
Submitted:
01 February 2024
Posted:
02 February 2024
You are already at the latest version
Abstract
The microalgae Phaeodactylum tricornutum (PT) is distinguished by its rich nutrient profile, characterized by well-documented neuroprotective activities, including fucoxanthin (FX), a major carotenoid, and polyunsaturated omega-3 fatty acids (n-3 PUFA). The current study aims to evaluate the protective effects of a standardized extract of PT (Mi136) containing 2% FX on cognitive function, oxidative stress, and inflammation parameters in a mouse model of accelerated aging. Seventy-two (72) male mice were randomly assigned to the blank control group (BC), negative control group (NC), and four similar microalgae extract of PT groups (branded as BrainPhytTM) with different human equivalent doses. From day 01 to day 51, mice in the BC group were injected with a 0.9% normal saline solution, while mice in all other groups were subcutaneously injected with D-galactose (D-Gal) at a dose of 150 mg/kg once per day, five days per week. Results indicated that, for the three higher microalgae extract of PT dose groups, spatial cognitive function, swim latency, and step-through latency impairments induced by chronic D-Gal intoxication were significantly and fully inhibited, with mean values similar to those in the BC group during each day of testing. Similar benefits were observed in biochemical analysis, specifically regarding brain and plasma levels of lipid peroxidation, TNF-α, and IL-6 markers. These data underscore the positive effects of a standardized extract of PT containing 2% FX on cognitive function parameters such as spatial working memory, long-term memory, and short-term memory through the regulation of oxidative stress and inflammation pathways.
Keywords:
microalgae
; Phaeodactylum tricornutum
; fucoxanthin
; cognitive function
; ageing
; inflammation
; oxidative stress
; brain
; memory
; omega-3
1. Introduction
In developed countries, the phenomenon of accelerated population aging has been extensively documented over the past few decades, leading to significant shifts in public health policies [1]. A consequential aspect of this demographic transformation is the impending challenge of preserving cognitive function in the years to come, posing substantial implications for both societal and economic realms. Age-related cognitive decline is characterized as a non-pathological reduction in cognitive capabilities, encompassing aspects such as information processing speed, attention span, and notably, working (or short-term) memory [2]. These alterations are rooted in normal and intricate physiological changes intricately linked to the aging process [3]. The onset of this cognitive decline remains a subject of contention, with studies reporting initiation as early as the third [4] or fourth decade [5], while others pinpoint the commencement in the sixth decade [6]. Regardless of the timing, cognitive impairment associated with aging is acknowledged as a primary symptom of degenerative brain disorders like Alzheimer's Disease or other forms of dementia [7]. Some scholars posit that brain disorders may exert a more significant impact than cardiovascular diseases and cancers combined in the forthcoming years [8]. Consequently, preventing age-related cognitive decline is of paramount importance to mitigate the risk of developing debilitating brain diseases. Although the precise physiological mechanisms are not fully elucidated, inflammatory and oxidative stress pathways appear to play a pivotal role in the aging process [9,10]. As summarized in McGrattan et al.'s recent review (2019) [11], microglial macrophages in the brain become chronically active during aging, leading to sustained production of pro-inflammatory cytokines such as interleukin-6 (IL-6) and tumor necrosis factor-α (TNF-α). Elevated levels of these cytokines can trigger a cascade of neuroinflammatory processes, including neuronal death, reduced brain volume, or cortical thinning [12]. Additionally, there is well-documented evidence of increased oxidative stress in the aging brain [10,13]. The oxidative stress induced by an excessive surge in free radicals initiates chain oxidation reactions, causing cell damage and aging, consequently resulting in significant cognitive impairment, given the brain's heightened susceptibility to free radical attacks. Given these potential mechanistic insights, compounds exhibiting antioxidant and/or anti-inflammatory properties may prove promising in preventing cognitive decline during aging, complementing existing pharmacological and behavioral approaches [8,11,14].
Within this domain, marine environments harbor an extensive biological diversity of microalgae, representing a vast reservoir of largely unexplored bioactive molecules, including carotenoids, pigments, fatty acids, peptides, and sterols, renowned for their antioxidant and anti-inflammatory properties. The cultivation of microalgae in photobioreactors, employing eco-friendly production processes, offers a sustainable alternative to address societal challenges, particularly in the preservation of biodiversity and landscapes [15,16]. Notably, the microalgae Phaeodactylum tricornutum (PT) stands out for its rich nutrient profile, featuring well-documented neuroprotective activities, such as fucoxanthin (FX), a major carotenoid, polyunsaturated omega-3 fatty acids (n-3 PUFA), including eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA), and phycoprostans [17,18,19,20,21,22]. Primarily, the preventive role of fucoxanthin in cognitive impairment is suggested by its robust singlet oxygen-quenching properties, mitigating oxidative stress and lipid peroxidation. Furthermore, this key carotenoid exhibits the ability to traverse the blood-brain barrier, modulating various signaling pathways (e.g., Nrf-ARE and Nrf2-autophagy) implicated in brain oxidative stress and inflammation associated with aging [9,20,21]. Secondly, concerning n-3 PUFA, a higher plasma EPA level has been correlated with reduced gray matter atrophy in several brain regions [23]. Moreover, EPA serves as an agonist of peroxisome proliferator-activated receptor alpha (PPARα), thereby activating or inhibiting various signaling pathways (e.g., decreasing neuroinflammation and oxidative stress, promoting autophagy and lipid metabolism), directly contributing to neuroprotection. As for DHA, its established benefits include improving cerebral blood flow and reducing inflammation [24]. Finally, phycoprostans exhibit a specific impact on neuronal mitochondria by diminishing membrane potential, resulting in reduced reactive oxygen species (ROS) production [22]. Thus, the different bioactivities of these diverse molecules give to microalgae extract of PT a substantial potential to enhance cognitive functions in aging. However, to our knowledge, no studies have yet validated this hypothesis. The present study aims precisely to assess the protective effects of a standardized extract of PT containing 2% FX on cognitive function, oxidative stress, and inflammation parameters in a mouse model of accelerated aging.
2. Results
2.1. Effect on Y-Maze spontaneous alternation per
2.1.1. Y-Maze spontaneous alternation performance
The spatial cognitive function was evaluated using Y-maze test, and the results are shown in Figure 1. The spatial cognitive function of mice was impaired by D-GAL chronic intoxication, and the results showed that the mean spontaneous alternation behavior of the NC group (41,16%) decreased approximatively 26,39% compared to that of the BC group (67,56%). Microalgae extract of PT low dose group (BP120) significantly but partially improve (57,15%) spatial cognitive function impairment induced by D-Gal chronic intoxication (p < 0.001). For the three higher microalgae extract of PT doses groups, the spatial cognitive function impairment induced by D-Gal chronic intoxication were significantly and fully inhibited. In contrast, Y-maze test results showed a similar number of total arm entries and indicated no differences in overall behavioral locomotor activity among all groups. Detailed individual values were presented in the Supplementary Material (Tables S1 to S6).
2.1.2. Place learning in the Morris Water-Maze
To evaluate the long-term learning and memory function, a Morris water maze test was conducted and the results were shown in Figure 2a, b. During the 5 days of testing sessions, the mean swim latency in the BC group gradually and significantly decreased from 82s to 18s. The results indicated significant long-term learning and memory impairment induced by D-GAL chronic intoxication. As shown in Figure 2a, the mean swim latency time in NC group (From 90s at day 1 to 52s at day 5) is significantly lower than in BC group in particular for sessions during days 3 to 5. Microalgae extract of PT low dose group (BP120) significantly but partially improve (204,60s) swim latency time increased by D-Gal chronic intoxication at days 3 to 5. For the three higher microalgae extract of PT doses groups, the swim latency impairment induced by D-Gal chronic intoxication were significantly and fully inhibited with similar mean value than in BC group during each day tests. Regarding data from probe test, the time spent in target quadrant is significantly decreased in NC group while there are no difference in all Microalgae extract of PT groups compared to BC group as shown in Figure 2b.
2.1.3. Passive avoidance test
To evaluate the short-term learning and memory function, a passive avoidance test was conducted and the results were shown in Figure 3a, b. The results indicated significant learning and memory impairment induced by D-GAL chronic intoxication. The mean NC group (124,70s) showed a 51,42% decreased step-through latency time compared to that of the BC group (256,70s). Regarding escape latency time, the mean value in NC group (90,27s) is significantly increased compared to BC group (20,83s). Microalgae extract of PT low dose group (BP120) significantly but partially improve (204,60s) step-through latency time decreased by D-Gal chronic intoxication (P<0.01). For the three higher microalgae extract of PT doses groups, the step-through latency impairment induced by D-Gal chronic intoxication were significantly and fully inhibited with similar mean value than in BC group. Regarding escape latency time, the benefits induced by each microalgae extract of PT doses evaluated are similar than for step-through latency time as shown in Figure 3b.
2.2. Effect on brain lipid peroxidation level
The levels of brain (hyppocampus) lipid peroxidation in mice, expressed in percentage of BC level, are shown in Figure 4. Compared with the BC group, the levels of brain lipid peroxidation in mice injected with D-gal were significantly increased (p < 0.0001). Microalgae extract of PT low dose group (BP120) very significantly but partially decreased the brain lipid peroxidation induced by D-Gal chronic intoxication (P<0.0001). For the three higher microalgae extract of PT doses groups, the increase of brain lipid peroxidation levels produced by D-Gal chronic intoxication were significantly and fully inhibited.
