids; therefore, Azospirillum inoculation induced greater formation of flavonoids both in the roots and shoots of plants. Flavonoids play an important antioxidant role, especially in complementing the action of antioxidant enzymes when they are inactivated or have insufficient activity to counteract ROS during stress conditions [47,48,49]. Highly hydroxylated flavonoids are induced by abiotic stress, and the presence of an additional free hydroxyl (-OH) on the C-3’ of the B ring contributes to a stronger elimination capacity under these conditions [50,51].
Another class of secondary metabolites related to plant defense responses against environmental stressors is phenolic compounds, which, due to their stable intermediate radicals, prevent lipid oxidation [52]. In this sense, the treatment that presented the highest number of phenolic compounds in plant shoots was T5, as it was under greater stress. In contrast, in the roots, T10 exhibited the highest amount of phenolic compounds, as plants perceive the initial inoculation with A. brasilense as a stress factor, along with a high Pb dosage [53].
According to the antioxidant activity (DPPH) analysis, treatments T1, T2 (0 and 50 Pb with no bacteria inoculation), T6, and T7 (0 and 50 Pb with no bacteria present) did not significantly differ from each other, showing higher antioxidant capacity than the other treatments. Lower Pb levels provided higher antioxidant activity regardless of the presence of the PGPB. However, A. brasilense is known for inducing production of antioxidant enzymes and reducing deleterious effects from reactive oxygen species (ROS) [22,24].
2.2. Response of Antioxidant Enzymes to Lead Stress
The capacity of antioxidant systems to detoxify ROS is closely related to plant tolerance to heavy metals [54]. In recent years, many plant species have been identified as accumulators, with the ability to accumulate heavy metals without impacting their growth and development [32,55]. The enzyme SOD, which constitutes the first line of defense against ROS and is responsible for dismutation of superoxide radicals (O2-.) into hydrogen peroxide (H2O2), showed a significant result (p≤0.05) only in treatment T3 (Figure 1).
CAT, an enzyme that degrades H2O2 without consuming reducing equivalents, responsible for peroxide removal in excess due to its low affinity for H2O2, showed higher activity in T2, T3, T7, and T8, which are treatments with 50 and 100 mg Pb kg–1 soil with and without inoculation of A. brasilense.
The response of APX, a key enzyme in the glutathione-ascorbate cycle that reduces H2O2 to H2O using ascorbate as an electron donor, with concomitant generation of dehydroascorbate, was prominent in T2, with 6282.96 μmol of ascorbic acid g–1 FW min–1. The observed pattern of enzymatic response showed that Pb concentrations between 50 and 100 mg kg–1 induce the synthesis of antioxidant enzymes in response to Pb stress. Inoculation with A. brasilense did not significantly affect the synthesis of the studied enzymes, although Wang et al. [56] reported that bacteria can secrete antioxidant enzymes to prevent oxidative damage.
Regarding proline content (Figure 2), the highest accumulation occurred in T5 (899.18 µg g–1 fresh mass). In metal accumulator plants like lemongrass, an amino acid proline synthesis mechanism is activated as a defense mechanism, which not only mitigates oxidative stress but also restores the osmotic balance of plant cells [57]. In the case of treatments T7 and T9, inoculated with A. brasilense, proline formation was lower compared to treatments with Pb and without inoculation, indicating that A. brasilense may have beneficial effects in reducing Pb stress in lemongrass. The bacterium has been shown to assist in abiotic stress tolerance, alleviating stress caused by heavy metals [19,58]. Among the mechanisms activated for stress relief is induced systemic tolerance (IST), which is mediated by antioxidants, phytohormone production, osmotic adjustment, and defense strategies such as pathogenesis-related (PR) gene expression [22].
2.3. Extraction, Evaluation of EO Content and Chemical Identification by GC/MS
Content of lemongrass EO ranged from 0.17 to 0.50% (Figure 3). These findings are similar to those found in the literature: variations between 0.21 and 0.69% [5], 0.20 to 0.75% [24,59] and 0.20 to 0.76% [60]. The highest EO content was observed in T5 (~0.5%), which received the highest Pb concentration in the soil and had no A. brasilense inoculation. Overall, treatments with A. brasilense inoculation showed lower EO levels; therefore, Pb stress increased EO production in non-inoculated plants, as stressed plants tend to produce higher EO concentrations. In medicinal plants, EO content is known to vary due to numerous factors biotic and abiotic, including levels of chemical elements in the soil, so the stress caused by these elements can increase EO production in plants [61,62]. In the present study, A. brasilense inoculation may have alleviated plant stress caused by Pb, resulting in lower EO production in inoculated plants compared to non-inoculated ones.
The chemical characterization of EO by GC/MS (Table 4) revealed the presence of 21 components, predominantly oxygenated monoterpenes, which greatly contribute to its fragrance [14]. The compounds neral (22.79% to 66.79%) and geranial (16.95% to 51.01%) were the main components. Extraction from fresh leaves allows for identification of the neral and geranial isomers (citral Z and E, respectively), which have deeply embedded structures that are difficult to access in dried leaves ([24,59].
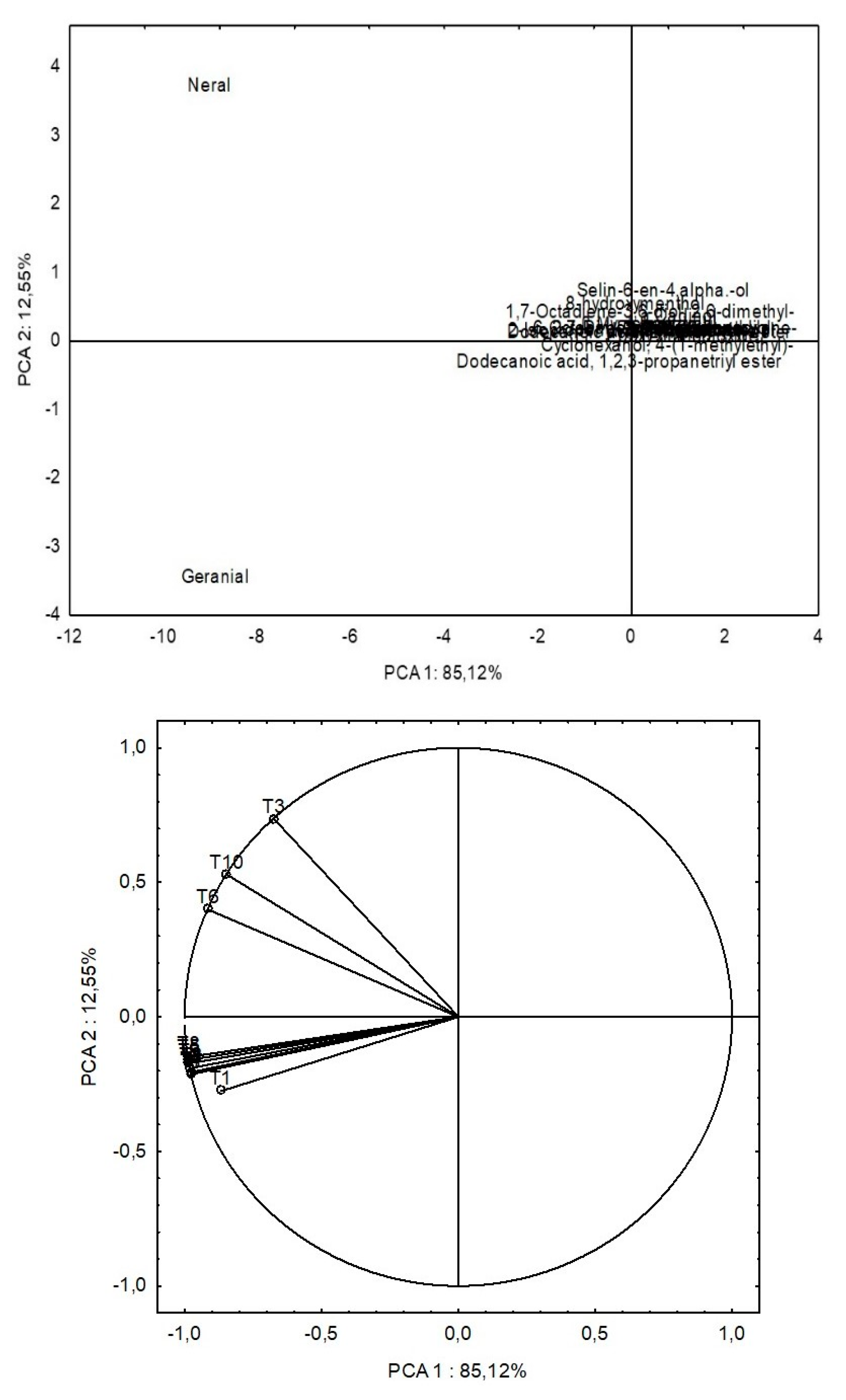
Principal component analysis (PCA) based on EO data from the ten different treatments showed that the components (PC1 and PC2) explained 85.12% and 12.55% of the variability among the chemical constituents of EO (Table 4). The PCA factorial loading graph (Figure 4) of EO constituents clearly separated treatments T3, T6, and T10, directly influencing the Neral component, from the other treatments (T1, T2, T4, T5, T7, T8, and T9), which were highly correlated with the production of Geranial. The concentration of Neral and Geranial behaved differently in relation to the Pb concentration in the soil [5], and the inoculation of A. brasilense also influencer this relationship between Neral and Geranial; this relationship is still not well understood between microorganisms and EO compound production production [24].
