
Submitted:
07 February 2024
Posted:
08 February 2024
You are already at the latest version
Abstract
This study aimed to investigate the effect of potassium poly-γ-glutamate (PGA-K) on mice fed a high-fat diet consisting of 60% of total calories for a duration of 12 weeks. PGA-K administration reduced the increase in body weight, epididymal fat, and liver weight caused by a high-fat diet compared to the obese group. The triglyceride, low-density lipoprotein cholesterol and high-density lipoprotein cholesterol levels, which are blood lipid indicators, were significantly increased in the obese group but were significantly decreased in the PGA-K-treated group. The administration of PGA-K resulted in a significant inhibition of pro-inflammatory cytokines, including tumor necrosis factor α and interleukin 6. Moreover, the levels of leptin and insulin, which are insulin resistance indicators, significantly increased in the obese group but were significantly decreased in the PGA-K-treated group. These results suggest that PGA-K exhibits a protective effect against obesity induced by a high-fat diet, underscoring its potential as a candidate for obesity treatment.
Keywords:
Potassium Poly-γ-glutamate
; Chungkookjang
; Obesity
; Inflammation
; Insulin resistance
1. Introduction
Obesity is recognized as a worldwide epidemic and is linked to a spectrum of health issues, such as fatty liver disease, type 2 diabetes mellitus (T2DM), hypertension, dyslipidemia, cardiovascular diseases, gallbladder disease, stroke, myocardial infarction, osteoarthritis, gout, mental disorders, and specific types of cancers [1,2]. Obesity is characterized by the conversion of excess energy from increased food intake into triglycerides (TGs), which are subsequently stored in adipose tissue, resulting in weight gain [3].
Obesity is associated with the expansion of fat cells (adipocytes) and inflammation in the surrounding adipose tissues [4]. Obesity-induced inflammation occurs because of continuous lipid accumulation within adipose tissues [5]. The release of pro-inflammatory factors from adipose tissue actively contributes to insulin resistance and increases the risk of metabolic disorders associated with obesity [6].
Several studies have suggested that natural products sourced from edible and medicinal plants have anti-obesity effects with minimal to no side effects [7]. Various studies have been undertaken to hinder the accumulation of lipids and the synthesis of pro-inflammatory cytokines using various food materials [8]. Ingredients commonly found in functional foods including curcumin, quercetin, resveratrol, extracts from Camellia sinensis, and green tea have been identified for their ability to mitigate lipid accumulation and inflammation associated with metabolic diseases such as obesity and hypertension [9]. In situations of excessive fat accumulation, dysfunctional expanded adipocytes release different pro-inflammatory adipokines, including interleukin (IL) 6 and IL-1β and tumor necrosis factor (TNF) α [10]. The expansion of adipocytes also increases adipocyte death and recruitment of macrophages to adipose tissue [11]. During this process of adipocyte remodeling, numerous crown-like structures are formed, characterized by macrophages surrounding dead adipocytes [12]. An increased prevalence of crown-like structures is associated with inflammation in adipose tissue [13].
Fermented soybean-based foods contain a diverse array of bioactive compounds that have potential therapeutic effects, particularly for metabolic disorders [14]. Chungkookjang, a traditional Korean fermented soybean product, is produced by fermenting boiled soybeans with rice straw, naturally incorporating Bacillus subtilis into the fermentation process [15]. During the fermentation of chungkookjang, soy protein decomposes into amino acids through the action of potent proteolytic enzymes produced by B. subtilis. This not only enhances digestibility but also contributes to an increase in the vitamin B2 and calcium content of the final product [16]. The primary component found in the viscous mucous substance produced during fermentation is poly-γ-glutamic acid (γ-PGA), along with modified isoflavone compounds. PGA is recognized for its health benefits as it contributes to the absorption of calcium in the body [17,18]. As the primary physiologically active substance in soybeans, isoflavones play a crucial role in enhancing the absorption and bioavailability of nutrients derived from fermented soybeans [19]. The fermentation process of chungkookjang, leads to the generation of enzymes and diverse physiologically active substances that are not present in raw soybeans. These include dietary fibers, phospholipids, isoflavones, phenolic acids, saponins, trypsin inhibitors, and phytic acids [20]. These components play a role in preventing atherosclerosis, heart disease and inflammation mediated by oxidative stress, obesity, diabetes, senile dementia, cancer, and osteoporosis [21]. Moreover, these components exhibit various beneficial activities including lipid-lowering, blood pressure-lowering, thrombolytic, anti-mutagenic, immunostimulatory, anti-atopic dermatitis, anti-allergic, anti-androgenetic, antibacterial anti-alopecia, and anti-asthmatic effects, as well as properties conducive to skin enhancement [22]. This study seeks to investigate the impact of γ-PGA potassium salt (PGA-K) produced by B. subtilis chungkookjang on obesity associated with inflammation induced by a high-fat diet (HFD) mouse model and to assess the potential of PGA-K for obesity treatment.
