
Submitted:
07 February 2024
Posted:
08 February 2024
You are already at the latest version
Abstract
There is sufficient knowledge of the beneficial effect of warm-dry storage on breaking the morpho-logical dormancy of the chincuya (Annona purpurea Moc. & Sessé ex Dunal) seed embryo. Chincuya is an underutilized, tropical wild species native to Mexico, with potential as a biopesticide.
The impact of warm-dry storage and gibberellic acid treatment on seed viability and germination was determined to improve that knowledge further. Also, the content and kinetics of fatty acids in chincuya seeds were profiled during warm-dry storage. Seed treatments included storage time (0, 3, 6, 9, and 12 months), seed imbibition for 72 h in gibberellic acid at 350 mg L-1, and distilled water im-bibition for 72 h for the control treatment. Seed viability was determined using the TTZ classic test. Imbibed seeds were germinated on paper towels and incubated at 30 °C during the day and 25 °C at night in a 12:12 h dark:light photoperiod. The lipid profile was obtained by gas chromatography. During the storage treatment, seeds stayed in darkness at 25 ± 3 °C. Results showed that high viability remained up to month 9 in storage when it significantly decreased. Freshly harvested seeds germi-nated at a statistically significantly lower rate (26.1 %) than six-month-old seeds (65.6 %). The profile of fatty acids breaks down as palmitic acid (33.41 %), stearic acid (7.72 %), and arachidic acid (1.30 %), for a total of 42.44 % of saturated fatty acids. The detected unsaturated fatty acids (52.15 %) were palmitoleic acid (1.52 %), oleic acid (43.37 %), and linoleic acid (7.24 %). No significant changes were detected in fatty acid content; however, the highest fatty acid content (92.84 %) occurred with the highest germination rate. Regression analysis showed significant content changes: palmitoleic and palmitic acid increased while oleic acid decreased during dry storage. Arachidic acid showed a slight downward trend. Also, fatty acid content significantly differed between intact seeds (unstored and unincubated) and latent seeds (unstored, incubated, and ungerminated). The total fatty acids content for latent seeds was significantly lower. These results show that Annona purpurea seeds present morphophysiological dormancy. Also, we document the highest germination rate reported (65.6 %) and report the presence of arachidic and palmitoleic acids in chincuya seeds.
Keywords:
germination
; dormancy
; endodormancy
; latency
; after ripening
; fatty acids
; hormonal regulators
; de-velopment regulators
; anonas soncoya
; arachidic acid
; palmitic acid
; palmitoleic acid
; linoleic aci
1. Introduction
Annona purpurea Moc. & Sessé ex Dunal. Chincuya is a native species of Mexico. According to Cabrera et al. [1], it is a tree that can grow up to 10 m tall and has a crown formed by scattered branches. The flowers are extra-axillary, solitary, and subsessile, with three triangular-ovate, acuminate sepals. The three external petals are thick and rigid, concave and usually acuminate, ferruginous-sericeous on the outside, and tinted with purple inside. The three smaller inner petals are imbricate, slender, and elliptic-oblong with a rounded apex.
Additionally, there are numerous stamens. The leaves are alternate, deciduous, and simple, measuring 12 to 30 cm in length and 6 to 14 cm in width. They are obovate to elliptic in shape and have short petioles. The fruit is oval to spherical and covered with numerous pyramidal protuberances. It has a felt-like tomentum. According to Vidal-Lezama et al. [2], the fruits are syncarpous and have numerous uniseminated carpels. They are characterized by being very firm at harvest. The fruit measures an average of 31.73 cm in length and 47.17 cm in width, weighs 1.23 kg, and contains an average of 138.4 seeds. Each seed weighs 1.51 g and measures 2.56 cm in length, 1.27 cm in width, and 0.78 cm in thickness. The fruit appears dull brown when harvested and has a woody appearance. It is hard and tough to open. The edible part of the fruit is composed of fibrous sarcotestas surrounding the seeds. This pulp is aromatic and has an intense orange color. The fruit skin is thick, rough, hard, and heavy, comprising 32.25% of the total weight. After being cut, the fruit ripens in 3 to 7 days in the shade at an average temperature of 25°C, causing the green color to fade and turn reddish-brown. The richness in the Annonacea family of secondary metabolites has led to numerous conclusive studies on the cytotoxic power of acetogenins, compounds contained in chincuya seeds [2].
Germination. Germination occurs when the seed imbibes and results in DNA transcription, translation, nucleic acid repair, cell elongation, and division. These processes induce protein and enzyme synthesis, activation of growth regulators, and mobilization of reserves, culminating in radicle protrusion. Recent findings indicate that reactive oxygen species and microRNAs also regulate seed dormancy and germination [3]. It's suggested that Chincuya might utilize an altered pathway for seed development. The initial phase of seed development is marked by significant mitotic activity alongside the accumulation of cytokinins, gibberellins, spermines, spermidines, and polyamines. During the intermediate stages, cell expansion and auxin production take precedence, accompanied by an increase in free and bound gibberellins; meanwhile, cytokinin levels drop, and abscisic acid remains undetectable. In the final phases, the seed undergoes preparation for dehydration. It enters a dormant state characterized by increased abscisic acid, reduced fresh weight, and peak accumulation of reserves, including Late Embryogenesis Abundant (LEA) proteins [4]. Thus, when seed maturation, both from morphological and physiological perspectives, concludes and the accumulation of dry matter ceases, the seed has reached maturity [5,6].
Dormancy. A viable seed is dormant when it cannot germinate despite being in favorable conditions. The morphological dormancy type is attributed to embryonic immaturity, a term repeatedly used to clarify why germination does not occur. Therefore, studies related to embryo growth must be carried out to demonstrate it [7)). Vidal-Lezama et al. [8] showed that chincuya seeds have morphological dormancy. The warm-dry storage treatment overcame this latency by causing morphological changes in the embryo and its growth.
Physiological dormancy. Primary dormancy or quiescence is obtained during seed ripeness and broken by environmental and endogenous factors [9]. Physiologically, dormant seeds are water-permeable and do not germinate; the embryo is impaired to grow. However, as dormancy interruption occurs, the growth potential increases to the point that germination is possible [7].
Morphophysiological dormancy. In this case, a combination of morphological and physiological dormancy causes delayed germination for more than 30 days [10]. Radicle protrusion in A. crassiflora seeds occurs 150 days after planting [11]. In Virola surinamensis (Rol.) Warb., seeds germinated between 94 and 124 days after planting [12]. It was determined that A. crassiflora Mart. shows morphophysiological dormancy, which disappears at low temperatures or when the temperature fluctuates before the rainy season [11]. The same authors point out that subsequent embryonic growth and endosperm digestion are probably controlled by gibberellins synthesized while overcoming physiological dormancy. In the case of A. macroprophyllata Donn. Smith, different authors agree that germination capacity increases when seeds are stored between 5 and 7 months [13,14,15]. Morphophysiological dormancy release happens when, in the imbibed seed, the embryo grows and differentiates. This type of dormancy is common in regions with humid seasonal climates worldwide [16].
Effect of warm-dry storage. After ripe seed dispersal, seed dormancy is released after an extended storage period under dry conditions. This process is called after-ripening. Several studies have shown that after-ripening is effective on seeds with moisture content between 5 and 18 % [17]. After-ripening treatments decrease dormancy and increase germination potential. Germination potential changes through a two-pronged pathway: increasing sensitivity to factors that promote germination, such as light and gibberellins, and decreasing sensitivity to factors that inhibit germination [17]. The available reports indicate that few gene expression changes occur after ripening. Seed dormancy release results from chemical transformations unrelated to normal cellular metabolic processes and affects the metabolic products present in the freshly produced seeds [9]. These authors report that non-enzymatic reactions play a role in seed dormancy release after ripening, like the production of reactive oxygen species. However, a significant decrease in the concentration of abscisic and salicylic acids and an increase of gibberellins, jasmonic acid, and isopentenyladenine was observed when imbibing Arabidopsis seeds treated under dry storage [18].
Effect of gibberellins. Gibberellins stimulate cell elongation and division. Biosynthesis of these substances occurs throughout the plant, including in germinating embryos and developing seeds. They can act in cells that do not produce them, leading to their migration from the production site to the action site. Gibberellins, which are endogenous, control their biosynthesis by altering gene transcription [19]. In imbibed seed embryos, gibberellins are produced, and they initiate the production of hydrolytic enzymes. These enzymes soften the seed coat, mobilize the nutrient reserves in the seed, promote embryo growth and the elongation of the hypocotyl, trigger meristem activity, and support the growth of new shoots and roots [20]. The process of breaking seed dormancy involves mechanisms that both degrade abscisic acid and encourage the biosynthesis of gibberellins [17]. AG1, AG20, and AG53 gibberellins were identified in A. purpurea in seeds stored for 3 to 4 months [21]. In the same way, in A. macroprophyllata and A. purpurea, abscisic acid and gibberellins were found in fresh seeds imbibed in water and unsoaked seeds [22]. Vidal et al. [13] and Ferreira et al. [22] have determined the promoting effect of gibberellic acid on germination in different species of Annonaceae at concentrations close to 350 ppm.
Lipids as a reserve substance. The high lipid content in seeds of some species could indicate a compensatory selection (greater energy/volume) towards the lightest or smallest seeds for better dispersal [23]. It has been mentioned that storage lipids are assembled into tiny spheres of 0.5 – 2.5 μm called oil or lipid bodies [24,25], which contain a triacylglycerol nucleus covered by a phospholipid layer with proteins (oleosin, caleosin, and steroleosin). These proteins are responsible for the stability of lipid bodies and prevent triacylglycerol storage from degrading until the seed germinates [26]. In oilseeds, lipases initiate the degradation of triacylglycerols in lipid bodies; these hydrolytic enzymes produce glycerol and free fatty acids in glyoxysomes. Glycerol enters the glycolytic pathway, which can also become pyruvate and then oxidize in the mitochondria through the Krebs Cycle. Then, oxidation reactions could degrade fatty acids to produce compounds containing fewer carbon atoms.