According to Bernstein et al. [63], hydrodistillation for EO extraction allows its extraction with nearly imperceptible levels of heavy metals, enabling its use in the food, pharmaceutical, and perfume industries, as citral is a raw material for production of ionone, vitamin A, and β-carotene. In addition to having a characteristic lemon scent, it can be combined in various applications [64].
According to our findings, lemongrass can be grown in Pb-contaminated environments, favoring soil phytoremediation, and inoculation with A. brasilense can help reduce chemical fertilizations, promoting the production system sustainability. This is expected to pave the way for further research to identify metabolic pathways and influence of microorganisms involved in EO production, as well as phytoremediation of contaminated soils. Our perspective is for lemongrass to be used as a biotechnological tool for eco-restoration of Pb-contaminated sites, bringing profitability to farmers.
2.4. Figures, Tables and Schemes

Table 1.
Chemical properties of the soil used in the experiment.
| pH (CaCl2) | P | C | Al3+ | H++Al3+ | Ca2+ | Mg2+ | K+ | SB | CTC | V | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| mg dm-3 | g dm-3 | -----------------------Cmolc dm-3------------------------- | % | ||||||||
| Soil | 6.07 | 1.54 | 4.09 | 0.00 | 1.89 | 1.38 | 0.75 | 0.05 | 2.18 | 4.07 | 53.52 |
| Ref1 | 3.8-6.6 | 16-24 | 0.8-15.9 | - | 0.6-5.0 | 0.3-7.2 | 0.3-3.3 | 0.1-0.7 | - | 2.2-12.5 | - |
EC=Cation exchange capacity; SB=Sum of bases; BS% = Base Saturation. *Methods: P. K extracted by Mehlich-I; Ca. Mg and Al – extracted by KCl 1 mol L−1; C – Dichromate / colorimetric. 1 Source: [65].
Table 2.
Total sugars in shoot (mg g−1 fresh mass) (TSS), total sugars in roots (mg g−1 fresh mass) (TSR), reducing sugars in shoot (µg g−1 fresh mass) (RSS) and reducing sugars in roots (µg g−1 fresh mass) (RSR) of lemongrass plants inoculated with Azospirillum® and grown in soils under different lead levels.
Table 2.
Total sugars in shoot (mg g−1 fresh mass) (TSS), total sugars in roots (mg g−1 fresh mass) (TSR), reducing sugars in shoot (µg g−1 fresh mass) (RSS) and reducing sugars in roots (µg g−1 fresh mass) (RSR) of lemongrass plants inoculated with Azospirillum® and grown in soils under different lead levels.
| Treatment | TSS | TSR | RSS | RSR |
|---|---|---|---|---|
| T1 | 184.56 ± 7.75d | 3849.19 ± 423.70b | 2337.80 ± 43.32cd | 2419.50 ± 45.82b |
| T2 | 108.66 ± 1.11e | 2676.01 ± 123.40c | 2780.62 ± 65.93a | 2178.07 ± 101.15c |
| T3 | 119.52 ± 3.24e | 4714.73 ± 329.25a | 2639.45 ± 69.21ab | 2183.84 ± 40.01c |
| T4 | 472.14 ± 11.63b | 1254.81 ± 35.89d | 2595.69 ± 106.51ab | 2368.12 ± 62.52bc |
| T5 | 117.85 ± 0.33e | 561.02 ± 68.17e | 2497.59 ± 20.95bc | 2389.64 ± 46.96bc |
| T6 | 788.39 ± 51.84a | 238.93 ± 1.99e | 2524.99 ± 121.51bc | 2317.78 ± 44.04bc |
| T7 | 125.56 ± 0.97e | 115.91 ± 1.11e | 2439.73 ± 86.55bc | 2415.42 ± 53.76b |
| T8 | 243.07 ± 1.59c | 200.51 ± 4.41e | 2216.46 ± 55.18d | 2361.74 ± 97.46bc |
| T9 | 106.13 ± 0.68e | 129.63 ± 0.82e | 2569.70 ± 70.05abc | 2764.81 ± 88.66a |
| T10 | 216.50 ± 3.25cd | 105.30 ± 0.60e | 2474.95 ± 33.65bc | 2492.77 ± 43.43b |
Mean values (n=6±standard error); different letters in the same column differ significantly by the Duncan test (p ≤ 0.05). T1: Control (autoclaved soil); T2: autoclaved soil + 50 mg Pb kg−1; T3: autoclaved soil + 100 mg Pb kg−1; T4: autoclaved soil + 300 mg Pb kg−1; T5: autoclaved soil + 500 mg Pb kg−1; T6: autoclaved soil + A. brasilense; T7: autoclaved soil + A. brasilense + 50 mg Pb kg−1 ; T8: autoclaved soil + A. brasilense + 100 mg Pb kg−1; T9: autoclaved soil + A. brasilense + 300 mg Pb kg−1; T10: autoclaved soil + A. brasilense + 500 mg Pb kg−1.
Table 3.
Total sugars in shoot (mg g−1 fresh mass) (TSS), total sugars in roots (mg g−1 fresh mass) (TSR), reducing sugars in shoot (µg g−1 fresh mass) (RSS) and reducing sugars in roots (µg g−1 fresh mass) (RSR) of lemongrass plants inoculated with Azospirillum® and grown in soils under different lead levels.
Table 3.
Total sugars in shoot (mg g−1 fresh mass) (TSS), total sugars in roots (mg g−1 fresh mass) (TSR), reducing sugars in shoot (µg g−1 fresh mass) (RSS) and reducing sugars in roots (µg g−1 fresh mass) (RSR) of lemongrass plants inoculated with Azospirillum® and grown in soils under different lead levels.
| Treatment | FlavS | FlavR | PhenS | PhenR | DPPH |
|---|---|---|---|---|---|
| T1 | 152.15 ± 5.81f | 32.34 ± 2.45f | 1207.59 ± 20.61e | 262.64 ± 4.97j | 36.91 ± 0.74a |
| T2 | 225.12 ± 1.16e | 40.12 ± 1.52ef | 1325.29 ± 34.80d | 309.28 ± 11.57i | 34.69 ± 0.07a |
| T3 | 224.81 ± 1.53e | 25.12 ± 4.38f | 1958.21 ± 7.87c | 348.88 ± 11.26h | 20.13 ± 0.28c |
| T4 | 236.78 ± 12.54e | 44.96 ± 2.51ef | 2479.73 ± 43.84b | 385.89 ± 3.56g | 12.70 ± 0.12d |
| T5 | 382.69 ± 9.29d | 68.30 ± 10.16e | 2771.76 ± 50.72a | 511.74 ± 1.23f | 12.64 ± 0.22d |
| T6 | 132.69 ± 1.34g | 434.81 ± 11.33d | 1284.57 ± 2.72de | 617.23 ± 2.39e | 35.50 ± 1.59a |
| T7 | 234.51 ± 2.71e | 468.45 ± 27.29c | 1330.47 ± 41.42d | 722.12 ± 22.95d | 34.53 ± 1.38a |
| T8 | 438.75 ± 2.29c | 584.72 ± 1.48b | 1351.20 ± 19.47d | 875.95 ± 13.61c | 15.07 ± 1.22d |
| T9 | 492.54 ± 2.55b | 604.36 ± 1.29b | 1893.07 ± 33.93c | 1081.00 ± 11.06b | 13.07 ± 0.10d |
| T10 | 674.06 ± 2.87a | 767.54 ± 1.24a | 1374.15 ± 28.82d | 2175.48 ± 2.96a | 28.21 ± 0.31b |
Mean values (n=6±standard error); different letters in the same column differ significantly by the Duncan test (p ≤ 0.05). T1: Control (autoclaved soil); T2: autoclaved soil + 50 mg Pb kg−1; T3: autoclaved soil + 100 mg Pb kg−1; T4: autoclaved soil + 300 mg Pb kg−1; T5: autoclaved soil + 500 mg Pb kg−1; T6: autoclaved soil + A. brasilense; T7: autoclaved soil + A. brasilense + 50 mg Pb kg−1 ; T8: autoclaved soil + A. brasilense + 100 mg Pb kg−1; T9: autoclaved soil + A. brasilense + 300 mg Pb kg−1; T10: autoclaved soil + A. brasilense + 500 mg Pb kg−1.
Table 4.
Chemical characterization (%) of the components of the essential oil of lemongrass plants inoculated with Azospirillum® and grown in soils under different lead levels.
Table 4.
