
Submitted:
05 February 2024
Posted:
06 February 2024
You are already at the latest version
Abstract
Intestinal epithelial cell activities during homeostasis and regeneration are well described, but their potential interactions with stromal cells remain unresolved. Exploring the functions of these heterogeneous intestinal mesenchymal stromal cells (iMSCs) remains challenging due to the lack of specific markers for most functionally homogenous subpopulations. In recent years, however, novel clustering techniques such as single-cell RNA sequencing (scRNA-seq), fluorescence-activated cell sorting (FACS), confocal microscope, and computational remodeling of intestinal anatomy have helped identify and characterize some specific iMSC subsets. These methods help researchers learn more about the localization and functions of iMSC populations during intestinal morphogenic and homeostatic conditions. Furthermore, it is imperative to understand their cellular pathways for activation and how they interact with surrounding cellular components, particularly during intestinal epithelial regeneration that follows mucosal injury. This review provides insights into the spatial distribution and functions of some identified iMSC subtypes in intestinal morphogenesis, homeostasis, and regeneration. We reviewed related signaling mechanisms implicated during epithelial and subepithelial stromal cell crosstalk. Future research should focus on elucidating the molecular intermediates of these regulatory pathways to open a new frontier for potential therapeutic targets that can alleviate intestinal mucosa-related injuries.
Keywords:
Intestinal stem cell niche
; mesenchymal stromal cells
; sub-epithelial gradient factors
; epithelial-mesenchymal interactions
1. Introduction
The continuous self-renewal of the intestinal stem cells (ISC) underpins the high turnover rate of differentiated epithelial cells [1,2,3,4]. Over a decade ago, Hans and colleagues demonstrated how ISCs self-renew and proliferate into progenitor cell populations before differentiating into specialized epithelium cells [5,6]. From their study, we understand that stem cell division is confined to the crypt. In contrast, proliferating and differentiated cells form single-layered monolayers of different cell types arranged from the crypt base toward the apical villi region [5,7,8]. The proliferating progenitor cells in the transit amplifying (TA) zone undergo asymmetrical cell divisions, such that the half-daughter cells differentiate into specialized lineages, such as the secretory (Paneth, tuft, goblet, and enteroendocrine cells) or the absorptive enterocytes that migrate and form the single epithelial layer of the villi compartment [9]. This crypt-villus axis is characterized by distinct cellular interaction and molecular signatures that maintain the homeostasis [10,11,12]. For example, previous studies demonstrated that Paneth cells and subcryptal stromal cells produce canonical Wnt ligands to maintain the stem cell niche [13,14,15,16,17]. Likewise, stromal cells close to the sub-villi have been shown to communicate with the differentiated cells by providing BMPs and non-canonical Wnt ligands [18,19,20].
During regeneration after mucosa injury, reserve stem cells are activated, and specialized cells dedifferentiate into active stem cell populations that proliferate to support epithelial regeneration in the damaged site [21]. Previous studies have suggested that injured intestinal epithelial cells activate subepithelial fibroblast via TGFβ to support its proliferation and migration. [22,23,24] However, the specific functions of various subepithelial stromal cell populations to epithelial regeneration remain unresolved [25,26]. On this account, this paper reviews the recently characterized subpopulations of iMSCs in mice and their interactions with epithelium, highlighting their role in intestinal epithelial morphogenesis, homeostasis, and regeneration.
2. Intestinal stem cell regulation
Neighboring epithelial and subepithelial cells secrete gradient factors that influence ISC fate. These cellular interactions maintain ISC stemness at the base of the crypt and aid intestinal cell proliferation and differentiation activities along the crypt-villus axis [4,27]. Though the specificity of active stem cells is debatable, they are marked by LGR5+, OLFM4, ASCL2, RNF43, SOX9, MSI1, and SMOC2, and are capable of self-renewal and replenish differentiated epithelial cells [9,11,27,28]. To regulate these ISC activities, Paneth cells are adjacently interspersed between ISCs and produce growth factors (Wnt, EGF, and Notch-associated ligands). For instance, Paneth cells as described by Sato, et al. [29] improved organoid formations. Yilmaz, et al. [30] also reported that caloric restrictions preserved ISC self-renewal, and their adaptation is likely coordinated by mTORC1 signaling in the neighboring Paneth cells. Despite their regulatory support to the ISC niche, debates remain on the contribution of these non-stem cell populations in the ISC niche. Paneth cell depletion did not significantly alter ISC fate in vivo [31], and their functions can be substituted with exogenous Wnt supplement in ex vivo enteroid culture [32]. Additionally, other epithelial cell populations also help maintain intestinal epithelial balance by providing cues, which are highlighted in Table 1. Specifically, non-active stem cells, found in +4 positions, were reported to replenish the active stem cell pool during injury recovery through YAP1-dependent transient expansion [33]. Likewise, Paneth cells in the small intestine or Reg4+ expressing cells in the colon secrete canonical Wnt factors (such as Wnt3, Wnt9b, and Wnt11) and growth factors (EGF) to support epithelial regenerations via Notch signaling activation [16,34,35,36,37]. Similarly, recent studies have shown that iMSCs play critical and indispensable roles in regulating intestinal morphogenesis, homeostasis, and regeneration by providing regulatory factors [38,39,40]. It is therefore vital to address how iMSCs regulate Wnt, EGF, and bone morphogenetic protein (BMP) signaling pathways that confine active stem cell renewal at the crypt base and differentiate progenitors into specialized cells [2].
2.1. Signaling pathways
2.1.1. Wnt pathway
ISC stemness, proliferation, and differentiation fate largely depend on ligand-mediated signaling processes. As demonstrated in previous studies, canonical Wnt (or Wnt/β-catenin) signaling is crucial for ISC self-renewal maintenance [44,45]. For example, knocking out Porcupine and Wntless, essential mediators of Wnt secretion, greatly reduced the ISC population [14,43,46]. Similarly, the adenoviral delivery of the Wnt signaling antagonist, Dkk1, inhibited intestinal proliferative marker, Ki67, and Wnt/β-catenin target genes, CD44 and EpbB2 [47]. These studies confirmed the critical roles of Wnt signaling in intestinal homeostasis [2].
Notably, the canonical Wnt/β-catenin activation, regarded as a major promoter of ISC stemness maintenance, is triggered by ligands (e.g., Wnt2b, Wnt3, etc.) secreted from the ISC’s neighboring cells. First, the Porcupine protein synthesized in the endoplasmic reticulum of the neighboring cells enables Wnt ligand secretions that bind on the Frizzled receptors of the ISC plasma membrane to simultaneously attach to the LRP5/6 co-receptors [48]. This Wnt ligand-receptor binding initiates Dsh protein recruitment, leading to LRP receptor phosphorylation and the disruption of the Axin-GSK3-CK1 complex that orchestrates the β-catenin ubiquitination [2,11]. As a result, β-catenins are stabilized and accumulated in the cytoplasm. They subsequently translocate into the ISC’s nucleus to interact with LEF/TCF factors, thereby promoting canonical Wnt target genes transcription as illustrated in Figure 1. Blocking Wnt ligands may compromise Lgr5+ stemness and induce premature lineage differentiation [49]. This is partly due to the essential role of Wnt in ISC stemness maintenance. In contrast, the expressions of Wnt signal-related promoters such as Wnt2b and Rspondins (Rsops), and Wnt target genes increased the Lgr5+ population and ISC diminished as the proliferating cells migrated apically in the transit-amplifying region [38].
Unlike the canonical Wnt ligands that support intestinal stemness via β-catenin stabilization by orchestrating Wnt target gene transcriptions, non-canonical Wnt pathways are β-catenin independent [50]. Non-canonical Wnt pathways regulate the Wnt/PCP (planar cell polarity) and Wnt/Ca2+ dependent signaling cascades to modulate the ISC fate [51,52,53]. Despite the extensive knowledge surrounding canonical Wnt signaling, the function of non-canonical Wnt signaling has not been thoroughly studied. Non-canonical Wnt signaling, triggered by non-canonical Wnt ligands, promotes cellular differentiation and regeneration to facilitate ISC migration. Non-canonical Wnt ligands, including Wnt4, Wnt5, Wnt5b, and Wnt16 were remarkably expressed along the villus axis to the tip [20,38]. Their presence indicates that some Wnt ligands support villus differentiation and homeostasis, which might complement BMPs that promote intestinal maturation [20,54]. Recent scRNA sequencings have identified some iMSCs clusters, that are abundant in the villus sub-region, express Wnt4, Wnt5, Wnt 16, and Egf [20,54]. This paradoxical revelation contradicts the classical roles of Wnt in maintaining the stemness [11,55]. For instance, ablation of villus tip subepithelial Lgr5+ iMSCs, which expressed high Wnt5a and Bmp4, resulted in enterocyte loss [54,56]. Additionally, the Wnt/Ca2+ pathway can independently stimulate stem cell proliferation when Wnt5a/Fzd6 interactions increase intracellular Ca2+ levels in the cytosol during gastric cancer. This leads to PKC and CAMK-mediated downstream cascades to promote cell migration [53,57,58]. In a related pattern, Wnt ligands could bind and activate Rho receptor complexes (Ror/Ryk) to trigger c-Jun N-terminal kinase, initiating JNK target gene transcription [57,59]. JNK target genes participate in cellular proliferation, migration, and regeneration by targeting exoskeleton proteins and adhesion molecules such as actin and Rho GTPases. These suggest that non-canonical Wnt signaling downstream could be a promising target for pharmaceutical drugs to improve intestinal injury recovery [60].
2.1.2. BMP pathways
The bone morphogenetic protein (BMP) pathway is an important signal opposing the Wnt mechanism that stimulates epithelial differentiation. Contrary to canonical Wnt signaling, BMP/Smad activities increase toward the apical region, indicating repressed stem cell activities along the villi [61,62,63]. An extensive review has addressed BMP/Smad regulation of intestinal epithelial differentiation and homeostasis [63]. The focus here provides insight into how Bmp factors mediate the interaction between the intestinal epithelium and iMSCs. Of note, BMP4, a ligand stimulating Bmp signaling, was reported as a transcriptional target gene of hedgehog-responsive iMSCs [61,63,64,65]. The interaction between iMSCs and epithelium has complex roles in establishing the distinct identities of the crypt and villi by supplying Wnt and Bmp gradients [62,65,66,67]. A recent study found that populations of PDGFRα+ iMSCs promote Wnt signaling, which can stimulate the expression of epithelial hedgehog ligands (SHH and IHH) during morphogenesis. [20]. In turn, epithelial-derived SHH ligands directly activate iMSC target gene transcriptions, including BMPs, that shape the developing intestinal villus formation [20,44,64,66,68]. Interestingly, sub-epithelial iMSC provides essential ligands that inhibit BMP activities in the crypt base where stem cell renewal and proliferation are at their peak. For instance, sub-cryptal iMSCs secrete Noggin and Gremlin1 to block BMP signaling, thereby supporting the ISC niche [20,69]. This means that epithelial-mesenchymal communications create a feedback loop to maintain intestinal homeostasis along the crypt-villus axis.
2.1.3. Other cellular signaling pathways
Subsequent to active ISC proliferation, cell lineage specialization fates are intricately regulated by the Notch pathway. Notch signaling actively commits progenitor cells to transform into differentiated specialized cells [36,70]. Moreso, increased Notch ligands from neighboring Paneth cells redirected proliferating cells toward enterocyte lineage by repressing Math1 transcriptions [7,71]. Paneth cells, which is a secretory lineage, could trigger negative feedback to limit secretory cell lineage commitment, thereby supporting the importance of stringent regulations of ISC activities to maintain intestinal epithelial homeostasis. Notwithstanding, other mechanistic pathways, such as EGF, contribute to ISC maintenance and are well documented in previous studies and reviews [2,11,27].