The primary oxidative process is β-oxidation. However, there are differences in the degradation routes of the fatty acid based on its degree of saturation. Yet, all must be converted to forms that can be degraded to form acetyl-CoA. Then, this acetyl-CoA is captured by the glyoxylate cycle and enters the gluconeogenic pathway in the cytoplasm to produce sugars [27,28]. Seeds with oil reserves, such as sunflower and canola seeds, limit germination and establishment of seedlings when the isocitrate lyase activity is inhibited, an essential enzyme during the glyoxylate cycle that plays a vital role in lipid metabolism. In the case of cereals, it is known that abscisic and gibberellic acids control lipid mobilization through the inhibition or induction of the isocitrate lyase activity [3]. Research has shown that for six grass species, the starch, protein, and fat reserves in their seeds did not impact the percentage or speed of germination. However, the levels of soluble sugars and protein did affect these factors. Additionally, there were no changes in lipid reserves throughout the germination process. The different composition of fatty acids means different susceptibility to peroxidation. In most plant species with oil-rich seeds, there is a risk of self-oxidation, where the degree of unsaturation significantly influences the degree of degradation [29]. The hypothesis proposed is that due to the morphological characteristics of the embryo and phylogenetics of the species, chincuya seeds present morphophysiological latency, which will be demonstrated by the increase in germination capacity and modifications in the content of existing fatty acids caused by dry storage conditions. Therefore, this study aimed to determine the effects of warm-dry storage and application of gibberellins on germination behavior and the results of warm-dry storage on lipid content in chincuya seeds.
2. Materials and Methods
2.1. Plant materials
In Las Salinas, Chicomuselo, Chiapas (15.74539 N, -92.28321 E), the ripe fruits were collected from different trees of tolerated vegetation in September and October. Seeds were extracted from these fruits. Initial moisture content was measured at extraction time in 4 repetitions of 10 seeds each. The fresh and dry weights (oven-dried at 110 °C for 10 days) from extracted seeds were obtained by analytical balance and recorded. The dry weight was divided by the fresh weight and multiplied by 100 to get the moisture percentage in the seeds [30]. The initial moisture content was 39%, but the seeds were kept in the laboratory, partially covered with black plastic, for 39 days until the moisture content reached 25%. At that moment, the dried seeds were put into a black plastic bag and placed in an incubator at a constant temperature (25 ± 3 °C), in darkness, and without added fungicides. This point marks the start of the warm-dry storage treatments.
2.2. Viability test
The stain test was conducted with 2,3,5-triphenyl-tetrazolium chloride to determine seed viability. Seeds with fully stained embryos and specifically marked unstained parts were considered viable according to the standards [30]. Annonaceae seeds have no staining reference standard, and due to the response diversity in staining, the following proposal was made (Figure 1). Seeds were observed with a magnifier and an integrated light. To interpret the test, only the stained portion of the embryo was considered, and endosperm staining was ignored as it stains intensely, making the test reading difficult (Figure 1). Only categories A, B, and C were considered viable seeds. Data was converted to percentages. Tetrazolium (Merck®) was prepared at 0.1 %, and 4 repetitions of 20 seeds were used. Seeds were conditioned by imbibing the whole seeds in distilled water for 24 hours at room temperature. The staining preparation consisted of removing the seed coat with tweezers. In the cleaned seed, the first transversal cut was made to shorten the length of the seed by half. The second cut was longitudinal, starting the cut in the hilum area without separating the halves, trying to cut deeply up to half of the seed, allowing the application of the solution and guaranteeing that if the embryo is alive, it will be stained. They were placed in Petri dishes with the tetrazolium solution and were kept in darkness at 30 °C for 24 hours. Subsequently, the solution was decanted, and the seeds were rinsed with distilled water. Throughout the observation, they were kept immersed in distilled water.
2.3. Treatments
Treatments that correspond to the warm-dry storage periods were defined as follows: when the seed was freshly extracted from the fruit, partially covered with black plastic, for 39 days, until 25 % moisture content and then, they were put into a black plastic bag and placed in an incubator at a constant temperature of 25 ± 3 ° C, in darkness and without adding fungicides; thus initiating warm-dry storage treatments, by 3, 6, 9, and 12 months of warm-dry storage. The control (time 0) was not stored under warm-dry conditions.
Seeds were treated with reagent-grade gibberellic acid (Merck®) at 350 mg L-1 and distilled water (control). Both treatments were applied by immersion for 72 hours at 25 °C with aeration for 10 min every 12 hours. Treatment immersion was applied to 4 repetitions of 20 seeds each. Before planting, seeds were soaked in 10 % (v/v) sodium hypochlorite solution (Cloralex®) for 3 min, followed by soaking in 10 % ethyl alcohol for 2 min and rinsed in distilled water. Seed incubation followed, with some modifications, the protocol by [27]. Absorbent paper towels were saturated with distilled water, and seeds were placed on them. The paper was rolled and placed into a partially closed plastic bag; seeds were continuously watered manually as needed. The plastic bags were put in a growth chamber (Shel Lab LI15 ®) with alternating temperatures of 30 °C during the day and 25 °C at night and 12:12 light:dark photoperiod. Throughout the test period, which lasted 40 days, the absorbent paper towels were changed three times, and washes in fungal preventive solutions were repeated during each change. Successful germination was considered when the radicle was visible (approx. 1 mm). Dormant seeds kept the initial appearance as when set to germinate, with no testa changes in odor, color, size, or texture. That is, they maintained the same appearance from the beginning to the end of the test, while rotten seeds showed signs of rot, such as loss of firmness and unpleasant odor. The studied variables were germination percentage and dormant and dead seeds percentage. To analyze lipids, three samples from each treatment were evaluated. They were deep-frozen at -20 ° C, then lyophilized for 14 days, and finally milled. They were kept in amber bottles, at a cool temperature, and in the shade until the analysis by gas chromatography.
2.4. Extraction of lipids
Lipid extraction was carried out according to the procedure by Reyes-Trejo et al. [31] followed. A. purpurea seed kernels (0.5 g) were placed in a 50 mL Erlenmeyer flask, and 20 mL of hexane were added. The samples were kept in maceration for three days. The procedure was repeated three times (20 mL × 3); subsequently, after the extraction was complete, the solvent was removed under reduced pressure using a rotary evaporator (BüchiR111, Waterbath B-461, Switzerland) at 45 °C until a light-yellow oily residue from both kernels, and their weights were recorded. For the analysis of fatty acid (FA) content in the seeds, FA methyl esters were prepared through a transesterification reaction in a basic medium. A mixture of KOH (1.0 g) was prepared in methanol (60 mL), and then 1.0 mL of this dilution was added to approximately 0.1 g of each sample of the obtained oil by maceration. The mixture was heated at 65 °C for 2 h with continuous stirring. This was followed by a conventional procedure consisting of separating phases, washing the resulting methyl esters with 0.1% aqueous citric acid to remove the basic catalyst, washing with water, and finally drying with anhydrous sodium sulfate. The excess methanol was eliminated under vacuum distillation to obtain yellow oil samples, and its weights were recorded to obtain the yield of this new Fatty Acid Methyl Esters (FAME). The calculation method to establish fatty acids composition was followed as published previously by Marroquin-Andrade et al. [32]: area percentage. Certified by the standard ISO 9001:2000 RSGC 238 with the following conditions: column AT- FAME 30m x 0.25mm x 0.25um film thickness; injector temperature 250 °C Temperature FID 250 ° C; oven 180 ° C [15 min] 10 °C /min to 230 ° C [3 min]; hydrogen flow 1.8 mL/min. H2 Split 100; temperature detector 275 ºC and injector 250 °C
2.5. Statistical analysis
The statistical analysis employed an analysis of variance of a completely randomized design and Tukey’s multiple comparison test, with a significance level of 5 %. Data on percentages were transformed before the analysis with the following equation: . The values reported in the Tables are the actual values. The SAS statistical program was used [33]. Simple linear regressions were also developed between storage treatments and concentrations of fatty acids.
3. Results and Discussion
3.1. Viability
The highest viability values corresponded at 6 and 9 months of storage, these results were significantly different. A sudden drop in viability occurred on the twelfth month of storage (Figure 2). Vidal-Lezama et al. noted that embryos grew during warm-dry storage until the sixth month [8], coinciding with the viability data observed here. Similarly, González-Esquinca et al. found that A. macroprophyllata seed viability did not change throughout the 8-month storage evaluation [11]. The difference in germination percentages is significant between freshly extracted seeds and those stored for three months, as shown in Table 1. After three months of storage, the germination percentage nearly doubles. The positive relationship between viability and germination remains for up to six months in dry storage. It might be possible that the viability test only in embryos (Figure 5) did not allow the detection of any level of deterioration, shown as lipid peroxidation in the endosperm [30].
3.2. Germination
The treatments favored final germination; consequently, dormant seeds decreased (Table 1). The coefficients of variation are high, which reflects the genetic variability of the species, given its limited domestication. The dead seeds percentage shows a highly erratic behavior. It was high in the twelfth month of storage, which might be explained by limited seed viability (Figure 2). After-ripening is adequate in seeds with moisture contents between 5–18 % [17]; however, chincuya seeds exceeded these values. Table 2 confirms the positive effect of applying gibberellic acid in germination. This same behavior has been observed in several Annona species (A. muricata, A. squamosa, A. cherimola, A. macroprophyllata, A. crassiflora, A. emarginata, and atemoya (Annona x atemoya Mabb.), including A. purpurea [13,15,17,21,22,34]. Until now, only exogenous applications of gibberellic acid, cytokines, and ethylene have been reported in Annonaceae [34]. In A. crassiflora germination, gibberellin promoted the production and reactivation of several hydrolytic enzymes involved in using endosperm reserves [12].
