
Submitted:
03 February 2024
Posted:
05 February 2024
You are already at the latest version
Abstract
The rapid and sensitive detection of pathogenic bacteria is becoming increasingly important for the timely prevention of contamination and treatment of infections. Biosensors based on nucleic acid aptamers integrat-ed with optical, electrochemical, and mass-sensitive analytical techniques have garnered intense interest for their versatility, cost-efficiency, and ability to exhibit high affinity and specificity in binding bacterial bi-omarkers, toxins, and whole cells. This review emphasizes the development of aptamers, their structural op-timization, and their use in bacterial sensing applications. Moreover, we explore barriers and develop per-spectives in applying aptamer-based bacterial detection.
Keywords:
Detection
; Biosensors
; Aptamers
; Bacterial pathogens
; Food security
1. Introduction
Pathogenic microorganisms have been a relentless challenge to human health and well-being throughout history. They encompass a wide range of bacteria, viruses, fungi, and parasites that can be transmitted through food, water, soil, and air or contact with infected individuals or animals. A survey for pathogens is one of the most important elements in tracing infectious diseases, determining the distribution of agents according to sites, and preventing their spreading. Among bacterial pathogens of particular concern are those resistant to antibacterial drugs commonly used to treat infections caused such as pathogenic strains designed by the acronym ESKAPE (Enterococcus faecium, Staphylococcus aureus, Klebsiella pneumoniae, Acinetobacter baumannii, Pseudomonas aeruginosa, and Enterobacter species). Multidrug-resistant bacterial strains cause the majority of nosocomial and community-acquired infections worldwide [1,2]. In addition, more than 200 identified bacterial pathogens are associated with foodborne illnesses that can occur through either foodborne infection (when ingested bacteria colonize and multiply in the human host) or foodborne intoxication (resulting from consuming food containing bacterial toxins). The most widespread foodborne and waterborne bacteria are Escherichia coli O157:H7, Salmonella spp., Clostridium perfringens, Cronobacter sakazakii, Bacillus cereus, Campylobacter jejuni, S. aureus, Listeria monocytogenes, Shigella spp., Clostridium botulinum, Vibrio spp. and Yersinia enterocoli [3,4]. Some foodborne bacteria, such as L. monocytogenes, pose a significant risk due to their high mortality rates, while others, such as Salmonella and C. jejuni, present high-frequency occurrence. In addition, some pathogenic bacteria are spore-forming (e.g. Clostridium botulinum, Bacillus cereus, Clostridium perfringens, Clostridioides difficile) [5,6,7]. In their sporulated form, bacteria are highly heat-resistant and survive most of the disinfection procedures used by the food industry or hospitals. Also, some bacterial toxins, like staphylococcal enterotoxin B (SEB) and cereulide are resistant to heat and may persist in food products for a long period. In 2019, it was estimated that 7.7 million deaths were caused by pathogenic bacteria [8].
Bacterial detection holds practical importance in the pharmaceutical industry, environmental control, and food safety. Foodborne infections and intoxication resulting from contaminated beverages or food are considered serious problems and have become more prevalent nowadays. In 2015, the World Health Organization (WHO) estimated that there were 600 million cases of foodborne illnesses, representing 11.6% of the world population and resulting in no fewer than 420,000 deaths. However, it is important to note that these data probably do not reflect the full scale of the problem. For instance, foodborne microorganisms can induce severe symptoms, such as putrefactive diarrhea, vomiting, abdominal cramps or fever and cause also debilitating diseases like meningitis in young children [9,10]. In the interest of public health and consumer protection, it is evident that there is a need to concentrate efforts on developing new methods for production control, including new detection techniques.
To reduce risks and control disease progression, diverse methods have been developed for pathogen detection including culture methods and molecular methods such as those based on nucleic acid Amplification (e.g. Polymerase Chain Reaction (PCR)) [11,12,13], and Enzyme-Linked ImmunoSorbent Assay (ELISA) [14]. Although they are the most frequently used methods for pathogen detection, they show many disadvantages to enable efficient health security and sanitary control. Methods based on cultivation are time-consuming, as they require several steps for bacterial enrichment, isolation, and identification. In addition, they can be performed only by qualified personnel and within a laboratory setting. Molecular methods are less time-consuming, but they also require a pre-enrichment step to increase the number of target bacterial cells and to prevent detection of dead bacteria. In addition, molecular methods can give false negative results because they rely on enzymes (DNA-polymerase) or antibodies that can be inhibited by some molecules or ions in food or biological samples [15].
The development and improvement of new diagnostic methods play a pivotal role in managing and mitigating the risks associated with pathogens. Fortunately, many new effective analytical techniques for rapid and sensitive detection of bacterial pathogens directly in complex matrices are under development. In 2003, the WHO established some criteria to ASSURED (Affordable, Sensitive, Specific, User-friendly, Rapid, Equipment-free, Delivered) an access to high-quality diagnostic devices that can be used across all levels of the healthcare system. Recently, two additional criteria, RE (Real-time and ease of specimen collection and environmental friendliness) were added creating the new acronym of REASSURED devices [16]. Biosensors as portable tools providing rapid, sensitive, and efficient detection of pathogens can reach the REASSURED criteria. Biosensors are analytical devices that integrate a bioreceptor, which assures the recognition of the target, to a transducer that converts the recognition event in a measurable signal enabling the detection of chemical or biological analytes [17]. Among recognition elements, aptamers, small artificial single-stranded DNA or RNA molecules (usually 20-100 nucleotides long) have emerged as specific sensing elements.
Aptamers have usually no biological function but exhibit a high affinity for a specific target, similar to the natural interactions observed between antigens and antibodies. Aptamers possess several advantages over expensive antibodies and stand out as strong candidates to replace them. They possess the ability to selectively recognize the desired target with high affinity and specificity, and they can be easily customized to accommodate changes [18,19,20]. Due to such properties, aptamers have gathered signification attention in recent years for their potential to revolutionize various fields, including diagnostics, therapeutics, as illustrated in Figure 1. The utilization of aptamers as recognition elements in biosensors offers several advantages for bacterial detection [21]. The primary advantages of aptamers include their bio- an chemical stability in complex matrices, ease of production due to their small size, and their low immunogenicity [10,22,23]. In addition, these small nucleic acid molecules has a relatively low cost of synthesis compared to antibodies [24]. Nonetheless, their binding affinity is comparable to that of the monoclonal antibodies. Moreover, various chemical modifications of aptamers can enhance both their stability and affinity to bind a specific target [25,26]. Recent years have seen an exponential increase in the development of aptamer-based systems for the monitoring, detection, and identification of pathogenic microorganisms. Figure 2 shows articles published for “aptamers” and “aptamers for bacteria”, along with future trends according to their publication rate in the last twenty years.
In this review, we present the latest advancements in aptamers for bacterial detection. First, we provide a short overview of the strategies for selecting aptamers targeting pathogenic bacteria. Second, we present their kinetics, thermodynamics, and structural characterization. Third, we focus on recent progress (2020–2023) and strategies utilizing aptamers for the detection of bacterial cells, biomarkers, and toxins. Alongside aptasensors for bacterial screening in food and clinical samples, we also highlight some aptamers developed for fundamental microbiological studies. Furthermore, we discuss the principal aspect of aptamers’ optimization. This involves a discussion of methodologies and techniques focusing on both kinetic and structural characterization, as well as chemical modifications ultimately enhancing the overall efficiency and functionality of aptamers for improved bacterial detection. It is noteworthy that some aspects of aptamer applications for bacterial detection, namely aptamer identification, optical and electrochemical transducer technologies used in aptasensors, the nanomaterials for aptasensors, and aptamer selection, have been comprehensively described in some recent reviews [27,28,29,30,31,32]. Finally, barriers and developing perspectives in the application of aptamer-based bacterial detection are analyzed.
2. Aptamer Selection Strategies for Bacterial Pathogen Detection
Aptamers, first developed in the early 1990s by two independent teams, Ellington and Szostak [33] and Tuerk and Gold [34], refer to distinct nucleic acid sequences with selective and specific binding to a target. These sequences are selected from a large pool of nucleic acids, using an in vitro selection technique known as Systematic Evolution of Ligands by Exponential Enrichment (SELEX) [35,36]. SELEX is a series of selection and amplification steps in which a large pool of nucleic acid molecules binds to desired target molecules under specific conditions, including temperature and salt concentration. The molecules that bind to the target are isolated from those that remain unbound, and then they are amplified to generate a newly enriched population. This cycle is repeated until a small population of highly specific molecules is obtained. The SELEX technique has undergone numerous modifications and improvements over time, making it more efficient, reliable, and affordable for the identification of aptamers [37,38]. Conventional SELEX methods have been extensively reviewed [39,40].
In the case of bacterial pathogen detection using aptamers, it can be achieved indirectly, where aptamers are designed against secretory molecules like toxins. A direct approach employs aptamers selected against either a specific molecule of bacteria or the whole-cell bacteria.
Indirect detection of bacteria is important for food, medical, and environmental safety. Indeed, some bacterial toxins can survive the pasteurization process and have a high degree of resistance to high temperatures (<100 °C), making them indicative of the past or current presence of bacteria. For instance, SELEX methods against toxins have been implemented, and aptamers have been discovered to identify toxins of S. aureus, Vibrio cholerae, and C. botulinum, named enterotoxins, cholera toxin, and botulinum toxins, respectively [41,42,43,44,45,46].
Direct detection of bacteria using aptamer selected against surface epitopes such as membrane and cell wall proteins or peptidoglycan has been developed. For instance, RNA aptamers have been reported to target S. aureus teichoic acid [47], a component of the cell wall of the majority of Gram-positive bacteria. Additionally, peptidoglycan-specific aptamers for the detection have also been developed [48]. More recently, aptamers that specifically target isdA, a crucial surface protein for S. aureus survival and colonization, have been successfully generated. Over time, this protein is gradually becoming a key marker for the detection of this bacterium [49,50].
Aptamers can also be selected to directly detect a specific bacterium by using the whole cells or spores as a target. This approach is termed cell-based aptamer selection or Cell-SELEX [51,52]. It was first introduced in 1998 using Human red blood cells by Morris et al. to generate aptamers capable of selectively binding to unidentified molecular biomarkers present on the surface of live cells [53].
Bacterial cell surfaces present a multitude of molecules, particularly proteins, which can serve as potential targets for aptamers. Many aptamers that selectively bind Gram-negative and Gram-positive bacteria such as E. coli [54,55], C. jejuni [56,57], Streptococcus pyogenes [58], S. aureus [59,60,61,62], B. cereus [63], Salmonella Typhimurium [64], or Listeria spp.[65], can be found in the literature. The Cell-SELEX was shown to be effective in generating specific aptamers against a particular bacterium or even a particular clone. Similarly, bacterial spores were used to develop specific aptamers through the Cell-SELEX procedure. In its sporulated form bacteria exhibit higher resistance to various stress factors such as UV radiation and heat compared to the corresponding growing cells [66,67]. The conventional microbiological methods to detect bacterial spores have lengthier procedures compared to those for vegetative cell detection. This is because spores are first subjected to germination, and then their vegetative cells are characterized. Therefore, conventional detection methods such as PCR or immunological assays, are not adapted to fully ensure safety. In contrast, aptamers that bind to the surface of bacterial spores offer a potential for rapid and direct detection. Particularly, aptamers have been developed and used in different aptasensors for the detection of spores from Bacillus species, which are ubiquitously present in air, water, and soil [68,69,70].
The procedure of Cell-SELEX does not require any purification or prior knowledge of a specific targeted surface epitope. Even more significant is that proteins on the cell surface represent a more physiological and natural folding condition than purified proteins. However, the Cell-SELEX might be difficult to perform, because the bacteria cell surface is negatively charged, leading to DNA repulsion from the cell surface [71]. Additionally, the process can introduce selection biases. For instance, dead cells in a suspension can result in a non-specific binding, strongly influencing the selection. Another challenge lies in identifying the specific target(s) of the selected aptamers. Much of the literature using Cell-SELEX lacks a comprehensive understanding of the targets.
Alternative in silico strategies have been developed in the last decades. These methods are based on a computational screening method of a large library of aptamer sequences to fit modeled target structures. These techniques require 3D structure predictions, through docking methodologies using software such as HADDOCK, AutoDock, Xdock, enabling the study of aptamer folding and selection of structural patterns responsible for the aptamer/target interaction based on predicted affinity and Gibbs free energy (∆G) [72,73,74]. The in silico method has been successfully conducted to select aptamers for bacterial surface proteins [75,76,77]. Another example is a recently employed computational screening method to select antimicrobial aptamers against the pathogen Pseudomonas aeruginosa [78]. In silico strategy is a powerful and complementary tool to SELEX, for developing new aptamers against pathogenic bacteria. As a computational-based approach, it offers timesaving and cost-effectiveness compared to SELEX. Nevertheless, in silico methods still require knowledge of aptamers' tridimensional (3D) structure and in vitro testing to confirm the binding interaction between selected aptamers and the targets.
3. Aptamer characterization: affinity, stability, and structure
Aptamers are typically short, single-stranded nucleic acids, either DNA or RNA molecules, that can fold into various 3D structures. Nucleic acids are chemically and structurally stable. Both share similar functionalities and have their advantages: RNA aptamers offer more diverse and complex 3D structures that can enhance binding specificity, and DNA is known for its stability and lower production costs [19]. They can recognize and bind their target with high specificity and affinity in the nano- and picomolar range. The structural conformation of an aptamer depends not only on its base composition but also on its environment including pH and salt type, ionic force. Those parameters have a significant influence on the formation and stability of specific conformations of aptamers, they are particularly true for intercalated motif (I-motif) or G-quadruplex structures [79,80,81,82].
The initial pool of nucleic acids used in SELEX contains a random nucleotide region flanked by two constant regions, the primers, that serve for the amplification in the SELEX process to enrich the sequences that bind to the target molecule. The primers are usually considered nonessential nucleotides and are not involved in the folding process, nor are they used for recognizing the target [83]. One study tried to explore the influence of the random-region length on aptamers, but they found no significant correlation between length and aptamer affinity [84]. However, a longer random region enhances the complexity of secondary and tertiary structures, while a shorter one provides better coverage and gives production advantages. [85]. One or a few sequences are identified at the end of the SELEX process. It is then necessary to determine its kinetics parameters, such as binding affinity (Kd).
Different methods have been used to evaluate the Kd, relying on either label-based and label-free techniques [86]. The label-based approaches, fluorescent or radioactive labels, lower the detection limit in the experiments, resulting in a more accurate determination of the Kd. Label-free methods, such as surface-based detection methods, are often used. One of the most commonly used methods is Surface plasmon resonance (SPR), an optical method to detect molecular binding interactions between one mobile and one fixed molecule. SPR is used for evaluating different kinetic parameters, such as the association and dissociation rate constant and the dissociation/association constant [87].
Those traditional methods are adapted to evaluate the binding affinity for an aptamer when the target is known, such as proteins or toxins that can be isolated or produced in a purified form. However, in the cell-based SELEX method, the target(s) for the aptamer is unknown. This, in turn, complicates the measurement of kinetics parameters because the number of targets and their concentration are unknown. This uncertainty makes it challenging to measure binding affinity accurately. On top of that, working with whole cells is technically challenging and requires complex protocols in terms of detection sensitivity and apparatus systems, such as microfluidics. Several studies have estimated the Kd using Fluorescence-Activated Cell Sorting (FACS) methods [49,88,89,90]. FACS offers the advantage of comparing different aptamer candidates for a whole cell biomarker. Although assessing the kinetics parameters is essential, the aptamer’s stability plays a crucial role in determining its overall suitability. Comprehensive descriptions of aptamers, including details on thermal stability, topology, and structure, are essential for their effective utilization in various applications and for ensuring their effectiveness in real-life scenarios.