Another notable mechanism that regulates intestinal cellular interactions is the Hedgehog (HH). HH ligands secreted by epithelium initiate negative regulation of its receptor (Ptch1) on neighboring iMSCs [65,72]. In the absence of HH ligands from epithelial cells, Ptch1 inhibits Smo signaling transduction, leading to Gli family phosphorylation in the cytoplasm by degradation complex including GSK3β, CKIα, and PKA. In contrast, when HH ligands bind on Ptch1 receptors, they release Smo to activate STK36, thereby inhibiting the degradation complex assembly. STK36 also phosphorylates SUFU complex to stabilize Gli and allow nuclear accumulation, which subsequently stimulates Gli-dependent target gene transcriptions Figure 2. Despite the extensive review on Hedgehog regulations, especially their intricate functions in fetal villigenesis and regenerations [20,45,65,69], their roles in promoting crypt stemness during homeostatic conditions are just gaining attention in recent studies [9,66,73]. This was best described when hedgehog-responsive mesenchymal cells were revealed to constitute a key colonic stem cell niche [73]. Consistently, the mesenchymal subsets that are localized in sub – and peri-cryptal regions form the main sources of the Wnt, Rspo, and Grem niche that supports the crypt stemness [38,39]. For example, activated HH pathway upregulated stromal Wnt expression which contributes to increased OLFM4 positive stem cell pool [74]. Likewise, Current data also suggest HH/Wnt pathways crosstalk potentially promote intestinal regeneration. Upon irradiation-induced intestinal epithelial injury, SHH was significantly upregulated, leading to increased production of Wnt ligands (Wnt2b, Wnt4, and Wnt5a) in underlying stromal cells – resulting in enhanced regeneration [67]. Together, these signaling networks complementarily or in opposing efforts regulate intestinal cell fate along the crypt-villi topologies during intestinal homeostasis and injury repair processes.
3. Intestinal cell plasticity and regeneration
The intestinal epithelium is a delicate but resilient organ. It undergoes rapid turnover of epithelial cells, predisposing it to luminal insults [75]. Epithelial cell exposure to luminal contents can cause severe damage induced by physical or pathogenic agents [21]. These perturbations necessitate addressing the concept of intestinal cell plasticity. While active Lgr5+ stem cells continuously proliferate to replenish epithelium during normal homeostatic conditions, the proliferating properties make them more susceptible to mucosal injuries, including irradiation or inflammation [76]. Studies showed that various mucosal injuries, such as inflammation, hypoxia, or irradiation caused Lgr5+ stem cell loss, thereby exacerbating intestinal epithelium regenerations [77,78,79,80]. Though the Lgr5+ stem cell pool is depleted during intestinal injury [78], another stem cell population (known as reserve stem cells) is activated to replenish the active stem cell pool [37,81,82]. This population of non-active Lgr5- ISC plays a crucial role in regeneration following intestinal mucosal damage and helps maintain intestinal homeostasis [33,83,84,85]. For example, both DSS (Dextran sulfate sodium) and irradiation-induced Lgr5+ cell loss caused the proliferation of quiescent Clu+ expressing cells to reconstitute columnar base Lgr5+ cells and replenish damaged epithelium during regenerations [33]. To buttress this idea, a previous study reported that the transitory loss of Lgr5+ stem cells did not disrupt intestinal architecture, and this could result from the reserved stem cells recruited following Lgr5+ stem cell loss [78,82]. This hypothesis corroborates the activation of putative reserve stem cells, including Bmi1, Hopx, and mTert, during Lgr5+ cell depletion [82,83,86,87]. It is suggested that quiescent cells are resistant to mucosal injury and play critical functions during the regeneration process [82,83]. Reserve stem cell populations are less proliferative and express high anti-apoptotic and DNA repair genes [33,88]. Though these properties make them a great candidate to replenish the active stem cell pool after mucosal injury [89,90], researchers still debate whether reserve stem cells at the +4 position of the crypt are responsible for regeneration [28].
Others proposed that several differentiated epithelial cells undergo fetal-like reprogramming and revert to active Lgr5+ cells during intestinal regeneration [23,24,67]. Secretary lineage seems to be the most prominent among the differentiated cell types that support intestinal regeneration during mucosal healing [91]. The dedifferentiation by these non-putative active stem cells to replenish the Lgr5+ pool is known as intestinal plasticity [28,82,92]. In the last decades, lineage tracing techniques using transgenic animals have provided the model to delineate cell migration [5,7]. This technique can track the specific cell lineage by editing the gene of interest. Van Es, et al. [42] explored this model to reveal that Dll1+, a secretory cell progenitor, can establish organoids containing Lgr5+ cells in in vitro culture. In a similar pattern, when irradiation depleted the stem cell pool in vivo, Dll1+ transformed into active Lgr5+ stem cells that proliferated into multiple progenitor lineages [42]. Another secretory progenitor cell, Atoh1+, was confirmed to generate Lgr5+ stem cells using different injury models such as irradiation and DTR (Diphtheria toxin receptor)-induced stem cell loss strategies [93]. To clarify the mechanisms behind the dedifferentiation phenomenon, several studies have tried to unravel the factors associated with cell plasticity and regeneration, but this area is still under active investigation. Recently, some studies suggested potential underlying mechanisms involved in the processes [21,28]. For instance, recombinant WNT3A supplements promote Dll1+ to produce organoids populated by Lgr5+ cells [94]. Furthermore, Ascl2, a Wnt target gene, was found to promote Paneth cell dedifferentiation following the DSS treatment [84]. According to Yu, et al. [95], Notch signaling target genes, particularly Hes1 and Notch1, are upregulated during Paneth cell dedifferentiation in irradiation-induced damage studies. Likewise, Yap1, a key hippo signaling target gene, was significantly upregulated in a DSS-induced study during epithelial regeneration [96]. Altogether, the crosstalk among Wnt, Notch, Hedgehog, and Hippo signalings needs further studies which will be relevant to understand their contributions to epithelial regeneration [2,97]. Unraveling these mechanisms could provide key insights into therapeutic targets to alleviate mucosal healing.
Non-epithelial cells, such as immune and iMSCs, also contribute to intestinal cell plasticity and regeneration. These cells secrete factors, including growth factors and cytokines, to stimulate mechanistic signalings that regulate regenerative processes [43,98,99]. They sense and are recruited to respond to injury repair. Immune cells specifically induced inflammation, and also recognized damaged associated molecular patterns released by apoptotic cells [100]. Besides immune cells, iMSCs are essential sub-epithelial cell types that are underappreciated during regeneration responses. Injured epithelial cells can send signals to iMSCs through hedgehog ligands, specifically Ihh and Shh, thereby enhancing ISC regenerations to support repair processes after epithelial injury [101,102,103]. Elevated levels of these ligands increased HH target gene transcription in iMSCs, including Cyclin D1, to promote epithelial regeneration [45,72,73]. Recent studies are unraveling potential iMSC subsets that are involved in intestinal epithelial repair functions. For instance, a study demonstrated that Gli+-expressing mesenchymal cells may secrete Rspo3 to support epithelial repair processes using the DSS-induced damage model [73]. Moreover, DSS-induced colitis increased fgf10, Vegf, Wnt2b, Grem1, and Rspo1 expressions in CD34+ cells, corroborating iMSCs roles during epithelial repair responses [45]. In conclusion, depleted active stem cell populations are certainly replenished during epithelial injury repair and regeneration, but there is still a knowledge gap on iMSC roles in supporting stem cell regenerative capacity. Understanding intestinal epithelial and MSC interaction could be the key target to treating relapsing intestinal diseases, such as inflammatory bowel diseases and necrotizing enterocolitis. Thus, it is important to elicit the contribution of multicellular crosstalk during intestinal cell plasticity and regeneration programs. We will discuss more details on the role of iMSCs during regeneration in the next chapter.
4. Subepithelial mesenchymal stromal cell characteristics
The intestinal mesenchymal stromal cells (iMSCs) are among the cell population localized in the lamina propria. The term “iMSCs” represents a group of non-epithelial, non-endothelial, non-neuronal, and non-hematopoietic cells that contribute growth factors and chemokines to regulate the intestinal epithelial homeostasis, regeneration, and immune functions [104,105,106]. These cells share surface markers with mesenchymal cells found in other tissues, giving them similar characteristics [107,108]. They were previously categorized by non-expressing α-smooth muscle actin (α-SMA-) fibroblast, α-SMA+ expressing myofibroblast, pericytes, and mesenchymal stem cells [55,66,107]. Before recent evidence, iMSCs were largely focused on their structural support functions to the epithelial architecture, as they form the largest component of the extracellular matrix (ECM) of the intestine [106]. Contrary to these past views, recent studies using immunostaining and single-cell RNA sequencing have revealed iMSCs as heterogeneous cell populations capable of dynamic multidirectional crosstalk with epithelial, hematopoietic, and immune cells [39,43,109]. Accordingly, iMSCs are dispersedly distributed along the crypt-villi sub-epithelial region in the lamina propria, muscularis mucosa, and deep in the submucosa with distinct morphology and functional support to epithelial cells.
4.1. Roles of intestinal mesenchymal stromal cells during prenatal intestinal morphogenesis
Intestinal epithelium develops from endodermal cells in the embryo [68]. During the early fetal stage, these cells in tube-like sheet layers invaginate to form pseudostratified epithelium (E9.5). This means that the proliferating epithelial cells are non-compartmentalized at the early embryonic stage [62,69,110]. Interestingly, cellular proliferation during early embryonic development survives in the absence of Wnt/β-catenin signaling despite the crucial role this mechanism plays in adult stem cell homeostasis [111]. Wnt target gene transcription factor (TCF) was found to be redundant during the pseudostratified epithelial development stage. Rather, mice lacking the Tcf7l2 gene had significant epithelial shortening only after villi had been formed completely [112]. In another study, conditional β-catenin deletion did not affect pseudostratified epithelium proliferation during the early fetal stage [113]. Together, the studies indicate that there might be distinct signaling mechanisms between early-phase and late-phase embryogenesis.
The pseudostratified intestinal epithelium and mesenchyme rapidly increase from E9.5 to E14.5. By E14.5, the pseudostratified epithelium undergoes extensive remodeling into columnar epithelium, which leads to the emergence of villus structure. To gain insights into the underlying factors that potentially shape fetal gut compartments at this stage, Maimets et al. reported that CD29 and PDGFRα+ cells are crucial for fetal gut vilification [20,46,69,111,114]. The CD29+ cells expressed high Acta2, Myl4, Des, and LRIG1 levels, which could be the progenitor for fetal gut muscularis mucosa cells. More importantly, they revealed how PDGFRα+cells guide the villigenesis of the pseudostratified epithelium [20,62]. This suggests that PDGFRα+cells could be a progenitor for stromal lineages because of their indispensable contributions to villi formation during morphogenesis, which aligns with previous studies that demonstrated iMSCs drive intestinal epithelial fate [20,66,115,116]. They hypothesized that PDGFRα+cells are crucial for villi emergence because they form clusters close to the expanding epithelium. This view is supported by their in vitro model study, which showed that PDGFRα+ cells isolated from the fetal gut can promote organoid growth independent of essential growth factor supplements. In contrast to the early fetal stage, they also found that inhibiting Porcn in pregnant dams (a Wnt ligand upstream regulator) between E12.5 and E16.5 had no effects on the fetal colon, but abrogated SI villi formation. Based on these data we could deduce that Wnt gradients become relevant from mid-stage morphogenesis onward for maintaining the crypt-villus compartment. To corroborate this hypothesis, other studies have also highlighted that the underlying mechanisms of early embryonic morphogenesis are biologically different from those that regulate postnatal intestinal epithelium [113,117].
Embryonic mice at E14.5 exhibit rapidly expanding epithelial cells. By E16.5, these proliferating cells are restricted to the intervillous domain between neighboring villi and differentiated absorptive and secretory cells in villi [68]. A previous report suggests that embryonic iMSCs expressing Dlk1 support rapid expansions of fetal gut morphogenesis, which differs from their role in adult mice [117]. To confirm this, PDGFRα+ cells, which are the dominant iMSC cluster in the late fetal phase, reportedly expressed DLK1, but this expression decreased toward late gestation [117]. For these reasons, the current challenge is to better understand the PDGFRα+ subset functional diversities as recent studies classified them as PDGFRαhi, PDGFRαmedium, and PDGFRαlo cells during morphogenesis [20,44]. These subsets showed distinct characteristics and potentially have unique signaling properties.
Studies have previously predicted the indispensable roles of hedgehog signaling, involving SHH upregulation during villi formation [20,62,68]. Consistently, Hedgehog target genes, especially Ptch1, were detected in the absence of Wnt signaling. Ptch1 is a key receptor, expressed by mesenchyme, for the SHH ligand, indicating that mesenchymal cells are indispensable during fetal gut formation. While the roles of non-Wnt pathways in fetal intestinal patterning during embryogenesis remain unclear, the available data suggest multiple signaling crosstalk such as Wnt, hedgehog, and Notch, among other factors produced by epithelial-mesenchymal cells regulate fetal gut morphogenesis [20,113].