2.3. Effect on brain and plasma inflammatory markers level
The levels of brain (hippocampus) and plasma inflammatory cytokines, expressed as percentage of BC levels group, in mice are shown in Figure 5. Compared with the BC group, the levels of brain TNF-α and IL-6 in mice injected with D-gal (NC group) were significantly increased (p < 0.0001). The two microalgae extract of PT lower doses groups (BP120 and BP235) very significantly but partially decrease the levels of brain TNF-α and IL-6 induced by D-Gal chronic intoxication while in microalgae extract of PT higher doses groups (BP295 and BP370) it is fully decreased. As shown in Figure 5c,d, the benefits of microalgae extract of PT on brain (hippocampus) and plasma IL-6 levels are similar than for TNF-α.
3. Discussion
Cognitive decline associated with aging is recognized as a significant public health concern, stemming from intricate physiological processes. Utilizing ingredients with antioxidant and/or anti-inflammatory functions emerges as a promising strategy for preventing cognitive decline in aging [8,11,14]. Microalgae have been identified as a rich source of bioactive molecules, encompassing carotenoids, pigments, fatty acids, peptides, and sterols, known for their neuroprotective effects through well-established signaling pathways. In this study, our aim was to investigate the effect of a standardized extract of Phaeodactylum tricornutum (PT) containing 2% fucoxanthin (FX) using a D-GAL mice model to assess its impact on cognitive function parameters through the regulation of oxidative stress and inflammation. Results revealed that the three higher doses of the microalgal extract of PT significantly and completely inhibited cognitive function impairments induced by chronic D-Gal intoxication. These inhibitory effects were comparable to the mean values observed in the control group (BC) during each daily test. Similar benefits were observed in biochemical analyses, demonstrating reductions in brain and plasma levels of lipid peroxidation, TNF-α, and IL-6 markers. These findings underscore the positive impact of the standardized extract of PT containing 2% FX on cognitive function parameters, including spatial working memory and short-term memory, achieved through the regulation of oxidative stress and inflammation pathways.
D-Galactose (D-Gal) mimics various behavioral and molecular features of brain aging in rodent models and is commonly employed to investigate the mechanisms of brain aging and anti-aging therapeutics in animal studies. As a physiological nutrient and reducing sugar, D-Gal reacts with the free amines of amino acids in proteins, leading to the formation of advanced glycation end products through nonenzymatic glycation. Excessive D-Galactose supply can contribute to the generation of reactive oxygen species (ROS) through oxidative metabolism and glycation end products. Rodents chronically injected with D-Gal for 6–10 weeks have been reported to exhibit a progressive decline in learning and memory abilities and increased free radical production in the brain [25,26]. Wu et al. (2008) demonstrated that chronic D-Gal supplementation induced a significant increase in latency to platform and a decrease in swim speed in mice models during the Morris Water Maze test. Moreover, Wei et al. (2017) reported that cognitive and behavioral parameters were impaired in a dose-dependent manner following chronic ingestion of D-Gal, justifying the dose of 150 mg/kg once per day, 5 days per week used in the present study. In our study, we observed significant cognitive impairments and elevated oxidative stress and inflammation markers in mice subjected to chronic D-Gal ingestion. The cognitive decline was particularly pronounced, with decreases ranging from 40% to 50% in various cognitive function tests compared to the placebo group. These alterations were paralleled by a notable increase of 200-300% in mean values of brain levels of lipid peroxidation, TNF-α, and IL-6 compared to the BC group. These results affirm the validity of the D-Gal mice model employed in our study and reinforce the positive benefits observed in all four experimental study groups. Indeed, the results showed that the three higher doses of the microalgae extract of PT fully inhibited impairments induced by chronic D-Gal intoxication in three major cognitive function tests. These results are comparable to the benefits induced by several interesting ingredients such as Astaxanthin, Spearmint, DHA, Ginkgo Biloba, or Fucoidan in different in-vivo cognitive-dysfunction models [27,28,29,30,31,32]. Ren et al. (2022) aimed to investigate the effects of different doses of a dietary supplement enriched with micronutrients, phosphatidylserine, and DHA on cognitive performance using a D-galactose (D-gal) induced aging rat model. Interestingly, the dietary supplement intervention significantly improved cognitive performance in the Morris water maze test with a similar dose-response kinetic than in the present study. Similar dose-response benefits were shown following 3 weeks of supplementation with Fucoidan, a sulfated fucose-containing polysaccharide extracted from brown algae, in a D-Gal mice model [32].
In addition to the observed improvements in behavioral cognitive function parameters, our study demonstrates that the microalgae extract of Phaeodactylum tricornutum (PT) regulates neuroinflammation and oxidative stress following D-GAL induction in mice, exhibiting a dose-response effect. Neurons, being metabolically robust and highly active consumers of oxygen, serve as major producers of reactive oxygen species (ROS) in their normal function [33,34]. A primary source of ROS is the mitochondrial respiratory complex, and due to its proximity to the site of production, mitochondrial DNA is particularly susceptible to oxidative damage (e.g. lipid peroxidation), leading to membrane fluidity [35]. Natural antioxidant processes in the nervous system are impaired during aging, mainly due to a downregulation of the Nrf2 pathway activity, resulting in redox potential imbalance. This imbalance leads to long-lasting accumulated cell damage (e.g., lipids, proteins, DNA) and has been identified as a major contributor to aging-related cognitive function decline [33,34,35]. Our study revealed a significant increase in lipid peroxidation levels in the mice hippocampus following D-GAL induction. The hippocampus, along with the frontal lobes, is a major brain region affected by oxidative stress with aging, directly contributing to alterations in learning, spatial working, and episodic memory [36,37]. Thus, the regulation and decrease of hippocampus lipid peroxidation levels observed following supplementation with the microalgae extract of PT may explain the benefits highlighted in cognitive function tests. Similar to mitochondrial dysfunction, cell senescence, or the general weakening of immune functions, the accumulation of oxidative stress is a major source of the upregulation of the production of pro-inflammatory cytokines. These, in turn, activate inducible nitric oxide synthase (iNOS) and NADPH oxidase (NOX) [38,39]. During aging, microglial macrophages, the resident innate immune cells of the central nervous system, become chronically active, inducing sustained production of pro-inflammatory cytokines such as interleukin-6 (IL-6), tumor necrosis factor-α (TNF-α), and proteins like C-reactive protein (CRP), well-recognized contributors to cognitive function decline [11]. The increase in these cytokines, initiated and regulated by the activation of the redox-sensitive transcription factor NF-kB, can induce a cycle of neuroinflammatory processes, including neuronal death, reduced brain volume, or cortical thinning [12]. Interestingly, an increase in peripheral inflammatory markers may activate neuroinflammation via neuronal and hormonal pathways [40,41]. As demonstrated in the present study, the microalgae extract of PT significantly decreased the levels of IL-6 and TNF-α at both systemic and hippocampus levels.
The microalgae extract of Phaeodactylum tricornutum (PT) contains several intriguing bioactive compounds, particularly fucoxanthin and omega-3 (Table 1), which may account for the observed benefits on oxidative stress and inflammation [20,33,42]. Firstly, fucoxanthin is a major carotenoid that exhibits antioxidant and anti-inflammatory activity, and its neuroprotective role has been highlighted in various in-vitro and in-vivo models [42,43,44]. Upon ingestion, fucoxanthin is rapidly metabolized by digestive enzymes to fucoxanthinol before absorption in the intestines, subsequently converting to amarouciaxanthin A in the liver. Apart from its ability to bypass the blood-brain barrier and strong singlet oxygen-quenching properties, which prevent oxidative stress by inhibiting H2O2-induced neurotoxicity (e.g., brain/systemic lipid peroxidation), fucoxanthin may prevent oxidative stress-induced neuronal death through the activation of the PI3 K/Akt/Nrf2 pathway. It can induce autophagy via the activation of BECLIN-1 and inhibit neuronal apoptosis through the inhibition of caspase-3. This process involves the translocation of the Nrf2 protein from the cytosol to the nucleus, facilitating the detachment of the keap1 protein. Nrf2 induces the expression of genes that activate the neuron's autophagy mechanism, eliminating intracellular waste caused by oxidative stress. Nrf2 also induces the expression of antioxidant enzymes such as HO-1 and NQO-1 by binding to the ARE in the promoter of antioxidant genes, preventing the accumulation of waste that amplifies oxidative stress [43]. Furthermore, fucoxanthin has been shown to inhibit the production of pro-inflammatory cytokines, including TNF-α, IL-6, and IL-1β, by inhibiting NF-κB and MAPK pathways in microglia, the major immune cells in the brain [42,44,45]. Secondly, eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA) are long-chain polyunsaturated fatty acids (LC-PUFAs) from the omega-3 family, essential for brain functions, given that the human brain comprises approximately 50-60% lipids [46]. EPA, in particular, acts as an agonist of the PPAR-α receptor (peroxisome proliferator-activated receptor alpha), allowing its activation and regulation of different signaling pathways. PPAR-α not only inhibits neuroinflammation and oxidative stress but also optimizes neurotransmission processes by altering membrane fluidity and increasing neurotransmitter release. LC-PUFAs also alleviate brain apoptosis through various mechanisms, such as reducing responses to reactive oxygen species, exerting an antiapoptotic action, or by up-regulating the expression of antiapoptotic proteins and down-regulating the expression of apoptotic proteins, thus slowing down the apoptosis reaction [46].