The stability of nucleic acid structures can be evaluated through thermal denaturation experiments using either UV-CD absorption techniques or fluorescence techniques such as FRET [91,92,93]. Isothermal titration calorimetry (ITC) is used to derive thermodynamic parameters and stoichiometry of the aptamer/target interactions. Several factors can influence the melting temperature of an aptamer, affecting its stability, such as the base composition, length, structures, ionic strength, and pH [94,95,96,97,98]. Moreover, the potential conformational change of aptamers upon binding with their targets are rarely explored and deserve better investigations, [99]. Resolving the high-resolution or even the topology of an aptamer could lead to a better understanding of target specificity and pave the way for further optimization of stability and specificity. It was shown that aptamers could adopt various tertiary structures that can be classified into three categories: Structures containing duplexes, loops, and bulges (e.g. Hairpins, pseudoknots, triplex), G-quadruplexes, and I-motif (Figure 3). The aptamer structure may include loops and bulge regions (Figure 3). They play a crucial role in their structure and function, influencing their binding affinity and specificity [100,101,102].
Structures containing a duplex stem with different loops and bulges are very diverse (loop length number and composition can vary), ranging from simple hairpins to pseudoknots (Figure 3A), all resulting in a spectrum of distinct conformations that can be used to bind a target selectively. G-quadruplexes are four-stranded non-canonical nucleic acid structures that can form under physiological conditions [103]. G-quadruplex structures are stacked G-tetrads formed by four hydrogen-bonded guanines stabilized by cations (Figure 3B). G-quadruplex structures are diverse in terms of strand orientations, loop length, and composition and have high structural variability, including interesting hybrid duplex-quadruplex structures [104]. They have shown promise in the bacterial detection [105]. The intercalated motif (I-motif) is a structure involving a hemiprotonated C-C+ base pair stabilized under acidic pH conditions [106] (Figure 3C). To our knowledge, no aptamer has been shown to form an I-motif, and none have been developed in the bacterial detection context. This may be explained by the lack of structural studies on the aptamers field, although several aptamers have C-rich sequences [61]. Aptamers forming an I-motif for bacterial detection are an area of potential future research. Prediction software has been overwhelmingly used to predict the secondary structure of an aptamer. However, those predictions consider simple rules of base pairing and free energy and fail to predict other secondary structures such as G-quadruplex or I-motif [107,108,109].
To have insight into the structure adopted by an aptamer, experimental data are required. Different experimental methods can be used to gain insight into the folding of aptamers. Shape-type experiments, which chemically probe unpaired or paired base pairs are used to determine the secondary structure of nucleic acids. These experiments are convenient for long sequences. Spectroscopic methods such as UV, circular dichroism, or fluorescence can be used to obtain information on the folding topology of nucleic acid [110]. For example, CD is widely used for G-quadruplex or I-motif to gain insights into the G-quadruplex topology [111]. Those relatively easy methods can be used to look at the dependence of the secondary structure, such as the ion type and concentration, pH, and buffer composition. This is particularly true for I-motif [112,113], and G-quadruplexes [114,115]. This information is crucial for establishing a protocol that ensures the correct folding and the monitoring of their stability over time. Three main experimental techniques have been used to resolve the structure of aptamers free or in complex with their target: Nuclear Magnetic Resonance (NMR), X-ray crystallography, and Cryogenic electron microscopy (cryo-EM). All the methods can give rise to high-resolution structures (see [116] for a comprehensive analysis of aptamers structure). NMR uniquely investigates nucleic acid conformational states and folding through simple 1D experiments. It monitors structures like I-motifs, G-quadruplexes, or hairpins, specifically through the imino-proton chemical shift [117]. Those methods provide detailed information and are becoming crucial for understanding the aptamer-binding mode. One notable example is an RNA aptamer's three-dimensional resolution that can recognize the bacterial protein Hfq of Bacilus subitilis [118]. Recently, there has been a growing interest in solving these aptamers' complex structures [116,119,120].
Aptamers' chemical modifications and optimizations are increasingly explored, serving as a crucial strategy to enhance binding affinity, thermal stability, and protection against nucleases. These properties directly influence the overall suitability of aptamers for biosensing applications. One significant disadvantage of aptamers as oligonucleotides is their high sensitivity to nuclease degradation, which affects detection in biological samples, especially in serum [121]. Temperature fluctuations can also impact aptamer binding capacity and susceptibility to enzymatic degradation by nucleases. To overcome these limitations, various chemical strategies have been devised to thermally stabilize, protect against degradation, and enhance the binding affinity of the active conformation of an aptamer.
Chemical modifications to the base, phosphate group, or sugar unit have been used to enhance the aptamer suitability [122,123,124]. For instance, incorporation of non-natural phosphorothioate into the aptamer backbone structure or modification of the 5’-end or sugar structure of strands were shown to slow down the hydrolysis rate by nucleases present in biological fluids while retaining their activity [125,126,127,128]. Another approach to avoiding the problems related to aptamer degradation is by incorporation of unnatural base pairs (such as 5-methyl-isocytidine) or utilization of L-DNA to make strands highly resistant to nuclease attacks [129]. The majority of such chemical modifications aim to improve the stability of nucleic acid structures that are highly vulnerable to nuclease digestion, particularly in single-stranded segments and internal nicks.
Aptamer can acquire a relative nuclease resistance when covalently linked to a protective group such as protein. For instance, DNA-type thrombin binding aptamer was shown to resist a variety of serum nucleases when it was tightly bound to its target protein [130]. Moreover, different protective coatings of DNA strands increase their nuclease resistance. Lacroix et al increased stability of the DNA strand by its coating with human serum albumin and DNA dendrites [131]. In another study, an oligolysine–PEG (polyethylene glycol) coating was shown to protect DNA structure from low-salt denaturation and nuclease degradation [132]. The stability can also be improved by ligation of strand termini or by the introduction of terminal functional groups, such as hexanediol or hexaethylene glycol, crosslinking of nucleic strands by click chemistry or through the formation of a thymidine dimer using ultraviolet irradiation [133,134]. These chemical modifications eliminate internal nicks once they are constructed.
In a different approach, El-Khoury and Damha used a chemical end-ligation to stabilize an intramolecular I-motif at both acidic et neutral pH using deoxy-2-fluoroarabinocytidine [135]. This method allows for the use of I-motif on a large scale for in vitro applications such as detection.
Finally, the effect of nucleases on the aptamer structure can also be minimized by applying conditions to denature nucleases or by using nuclease inhibitors. For instance, the detection in serum can be performed after serum heat pretreatment at 75°C for 10 min, thus rendering nucleases inactive [136].
4. Bacterial aptasensors
To advance the development of diagnostic tests to detect pathogenic bacteria, significant focus has been directed toward biosensors paired with aptamers, named aptasensors. They are devices that incorporate one or several aptamers and a signal transducer that can be electrochemical, optical, or mass-sensitive. Infectious bacterial agents differ in numerous aspects, comprising their virulence factors, contagion, and transmission. All these factors and molecules playing a role in the bacterial life cycle can be targeted by aptamers to enable specific and efficient bacterial detection and identification using aptasensors. Many innovative aptasensors have been developed recently and can be divided into aptasensors targeting whole bacterial cells, bacterial toxins, and specific biomarkers. Table 1 summarizes recently reported aptamer-based biosensors with various detection methods and their characteristics. It illustrates the remarkable performances of aptasensors for bacterial detection.
4.1. Detection of bacterial cell
There are significant practical advantages in developing aptasensors for detection of whole bacterial cells. The diversity of biomarkers and their abundance on a cell surface provide the potential for higher sensitivity than targeting a single protein expressed at a low level. In addition, targeting whole cells facilitates the overall detection procedure since the sample preparation prior to analysis is highly simplified [164]. In most traditional and molecular methods for bacterial detection, sample preparation is the most expensive and the most time-consuming step. Finally, the selection of specific aptamers is also facilitated because there is no need for the lengthy process of purifying a specific target biomarker [165]. However, bacterial surfaces can vary in structure and composition depending on the environmental condition. This bacterial adaptation to external conditions may modify their surface molecule composition, which influences the reliability of diagnostic tests. This problem can be overcome by the utilization of a mixture of aptamers that target different surface epitopes of the target bacterium [166,167].
One recent development of bacterial electrochemical aptasensor was designed for selective and ultrasensitive detection of S. aureus at the single-cell level using a sandwich assay [168]. The aptasensor relied on dual recognition through a bacteria-imprinted polymer film (BIF) and an aptamer (Figure 4). The BIF, which served as a capture probe, was fabricated on a glass carbon electrode, whereas the aptamer, which served as a detection probe, was functionalized with 6-Ferrocenyl-hexanediol on gold nanoparticles (AuNPs). Aptamer was produced with a thiol on its 5’ to allow its covalent binding to AuNPs. After anchoring S. aureus on the BIF-modified electrode, AuNPs-aptamers were introduced to bind the captured bacteria, leading to an amplified current signal. Obtained ultrasensitive detection enabled sensing of a single S. aureus cell in a buffer. The limit of detection (LoD) of the aptasensor was 10 CFU/mL when used in complex, lipid-rich media like milk. This high sensitivity suggests the sensor's applicability in food safety and prevention.
In another recent example, Ding et al. [169] developed a novel ratiometric dual-signal electrochemical aptasensor for pathogenic bacteria at the single-cell level. This sensor was based on the aptamer recognition-induced Rolling Circle Amplification/G-quadruplex strategy [170,171,172]. G-quadruplex can interact with various signaling molecules, including electrochemical signal redox indicator methylene blue (MB). In this study, the signal amplification was achieved through the utilization of two probes—one being an aptamer modified with ferrocene for the target bacteria and the other being the primer sequence for rolling circle amplification (RCA) anchored to the gold electrode with a sulfhydryl probe (Figure 5). These two probes can form a complex disrupted by the presence of the target bacteria S. aureus. Consequently, when contaminated samples were tested, only the RCA probe remained on the gold surface. To stimulate the RCA reaction, a C-base-rich circular template was introduced to the reaction mixture, generating a multitude of G-base-rich nucleic acid sequences that formed RCA products into a G-quadruplex. Upon the addition of MB, there was a substantial cascade amplification of electrochemical signals due to its strong affinity of the G-quadruplex [173].
Bacterial spores remain the major concern in food industry and hospitals since they are remarkably resistance to high temperatures, chemical agents, radiation, and harsh physical conditions. In sporulated form bacteria exhibit resilience to unfavorable environmental conditions, making them the most resistant life forms known to date. Typically, they can withstand temperatures 40-45 °C higher than their corresponding vegetative cells. Additionally, spores can enter dormancy and survive in a wet state for thousands of years. Furthermore, spores are regarded as potential biological weapons. Indeed, Bacillus anthracis was initially used during the First World War. The dispersal of spores in the ambient air can result in the development of the respiratory form of anthrax. Due to these unique characteristics, for biosecurity and food safety, researchers have developed aptasensors specifically designed for the detection of target bacteria in their sporulated form [5,68,174]. Recently, Rizzotto et al. developed a colorimetric assay for the detection of Bacillus cytotoxicus spores in food [29]. B. cytotoxicus belongs to a large group of bacteria commonly named the Bacillus cereus group [6,167]. The group is constituted of different species some of them are human pathogen (such as B. anthracis, B. cereus sensu stricto or B. cytotoxicus) but others are harmless (such as B. thuringiensis, B. weihenstephanensis and B. mycoides). The detection test relied on the spore-enhanced peroxidase-like catalytic activity of gold nanoparticles (Figure 6). The test was performed in a simple microtube containing AuNPs and magnetic beads (MP) both conjugated with a specific aptamer BAS6R as a recognition element for B. cytotoxicus spores. The presence of spores was detected with the naked eye due to the change in color in the solution (blue to dark blue) upon oxidation of tetramethylbenzidine (TMB) with H2O2. This method has the advantage of the aptamer binding to an unknown epitope on the spore surface, eliminating the need to extract, purify, and amplify a specific biomarker. However, BAS6R was shown to bind to spores of different B. cereus species. It cannot thus distinguish between pathogenic and non-pathogenic strains of this group of bacteria.
In an another study, Zhou et al, developed a dual-aptamer microfluidic chip for detection of both B. cereus vegetative cells and spores [156]. To improve the selectivity and sensitivity of detection, aptamers selected through the Cell-SELEX were additionally, step-by-step, tailored and optimized using molecular docking. Based on this dumbbell aptamer design, two best aptamers were obtained and integrated into a microfluidic chip biosensor. These two aptamers had repeated G-based both with a high affinity to bind the polar amino-acids in the α-helix of epiprotein of B. cereus. The biosensor applied directly in food, such as milk and rice, provided a complete analysis within 1 hour and showed a LoD of only 9.27 CFU/mL.
4.2. Bacterial toxin detection
Detecting bacteria is not always sufficient. Many bacterial pathogens such as S. aureus, E. coli, C. botulinum, C. perfringens, C. difficile, B. cereus, C. jejuni, L. monocytogenes, Yersinia enterocolitica, and Salmonella spp. produce toxins [175,176,177,178,179,180,181,182,183]. Ingested bacteria toxins cause various effects, ranging from disrupting cell membranes and causing cell death to interfering with the host immune response. They play a crucial role in the pathogenicity of many bacterial species, contributing to the symptoms and severity of bacterial infections [184]. For instance, C. botulinum toxins can cause botulism, a severe illness characterized by muscle paralysis, general weakness and potentially fatal respiratory failure. S. aureus enterotoxins in food can lead to symptoms such as nausea, vomiting, huge diarrhea. As already mentioned, these toxins are thermostable and can hardly be eliminated by food treatment or preparation like cooking, frying or freezing that eliminate bacterial cells. Moreover, toxins can go undetected during classic method pathogen detection. Usually when strains are isolated, the presence of toxins is tested by PCR amplification of genes that encode them. This additionally step prolongate the procedure and cannot be performed in the field. In addition, the presence of gene does not necessary yield to the toxin production. There is, thus, a need to directly detect the presence of bacterial toxins in food and biological samples.
An electrochemical aptasensors was designed for the detection of staphylococcal enterotoxin B (SEB), [100] a potent bacterial toxin responsible for severe food poisoning [185,186]. The sensor utilized a commercial carbon screen-printed electrode modified with reduce graphene oxide (rGO) and gold nano-urchins (AuNUs). The electrode was further functionalized with a single-stranded DNA probe, followed by the attachment of a specific aptamer. Upon encountering SEB molecules, the aptamer detached from the electrode surface. Following this, an electrochemical signal of the redox probe, hematoxylin, decreased significantly (Figure 7). The aptasensor analytical performance was validated with food samples, serving as simulated real samples. This tool surpasses a commercial ELISA kit for SEB detection. By employing this method, a promising application of the aptasensor was demonstrated for food sample screening.
To assess food safety, a multiplex aptasensors were developed for the detection of different targets in a food sample. Jin and al. proposed a lateral-flow strips (LFT) aptasensor for Salmonella, Hg2+-ions and ochratoxin A (OTA) using an upconversion nanoparticles (UCNP) as a signal source and a smartphone equipped with a CCD camera for the detection [187]. This approach ensures a fast and portable detection process. As illustrated in Figure 8 three different aptamers were used in the sensor, each grafted on the surface of different color UCNPs and hybridized with their complementary DNA (cDNA) fixed on the nitrocellulose membrane. In the presence of bacteria, ions, and small molecules, the three aptamers preferentially bond to their respective targets. Consequently, the liberated cDNA decreases the fluorescence from the UCNP. It was demonstrated that this device had no non-specific or cross-reactivity due to the separate color channels providing detection signal. This feature was crucial to enable multiplexing. The method of detection holds promising prospects for the simultaneous detection of multiple targets in food samples for enhanced security.
4.3. Bacterial biomarker detection
A bacterial biological marker or bacterial biomarker can be defined as a specific characteristic that is measured as an indicator of bacterial presence, their biological processes, or responses to an external exposure or intervention, including therapeutic interventions. Detection of biomarkers enables various applications, such as monitoring of infection and therapy efficiency, risk estimation, diagnosis, or estimation of bacterial adaptation to environmental signals. Numerous aptamers have been developed to target a wide array of bacterial biomarkers although targeting the entire cell is more common. Some aptamers have been developed for purposes other than detection, such as therapeutic or basic research applications. This technology opened the way for innovative possibilities that are not only cost-effective but also capable of overcoming the limitations posed by conventional antibodies or various probes and drugs.