4.2. Roles of intestinal mesenchymal stromal cells during intestinal homeostasis
The neonatal and mature intestinal epithelium is supported by heterogeneous stromal cell populations [38,39,44]. By the time of birth, the neonate gut will transition into a compartmentalized intestine driven by polarized signaling mechanisms. Specifically, the crypt and Paneth cells in the small intestine will start to emerge by the 14th postnatal day. By postnatal day 28, crypts would have rapidly expanded and matured, forming the intestinal epithelial crypt-villus structure [68,118,119]. Adult mice maintain intestinal homeostasis through several cellular programming, including ISC stemness, TA proliferation, and differentiation [9]. Each process requires unique gradient factors to keep the intestinal compartments in normal condition, in which iMSCs serve as a crucial source for all the essential ligands required for regional specificity of epithelial cell integrity. Remarkably, iMSC subtype heterogeneity increases as gut maturation progresses during postnatal development [39,44].
4.2.1. Recent classification of iMSC lineage
Although there has been substantial progress in characterizing epithelial cells [21], subepithelial iMSCs classifications, their cellular diversities, and functionalities remain challenging due to a lack of unique surface markers. Nevertheless, they are broadly categorized according to their regional specificity and functional diversity along the intestinal region using immunostaining, FACS, and transgenic mouse models. iMSCs expressing PDGFRαhi, ACTA2lo myofibroblast, FOXL1+, and GLI1+ but not vascular CD31+ markers are loosely regarded as telocytes due to their proximity to epithelial cells in the villus region [69,120,121]. Most cryptal iMSCs uniquely express CD34+/GP38+ co-localizing with CD81+ and other PDGFRαlow cells, forming sub- and peri-cryptal stromal populations [44,45,46,69]. The subcryptal stromal populations produce trophic factors to support intestinal stem cell niche, and they are distinguished from muscularis mucosa cells due to their lack of Myh11+ gene expression [43]. Despite being isolated and studied, those cells still display heterogeneity and remain undifferentiated in terms of their origin and possible lineages.
Recent studies using scRNA-seq assay have revealed a robust description of possible lineage, anatomic and physiologic heterogeneity of previously obscure iMSC subtypes [38,39,43]. One common proposition is that adult iMSC subsets originate from a similar embryonic precursor identified as Gli-expressing cells, which is still debatable [39]. For instance, most adult iMSCs express PDGFRα+ at varying magnitudes which suggests PDGFRα+ as another potential progenitor – more details on how iMSCs support embryogenesis are presented in the morphogenesis section [20,62]. Their localization probing and gene expression profiling showed a diverse population of iMSCs despite sharing the same lineage.
iMSC subsets that are sparsely localized beneath the epithelium produce gradient factors that shape the crypt/villi compartments. To appreciate the spatial distributions of these subsets, we reviewed recently characterized iMSCs and schematically illustrated them in Figure 3. These studies revealed distinct iMSC subsets that are classed under PDGFRαlo (CD34hiCD81+, CD34hiIgfbp5+, CD34loFgfr2+) and PDGFRαhi (CD9hiCD141-, CD9loCD141+, CD141int) [39,66,122].
Functions of pericryptal (PDGFRαlo or CD34+Gp38+) subpopulations
PDGFRαlo subsets are found beneath the muscularis mucosae (CD81+), pericryptal (Igfbp5+), and the lamina propria (Fgfr2+). This idea is strengthened by the data reported in recent studies [38,39]. The studies suggested that gradients regulating ISC niche and terminal differentiation support the intestinal epithelial regional specificity.
To confirm the functional characteristics of these PDGFRαlo subsets, clusters of PDGFRαlo cells that expressed pericryptal CD34+Gp38+ only emerged after birth and they are found to promote intestinal stemness [45]. The study demonstrated that CD34+Gp38+ cells are the main Wnt ligand contributors to adult ISC homeostasis when the crypt is formed. This indicates that the intestinal epithelium requires a specific iMSC sub-population to reach maturation postnatally [45]. The idea corresponds with data showing that CD81+ cells located in the muscularis mucosae, a subset of the CD34+ population, are the key producer of Grem1 that inhibits Bmp activities in the crypt [39,45]. PDGFRαloCD81+ Ackr4+ (trophocytes) and PDGFRαloCD81-CD55hi cells are subsets of the CD34+ population that produce BMP antagonists for stem cell niche to promote ISC stemness near the crypt base [38]. The Shivdasani group also reported that PDGFRαloCD81+ cells expressed high Wnt2b, R-spos, and Grem1 levels [55,66,104]. The depletion of Rspo3-secreting iMSCs that caused delayed gut maturation and reduced ISCs buttresses the functional specificity of PDGFRαloCD81+ cells [46,49,123].
The cryptal cells transitioned into differentiated cells just above the villus base. iMSCs that are found beneath the villus base and the corresponding lamina propria region produce pro-BMP factors [38,39]. For example, PDGFRαloCD81-CD55lo and CD34loFgfr2+ cells expressed non-canonical Wnt4 near the top of the colon crypt base, suppressed trophic factor effects, and also reinforced the BMP gradient activities, suggesting their roles in promoting terminal differentiation in the TA domain toward the villus region. Though they are PDGFRαlo subsets, their transcriptomic data revealed they expressed a low level of Wnt2b. This is consistent with previous studies that showed BMP gradients increased apically in the SI villi or the colon crypt top [38,66]. In contrast, BMP inhibitors increased distally, especially from pericryptal PDGFRαloCD81- cell to PDGFRαloCD81+ trophocytes beneath the muscularis mucosae [39,44,45]. Though it is currently difficult to sort, culture, and investigate all of these distinct iMSC sub-populations, studies have shown that they may functionally overlap, express related molecular profiles to perform complementary roles, and in some cases, provide opposing cues to maintain intestinal homeostasis and development (Table 2). Collectively, the current observations depict that epithelial development relies heavily on gradients from diverse iMSCs to maintain homeostasis despite the high turnover rate of epithelial cells [104,124,125].
Functions of PDGFRαhi subpopulations
PDGFRαhiFoxl1+ cells are reported to be a good source of BMP-promoting factors, including Bmp4, Bmp5, and Bmp7 [38,39,45,66]. Paerregaard et al. reported that the three PDGFRαhi subsets expressed Foxl1. 1) Nrg1 was expressed in CD9hiCD141-; 2) Cxcl12, and Acta2 expressed in CD9loCD141+; and 3) Adamdec1, Wnt4, and Acta2+ were expressed in CD141int cells [39]. While all these PDGFRαhi subpopulations produce BMP (Bmp5, and Bmp7) gradients, CD9loCD141+ had the highest levels of Wif1, Bmp3, and Bmp4, contributing to terminal differentiation of intestinal epithelial progenitor cells. Upregulations of Wif1, a protein that binds and inhibits canonical and non-canonical Wnt ligands (such as Wnt3a, Wnt4, and Wnt5a) [127], corroborated the hypothesis that Wnt signaling activities are repressed from the villus base toward the tip. Interestingly, using RNA velocity analysis, these three PDGFRαhi subsets were predicted to originate from CD34loFgfr2+ found along the lamina propria of the small intestine in mice.
According to a recent study, post-natal intestinal epithelium required PDGFRα+ cell-dependent maturation to shift from immature proliferative compartments into distinctly functional crypt-villi regions [44]. In their study, they generated reporter and inducible lineage tracing models for lymphotoxin beta receptor (LTβR) cells by crossing LTβRtTA mice with Rosa26floxSTOP-YFP mice. Using the inducible lineage tracing technique, some fractions of PDGFRα+ subepithelial stromal cells developed from LTβRYFP progenitor cell lineage before the mice reached weaning age. To confirm this hypothesis, they also generated a direct LTβRGFP reporter model to show that PDGFRαhi cells, found close to the villus epithelial cells, expressed GFP [44]. Additional transcriptomic analysis revealed that LTβR+PDGFRα+ expressing cells showed upregulated levels of hedgehog and pro-differentiation gene markers that are specific to PDGFRαhi expressing cell population, especially Ptch1, Foxf2, Gli1, Bmp4, and Bmp5. In contrast, the pro-stem cell niche gene markers (i.e., Cd34, Cd81, Rspo2/3, and Grem1/2) were downregulated in the transcriptome data of LTβR+PDGFRα+ cells [39,44,66]. Inducible conditional ablation of PDGFRα in LTβR lineage (LTβRΔPDGFRα mice) caused increased ISC markers (Olfm4+ and Lgr5+) in the gut compared with the wild type. Consistently, LTβR stromal lineage from LTβRΔPDGFRα mice had decreased pro-differentiating factor (specifically, Bmp2) and increased pro-stemness factor (Grem2 and Chrdl1) levels in their stromal cell transcriptome data. Collectively, these observations demonstrated in their studies suggest that the LTβR+PDGFRα+ cells expressing pro-differentiation signals share similar characteristics with the PDGFRαhi subsets reported by Paerregaard et al., and they are essential for intestinal epithelial maturation before weaning [39,44,45]. More importantly, their results showed that the appearance of PDGFRαhi cells not only influences epithelial cell differentiations but is also essential for the transcriptomic switch of functionally distinct LTβR+ stromal lineage cells towards maturation and localization during early postnatal gut development [44]. Accordingly, different iMSC subsets create a functionally distinct enabling environment for polarized signaling crosstalk to regulate epithelial cell fate during development. To sum it up, iMSC populations are less diverse during intestinal morphogenesis but become heterogeneous postnatally to support mature intestinal epithelial homeostasis [38,39,43,69].
4.3. Roles of intestinal mesenchymal stromal cells during intestinal injury and repair
iMSCs are not only crucial for intestinal morphogenesis and homeostasis; recent reports have also revealed their modulatory functions using DSS, irradiation, and DTR-induced injuries in transgenic mice. For example, RBP1+ cells, confirmed to be a subset of GLI1+ cells, are suggested to sense DSS-induced colitis injury, thereby stimulating Rspo3 production to promote the injury recovery process [73]. Likewise, conditional deletion of the Wntless allele in a Villin-WlscKO, a protein required for Wnt secretion, triggered GLI1+ cell expansion during injury. This observation was confirmed using recombinant Shh ligand to promote GLI1+ cell response to injury by compensating for epithelial Wnt loss. The results suggested that SHH signaling could be a potential regulatory target for enhancing mucosal healing by activating iMSCs. In another study, pericryptal CD34+ cells, in response to DSS treatment, were reported to migrate and localize under the colonic crypt–apical epithelial cells [128]. Following the induced epithelial injury, the mRNA data revealed upregulated levels of Bmp2, Bmp3, Bmp7, and Wnt5a, indicating the plasticity potential of CD34+ to support reepithelization during colon regeneration [128].
According to the reviewed studies above, mesenchymal cells support epithelial regeneration by secreting growth factors and other signaling molecules that promote the proliferation and differentiation of epithelial cells. scRNA seq analyses showed that iMSCs are intricately involved in epithelial cell response to injury recovery processes. iMSCs such as CD81+ and CD81- subsets play significant roles during injury states, by increasing the production of Wnt-promoting factors [45,73]. For instance, Grem1 and Rspo3 reportedly increase in PDGFRαlo cells during colitis-induced conditions, suggesting that these iMSCs support epithelial regeneration [116]. In another study, non-coding RNAs (miR-143/145) specific to iMSCs were also involved in regulating IGF1 signaling response to repair DSS-induced epithelial damage [129]. In all, iMSCs are essential for the repair and regeneration of the intestinal epithelium following injury. However, more studies are needed to clarify the relevance of specific iMSC subsets that engage in different disease resolution processes.
5. Conclusion and Future Perspectives
The iMSC populations play a crucial role in maintaining intestinal morphogenesis, homeostasis, and regeneration by providing support and signaling to the epithelial cells. Recent studies have shown that iMSCs belong to different subsets that have distinct regulatory functions along the crypt-villi axis. However, several obstacles need to be resolved to fully understand their specific characteristics, functions, and responses during homeostasis and epithelial regeneration. To advance our understanding of iMSCs, three critical areas require immediate attention.
Firstly, researchers need to develop specific markers to further characterize homogenous iMSC subsets within clusters of notable iMSC subpopulations addressed in this paper. This approach will help to address the conflicting results on the iMSC population that have been reported to provide opposing signaling factors they secrete at distinct locations along the crypt-villi axis. Improved iMSC sub-type sorting techniques would clarify the unique functional diversity of specific homogenous iMSCs and how each distinct subset interacts with different epithelial cells. As such, researchers will be able to define the contributions of the overlapping molecular signatures by the heterogeneous iMSCs clusters during normal and regeneration conditions. Thus, this is a call to develop novel iMSCs surface markers for cell sorting assays which will advance the current knowledge about epithelial and mesenchymal crosstalk.