Overall, these data are promising, advocating for the promotion of the microalgae extract of Phaeodactylum tricornutum (PT) as a natural and viable alternative in the dietary supplement arena for improving cognitive function in humans. Notably, Leonard et al. (2023) demonstrated that both acute and 30-day ingestion of two different doses of the same microalgae extract of PT tested in the present study, combined with guarana in equal quantities across the experimental groups, improved reaction times, reasoning, learning, executive control, attention shifting (cognitive flexibility), and impulsiveness in e-gamers [47]. Our research team is currently conducting two clinical studies to evaluate the efficacy of PT microalgae on the cognitive function of healthy elderly individuals (NCT04832412 and NCT05759910). While additional pre-clinical studies are necessary to better comprehend and refine the underlying mechanisms of action, it would be intriguing to investigate the impact of this microalgae extract on the gut-brain axis. Over the past several years, a growing body of literature has underscored the bidirectional relationship between the gut and the brain [48,49]. Specifically, disruptions in gut homeostasis, influenced by various factors such as diet, infections, aging, or antibiotic exposure, can lead to compromised intestinal barrier integrity through tight junction impairment and apoptosis of gut epithelial cells [49]. Subsequently, chronic systemic inflammation and oxidative stress play a pivotal role in the brain by regulating the activity of multiple metabolites and neurotransmitters associated with specific signaling pathways [48,49]. As outlined in the review by Fakhri et al. (2021), marine-derived natural ingredients extracted from various sources, including microalgae, present a promising approach to positively modulate intestinal disorders, inflammatory mediators, apoptosis, and oxidative stress, thereby contributing to neuroprotective properties."
4. Materials and Methods
4.1. Animals, diet and treatment groups
Seventy-two (72) male swiss mice, weighing 30-35 g, from JANVIER company (Saint Berthevin, France), were kept for housing and experiments took place in Amylgen (Direction Régionale de l'Alimentation, de l'Agriculture et de la Forêt du Languedoc-Roussillon, agreement #A 34-169-002 from May 02, 2014). Animals were housed in groups with access to food and water ad libitum, except during behavioral experiments. They were kept in a temperature and humidity-controlled animal facility on a 12 h/12 h light/dark cycle (lights off at 07:00 pm). Mice were numbered by marking their tail with permanent markers. All animal procedures were conducted in strict adherence to the European Union directive of September 22, 2010 (2010/63/UE). The experiment design (protocol #02441) was approved by the Mediterranée Ethics Committee in January 2015.
Mice were randomly divided into blank control group (BC), negative control group (NC) and 4 microalgae extract of PT (branded name: BrainPhytTM) groups with different human equivalent doses evaluated (BP120, dose of 120 mg/day/70kg; BP235, dose of 235 mg/day/70kg; BP295, dose of 295 mg/day/70kg; BP370, dose of 370 mg/day/70kg) with 12 mice in each group. Microalgae extract of PT supplementation has been administered by incorporation into food pellets during all the study duration (between day -28 to day 51, total of 79 days) by Safe company (Augy, France) and kept at 4°C. BC and NC groups received normal food pellets. Between day 01 to day 51, mice in BC group were injected with 0.9% normal saline while mice in all other groups were subcutaneously injected with D-galactose (D-Gal) at a dose of 150 mg/kg once per day, 5 days per week.
Health of animals was checked daily as well as general aspect of animals, activity and weight were observed daily. Acute or delayed mortality was checked daily.
4.2. Microalgae extract of PT production and characterization
The microalgae extract of PT production process was based in microalgae cultures in industrial photobioreactors and all steps were realised in Microphyt (Baillargues, FRANCE). Microphyt has developed and operates its own proprietary production technology called CAMARGUE® technology which allows controlled microalgae biomass production. CAMARGUE® technology consists of a 1,200 m serpentine glass piping circuit, with a 76 mm inside diameter and 4.5 mm wall thickness, folded horizontally in 24 straight lines with a vertical height of 3 m, 0.3 m wide realising a 50 m long tubular fence. The global description of product manufacturer is presented on the schema below knowing that the first step consisted of biomass production from the microalgae Phaeodactylum tricornotum:
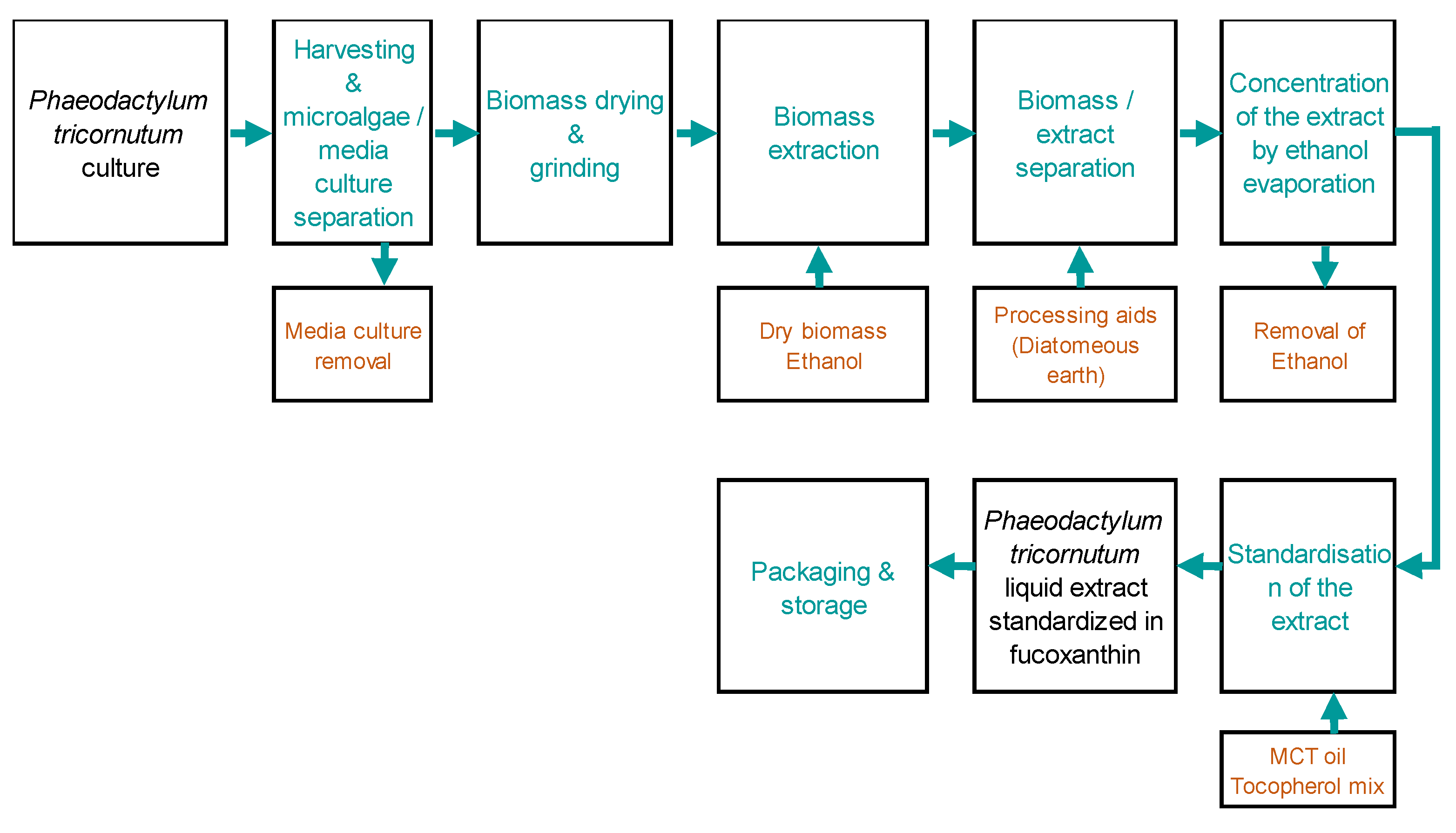
Figure 6.
Microalgae extract of PT production process by Microphyt company.
The crude ethanolic extract is standardised with regard to actives content by dilution with Medium Chain Triglycerides (MCT) and stabilised with antioxidants. Thus, the final product called microalgae extract of PT, is firstly standardised to contain 2% of FX. The global composition of microalgae extract of PT is presented on Table 1.

Table 1.
Microalgae extract of PT global specifications.
| Composition (%) | |
|---|---|
| Phaeodactylum tricornutum extract | 40 – 70 (w:w) |
| MCT oil based on coconut oil | 30 – 60 (w:w) |
| Mix on non-GMO tocopherols | 0.45 – 0.55 (w:w) |
| Total lipids | 60 – 90 (w:w) |
| Proteins | 5 – 15 (w:w) |
| Humidity | <2 (w:w) |
| Ashes | <10 (w:w) |
| Carbohydrates | 0.5 – 20 (w:w) |
| Total PUFAs-w3 | ≥4 (w:w) |
| All-trans-fucoxanthin | 2.0 ± 0.4% (w:w) |
The detailled composition of microalgae extract of PT regarding total chlorophylls, carotenoids and fatty acids contents is presented on Table 2. Total chlorophylls and carotenoids contents were determined by UV-Vis spectrophotometer while omega 3 fatty acids were quantified by gas chromatography.
4.3. Y-Maze spontaneous alternation performance
On day 43, all animals were tested for spontaneous alternation performance in the Y-maze, an index of spatial working memory. The Y-maze was designed according to Hiramatsu and al. (1999) and is made of grey polyvinylchloride [50]. Each arm is 40 cm long, 13 cm high, 3 cm wide at the bottom, 10 cm wide at the top, and converging at an equal angle. Each mice was placed at the end of one arm and allowed to move freely through the maze during an 8 min session. The series of arm entries, including possible returns into the same arm, were checked visually. An alternation was defined as entries into all three arms on consecutive occasions. The number of maximum alternations is therefore the total number of arm entries minus two and the percentage of alternation was calculated as (actual alternations / maximum alternations) × 100. Parameters included the percentage of alternation (memory index) and total number of arm entries (exploration index). Animals showing an extreme behavior (Alternation percentage < 20% or > 90% or number of arm entries < 10) were discarded from the calculation. No animal was discarded accordingly.