The methyl erythritol phosphate (MEP) metabolic pathway of isoprenoid biosynthesis is essential for many pathogenic bacteria, both gram-positive and gram-negative, such as B. anthracis, Clostridium spp., L. monocytogenes, S. enterica, Vibrio cholerae, Shigella spp, E. coli and Pseudomonas aeruginosa. Recently a DNA aptamer, D10, was develop against 1-deoxy-D-xylulose-5-phosphate reductoisomerase (DXR), the second enzyme of MEP pathway [188]. This aptamer modified with a fluorescent label allowed a strong specific staining of MEP+ bacterial cells (E. coli and P. aeruginosa) while no staining was observed with bacterial lacking the DXR enzyme (e.g., E. faecalis). It presents, thus, a potential to replace the specific antibody for future therapeutic and diagnostic applications.
More recently, an antagonist aptamer for SEB neutralization was selected through the SELEX [189]. SEB is a critical virulence factor in staphylococcal toxic shock syndrome (TSS), triggering a robust release of proinflammatory cytokines by activating T lymphocytes. The aptamer demonstrated effectiveness in inhibiting SEB-induced proliferation and cytokine secretion in human peripheral blood mononuclear cells. Their results highlight the potential therapeutic value of the novel aptamer antagonist in mitigating SEB-mediated TSS, and presenting a promising avenue for future therapeutic strategies.
Another expanding aptamer application was described by Stoltenburg and his co-workers [105], who explored an alternative approach develop aptamers that bind a bacterial surface protein. In contrast to the Cell-SELEX which selects specific aptamers that bind unknow target at the bacterial cell surface, they procedure was based on selecting aptamers that recognize a purified surface protein of S. aureus, called protein A. This protein A is a major component of the cell wall of S. aureus. Obtained selected protein-A binding G-quadruplex aptamer was integrated into Enzyme-linked OligoNucleotide Assay (ELONA). They demonstrated that the aptamer and its truncated versions bound to the Protein A expressed by intact cells of S. aureus. Some structural information of the aptamer used was also gathered because the aptamer functionality was shown to be tightly linked to their structure. Such insights of the structural basis of the aptamer functionality enhance our understanding of the molecular bases of interactions and pave the way for the development of novel more robust aptamer-based diagnostic tools.
Many bacterial virulence factors are exported by external vesicles. All Gram-negative bacteria secrete outer membrane vesicles (OMV)s, proteoliposomes derived from their outer membrane with sizes between 20 and 250 nm. OMVs play crucial roles during host-microbe interactions because they transport a broad range of molecularly diverse cargoes, including adhesins, toxins and membrane embedded and associated proteins, small molecules, lipids, peptidoglycan and nucleic acids. Shin et al, have selected aptamers that bind OMVs of multiple Gram-negative bacteria [147]. The aptamers selected through the Cell-SELEX showing the highest affinity to bind Gram-negative bacterial cells were conjugated with horseradish peroxidase (HRP) and used in an enzyme-linked aptamer essay (ELAA) to detect OMVs. A LoD of 25 ng/mL was obtained. The recognition of OMVs was expected because secreted vesicles contain similar proteolipids as their cell of origin. They suggested that detection of bacterial OMVs was more effective then detection of bacterial cells in clinical samples.
5. Conclusions
In the realm of pathogenic bacteria, biosensors for their rapid and accurate identification and monitoring are important added-value in preventing spreading, reducing treatment expenses and ensuring food safety. Biosensors offer numerous advantages compared to traditional microbiological methods for their enhanced sensitivity and rapidity. On the most significant challenges in bacterial pathogen control is the time required for the detection and identification of particular agents. Currently, identification of a microorganism typically takes between one and several days, when plating or PCR based protocols are employed. In contrast, biosensors may provide a complete microbiological analysis within several hours or even minutes [190,191]. Biosensors also hold the potential to be portable, multiplex and accessible to a wide range of users [192,193]. Biosensors are thus the most promising tool for efficient microbial detection and their point-of-need testing [194].
Aptamers have been used extensively in recent years as an alternative to conventional antibodies in various biosensing strategies including electrochemical, optical and microfluidic chip-based platforms. Compared to antibodies, aptamers have a shorter production time, lower cost, higher stability in complex matrices, and possess ability to bind a variety of specific targets which enable their tailored applications. As outlined here, various aptamers are available for different bacterial targets, including whole cells, toxins, intracellular and secreted biomarkers. Regarding the latest research, trends in aptasenors for bacterial pathogens detection include development of portable biosensors, such as electrochemical or paper-strip based devices coupled to easy or no sample preparation and high cost-affordability biosensing strategies. The specificity of detection and accuracy of overall analysis may be improved by using dual aptamer probes, where the first aptamer is capturing probe and second is detection probe. This configuration allows the use aptamers with different chemical modifications i.e., the capturing probe may carry a thiol for its covalent immobilization onto the gold surface of the biosensor, while the detection probe may be conjugated with a fluorescent or redox probe to enable signal read-out. Moreover, a mix of various aptamers, each targeting different epitopes of the same bacterium can be employed to achieve higher accuracy of detection.
Signal amplification strategies employ aptasensors coupled with different nanomaterials, catalysis amplification, hybridization chain reaction, and rolling circle amplification [30]. Alternatively, aptamers adopting specific structural 3D conformations, such as G-quadruplex, are shown to provide high signal intensities. The utilization of aptamers of specific 3D structures as recognition elements seems to be a smart and cost-effective approach to develop sensitive biosensors [195]. In the same time, aptamers structural changes upon target binding may induce a signal transduction and/or result in target triggered amplification, which highly simplify the device development [196]. Interestingly, although primary sequence of many validated and published aptamers suggests that they adopt a specific 3D structure (G-rich or C-rich sequences), their structure was usually not investigated. It is especially case for aptamers adopting I-motive (C-rich sequence) [61].
6. Future directions
Aptasensors still have to cross a long way to become a standard technique for microbiological applications. Indeed, there is a significant number of papers presenting proof-of-concept, but still, there are only a few commercially available aptasensors that can be utilized at the level of the food industry or hospitals. First, aptamer technology has some shortcomings. Most SELEX procedures are performed using the same oligonucleotide pool, and not all selected aptamers are very selective. Second, the selection and structural characterization of aptamers are usually performed under defined conditions, but their configuration and binding properties can be modified upon temperature, ionic straight or pH variations, resulting in a significant reduction of binding affinities. Consequently, although some aptasensors provide efficient bacterial detection in the buffer, they are inefficient when applied in biological or food samples. Still, aptamer active conformation can be stabilized through chemical modifications. However, there are only few reports where such optimization was performed. Third, aptamers as oligonucleotides can be degraded in biological matrices containing DNAse or RNAse enzymes. Again, chemical modification of aptamer molecules can be performed to prevent such degradations, but are rarely done. The future trend in aptamer selection, characterization and structural optimization will require closer collaboration between molecular biophysicists, microbiologist and biosensor developers.
Author Contributions
All authors contributed equally.
Funding
This work was supported in part by the French National Agency for Research (ANR-21-CE21-009 “Siena”). V.L. was supported by a ‘DIM1Health’ doctoral fellowship awarded by the Conseil Régional d’Ile-de- France.
Conflicts of Interest
The authors declare no conflict of interest.
References
- J. C. de M Campos, L. C. Antunes, and R. B. Ferreira, “Global priority pathogens: virulence, antimicrobial resistance and prospective treatment options,” Future Microbiology, vol. 15, no. 8, pp. 649–677, May 2020. [CrossRef]
- “WHO. 2022.” [Online]. Available: https://cdn.who.int/media/docs/default-source/gho-documents/world-health-statistic-reports/worldhealthstatistics_2022.pdf.
- P. S. Mead et al., “Food-Related Illness and Death in the United States,” Emerging Infectious Diseases, vol. 5, no. 5, pp. 607–625, 1999. [CrossRef]
- M. Marin, M. V. Nikolic, and J. Vidic, “Rapid point-of-need detection of bacteria and their toxins in food using gold nanoparticules,” Comp Rev Food Sci Food Saf., vol. 20, no. 6, pp. 5880–5900, Oct. 2021. [CrossRef]
- J. Vidic, C. Chaix, M. Manzano, and M. Heyndrickx, “Food Sensing: Detection of Bacillus cereus Spores in Dairy Products,” Biosensors, vol. 10, no. 3, p. 15, Feb. 2020. [CrossRef]
- N. Ramarao, S.-L. Tran, M. Marin, and J. Vidic, “Advanced Methods for Detection of Bacillus cereus and Its Pathogenic Factors,” Sensors, vol. 20, no. 9, p. 2667, May 2020. [CrossRef]
- P. Setlow and E. A. Johnson, “Spores and Their Significance,” in Food Microbiology, M. P. Doyle, F. Diez-Gonzalez, and C. Hill, Eds., Washington, DC, USA: ASM Press, 2019, pp. 23–63. [CrossRef]
- K. S. Ikuta et al., “Global mortality associated with 33 bacterial pathogens in 2019: a systematic analysis for the Global Burden of Disease Study 2019,” The Lancet, vol. 400, no. 10369, pp. 2221–2248, Dec. 2022. [CrossRef]
- E. J. Scallan Walter, H. Q. McLean, and P. M. Griffin, “Hospital Discharge Data Underascertain Enteric Bacterial Infections Among Children,” Foodborne Pathogens and Disease, vol. 17, no. 9, pp. 530–532, Sep. 2020. [CrossRef]
- R. Chowdary Akkina, V. Payala, and S. Sushma Maganti, “Tools for Rapid Detection and Control of Foodborne Microbial Pathogens,” in Foodborne Pathogens - Recent Advances in Control and Detection, A. Lamas, C. Manuel Franco, and P. Regal, Eds., IntechOpen, 2023. [CrossRef]
- K. Mullis, F. Faloona, S. Scharf, R. Saiki, G. Horn, and H. Erlich, “Specific Enzymatic Amplification of DNA In Vitro: The Polymerase Chain Reaction,” Cold Spring Harbor Symposia on Quantitative Biology, vol. 51, no. 0, pp. 263–273, Jan. 1986. [CrossRef]
- B. Malorny, P. T. Tassios, P. Rådström, N. Cook, M. Wagner, and J. Hoorfar, “Standardization of diagnostic PCR for the detection of foodborne pathogens,” International Journal of Food Microbiology, vol. 83, no. 1, pp. 39–48, May 2003. [CrossRef]
- S. Toze, “PCR and the detection of microbial pathogens in water and wastewater,” Water Research, vol. 33, no. 17, pp. 3545–3556, Dec. 1999. [CrossRef]
- J. E. Butler, “Enzyme-Linked Immunosorbent Assay,” Journal of Immunoassay, vol. 21, no. 2–3, pp. 165–209, May 2000. [CrossRef]
- A. Klancnik, M. Kovac, N. Toplak, S. Piskernik, and B. Jersek, “PCR in Food Analysis,” in Polymerase Chain Reaction, P. Hernandez-Rodriguez, Ed., InTech, 2012. [CrossRef]
- K. J. Land, D. I. Boeras, X.-S. Chen, A. R. Ramsay, and R. W. Peeling, “REASSURED diagnostics to inform disease control strategies, strengthen health systems and improve patient outcomes,” Nat Microbiol, vol. 4, no. 1, pp. 46–54, Dec. 2018. [CrossRef]
- D. R. Thévenot, K. Toth, R. A. Durst, and G. S. Wilson, “ELECTROCHEMICAL BIOSENSORS: RECOMMENDED DEFINITIONS AND CLASSIFICATION *,” Analytical Letters, vol. 34, no. 5, pp. 635–659, Mar. 2001. [CrossRef]
- K.-Y. Wang, Y.-L. Zeng, X.-Y. Yang, W.-B. Li, and X.-P. Lan, “Utility of aptamer-fluorescence in situ hybridization for rapid detection of Pseudomonas aeruginosa,” Eur J Clin Microbiol Infect Dis, vol. 30, no. 2, pp. 273–278, Feb. 2011. [CrossRef]