2. Materials and Methods
2.1. Preparation of PGA-K
PGA-K was prepared by B. subtilis chungkookjang. B. subtilis chungkookjang (KCTC 0697BP) was inoculated into a preparative basic medium [GS basic medium with 5% l-glutamic acid, 5% glucose, 1%, (NH4)2SO4, 0.27% KH2PO4, 0.42% Na2HPO4·12H2O, 0.05% NaCl 0.05%, pH 6.8] and cultured at 150 rpm stirring rate, an aeration rate of 1vvm, and 37°C for 36 h. Then, 2N hydrochloric acid was added to the solution and stand at 10°C for 12 h to obtain γ-PGA precipitate. After 12h, the solution was filtered through a Nutsche filter, and the filtered PGA precipitate was thoroughly washed with distilled. γ-PGA has a molecular mass of 1–15,000 kDa, and separate experiments were performed on subfractions with different molecular masses. To obtain γ-PGA potassium salt, γ-PGA was solubilized in 5N KOH. The molecular mass of PGA-K was determined by gel permeation chromatography. Briefly, PGA-K solution was diluted with 0.1 M NaNO3 and injected into the gel permeation chromatograph equipped with a ViscoGel GMPWXL xl column (7.8 mm × 30 cm; Viscotek, Houston, TX, USA), which was equilibrated with 0.1 M NaNO3 at 40°C with a flow rate of 0.8 mL/min, and a Viscotek LR25 laser refractometer (Viscotek).
2.2. Animals
Four-week-old specific pathogen-free-grade male C57BL/6 mice were purchased from Damul Science (Daejeon, Korea) and acclimated for 1 week. During the study, the mice were accommodated in a mouse cage under controlled conditions of a 12-hour light/dark cycle at a temperature of 22°C ± 2°C and a relative humidity of 55% ± 5%. All experimental procedures were conducted with approval from the Animal Care Committee of Jeonju AgroBio-Materials Institute, Jeonju, Korea (approval number: JAMI IACUC 2023003) prior to the commencement of the study.
2.3. Experimental Groups
The mice were divided into five groups, normal group (N), the high-fat diet-induced obesity group (HFD), the positive control group (PC), and the PGA-K administration group (PGA-K), each consisting of 7 ~ 8 mice. To induce obesity, the HFD, PC, and PGA-K groups were fed a 60% kcal fat diet for 12 weeks, and the N group was fed a normal diet (10% kcal fat). The PC and PGA-K groups were orally administered garcinia (300 mg/kg) and PGA-K (100 mg/kg and 200 mg/kg), respectively, for 12 weeks. N and HFD groups were administered vehicle (distilled water). The N and PC groups were consisted 7 mice per group, and the HFD and PGA-K groups were consisted 8 mice per group.
2.4. Evaluation of Biomarkers in Serum
Using the ELISA kit from R&D Systems (Abingdon, UK), the amounts of TNF-α, IL-6, and leptin in serum were determined. Concentrations of Triglyceride (TG), total cholesterol (TC) and high-density lipoprotein (HDL) were determined using the kit provided by Asan Pharm (Seoul, Korea), and low-density lipoprotein (LDL) and insulin levels were measured using kits provided by CrystalChem (Elk Grove, CA, USA). The manufacturer’s instructions were followed for all the measurements.