Second, to unravel the roles of characterized iMSC subsets during morphogenesis and disease development, researchers should improve lineage tracing techniques that focus on examining the response of iMSCs to regeneration. For example, iMSCs are currently proposed to have multidirectional relationships with both epithelial and immune cells. While iMSCs supply Wnt and Bmp agonists during normal conditions, it remained obscure whether (1) they directly provide regenerative factors to repair epithelial cells, (2) adopt homing effects to replace damaged epithelial cells, or [41] indirectly activate anti-inflammatory immune cell responses to restore intestinal homeostasis after injury. Future research directions in this field could include investigating the mechanisms by which subepithelial stromal cells regulate stem cell behavior and the role of these cells in chronic diseases of the intestine, such as inflammatory bowel disease. Additionally, understanding the interactions between subepithelial stromal cells and other cell types may lead to the development of novel therapeutic strategies for promoting intestinal regeneration and repair.
Finally, despite the difficulty relating to investigating functional studies in vivo due to their delicate nature, the future approach needs to improve scRNA seq techniques, develop special iMSCs culture conditions that can support the viability of FACS cells, and improve co-culture models to better understand the complexity of mesenchymal and epithelial crosstalk.
In conclusion, this paper has provided a comprehensive review of the recent evidence about iMSCs heterogeneity, suggesting their spatial distribution which enables them to perform distinct or overlapping functions in maintaining epithelial integrity. Future studies should reconstruct 3D organoid co-culture set-ups, such as transwell and scaffold models, to elucidate the spatial organization of iMSCs in the intestine for different developmental stages and address potential communications between epithelial and sorted iMSC subsets. By addressing these challenges, we can gain a better understanding of the complexity of the iMSC niche and develop novel therapeutic strategies for promoting intestinal regeneration and repair.
Author Contributions
HA: conceptualization, image design, original writing draft, revision; EML: revision; YJ: conceptualization, image design, revision, funding.
Acknowledgments
The authors appreciate the financial support from the Maryland Stem Cell Research Fund and MAES Competitive Grant Program at the University of Maryland, College Park, USA. We used Biorender to illustrate the images presented in this paper.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Corominas-Murtra, B.; Hannezo, E. Modelling the dynamics of mammalian gut homeostasis. In Proceedings of the Seminars in Cell & Developmental Biology; 2022. [Google Scholar]
- Gehart, H.; Clevers, H. Tales from the crypt: new insights into intestinal stem cells. Nat Rev Gastroenterol Hepatol 2019, 16, 19–34. [Google Scholar] [CrossRef]
- Guiu, J.; Hannezo, E.; Yui, S.; Demharter, S.; Ulyanchenko, S.; Maimets, M.; Jorgensen, A.; Perlman, S.; Lundvall, L.; Mamsen, L.S.; et al. Tracing the origin of adult intestinal stem cells. Nature 2019, 570, 107–111. [Google Scholar] [CrossRef] [PubMed]
- Mannino, G.; Russo, C.; Maugeri, G.; Musumeci, G.; Vicario, N.; Tibullo, D.; Giuffrida, R.; Parenti, R.; Lo Furno, D. Adult stem cell niches for tissue homeostasis. J Cell Physiol 2022, 237, 239–257. [Google Scholar] [CrossRef] [PubMed]
- Barker, N.; van Es, J.H.; Kuipers, J.; Kujala, P.; van den Born, M.; Cozijnsen, M.; Haegebarth, A.; Korving, J.; Begthel, H.; Peters, P.J.; et al. Identification of stem cells in small intestine and colon by marker gene Lgr5. Nature 2007, 449, 1003–1007. [Google Scholar] [CrossRef] [PubMed]
- Buczacki, S.J.; Zecchini, H.I.; Nicholson, A.M.; Russell, R.; Vermeulen, L.; Kemp, R.; Winton, D.J. Intestinal label-retaining cells are secretory precursors expressing Lgr5. Nature 2013, 495, 65–69. [Google Scholar] [CrossRef] [PubMed]
- Clevers, H. The intestinal crypt, a prototype stem cell compartment. Cell 2013, 154, 274–284. [Google Scholar] [CrossRef]
- van der Flier, L.G.; van Gijn, M.E.; Hatzis, P.; Kujala, P.; Haegebarth, A.; Stange, D.E.; Begthel, H.; van den Born, M.; Guryev, V.; Oving, I.; et al. Transcription factor achaete scute-like 2 controls intestinal stem cell fate. Cell 2009, 136, 903–912. [Google Scholar] [CrossRef] [PubMed]
- Beumer, J.; Clevers, H. Cell fate specification and differentiation in the adult mammalian intestine. Nat Rev Mol Cell Biol 2021, 22, 39–53. [Google Scholar] [CrossRef]
- Hu, D.; Yan, H.; He, X.C.; Li, L. Recent advances in understanding intestinal stem cell regulation. F1000Res 2019, 8. [Google Scholar] [CrossRef]
- Santos, A.J.M.; Lo, Y.H.; Mah, A.T.; Kuo, C.J. The Intestinal Stem Cell Niche: Homeostasis and Adaptations. Trends Cell Biol 2018, 28, 1062–1078. [Google Scholar] [CrossRef]
- Spit, M.; Koo, B.K.; Maurice, M.M. Tales from the crypt: intestinal niche signals in tissue renewal, plasticity and cancer. Open Biol 2018, 8, 180120. [Google Scholar] [CrossRef]
- Felsenthal, N.; Vignjevic, D.M. Stand by me: Fibroblasts regulation of the intestinal epithelium during development and homeostasis. Curr Opin Cell Biol 2022, 78, 102116. [Google Scholar] [CrossRef]
- Kabiri, Z.; Greicius, G.; Madan, B.; Biechele, S.; Zhong, Z.; Zaribafzadeh, H.; Edison; Aliyev, J.; Wu, Y.; Bunte, R.; et al. Stroma provides an intestinal stem cell niche in the absence of epithelial Wnts. Development 2014, 141, 2206–2215. [Google Scholar] [CrossRef]
- Miyoshi, H. Wnt-expressing cells in the intestines: guides for tissue remodeling. J Biochem 2017, 161, 19–25. [Google Scholar] [CrossRef]
- Sasaki, N.; Sachs, N.; Wiebrands, K.; Ellenbroek, S.I.; Fumagalli, A.; Lyubimova, A.; Begthel, H.; van den Born, M.; van Es, J.H.; Karthaus, W.R.; et al. Reg4+ deep crypt secretory cells function as epithelial niche for Lgr5+ stem cells in colon. Proc Natl Acad Sci U S A 2016, 113, E5399–5407. [Google Scholar] [CrossRef] [PubMed]
- Zou, W.Y.; Blutt, S.E.; Zeng, X.L.; Chen, M.S.; Lo, Y.H.; Castillo-Azofeifa, D.; Klein, O.D.; Shroyer, N.F.; Donowitz, M.; Estes, M.K. Epithelial WNT Ligands Are Essential Drivers of Intestinal Stem Cell Activation. Cell Rep 2018, 22, 1003–1015. [Google Scholar] [CrossRef] [PubMed]
- Bahar Halpern, K.; Massalha, H.; Zwick, R.K.; Moor, A.E.; Castillo-Azofeifa, D.; Rozenberg, M.; Farack, L.; Egozi, A.; Miller, D.R.; Averbukh, I.; et al. Lgr5+ telocytes are a signaling source at the intestinal villus tip. Nat Commun 2020, 11, 1936. [Google Scholar] [CrossRef] [PubMed]
- Bernier-Latmani, J.; Mauri, C.; Marcone, R.; Renevey, F.; Durot, S.; He, L.; Vanlandewijck, M.; Maclachlan, C.; Davanture, S.; Zamboni, N.; et al. ADAMTS18(+) villus tip telocytes maintain a polarized VEGFA signaling domain and fenestrations in nutrient-absorbing intestinal blood vessels. Nat Commun 2022, 13, 3983. [Google Scholar] [CrossRef]
- Maimets, M.; Pedersen, M.T.; Guiu, J.; Dreier, J.; Thodberg, M.; Antoku, Y.; Schweiger, P.J.; Rib, L.; Bressan, R.B.; Miao, Y.; et al. Mesenchymal-epithelial crosstalk shapes intestinal regionalisation via Wnt and Shh signalling. Nat Commun 2022, 13, 715. [Google Scholar] [CrossRef]
- Kurokawa, K.; Hayakawa, Y.; Koike, K. Plasticity of Intestinal Epithelium: Stem Cell Niches and Regulatory Signals. Int J Mol Sci 2020, 22. [Google Scholar] [CrossRef] [PubMed]
- Onfroy-Roy, L.; Hamel, D.; Malaquin, L.; Ferrand, A. Colon Fibroblasts and Inflammation: Sparring Partners in Colorectal Cancer Initiation? Cancers (Basel) 2021, 13, 1749. [Google Scholar] [CrossRef]
- Chen, L.; Dupre, A.; Qiu, X.; Pellon-Cardenas, O.; Walton, K.D.; Wang, J.; Perekatt, A.O.; Hu, W.; Spence, J.R.; Verzi, M.P. TGFB1 Induces Fetal Reprogramming and Enhances Intestinal Regeneration. bioRxiv 2021. [Google Scholar] [CrossRef]
- Iqbal, S.; Andersson, S.; Nestaite, E.; Pentinmikko, N.; Kumar, A.; Borshagovski, D.; Webb, A.; Saarinen, T.; Juuti, A.; Ori, A.; et al. Fetal-like reversion in the regenerating intestine is regulated by mesenchymal Asporin. bioRxiv 2021. [Google Scholar] [CrossRef]
- Brugger, M.D.; Basler, K. The diverse nature of intestinal fibroblasts in development, homeostasis, and disease. Trends Cell Biol 2023, 33, 834–849. [Google Scholar] [CrossRef] [PubMed]
- Fink, M.; Wrana, J.L. Regulation of homeostasis and regeneration in the adult intestinal epithelium by the TGF-beta superfamily. Dev Dyn 2023, 252, 445–462. [Google Scholar] [CrossRef] [PubMed]
- Baulies, A.; Angelis, N.; Li, V.S.W. Hallmarks of intestinal stem cells. Development 2020, 147. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Chen, Y.G. Intestinal epithelial plasticity and regeneration via cell dedifferentiation. Cell Regen 2020, 9, 14. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; van Es, J.H.; Snippert, H.J.; Stange, D.E.; Vries, R.G.; van den Born, M.; Barker, N.; Shroyer, N.F.; van de Wetering, M.; Clevers, H. Paneth cells constitute the niche for Lgr5 stem cells in intestinal crypts. Nature 2011, 469, 415–418. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, O.H.; Katajisto, P.; Lamming, D.W.; Gultekin, Y.; Bauer-Rowe, K.E.; Sengupta, S.; Birsoy, K.; Dursun, A.; Yilmaz, V.O.; Selig, M.; et al. mTORC1 in the Paneth cell niche couples intestinal stem-cell function to calorie intake. Nature 2012, 486, 490–495. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.H.; Escudero, S.; Shivdasani, R.A. Intact function of Lgr5 receptor-expressing intestinal stem cells in the absence of Paneth cells. Proc Natl Acad Sci U S A 2012, 109, 3932–3937. [Google Scholar] [CrossRef]