- J. Zhou and J. Rossi, “Aptamers as targeted therapeutics: current potential and challenges,” Nat Rev Drug Discov, vol. 16, no. 3, pp. 181–202, Mar. 2017. [CrossRef]
- S. Y. Toh, M. Citartan, S. C. B. Gopinath, and T.-H. Tang, “Aptamers as a replacement for antibodies in enzyme-linked immunosorbent assay,” Biosensors and Bioelectronics, vol. 64, pp. 392–403, Feb. 2015. [CrossRef]
- E. Dausse, S. Da Rocha Gomes, and J.-J. Toulmé, “Aptamers: a new class of oligonucleotides in the drug discovery pipeline?,” Current Opinion in Pharmacology, vol. 9, no. 5, pp. 602–607, Oct. 2009. [CrossRef]
- T. Wandtke, E. Wędrowska, M. Szczur, G. Przybylski, M. Libura, and P. Kopiński, “Aptamers—Diagnostic and Therapeutic Solution in SARS-CoV-2,” IJMS, vol. 23, no. 3, p. 1412, Jan. 2022. [CrossRef]
- T. Wandtke, J. Woźniak, and P. Kopiński, “Aptamers in Diagnostics and Treatment of Viral Infections,” Viruses, vol. 7, no. 2, pp. 751–780, Feb. 2015. [CrossRef]
- T. Wang, C. Chen, L. M. Larcher, R. A. Barrero, and R. N. Veedu, “Three decades of nucleic acid aptamer technologies: Lessons learned, progress and opportunities on aptamer development,” Biotechnology Advances, vol. 37, no. 1, pp. 28–50, Jan. 2019. [CrossRef]
- P. Röthlisberger and M. Hollenstein, “Aptamer chemistry,” Advanced Drug Delivery Reviews, vol. 134, pp. 3–21, Sep. 2018. [CrossRef]
- K. Y. Chan, A. B. Kinghorn, M. Hollenstein, and J. A. Tanner, “Chemical Modifications for a Next Generation of Nucleic Acid Aptamers,” ChemBioChem, vol. 23, no. 15, p. e202200006, Aug. 2022. [CrossRef]
- N. E. Trunzo and K. L. Hong, “Recent Progress in the Identification of Aptamers Against Bacterial Origins and Their Diagnostic Applications,” IJMS, vol. 21, no. 14, p. 5074, Jul. 2020. [CrossRef]
- J. Yi et al., “The research of aptamer biosensor technologies for detection of microorganism,” Appl Microbiol Biotechnol, vol. 104, no. 23, pp. 9877–9890, Dec. 2020. [CrossRef]
- F. Rizzotto, M. Marin, C. Péchoux, S. Auger, and J. Vidic, “Colorimetric aptasensor for detection of Bacillus cytotoxicus spores in milk and ready-to-use food,” Heliyon, vol. 9, no. 7, p. e17562, Jul. 2023. [CrossRef]
- I. Bobrinetskiy et al., “Advances in Nanomaterials-Based Electrochemical Biosensors for Foodborne Pathogen Detection,” Nanomaterials, vol. 11, no. 10, p. 2700, Oct. 2021. [CrossRef]
- S. Sharifi et al., “Detection of pathogenic bacteria via nanomaterials-modified aptasensors,” Biosensors and Bioelectronics, vol. 150, p. 111933, Feb. 2020. [CrossRef]
- M. Liu, F. Yue, Q. Kong, Z. Liu, Y. Guo, and X. Sun, “Aptamers against Pathogenic Bacteria: Selection Strategies and Apta-assay/Aptasensor Application for Food Safety,” Agricultural and food chemistry, vol. 70, no. 18, pp. 5477–5498, May 2022. [CrossRef]
- Andrew D. Ellington and Jack W. Szostak, “In vitro selection of RNA molecules that bind specific ligands,” Nature, vol. 346, no. 6287, pp. 818–822, Aug. 1990. [CrossRef]
- C. Tuerk and L. Gold, “Systematic Evolution of Ligands by Exponential Enrichment: RNA Ligands to Bacteriophage T4 DNA Polymerase,” Science, New Series, vol. 249, no. 4968, pp. 505–510, 1990. [CrossRef]
- T. Sampson, “Aptamers and SELEX: the technology,” World Patent Information, vol. 25, no. 2, pp. 123–129, Jun. 2003. [CrossRef]
- E. Dausse et al., “Aptamer selection by direct microfluidic recovery and surface plasmon resonance evaluation,” Biosensors and Bioelectronics, vol. 80, pp. 418–425, Jun. 2016. [CrossRef]
- N. Komarova and A. Kuznetsov, “Inside the Black Box: What Makes SELEX Better?,” Molecules, vol. 24, no. 19, p. 3598, Oct. 2019. [CrossRef]
- M. Kohlberger and G. Gadermaier, “SELEX: Critical factors and optimization strategies for successful aptamer selection,” Biotech and App Biochem, vol. 69, no. 5, pp. 1771–1792, Oct. 2022. [CrossRef]
- C. Zhu et al., “Recent progress of SELEX methods for screening nucleic acid aptamers,” Talanta, vol. 266, p. 124998, Jan. 2024. [CrossRef]
- K. Wang, M. Wang, T. Ma, W. Li, and H. Zhang, “Review on the Selection of Aptamers and Application in Paper-Based Sensors,” Biosensors, vol. 13, no. 1, p. 39, Dec. 2022. [CrossRef]
- B. Mondal, S. Ramlal, P. S. Lavu, B. N, and J. Kingston, “Highly Sensitive Colorimetric Biosensor for Staphylococcal Enterotoxin B by a Label-Free Aptamer and Gold Nanoparticles,” Front. Microbiol., vol. 9, p. 179, Feb. 2018. [CrossRef]
- Y. Huang et al., “A multicolor time-resolved fluorescence aptasensor for the simultaneous detection of multiplex Staphylococcus aureus enterotoxins in the milk,” Biosensors and Bioelectronics, vol. 74, pp. 170–176, Dec. 2015. [CrossRef]
- E. Frohnmeyer, F. Frisch, S. Falke, C. Betzel, and M. Fischer, “Highly affine and selective aptamers against cholera toxin as capture elements in magnetic bead-based sandwich ELAA,” Journal of Biotechnology, vol. 269, pp. 35–42, Mar. 2018. [CrossRef]
- J. G. Bruno, A. M. Richarte, M. P. Carrillo, and A. Edge, “An aptamer beacon responsive to botulinum toxins,” Biosensors and Bioelectronics, vol. 31, no. 1, pp. 240–243, Jan. 2012. [CrossRef]
- A. Subekin, R. Alieva, V. Kukushkin, I. Oleynikov, and E. Zavyalova, “Rapid SERS Detection of Botulinum Neurotoxin Type A,” Nanomaterials, vol. 13, no. 18, p. 2531, Sep. 2023. [CrossRef]
- N. A. Molejon et al., “Selection of G-rich ssDNA aptamers for the detection of enterotoxins of the cholera toxin family,” Analytical Biochemistry, vol. 669, p. 115118, May 2023. [CrossRef]
- S. R. Han and S.-W. Lee, “In vitro selection of RNA aptamer specific to Staphylococcus aureus,” Ann Microbiol, vol. 64, no. 2, pp. 883–885, Jun. 2014. [CrossRef]
- I. M. Ferreira, C. M. De Souza Lacerda, L. S. De Faria, C. R. Corrêa, and A. S. R. De Andrade, “Selection of Peptidoglycan-Specific Aptamers for Bacterial Cells Identification,” Appl Biochem Biotechnol, vol. 174, no. 7, pp. 2548–2556, Dec. 2014. [CrossRef]
- J. Moon, G. Kim, S. Park, J. Lim, and C. Mo, “Comparison of Whole-Cell SELEX Methods for the Identification of Staphylococcus Aureus-Specific DNA Aptamers,” Sensors, vol. 15, no. 4, pp. 8884–8897, Apr. 2015. [CrossRef]
- K. M. Wijesinghe, G. Sabbih, C. H. Algama, R. Syed, M. K. Danquah, and S. Dhakal, “FRET-Based Single-Molecule Detection of Pathogen Protein IsdA Using Computationally Selected Aptamers,” Anal. Chem., vol. 95, no. 26, pp. 9839–9846, Jul. 2023. [CrossRef]
- S. Ohuchi, “Cell-SELEX Technology,” BioResearch Open Access, vol. 1, no. 6, pp. 265–272, Dec. 2012. [CrossRef]
- K. Sefah, D. Shangguan, X. Xiong, M. B. O’Donoghue, and W. Tan, “Development of DNA aptamers using Cell-SELEX,” Nat Protoc, vol. 5, no. 6, pp. 1169–1185, Jun. 2010. [CrossRef]
- K. N. Morris, K. B. Jensen, C. M. Julin, M. Weil, and L. Gold, “High affinity ligands from in vitro selection: Complex targets,” Proc. Natl. Acad. Sci. U.S.A., vol. 95, no. 6, pp. 2902–2907, Mar. 1998. [CrossRef]
- Y.-W. Zhao, H.-X. Wang, G.-C. Jia, and Z. Li, “Application of Aptamer-Based Biosensor for Rapid Detection of Pathogenic Escherichia coli,” Sensors, vol. 18, no. 8, p. 2518, Aug. 2018. [CrossRef]
- W. Zhao et al., “Ultrasensitive dual-enhanced sandwich strategy for simultaneous detection of Escherichia coli and Staphylococcus aureus based on optimized aptamers-functionalized magnetic capture probes and graphene oxide-Au nanostars SERS tags,” Journal of Colloid and Interface Science, vol. 634, pp. 651–663, Mar. 2023. [CrossRef]
- H. P. Dwivedi, R. D. Smiley, and L.-A. Jaykus, “Selection and characterization of DNA aptamers with binding selectivity to Campylobacter jejuni using whole-cell SELEX,” Appl Microbiol Biotechnol, vol. 87, no. 6, pp. 2323–2334, Aug. 2010. [CrossRef]
- J. G. Bruno, T. Phillips, M. P. Carrillo, and R. Crowell, “Plastic-Adherent DNA Aptamer-Magnetic Bead and Quantum Dot Sandwich Assay for Campylobacter Detection,” J Fluoresc, vol. 19, no. 3, pp. 427–435, May 2009. [CrossRef]
- M. Alibolandi et al., “Smart AS1411-aptamer conjugated pegylated PAMAM dendrimer for the superior delivery of camptothecin to colon adenocarcinoma in vitro and in vivo,” International Journal of Pharmaceutics, vol. 519, no. 1–2, pp. 352–364, Mar. 2017. [CrossRef]
- X. Cao et al., “Combining use of a panel of ssDNA aptamers in the detection of Staphylococcus aureus,” Nucleic Acids Research, vol. 37, no. 14, pp. 4621–4628, Aug. 2009. [CrossRef]
- T. T.-Q. Nguyen, E. R. Kim, and M. B. Gu, “A new cognate aptamer pair-based sandwich-type electrochemical biosensor for sensitive detection of Staphylococcus aureus,” Biosensors and Bioelectronics, vol. 198, p. 113835, Feb. 2022. [CrossRef]
- Y.-C. Chang, C.-Y. Yang, R.-L. Sun, Y.-F. Cheng, W.-C. Kao, and P.-C. Yang, “Rapid single cell detection of Staphylococcus aureus by aptamer-conjugated gold nanoparticles,” Sci Rep, vol. 3, no. 1, p. 1863, May 2013. [CrossRef]
- S. H. Lim, Y. C. Ryu, and B. H. Hwang, “Aptamer-immobilized Gold Nanoparticles Enable Facile and On-site Detection of Staphylococcus aureus,” Biotechnol Bioproc E, vol. 26, no. 1, pp. 107–113, Feb. 2021. [CrossRef]
- A. Manfredini, E. Malusà, and L. Canfora, “Aptamer-based technology for detecting Bacillus subtilis in soil,” Appl Microbiol Biotechnol, vol. 107, no. 22, pp. 6963–6972, Nov. 2023. [CrossRef]
- H. P. Dwivedi, R. D. Smiley, and L.-A. Jaykus, “Selection of DNA aptamers for capture and detection of Salmonella Typhimurium using a whole-cell SELEX approach in conjunction with cell sorting,” Appl Microbiol Biotechnol, vol. 97, no. 8, pp. 3677–3686, Apr. 2013. [CrossRef]
- S. H. Suh, H. P. Dwivedi, S. J. Choi, and L.-A. Jaykus, “Selection and characterization of DNA aptamers specific for Listeria species,” Analytical Biochemistry, vol. 459, pp. 39–45, Aug. 2014. [CrossRef]
- P. Setlow, “Resistance of spores of Bacillus species to ultraviolet light,” Environ and Mol Mutagen, vol. 38, no. 2–3, pp. 97–104, Jan. 2001. [CrossRef]
- P. Setlow and G. Christie, “New Thoughts on an Old Topic: Secrets of Bacterial Spore Resistance Slowly Being Revealed,” Microbiol Mol Biol Rev, vol. 87, no. 2, pp. e00080-22, Jun. 2023. [CrossRef]
- V. Mazzaracchio et al., “A label-free impedimetric aptasensor for the detection of Bacillus anthracis spore simulant,” Biosensors and Bioelectronics, vol. 126, pp. 640–646, Feb. 2019. [CrossRef]
- C. Zhou et al., “Aptamer-Conjugated Polydiacetylene Colorimetric Paper Chip for the Detection of Bacillus thuringiensis Spores,” Sensors, vol. 20, no. 11, p. 3124, Jun. 2020. [CrossRef]
- M. Ikanovic et al., “Fluorescence Assay Based on Aptamer-Quantum Dot Binding to Bacillus thuringiensis Spores,” J Fluoresc, vol. 17, no. 2, pp. 193–199, Feb. 2007. [CrossRef]
- A. Asif, H. Mohsin, R. Tanvir, and Y. Rehman, “Revisiting the Mechanisms Involved in Calcium Chloride Induced Bacterial Transformation,” Front. Microbiol., vol. 8, p. 2169, Nov. 2017. [CrossRef]
- K. Raval and T. Ganatra, “Basics, types and applications of molecular docking: A review,” IJCAAP, vol. 7, no. 1, pp. 12–16, Mar. 2022. [CrossRef]
- C. Dominguez, R. Boelens, and A. M. J. J. Bonvin, “HADDOCK: A Protein−Protein Docking Approach Based on Biochemical or Biophysical Information,” J. Am. Chem. Soc., vol. 125, no. 7, pp. 1731–1737, Feb. 2003. [CrossRef]
- G. M. Morris, R. Huey, and A. J. Olson, “Using AutoDock for Ligand-Receptor Docking,” CP in Bioinformatics, vol. 24, no. 1, Dec. 2008. [CrossRef]
- S. Soon and N. Aina Nordin, “In silico predictions and optimization of aptamers against Streptococcus agalactiae surface protein using computational docking,” Materials Today: Proceedings, vol. 16, pp. 2096–2100, 2019. [CrossRef]
- A. Escamilla-Gutiérrez, M. G. Córdova-Espinoza, A. Sánchez-Monciváis, B. Tecuatzi-Cadena, A. G. Regalado-García, and K. Medina-Quero, “In silico selection of aptamers for bacterial toxins detection,” Journal of Biomolecular Structure and Dynamics, vol. 41, no. 20, pp. 10909–10918, Dec. 2023. [CrossRef]
- M. Moradi, H. Mohabatkar, M. Behbahani, and G. Dini, “Application of G-quadruplex aptamer conjugated MSNs to deliver ampicillin for suppressing S. aureus biofilm on mice bone,” Arabian Journal of Chemistry, vol. 15, no. 11, p. 104274, Nov. 2022. [CrossRef]
- R. Selvam, I. H. Y. Lim, J. C. Lewis, C. H. Lim, M. K. K. Yap, and H. S. Tan, “Selecting antibacterial aptamers against the BamA protein in Pseudomonas aeruginosa by incorporating genetic algorithm to optimise computational screening method,” Sci Rep, vol. 13, no. 1, p. 7582, May 2023. [CrossRef]
- K. Yuan et al., “Extracellular Milieu and Membrane Receptor Dual-Driven DNA Nanorobot for Accurate in Vivo Tumor Imaging,” CCS Chem, vol. 4, no. 5, pp. 1597–1609, May 2022. [CrossRef]