2.5. Histology
Mouse epididymal adipose and liver tissues were fixed in 4% paraformaldehyde and embedded in paraffin. Tissue sections, with a thickness of 4 μm, were subjected to staining using Oil Red O (for liver tissues) and hematoxylin and eosin (H&E) (for adipose tissues).
2.6. Statistical Analyses
The data are presented as means ± standard deviation, and all statistical analyses were conducted using Sigmaplot v16.0 (Systat Software Inc., San Jose, CA, USA). Statistical analysis was performed to identify differences, followed by one-way analysis of variance and Duncan’s multiple comparison test. These analyses were employed to assess variations among three or more groups across all measured parameters. Statistical significance was defined as a difference with p < 0.05.
3. Results
3.1. Effects of PGA-K Administration on Body Weight and Food Intake in HFD-Fed Mice
There were no significant differences observed in the initial body weight across the groups. From the 1st week of the experiment, the HFD group exhibited a notable rise in body weight compared to the N group. After 12 weeks, the final body weight of the HFD group showed a significant increased to 44.26 ± 3.6 g, which is about 1.4 times that of the N group, while the PGA-K administration group had a relatively low increase in body weight, and the 100 mg/kg PGA-K group showed a statistically significant effect (Figure 1A and 1C). Epididymal white adipose tissue (eWAT) weight also significantly increased in the HFD group compared to the N group after 12 weeks, but the PGA-K group showed a decrease in eWAT weight (Figure 1D).
Moreover, the eWAT/body weight ratio was decreased by treatment with 100 and 200 mg/kg PGA-K (Figure 1E). Throughout the experimental period, no significant differences noted in food intake across the various experimental groups (Figure 1B). Hence, it can be inferred that PGA-K has therapeutic potential for HFD-induced obesity.
3.2. PGA-K Prevents Adipogenesis and Lipid Accumulation in HFD-Fed Mice
For histological analysis, eWAT tissue was stained with H&E. In the epididymal fat of obese mice, adipocytes were abnormally enlarged and some adipocytes were damaged (Figure 2A). In the group administered with doses of 100 and 200 mg/kg of PGA-K, adipocyte heterogeneity and hypertrophy were reduced compared to the HFD group (Figure 2A). To evaluate whether PGA-K inhibits HFD-induced hepatic steatosis, histological analysis was performed on Oil Red O-stained liver sections. The liver tissue of the HFD group showed numerous red lipid droplets with incomplete cellular structure and fat accumulation, which suggested that substantial accumulation of lipid occurred in the liver tissue (Figure 2B). On the other hand, both groups administered 100 and 200 mg/kg of PGA-K showed decreased cell deformation and lipid accumulation in liver tissue (Figure 2B). These results showed that PGA-K had a preventive effect on adipocyte hypertrophy and intrahepatic lipid accumulation in HFD-induced obese mice.
3.3. Effects of PGA-K on Serum Lipid Profiles in HFD-Fed Mice
The serum TG, TC, HDL, and LDL levels of mice in each group were measured using ELISA. Compared to those in the N group, the levels were significantly increased in the HFD group. After PGA-K administration for 12 weeks, the levels of TG (Figure 3A), TC (Figure 3B), HDL (Figure 3C), and LDL (Figure 3D) decreased.
3.4. Effects of PGA-K on Liver Damage in HFD-Fed Mice
To test whether PGA-K inhibits liver damage, along with serum levels of alanine aminotransferase (ALT) and aspartate aminotransferase (AST) were quantified. HFD-induced obese mice had significantly increased liver weight and serum ALT and AST levels. Liver weight was decreased in the PGA-K 100 mg/kg group (Figure 4A), and serum ALT was significantly decreased in both PGA-K 100 and 200 mg/kg groups (Figure 4B). In the case of serum AST levels, the average value seemed to decrease only in the PGA-K 100 mg/kg group. Nevertheless, no significant difference was observed between the HFD and PGA-K groups (Figure 4C).