Chemical characterization (%) of the components of the essential oil of lemongrass plants inoculated with Azospirillum® and grown in soils under different lead levels.
| Peak | 1RI | Component | T1 | T2 | T3 | T4 | T5 | T6 | T7 | T8 | T9 | T10 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 8.342 | β Myrcene | 2.46 | 3.79 | 8.04 | 4.73 | 6.63 | 3.66 | 7.31 | 8.05 | 4.87 | 1.79 |
| 2 | 14.568 | Neral (Citral Z) | 22.79 | 33.02 | 63.18 | 33.83 | 34.54 | 33.31 | 33.69 | 33.58 | 33.59 | 66.79 |
| 3 | 15.265 | Geranial (Citral E) | 35.7 | 48.02 | t | 50.44 | 47.03 | 49.31 | 50.79 | 44.74 | 51.01 | 16.95 |
| 4 | 15.550 | Epoxy-linalooloxide | 2.66 | t | t | t | t | t | t | t | t | t |
| 5 | 16.856 | Dodecanoic acid. 2-hexen-1-yl ester | 3.86 | t | 2.9 | t | t | t | 2.43 | t | t | t |
| 6 | 17.664 | Cyclohexanol. 4-(1-methylethyl)- | 5.97 | t | t | t | t | t | t | t | t | t |
| 7 | 42.681 | Dodecanoic acid. 1.2.3-propanetriyl ester | 19.87 | t | t | t | t | t | t | t | t | t |
| 8 | 8.280 | methyl heptenone | t | 0.34 | t | t | t | t | t | t | t | t |
| 9 | 10.908 | cis-Myroxide | t | 0.39 | 0.75 | 0.5 | 0.64 | 0.47 | 0.85 | 0.96 | 0.54 | t |
| 10 | 11.179 | Linalool | t | 0.53 | 0.98 | 0.51 | 0.6 | t | t | 0.56 | t | 0.51 |
| 11 | 12.202 | 6-Octenal. 7-methyl-3-methylene- | t | 0.39 | 0.75 | 0.4 | 0.42 | 0.39 | t | 0.43 | t | 0.38 |
| 12 | 12.689 | Isoneral | t | 0.7 | 2.34 | 0.76 | 0.82 | 0.7 | t | 0.8 | 0.62 | 0.58 |
| 13 | 13.125 | Isogeranial | t | 0.89 | 2.68 | 1 | 1.12 | 0.92 | 0.85 | 1.07 | 0.91 | 0.85 |
| 14 | 15.802 | 2-Undecanone | t | 0.3 | 0.52 | 0.3 | t | 0.38 | t | 0.36 | 0.28 | |
| 15 | 16.947 | 2.7-Dimethyl-2.7-octanediol | t | 2.41 | 2.34 | 2.3 | 2.96 | t | 3.19 | 2.5 | 2.16 | |
| 16 | 17.783 | 8-hydroxymenthol | t | 4.31 | 6.94 | 4.21 | 3.72 | 5.13 | 2.79 | 5.13 | 4.32 | 3.85 |
| 17 | 22.920 | Selin-6-en-4.alpha.-ol | t | 3.05 | 7.02 | 0.97 | 1.56 | 1.59 | t | 0.51 | 1.08 | 3.28 |
| 18 | 23.580 | α Cadinol | t | 1.16 | 3.64 | t | 0.6 | 0.59 | t | t | t | 1.31 |
| 19 | 15.602 | 1-Undecyne | t | t | t | t | t | 0.27 | t | 0.24 | t | t |
| 20 | 15.675 | 2-Isopropenyl-5-methylhex-4-enal | t | t | t | t | t | 0.31 | t | 0.35 | t | t |
| 21 | 20.070 | 2-Tridecanone | t | t | t | t | t | t | t | t | 0.43 | 0.56 |
| Total | 93.31 | 99.3 | 99.74 | 99.99 | 99.98 | 99.99 | 98.71 | 99.97 | 99.87 | 99.29 |
t: trace. 1Identification based on retention index (RI). T1: Control (autoclaved soil); T2: autoclaved soil + 50 mg Pb kg−1; T3: autoclaved soil + 100 mg Pb kg−1; T4: autoclaved soil + 300 mg Pb kg−1; T5: autoclaved soil + 500 mg Pb kg−1; T6: autoclaved soil + A. brasilense; T7: autoclaved soil + A. brasilense + 50 mg Pb kg−1 ; T8: autoclaved soil + A. brasilense + 100 mg Pb kg−1; T9: autoclaved soil + A. brasilense + 300 mg Pb kg−1; T10: autoclaved soil + A. brasilense + 500 mg Pb kg−1.
3. Materials and Methods
3.1. Experimental Design
The soil used in the experiment was characterized as Dark Red Latosol with medium texture and was collected from the experimental farm of the Universidade Paranaense (UNIPAR), Umuarama-PR, Brazil (Latitude: 23° 45’ 51’’ South, Longitude: 53° 19’ 6’’ West), at a depth of 0 to 20 cm. For chemical characterization, a soil sample was sent to the Laboratory of Soil Fertility located in Umuarama, PR (Table 1).
The experimental unit consisted of a polyethylene pot with a capacity of 3 liters of soil. The soil was sieved through a 4 mm mesh and sterilized in an autoclave for 1 h at 120 °C, twice, with a 24-h interval, and allowed to cool for three days before setting up the experiment. Young lemongrass seedlings, approximately 20 cm in height, were collected from the medicinal garden of the UNIPAR and washed with running water. Two disinfected seedlings were transplanted into each pot after being previously disinfected in 70% alcohol for one minute.
The experimental design used was completely randomized in a 2 x 5 factorial scheme: two levels of A. brasilense (absence or presence [1 mL plant–1]) strains Ab-V5 and Ab-V6 form a commercial and registered inoculate used in Brazil [23] and five levels of lead (Pb), totaling 60 experimental units in ten treatments with six replications conducted in a greenhouse. The treatments were as follows:
Treatment 1: soil autoclaved + 0 Pb (mg Pb kg–1 of soil)
Treatment 2: soil autoclaved + 50 Pb (mg Pb kg–1 of soil)
Treatment 3: soil autoclaved + 100 Pb (mg Pb kg–1 of soil)
Treatment 4: soil autoclaved + 300 Pb (mg Pb kg–1 of soil)
Treatment 5: soil autoclaved + 500 Pb (mg Pb kg–1 of soil)
Treatment 6: soil autoclaved + A. brasilense + 0 Pb (mg Pb kg–1 of soil)
Treatment 7: soil autoclaved + A. brasilense + 50 Pb (mg Pb kg–1 of soil)
Treatment 8: soil autoclaved + A. brasilense + 100 Pb (mg Pb kg–1 of soil)
Treatment 9: soil autoclaved + A. brasilense + 300 Pb (mg Pb kg–1 of soil)
Treatment 10: soil autoclaved + A. brasilense + 500 Pb (mg Pb kg–1 of soil)
All treatments were irrigated every two days for a period of four months with half-strength solution developed by Hoagland and Arnon [66].
3.2. Determination of Total and Reducing Sugars
Total sugars (glucose, fructose, mannose, and sucrose) were quantified using the phenol-sulfuric acid method by reading at 540 nm spectrophotometer [67]. Reducing sugars were quantified using the Dinitrosalicylic Acid (DNS) method adapted for microplates, and the samples were read at 490 nm [68]. A calibration curve was established using glucose as a standard. Both total and reducing sugar quantifications were performed with three biological replicates, in triplicate.
3.3. Determination of Total Phenolic Content, Flavonoids and DPPH Antioxidant Activity
Total phenolics were determined colorimetrically using the Folin-Ciocalteu reagent, as described by [69], with readings at 760 nm (R2: 0.9962). The total flavonoid content was also determined spectrophotometrically at 425 nm (R2: 0.9917), using a method described by [70] based on the formation of an aluminum-flavonoid complex. The antioxidant activity of fresh plant leaf extracts and standard antioxidants was evaluated based on the DPPH (2,2-diphenyl-1-picrylhydrazyl) free radical scavenging effect, measured spectrophotometrically at 515 nm [71]. All analyses were performed with three biological replicates, in triplicate.
3.4. Antioxidant Enzymes
Fresh plant tissues were macerated in liquid nitrogen and then approximately 0.3 g samples were homogenized in 1.5 mL of 200 mM potassium phosphate buffer (pH 7.8) containing 10 mM EDTA, 200 mM ascorbic acid, and 10% polyvinylpyrrolidone (PVPP) using a mortar and pestle. The homogenate was centrifuged at 16128 G-force for 20 min at 4 °C, and the supernatant was collected and stored in an ultra-freezer (-80 °C) until analysis. The extracts were used to test the antioxidant enzymes superoxide dismutase (SOD), catalase (CAT), and ascorbate peroxidase (APX). All assays were performed with three biological replicates, in triplicate [72].
3.4.1. Superoxide Dismutase (SOD, EC 1.15.1.1)
The SOD activity was determined by its ability to inhibit reduction of Nitroblue tetrazolium (NBT) by superoxide, forming blue formazan [73]. The reaction medium (1 mL) consisted of 50 μL of the crude sample extract, 50 mM KPO4 buffer (pH 7.8), 13 mM methionine, 0.1 μM EDTA, 75 μM NBT, and 2 μM riboflavin. The SOD activity was determined by spectrophotometry (560 nm) and expressed as U SOD g–1 FW min–1, where one unit of SOD activity (U) was defined as the amount of enzyme required to inhibit 50% of NBT reduction.