- S. B. Ebrahimi, D. Samanta, H. F. Cheng, L. I. Nathan, and C. A. Mirkin, “Forced Intercalation (FIT)-Aptamers,” J. Am. Chem. Soc., vol. 141, no. 35, pp. 13744–13748, Sep. 2019. [CrossRef]
- G. N. Parkinson, M. P. H. Lee, and S. Neidle, “Crystal structure of parallel quadruplexes from human telomeric DNA,” Nature, vol. 417, no. 6891, pp. 876–880, Jun. 2002. [CrossRef]
- R. F. Macaya, P. Schultze, F. W. Smith, J. A. Roe, and J. Feigon, “Thrombin-binding DNA aptamer forms a unimolecular quadruplex structure in solution.,” Proc. Natl. Acad. Sci. U.S.A., vol. 90, no. 8, pp. 3745–3749, Apr. 1993. [CrossRef]
- M. C. Cowperthwaite and A. D. Ellington, “Bioinformatic Analysis of the Contribution of Primer Sequences to Aptamer Structures,” J Mol Evol, vol. 67, no. 1, pp. 95–102, Jul. 2008. [CrossRef]
- M. McKeague et al., “Analysis of In Vitro Aptamer Selection Parameters,” J Mol Evol, vol. 81, no. 5–6, pp. 150–161, Dec. 2015. [CrossRef]
- P. C. Sabeti, P. J. Unrau, and D. P. Bartel, “Accessing rare activities from random RNA sequences: the importance of the length of molecules in the starting pool,” Chemistry & Biology, vol. 4, no. 10, pp. 767–774, Oct. 1997. [CrossRef]
- R. Thevendran and M. Citartan, “Assays to Estimate the Binding Affinity of Aptamers,” Talanta, vol. 238, p. 122971, Feb. 2022. [CrossRef]
- H. Kaur and L.-Y. L. Yung, “Probing High Affinity Sequences of DNA Aptamer against VEGF165,” PLoS ONE, vol. 7, no. 2, p. e31196, Feb. 2012. [CrossRef]
- J. Moon, G. Kim, S. Lee, and S. Park, “Identification of Salmonella Typhimurium-specific DNA aptamers developed using whole-cell SELEX and FACS analysis,” Journal of Microbiological Methods, vol. 95, no. 2, pp. 162–166, Nov. 2013. [CrossRef]
- N. Duan, M. Ye, M. Lu, X. Chen, and S. Wu, “DNA aptamers selection and characterization for development of impedimetric aptasensor for Bacillus cereus at different growing stages,” Advanced Agrochem, vol. 2, no. 3, pp. 284–290, Sep. 2023. [CrossRef]
- D. Yılmaz, T. Muslu, A. Parlar, H. Kurt, and M. Yüce, “SELEX against whole-cell bacteria resulted in lipopolysaccharide binding aptamers,” Journal of Biotechnology, vol. 354, pp. 10–20, Aug. 2022. [CrossRef]
- J.-L. Mergny and L. Lacroix, “Analysis of Thermal Melting Curves,” Oligonucleotides, vol. 13, no. 6, pp. 515–537, Dec. 2003. [CrossRef]
- Y. Luo, A. Granzhan, D. Verga, and J.-L. Mergny, “FRET-MC: A fluorescence melting competition assay for studying G4 structures in vitro,” Biopolymers, vol. 112, no. 4, Dec. 2020. [CrossRef]
- V. Esposito et al., “A straightforward modification in the thrombin binding aptamer improving the stability, affinity to thrombin and nuclease resistance,” Org. Biomol. Chem., vol. 12, no. 44, pp. 8840–8843, 2014. [CrossRef]
- J.-L. Mergny, “Thermal difference spectra: a specific signature for nucleic acid structures,” Nucleic Acids Research, vol. 33, no. 16, pp. e138–e138, Sep. 2005. [CrossRef]
- I. Smirnov and R. H. Shafer, “Effect of Loop Sequence and Size on DNA Aptamer Stability,” Biochemistry, vol. 39, no. 6, pp. 1462–1468, Feb. 2000. [CrossRef]
- S. M. Reilly, R. K. Morgan, T. A. Brooks, and R. M. Wadkins, “Effect of Interior Loop Length on the Thermal Stability and p K a of i-Motif DNA,” Biochemistry, vol. 54, no. 6, pp. 1364–1370, Feb. 2015. [CrossRef]
- S. Ahmed, M. Kaushik, S. Chaudhary, and S. Kukreti, “Structural polymorphism of a cytosine-rich DNA sequence forming i-motif structure: Exploring pH based biosensors,” International Journal of Biological Macromolecules, vol. 111, pp. 455–461, May 2018. [CrossRef]
- B. I. Kankia and L. A. Marky, “Folding of the Thrombin Aptamer into a G-Quadruplex with Sr 2+ : Stability, Heat, and Hydration,” J. Am. Chem. Soc., vol. 123, no. 44, pp. 10799–10804, Nov. 2001. [CrossRef]
- N. Zhang et al., “Structural Biology for the Molecular Insight between Aptamers and Target Proteins,” IJMS, vol. 22, no. 8, p. 4093, Apr. 2021. [CrossRef]
- D. Shangguan, Z. Tang, P. Mallikaratchy, Z. Xiao, and W. Tan, “Optimization and Modifications of Aptamers Selected from Live Cancer Cell Lines,” ChemBioChem, vol. 8, no. 6, pp. 603–606, Apr. 2007. [CrossRef]
- T. Bing, X. Yang, H. Mei, Z. Cao, and D. Shangguan, “Conservative secondary structure motif of streptavidin-binding aptamers generated by different laboratories,” Bioorganic & Medicinal Chemistry, vol. 18, no. 5, pp. 1798–1805, Mar. 2010. [CrossRef]
- G. Xu et al., “Structural basis for high-affinity recognition of aflatoxin B1 by a DNA aptamer,” Nucleic Acids Research, vol. 51, no. 14, pp. 7666–7674, Aug. 2023. [CrossRef]
- M. Gellert, M. N. Lipsett, and D. R. Davies, “HELIX FORMATION BY GUANYLIC ACID,” Proc. Natl. Acad. Sci. U.S.A., vol. 48, no. 12, pp. 2013–2018, Dec. 1962. [CrossRef]
- K. W. Lim and A. T. Phan, “Structural Basis of DNA Quadruplex–Duplex Junction Formation,” Angew Chem Int Ed, vol. 52, no. 33, pp. 8566–8569, Aug. 2013. [CrossRef]
- R. Stoltenburg, P. Krafčiková, V. Víglaský, and B. Strehlitz, “G-quadruplex aptamer targeting Protein A and its capability to detect Staphylococcus aureus demonstrated by ELONA,” Sci Rep, vol. 6, no. 1, p. 33812, Sep. 2016. [CrossRef]
- K. Gehring, J.-L. Leroy, and M. Gueron, “A tetrameric DNA structure with protonated cytosine· cytosine base pairs,” vol. 363, 1993. [CrossRef]
- X.-J. Lu, “3DNA: a software package for the analysis, rebuilding and visualization of three-dimensional nucleic acid structures,” Nucleic Acids Research, vol. 31, no. 17, pp. 5108–5121, Sep. 2003. [CrossRef]
- A. R. Gruber, R. Lorenz, S. H. Bernhart, R. Neubock, and I. L. Hofacker, “The Vienna RNA Websuite,” Nucleic Acids Research, vol. 36, no. Web Server, pp. W70–W74, May 2008. [CrossRef]
- M. Biesiada, K. J. Purzycka, M. Szachniuk, J. Blazewicz, and R. W. Adamiak, “Automated RNA 3D Structure Prediction with RNAComposer,” in RNA Structure Determination, vol. 1490, D. H. Turner and D. H. Mathews, Eds., in Methods in Molecular Biology, vol. 1490., New York, NY: Springer New York, 2016, pp. 199–215. [CrossRef]
- B. Gao, Y.-T. Zheng, A.-M. Su, B. Sun, X.-G. Xi, and X.-M. Hou, “Remodeling the conformational dynamics of I-motif DNA by helicases in ATP-independent mode at acidic environment,” iScience, vol. 25, no. 1, p. 103575, Jan. 2022. [CrossRef]
- J. Kypr, I. Kejnovska, D. Renciuk, and M. Vorlickova, “Circular dichroism and conformational polymorphism of DNA,” Nucleic Acids Research, vol. 37, no. 6, pp. 1713–1725, Jan. 2009. [CrossRef]
- P. Bielecka, A. Dembska, and B. Juskowiak, “Monitoring of pH Using an i-Motif-Forming Sequence Containing a Fluorescent Cytosine Analogue, tC,” Molecules, vol. 24, no. 5, p. 952, Mar. 2019. [CrossRef]
- C. Chen et al., “Study of pH-Induced Folding and Unfolding Kinetics of the DNA i-Motif by Stopped-Flow Circular Dichroism,” Langmuir, vol. 28, no. 51, pp. 17743–17748, Dec. 2012. [CrossRef]
- T. Santos et al., “Stabilization of a DNA aptamer by ligand binding,” Biochimie, vol. 200, pp. 8–18, Sep. 2022. [CrossRef]
- P.-H. Lin, R.-H. Chen, C.-H. Lee, Y. Chang, C.-S. Chen, and W.-Y. Chen, “Studies of the binding mechanism between aptamers and thrombin by circular dichroism, surface plasmon resonance and isothermal titration calorimetry,” Colloids and Surfaces B: Biointerfaces, vol. 88, no. 2, pp. 552–558, Dec. 2011. [CrossRef]
- R. Troisi, N. Balasco, I. Autiero, L. Vitagliano, and F. Sica, “Structural Insights into Protein–Aptamer Recognitions Emerged from Experimental and Computational Studies,” IJMS, vol. 24, no. 22, p. 16318, Nov. 2023. [CrossRef]
- S. S. Wijmenga and B. N. M. Van Buuren, “The use of NMR methods for conformational studies of nucleic acids,” Progress in Nuclear Magnetic Resonance Spectroscopy, vol. 32, no. 4, pp. 287–387, Jun. 1998. [CrossRef]
- T. Someya, S. Baba, M. Fujimoto, G. Kawai, T. Kumasaka, and K. Nakamura, “Crystal structure of Hfq from Bacillus subtilis in complex with SELEX-derived RNA aptamer: insight into RNA-binding properties of bacterial Hfq,” Nucleic Acids Research, vol. 40, no. 4, pp. 1856–1867, Feb. 2012. [CrossRef]
- E. Menichelli et al., “Discovery of small molecules that target a tertiary-structured RNA,” Proc. Natl. Acad. Sci. U.S.A., vol. 119, no. 48, p. e2213117119, Nov. 2022. [CrossRef]
- R. Troisi et al., “Steric hindrance and structural flexibility shape the functional properties of a guanine-rich oligonucleotide,” Nucleic Acids Research, vol. 51, no. 16, pp. 8880–8890, Sep. 2023. [CrossRef]
- C. Kratschmer and M. Levy, “Effect of Chemical Modifications on Aptamer Stability in Serum,” Nucleic Acid Therapeutics, vol. 27, no. 6, pp. 335–344, Dec. 2017. [CrossRef]
- C. G. Peng and M. J. Damha, “G-quadruplex induced stabilization by 2′-deoxy-2′-fluoro-d-arabinonucleic acids (2′F-ANA),” Nucleic Acids Research, vol. 35, no. 15, pp. 4977–4988, Aug. 2007. [CrossRef]
- J. P. Elskens, J. M. Elskens, and A. Madder, “Chemical Modification of Aptamers for Increased Binding Affinity in Diagnostic Applications: Current Status and Future Prospects,” International Journal of Molecular Sciences, vol. 21, no. 12, p. 4522, Jun. 2020. [CrossRef]
- M. R. Dunn, R. M. Jimenez, and J. C. Chaput, “Analysis of aptamer discovery and technology,” Nat Rev Chem, vol. 1, no. 10, p. 0076, Oct. 2017. [CrossRef]
- Y. Kasahara and M. Kuwahara, “Artificial Specific Binders Directly Recovered from Chemically Modified Nucleic Acid Libraries,” Journal of Nucleic Acids, vol. 2012, pp. 1–13, 2012. [CrossRef]
- J. Byun, “Recent Progress and Opportunities for Nucleic Acid Aptamers,” Life, vol. 11, no. 3, p. 193, Feb. 2021. [CrossRef]
- F. Odeh et al., “Aptamers Chemistry: Chemical Modifications and Conjugation Strategies,” Molecules, vol. 25, no. 1, p. 3, Dec. 2019. [CrossRef]
- A. R. Chandrasekaran, “Nuclease resistance of DNA nanostructures,” Nat Rev Chem, vol. 5, no. 4, pp. 225–239, Feb. 2021. [CrossRef]
- Q. Liu et al., “Enhanced Stability of DNA Nanostructures by Incorporation of Unnatural Base Pairs,” ChemPhysChem, vol. 18, no. 21, pp. 2977–2980, Nov. 2017. [CrossRef]
- Y. Tabuchi, J. Yang, and M. Taki, “Relative Nuclease Resistance of a DNA Aptamer Covalently Conjugated to a Target Protein,” IJMS, vol. 23, no. 14, p. 7778, Jul. 2022. [CrossRef]
- A. Lacroix, T. G. W. Edwardson, M. A. Hancock, M. D. Dore, and H. F. Sleiman, “Development of DNA Nanostructures for High-Affinity Binding to Human Serum Albumin,” J. Am. Chem. Soc., vol. 139, no. 21, pp. 7355–7362, May 2017. [CrossRef]
- N. Ponnuswamy et al., “Oligolysine-based coating protects DNA nanostructures from low-salt denaturation and nuclease degradation,” Nat Commun, vol. 8, no. 1, p. 15654, May 2017. [CrossRef]
- J.-W. Keum and H. Bermudez, “Enhanced resistance of DNA nanostructures to enzymatic digestion,” Chem. Commun., no. 45, p. 7036, 2009. [CrossRef]
- T. Gerling, M. Kube, B. Kick, and H. Dietz, “Sequence-programmable covalent bonding of designed DNA assemblies,” Sci. Adv., vol. 4, no. 8, p. eaau1157, Aug. 2018. [CrossRef]
- R. El-Khoury and M. J. Damha, “End-ligation can dramatically stabilize i-motifs at neutral pH,” Chem. Commun., vol. 59, no. 25, pp. 3715–3718, 2023. [CrossRef]
- J. Hahn, S. F. J. Wickham, W. M. Shih, and S. D. Perrault, “Addressing the Instability of DNA Nanostructures in Tissue Culture,” ACS Nano, vol. 8, no. 9, pp. 8765–8775, Sep. 2014. [CrossRef]
- Y. Lian, F. He, H. Wang, and F. Tong, “A new aptamer/graphene interdigitated gold electrode piezoelectric sensor for rapid and specific detection of Staphylococcus aureus,” Biosensors and Bioelectronics, vol. 65, pp. 314–319, Mar. 2015. [CrossRef]
- Q. Kang et al., “A novel Aptamer-induced CHA amplification strategy for ultrasensitive detection of Staphylococcus aureus and NIR-triggered photothermal bactericidal Activity based on aptamer-modified magnetic Fe3O4@AuNRs,” Sensors and Actuators B: Chemical, vol. 382, p. 133554, May 2023. [CrossRef]
- A. Abbaspour, F. Norouz-Sarvestani, A. Noori, and N. Soltani, “Aptamer-conjugated silver nanoparticles for electrochemical dual-aptamer-based sandwich detection of staphylococcus aureus,” Biosensors and Bioelectronics, vol. 68, pp. 149–155, Jun. 2015. [CrossRef]
- M. Marin, F. Rizzotto, V. Léguillier, C. Péchoux, E. Borezee-Durant, and J. Vidic, “Naked-eye detection of Staphylococcus aureus in powdered milk and infant formula using gold nanoparticles,” Journal of Microbiological Methods, vol. 201, p. 106578, Oct. 2022. [CrossRef]
- Y. S. Kim, M. Y. Song, J. Jurng, and B. C. Kim, “Isolation and characterization of DNA aptamers against Escherichia coli using a bacterial cell–systematic evolution of ligands by exponential enrichment approach,” Analytical Biochemistry, vol. 436, no. 1, pp. 22–28, May 2013. [CrossRef]