Storage increased the viability of the chincuya seed (Figure 2). The germination increase by the dry storage period might occur because of changes in sensitivity to gibberellin or germination inhibitors [17]. Imbibed chincuya seeds contain gibberellins and abscisic acid [21,22]. Also, Bewley et al. mentioned that imbibed seeds synthesize gibberellins [20]. Bazin et al. discovered that the release of dormancy in post-ripened sunflower seeds is linked to the availability of free water and the movement of molecules within the seeds' embryonic areas [35]. These authors suggest that changes in the bonding properties of water, resulting from oxidative processes, enable metabolic activities. Other mechanisms that might explain the after-ripening effect include non-enzymatic reactions that eliminate germination inhibitors, reactive oxygen species, and antioxidants [36], membrane alterations [37], and specific protein degradation through the proteasome [38]. Taiz and Zeiger mention that developing seeds synthesize gibberellins [19], and following the results found by Vidal-Lezama et al., who pointed out that chincuya embryos grow during warm-dry storage [8], it is possible to assume a continuum in the synthesis of gibberellins (although not exclusively) in the seed.
On the other hand, the location of the species in the phylogenetic tree and its relationship with dormancy should be noted. According to Chatrou, the Annonaceae family is very ancient [38], and Forbis et al. recognized that primitive angiosperms have seeds with tiny embryos and abundant endosperm around them [39]. Likewise, other researchers mentioned that morphophysiological dormancy is a plesiomorphic character [40]. However, some seeds have no dormant period [41]; for example, the seed may delay germination in Annona after its morphological and physiological ripening. In contrast with other authors, Duke considers the absence of dormancy as a primitive character [42]. Using phylogenetic analysis tools for 216,000 legume observations worldwide, researchers found that seeds without dormancy evolved in climates with long growing seasons and in families that produce larger seeds [43].
On the other hand, dormancy matches families of species of temperate origin with smaller seeds. When the favorable growing season is short, dormancy is the only adaptation and survival strategy. Previous research indicates that most non-dormant species are in tropical forests [10,16].
Considering the above information, the sampled chincuya trees grow in a sub-humid warm climate (Chicomuselo, Chiapas, Mexico.). In the preceding years, according to the sampling recollection, rainfall was mainly concentrated between May and October (1 331.3 - 2 432.2 mm), up to 89.2% of the annual rainfall. September was the rainiest month, with an average of 27.4 % of the rainy months. The large seed size is due to the species' habitat [8] compared to other Annona species and rainfall seasonality. The functional strategy of the seeds is likely to maintain metabolic activity at a lower rate than when they were inside the fruit and attached to the tree; for this, the dormancy manifests as a germination delay. At the same time, they acquire the germination capacity as time passes and the rainy season arrives. Chincuya seeds naturally remain in the soil for seven months and germinate when the first rains of May come. Results of Table 1 pointed out that six-month storage is the best time to reach germination, and in natural conditions, seeds remain for a remarkably similar time. Likewise, the temperature during the applied storage was similar (25 ° C) to that of their natural condition, where the 4-year average was 23.8 ° C.
3.3. Fatty acids content
The analysis of fatty acids revealed high lipid values in the seeds (Table 3). When oxidized, the lipid reserves of seeds produce more than twice as much energy per volume unit as the hydrolysis of proteins or carbohydrates [6], which results in an advantage for the germination process. Unsaturated fatty acids (UFA) were slightly higher than saturated fatty acids (SFA). The concentration of oleic acid as UFA and palmitic acid as SFA are highlighted. It is known that oleic acid plays a vital role in the anabolism of plant fatty acids as the precursor of the primary unsaturated fatty acids. Their high content can then be explained. Table 4 shows that chincuya seeds have more fatty acids than other species. For palmitic and stearic acids, chincuya seeds had the highest values.
In A. purpurea, Pérez-Amador et al. identified the same fatty acids in this research, except for arachidic and palmitoleic acids [44]. The concentrations reported by these authors were palmitic acid at 26.38 %, stearic acid at 3.7 %, oleic acid at 43.67 %, and linoleic acid at 26.25 %; oleic acid concentration was the only coincidence. Arachidic acid is only reported in sugar apples (A. squamosa) (Table 4), and the content is higher in chincuya. Palmitic acid is not mentioned in other species, except for soursop (A. muricata), which has a similar concentration. The relation between UFA/SFA (1.22) in chincuya seeds (Table 3) shows that the total of saturated and unsaturated acids is not entirely different; they are balanced, in contrast to what is observed in other species of the genus Annona (Table 4). Figure 6 shows no significant changes in the studied fatty acids content in response to storage time.
The fatty acid content did not decrease significantly after 12 months (Figure 3). However, it is known that deterioration can occur due to prolonged storage, causing oxidation of the fatty acids present in the seed, as it is pointed out in Tabebuia roseoalba (Ridl.) Sandwith seeds stored for 24 months [48]. The different composition of fatty acids means different susceptibility to peroxidation. High saturated-fatty acid contents speed the deterioration process of recalcitrant seeds during their storage in American oak (Quercus rubra L.) [49] and mahogany (Swietenia macrophylla King) [50]
Table 5 shows the values of the fatty acids evaluated in the freshly extracted seeds (without storage) but without having been placed under germination conditions (intact) and newly extracted seeds but imbibed and incubated to promote germination, but in which no radicle protrusion was observed. Statistically meaningful differences are seen in the content of stearic and total fatty acids.
These data and those already mentioned indicate that the incubated seeds reactivated their metabolism using fat reserves even when they did not show radicle protrusion. In physiological dormancy, seeds are water-permeable, and the inhibiting mechanism is in the embryo. Seeds do not germinate because the embryo has a physiological impairment that causes low growth potential. However, as seed dormancy is released, the potential increases to the point that germination is possible [7]. Holdsworth et al. [9] have indicated that the release of physiological dormancy results from chemical transformations, which affect the metabolic products present in the seeds.
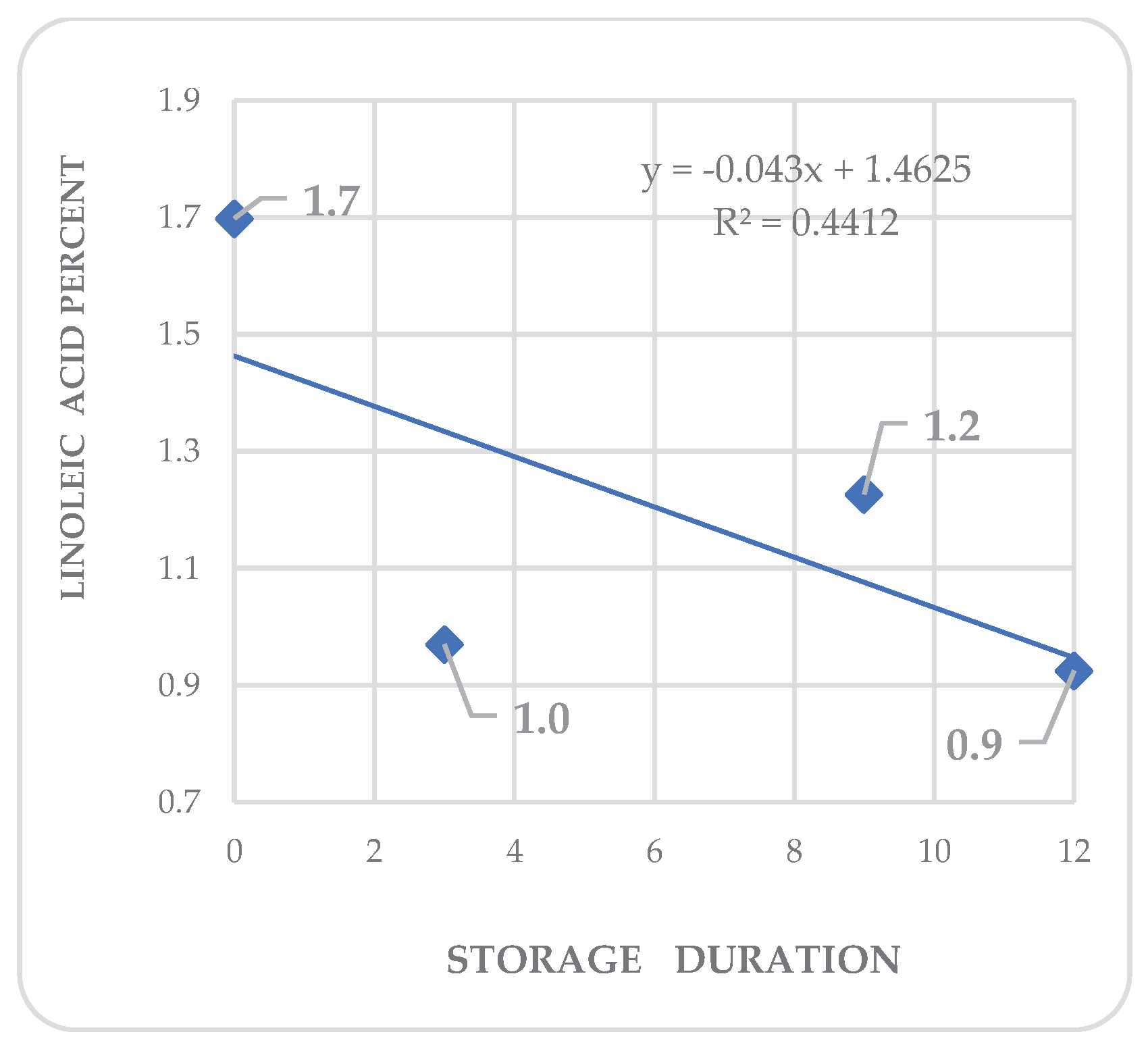
Regression analysis shows trends in differential variations depending on the acid. Fatty acid kinetics reveal that palmitic (Figure 4) and palmitoleic acids (Figure 6) contents increase as the storage time increases, while oleic acid (Figure 5) has a different behavior. In the three cases, R2 values are high for these trends. Also, stearic acid (Figure 8) displays an increasing tendency as storage time increases, but the corresponding R2 value is outstandingly low. On the other hand, arachidic (Figure 7) and linoleic acids (Figure 9) show a downward trend as the storage period increases; however, R2 values are also low.