3.5. Effects of PGA-K on HFD-Induced Pro-Inflammatory Cytokines
To understand the mechanisms underlying obesity-induced lipid metabolism and inflammatory responses, we measured the levels of inflammatory cytokines. Persistent inflammation in obesity is characterized by abnormal expression of genes encoding pro-inflammatory cytokines [23]. Elevated levels of pro-inflammatory cytokines, such as TNF-α and IL-6, occur concomitantly with an increase in lipid content in white adipose tissue (WAT), contributing to the development of complications associated with obesity [24]. In this study, TNF-α and IL-6 levels were measured in serum using ELISA. The levels of serum TNF-α and IL-6 were notably elevated in the HFD group compared to the normal diet group (N). After administration of PGA-K for 12 weeks, TNF-α (Figure 5A) and IL-6 production (Figure 5B) were decreased.
3.6. Effects of PGA-K on HFD-Induced Insulin Resistance
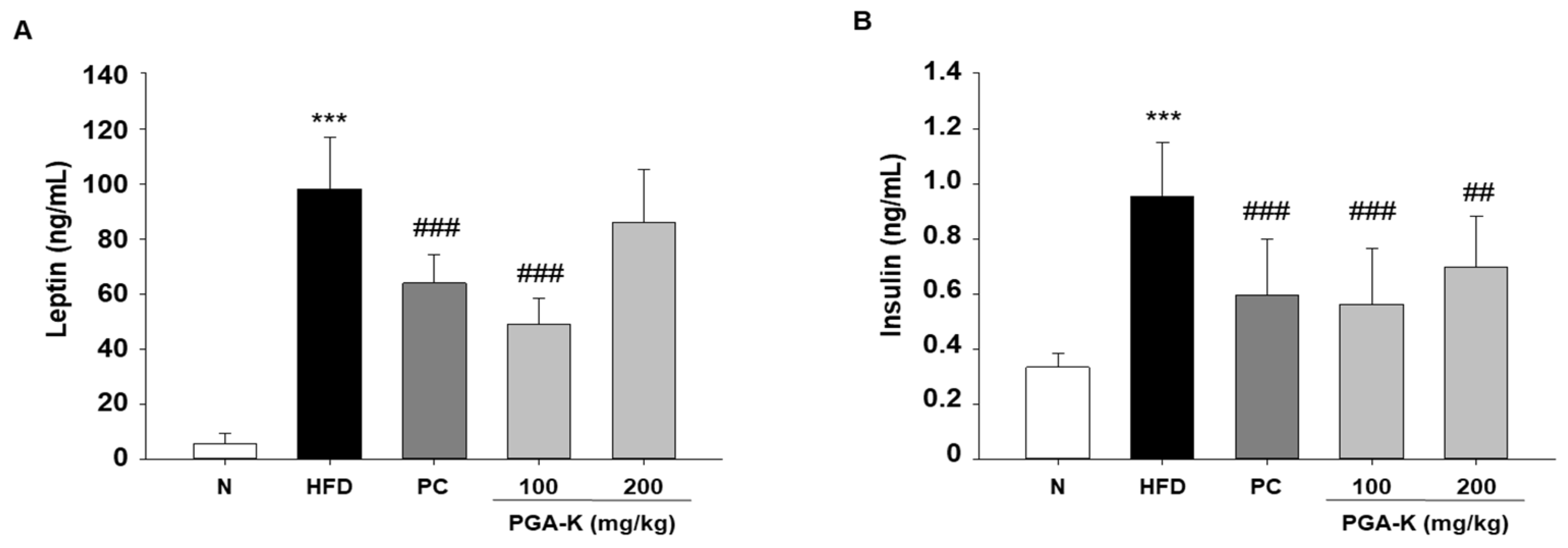
The production of pro-inflammatory molecules by adipose tissue plays an active role in fostering insulin resistance and elevating the susceptibility to metabolic diseases associated with obesity [25]. The induction of systemic low-grade inflammation by obesity increases the risk of developing T2DM [26]. In addition, insulin resistance in obese individuals is often accompanied by hyperinsulinemia, which is associated with obesity, dyslipidemia, and glucose intolerance [27]. To examine whether PGA-K is effective against insulin resistance, we determined the levels of leptin and insulin in serum using ELISA. HFD significantly increased leptin and insulin levels. After administration of PGA-K for 12 weeks, the production of leptin (Figure 6A) and insulin (Figure 6B) decreased.