3.4.2. Catalase (CAT, EC 1.11.1.6)
The CAT activity was determined according to Havir and McHale [74]. The reaction medium (1 mL) consisted of 50 μL of the crude sample extract, 200 mM KPO4 buffer (pH 7.0), and 20 mM H2O2. The consumption of H2O2 was used to measure CAT activity by spectrophotometry (240 nm) for 1 min and then quantified using the molar extinction coefficient of 36 M–1 cm–1 [75]. The CAT activity was expressed as μmol H2O2 g–1 FW min–1.
3.4.3. Ascorbate Peroxidase (APX, EC 1.11.1.11)
The reduction of H2O2 to H2O oxidizing ascorbic acid is catalyzed by ascorbate peroxidase (APX). The method proposed by Nakano and Asada [76] was used to determine APX activity. The reaction medium (1 mL) consisted of 50 μL of the crude sample extract, 50 mM KPO4 buffer (pH 7.0), 10 mM ascorbic acid, and 1 mM H2O2. APX activity was determined by H2O2 degradation monitored through spectrophotometry (290 nm) for 1 min and quantified using the molar extinction coefficient of 2.8 mM–1 cm–1. APX activity was expressed as μmol ascorbic acid g–1 FW min–1.
3.5. Proline
Proline content was determined following the method proposed by Bates et al. [77]. Free proline contents in plant shoots were determined using fresh leaves (0.5 g) that were crushed with liquid nitrogen and mixed with 5 mL of a 3% sulfosalicylic acid solution. After centrifugation for 10 min at 16128 G-force, 2 mL of the resulting filtrate was combined with 2 mL of ninhydrin and 2 mL of glacial acetic acid in a test tube. The mixture was then heated in a water bath at 100 °C for 1 h and cooled to room temperature. Afterward, 4 mL of toluene was used to extract the mixture, and the absorbance was measured at 520 nm. This assay was conducted with five biological replicates, each performed in triplicate, and the proline content was calculated using a pre-established proline standard curve (R2: 0.9958).
3.6. Essential Oil Extraction and Yield Evaluation
Essential oil (EO) was extracted by hydrodistillation using a modified Clevenger apparatus for 3 h, according to Cruz et al. [24]. After extraction, it was transferred to amber bottles and allowed to evaporate the solvent to calculate the content (m/m %), considering the plant mass versus the EO mass. The EO was stored in a freezer (-20 °C) until the chemical characterization of the EO.
3.7. Chemical Identification of Essential Oil by GC/MS
EO was chemically identified using gas chromatography GC-MS QP 2010 SE (Shimadzu). Ten μL of the samples were diluted in 1000 μL of anhydrous dichloromethane before being injected into an SH-RTx-5MS column (Shimadzu, 5% phenylmethyl siloxane, 30 mx 0.25 mm id, 0.25 μm) using an autosampler (Shimadzu AOC-20i). Helium was used as carrier gas at a flow rate of 1.0 mL per min, with a split ratio of 2:1 and a sample injection amount of 1 μL. The column temperature was initially programmed at 40 °C, increasing at 8 °C per min to a final temperature of 300 °C. The injector and GC-MS interface temperatures were maintained at 250 °C. Mass spectra were recorded at 70 eV with a mass range of m/z 50 to 550 amu. The chemical compounds in the EO were identified based on library and GC–MS Postrun Analysis software.
3.8. Statistical Analysis
Data were subjected to analysis of variance (ANOVA), with means being compared by the Duncan’s test (p≤0.05) through the SPSS version 22.0 statistical program for Windows (SPSS Inc., Chicago, IL, USA). Principal Component Analysis (PCA) was performed to discriminate EO composition as a function of each treatment. All variables were analyzed using the Statistica v 13.0 software [78].
5. Conclusions
A. brasilense inoculation was effective in reducing high Pb concentration-related stress in the soil. Inoculated plants had lower sugar contents in the roots, even in the presence of Pb in the soil, and showed increased flavonoid formation. Enzymatic responses were mainly affected by Pb concentrations between 50 and 100 mg kg–1 in the soil.
EO content increased in plants grown in the soil with the highest Pb levels and without inoculation with A. brasilense. However, inoculated plants did not exhibit an increase in EO content, possibly due to the reduction of Pb-induced stress, as stressed plants typically produce higher EO concentrations. In conclusion, cultivating lemongrass in Pb-contaminated areas seems to be a promising alternative for EO production, as it did not significantly alter its composition.
6. Patents
Author Contributions
Rayane Monique Sete da Cruz: Methodology, Investigation, Data curation, Writing – original draft. Henrique Ferreira: Methodology, Investigation. Jonas Marcelo Jaski: Methodology, Data curation. Marcelo Coelho Esperança Vieira: Investigation, Data curation. Mariana Moraes Pinc: Investigation, Methodology, Data curation. Silvia Graciele Hülse de Souza: Methodology, Investigation, Data curation, Writing - review & editing. Odair Alberton: Supervision, Funding acquisition, Project administration, Writing – original draft, Writing - review & editing. All authors have read and agreed to the published version of the manuscript.
Data Availability Statement
Data will be made available on request.
Acknowledgments
The authors acknowledge Universidade Paranaense (UNIPAR) for the financial support. Rayane M. S. da Cruz thanks PROSUP/CAPES and the CNPq (National Council for Scientific and Technological Development) for the scholarship. Mariana M. Pinc thanks CAPES/PDPG Emergencial de Consolidação Estratégica dos Programas de Pós-Graduação for the scholarship. Odair Alberton acknowledge a research fellowship for the CNPq.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bharagava, R.N.; Saxena, G.; Mulla, S.I. Introduction to Industrial Wastes Containing Organic and Inorganic Pollutants and Bioremediation Approaches for Environmental Management. In Bioremediation of Industrial Waste for Environmental Safety; Springer Singapore: Singapore, 2020; pp. 1–18. [Google Scholar]
- Wei, B.; Yang, L. A Review of Heavy Metal Contaminations in Urban Soils, Urban Road Dusts and Agricultural Soils from China. Microchem. J. 2010, 94, 99–107. [Google Scholar] [CrossRef]
- Agwu, K.K.; Okoye, C.M.I.; Okeji, M.C.; Clifford, E.O. Potential Health Impacts of Heavy Metal Concentrations in Fresh and Marine Water Fishes Consumed in Southeast, Nigeria. Pakistan J. Nutr. 2018, 17, 647–653. [Google Scholar] [CrossRef]
- Manoj, M.C.; Kawsar, M. Metal Contamination Assessment in a Sediment Core from Vagamon Lake, Southwest India: Natural/Anthropogenic Impact. Environ. Nanotechnology, Monit. Manag. 2020, 14, 100362. [Google Scholar] [CrossRef]
- Lermen, C.; Morelli, F.; Gazim, Z.C.; Silva, A.P. da; Gonçalves, J.E.; Dragunski, D.C.; Alberton, O. Essential Oil Content and Chemical Composition of Cymbopogon Citratus Inoculated with Arbuscular Mycorrhizal Fungi under Different Levels of Lead. Ind. Crops Prod. 2015, 76, 734–738. [Google Scholar] [CrossRef]
- Sharma, P.; Dubey, R.S. Lead Toxicity Im Plants. Brazilian J. plant Physiol. 2005, 17, 35–52. [Google Scholar] [CrossRef]
- Kou, M.; Xiong, J.; Li, M.; Wang, M.; Tan, W. Interactive Effects of Cd and Pb on the Photosynthesis Efficiency and Antioxidant Defense System of Capsicum Annuum L. Bull. Environ. Contam. Toxicol. 2022. [Google Scholar] [CrossRef] [PubMed]
- Huihui, Z.; Xin, L.; Zisong, X.; Yue, W.; Zhiyuan, T.; Meijun, A.; Yuehui, Z.; Wenxu, Z.; Nan, X.; Guangyu, S. Toxic Effects of Heavy Metals Pb and Cd on Mulberry (Morus Alba L.) Seedling Leaves: Photosynthetic Function and Reactive Oxygen Species (ROS) Metabolism Responses. Ecotoxicol. Environ. Saf. 2020, 195, 110469. [Google Scholar] [CrossRef]
- Ali, M.; Nas, F.S. The Effect of Lead on Plants in Terms of Growing and Biochemical Parameters: A Review. MOJ Ecol. Environ. Sci. 2018, 3, 265–268. [Google Scholar] [CrossRef]
- Al-Khatib, I.A.; Anayah, F.M.; Al-Sari, M.I.; Al-Madbouh, S.; Salahat, J.I.; Jararaa, B.Y.A. Assessing Physiochemical Characteristics of Agricultural Waste and Ready Compost at Wadi Al-Far’a Watershed of Palestine. J. Environ. Public Health 2023, 2023, 1–13. [Google Scholar] [CrossRef]
- Zago, V.C.P.; das Dores, N.C.; Watts, B.A. Strategy for Phytomanagement in an Area Affected by Iron Ore Dam Rupture: A Study Case in Minas Gerais State, Brazil. Environ. Pollut. 2019, 249, 1029–1037. [Google Scholar] [CrossRef]
- Zakka Israila, Y. The Effect of Application of EDTA on the Phytoextraction of Heavy Metals by Vetivera Zizanioides, Cymbopogon Citratus and Helianthus Annuls. Int. J. Environ. Monit. Anal. 2015, 3, 38. [Google Scholar] [CrossRef]
- Oladeji, O.S.; Adelowo, F.E.; Ayodele, D.T.; Odelade, K.A. Phytochemistry and Pharmacological Activities of Cymbopogon Citratus: A Review. Sci. African 2019, 6, e00137. [Google Scholar] [CrossRef]
- Ajayi, E.O.; Sadimenko, A.P.; Afolayan, A.J. GC–MS Evaluation of Cymbopogon Citratus (DC) Stapf Oil Obtained Using Modified Hydrodistillation and Microwave Extraction Methods. Food Chem. 2016, 209, 262–266. [Google Scholar] [CrossRef]
- Balakrishnan, B.; Paramasivam, S.; Arulkumar, A. Evaluation of the Lemongrass Plant (Cymbopogon Citratus) Extracted in Different Solvents for Antioxidant and Antibacterial Activity against Human Pathogens. Asian Pacific J. Trop. Dis. 2014, 4, S134–S139. [Google Scholar] [CrossRef]
- Olorunnisola, S.K.; Asiyanbi, H.T.; Hammed, A.M.; Simsek, S. Biological Properties of Lemongrass: An Overview. Int. Food Res. J. 2014, 21, 455–462. [Google Scholar]
- Galanakis, C.M. The Food Systems in the Era of the Coronavirus (COVID-19) Pandemic Crisis. Foods 2020, 9, 523. [Google Scholar] [CrossRef] [PubMed]
- Kanyinda, J.-N.M. Preparation of Papers for European Journal of Medical and Health Sciences (EJMED). Eur. J. Med. Heal. Sci. 2020, 2, 3–6. [Google Scholar]
- Bashan, Y.; De-Bashan, L.E. How the Plant Growth-Promoting Bacterium Azospirillum Promotes Plant Growth—A Critical Assessment. In; 2010; pp. 77–136.