Figure 4.
Kinetics of palmitic acid in Annona purpurea (chincuya) seeds, in response to warm-dry storage.
Figure 4.
Kinetics of palmitic acid in Annona purpurea (chincuya) seeds, in response to warm-dry storage.
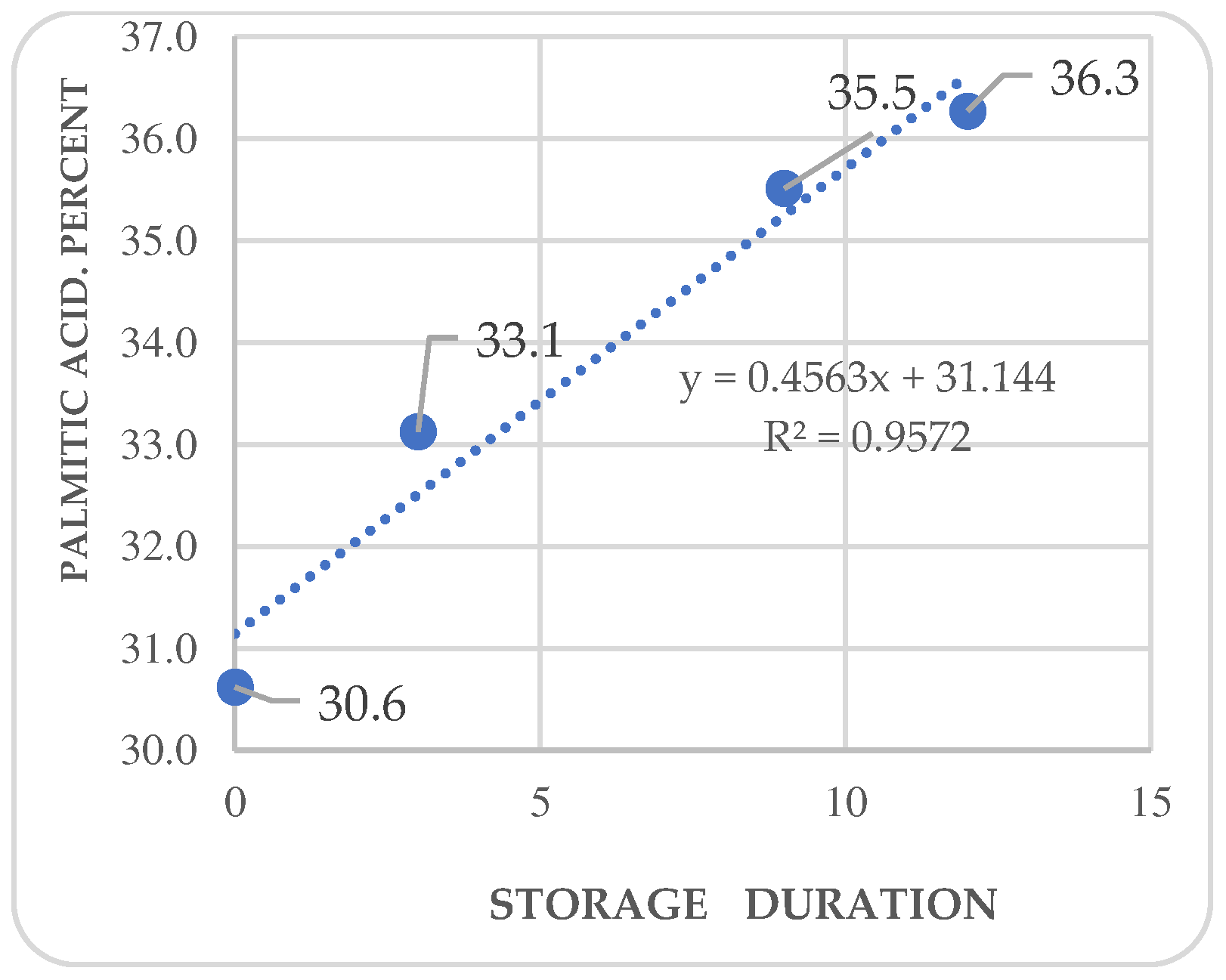
Figure 5.
Kinetics of oleic acid in Annona purpurea (chincuya) seeds, in response to warm-dry storage.
Figure 5.
Kinetics of oleic acid in Annona purpurea (chincuya) seeds, in response to warm-dry storage.
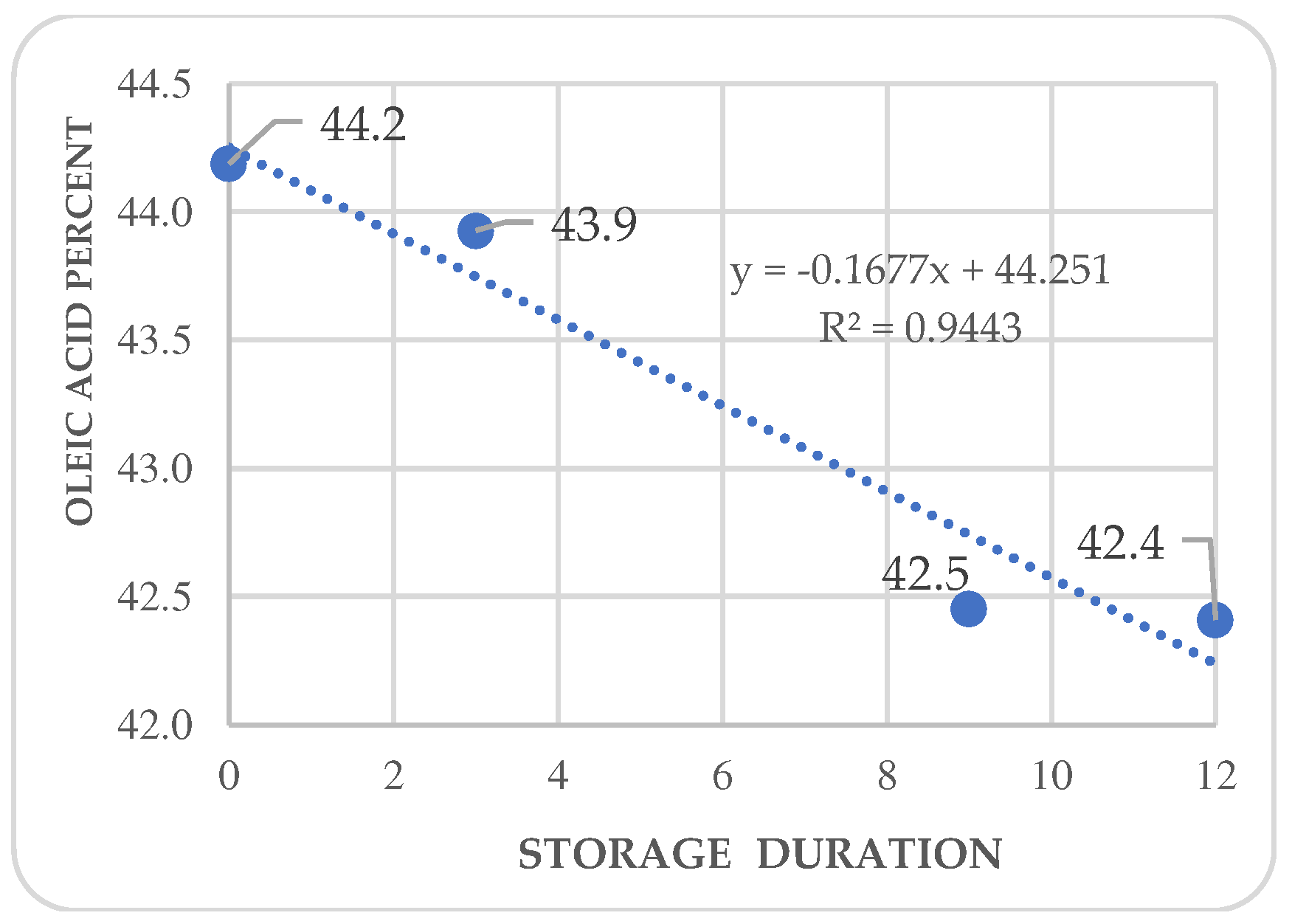
Figure 6.
Kinetics of palmitoleic acid in Annona purpurea (chincuya) seeds, in response to warm-dry storage.
Figure 6.
Kinetics of palmitoleic acid in Annona purpurea (chincuya) seeds, in response to warm-dry storage.