- Durand, A.; Donahue, B.; Peignon, G.; Letourneur, F.; Cagnard, N.; Slomianny, C.; Perret, C.; Shroyer, N.F.; Romagnolo, B. Functional intestinal stem cells after Paneth cell ablation induced by the loss of transcription factor Math1 (Atoh1). Proc Natl Acad Sci U S A 2012, 109, 8965–8970. [Google Scholar] [CrossRef] [PubMed]
- Ayyaz, A.; Kumar, S.; Sangiorgi, B.; Ghoshal, B.; Gosio, J.; Ouladan, S.; Fink, M.; Barutcu, S.; Trcka, D.; Shen, J.; et al. Single-cell transcriptomes of the regenerating intestine reveal a revival stem cell. Nature 2019, 569, 121–125. [Google Scholar] [CrossRef] [PubMed]
- Abud, H.E.; Chan, W.H.; Jarde, T. Source and Impact of the EGF Family of Ligands on Intestinal Stem Cells. Front Cell Dev Biol 2021, 9, 685665. [Google Scholar] [CrossRef]
- Jones, J.C.; Brindley, C.D.; Elder, N.H.; Myers, M.G., Jr.; Rajala, M.W.; Dekaney, C.M.; McNamee, E.N.; Frey, M.R.; Shroyer, N.F.; Dempsey, P.J. Cellular Plasticity of Defa4(Cre)-Expressing Paneth Cells in Response to Notch Activation and Intestinal Injury. Cell Mol Gastroenterol Hepatol 2019, 7, 533–554. [Google Scholar] [CrossRef]
- Koch, U.; Lehal, R.; Radtke, F. Stem cells living with a Notch. Development 2013, 140, 689–704. [Google Scholar] [CrossRef]
- Meyer, A.R.; Brown, M.E.; McGrath, P.S.; Dempsey, P.J. Injury-Induced Cellular Plasticity Drives Intestinal Regeneration. Cell Mol Gastroenterol Hepatol 2022, 13, 843–856. [Google Scholar] [CrossRef]
- Kraiczy, J.; McCarthy, N.; Malagola, E.; Tie, G.; Madha, S.; Boffelli, D.; Wagner, D.E.; Wang, T.C.; Shivdasani, R.A. Graded BMP signaling within intestinal crypt architecture directs self-organization of the Wnt-secreting stem cell niche. Cell Stem Cell 2023, 30, 433–449 e438. [Google Scholar] [CrossRef]
- Paerregaard, S.I.; Wulff, L.; Schussek, S.; Niss, K.; Morbe, U.; Jendholm, J.; Wendland, K.; Andrusaite, A.T.; Brulois, K.F.; Nibbs, R.J.B.; et al. The small and large intestine contain related mesenchymal subsets that derive from embryonic Gli1(+) precursors. Nat Commun 2023, 14, 2307. [Google Scholar] [CrossRef] [PubMed]
- Sylvestre, M.; Di Carlo, S.E.; Peduto, L. Stromal regulation of the intestinal barrier. Mucosal Immunol 2023, 16, 221–231. [Google Scholar] [CrossRef] [PubMed]
- Pentinmikko, N.; Iqbal, S.; Mana, M.; Andersson, S.; Cognetta, A.B., 3rd; Suciu, R.M.; Roper, J.; Luopajarvi, K.; Markelin, E.; Gopalakrishnan, S.; et al. Notum produced by Paneth cells attenuates regeneration of aged intestinal epithelium. Nature 2019, 571, 398–402. [Google Scholar] [CrossRef]
- Van Es, J.H.; Sato, T.; Van De Wetering, M.; Lyubimova, A.; Yee Nee, A.N.; Gregorieff, A.; Sasaki, N.; Zeinstra, L.; Van Den Born, M.; Korving, J. Dll1+ secretory progenitor cells revert to stem cells upon crypt damage. Nature cell biology 2012, 14, 1099–1104. [Google Scholar] [CrossRef]
- Das, S.; Feng, Q.; Balasubramanian, I.; Lin, X.; Liu, H.; Pellon-Cardenas, O.; Yu, S.; Zhang, X.; Liu, Y.; Wei, Z.; et al. Colonic healing requires Wnt produced by epithelium as well as Tagln+ and Acta2+ stromal cells. Development 2022, 149. [Google Scholar] [CrossRef] [PubMed]
- Jacob, J.M.; Di Carlo, S.E.; Stzepourginski, I.; Lepelletier, A.; Ndiaye, P.D.; Varet, H.; Legendre, R.; Kornobis, E.; Benabid, A.; Nigro, G.; et al. PDGFRalpha-induced stromal maturation is required to restrain postnatal intestinal epithelial stemness and promote defense mechanisms. Cell Stem Cell 2022, 29, 856–868 e855. [Google Scholar] [CrossRef] [PubMed]
- Stzepourginski, I.; Nigro, G.; Jacob, J.M.; Dulauroy, S.; Sansonetti, P.J.; Eberl, G.; Peduto, L. CD34+ mesenchymal cells are a major component of the intestinal stem cells niche at homeostasis and after injury. Proc Natl Acad Sci U S A 2017, 114, E506–E513. [Google Scholar] [CrossRef] [PubMed]
- Greicius, G.; Kabiri, Z.; Sigmundsson, K.; Liang, C.; Bunte, R.; Singh, M.K.; Virshup, D.M. PDGFRalpha(+) pericryptal stromal cells are the critical source of Wnts and RSPO3 for murine intestinal stem cells in vivo. Proc Natl Acad Sci U S A 2018, 115, E3173–E3181. [Google Scholar] [CrossRef] [PubMed]
- Kuhnert, F.; Davis, C.R.; Wang, H.T.; Chu, P.; Lee, M.; Yuan, J.; Nusse, R.; Kuo, C.J. Essential requirement for Wnt signaling in proliferation of adult small intestine and colon revealed by adenoviral expression of Dickkopf-1. Proc Natl Acad Sci U S A 2004, 101, 266–271. [Google Scholar] [CrossRef] [PubMed]
- Hausmann, G.; Banziger, C.; Basler, K. Helping Wingless take flight: how WNT proteins are secreted. Nat Rev Mol Cell Biol 2007, 8, 331–336. [Google Scholar] [CrossRef] [PubMed]
- Yan, K.S.; Janda, C.Y.; Chang, J.; Zheng, G.X.Y.; Larkin, K.A.; Luca, V.C.; Chia, L.A.; Mah, A.T.; Han, A.; Terry, J.M.; et al. Non-equivalence of Wnt and R-spondin ligands during Lgr5(+) intestinal stem-cell self-renewal. Nature 2017, 545, 238–242. [Google Scholar] [CrossRef] [PubMed]
- Flanagan, D.J.; Austin, C.R.; Vincan, E.; Phesse, T.J. Wnt Signalling in Gastrointestinal Epithelial Stem Cells. Genes (Basel) 2018, 9, 178. [Google Scholar] [CrossRef]
- Bottcher, A.; Buttner, M.; Tritschler, S.; Sterr, M.; Aliluev, A.; Oppenlander, L.; Burtscher, I.; Sass, S.; Irmler, M.; Beckers, J.; et al. Author Correction: Non-canonical Wnt/PCP signalling regulates intestinal stem cell lineage priming towards enteroendocrine and Paneth cell fates. Nat Cell Biol 2021, 23, 566–576. [Google Scholar] [CrossRef]
- Flores-Hernandez, E.; Velazquez, D.M.; Castaneda-Patlan, M.C.; Fuentes-Garcia, G.; Fonseca-Camarillo, G.; Yamamoto-Furusho, J.K.; Romero-Avila, M.T.; Garcia-Sainz, J.A.; Robles-Flores, M. Canonical and non-canonical Wnt signaling are simultaneously activated by Wnts in colon cancer cells. Cell Signal 2020, 72, 109636. [Google Scholar] [CrossRef]
- Werner, J.; Boonekamp, K.E.; Zhan, T.; Boutros, M. The Roles of Secreted Wnt Ligands in Cancer. Int J Mol Sci 2023, 24, 5349. [Google Scholar] [CrossRef]
- Halpern, K.B.; Massalha, H.; Zwick, R.K.; Moor, A.E.; Castillo-Azofeifa, D.; Rozenberg, M.; Farack, L.; Egozi, A.; Miller, D.R.; Averbukh, I.; et al. Lgr5+ telocytes are a signaling hub at the intestinal villus tip. bioRxiv 2020, 850909. [Google Scholar] [CrossRef]
- McCarthy, N.; Kraiczy, J.; Shivdasani, R.A. Cellular and molecular architecture of the intestinal stem cell niche. Nat Cell Biol 2020, 22, 1033–1041. [Google Scholar] [CrossRef] [PubMed]
- Bonis, V.; Rossell, C.; Gehart, H. The Intestinal Epithelium - Fluid Fate and Rigid Structure From Crypt Bottom to Villus Tip. Front Cell Dev Biol 2021, 9, 661931. [Google Scholar] [CrossRef]
- Corda, G.; Sala, A. Non-canonical WNT/PCP signalling in cancer: Fzd6 takes centre stage. Oncogenesis 2017, 6, e364. [Google Scholar] [CrossRef] [PubMed]
- De, A. Wnt/Ca2+ signaling pathway: a brief overview. Acta Biochim Biophys Sin (Shanghai) 2011, 43, 745–756. [Google Scholar] [CrossRef] [PubMed]
- Mehta, S.; Hingole, S.; Chaudhary, V. The Emerging Mechanisms of Wnt Secretion and Signaling in Development. Front Cell Dev Biol 2021, 9, 714746. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Singh, U.K.; Kini, S.G.; Garg, V.; Agrawal, S.; Tomar, P.K.; Pathak, P.; Chaudhary, A.; Gupta, P.; Malik, A. JNK pathway signaling: a novel and smarter therapeutic targets for various biological diseases. Future Med Chem 2015, 7, 2065–2086. [Google Scholar] [CrossRef] [PubMed]
- Walton, K.D.; Freddo, A.M.; Wang, S.; Gumucio, D.L. Generation of intestinal surface: an absorbing tale. Development 2016, 143, 2261–2272. [Google Scholar] [CrossRef]
- Walton, K.D.; Kolterud, A.; Czerwinski, M.J.; Bell, M.J.; Prakash, A.; Kushwaha, J.; Grosse, A.S.; Schnell, S.; Gumucio, D.L. Hedgehog-responsive mesenchymal clusters direct patterning and emergence of intestinal villi. Proc Natl Acad Sci U S A 2012, 109, 15817–15822. [Google Scholar] [CrossRef]
- Wang, S.; Chen, Y.G. BMP signaling in homeostasis, transformation and inflammatory response of intestinal epithelium. Sci China Life Sci 2018, 61, 800–807. [Google Scholar] [CrossRef]
- Madison, B.B.; Braunstein, K.; Kuizon, E.; Portman, K.; Qiao, X.T.; Gumucio, D.L. Epithelial hedgehog signals pattern the intestinal crypt-villus axis. Development 2005, 132, 279–289. [Google Scholar] [CrossRef]
- Walton, K.D.; Gumucio, D.L. Hedgehog Signaling in Intestinal Development and Homeostasis. Annu Rev Physiol 2021, 83, 359–380. [Google Scholar] [CrossRef]
- McCarthy, N.; Manieri, E.; Storm, E.E.; Saadatpour, A.; Luoma, A.M.; Kapoor, V.N.; Madha, S.; Gaynor, L.T.; Cox, C.; Keerthivasan, S.; et al. Distinct Mesenchymal Cell Populations Generate the Essential Intestinal BMP Signaling Gradient. Cell Stem Cell 2020, 26, 391–402 e395. [Google Scholar] [CrossRef]