4.4. Place learning in the Morris Water-Maze
From day 44 up to day 49 all animals performed the Morris water-maze to test spatial working memory. The water maze consists of a circular pool (diameter 140 cm, height 40 cm) filled with water at temperature of 23 ± 1°C placed in a room with controlled light intensity, external cues (sink, contrasted posters, shelves), and water opacity rigorously reproduced. A transparent Plexiglas non-slippery platform (diameter 10 cm) is immersed under the water surface during acquisition. Training consisted in 3 swims per day for 5 days, performed between days 44 to 48, with 20 min inter-trial time. Animals were tested by batch of 12 individuals. Start positions, set at each limit between quadrants, were randomly selected each day and each animal was allowed a 90 s swim in order to find the platform and it was left on it for 20 s. The median latency, expressed as mean ± S.E.M., was calculated for each training day. Working memory was specifically assayed by changing the platform location every day. A probe test (PT) was performed 24 h after the last swim, on day 49. The platform was removed and each animal was allowed a free 60 s swim. The start position for each mice corresponds to one of two positions remote from the platform location in counterbalanced order. The platform quadrant was termed the training (T) quadrant and others opposite (O), adjacent right (R), and adjacent left (L) during retention. The time spent in each quadrant was determined. The results were expressed as time spent in target quadrant (T) compared to the mean of the three other quadrants (O).
4.5. Passive avoidance test
On days 50 and 51, all animals were tested for the STPA task. The apparatus is a two compartments (15 × 20 × 15 cm high) box, whereas one is illuminated with white polyvinylchloride walls and the other is darkened with black polyvinylchloride walls and a grid floor. A guillotine door separates each compartment. A 60 W lamp positioned 40 cm above the apparatus lights up the white compartment during the experiment. Scrambled foo shocks (0.3 mA for 3 s) were delivered to the grid floor using a shock generator scrambler (Lafayette Instruments, Lafayette, USA). The guillotine door was initially closed during the training session. During training session, each mice was placed into the white compartment. After 5 s, the door was raised. When the mice entered into the dark compartment and placed all its paws on the grid floor, the door was closed and the foot-shock delivered for 3 s. The step through latency, that is the latency spent to enter into the dark compartment, and the number of vocalizations were recorded. The retention test was carried out 24 h after training. Each mice was placed again into the white compartment. After 5 s, the door was raised. The step-through and escape latencies (corresponding to the re-exit from the dark compartment) were recorded up to 300 s. Animals that show latencies during the training and retention session both lower than 10 s are considered as failing to respond to the procedure and are discarded from the calculations. In this study, no animal was discarded accordingly.
4.6. Biochemical analysis
On day 51, all animals were sacrificed by decapitation without prior anesthesia after the behavioral session. For all animals, trunk blood was collected into EDTA-coated tubes that were inverted several times and placed on ice until plasma separation through centrifugation (2000 g for 10 min at 4°C). Brain was quickly removed and hippocampus and frontal cortex dissected out on an ice-cold metal plate. On n = 6 animals per group, hippocampus was used to determine the lipid peroxidation levels (LPO) and the cortex was used for measuring TNFα and IL-6. The other brain structures and plasma were frozen on dry ice and stored at -80°C at Amylgen for two months and available for supplementary biochemical assays.
Regarding LPO analysis, the procedure was modified and adapted from Hermes-Lima and al., (1995) [51]. This method measures the ability of brain peroxidized lipids to oxidize a ferrous oxide / xylenol orange complex. This procedure was named FOX assay (ferrous oxidation/xylenol orange method) and was shown to be a fast and accurate method to measure lipid peroxides. A comparison between the FOX method and the TBARS (thiobarbituric acid reactive substances) assay was made previously by Hermes-Lima et al. (1995) The authors determined that the TBARS assay detects products at different stages of lipid peroxidation, whereas the FOX assay has the advantage of direct detection of lipid peroxides. Six (6) hippocampi from each group were used. After thawing on ice, samples were homogenized by ultrasound (1 kJ, 10 seconds, Sonopuls, Bandelin, France) in cold methanol (1/10 w/v), centrifuged at 1,000 g during 5 min and the supernatant placed in Eppendorf tubes. The reaction volume of each homogenate was added to FeSO4 1 mM, H2SO4 0.25 M, xylenol orange 1 mM and incubated for 30 min at room temperature. After reading the absorbance at 580 nm (A5801), 10 µl of cumene hydroperoxyde (CHP) 1 mM were added to the sample and incubated for 30 min at room temperature, to determine the maximal oxidation level. The absorbance was measured at 580 nm (A5802). The level of lipid peroxidation is determined as CHP equivalents according to: CHPE = A5801/A5802 x [CHP (nmol)] and expressed as CHP equivalents per wet weight of tissue and as percentage of the mean BC group data.
Regarding brain and plasma inflammatory markers level, contents in IL6 and TNFα were analyzed by ELISA assays. For all assays, the cortex was homogenized after thawing in 50 mM Tris-150 mM NaCl buffer, pH 7.5, and sonicated for 20 s. After centrifugation (16,100 g for 15 min, 4°C), supernatant or plasma was used for ELISA assays according to the manufacturer instructions. For each assay, absorbance was read at 450 nm and sample concentration was calculated using the standard curve. Results are expressed in pg of marker per mg of tissue. Then, the results were expressed as percentage of the mean BC group data. N = 10 per group were assayed.
4.7. Statistical analysis
All values, except passive avoidance latencies, were expressed as mean ± S.D. (Standard Deviation). Statistical analyses were performed separately for each compound using one-way ANOVA (F value), followed by the Tukey’s post-hoc multiple comparison test. Passive avoidance latencies do not follow a Gaussian distribution, since upper cut-off times are set. They were therefore analyzed using a Kruskal-Wallis non-parametric ANOVA (H value), followed by a Tukey’s multiple comparison test. Values with p < 0.05 were considered as statistically significant. For Morris Water Maze data analysis, a Tukey’s multiple comparison test was performed after two-way ANOVA.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, R.P. and A.D.; methodology, R.P., A.D. and V.V.; validation, R.P., V.V.; formal analysis, R.P., V.V.; investigation, J.M.; writing—original draft preparation, J.M.; writing—review and editing, J.M., R.P., A.D. and V.V.; supervision, R.P. and A.D. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by MICROPHYT company (Baillargues, FRANCE) as a fee for service project awarded to the Amylgen SAS (Montferrier-sur-Lez, FRANCE) and was conducted by their staff in their facilities.