- Vejan, P.; Abdullah, R.; Khadiran, T.; Ismail, S.; Nasrulhaq Boyce, A. Role of Plant Growth Promoting Rhizobacteria in Agricultural Sustainability—A Review. Molecules 2016, 21, 573. [Google Scholar] [CrossRef]
- Zeffa, D.M.; Perini, L.J.; Silva, M.B.; de Sousa, N.V.; Scapim, C.A.; Oliveira, A.L.M. de; Amaral Júnior, A.T. do; Azeredo Gonçalves, L.S. Azospirillum Brasilense Promotes Increases in Growth and Nitrogen Use Efficiency of Maize Genotypes. PLoS One 2019, 14, e0215332. [Google Scholar] [CrossRef]
- Fukami, J.; Cerezini, P.; Hungria, M. Azospirillum: Benefits That Go Far beyond Biological Nitrogen Fixation. AMB Express 2018, 8, 73. [Google Scholar] [CrossRef]
- Fukami, J.; Ollero, F.J.; Megías, M.; Hungria, M. Phytohormones and Induction of Plant-Stress Tolerance and Defense Genes by Seed and Foliar Inoculation with Azospirillum Brasilense Cells and Metabolites Promote Maize Growth. AMB Express 2017, 7, 153. [Google Scholar] [CrossRef] [PubMed]
- da Cruz, R.M.S.; Alberton, O.; da Silva Lorencete, M.; da Cruz, G.L.S.; Gasparotto-Junior, A.; Cardozo-Filho, L.; de Souza, S.G.H. Phytochemistry of Cymbopogon Citratus (D.C.) Stapf Inoculated with Arbuscular Mycorrhizal Fungi and Plant Growth Promoting Bacteria. Ind. Crops Prod. 2020, 149, 112340. [Google Scholar] [CrossRef]
- Li, X.; Zhou, Y.; Yang, Y.; Yang, S.; Sun, X.; Yang, Y. Physiological and Proteomics Analyses Reveal the Mechanism of Eichhornia Crassipes Tolerance to High-Concentration Cadmium Stress Compared with Pistia Stratiotes. PLoS One 2015, 10, e0124304. [Google Scholar] [CrossRef] [PubMed]
- Basu, S.; Roychoudhury, A.; Saha, P.P.; Sengupta, D.N. Differential Antioxidative Responses of Indica Rice Cultivars to Drought Stress. Plant Growth Regul. 2010, 60, 51–59. [Google Scholar] [CrossRef]
- Chaoui, A.; Mazhoudi, S.; Ghorbal, M.H.; El Ferjani, E. Cadmium and Zinc Induction of Lipid Peroxidation and Effects on Antioxidant Enzyme Activities in Bean (Phaseolus Vulgaris L.). Plant Sci. 1997, 127, 139–147. [Google Scholar] [CrossRef]
- Yang, J.; Ye, Z. Antioxidant Enzymes and Proteins of Wetland Plants: Their Relation to Pb Tolerance and Accumulation. Environ. Sci. Pollut. Res. 2015, 22, 1931–1939. [Google Scholar] [CrossRef] [PubMed]
- Keser, G.; Saygideger, S. Effects of Lead on the Activities of Antioxidant Enzymes in Watercress, Nasturtium Officinale R. Br. Biol. Trace Elem. Res. 2010, 137, 235–243. [Google Scholar] [CrossRef]
- Guo, Z.; Gao, Y.; Cao, X.; Jiang, W.; Liu, X.; Liu, Q.; Chen, Z.; Zhou, W.; Cui, J.; Wang, Q. Phytoremediation of Cd and Pb Interactive Polluted Soils by Switchgrass (Panicum Virgatum L.). Int. J. Phytoremediation 2019, 21, 1486–1496. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Han, X.; Liang, Y.; Ghosh, A.; Chen, J.; Tang, M. The Combined Effects of Arbuscular Mycorrhizal Fungi (AMF) and Lead (Pb) Stress on Pb Accumulation, Plant Growth Parameters, Photosynthesis, and Antioxidant Enzymes in Robinia Pseudoacacia L. PLoS One 2015, 10, e0145726. [Google Scholar] [CrossRef]
- Etesami, H. Bacterial Mediated Alleviation of Heavy Metal Stress and Decreased Accumulation of Metals in Plant Tissues: Mechanisms and Future Prospects. Ecotoxicol. Environ. Saf. 2018, 147, 175–191. [Google Scholar] [CrossRef]
- Gibson, S.I. Control of Plant Development and Gene Expression by Sugar Signaling. Curr. Opin. Plant Biol. 2005, 8, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Lastdrager, J.; Hanson, J.; Smeekens, S. Sugar Signals and the Control of Plant Growth and Development. J. Exp. Bot. 2014, 65, 799–807. [Google Scholar] [CrossRef] [PubMed]
- Nelson, D.L.; Cox, M.M. Princípios de Bioquímica de Lehninger; Artmed, 2018. [Google Scholar]
- Kunz, S.; Pesquet, E.; Kleczkowski, L.A. Functional Dissection of Sugar Signals Affecting Gene Expression in Arabidopsis Thaliana. PLoS One 2014, 9, e100312. [Google Scholar] [CrossRef] [PubMed]
- Sakr, S.; Wang, M.; Dédaldéchamp, F.; Perez-Garcia, M.-D.; Ogé, L.; Hamama, L.; Atanassova, R. The Sugar-Signaling Hub: Overview of Regulators and Interaction with the Hormonal and Metabolic Network. Int. J. Mol. Sci. 2018, 19, 2506. [Google Scholar] [CrossRef] [PubMed]
- Schubert, K.R.; Evans, H.J. Hydrogen Evolution: A Major Factor Affecting the Efficiency of Nitrogen Fixation in Nodulated Symbionts. Proc. Natl. Acad. Sci. 1976, 73, 1207–1211. [Google Scholar] [CrossRef] [PubMed]
- Taiz, L.; Zeiger, E. Fisiologia e Desenvolvimento Vegetal; 6th ed.; 2017.