- Orzechowska-Licari, E.J.; Bialkowska, A.B.; Yang, V.W. Sonic Hedgehog and WNT Signaling Regulate a Positive Feedback Loop Between Intestinal Epithelial and Stromal Cells to Promote Epithelial Regeneration. Cell Mol Gastroenterol Hepatol 2023, 16, 607–642. [Google Scholar] [CrossRef] [PubMed]
- Chin, A.M.; Hill, D.R.; Aurora, M.; Spence, J.R. Morphogenesis and maturation of the embryonic and postnatal intestine. Semin Cell Dev Biol 2017, 66, 81–93. [Google Scholar] [CrossRef] [PubMed]
- Greicius, G.; Virshup, D.M. Stromal control of intestinal development and the stem cell niche. Differentiation 2019, 108, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Pellegrinet, L.; Rodilla, V.; Liu, Z.; Chen, S.; Koch, U.; Espinosa, L.; Kaestner, K.H.; Kopan, R.; Lewis, J.; Radtke, F. Dll1- and dll4-mediated notch signaling are required for homeostasis of intestinal stem cells. Gastroenterology 2011, 140, 1230–1240 e1231-1237. [Google Scholar] [CrossRef] [PubMed]
- Liang, S.J.; Li, X.G.; Wang, X.Q. Notch Signaling in Mammalian Intestinal Stem Cells: Determining Cell Fate and Maintaining Homeostasis. Curr Stem Cell Res Ther 2019, 14, 583–590. [Google Scholar] [CrossRef] [PubMed]
- Geyer, N.; Gerling, M. Hedgehog signaling in colorectal cancer: All in the stroma? International Journal of Molecular Sciences 2021, 22, 1025. [Google Scholar] [CrossRef] [PubMed]
- Degirmenci, B.; Valenta, T.; Dimitrieva, S.; Hausmann, G.; Basler, K. GLI1-expressing mesenchymal cells form the essential Wnt-secreting niche for colon stem cells. Nature 2018, 558, 449–453. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.E.; Fei, L.; Yin, W.C.; Coquenlorge, S.; Rao-Bhatia, A.; Zhang, X.; Shi, S.S.W.; Lee, J.H.; Hahn, N.A.; Rizvi, W.; et al. Single cell and genetic analyses reveal conserved populations and signaling mechanisms of gastrointestinal stromal niches. Nat Commun 2020, 11, 334. [Google Scholar] [CrossRef]
- Rees, W.D.; Tandun, R.; Yau, E.; Zachos, N.C.; Steiner, T.S. Regenerative Intestinal Stem Cells Induced by Acute and Chronic Injury: The Saving Grace of the Epithelium? Front Cell Dev Biol 2020, 8, 583919. [Google Scholar] [CrossRef] [PubMed]
- Baghdadi, M.B.; Ayyaz, A.; Coquenlorge, S.; Chu, B.; Kumar, S.; Streutker, C.; Wrana, J.L.; Kim, T.H. Enteric glial cell heterogeneity regulates intestinal stem cell niches. Cell Stem Cell 2022, 29, 86–100 e106. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, M.; Schewe, M.; Sacchetti, A.; Feijtel, D.; van de Geer, W.S.; Teeuwssen, M.; Sleddens, H.F.; Joosten, R.; van Royen, M.E.; van de Werken, H.J.G.; et al. Paneth Cells Respond to Inflammation and Contribute to Tissue Regeneration by Acquiring Stem-like Features through SCF/c-Kit Signaling. Cell Rep 2018, 24, 2312–2328 e2317. [Google Scholar] [CrossRef]
- Metcalfe, C.; Kljavin, N.M.; Ybarra, R.; de Sauvage, F.J. Lgr5+ stem cells are indispensable for radiation-induced intestinal regeneration. Cell Stem Cell 2014, 14, 149–159. [Google Scholar] [CrossRef]
- Wang, Y.; Chiang, I.L.; Ohara, T.E.; Fujii, S.; Cheng, J.; Muegge, B.D.; Ver Heul, A.; Han, N.D.; Lu, Q.; Xiong, S.; et al. Long-Term Culture Captures Injury-Repair Cycles of Colonic Stem Cells. Cell 2019, 179, 1144–1159 e1115. [Google Scholar] [CrossRef]
- Rees, W.D.; Telkar, N.; Lin, D.T.S.; Wong, M.Q.; Poloni, C.; Fathi, A.; Kobor, M.; Zachos, N.C.; Steiner, T.S. An in vitro chronic damage model impairs inflammatory and regenerative responses in human colonoid monolayers. Cell Rep 2022, 38, 110283. [Google Scholar] [CrossRef]
- Barry, E.R.; Morikawa, T.; Butler, B.L.; Shrestha, K.; de la Rosa, R.; Yan, K.S.; Fuchs, C.S.; Magness, S.T.; Smits, R.; Ogino, S.; et al. Restriction of intestinal stem cell expansion and the regenerative response by YAP. Nature 2013, 493, 106–110. [Google Scholar] [CrossRef]
- Yan, K.S.; Chia, L.A.; Li, X.; Ootani, A.; Su, J.; Lee, J.Y.; Su, N.; Luo, Y.; Heilshorn, S.C.; Amieva, M.R.; et al. The intestinal stem cell markers Bmi1 and Lgr5 identify two functionally distinct populations. Proc Natl Acad Sci U S A 2012, 109, 466–471. [Google Scholar] [CrossRef]
- Tian, H.; Biehs, B.; Warming, S.; Leong, K.G.; Rangell, L.; Klein, O.D.; de Sauvage, F.J. A reserve stem cell population in small intestine renders Lgr5-positive cells dispensable. Nature 2011, 478, 255–259. [Google Scholar] [CrossRef]
- Murata, K.; Jadhav, U.; Madha, S.; van Es, J.; Dean, J.; Cavazza, A.; Wucherpfennig, K.; Michor, F.; Clevers, H.; Shivdasani, R.A. Ascl2-Dependent Cell Dedifferentiation Drives Regeneration of Ablated Intestinal Stem Cells. Cell Stem Cell 2020, 26, 377–390 e376. [Google Scholar] [CrossRef] [PubMed]
- Bankaitis, E.D.; Ha, A.; Kuo, C.J.; Magness, S.T. Reserve Stem Cells in Intestinal Homeostasis and Injury. Gastroenterology 2018, 155, 1348–1361. [Google Scholar] [CrossRef]
- Montgomery, R.K.; Carlone, D.L.; Richmond, C.A.; Farilla, L.; Kranendonk, M.E.; Henderson, D.E.; Baffour-Awuah, N.Y.; Ambruzs, D.M.; Fogli, L.K.; Algra, S.; et al. Mouse telomerase reverse transcriptase (mTert) expression marks slowly cycling intestinal stem cells. Proc Natl Acad Sci U S A 2011, 108, 179–184. [Google Scholar] [CrossRef] [PubMed]
- Yousefi, M.; Li, N.; Nakauka-Ddamba, A.; Wang, S.; Davidow, K.; Schoenberger, J.; Yu, Z.; Jensen, S.T.; Kharas, M.G.; Lengner, C.J. Msi RNA-binding proteins control reserve intestinal stem cell quiescence. J Cell Biol 2016, 215, 401–413. [Google Scholar] [CrossRef] [PubMed]
- Suh, H.N.; Kim, M.J.; Jung, Y.S.; Lien, E.M.; Jun, S.; Park, J.I. Quiescence Exit of Tert(+) Stem Cells by Wnt/beta-Catenin Is Indispensable for Intestinal Regeneration. Cell Rep 2017, 21, 2571–2584. [Google Scholar] [CrossRef]
- Hageman, J.H.; Heinz, M.C.; Kretzschmar, K.; van der Vaart, J.; Clevers, H.; Snippert, H.J.G. Intestinal Regeneration: Regulation by the Microenvironment. Dev Cell 2020, 54, 435–446. [Google Scholar] [CrossRef]
- Sprangers, J.; Zaalberg, I.C.; Maurice, M.M. Organoid-based modeling of intestinal development, regeneration, and repair. Cell Death Differ 2021, 28, 95–107. [Google Scholar] [CrossRef]
- Yan, K.S.; Gevaert, O.; Zheng, G.X.Y.; Anchang, B.; Probert, C.S.; Larkin, K.A.; Davies, P.S.; Cheng, Z.F.; Kaddis, J.S.; Han, A.; et al. Intestinal Enteroendocrine Lineage Cells Possess Homeostatic and Injury-Inducible Stem Cell Activity. Cell Stem Cell 2017, 21, 78–90 e76. [Google Scholar] [CrossRef]
- Shivdasani, R.A.; Clevers, H.; de Sauvage, F.J. Tissue regeneration: Reserve or reverse? Science 2021, 371, 784–786. [Google Scholar] [CrossRef]
- Tomic, G.; Morrissey, E.; Kozar, S.; Ben-Moshe, S.; Hoyle, A.; Azzarelli, R.; Kemp, R.; Chilamakuri, C.S.R.; Itzkovitz, S.; Philpott, A.; et al. Phospho-regulation of ATOH1 Is Required for Plasticity of Secretory Progenitors and Tissue Regeneration. Cell Stem Cell 2018, 23, 436–443 e437. [Google Scholar] [CrossRef]
- Castillo-Azofeifa, D.; Fazio, E.N.; Nattiv, R.; Good, H.J.; Wald, T.; Pest, M.A.; de Sauvage, F.J.; Klein, O.D.; Asfaha, S. Atoh1(+) secretory progenitors possess renewal capacity independent of Lgr5(+) cells during colonic regeneration. EMBO J 2019, 38, e99984. [Google Scholar] [CrossRef]
- Yu, S.; Tong, K.; Zhao, Y.; Balasubramanian, I.; Yap, G.S.; Ferraris, R.P.; Bonder, E.M.; Verzi, M.P.; Gao, N. Paneth Cell Multipotency Induced by Notch Activation following Injury. Cell Stem Cell 2018, 23, 46–59 e45. [Google Scholar] [CrossRef]
- Deng, F.; Peng, L.; Li, Z.; Tan, G.; Liang, E.; Chen, S.; Zhao, X.; Zhi, F. YAP triggers the Wnt/beta-catenin signalling pathway and promotes enterocyte self-renewal, regeneration and tumorigenesis after DSS-induced injury. Cell Death Dis 2018, 9, 153. [Google Scholar] [CrossRef]
- Hong, A.W.; Meng, Z.; Guan, K.L. The Hippo pathway in intestinal regeneration and disease. Nat Rev Gastroenterol Hepatol 2016, 13, 324–337. [Google Scholar] [CrossRef] [PubMed]
- Palikuqi, B.; Rispal, J.; Klein, O. Good Neighbors: The Niche that Fine Tunes Mammalian Intestinal Regeneration. Cold Spring Harb Perspect Biol 2022, 14, a040865. [Google Scholar] [CrossRef]
- Chalkidi, N.; Paraskeva, C.; Koliaraki, V. Fibroblasts in intestinal homeostasis, damage, and repair. Front Immunol 2022, 13, 924866. [Google Scholar] [CrossRef] [PubMed]
- Nusse, Y.M.; Savage, A.K.; Marangoni, P.; Rosendahl-Huber, A.K.; Landman, T.A.; De Sauvage, F.J.; Locksley, R.M.; Klein, O.D. Parasitic helminths induce fetal-like reversion in the intestinal stem cell niche. Nature 2018, 559, 109–113. [Google Scholar] [CrossRef] [PubMed]
- van Dop, W.A.; Heijmans, J.; Buller, N.V.; Snoek, S.A.; Rosekrans, S.L.; Wassenberg, E.A.; van den Bergh Weerman, M.A.; Lanske, B.; Clarke, A.R.; Winton, D.J.; et al. Loss of Indian Hedgehog activates multiple aspects of a wound healing response in the mouse intestine. Gastroenterology 2010, 139, 1665–1676.e10. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.; Peng, Y.; Li, H. The Injury-Related Activation of Hedgehog Signaling Pathway Modulates the Repair-Associated Inflammation in Liver Fibrosis. Front Immunol 2017, 8, 1450. [Google Scholar] [CrossRef]
- Xie, Z.; Zhang, M.; Zhou, G.; Lin, L.; Han, J.; Wang, Y.; Li, L.; He, Y.; Zeng, Z.; Chen, M.; et al. Emerging roles of the Hedgehog signalling pathway in inflammatory bowel disease. Cell Death Discov 2021, 7, 314. [Google Scholar] [CrossRef]