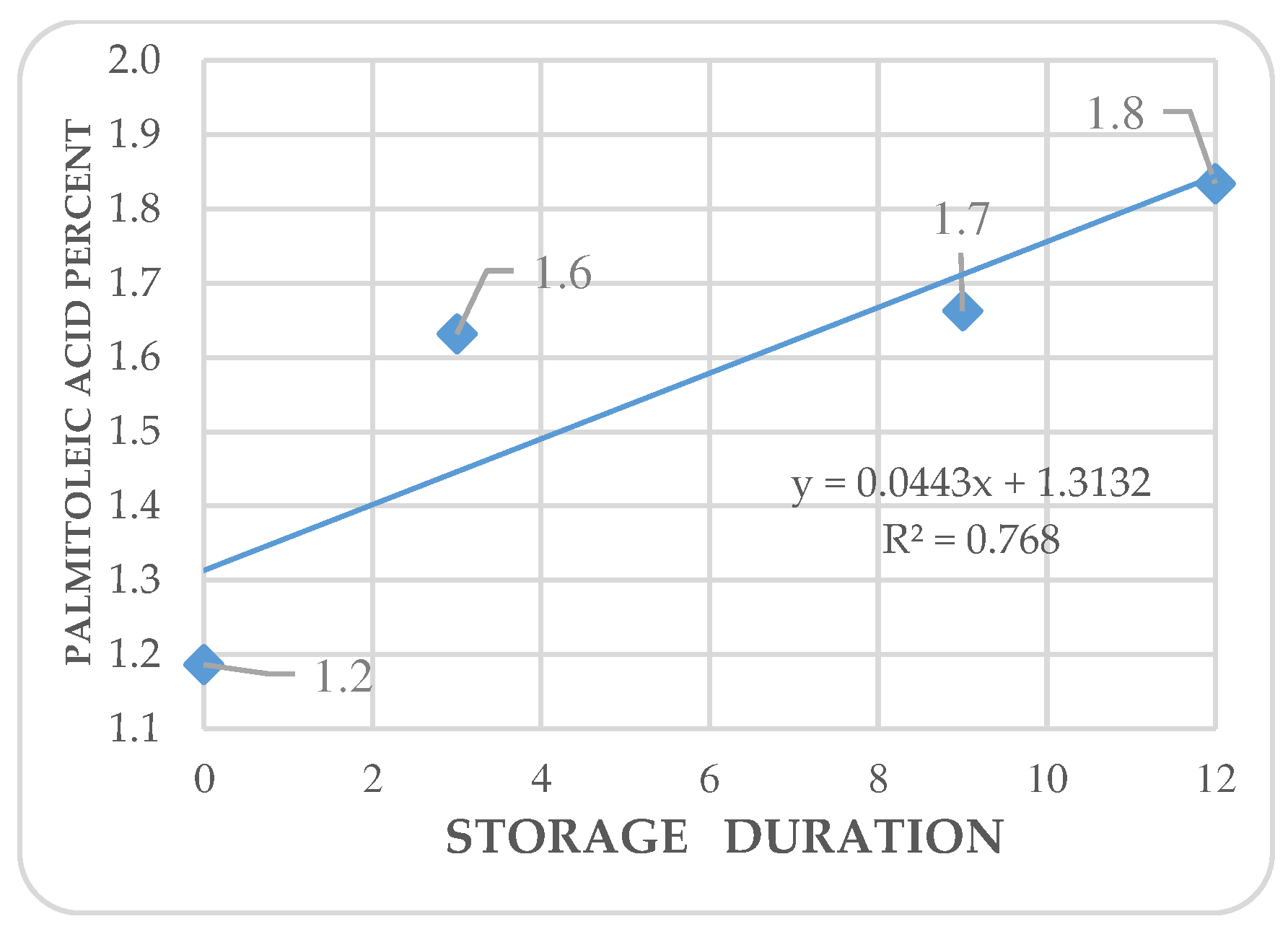
Figure 7.
Kinetics of arachidic acid in Annona purpurea (chincuya) seeds, in response to warm-dry storage.
Figure 7.
Kinetics of arachidic acid in Annona purpurea (chincuya) seeds, in response to warm-dry storage.
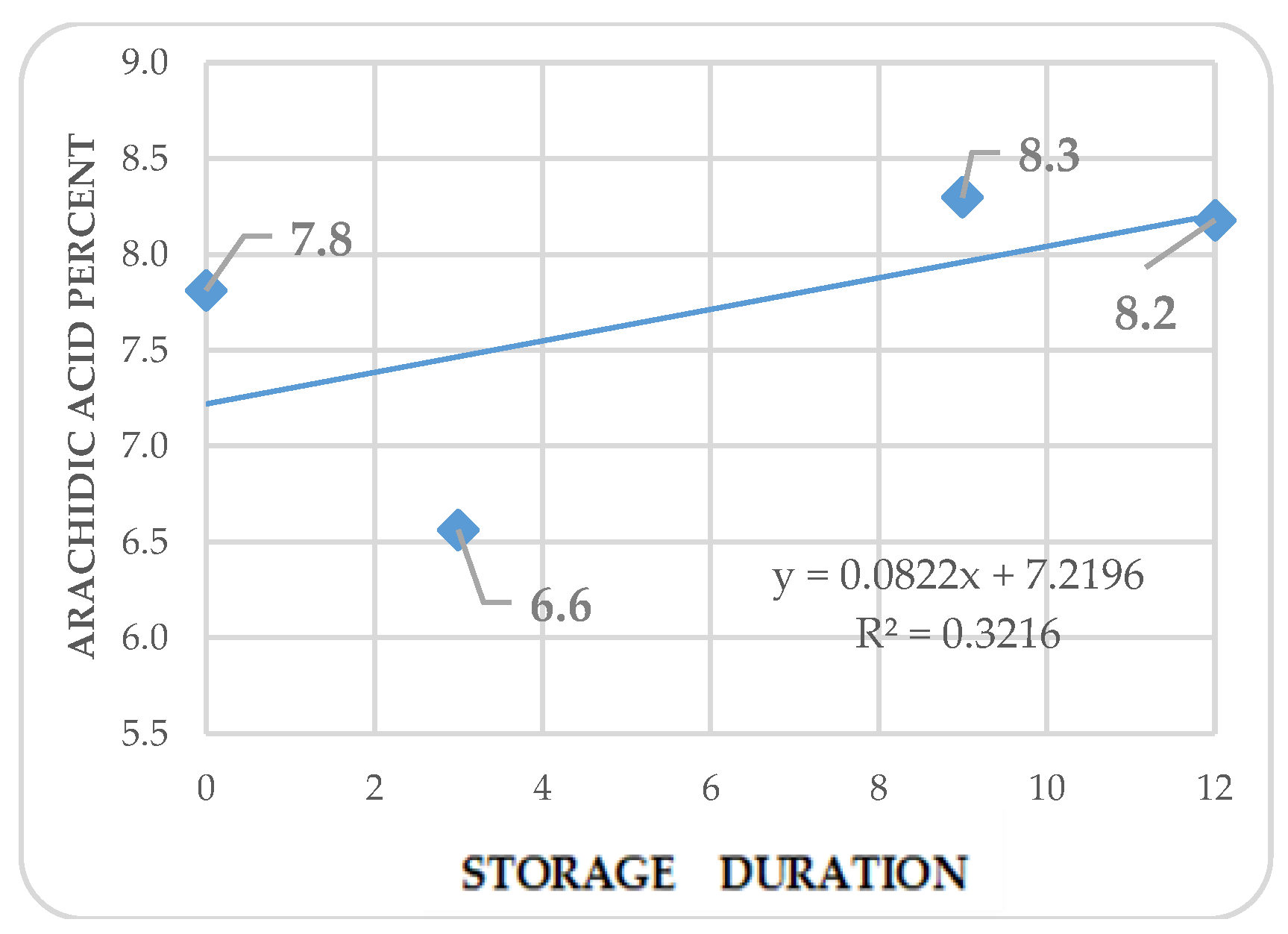
Figure 8.
Kinetics of stearic acid in Annona purpurea (chincuya) seeds, in response to warm-dry storage.
Figure 8.
Kinetics of stearic acid in Annona purpurea (chincuya) seeds, in response to warm-dry storage.
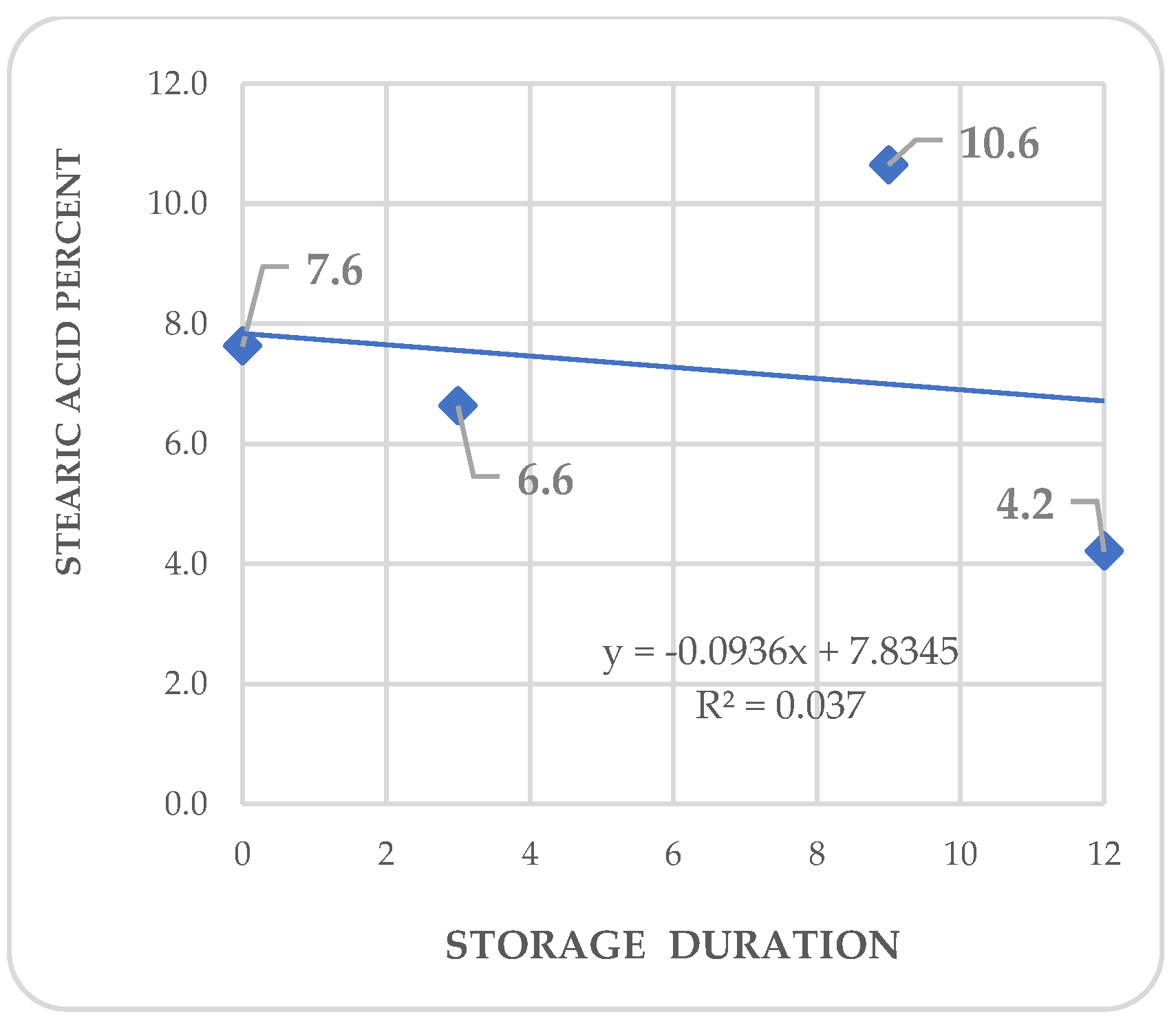
Figure 9.