Institutional Review Board Statement
The animal study protocol was approved by the Ethics Committee of Mediterranean (protocol #02441 approved in January 2015).
Data Availability Statement
Raw data are available from corresponding author upon justified requests
Conflicts of Interest
J.M., A.D. and R.P. are employees of Microphyt company but they had no role in the collection, analyses and interpretation of data. They contributed only in the study design writing of the manuscript and in the decision to publish the results.
References
- World Population Prospects, the 2012 Revision; 2013.
- Salthouse, T. Consequences of Age-Related Cognitive Declines. Annu Rev Psychol 2012, 63, 201–226. [Google Scholar] [CrossRef] [PubMed]
- Salthouse, T.A. When Does Age-Related Cognitive Decline Begin? Neurobiol Aging 2009, 30, 507–514. [Google Scholar] [CrossRef] [PubMed]
- Singh-Manoux, A.; Kivimaki, M.; Glymour, M.M.; Elbaz, A.; Berr, C.; Ebmeier, K.P.; Ferrie, J.E.; Dugravot, A. Timing of Onset of Cognitive Decline: Results from Whitehall II Prospective Cohort Study. BMJ 2012, 344, d7622. [Google Scholar] [CrossRef] [PubMed]
- Hedden, T.; Gabrieli, J.D.E. Insights into the Ageing Mind: A View from Cognitive Neuroscience. Nat Rev Neurosci 2004, 5, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Freemantle, E.; Vandal, M.; Tremblay-Mercier, J.; Tremblay, S.; Blachère, J.-C.; Bégin, M.E.; Brenna, J.T.; Windust, A.; Cunnane, S.C. Omega-3 Fatty Acids, Energy Substrates, and Brain Function during Aging. Prostaglandins Leukot Essent Fatty Acids 2006, 75, 213–220. [Google Scholar] [CrossRef] [PubMed]
- Andlin-Sobocki, P.; Jönsson, B.; Wittchen, H.-U.; Olesen, J. Cost of Disorders of the Brain in Europe. Eur J Neurol 2005, 12 Suppl 1, 1–27. [Google Scholar] [CrossRef]
- Harada, C.N.; Natelson Love, M.C.; Triebel, K.L. Normal Cognitive Aging. Clin Geriatr Med 2013, 29, 737–752. [Google Scholar] [CrossRef]
- Scarmeas, N.; Anastasiou, C.A.; Yannakoulia, M. Nutrition and Prevention of Cognitive Impairment. Lancet Neurol 2018, 17, 1006–1015. [Google Scholar] [CrossRef]
- Tönnies, E.; Trushina, E. Oxidative Stress, Synaptic Dysfunction, and Alzheimer’s Disease. J Alzheimers Dis 2017, 57, 1105–1121. [Google Scholar] [CrossRef]
- McGrattan, A.M.; McGuinness, B.; McKinley, M.C.; Kee, F.; Passmore, P.; Woodside, J.V.; McEvoy, C.T. Diet and Inflammation in Cognitive Ageing and Alzheimer’s Disease. Curr Nutr Rep 2019, 8, 53–65. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Vorburger, R.; Scarmeas, N.; Luchsinger, J.A.; Manly, J.J.; Schupf, N.; Mayeux, R.; Brickman, A.M. Circulating Inflammatory Biomarkers in Relation to Brain Structural Measurements in a Non-Demented Elderly Population. Brain Behav Immun 2017, 65, 150–160. [Google Scholar] [CrossRef] [PubMed]
- Swomley, A.M.; Butterfield, D.A. Oxidative Stress in Alzheimer Disease and Mild Cognitive Impairment: Evidence from Human Data Provided by Redox Proteomics. Arch Toxicol 2015, 89, 1669–1680. [Google Scholar] [CrossRef] [PubMed]
- Valls-Pedret, C.; Sala-Vila, A.; Serra-Mir, M.; Corella, D.; de la Torre, R.; Martínez-González, M.Á.; Martínez-Lapiscina, E.H.; Fitó, M.; Pérez-Heras, A.; Salas-Salvadó, J.; et al. Mediterranean Diet and Age-Related Cognitive Decline: A Randomized Clinical Trial. JAMA Intern Med 2015, 175, 1094–1103. [Google Scholar] [CrossRef] [PubMed]
- Johnson, T.J.; Katuwal, S.; Anderson, G.A.; Gu, L.; Zhou, R.; Gibbons, W.R. Photobioreactor Cultivation Strategies for Microalgae and Cyanobacteria. Biotechnol Prog 2018, 34, 811–827. [Google Scholar] [CrossRef] [PubMed]
- Delbrut, A.; Albina, P.; Lapierre, T.; Pradelles, R.; Dubreucq, E. Fucoxanthin and Polyunsaturated Fatty Acids Co-Extraction by a Green Process. Molecules 2018, 23, E874. [Google Scholar] [CrossRef] [PubMed]
- Salem, N.; Vandal, M.; Calon, F. The Benefit of Docosahexaenoic Acid for the Adult Brain in Aging and Dementia. Prostaglandins Leukot Essent Fatty Acids 2015, 92, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Yurko-Mauro, K.; McCarthy, D.; Rom, D.; Nelson, E.B.; Ryan, A.S.; Blackwell, A.; Salem, N.; Stedman, M. MIDAS Investigators Beneficial Effects of Docosahexaenoic Acid on Cognition in Age-Related Cognitive Decline. Alzheimers Dement 2010, 6, 456–464. [Google Scholar] [CrossRef] [PubMed]
- Aziz, E.; Batool, R.; Akhtar, W.; Rehman, S.; Shahzad, T.; Malik, A.; Shariati, M.A.; Laishevtcev, A.; Plygun, S.; Heydari, M.; et al. Xanthophyll: Health Benefits and Therapeutic Insights. Life Sci 2020, 240, 117104. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Chen, W.; Tian, F.; Yuan, C.; Wang, H.; Yue, H. Neuroprotective Role of Fucoxanthin against Cerebral Ischemic/Reperfusion Injury through Activation of Nrf2/HO-1 Signaling. Biomed Pharmacother 2018, 106, 1484–1489. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, Y.; Chen, L.; Cheng, W.; Liu, T. Combined Production of Fucoxanthin and EPA from Two Diatom Strains Phaeodactylum Tricornutum and Cylindrotheca Fusiformis Cultures. Bioprocess Biosyst Eng 2018, 41, 1061–1071. [Google Scholar] [CrossRef] [PubMed]
- Minghetti, L.; Salvi, R.; Lavinia Salvatori, M.; Ajmone-Cat, M.A.; De Nuccio, C.; Visentin, S.; Bultel-Poncé, V.; Oger, C.; Guy, A.; Galano, J.-M.; et al. Nonenzymatic Oxygenated Metabolites of α-Linolenic Acid B1- and L1-Phytoprostanes Protect Immature Neurons from Oxidant Injury and Promote Differentiation of Oligodendrocyte Progenitors through PPAR-γ Activation. Free Radic Biol Med 2014, 73, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Fotuhi, M.; Mohassel, P.; Yaffe, K. Fish Consumption, Long-Chain Omega-3 Fatty Acids and Risk of Cognitive Decline or Alzheimer Disease: A Complex Association. Nat Clin Pract Neurol 2009, 5, 140–152. [Google Scholar] [CrossRef] [PubMed]
- Dyall, S.C. Long-Chain Omega-3 Fatty Acids and the Brain: A Review of the Independent and Shared Effects of EPA, DPA and DHA. Front Aging Neurosci 2015, 7, 52. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Li, L.; Song, Q.; Ai, H.; Chu, J.; Li, W. Behavioural Study of the D-Galactose Induced Aging Model in C57BL/6J Mice. Behav Brain Res 2005, 157, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Lu, J.; Zheng, Y.; Zhou, Z.; Shan, Q.; Ma, D. Purple Sweet Potato Color Repairs D-Galactose-Induced Spatial Learning and Memory Impairment by Regulating the Expression of Synaptic Proteins. Neurobiol Learn Mem 2008, 90, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Grimmig, B.; Hudson, C.; Moss, L.; Peters, M.; Subbarayan, M.; Weeber, E.J.; Bickford, P.C. Astaxanthin Supplementation Modulates Cognitive Function and Synaptic Plasticity in Young and Aged Mice. Geroscience 2019, 41, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Yook, J.S.; Okamoto, M.; Rakwal, R.; Shibato, J.; Lee, M.C.; Matsui, T.; Chang, H.; Cho, J.Y.; Soya, H. Astaxanthin Supplementation Enhances Adult Hippocampal Neurogenesis and Spatial Memory in Mice. Mol Nutr Food Res 2016, 60, 589–599. [Google Scholar] [CrossRef] [PubMed]
- Farr, S.A.; Niehoff, M.L.; Ceddia, M.A.; Herrlinger, K.A.; Lewis, B.J.; Feng, S.; Welleford, A.; Butterfield, D.A.; Morley, J.E. Effect of Botanical Extracts Containing Carnosic Acid or Rosmarinic Acid on Learning and Memory in SAMP8 Mice. Physiol Behav 2016, 165, 328–338. [Google Scholar] [CrossRef] [PubMed]