- Pasztoi, M.; Ohnmacht, C. Tissue Niches Formed by Intestinal Mesenchymal Stromal Cells in Mucosal Homeostasis and Immunity. Int J Mol Sci 2022, 23, 5181. [Google Scholar] [CrossRef]
- Owens, B.M. Inflammation, Innate Immunity, and the Intestinal Stromal Cell Niche: Opportunities and Challenges. Front Immunol 2015, 6, 319. [Google Scholar] [CrossRef]
- Roulis, M.; Flavell, R.A. Fibroblasts and myofibroblasts of the intestinal lamina propria in physiology and disease. Differentiation 2016, 92, 116–131. [Google Scholar] [CrossRef] [PubMed]
- Powell, D.W.; Pinchuk, I.V.; Saada, J.I.; Chen, X.; Mifflin, R.C. Mesenchymal cells of the intestinal lamina propria. Annu Rev Physiol 2011, 73, 213–237. [Google Scholar] [CrossRef] [PubMed]
- Bonnans, C.; Chou, J.; Werb, Z. Remodelling the extracellular matrix in development and disease. Nat Rev Mol Cell Biol 2014, 15, 786–801. [Google Scholar] [CrossRef]
- Fawkner-Corbett, D.; Antanaviciute, A.; Parikh, K.; Jagielowicz, M.; Geros, A.S.; Gupta, T.; Ashley, N.; Khamis, D.; Fowler, D.; Morrissey, E.; et al. Spatiotemporal analysis of human intestinal development at single-cell resolution. Cell 2021, 184, 810–826 e823. [Google Scholar] [CrossRef] [PubMed]
- Sherwood, R.I.; Maehr, R.; Mazzoni, E.O.; Melton, D.A. Wnt signaling specifies and patterns intestinal endoderm. Mech Dev 2011, 128, 387–400. [Google Scholar] [CrossRef]
- Chen, L.; Acciani, T.; Le Cras, T.; Lutzko, C.; Perl, A.K. Dynamic regulation of platelet-derived growth factor receptor alpha expression in alveolar fibroblasts during realveolarization. Am J Respir Cell Mol Biol 2012, 47, 517–527. [Google Scholar] [CrossRef]
- Korinek, V.; Barker, N.; Moerer, P.; van Donselaar, E.; Huls, G.; Peters, P.J.; Clevers, H. Depletion of epithelial stem-cell compartments in the small intestine of mice lacking Tcf-4. Nat Genet 1998, 19, 379–383. [Google Scholar] [CrossRef]
- Chin, A.M.; Tsai, Y.H.; Finkbeiner, S.R.; Nagy, M.S.; Walker, E.M.; Ethen, N.J.; Williams, B.O.; Battle, M.A.; Spence, J.R. A Dynamic WNT/beta-CATENIN Signaling Environment Leads to WNT-Independent and WNT-Dependent Proliferation of Embryonic Intestinal Progenitor Cells. Stem Cell Reports 2016, 7, 826–839. [Google Scholar] [CrossRef]
- Sun, H.; Tan, J.; Chen, H.; Wu, N.; Su, B. Immune niches orchestrated by intestinal mesenchymal stromal cells lining the crypt-villus. Front Immunol 2022, 13, 1057932. [Google Scholar] [CrossRef] [PubMed]
- Haffen, K.; Kedinger, M.; Simon-Assmann, P. Mesenchyme-dependent differentiation of epithelial progenitor cells in the gut. J Pediatr Gastroenterol Nutr 1987, 6, 14–23. [Google Scholar] [CrossRef] [PubMed]
- Fazilaty, H.; Brugger, M.D.; Valenta, T.; Szczerba, B.M.; Berkova, L.; Doumpas, N.; Hausmann, G.; Scharl, M.; Basler, K. Tracing colonic embryonic transcriptional profiles and their reactivation upon intestinal damage. Cell Rep 2021, 36, 109484. [Google Scholar] [CrossRef] [PubMed]
- Ichinose, M.; Suzuki, N.; Wang, T.; Wright, J.A.; Lannagan, T.R.M.; Vrbanac, L.; Kobayashi, H.; Gieniec, K.A.; Ng, J.Q.; Hayakawa, Y.; et al. Stromal DLK1 promotes proliferation and inhibits differentiation of the intestinal epithelium during development. Am J Physiol Gastrointest Liver Physiol 2021, 320, G506–G520. [Google Scholar] [CrossRef] [PubMed]
- Bry, L.; Falk, P.; Huttner, K.; Ouellette, A.; Midtvedt, T.; Gordon, J.I. Paneth cell differentiation in the developing intestine of normal and transgenic mice. Proc Natl Acad Sci U S A 1994, 91, 10335–10339. [Google Scholar] [CrossRef] [PubMed]
- Dehmer, J.J.; Garrison, A.P.; Speck, K.E.; Dekaney, C.M.; Van Landeghem, L.; Sun, X.; Henning, S.J.; Helmrath, M.A. Expansion of intestinal epithelial stem cells during murine development. PLoS One 2011, 6, e27070. [Google Scholar] [CrossRef] [PubMed]
- Vannucchi, M.G.; Traini, C.; Manetti, M.; Ibba-Manneschi, L.; Faussone-Pellegrini, M.S. Telocytes express PDGFRalpha in the human gastrointestinal tract. J Cell Mol Med 2013, 17, 1099–1108. [Google Scholar] [CrossRef] [PubMed]
- Cretoiu, D.; Cretoiu, S.M.; Simionescu, A.A.; Popescu, L.M. Telocytes, a distinct type of cell among the stromal cells present in the lamina propria of jejunum. Histol Histopathol 2012, 27, 1067–1078. [Google Scholar] [CrossRef]
- Hong, S.P.; Yang, M.J.; Cho, H.; Park, I.; Bae, H.; Choe, K.; Suh, S.H.; Adams, R.H.; Alitalo, K.; Lim, D.; et al. Distinct fibroblast subsets regulate lacteal integrity through YAP/TAZ-induced VEGF-C in intestinal villi. Nat Commun 2020, 11, 4102. [Google Scholar] [CrossRef]
- VanDussen, K.L.; Carulli, A.J.; Keeley, T.M.; Patel, S.R.; Puthoff, B.J.; Magness, S.T.; Tran, I.T.; Maillard, I.; Siebel, C.; Kolterud, A.; et al. Notch signaling modulates proliferation and differentiation of intestinal crypt base columnar stem cells. Development 2012, 139, 488–497. [Google Scholar] [CrossRef] [PubMed]
- Valenta, T.; Degirmenci, B.; Moor, A.E.; Herr, P.; Zimmerli, D.; Moor, M.B.; Hausmann, G.; Cantu, C.; Aguet, M.; Basler, K. Wnt Ligands Secreted by Subepithelial Mesenchymal Cells Are Essential for the Survival of Intestinal Stem Cells and Gut Homeostasis. Cell Rep 2016, 15, 911–918. [Google Scholar] [CrossRef]
- Lahar, N.; Lei, N.Y.; Wang, J.; Jabaji, Z.; Tung, S.C.; Joshi, V.; Lewis, M.; Stelzner, M.; Martin, M.G.; Dunn, J.C. Intestinal subepithelial myofibroblasts support in vitro and in vivo growth of human small intestinal epithelium. PLoS One 2011, 6, e26898. [Google Scholar] [CrossRef]
- Guenin-Mace, L.; Konieczny, P.; Naik, S. Immune-Epithelial Cross Talk in Regeneration and Repair. Annu Rev Immunol 2023, 41, 207–228. [Google Scholar] [CrossRef]
- Poggi, L.; Casarosa, S.; Carl, M. An Eye on the Wnt Inhibitory Factor Wif1. Front Cell Dev Biol 2018, 6, 167. [Google Scholar] [CrossRef] [PubMed]
- Melissari, M.T.; Henriques, A.; Tzaferis, C.; Prados, A.; Sarris, M.E.; Chalkidi, N.; Mavroeidi, D.; Chouvardas, P.; Grammenoudi, S.; Kollias, G.; et al. Col6a1(+)/CD201(+) mesenchymal cells regulate intestinal morphogenesis and homeostasis. Cell Mol Life Sci 2021, 79, 1. [Google Scholar] [CrossRef]
- Chivukula, R.R.; Shi, G.; Acharya, A.; Mills, E.W.; Zeitels, L.R.; Anandam, J.L.; Abdelnaby, A.A.; Balch, G.C.; Mansour, J.C.; Yopp, A.C.; et al. An essential mesenchymal function for miR-143/145 in intestinal epithelial regeneration. Cell 2014, 157, 1104–1116. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Multiple signaling pathways regulating intestinal epithelial–mesenchymal crosstalk in the crypt. The diagram provides an overview of the intricate mechanisms involved in the interactions between crypt-based epithelial cells and the neighboring iMSCs. This interplay controls intestinal stem cell (ISC) homeostasis and epithelial differentiation. The activation of Wnt-promoting pathways and inhibition of Bmp/Bmpr binding orchestrate ISC stemness in the crypt base. Paneth cells secrete Wnt, Notch, and epithelial growth factors (EGF) ligands that induce Wnt target gene transcription in ISC. Wnt factors secreted by subcryptal iMSCs bind FZD and LRPs co-receptors and stabilize β-catenin in the Wnt signaling cascade to promote ISC-related gene transcription. Rspos binding LGR family receptors stabilize FZD expressions, contributing to WNT pathway activation. In addition, subcryptal iMSCs secrete Bmp antagonists such as Gremlin to maintain Wnt activities in the ISC niche.
Figure 1.
Multiple signaling pathways regulating intestinal epithelial–mesenchymal crosstalk in the crypt. The diagram provides an overview of the intricate mechanisms involved in the interactions between crypt-based epithelial cells and the neighboring iMSCs. This interplay controls intestinal stem cell (ISC) homeostasis and epithelial differentiation. The activation of Wnt-promoting pathways and inhibition of Bmp/Bmpr binding orchestrate ISC stemness in the crypt base. Paneth cells secrete Wnt, Notch, and epithelial growth factors (EGF) ligands that induce Wnt target gene transcription in ISC. Wnt factors secreted by subcryptal iMSCs bind FZD and LRPs co-receptors and stabilize β-catenin in the Wnt signaling cascade to promote ISC-related gene transcription. Rspos binding LGR family receptors stabilize FZD expressions, contributing to WNT pathway activation. In addition, subcryptal iMSCs secrete Bmp antagonists such as Gremlin to maintain Wnt activities in the ISC niche.
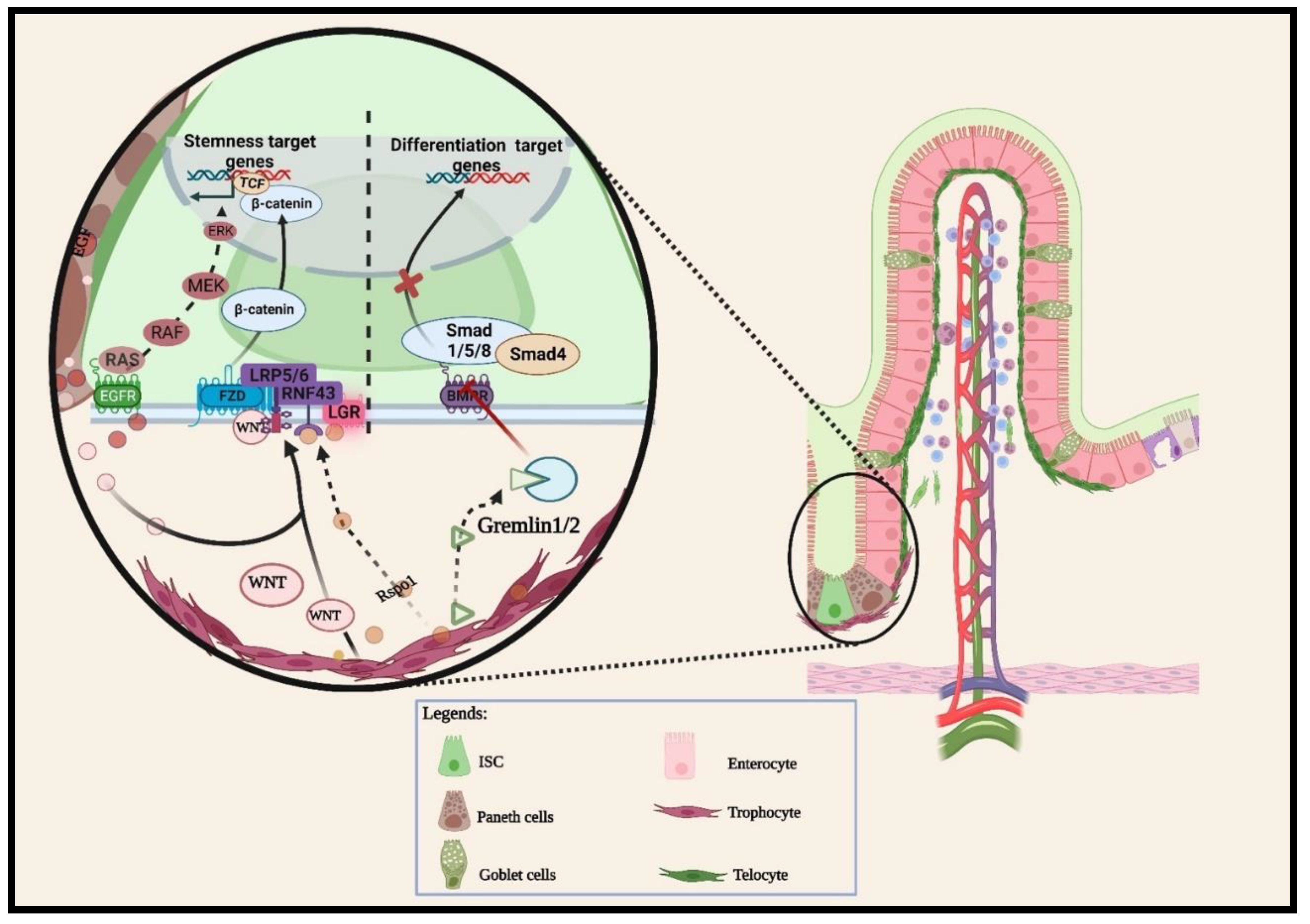
Figure 2.
Intestinal villus base epithelial secreted ligands activate hedgehog signaling in neighboring mesenchymal cells. These epithelial–mesenchymal interactions at the villus base stimulate intestinal epithelial cell terminal differentiation and coordinate the migration of differentiated cells toward the villus tip. Hedgehog ligands produced by epithelial cells bind on the Pitch1 receptor of PDGFRαhi cells located at the villus base. This binding triggers the transcription of hedgehog target genes. The genes transcribed by the Gli family of PDGFRαhi cells include Bmps. Secreted Bmp ligands from these iMSCs bind with the Bmp receptor (Bmpr) on adjacent epithelial cells. The Bmp and Bmpr binding subsequently phosphorylate the Smad family transcription factor, which in turn induces the differentiation of villus epithelial cells.