Kinetics of linoleic acid in Annona purpurea (chincuya) seeds, in response to warm-dry storage.
Figure 9.
Kinetics of linoleic acid in Annona purpurea (chincuya) seeds, in response to warm-dry storage.
Expectations about future research. From the human nutrition and energy sustainability points of view, fatty acids content in the chincuya seed is elemental because the number of seeds per fruit is high; it may be feasible to promote its cultivation for biofuel production. The chincuya seed values in saturated and unsaturated fatty acids are high. Given that the consumption of unsaturated fatty acids supports vascular health, consumption as a source of edible oil could be feasible. However, developing the necessary tests to determine safety is essential due to the acetogenin content and other metabolites recognized as cytotoxic [2].
4. Conclusions
The fatty acids identified were palmitic acid, stearic acid, oleic acid, linoleic acid, and, for the first time, arachidic acid and palmitoleic acid. The use of gibberellic acid and warm-dry storage improved germination, alongside the observed change in fatty acid content during such storage. Coupled with previously reported findings on morphological transformations and embryonic growth resulting from the same storage conditions, it is confirmed that Annona purpurea seeds exhibit morphophysiological dormancy.
Author Contributions
Conceptualization, E.V. L.; methodology, E.V.L.; B.R.T.; A.V.M.; A.R.P.; software, H.V.V.; A.M.P.; validation, E.V.L.; A.V.M.; B.R.T.; analysis, H.V.H.; G.F.; investigation, A.M. P.; G.F.; A.M.P.; resources, E.V.L.; A.V.M.; data curation, H.V.H.; A.R. P.; writing—original draft preparation, E.V.L.; writing—review and editing, A.V.M.; B.R.T.; G.F.; visualization, A.M.P.; supervision, A.V.M.; project administration, E.V.L.; funding acquisition, A.R.P.; A.V.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Colegio de Postgraduados and Universidad Autónoma Chapingo (2016 Project. General Management of Research and Postgraduate Studies).
Institutional Review Board Statement
Not applicable.
Data Availability Statement
The data supporting this study's findings are available from the corresponding author upon reasonable request.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Cabrera, C.E.F., Hernández, M. E., H., Salvador, F. J., Salazar, G. C. Annonaceae de la Península de Yucatán. Taxonomía, Florística y Etnobotánica. Etnoflora Yucatenense. 2004. Fascículo 21. José Salvador Flores (Ed.). Universidad Autónoma de Yucatán. Facultad de Medicina Veterinaria y Zootecnia. pp.64.
- Vidal-Lezama, Eloísa; Villegas-Monter, Ángel; Vaquera-Huerta, Humberto; Robledo-Paz, Alejandrina; Martínez-Palacios, Alejandro. Annona purpurea Moc. & Sessé ex Dunal especie nativa de México, subutilizada. AgroProductividad, 2019, 12, 9-15.
- Li Z.; Gao Y.; Zhang Y.; Lin C.; Gong D.; Guan Y.; Hu J. Reactive oxygen species and gibberellin acid mutual induction to regulate tobacco seed Germination. Frontiers Plant Science, 2018, 9, 1279.
- Matilla, A. Germinación y dormición de las semillas. In Fundamentos de Fisiología Vegetal. Azcón-Bieto, J.; Talón, M.. McGraw-Hill Interamericana. Madrid, España, 2000; pp. 435-450.
- Sano, N.; Rajjou, L.; North, H. M.; Debeaujon, I.; Marion-Poll, A.; Seo, M. Staying alive: molecular aspects of seed longevity. Plant and Cell Physiology, 2015, 57, 660-674. 57. [CrossRef]
- Costa, M. C. D.; Cooper, K.; Hilhorst, H. W.; Farrant, J. M. Orthodox seeds and resurrection plants: Two of a kind. Plant physiology, 2017, 175, 589-599. [CrossRef]
- Baskin, C. C.; Baskin, J. M. Seeds: ecology, biogeography, and evolution of dormancy and germination. 2a ed; Elsevier, 2014; 14.
- Vidal-Lezama, E. , Villegas-Monter A., Vaquera-Huerta, H., Robledo-Paz A., Martínez-Palacios, A., Ferreira, G. Morphometry of chincuya seeds (Annona purpurea Moc. & Sessé ex Dunal) and embryonic growth under dry warm storage. Revista Brasileira de Fruticultura, 2023. v.45, e-042. [CrossRef]
- Holdsworth, M. J.; Bentsink, L.; Soppe, W. J. Molecular networks regulating Arabidopsis seed maturation, after-ripening, dormancy and germination. New Phytologist, 2008, 179, 33-54. [CrossRef]
- Fogliani, B.; Gateblé, G.; Villegente, M. Fabre, L.; Klein, N.; Anger, N.; Baskin, C. C.; Seutt, C. P. The morphophysiological dormancy in Amborella trichopoda seeds is a pleisiomorphic trait in angiosperms. Annals of Botany, 2017, 119, 581-590.
- Cruz, E.; Barros, H. Germinação de sementes de espécies amazônicas: ucuúba [Virola surinamensis (Rol. ex Rottb.) Warb]; Embrapa: Amazônia Oriental, Brasil 2016; 4.
- Da Silva, E. A.; De Melo, D. L.; Davide, A. C., De Bode, N.; Abreu, G. B.; Faria, J. M.; Hilhorst, H. W. Germination ecophysiology of Annona crassiflora seeds. Annals of botany, 2007, 99, 823-830. [CrossRef]
- Vidal-Lezama, E.; Marroquín-Andrade, L.; Gómez, R. S. Efecto del almacenamiento y tratamientos pregerminativos en semillas de Annona muricata L., Annona diversifolia Saff. y Annona spp. Proc. Inter. Soc. for Trop. Hort, 2008, 52, 203-209.
- González-Esquinca, A. R.; De-La-Cruz-Chacón, I.; Domínguez-Gutú, L. M. Dormancy and germination of Annona macroprophyllata (Annonaceae): the importance of the micropylar plug and seed position in the fruits. Botanical Sciences, 2015, 93, 509-515. [CrossRef]
- Ferreira, G.; De-La-Cruz-Chacón, I.; González-Esquinca, A. R. Overcoming seed dormancy in Annona macroprophyllata and Annona purpurea using plant growth regulator. Revista Brasileira de Fruticultura, 2016, 38, e-234. [CrossRef]
- Fenner, M.; Thompson, K. The ecology of seeds. Cambridge University Press, New York,USA, 2005.
- Finch-Savage, W. E.; And Leubner-Metzger, G. Seed dormancy and the control of germination. New Phytologist, 2006, 171, 501-523.
- Yano, R.; Kanno, Y.; Jikumaru, Y.; Nakabayashi, K.; Kamiya, Y.; Nambara, E. CHOTTO1, a putative double APETALA2 repeat transcription factor, is involved in ABA-mediated repression of gibberellin biosynthesis during seed germination in Arabidopsis. Plant Physiol, 2009, 151, 641-654.
- Taiz, L. And Zeiger, E. Plant Physiology. 5a ed.; Sinauer Associates Inc., Sunderland, UK, 2010. 2010.
- Bewley, J.; M. Black. Seeds: Physiology of Development and Germination. Plenum Press, New York and London, 1994.
- Gómez-Castañeda, J. A.; Ramírez, H.; Benavides-Mendoza, A.; Encina-Rodríguez, I. Germination and seedling development of soncoya (Annona purpurea Moc & Sessé) in relation to gibberellins and abscisic levels. Revista Chapingo Serie Horticultura, 2003, 9, 243-253.
- Ferreira, G. Reguladores vegetais na superação da dormência, balanço hormonal e degradação de reservas em sementes de Annona diversifolia Saff. e A. purpurea Moc. & Sessé ex Dunal (Annonaceae). Tese (Livre-Docência). 2011. Universidade Estadual Paulista, Instituto De Biociências. Campus de Botucatu, Sao Paulo, Brasil;.