- Abdelmeguid, N.E.; Khalil, M.I.M.; Elhabet, R.; Sultan, A.S.; Salam, S.A. Combination of Docosahexaenoic Acid and Ginko Biloba Extract Improves Cognitive Function and Hippocampal Tissue Damages in a Mouse Model of Alzheimer’s Disease. J Chem Neuroanat 2021, 116, 101995. [Google Scholar] [CrossRef] [PubMed]
- Ren, Q.; Sun, J.; Xu, D.; Xie, H.; Ye, M.; Zhao, Y. A Dietary Supplement Containing Micronutrients, Phosphatidylserine, and Docosahexaenoic Acid Counteracts Cognitive Impairment in D-Galactose-Induced Aged Rats. Frontiers in Nutrition 2022, 9. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Gao, Z.; Zheng, L.; Zhang, C.; Liu, Z.; Yang, Y.; Teng, H.; Hou, L.; Yin, Y.; Zou, X. Protective Effects of Fucoidan on Aβ25–35 and d-Gal-Induced Neurotoxicity in PC12 Cells and d-Gal-Induced Cognitive Dysfunction in Mice. Mar Drugs 2017, 15, 77. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Xuan, Z.; Wang, Q.; Yan, S.; Zhou, D.; Naman, C.B.; Zhang, J.; He, S.; Yan, X.; Cui, W. Fucoxanthin Has Potential for Therapeutic Efficacy in Neurodegenerative Disorders by Acting on Multiple Targets. Nutritional Neuroscience 2022, 25, 2167–2180. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Kukreti, R.; Saso, L.; Kukreti, S. Oxidative Stress: A Key Modulator in Neurodegenerative Diseases. Molecules 2019, 24, 1583. [Google Scholar] [CrossRef] [PubMed]
- Andronie-Cioara, F.L.; Ardelean, A.I.; Nistor-Cseppento, C.D.; Jurcau, A.; Jurcau, M.C.; Pascalau, N.; Marcu, F. Molecular Mechanisms of Neuroinflammation in Aging and Alzheimer’s Disease Progression. Int J Mol Sci 2023, 24, 1869. [Google Scholar] [CrossRef] [PubMed]
- Peters, M.M.; Hill, K.E.; Burk, R.F.; Weeber, E.J. Altered Hippocampus Synaptic Function in Selenoprotein P Deficient Mice. Molecular Neurodegeneration 2006, 1, 12. [Google Scholar] [CrossRef] [PubMed]
- Papp, L.; Vizi, E.S.; Sperlágh, B. P2X7 Receptor Mediated Phosphorylation of p38MAP Kinase in the Hippocampus. Biochemical and Biophysical Research Communications 2007, 355, 568–574. [Google Scholar] [CrossRef] [PubMed]
- Sanada, F.; Taniyama, Y.; Muratsu, J.; Otsu, R.; Shimizu, H.; Rakugi, H.; Morishita, R. Source of Chronic Inflammation in Aging. Front Cardiovasc Med 2018, 5, 12. [Google Scholar] [CrossRef] [PubMed]
- Kobylarek, D.; Iwanowski, P.; Lewandowska, Z.; Limphaibool, N.; Szafranek, S.; Labrzycka, A.; Kozubski, W. Advances in the Potential Biomarkers of Epilepsy. Front Neurol 2019, 10, 685. [Google Scholar] [CrossRef] [PubMed]
- Koyama, A.; O’Brien, J.; Weuve, J.; Blacker, D.; Metti, A.L.; Yaffe, K. The Role of Peripheral Inflammatory Markers in Dementia and Alzheimer’s Disease: A Meta-Analysis. J Gerontol A Biol Sci Med Sci 2013, 68, 433–440. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C. Omega-3 Fatty Acids and Inflammatory Processes: From Molecules to Man. Biochem Soc Trans 2017, 45, 1105–1115. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Wang, H.; Fan, Y.; Gao, Y.; Li, X.; Hu, Z.; Ding, K.; Wang, Y.; Wang, X. Fucoxanthin Provides Neuroprotection in Models of Traumatic Brain Injury via the Nrf2-ARE and Nrf2-Autophagy Pathways. Sci Rep 2017, 7, 46763. [Google Scholar] [CrossRef] [PubMed]
- Xiang, S.; Liu, F.; Lin, J.; Chen, H.; Huang, C.; Chen, L.; Zhou, Y.; Ye, L.; Zhang, K.; Jin, J.; et al. Fucoxanthin Inhibits β-Amyloid Assembly and Attenuates β-Amyloid Oligomer-Induced Cognitive Impairments. J Agric Food Chem 2017, 65, 4092–4102. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.-N.; Heo, S.-J.; Yoon, W.-J.; Kang, S.-M.; Ahn, G.; Yi, T.-H.; Jeon, Y.-J. Fucoxanthin Inhibits the Inflammatory Response by Suppressing the Activation of NF-κB and MAPKs in Lipopolysaccharide-Induced RAW 264.7 Macrophages. Eur J Pharmacol 2010, 649, 369–375. [Google Scholar] [CrossRef]
- Subhramanyam, C.S.; Wang, C.; Hu, Q.; Dheen, S.T. Microglia-Mediated Neuroinflammation in Neurodegenerative Diseases. Semin Cell Dev Biol 2019, 94, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Dighriri, I.M.; Alsubaie, A.M.; Hakami, F.M.; Hamithi, D.M.; Alshekh, M.M.; Khobrani, F.A.; Dalak, F.E.; Hakami, A.A.; Alsueaadi, E.H.; Alsaawi, L.S.; et al. Effects of Omega-3 Polyunsaturated Fatty Acids on Brain Functions: A Systematic Review. Cureus 2022, 14, e30091. [Google Scholar] [CrossRef] [PubMed]
- Leonard, M.; Maury, J.; Dickerson, B.; Gonzalez, D.E.; Kendra, J.; Jenkins, V.; Nottingham, K.; Yoo, C.; Xing, D.; Ko, J.; et al. Effects of Dietary Supplementation of a Microalgae Extract Containing Fucoxanthin Combined with Guarana on Cognitive Function and Gaming Performance. Nutrients 2023, 15, 1918. [Google Scholar] [CrossRef] [PubMed]
- Fakhri, S.; Yarmohammadi, A.; Yarmohammadi, M.; Farzaei, M.H.; Echeverria, J. Marine Natural Products: Promising Candidates in the Modulation of Gut-Brain Axis towards Neuroprotection. Mar Drugs 2021, 19, 165. [Google Scholar] [CrossRef] [PubMed]
- Mou, Y.; Du, Y.; Zhou, L.; Yue, J.; Hu, X.; Liu, Y.; Chen, S.; Lin, X.; Zhang, G.; Xiao, H.; et al. Gut Microbiota Interact With the Brain Through Systemic Chronic Inflammation: Implications on Neuroinflammation, Neurodegeneration, and Aging. Front Immunol 2022, 13, 796288. [Google Scholar] [CrossRef] [PubMed]
- Hiramatsu, M.; Inoue, K. Nociceptin/Orphanin FQ and Nocistatin on Learning and Memory Impairment Induced by Scopolamine in Mice. Br J Pharmacol 1999, 127, 655–660. [Google Scholar] [CrossRef] [PubMed]
- Hermes-Lima, M.; Willmore, W.G.; Storey, K.B. Quantification of Lipid Peroxidation in Tissue Extracts Based on Fe(III)Xylenol Orange Complex Formation. Free Radic Biol Med 1995, 19, 271–280. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Effects of microalgae extract of PT on D-Gal-induced spontaneous alternation deficits in mice. n is between 11 and 12 depending on the groups; * p < 0.05, **** p < 0.0001 vs. the Saline solution / Normal food group (BC group), ### p < 0.001, #### p < 0.0001 vs. the D-GAL 150 / Veh group (NC group); One-way ANOVA (F value), followed by the Tukey’s post-hoc multiple comparison test.
Figure 1.
Effects of microalgae extract of PT on D-Gal-induced spontaneous alternation deficits in mice. n is between 11 and 12 depending on the groups; * p < 0.05, **** p < 0.0001 vs. the Saline solution / Normal food group (BC group), ### p < 0.001, #### p < 0.0001 vs. the D-GAL 150 / Veh group (NC group); One-way ANOVA (F value), followed by the Tukey’s post-hoc multiple comparison test.
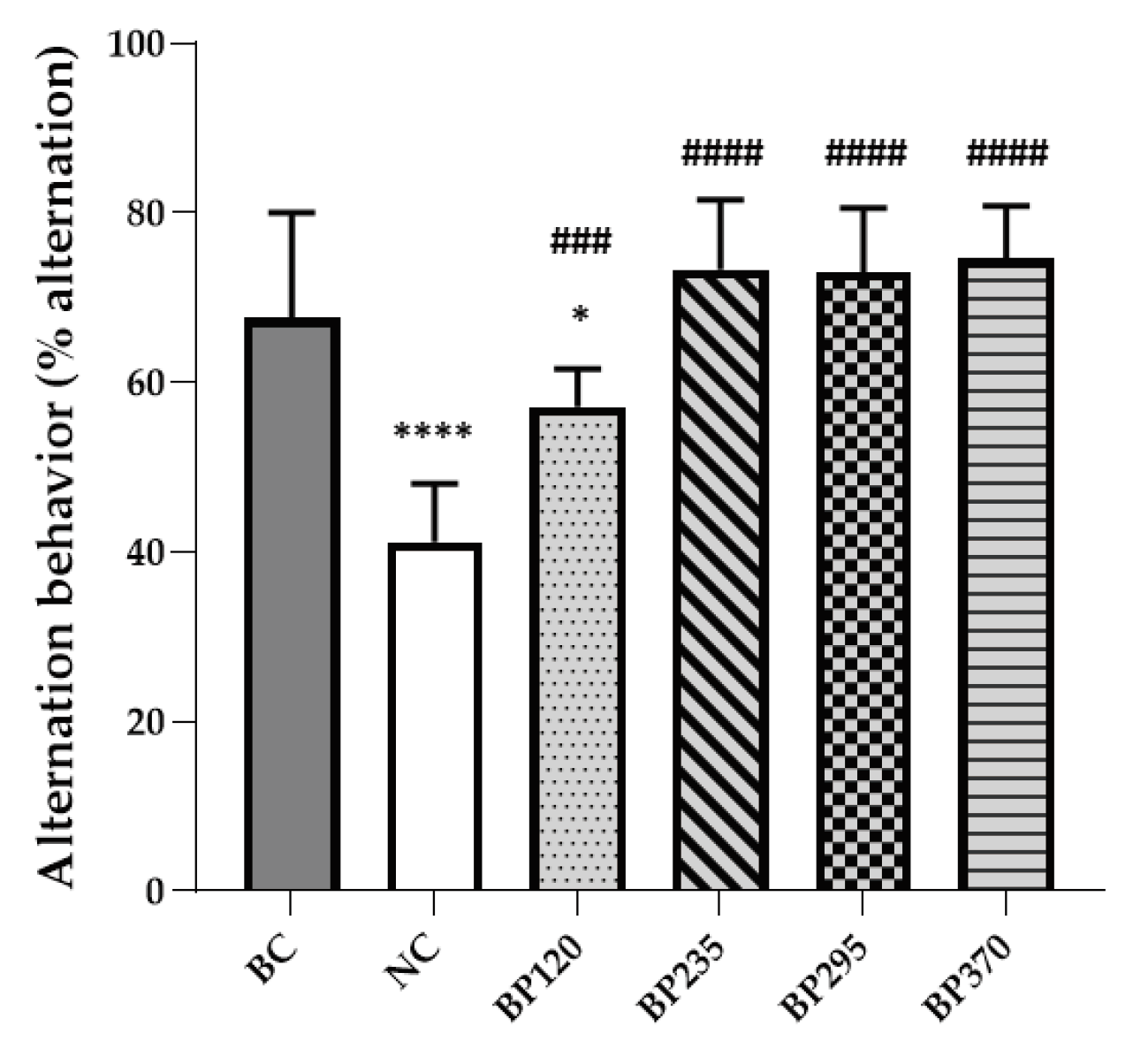
Figure 2.
Effects of microalgae extract of PT on D-Gal-induced learning deficits in mice. (a) swin latency (seconds). (b) time spent in target quadrant (seconds). n is between 11 and 12 depending on the groups; * p < 0.05 vs. the Saline solution / Normal Food group (BC group); p < 0.0001 vs. the D-GAL 150 / Veh group (NC group); Tukey’s multiple comparison test after two-way ANOVA.
Figure 2.
Effects of microalgae extract of PT on D-Gal-induced learning deficits in mice. (a) swin latency (seconds). (b) time spent in target quadrant (seconds). n is between 11 and 12 depending on the groups; * p < 0.05 vs. the Saline solution / Normal Food group (BC group); p < 0.0001 vs. the D-GAL 150 / Veh group (NC group); Tukey’s multiple comparison test after two-way ANOVA.