- Rolland, F.; Moore, B.; Sheen, J. Sugar Sensing and Signaling in Plants. Plant Cell 2002, 14, S185–S205. [Google Scholar] [CrossRef] [PubMed]
- Sami, F.; Yusuf, M.; Faizan, M.; Faraz, A.; Hayat, S. Role of Sugars under Abiotic Stress. Plant Physiol. Biochem. 2016, 109, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Srivastava, P.K.; Kumar, D.; Tripathi, D.K.; Chauhan, D.K.; Prasad, S.M. Morpho-Anatomical and Biochemical Adapting Strategies of Maize (Zea Mays L.) Seedlings against Lead and Chromium Stresses. Biocatal. Agric. Biotechnol. 2015, 4, 286–295. [Google Scholar] [CrossRef]
- Keunen, E.; Peshev, D.; Vangronsveld, J.; Van Den Ende, W.; Cuypers, A. Plant Sugars Are Crucial Players in the Oxidative Challenge during Abiotic Stress: Extending the Traditional Concept. Plant. Cell Environ. 2013, 36, 1242–1255. [Google Scholar] [CrossRef]
- Hu, M.; Shi, Z.; Zhang, Z.; Zhang, Y.; Li, H. Effects of Exogenous Glucose on Seed Germination and Antioxidant Capacity in Wheat Seedlings under Salt Stress. Plant Growth Regul. 2012, 68, 177–188. [Google Scholar] [CrossRef]
- Shimoi, K.; Masuda, S.; Shen, B.; Furugori, M.; Kinae, N. Radioprotective Effects of Antioxidative Plant Flavonoids in Mice. Mutat. Res. Mol. Mech. Mutagen. 1996, 350, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Franco; Navarro; Martínez-Pinilla Hormetic and Mitochondria-Related Mechanisms of Antioxidant Action of Phytochemicals. Antioxidants 2019, 8, 373. [CrossRef] [PubMed]
- Hatier, J.-H.B.; Gould, K.S. Foliar Anthocyanins as Modulators of Stress Signals. J. Theor. Biol. 2008, 253, 625–627. [Google Scholar] [CrossRef] [PubMed]
- Di Ferdinando, M.; Brunetti, C.; Fini, A.; Tattini, M. Flavonoids as Antioxidants in Plants Under Abiotic Stresses. In Abiotic Stress Responses in Plants; Springer New York: New York, NY, 2012; pp. 159–179. [Google Scholar]
- D’Amelia, V.; Aversano, R.; Chiaiese, P.; Carputo, D. The Antioxidant Properties of Plant Flavonoids: Their Exploitation by Molecular Plant Breeding. Phytochem. Rev. 2018, 17, 611–625. [Google Scholar] [CrossRef]
- Tattini, M.; Galardi, C.; Pinelli, P.; Massai, R.; Remorini, D.; Agati, G. Differential Accumulation of Flavonoids and Hydroxycinnamates in Leaves of Ligustrum Vulgare under Excess Light and Drought Stress. New Phytol. 2004, 163, 547–561. [Google Scholar] [CrossRef] [PubMed]
- Agati, G.; Azzarello, E.; Pollastri, S.; Tattini, M. Flavonoids as Antioxidants in Plants: Location and Functional Significance. Plant Sci. 2012, 196, 67–76. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a Free Radical Method to Evaluate Antioxidant Activity. LWT - Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Soobrattee, M.A.; Neergheen, V.S.; Luximon-Ramma, A.; Aruoma, O.I.; Bahorun, T. Phenolics as Potential Antioxidant Therapeutic Agents: Mechanism and Actions. Mutat. Res. Mol. Mech. Mutagen. 2005, 579, 200–213. [Google Scholar] [CrossRef]
- Schutzendubel, A.; Polle, A. Plant Responses to Abiotic Stresses: Heavy Metal-Induced Oxidative Stress and Protection by Mycorrhization. J. Exp. Bot. 2002, 53, 1351–1365. [Google Scholar] [CrossRef]
- Sharma, P. Efficiency of Bacteria and Bacterial Assisted Phytoremediation of Heavy Metals: An Update. Bioresour. Technol. 2021, 328, 124835. [Google Scholar] [CrossRef]
- Wang, Q.; Xiong, D.; Zhao, P.; Yu, X.; Tu, B.; Wang, G. Effect of Applying an Arsenic-Resistant and Plant Growth-Promoting Rhizobacterium to Enhance Soil Arsenic Phytoremediation by Populus Deltoides LH05-17. J. Appl. Microbiol. 2011, 111, 1065–1074. [Google Scholar] [CrossRef]
- Gupta, D.K.; Huang, H.G.; Corpas, F.J. Lead Tolerance in Plants: Strategies for Phytoremediation. Environ. Sci. Pollut. Res. 2013, 20, 2150–2161. [Google Scholar] [CrossRef]
- Bacilio, M.; Vazquez, P.; Bashan, Y. Alleviation of Noxious Effects of Cattle Ranch Composts on Wheat Seed Germination by Inoculation with Azospirillum Spp. Biol. Fertil. Soils 2003, 38, 261–266. [Google Scholar] [CrossRef]
- Lewinsohn, E. Histochemical Localization of Citral Accumulation in Lemongrass Leaves (Cymbopogon Citratus(DC.) Stapf., Poaceae). Ann. Bot. 1998, 81, 35–39. [Google Scholar] [CrossRef]
- de Souza, B.C.; da Cruz, R.M.S.; Lourenço, E.L.B.; Pinc, M.M.; Dalmagro, M.; da Silva, C.; Nunes, M.G.I.F.; de Souza, S.G.H.; Alberton, O. Inoculation of Lemongrass with Arbuscular Mycorrhizal Fungi and Rhizobacteria Alters Plant Growth and Essential Oil Production. Rhizosphere 2022, 22, 100514. [Google Scholar] [CrossRef]
- Ben Taarit, M.; Msaada, K.; Hosni, K.; Hammami, M.; Kchouk, M.E.; Marzouk, B. Plant Growth, Essential Oil Yield and Composition of Sage (Salvia Officinalis L.) Fruits Cultivated under Salt Stress Conditions. Ind. Crops Prod. 2009, 30, 333–337. [Google Scholar] [CrossRef]
- Abdelmajeed, N.A.; Danial, E.N.; Ayad, H.S. The Effect of Environmental Stress on Qualitative and Quantitative Essential Oil of Aromatic and Medicinal Plants. Arch. Des Sci. 2013, 66, 100. [Google Scholar]
- Bernstein, N.; Chaimovitch, D.; Dudai, N. Effect of Irrigation with Secondary Treated Effluent on Essential Oil, Antioxidant Activity, and Phenolic Compounds in Oregano and Rosemary. Agron. J. 2009, 101, 1–10. [Google Scholar] [CrossRef]
- Shahi, A.K.; Kaul, M.K.; Gupta, R.; Dutt, P.; Chandra, S.; Qazi, G.N. Determination of Essential Oil Quality Index by Using Energy Summation Indices in an Elite Strain of Cymbopogon Citratus (DC) Stapf [RRL(J)CCA12]. Flavour Fragr. J. 2005, 20, 118–121. [Google Scholar] [CrossRef]
- Sambatti, J.A.; Souza Junior, I.G.; Costa, A.C.S.; Tormena, C.A. Estimativa Da Acidez Potencial Pelo Método Do PH SMP Em Solos Da Formação Caiuá: Noroeste Do Estado Do Paraná. Rev. Bras. Ciência do Solo 2003, 27, 257–264. [Google Scholar] [CrossRef]
- Hoagland, D.R.; Arnon, D.I. The Water-Culture Method for Growing Plants without Soil. Circ. Calif. Agric. Exp. Stn. 1950, 347. [Google Scholar]
- Miller, G.L. Use of Dinitrosalicylic and for Determination of Reducing Sugar. Anal. Chem. 1959, 426–428. [Google Scholar] [CrossRef]
- Santos, A.A. dos; Deoti, J.R.; Müller, G.; Dário, M.G.; Stambuk, B.U.; Alves Junior, S.L. Dosagem de Açúcares Redutores Com o Reativo DNS Em Microplaca. Brazilian J. Food Technol. 2017, 20. [Google Scholar] [CrossRef]
- McDonald, S.; Prenzler, P.D.; Antolovich, M.; Robards, K. Phenolic Content and Antioxidant Activity of Olive Extracts. Food Chem. 2001, 73, 73–84. [Google Scholar] [CrossRef]
- Zhishen, J.; Mengcheng, T.; Jianming, W. The Determination of Flavonoid Contents in Mulberry and Their Scavenging Effects on Superoxide Radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Koleva, I.I.; van Beek, T.A.; Linssen, J.P.H.; Groot, A. de; Evstatieva, L.N. Screening of Plant Extracts for Antioxidant Activity: A Comparative Study on Three Testing Methods. Phytochem. Anal. 2002, 13, 8–17. [Google Scholar] [CrossRef]
- Bonacina, C.; da Cruz, R.M.S.; Nascimento, A.B.; Barbosa, L.N.; Gonçalves, J.E.; Gazim, Z.C.; Magalhães, H.M.; de Souza, S.G.H. Salinity Modulates Growth, Oxidative Metabolism, and Essential Oil Profile in Curcuma Longa L. (Zingiberaceae) Rhizomes. South African J. Bot. 2022, 146, 1–11. [Google Scholar] [CrossRef]
- Giannopolitis, C.N.; Ries, S.K. Superoxide Dismutases. Plant Physiol. 1977, 59, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Havir, E.A.; McHale, N.A. Biochemical and Developmental Characterization of Multiple Forms of Catalase in Tobacco Leaves. Plant Physiol. 1987, 84, 450–455. [Google Scholar] [CrossRef]
- Anderson, M.D.; Prasad, T.K.; Stewart, C.R. Changes in Isozyme Profiles of Catalase, Peroxidase, and Glutathione Reductase during Acclimation to Chilling in Mesocotyls of Maize Seedlings. Plant Physiol. 1995, 109, 1247–1257. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Hydrogen Peroxide Is Scavenged by Ascorbate-Specific Peroxidase in Spinach Chloroplasts. Plant Cell Physiol. 1981. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid Determination of Free Proline for Water-Stress Studies. 1973, 207, 205–207. [Google Scholar] [CrossRef]
- Statsoft Statsoft. Eletronic Statistics Textbook. Tulsa: StatSoft 2017.
Figure 1.