- J. A. DeGrasse, “A Single-Stranded DNA Aptamer That Selectively Binds to Staphylococcus aureus Enterotoxin B,” PLoS ONE, vol. 7, no. 3, p. e33410, Mar. 2012. [CrossRef]
- N. Alizadeh, M. Y. Memar, B. Mehramuz, S. S. Abibiglou, F. Hemmati, and H. Samadi Kafil, “Current advances in aptamer-assisted technologies for detecting bacterial and fungal toxins,” J Appl Microbiol, vol. 124, no. 3, pp. 644–651, Mar. 2018. [CrossRef]
- J. Chen et al., “Multichannel-Structured Three-Dimensional Chip for Highly Sensitive Pathogenic Bacteria Detection Based on Fast DNA-Programmed Signal Polymerization,” Anal. Chem., vol. 90, no. 20, pp. 12019–12026, Oct. 2018. [CrossRef]
- F. Bakhshandeh et al., “A universal bacterial sensor created by integrating a light modulating aptamer complex with photoelectrochemical signal readout,” Biosensors and Bioelectronics, vol. 235, p. 115359, Sep. 2023. [CrossRef]
- Y. Zhang, Y. Liu, Y. Yang, L. Li, X. Tao, and E. Song, “Rapid detection of pathogenic bacteria based on a universal dual-recognition FRET sensing system constructed with aptamer-quantum dots and lectin-gold nanoparticles,” Chinese Chemical Letters, vol. 34, no. 8, p. 108102, Aug. 2023. [CrossRef]
- H.-S. Shin, V. Gedi, J.-K. Kim, and D. Lee, “Detection of Gram-negative bacterial outer membrane vesicles using DNA aptamers,” Sci Rep, vol. 9, no. 1, p. 13167, Sep. 2019. [CrossRef]
- J. C. Niles and M. A. Marletta, “Utilizing RNA Aptamers To Probe a Physiologically Important Heme-Regulated Cellular Network,” ACS Chem. Biol., vol. 1, no. 8, pp. 515–524, Sep. 2006. [CrossRef]
- J. G. Bruno, M. P. Carrillo, T. Phillips, and C. J. Andrews, “A Novel Screening Method for Competitive FRET-Aptamers Applied to E. coli Assay Development,” J Fluoresc, vol. 20, no. 6, pp. 1211–1223, Nov. 2010. [CrossRef]
- R. B. Queirós, N. de-los-Santos-Álvarez, J. P. Noronha, and M. G. F. Sales, “A label-free DNA aptamer-based impedance biosensor for the detection of E. coli outer membrane proteins,” Sensors and Actuators B: Chemical, vol. 181, pp. 766–772, May 2013. [CrossRef]
- D. Jiang et al., “Ultra-sensitive photoelectrochemical aptamer biosensor for detecting E. coli O157:H7 based on nonmetallic plasmonic two-dimensional hydrated defective tungsten oxide nanosheets coupling with nitrogen-doped graphene quantum dots (dWO3•H2O@N-GQDs),” Biosensors and Bioelectronics, vol. 183, p. 113214, Jul. 2021. [CrossRef]
- H. Kaur, M. Shorie, and P. Sabherwal, “Biolayer interferometry-SELEX for Shiga toxin antigenic-peptide aptamers & detection via chitosan-WSe2 aptasensor,” Biosensors and Bioelectronics, vol. 167, p. 112498, Nov. 2020. [CrossRef]
- F. R. W. Schmitz, K. Cesca, A. Valério, D. De Oliveira, and D. Hotza, “Colorimetric detection of Pseudomonas aeruginosa by aptamer-functionalized gold nanoparticles,” Appl Microbiol Biotechnol, vol. 107, no. 1, pp. 71–80, Jan. 2023. [CrossRef]
- H. Zheng, R. Sheng, H. Li, W. Ahmad, and Q. Chen, “Rapid and selective detection of Bacillus cereus in food using cDNA-based up-conversion fluorescence spectrum copy and aptamer modified magnetic separation,” Spectrochimica Acta Part A: Molecular and Biomolecular Spectroscopy, vol. 267, p. 120618, Feb. 2022. [CrossRef]
- J. Yan et al., “Dual recognition strategy for the rapid and precise detection of Bacillus cereus using post-modified nano-MOF and aptamer,” Sensors and Actuators B: Chemical, vol. 386, p. 133745, Jul. 2023. [CrossRef]
- Z. Zhou et al., “Portable dual-aptamer microfluidic chip biosensor for Bacillus cereus based on aptamer tailoring and dumbbell-shaped probes,” Journal of Hazardous Materials, vol. 445, p. 130545, Mar. 2023. [CrossRef]
- C. Y. Effah et al., “A SERS bioassay based on vancomycin-modified PEI-interlayered nanocomposite and aptamer-functionalized SERS tags for synchronous detection of Acinetobacter baumannii and Klebsiella pneumoniae,” Food Chemistry, vol. 423, p. 136242, Oct. 2023. [CrossRef]
- R. Abedi, J. B. Raoof, M. Mohseni, and A. Bagheri Hashkavayi, “Development of a label-free impedimetric aptasensor for the detection of Acinetobacter baumannii bacteria,” Analytical Biochemistry, vol. 679, p. 115288, Oct. 2023. [CrossRef]
- A. Vishwakarma, Y. Meganathan, and M. Ramya, “Aptamer-based assay for rapid detection, surveillance, and screening of pathogenic Leptospira in water samples,” Sci Rep, vol. 13, no. 1, p. 13379, Aug. 2023. [CrossRef]
- M. Sun, N. Ma, H. Shi, L.-Z. Cheong, W. Yang, and Z. Qiao, “A HCR based multivalent aptamer amplifier for ultrasensitive detection of Salmonella,” Sensors and Actuators B: Chemical, vol. 375, p. 132860, Jan. 2023. [CrossRef]
- M. Tavassoli, A. Khezerlou, H. Hamishehkar, A. Ehsani, and B. Khalilzadeh, “An ultrasensitive aptamer-based fluorescent on/off system for trace amount evaluation of Yersinia enterocolitica in food samples,” Microchimica Acta, vol. 190, no. 253, Jun. 2023. [CrossRef]
- P. Luo, Y. Liu, Y. Xia, H. Xu, and G. Xie, “Aptamer biosensor for sensitive detection of toxin A of Clostridium difficile using gold nanoparticles synthesized by Bacillus stearothermophilus,” Biosensors and Bioelectronics, vol. 54, pp. 217–221, Apr. 2014. [CrossRef]
- J. Hu, Z. Shen, L. Tan, J. Yuan, and N. Gan, “Electrochemical aptasensor for simultaneous detection of foodborne pathogens based on a double stirring bars-assisted signal amplification strategy,” Sensors and Actuators B: Chemical, vol. 345, p. 130337, Oct. 2021. [CrossRef]
- J. Vidic et al., “Point-of-Need DNA Testing for Detection of Foodborne Pathogenic Bacteria,” Sensors, vol. 19, no. 5, p. 1100, Mar. 2019. [CrossRef]
- M. Majdinasab, A. Hayat, and J. L. Marty, “Aptamer-based assays and aptasensors for detection of pathogenic bacteria in food samples,” TrAC Trends in Analytical Chemistry, vol. 107, pp. 60–77, Oct. 2018. [CrossRef]
- C. Fischer et al., “Food Sensing: Aptamer-Based Trapping of Bacillus cereus Spores with Specific Detection via Real Time PCR in Milk,” J. Agric. Food Chem., vol. 63, no. 36, pp. 8050–8057, Sep. 2015. [CrossRef]
- Y. S. Kim, J. Chung, M. Y. Song, J. Jurng, and B. C. Kim, “Aptamer cocktails: Enhancement of sensing signals compared to single use of aptamers for detection of bacteria,” Biosensors and Bioelectronics, vol. 54, pp. 195–198, Apr. 2014. [CrossRef]
- X. Lin, P. P. Liu, J. Yan, D. Luan, T. Sun, and X. Bian, “Dual Synthetic Receptor-Based Sandwich Electrochemical Sensor for Highly Selective and Ultrasensitive Detection of Pathogenic Bacteria at the Single-Cell Level,” Anal. Chem., vol. 95, no. 13, pp. 5561–5567, Apr. 2023. [CrossRef]
- Z. Ding, et al., “Rolling Circle Amplification/G-Quadruplex-Based Dual Signal Ratiometric Electrochemical Aptasensor for Ultrasensitive Detection of Pathogenic Bacteria,” ChemElectroChem, vol. 10, no. e202300257, Aug. 2023. [CrossRef]
- V. V. Demidov, “Rolling-circle amplification in DNA diagnostics: the power of simplicity,” Expert Review of Molecular Diagnostics, vol. 2, no. 6, pp. 542–548, Nov. 2002. [CrossRef]
- W. Zhao, M. M. Ali, M. A. Brook, and Y. Li, “Rolling Circle Amplification: Applications in Nanotechnology and Biodetection with Functional Nucleic Acids,” Angew Chem Int Ed, vol. 47, no. 34, pp. 6330–6337, Aug. 2008. [CrossRef]
- M. M. Ali et al., “Rolling circle amplification: a versatile tool for chemical biology, materials science and medicine,” Chem. Soc. Rev., vol. 43, no. 10, p. 3324, 2014. [CrossRef]
- F.-T. Zhang, J. Nie, D.-W. Zhang, J.-T. Chen, Y.-L. Zhou, and X.-X. Zhang, “Methylene Blue as a G-Quadruplex Binding Probe for Label-Free Homogeneous Electrochemical Biosensing,” Anal. Chem., vol. 86, no. 19, pp. 9489–9495, Oct. 2014. [CrossRef]
- J. G. Bruno and M. P. Carrillo, “Development of Aptamer Beacons for Rapid Presumptive Detection of Bacillus Spores,” J Fluoresc, vol. 22, no. 3, pp. 915–924, May 2012. [CrossRef]
- T. Martinović, U. Andjelković, M. Š. Gajdošik, D. Rešetar, and D. Josić, “Foodborne pathogens and their toxins,” Journal of Proteomics, vol. 147, pp. 226–235, Sep. 2016. [CrossRef]
- M. Otto, “Staphylococcus aureus toxins,” Current Opinion in Microbiology, vol. 17, pp. 32–37, Feb. 2014. [CrossRef]
- A.-M. Tătaru, A. Canciu, M. Tertiș, C. Cristea, and A. Cernat, “Staphylococcus aureus – Review on potential targets for sensors development,” Bioelectrochemistry, vol. 153, p. 108492, Oct. 2023. [CrossRef]
- J. Mainil, “Escherichia coli virulence factors,” Veterinary Immunology and Immunopathology, vol. 152, no. 1–2, pp. 2–12, Mar. 2013. [CrossRef]
- W. K. Smits, D. Lyras, D. B. Lacy, M. H. Wilcox, and E. J. Kuijper, “Clostridium difficile infection,” Nat Rev Dis Primers, vol. 2, no. 1, p. 16020, Apr. 2016. [CrossRef]
- C. L. Pickett, E. C. Pesci, D. L. Cottle, G. Russell, A. N. Erdem, and H. Zeytin, “Prevalence of cytolethal distending toxin production in Campylobacter jejuni and relatedness of Campylobacter sp. cdtB gene,” Infect Immun, vol. 64, no. 6, pp. 2070–2078, Jun. 1996. [CrossRef]
- R. L. T. Churchill, H. Lee, and J. C. Hall, “Detection of Listeria monocytogenes and the toxin listeriolysin O in food,” Journal of Microbiological Methods, vol. 64, no. 2, pp. 141–170, Feb. 2006. [CrossRef]
- A. Platt-Samoraj, “Toxigenic Properties of Yersinia enterocolitica Biotype 1A,” Toxins, vol. 14, no. 2, p. 118, Feb. 2022. [CrossRef]
- G. Sakaguchi, “Clostridium botulinum toxin,” Pharmacology & therapeutics, vol. 19, no. 2, pp. 165–194, 1983. [CrossRef]
- A. Rajkovic, “Microbial toxins and low level of foodborne exposure,” Trends in Food Science & Technology, vol. 38, no. 2, pp. 149–157, Aug. 2014. [CrossRef]
- S. Mousavi Nodoushan, N. Nasirizadeh, J. Amani, R. Halabian, and A. A. Imani Fooladi, “An electrochemical aptasensor for staphylococcal enterotoxin B detection based on reduced graphene oxide and gold nano-urchins,” Biosensors and Bioelectronics, vol. 127, pp. 221–228, Feb. 2019. [CrossRef]
- M. Gholamzad, M. R. Khatami, S. Ghassemi, Z. Vaise Malekshahi, and M. B. Shooshtari, “Detection of Staphylococcus Enterotoxin B (SEB) Using an Immunochromatographic Test Strip,” Jundishapur J Microbiol, vol. 8, no. 9, Sep. 2015. [CrossRef]
- B. Jin et al., “Lateral flow aptamer assay integrated smartphone-based portable device for simultaneous detection of multiple targets using upconversion nanoparticles,” Sensors and Actuators B: Chemical, vol. 276, pp. 48–56, Dec. 2018. [CrossRef]
- C. Roca et al., “Selection of an Aptamer against the Enzyme 1-deoxy-D-xylulose-5-phosphate Reductoisomerase from Plasmodium falciparum,” Pharmaceutics, vol. 14, no. 11, p. 2515, Nov. 2022. [CrossRef]
- K. Wang et al., “Neutralization of Staphylococcal Enterotoxin B by an Aptamer Antagonist,” Antimicrob Agents Chemother, vol. 59, no. 4, pp. 2072–2077, Apr. 2015. [CrossRef]
- L. Rotariu, F. Lagarde, N. Jaffrezic-Renault, and C. Bala, “Electrochemical biosensors for fast detection of food contaminants – trends and perspective,” TrAC Trends in Analytical Chemistry, vol. 79, pp. 80–87, May 2016. [CrossRef]
- L. Xu, D. Li, S. Ramadan, Y. Li, and N. Klein, “Facile biosensors for rapid detection of COVID-19,” Biosensors and Bioelectronics, vol. 170, p. 112673, Dec. 2020. [CrossRef]
- Y. Lu, Z. Shi, and Q. Liu, “Smartphone-based biosensors for portable food evaluation,” Current Opinion in Food Science, vol. 28, pp. 74–81, Aug. 2019. [CrossRef]
- J. R. Choi, “Development of Point-of-Care Biosensors for COVID-19,” Front. Chem., vol. 8, p. 517, May 2020. [CrossRef]
- V. Soheili, S. M. Taghdisi, K. Abnous, and M. Ebrahimi, “Point-of-care detection of Escherichia coli O157:H7 in water using AuNPs-based aptasensor,” Iran J Basic Med Sci, vol. 23, no. 7, 2020. [CrossRef]
- H. Yang, Y. Zhou, and J. Liu, “G-quadruplex DNA for construction of biosensors,” TrAC Trends in Analytical Chemistry, vol. 132, p. 116060, Nov. 2020. [CrossRef]
- J. Dejeu, A. Van Der Heyden, N. Spinelli, E. Defrancq, and L. Coche-Guérente, “Recent progress in the design of G-quadruplex–based electrochemical aptasensors,” Current Opinion in Electrochemistry, vol. 30, p. 100812, Dec. 2021. [CrossRef]
Figure 1.
Major aptamer characteristics and applications.
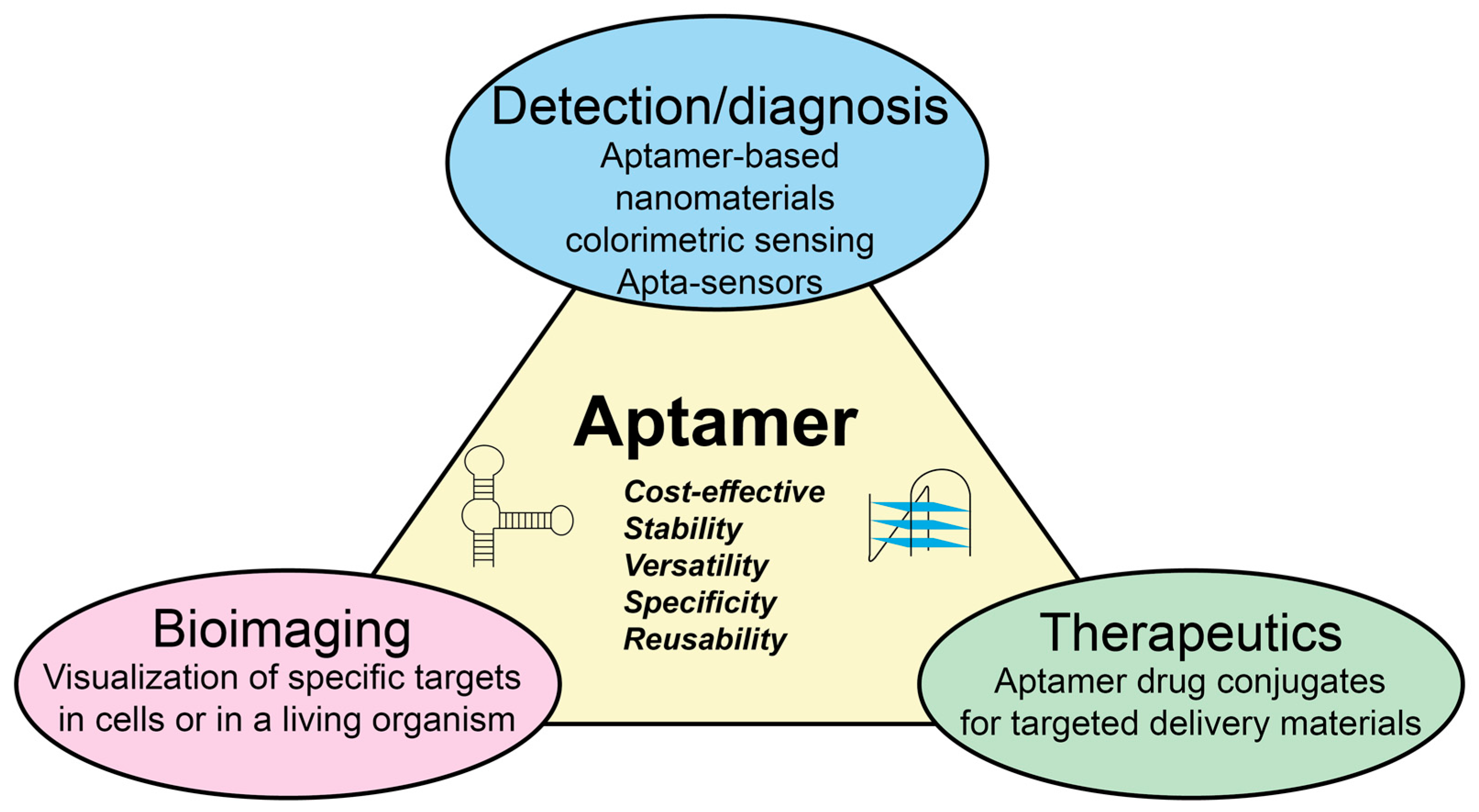
Figure 2.
Aptamer development and their application in bacteriology in the literature in the period of 1990–2021. Values were obtained by searching “aptamer” and “aptamer & bacteria” keywords in Scopus. Trends obtained by fitting a tendency curve and projecting it for the next 4 years.
Figure 2.
Aptamer development and their application in bacteriology in the literature in the period of 1990–2021. Values were obtained by searching “aptamer” and “aptamer & bacteria” keywords in Scopus. Trends obtained by fitting a tendency curve and projecting it for the next 4 years.