- Baskin, C. C.; Baskin, J. M. Seeds. Ecology, Biogeography, and Evolution of Dormancy and Germination. Academic Press: San Diego, California, 1998.
- Zienkiewicz, A.; Zienkiewicz, K.; Rejón, J. D.; De Dios Alché, J., Castro, A. J.; Rodríguez-García, M. I. Olive seed protein bodies store degrading enzymes involved in mobilization of oil bodies. Journal of Experimental Botany, 2013, 65, 103-115. 65. [CrossRef]
- Murphy, D. J. The dynamic roles of intracellular lipid droplets: from archaea to mammals. Protoplasma, 2012, 249, 541-585. [CrossRef]
- Poxleitner, M.; Rogers, S. W.; Samuels, A. L.; Browse, J.; Rogers J. C. A role of caleosin in degradation of oil-body storage lipids during seed germination. The Plant Journal, 2006, 47, 917-933. 47. [CrossRef]
- Bewley, J. D.; Black, M. Physiology and biochemistry of seeds in relation to germination: volume 2: viability, dormancy, and environmental control. Springer Science & Business Media, 1982.
- Azcón-Bieto, J.; Talón, M. Fundamentos de Fisiología Vegetal. McGraw-Hill Interamericana: Madrid, España; 2000.
- Zhao, M.; Zhang, H.; Yan, H.; Qiu, L.; Baskin, C. C. Mobilization and role of starch, protein, and fat reserves during seed germination of six wild grassland species. Frontiers Plant Science, 2018, 9, 234. 9.
- International Seed Testing Association. International Rules for Seed Testing. 2013, ISTA, Zurich, Suiza.
- Reyes-Trejo, B., Guerra-Ramírez, D., Zuleta-Prada, H., Cuevas-Sánchez, J. A., Reyes, L., Reyes-Chumacero, A., & Rodríguez-Salazar, J. A. Annona diversifolia seed oil as a promising non-edible feedstock for biodiesel production. Industrial Crops and Products, 2014, 52, 400-404. [CrossRef]
- Marroquín-Andrade, L., Cuevas-Sánchez, J.A., Guerra-Ramírez, D., Reyes, L., Reyes Chumacero, A., Reyes-Trejo, B.,. Proximate composition, mineral nutrient and fatty acids of the seed of ilama, Annona diversifolia Saff. Sci. Res. Essays, 2011 6, 3089–3093.
- SAS Institute Inc., SYSTEM 2000® Software: Product Support Manual, Version 1, First edition, 2000, SAS Institute Inc., Cary, NC;; 293.
- Ferreira, G.; De-La-Cruz-Chacón, I.; Boaro, C. S. F.; Baron, D.; Lemos, E. E. P. D. Propagation of Annonaceous plants. Revista Brasileira de Fruticultura, 2019, 41, e-500.
- Bazin, J.; Batlla, D.; Dussert, S.; El-Maarouf-Bouteau, H.; Bailly, C. Role of relative humidity, temperature, and water status in dormancy alleviation of sunflower seeds during dry after-ripening. Journal of Experimental Botany, 2010, 62, 627-640. 62. [CrossRef]
- Bailly, C. Active oxygen species and antioxidants in seed biology. Seed Science Research, 2004. 14, 93-107.
- Hallett B. P.; Bewley J. D. Membranes and seed dormancy: beyond the anaesthetic hypothesis. Seed Science Research, 2002, 12, 69-82. [CrossRef]
- Skoda, B.; Malek, L. Dry pea seed proteasome. Plant physiology,1992, 99, 1515-1519. [CrossRef]
- Chatrou, L.W. The Annonaceae and the Annonaceae project: a brief overview of the state of affairs. Acta Hortic., 1999, 497, 43-58. [CrossRef]
- Forbis, T. A.; Floyd, S. K.; And De Queiroz A. The evolution of embryo size in angiosperms and other seed plants: implications for the evolution of seed dormancy. Evolution, 2002, 56, 2112-25.
- Sautu, A.; Baskin, J. M.; Baskin, C. C.; Deago, J.; Condit, R. Classification and ecological relationships of seed dormancy in a seasonal moist tropical forest, Panama, Central America. Seed Science Research, 2007, Cambridge, , 17, 127-140. [CrossRef]
- Duke, J. A. On tropical tree seedlings I. Seeds, seedlings, systems, and systematics. Annals of the Missouri Botanical Garden, Missouri, 1969, 56, 125-161.
- Rubio De Casas, R., Willis, C. G., Pearse, W. D., Baskin, C. C., Baskin, J. M.; Cavender-Bares, J. Global biogeography of seed dormancy is determined by seasonality and seed size: a case study in the legumes. New Phytologist, 2017, 214, 1527-1536. [CrossRef]
- Pérez-Amador, M. C.; González-Esquinca, A.; García-Argaez, A.; Bratoeff, E.; Labastida, C. Oil composition and flavonoid profiles of the seeds of three Annona species. Phyton., 1997, 61, 77-80.
- Abbade, L. C.; Takaki, M. Biochemical and physiological changes of Tabebuia roseoalba (Ridl.) Sandwith (Bignoniaceae) seeds under storage. Journal of Seed Science 2014, 36, 100-107. [CrossRef]
- Sun, W. Q. State and phase transition behaviors of Quercus rubra seed axes and cotyledonary tissues: Relevance to the desiccation sensitivity and cryopreservation of recalcitrant seeds. Cryobiol., 1999, 38, 372-385. [CrossRef]
- Gómez T. J.; Jasso-Mata J. J.; Vargas-Hernández J. J.; Soto-Hernández R. M. Deterioro de semilla de dos procedencias de Swietenia macrophylla King., bajo distintos métodos de almacenamiento. Ra Ximhai, 2006, 2, 223-239.
- Solís-Fuentes, J. A.; Amador-Hernández, C.; Hernández-Medel, M. R.; Durán-De-Bazúa, M. C. Caracterización fisicoquímica y comportamiento térmico del aceite de "almendra" de guanábana (Annona muricata, L). Grasas y aceites, 2010, 61, 58-66.
- Branco, P. C.; Castilho, P. C.; Rosa, M. F.; Ferreira, J. Characterization of Annona cherimola Mill. seed oil from Madeira Island: a possible biodiesel feedstock. Journal of the American Oil Chemists' Society, 2010, 87, 429-436. [CrossRef]
- Yathish, K. V.; Omkaresh, B. R.; Suresh, R. Biodiesel production from custard apple seed (Annona squamosa) oil and its characteristics study., IJERT, 201510, 2, 1938-1942.
Figure 1.
Staining pattern to assess seed viability. Where: A. Viable. The embryo is stained intensely (+++) throughout and evenly. B. Viable. The embryo is stained, but the staining in the hypocotyl root axis is more intense (+++) than in cotyledons (++). C. Viable. The embryo is stained, but the staining in the hypocotyl root axis is less intense (++) than in B. D. Not viable. The embryo is stained (++), but the hypocotyl root axis remains unstained. E. Not viable. The embryo is not stained.
Figure 1.
Staining pattern to assess seed viability. Where: A. Viable. The embryo is stained intensely (+++) throughout and evenly. B. Viable. The embryo is stained, but the staining in the hypocotyl root axis is more intense (+++) than in cotyledons (++). C. Viable. The embryo is stained, but the staining in the hypocotyl root axis is less intense (++) than in B. D. Not viable. The embryo is stained (++), but the hypocotyl root axis remains unstained. E. Not viable. The embryo is not stained.

Figure 2.
Annona purpurea (chincuya) seed viability is due to warm-dry storage.
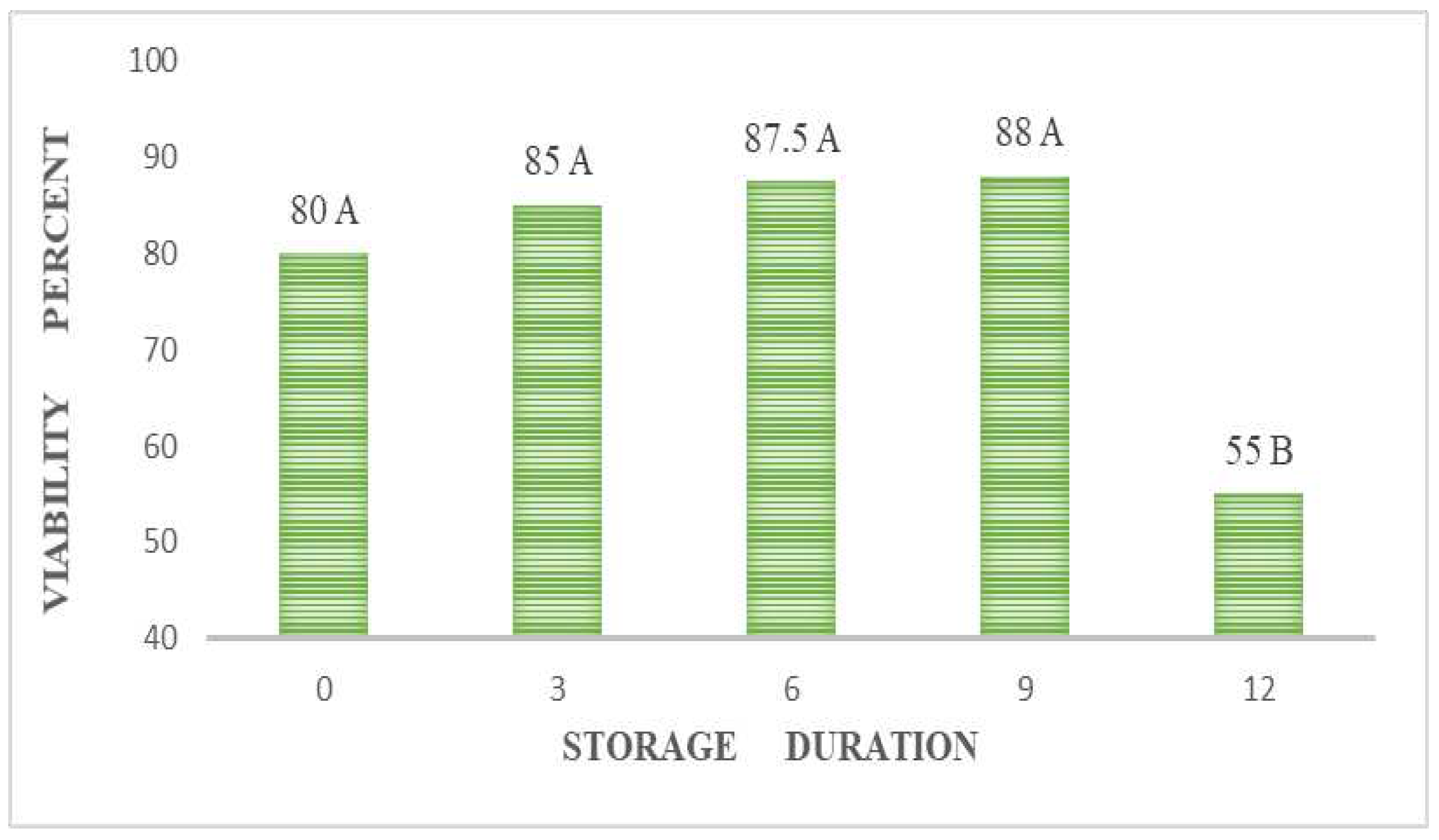
Figure 3.