Antioxidant enzymes activity of superoxide dismutase (SOD (A), catalase (CAT) (B) and ascorbate peroxidase (APX) (C) of lemongrass plants inoculated with Azospirillum® and grown in soils under different lead levels. Mean values (n=6±standard error); different letters in the same column differ significantly by the Duncan test (p ≤ 0.05). T1: Control (autoclaved soil); T2: autoclaved soil + 50 mg Pb kg−1; T3: autoclaved soil + 100 mg Pb kg−1; T4: autoclaved soil + 300 mg Pb kg−1; T5: autoclaved soil + 500 mg Pb kg−1; T6: autoclaved soil + A. brasilense; T7: autoclaved soil + A. brasilense + 50 mg Pb kg−1 ; T8: autoclaved soil + A. brasilense + 100 mg Pb kg−1; T9: autoclaved soil + A. brasilense + 300 mg Pb kg−1; T10: autoclaved soil + A. brasilense + 500 mg Pb kg−1.
Figure 1.
Antioxidant enzymes activity of superoxide dismutase (SOD (A), catalase (CAT) (B) and ascorbate peroxidase (APX) (C) of lemongrass plants inoculated with Azospirillum® and grown in soils under different lead levels. Mean values (n=6±standard error); different letters in the same column differ significantly by the Duncan test (p ≤ 0.05). T1: Control (autoclaved soil); T2: autoclaved soil + 50 mg Pb kg−1; T3: autoclaved soil + 100 mg Pb kg−1; T4: autoclaved soil + 300 mg Pb kg−1; T5: autoclaved soil + 500 mg Pb kg−1; T6: autoclaved soil + A. brasilense; T7: autoclaved soil + A. brasilense + 50 mg Pb kg−1 ; T8: autoclaved soil + A. brasilense + 100 mg Pb kg−1; T9: autoclaved soil + A. brasilense + 300 mg Pb kg−1; T10: autoclaved soil + A. brasilense + 500 mg Pb kg−1.
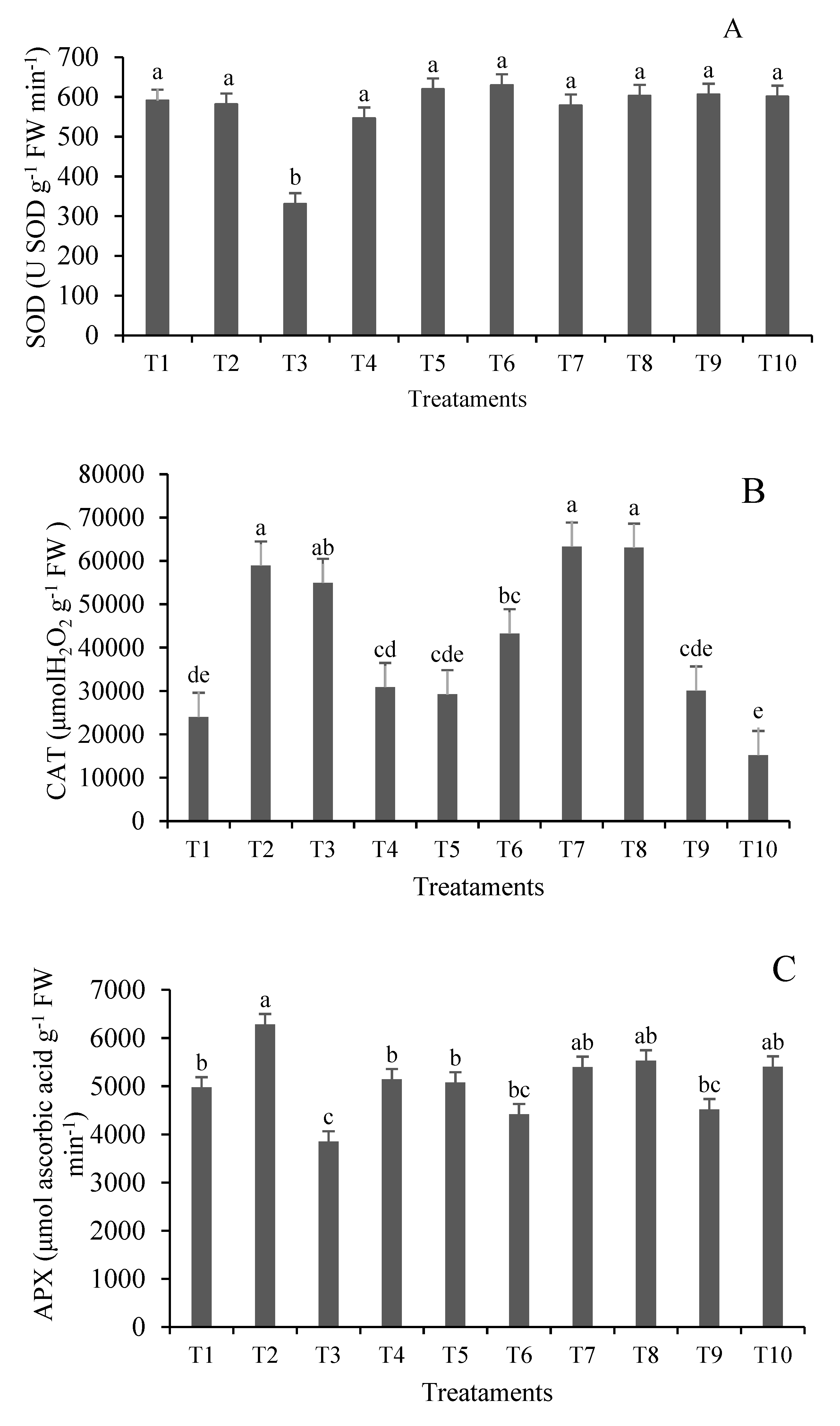
Figure 2.
Proline content (µg g−1 fresh leaves (FW)) in shoots of lemongrass plants inoculated with Azospirillum® and grown in soils under different lead levels. Mean values (n=6±standard error); different letters in the same column differ significantly by the Duncan test (p ≤ 0.05). T1: Control (autoclaved soil); T2: autoclaved soil + 50 mg Pb kg−1; T3: autoclaved soil + 100 mg Pb kg−1; T4: autoclaved soil + 300 mg Pb kg−1; T5: autoclaved soil + 500 mg Pb kg−1; T6: autoclaved soil + A. brasilense; T7: autoclaved soil + A. brasilense + 50 mg Pb kg−1 ; T8: autoclaved soil + A. brasilense + 100 mg Pb kg−1; T9: autoclaved soil + A. brasilense + 300 mg Pb kg−1; T10: autoclaved soil + A. brasilense + 500 mg Pb kg−1.
Figure 2.
Proline content (µg g−1 fresh leaves (FW)) in shoots of lemongrass plants inoculated with Azospirillum® and grown in soils under different lead levels. Mean values (n=6±standard error); different letters in the same column differ significantly by the Duncan test (p ≤ 0.05). T1: Control (autoclaved soil); T2: autoclaved soil + 50 mg Pb kg−1; T3: autoclaved soil + 100 mg Pb kg−1; T4: autoclaved soil + 300 mg Pb kg−1; T5: autoclaved soil + 500 mg Pb kg−1; T6: autoclaved soil + A. brasilense; T7: autoclaved soil + A. brasilense + 50 mg Pb kg−1 ; T8: autoclaved soil + A. brasilense + 100 mg Pb kg−1; T9: autoclaved soil + A. brasilense + 300 mg Pb kg−1; T10: autoclaved soil + A. brasilense + 500 mg Pb kg−1.