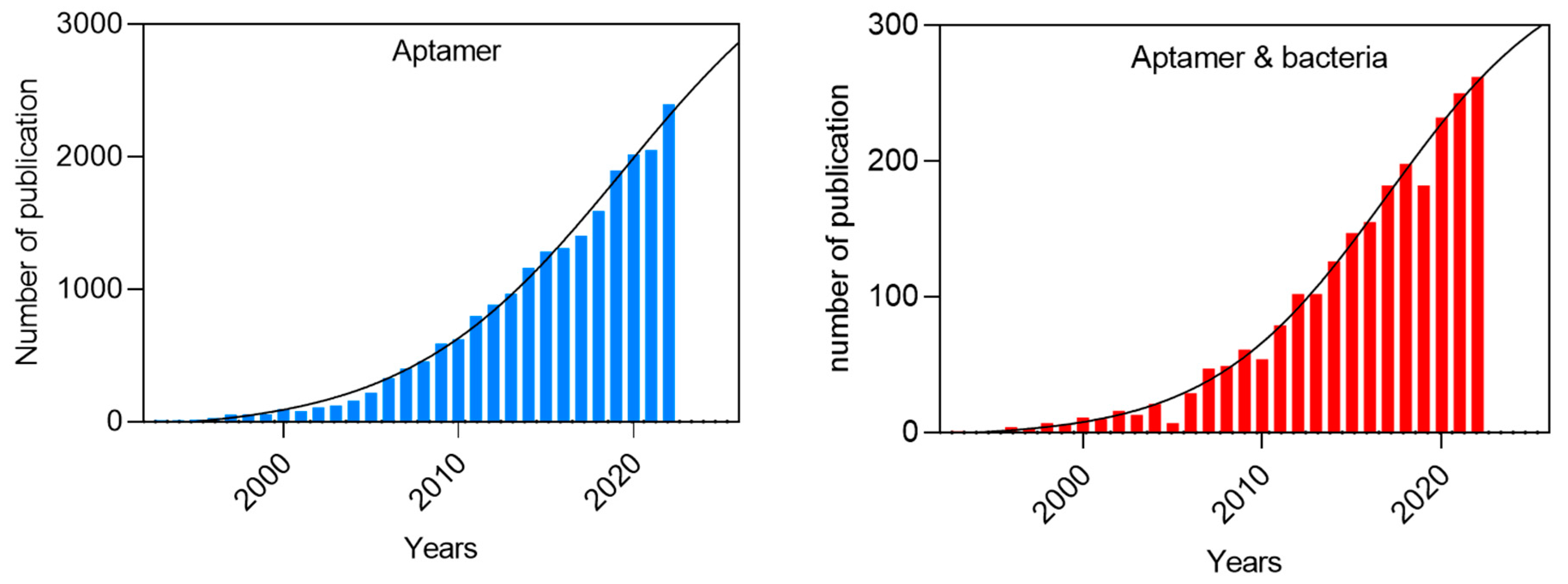
Figure 3.
Schematic of non-canonical aptamer structures: hairpins and stem-loop (A), G-quadruplex (B), and I-motif (C). .
Figure 3.
Schematic of non-canonical aptamer structures: hairpins and stem-loop (A), G-quadruplex (B), and I-motif (C). .
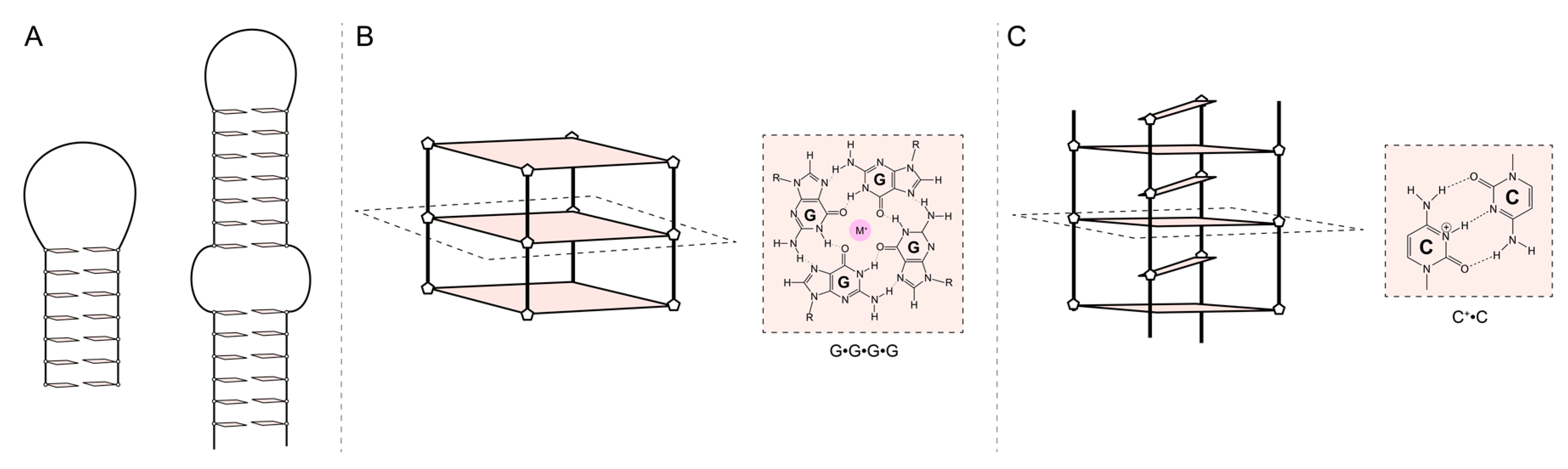
Figure 4.
Schematic illustration presenting (A) Preparation of the BIF-modified glassy carbon electrode and (B) Preparation of aptamer carrying signal nanoprobe; and (C) Dual BIF-aptamer based electrochemical sandwich detection of S. aureus cells. Reprinted with permission from [168]. Copyright 2023. American Chemical Society.
Figure 4.
Schematic illustration presenting (A) Preparation of the BIF-modified glassy carbon electrode and (B) Preparation of aptamer carrying signal nanoprobe; and (C) Dual BIF-aptamer based electrochemical sandwich detection of S. aureus cells. Reprinted with permission from [168]. Copyright 2023. American Chemical Society.
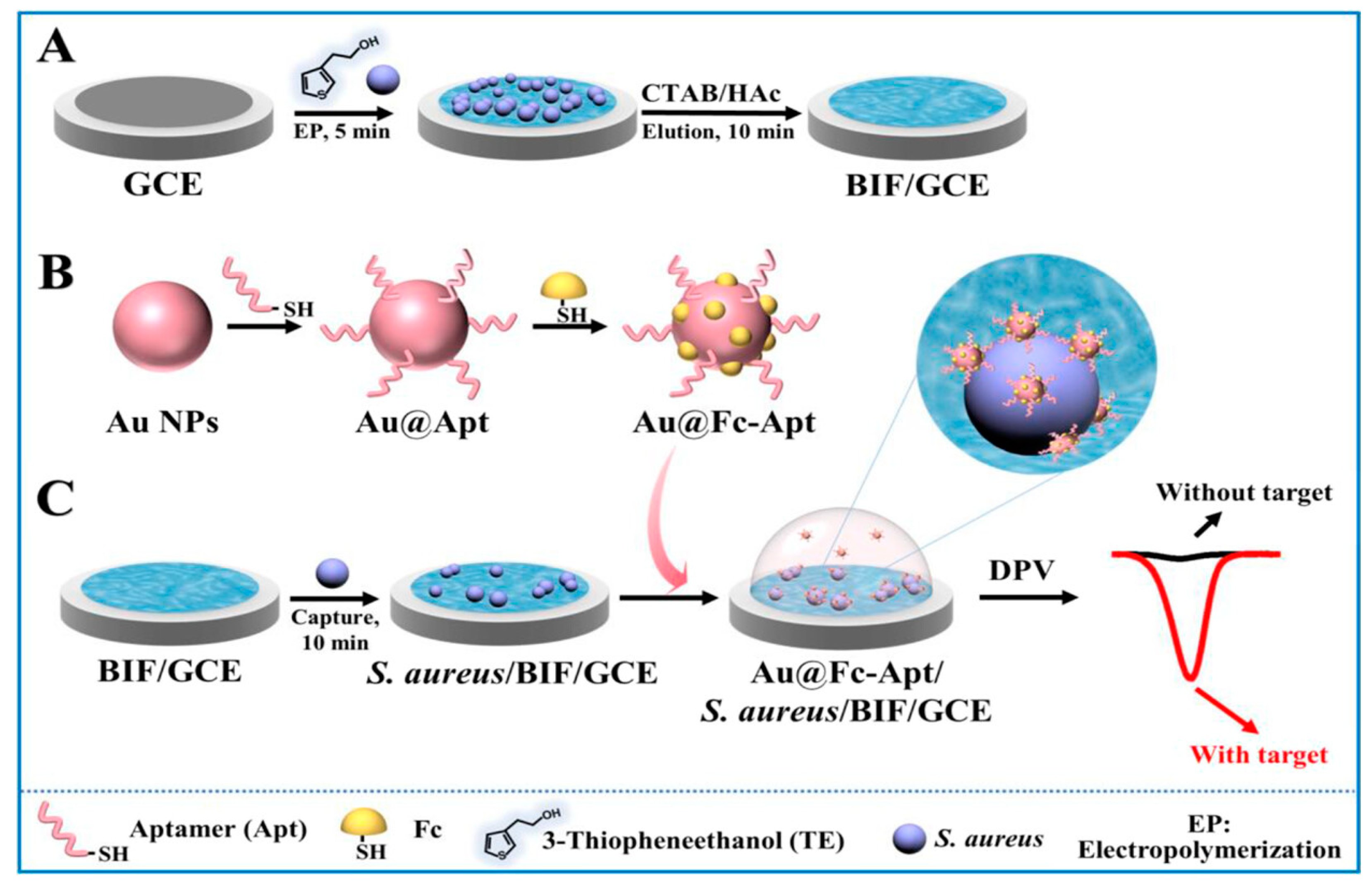
Figure 5.
Schematic illustration presenting ratiometric dual-signal electrochemical biosensor for ultrasensitive detection of pathogenic bacteria based on aptamer recognition-induced rolling circle amplification (RCA)/G-quadruplex strategy. Adapted with permission from [169].
Figure 5.
Schematic illustration presenting ratiometric dual-signal electrochemical biosensor for ultrasensitive detection of pathogenic bacteria based on aptamer recognition-induced rolling circle amplification (RCA)/G-quadruplex strategy. Adapted with permission from [169].
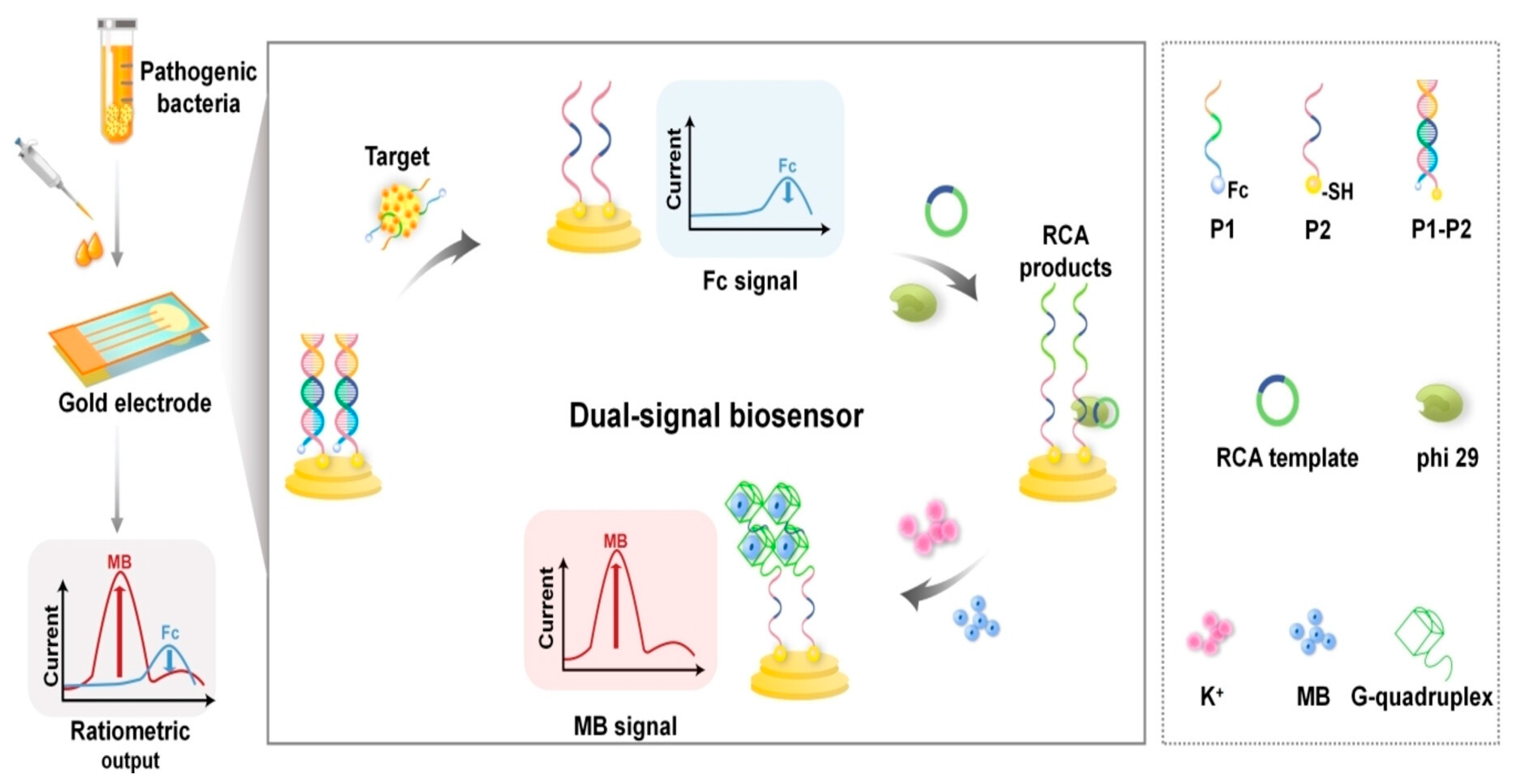
Figure 6.
Schematic illustration presenting colorimetric biosensor for detection of B. cytotoxicus spores based on spore-enhanced peroxidase-like catalytic activity of AuNPs. Adapted with permission from [29].
Figure 6.
Schematic illustration presenting colorimetric biosensor for detection of B. cytotoxicus spores based on spore-enhanced peroxidase-like catalytic activity of AuNPs. Adapted with permission from [29].