Kinetics of total fatty acids in Annona purpurea (chincuya) seeds in response to warm-dry storage. Means with the same letter between columns are not significantly different, at p ≤ 0.05 according to the Tukey test. LSD = Least significant difference.
Figure 3.
Kinetics of total fatty acids in Annona purpurea (chincuya) seeds in response to warm-dry storage. Means with the same letter between columns are not significantly different, at p ≤ 0.05 according to the Tukey test. LSD = Least significant difference.
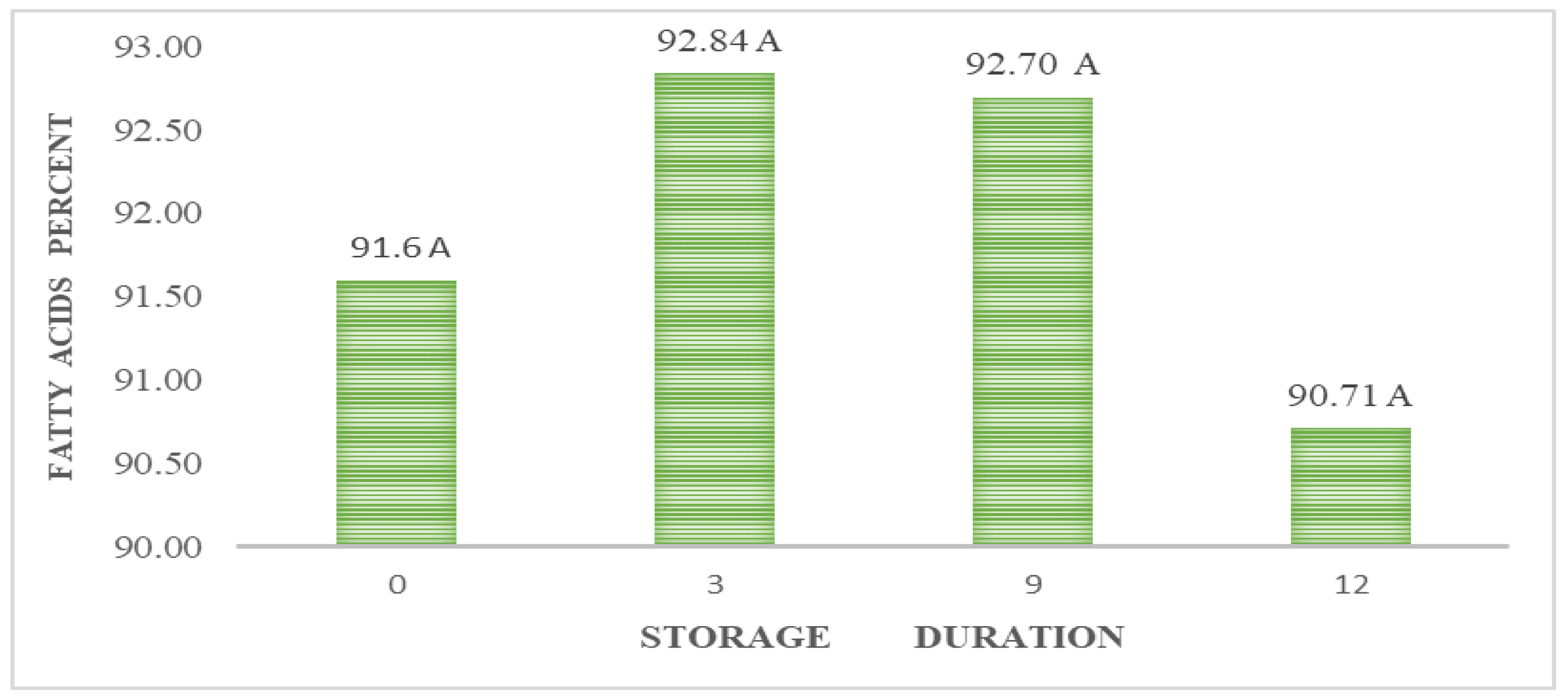

Table 1.
Effect of warm-dry storage on the germination of Annona purpurea (chincuya) seeds.
| Storage duration (months) |
Germinated seeds (%) |
Dormant seeds (%) |
Dead seeds (%) |
|---|---|---|---|
| 0 | 26.12 B | 59.13 A | 14.75 C |
| 3 | 52.16 A | 25.50 B | 22.33 BC |
| 6 | 65.62 A | 18.13 B | 16.25 C |
| 9 | 13.75 BC | 43.13 AB | 43.12 B |
| 12 | 2.00 C | 19.50 B | 78.50 A |
| LSD | 14.48 | 28.91 | 21.52 |
Means with the same letter between columns are not significantly different, at p ≤ 0.05 according to the Tukey test. LSD = Least significant difference.
Table 2.
Effect of imbibing Annona purpurea (chincuya) seeds in gibberellic acid on germination behavior.
Table 2.
Effect of imbibing Annona purpurea (chincuya) seeds in gibberellic acid on germination behavior.
| Treatment | Germination (%) |
Latent Seeds (%) |
Dead Seeds (%) |
|---|---|---|---|
| Gibberellic acid | 40.25 A | 34.54 B | 25.19 A |
| Control | 17.80 B | 52.25 A | 29.93 A |
| LSD | 5.456 | 10.896 | 8.109 |
Means with the same letter between columns are not significantly different, at p ≤ 0.05 according to the Tukey test. LSD = Least significant difference.
Table 3.
Fatty acid content of seeds of Annona purpurea (chincuya).
| Fatty acid percent | ||||||||
|---|---|---|---|---|---|---|---|---|
| Saturated | Unsaturated | |||||||
| Palmitic Acid (C16:0) | Estearic Acid (C18:0) | Arachidic Acid (C20:0) | Total Satured Fatty Acid (SFA) | Oleic Acid (C18:1) |
Linoleic Acid (C18:2) |
Palmitoleic Acid (C16:0) | Total Unsatured Fatty Acids (UFA) |
UFA: SFA ratio |
| 33.41±6.02 | 7.72±1.20 | 1.30±0.57 | 42.44 | 43.37±3.67 | 7.24±7.18 | 1.52±0.52 | 52.15 | 1.22 |
Table 4.
Fatty acid percentage of four seeds of the Annona genus.
| Fatty acid | Soursoup (%) (A. muricata) Z |
Cherimola (%) (A. cherimola) Y |
Ilama (%) (A. macroprophyllata) X |
Sugar apple (%) (A. squamosa) W |
|---|---|---|---|---|
| Palmitic | 25.5 | 19.99 | 16.4 | 17.79 |
| Palmitoleic | 1.5 | -- | --- | -- |
| Estearic | 6.0 | 4.16 | 5.22 | 4.29 |
| Oleic | 39.5 | 38.58 | 70.42 | 39.72 |
| Linoleic | 27.0 | 36.97 | 7.97 | 29.13 |
| Arachidic | ---- | -- | -- | 1.06 |
| Relationship U:S U | 2.44 | 2.21 | 3.62 | 2.97 |
Table 5.
The fatty acid content of Annona purpurea (chincuya) seeds: recently extracted, without incubating (intact) and recently extracted, incubated (imbibed).
Table 5.
The fatty acid content of Annona purpurea (chincuya) seeds: recently extracted, without incubating (intact) and recently extracted, incubated (imbibed).
| Percentage of fatty acids | |||||||
|---|---|---|---|---|---|---|---|
| Seed Condition |
Palmitic | Palmitoleic | Stearic | Oleic | Arachidic | Linoleic | TOTAL |
| Recently extracted without incubating (intact) |
30.62 Az |
1.71 A |
7.81 A |
47.05 A |
1.69 A |
12.75 A |
98.15 A |
| Recently extracted incubated (imbibed) latent | 29.61 A | 1.18 A | 6.03 B | 44.19 A | 0.98 A | 7.62 A | 91.60 B |
| LSD | 9.00 | 0.82 | 1.60 | 6.17 | 0.77 | 11.51 | 5.92 |
Z According to the Tukey test, means with the same letter between columns are not significantly different, at p ≤ 0.05. LSD = Least significant difference.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