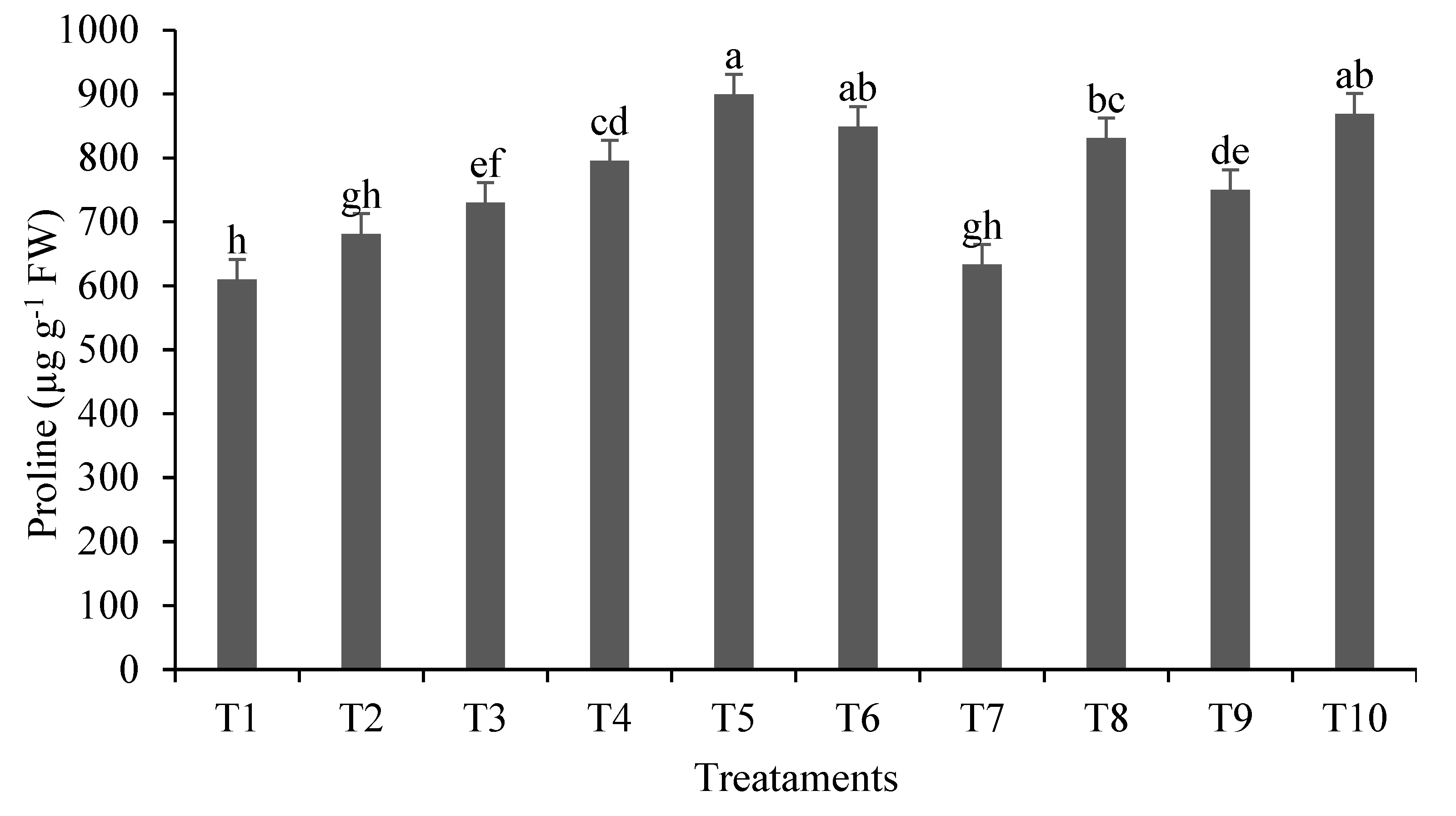
Figure 3.
Essential oil content (%) of lemongrass plants inoculated with Azospirillum® and grown in soils under different lead levels. Mean values (n=3±standard error); different letters in the same column differ significantly by the Duncan test (p ≤ 0.05). T1: Control (autoclaved soil); T2: autoclaved soil + 50 mg Pb kg−1; T3: autoclaved soil + 100 mg Pb kg−1; T4: autoclaved soil + 300 mg Pb kg−1; T5: autoclaved soil + 500 mg Pb kg−1; T6: autoclaved soil + A. brasilense; T7: autoclaved soil + A. brasilense + 50 mg Pb kg−1 ; T8: autoclaved soil + A. brasilense + 100 mg Pb kg−1; T9: autoclaved soil + A. brasilense + 300 mg Pb kg−1; T10: autoclaved soil + A. brasilense + 500 mg Pb kg−1.
Figure 3.
Essential oil content (%) of lemongrass plants inoculated with Azospirillum® and grown in soils under different lead levels. Mean values (n=3±standard error); different letters in the same column differ significantly by the Duncan test (p ≤ 0.05). T1: Control (autoclaved soil); T2: autoclaved soil + 50 mg Pb kg−1; T3: autoclaved soil + 100 mg Pb kg−1; T4: autoclaved soil + 300 mg Pb kg−1; T5: autoclaved soil + 500 mg Pb kg−1; T6: autoclaved soil + A. brasilense; T7: autoclaved soil + A. brasilense + 50 mg Pb kg−1 ; T8: autoclaved soil + A. brasilense + 100 mg Pb kg−1; T9: autoclaved soil + A. brasilense + 300 mg Pb kg−1; T10: autoclaved soil + A. brasilense + 500 mg Pb kg−1.
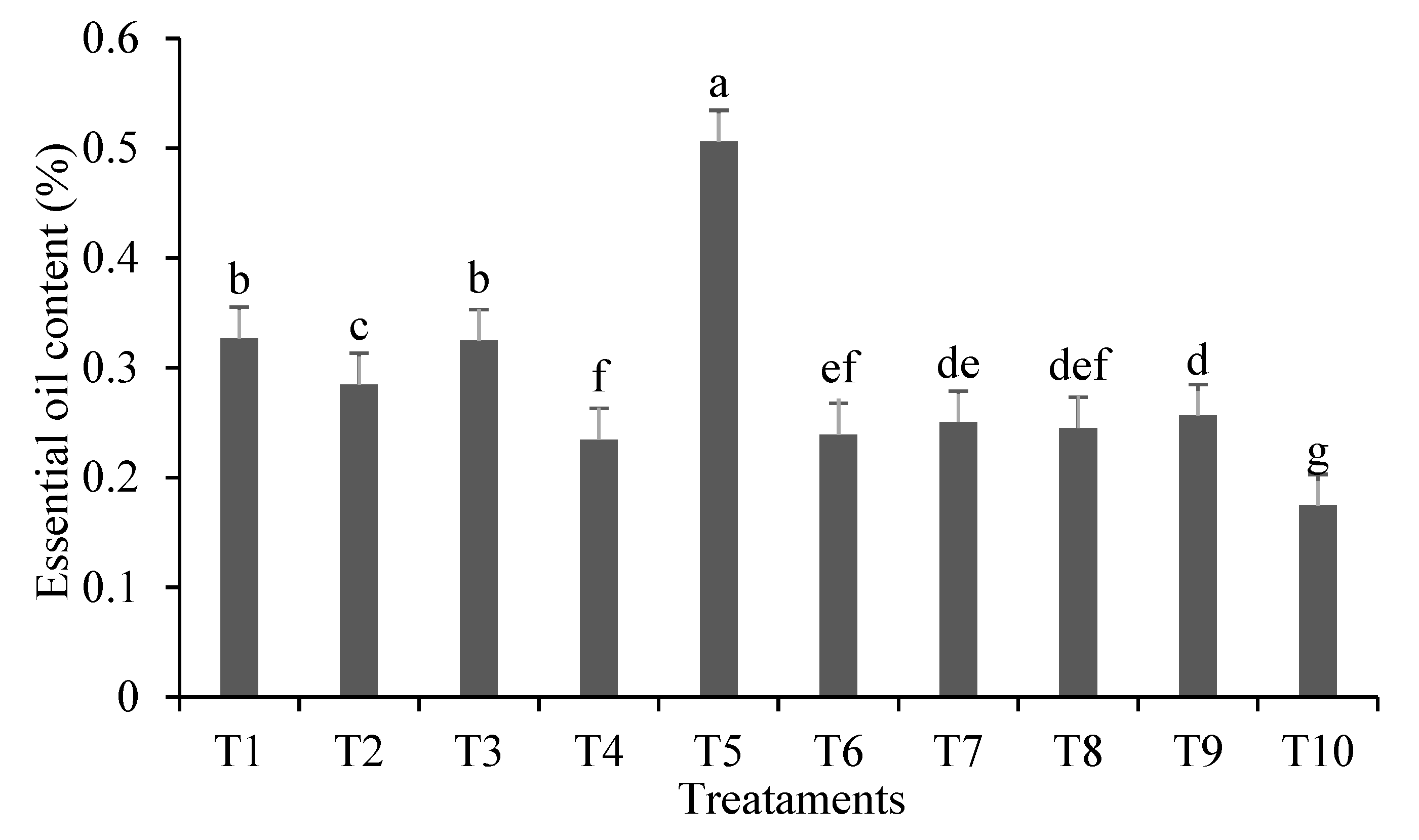
Figure 4.
Biplot representation of a PCA (Principal Component Analysis) performed on essential oils of lemon of lemongrass plants inoculated with Azospirillum® and grown in soils under different lead levels. T1: Control (autoclaved soil); T2: autoclaved soil + 50 mg Pb kg−1; T3: autoclaved soil + 100 mg Pb kg−1; T4: autoclaved soil + 300 mg Pb kg−1; T5: autoclaved soil + 500 mg Pb kg−1; T6: autoclaved soil + A. brasilense; T7: autoclaved soil + A. brasilense + 50 mg Pb kg−1 ; T8: autoclaved soil + A. brasilense + 100 mg Pb kg−1; T9: autoclaved soil + A. brasilense + 300 mg Pb kg−1; T10: autoclaved soil + A. brasilense + 500 mg Pb kg−1.
Figure 4.
Biplot representation of a PCA (Principal Component Analysis) performed on essential oils of lemon of lemongrass plants inoculated with Azospirillum® and grown in soils under different lead levels. T1: Control (autoclaved soil); T2: autoclaved soil + 50 mg Pb kg−1; T3: autoclaved soil + 100 mg Pb kg−1; T4: autoclaved soil + 300 mg Pb kg−1; T5: autoclaved soil + 500 mg Pb kg−1; T6: autoclaved soil + A. brasilense; T7: autoclaved soil + A. brasilense + 50 mg Pb kg−1 ; T8: autoclaved soil + A. brasilense + 100 mg Pb kg−1; T9: autoclaved soil + A. brasilense + 300 mg Pb kg−1; T10: autoclaved soil + A. brasilense + 500 mg Pb kg−1.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