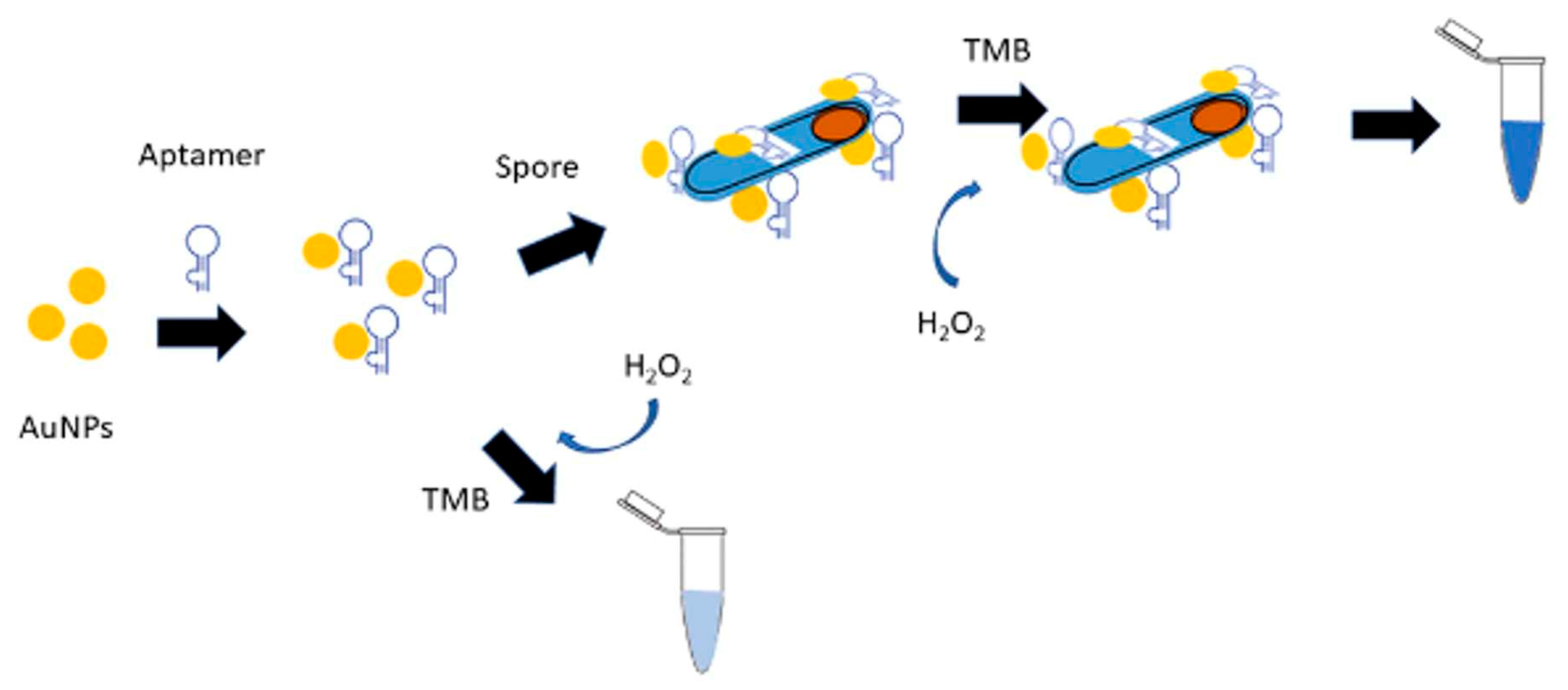
Figure 7.
Schematic illustration of fabrication process of SEB aptasensor using reduced graphene oxide and gold nano-urchins. Adapted with permission from [185].
Figure 7.
Schematic illustration of fabrication process of SEB aptasensor using reduced graphene oxide and gold nano-urchins. Adapted with permission from [185].
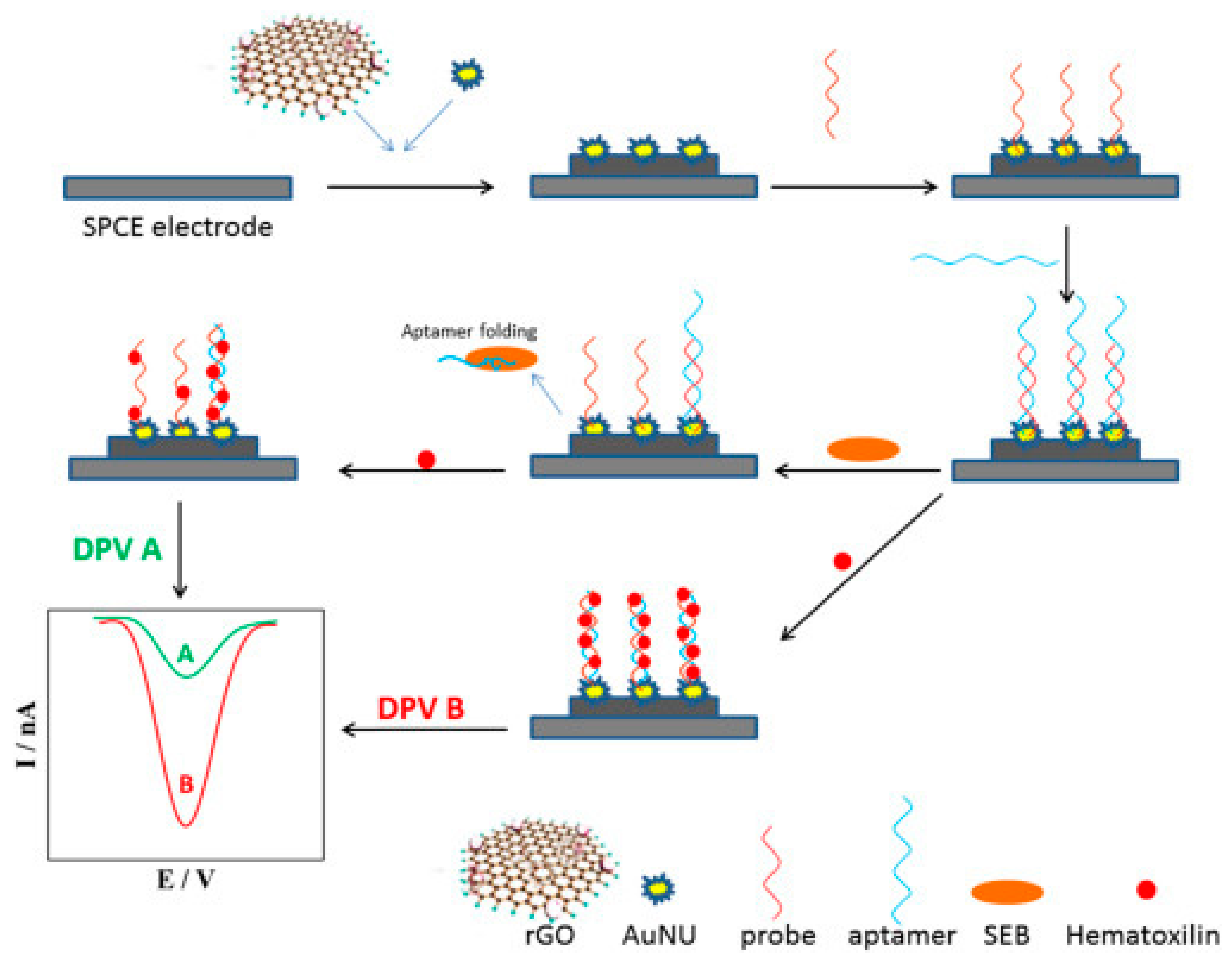
Figure 8.
Schematic illustration of lateral flow assay based on 3 different aptamers for simultaneous multiple targets detection. Adapted with permission from [187].
Figure 8.
Schematic illustration of lateral flow assay based on 3 different aptamers for simultaneous multiple targets detection. Adapted with permission from [187].
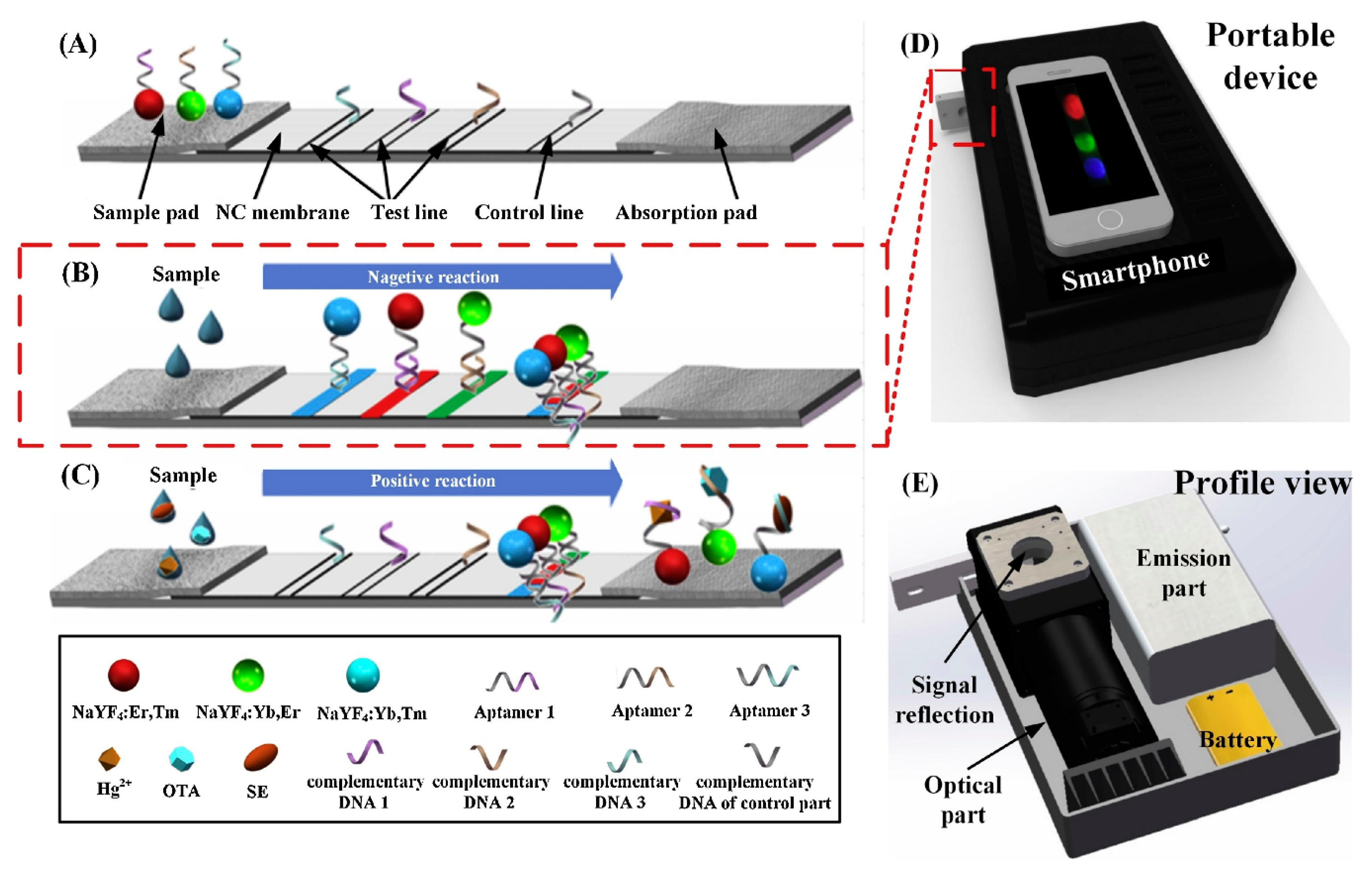

Table 1.
Aptamer-based biosensors for bacterial detection. Aptamer sequences are provided in Supplementary data.
Table 1.
Aptamer-based biosensors for bacterial detection. Aptamer sequences are provided in Supplementary data.
| Bacterium | Aptamer | DNA or RNA | Target | Kd (nM) | Detection method | LOD | Linear range (CFU/mL) | Ref |
|---|---|---|---|---|---|---|---|---|
| S. aureus | SA20 SA23 SA34 SA31 SA43 |
DNA | Whole cell | 70.86±39.22 61.50±22.43 72.42±35.23 82.86±33.20 210.7±135.9 |
Fluorescence |
/ | / | [59] |
| S. aureus | T1 T2 T3 A14 |
RNA & DNA |
IsdA | 2.2 ± 0.5 1.0 ± 0.3 0.7 ± 0.4 4 ± 2 |
FRET-switch | 113 pM 17 pM 11 pM 485 pM |
/ | [50] |
| S. aureus | H1 | DNA | / | / | Piezoelectric sensor | 41 CFU/mL | 4.1 x 101 to 4.1 x 105 | [137] |
| S. aureus | H1 H2 cApt |
DNA | / | / | Fluorescence | 101 CFU/mL | 102-106 | [138] |
| S. aureus | Apt1 Apt2 |
DNA | Whole cell | 35 129 |
Colorimetric | 7,5 – 8,4 * 104 CFU/mL | 104 - 108 | [139,140] |
| S. aureus | SH-Apt2 | DNA | Whole cell | 210.7 | Surface-enhanced Raman scattering (SERS) | / | / | [55,141] |
| S. aureus | APTseb1 | DNA | Staphylococcal Enterotoxin B (SEB) | / | SDS-Page | / | / | [142] |
| S. aureus | G1 #2 #18 |
RNA | Teichoic acid | / | SDS-Page | / | / | [47] |
| S. aureus | AT-27 AT-33 AT-36 AT-49 |
DNA | α-toxin | / | Neutralization | / | / | [143] |
| S. aureus | H1 H2 |
DNA | Whole cell | / | Fluorescence | 4-8 CFU/mL | 45–4.5 × 106 | [144] |
| S. aureus | Antibac1&2 | DNA | Peptidoglycan | 415 + 0.047 1261 + 0.280 |
Photoelectrochemical sensor | 82 pg/mL | / | [48,145] |
| E. coli | SH-Apt1 | DNA | Whole cell | 25,2 | Surface-enhanced Raman scattering (SERS) | / | / | [55,123] |
| E.coli | / | DNA | Whole cell | / | Optical | 45 CFU/mL | 102 -10e8 | [146] |
| E. coli | GN6 GN12 |
DNA | Outer membrane vesicles (OMV) | 29,94 20.36 59.70 24.80 38,98 53.83 |
Enzyme-linked aptamer assay (ELAA) | / | / | [147] |
| E. coli | 6-3 8-1 8-7 8-8 8-12 8-13 8-19 8-35 |
RNA | Heme | 188 309 256 371 445 425 |
Functional assay | / | / | [148] |
| E. coli | ECA I ECA II |
DNA | Outer membrane proteins (OMP) | / | Electrochemistry | / | 1*10-7–2*10-6M | [149,150] |
| E. coli | / | DNA | Whole cell | / | Plasmonic sensor | 0.05 CFU/mL | 0.1-104 | [151] |
| E. coli | Stx1 stx2 |
DNA | Shiga toxin Viz, stx1 & stx2 |
47 pM 29 pM |
/ | 44,5 pg/ml 41,3 pg/mg |
50 pg/ml – 100 ng/mg | [152] |
| P. aeroginosa | F23 | DNA | Whole cell | 17.27 ± 5.00 | colorimetric | 104 CFU/mL | / | [153] |
| B. cereus | / | DNA | Whole cell | / | Fluorescence | 22 CFU/mL | 49-49*106 | [154] |
| B. cereus | / | DNA | Whole Cell | / | Fluorescence | 4 CFU/mL | 20-2*108 | [155] |
| B. cereus | B15 B16 |
DNA | Whole cell | 16,13 20.67 |
Electrochemical sensor | 10 CFU/mL | / | [89] |
| B. cereus | 13-18 13-24 |
DNA | Whole cell | 22.75 36.72 |
Microfluidic chip | 9.27 CFU/mL | / | [156] |
| Acinetobacter baumannii | AB K2 |
DNA | Whole cell | 5.377 6.8 |
SERS | 10 CFU/mL | 10–105 | [157] |
| Acinetobacterer baumanni | / | DNA | Whole cell | / | Electrochemical sensor | 150 CFU/mL | 1*103–1.0*108 | [158] |
| Klebsiella pneumoniae | K2 | DNA | Whole cell | / | SERS | 10 CFU/mL | 10–105 | [157] |
| Leptospira interrogans | LAP3 | DNA | Outer Membrane protein | 133,13 | Colorimetric | 57 CFU/mL | 60–6*105 | [159] |
| Salmonella | Multi-apt | DNA | Multi | 11.72 | Fluorescence | 7 CFU/mL | 10-107 | [160] |
| Yersinia enterocolitica | / | DNA | Whole cell | / | Fluorescence | 3 CFU/mL | 10–109 | [161] |
| Bacillus cytotoxicus | BAS6R | DNA | Spore | / | Colorimetric | 102 – 104 CFU/mL | 103 - 104 | [29] |
| Vibrio cholerae | CT916 | Cholera toxin (CT) | 48,5 | ELAA | 2,1 – 2,4 ng/ml | 0-10 ng/mL | [43] | |
| Clostridium difficile | No name | DNA G-quadruplex |
Toxin A (TOA) | Electrochemistry | 1 nM | 0-200 ng/mL | [162] | |
| S. typhimurium | H2 | DNA | Whole cell | / | Fluorescence | 4-8 CFU/mL | 36–3.6 × 106 | [144] |
| S. Typhimurium | Apt S.T | DNA | Whole cell | 10 | Fluorescence | 30 CFU/mL | 102-106 | [163] |
| Vibrio parahaemolyticus | Apt VP | DNA | Whole cell | 16,88 | Fluorescence | 10 CFU/mL | 102-106 | [163] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