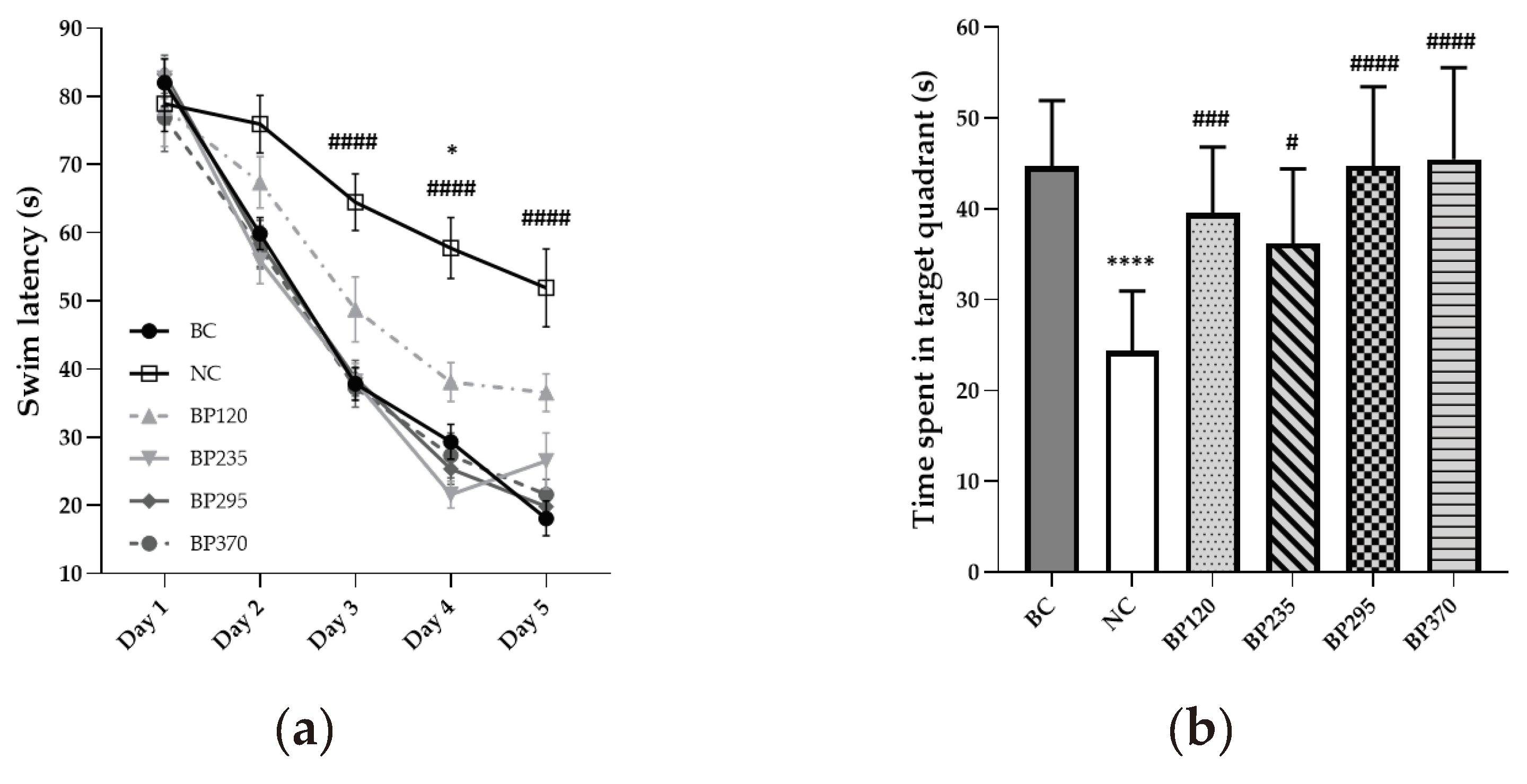
Figure 3.
Effects of Microalgae extract of PT on D-Gal-induced contextual long-term memory deficits in mice. (a) Step through latency (seconds). (b) Escape latency (seconds). n is between 11 and 12 depending on the groups; ** p < 0.01, **** p < 0.0001 vs. the Saline solution / Normal Food group (BC group), ## p < 0.01, #### p < 0.0001 vs. the D-GAL 150 / Veh group (NC group); One-way ANOVA (F value), followed by the Tukey’s post-hoc multiple comparison test.
Figure 3.
Effects of Microalgae extract of PT on D-Gal-induced contextual long-term memory deficits in mice. (a) Step through latency (seconds). (b) Escape latency (seconds). n is between 11 and 12 depending on the groups; ** p < 0.01, **** p < 0.0001 vs. the Saline solution / Normal Food group (BC group), ## p < 0.01, #### p < 0.0001 vs. the D-GAL 150 / Veh group (NC group); One-way ANOVA (F value), followed by the Tukey’s post-hoc multiple comparison test.
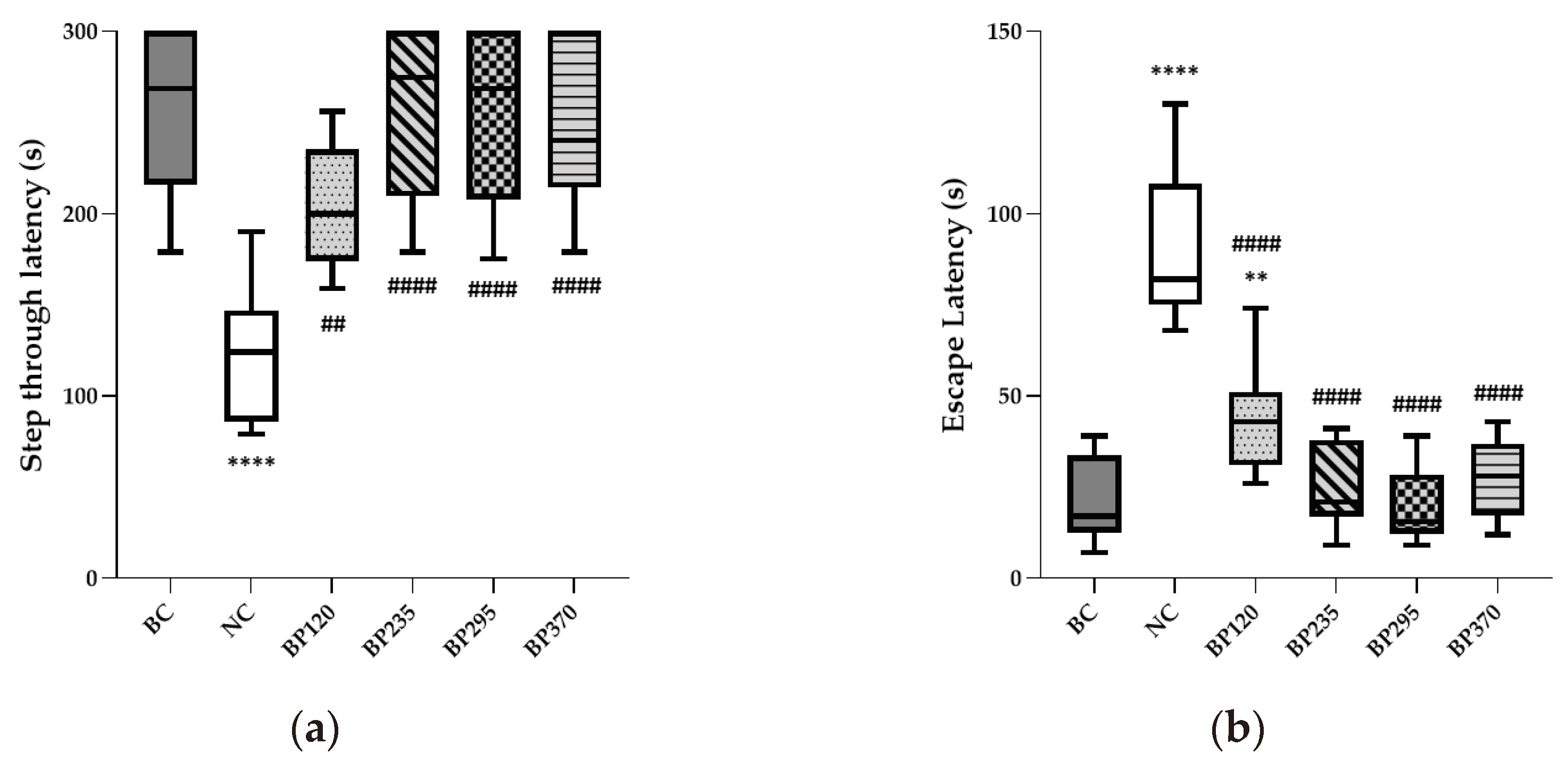
Figure 4.
Effects of microalgae extract of PT on D-Gal-induced lipid peroxidation increase in mice hippocampus. n is 6 per groups; * p < 0.05, **** p < 0.0001 vs. the Saline solution/Normal Food group (BC group), ### p < 0.001, #### p < 0.001 vs. the D-GAL 150/Veh group (NC group); One-way ANOVA (F value), followed by the Tukey’s post-hoc multiple comparison test.
Figure 4.
Effects of microalgae extract of PT on D-Gal-induced lipid peroxidation increase in mice hippocampus. n is 6 per groups; * p < 0.05, **** p < 0.0001 vs. the Saline solution/Normal Food group (BC group), ### p < 0.001, #### p < 0.001 vs. the D-GAL 150/Veh group (NC group); One-way ANOVA (F value), followed by the Tukey’s post-hoc multiple comparison test.