Figure 2.
Intestinal villus base epithelial secreted ligands activate hedgehog signaling in neighboring mesenchymal cells. These epithelial–mesenchymal interactions at the villus base stimulate intestinal epithelial cell terminal differentiation and coordinate the migration of differentiated cells toward the villus tip. Hedgehog ligands produced by epithelial cells bind on the Pitch1 receptor of PDGFRαhi cells located at the villus base. This binding triggers the transcription of hedgehog target genes. The genes transcribed by the Gli family of PDGFRαhi cells include Bmps. Secreted Bmp ligands from these iMSCs bind with the Bmp receptor (Bmpr) on adjacent epithelial cells. The Bmp and Bmpr binding subsequently phosphorylate the Smad family transcription factor, which in turn induces the differentiation of villus epithelial cells.
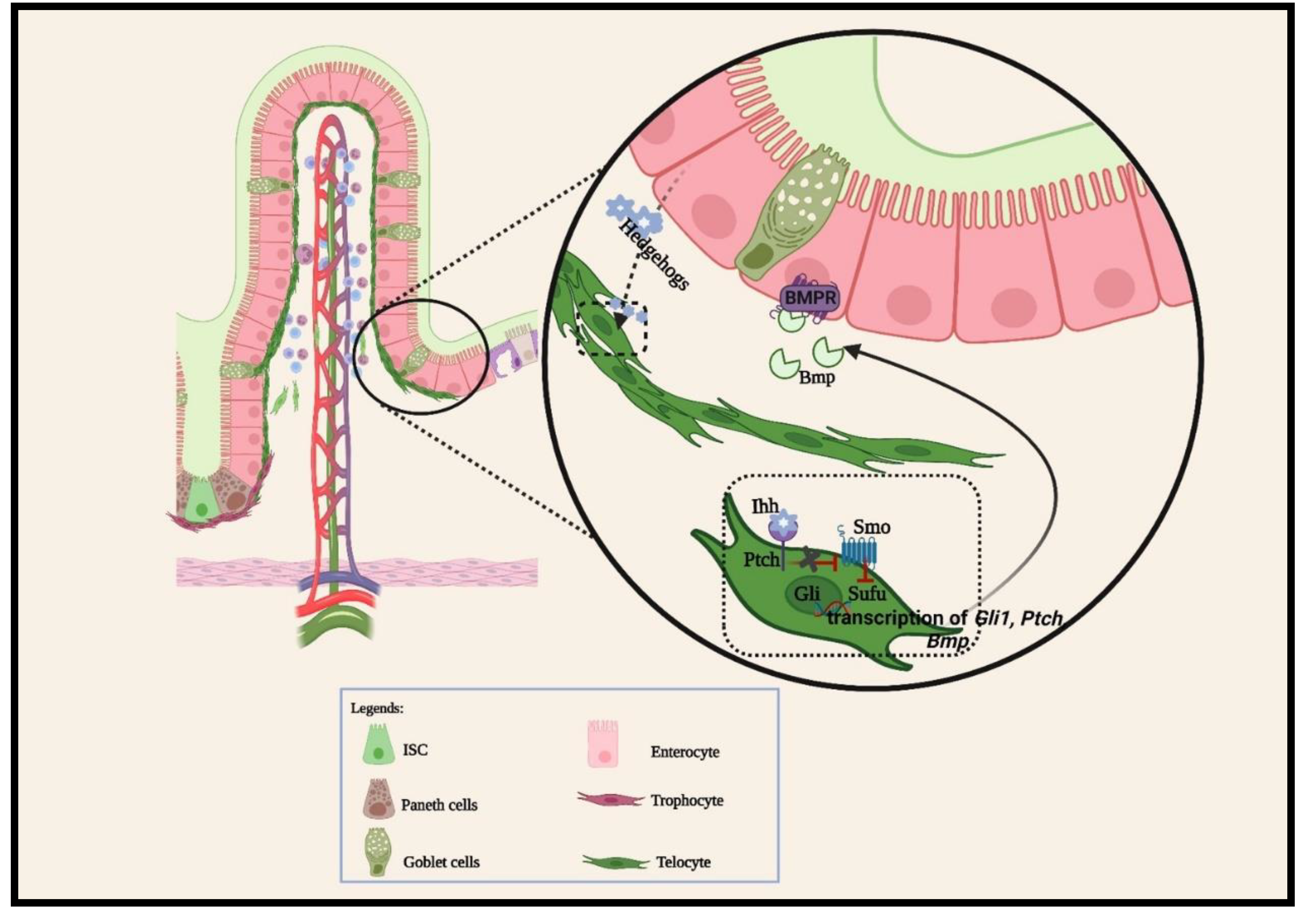
Figure 3.
Schematic representation of intestinal mesenchymal stromal cell subsets and their location. The illustration here describes the subpopulations of PDGFRα expressing cells, including telocytes, CD81- cells, and trophocytes. The trophocyte cluster group, CD81+, is confined to the subcryptal domain, beneath the muscularis mucosa, to secrete Wnt promoting factors that support the ISC niche. Other PDGFRαlo subsets, including CD55hi and Fgfr2+ cells, localize in the lamina propria and extend upward to the TA domain/villi trunk to initiate terminal differentiation. They switch the signal gradients from Wnt-promoting factors to Bmp agonists. PDGFRαhi subsets form the subepithelial stromal populations that are localized in the villi core in the small intestine and the colon top [38,39].
Figure 3.
Schematic representation of intestinal mesenchymal stromal cell subsets and their location. The illustration here describes the subpopulations of PDGFRα expressing cells, including telocytes, CD81- cells, and trophocytes. The trophocyte cluster group, CD81+, is confined to the subcryptal domain, beneath the muscularis mucosa, to secrete Wnt promoting factors that support the ISC niche. Other PDGFRαlo subsets, including CD55hi and Fgfr2+ cells, localize in the lamina propria and extend upward to the TA domain/villi trunk to initiate terminal differentiation. They switch the signal gradients from Wnt-promoting factors to Bmp agonists. PDGFRαhi subsets form the subepithelial stromal populations that are localized in the villi core in the small intestine and the colon top [38,39].
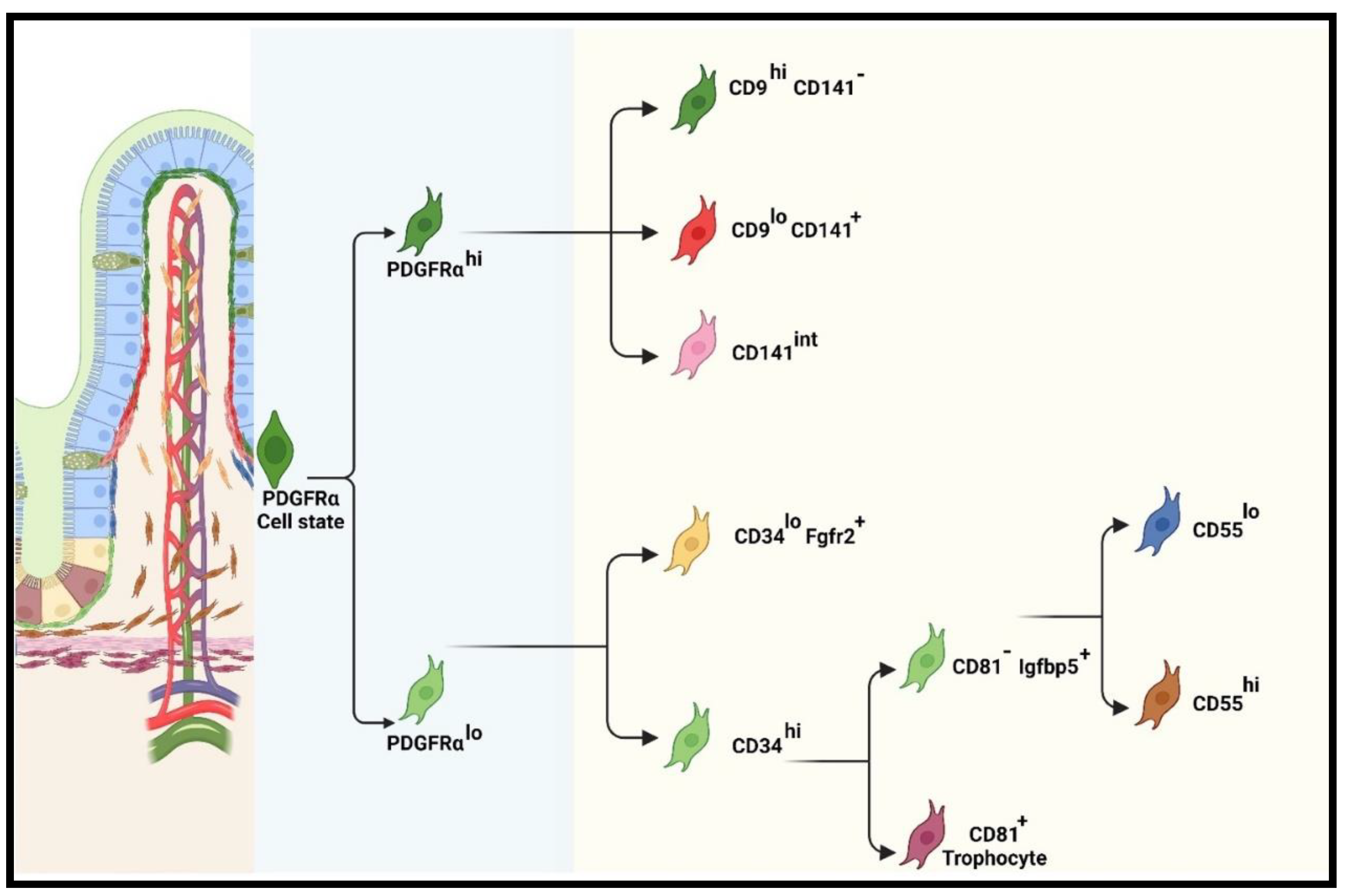

| Epithelial cells | Cell Markers | Ligands | Functions | Signaling pathways | References |
|---|---|---|---|---|---|
| Paneth cells | Defa4 expressing cell | Wnt3a, Wnt9b, Wnt11 | Support regeneration | Notch | [35] |
| pS6+ | Notum | Wnt inhibitor | Non-canonical Wnt/mTORC1 | [41] | |
| Progenitor cells | Dll+ | Dll1, dll4 | Support regeneration | Notch | [42] |
| Secretory lineage | Unknown | Egf, Tgfa, | Promote IEC homeostasis | EGFR/RAS | [34] |
| Colonic Paneth cells | REG4+ | Dll1, Egf | Promote stemness | Notch | [43] |
| +4 quiescent cells | BMI1, HOPX, MTERT | Unknown | Support regeneration | Hippo | [21,33] |
| Tuft cells | DCLK1 | Dll1 | Promote regeneration | Notch | [37] |
Table 2.
Distinct intestinal stromal subsets supporting intestinal balance regulations.
| Non-epithelial cells | Cell Markers | Ligands | Functions | Signaling pathways | References |
|---|---|---|---|---|---|
| PDGFRαlo Cells | CD81+, CD55hi | Wnt2b, Gremlin1/2, Rspo3 | Wnt promoters | Wnt/β-catenin | [38,66] |
| Fgfr2+, CD55lo | Wnt4, Frzb, Sfrp1 | Wnt repressors | Non-canonical Wnt/Bmp | [38,39] | |
| PDGFRαhi Cells | FOXL1+, CD9hiCD141-, CD9loCD141+, CD141int | Bmp3/4, Wnt5a/b, Dkk | BMP agonists,Wnt inhibitor | Non-canonical Wnt/Bmp | [39] |
| PDGFRα+ | PDGFRα+DLK1+ | Dlk1 | Embryonic morphogenesis | Notch | [117] |
| LTβR+ | LTβR+PDGFRαhi | Pdgf | Stromal maturation | Bmp activation | [44] |
| Smooth muscle cells | Tagln+, Acta2+,Myh11+ | Wnts | ISC integrity and wound healing | Wnt/β-catenin | [43] |
| Immune cells | ILC2, ILC3 | Il13, Il22 | Promote regeneration | Wnt/β-catenin | [126] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