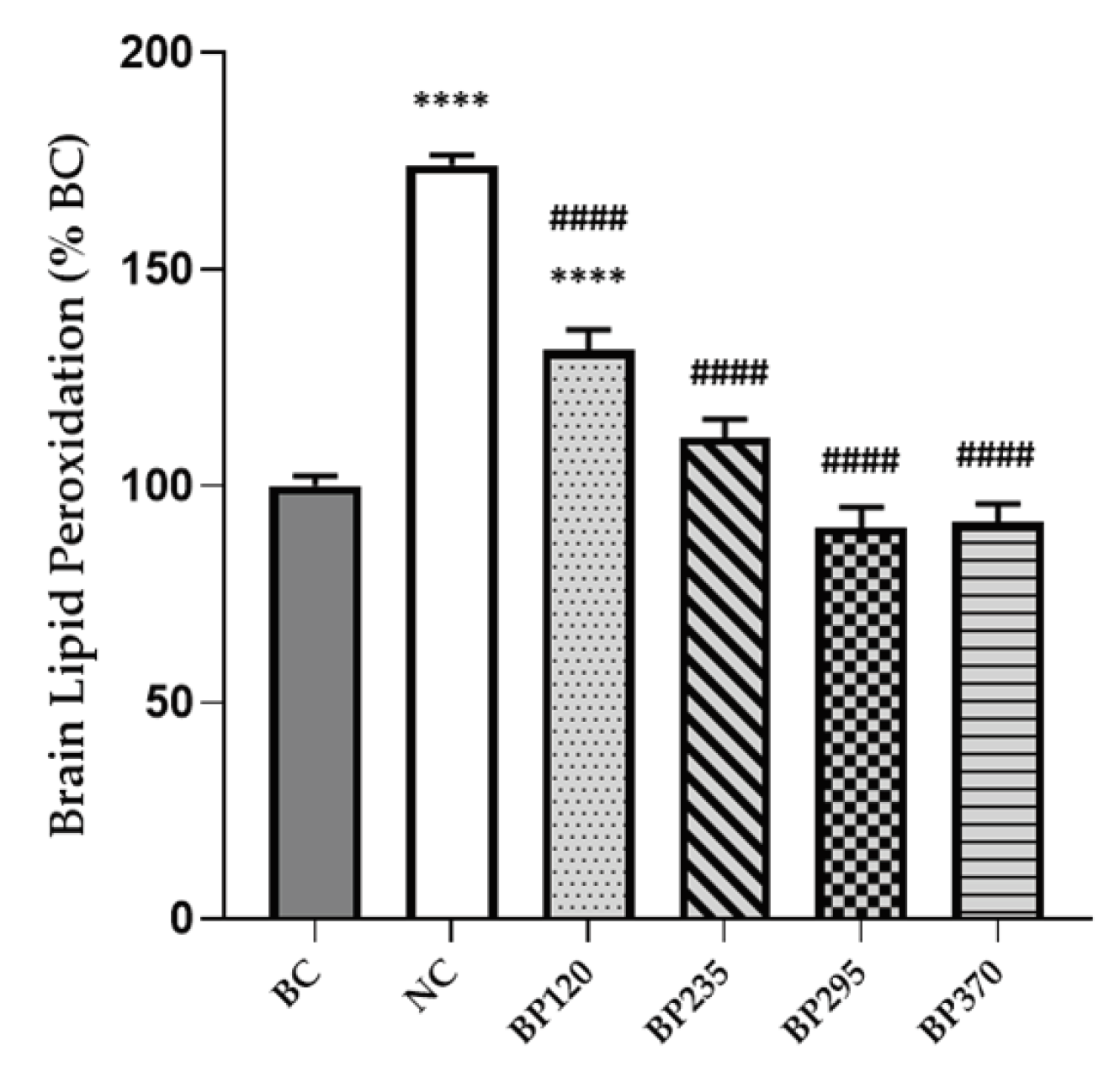
Figure 5.
Effects of microalgae extract of PT on D-Gal-induced brain and plasma inflammatory markers increase in mice. (a) TNF-α brain content (% BC). (b) TNF-α plasma content (% BC). (c) IL-6 brain content (% BC). (d) IL-6 plasma content (% BC). n is 10 per groups; ** p < 0.01, **** p < 0.0001 vs. the Saline solution / Normal Food group (BC group), ## p < 0.01, ### p < 0.001, #### p < 0.0001 vs. the D-GAL 150 / Veh group (NC group); One-way ANOVA (F value), followed by the Tukey’s post-hoc multiple comparison test.
Figure 5.
Effects of microalgae extract of PT on D-Gal-induced brain and plasma inflammatory markers increase in mice. (a) TNF-α brain content (% BC). (b) TNF-α plasma content (% BC). (c) IL-6 brain content (% BC). (d) IL-6 plasma content (% BC). n is 10 per groups; ** p < 0.01, **** p < 0.0001 vs. the Saline solution / Normal Food group (BC group), ## p < 0.01, ### p < 0.001, #### p < 0.0001 vs. the D-GAL 150 / Veh group (NC group); One-way ANOVA (F value), followed by the Tukey’s post-hoc multiple comparison test.
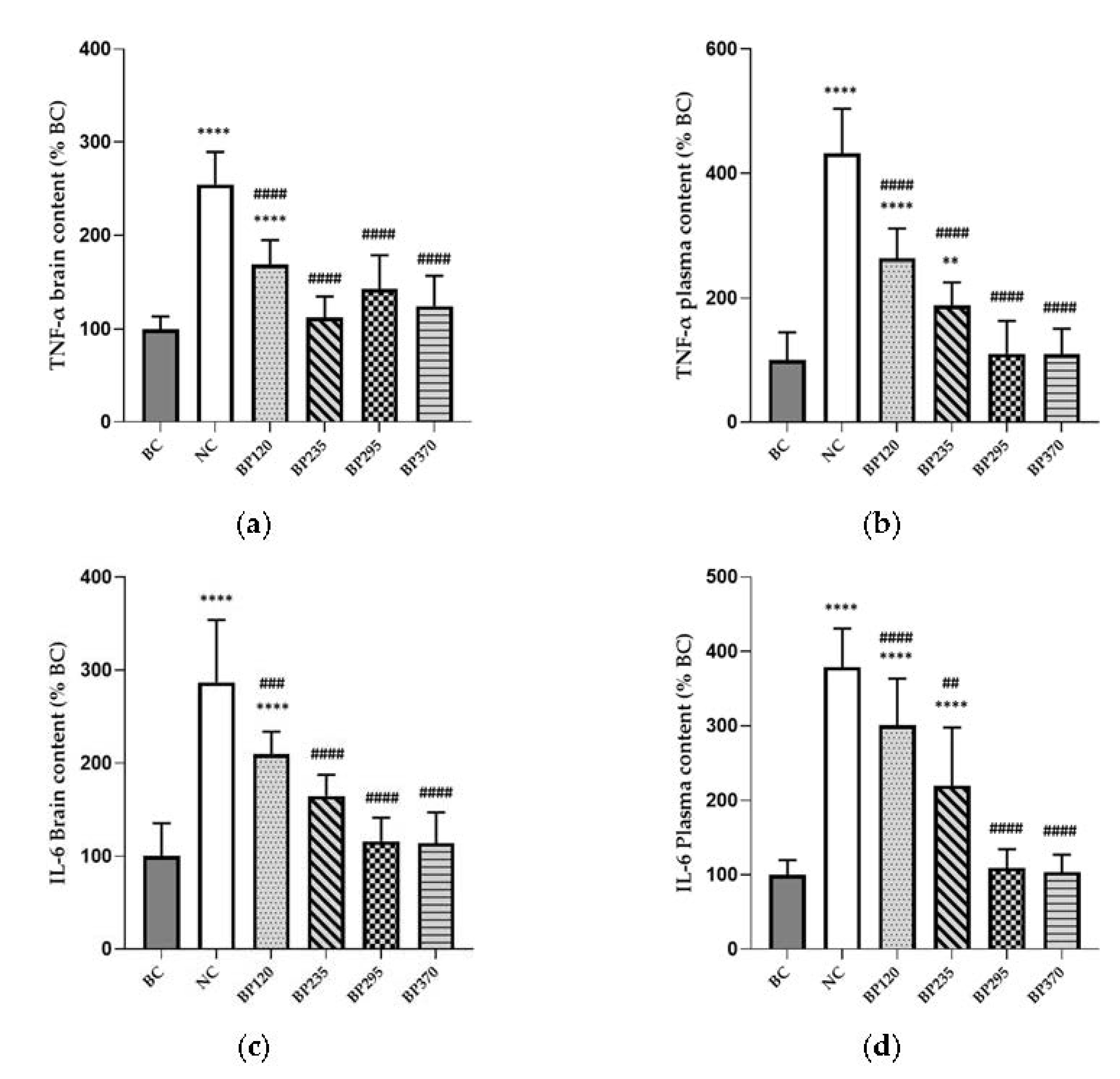
Table 2.
Main chlorophylls, total carotenoids and fatty acids composition of microalgae extract of PT. Data are presented as mean ± SD from six independent industrial-scale batches.
Table 2.
Main chlorophylls, total carotenoids and fatty acids composition of microalgae extract of PT. Data are presented as mean ± SD from six independent industrial-scale batches.
| Concentration | |
|---|---|
| Chlorophylls (g equivalent chloropyll A/100g) | |
| Chlorophyll C | 3.46 ± 0.45 |
| Σ Chlorophyll B | ND |
| Σ Chlorophyll A | 0.24 ± 0.25 |
| Total carotenoids | |
| Β-carotene (g equivalent Β-carotene / 100g) | 0.09 ± 0.08 |
| All trans Fucoxanthin (g equivalent fucoxanthin/100g) | 1.99 ± 1.01 |
| Total unsaturated fatty acids omega 3 (g/100g of product) | 6.51 ± 1.17 |
| Eicosapentaenoic acid (EPA) | 7.99 ± 6.14 |
| Docosahexaenoic acid (DHA) | 0.23 ± 0.20 |
| Alpha-linolenic acid (ALA) | 0.24 ± 0.17 |
| Stearidonic acid (SDA) | 0.07 ± 0.08 |
| Total unsaturated fatty acids omega 6 (g/100g of product) | 2.56 ± 0.19 |
| Hexadecadienoic acid | 2.01 ± 1.38 |
| Linoleic acid | 0.83 ± 0.66 |
| Arachidonic acid | 0.52 ± 0.44 |
| Eicosatetraenoic acid | 0.14 ± 0.12 |
| Total saturated fatty acids (g/100g of product) | 45.42 ± 8.49 |
| Caprylic acid | 23.24 ± 5.35 |
| Capric acid | 18.23 ± 4.06 |
| Palmitic acid | 2.53 ± 0.60 |
| Myristic acid | 0.90 ± 0.25 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