4. Discussion
Excessive body fat accumulation delineates the hallmark of obesity. Inflammation induced by obesity arises from the accumulation of lipids in adipose tissue [28]. This inflammatory process is characterized by an augmentation in both the quantity (hyperplasia) and size (hypertrophy) of adipocytes, ultimately resulting in the expansion of adipose tissue mass [29]. Adipose tissue functions as a biological calorie reservoir, expanding in response to an abundance of nutrients and storing lipids during periods of energy surplus, then releasing them during energy deficiency [30]. Nevertheless, excessive fat accumulation and dysfunction in adipocytes result in changes in plasma lipid and lipoprotein levels. This includes an increase in LDL cholesterol levels and TG, along with a decrease in HDL cholesterol levels, contributing to the development of obesity-related disorders [31].
This study provides scientific evidence supporting the anti-obesity function of PGA-K in HFD-fed mice, with a focus on adipose tissue dysfunction. The results of this study indicate that PGA-K significantly reduced HFD-induced body weight, especially adipose tissue hypertrophy. PGA-K administration to HFD-induced obese mice significantly reduced relative liver size and fat pad size. The relative fat pad size of HFD-induced obese mice administered PGA-K was reduced to the level of mice fed a normal fat diet. Although no significant effects of PGA-K on food intake were observed, body weight gain and white adipose fat mass were lower in PGA-K-treated mice than in untreated mice. These findings suggest that alterations in caloric intake do not necessarily correspond to changes in adipose tissue mass. In addition, PGA-K ameliorated HFD-induced obesity-related adipose tissue disorders, such as elevated inflammation, indicating its ability to attenuate obesity-related metabolic disorders. Moreover, PGA-K reduced the levels of insulin resistance markers, including insulin and leptin. Thus, B. subtilis chungkookjang-derived PGA-K is a potential candidate for reducing and managing obesity and obesity-related diseases.
The field of natural product research is continuously expanding, with ongoing efforts aimed at enhancing the biological activities of these compounds to address diverse diseases through treatment and prevention. Despite significant research efforts into the pathogenesis of obesity and ongoing exploration of novel treatment strategies, a definitive cure remains elusive. Treatment medications for obesity frequently have potential side effects and complications. Several anti-obesity medications, including liraglutide, naltrexone-bupropion, orlistat, and phentermine-topiramate have been approved by the US Food and Drug Administration (FDA) [32]. Meanwhile, numerous stimulant-type weight loss medications, such as phentermine and diethylpropion, are typically recommended for short-term use because of the risk of dependence and other potential side effects [33]. Certain drugs used in the clinical treatment of obesity are associated with adverse effects, including nausea, insomnia, constipation, gastrointestinal problems, and potential cardiovascular complications [34]. Therefore, numerous initiatives are underway to explore and cultivate anti-obesity foods and food ingredients that can effectively reduce body fat accumulation, decrease the risk of obesity-related chronic diseases, and reduce potential side effects associated with clinical treatments [35].
Additionally, some studies have linked these effects to the bioactive phytochemical content of fermented soybean foods, such as α-amylase and α-glucosidase inhibitors, protease inhibitors, hemagglutinin, and crude fibers. These components have the potential to disrupt normal metabolism and aid in the management of obesity and various metabolic disorders [36]. This phase is linked with a heightened risk of various diseases and is often attributed to the lack of hormonal regulation. These include accumulation of abdominal fat, hypertriglyceridemia, increased LDL cholesterol, decreased HDL cholesterol, elevated blood pressure, and impaired glucose tolerance/diabetes [37]. There is a correlation between obesity and the transition from premenopause to postmenopause in women. The intake of isoflavones has been linked to an increase in HDL cholesterol and a decrease in TC, LDL, and TGs. In addition to isoflavones, soy proteins and peptides are active ingredients that lower LDL cholesterol and triacylglycerols [38]. Research indicates that the consumption of fermented soybean foods rich in isoflavones has positive effects on the distribution of body fat and lipid profiles in menopausal women. This is attributed to the structural similarity of these compounds to estrogen, their higher affinity for estrogen receptors, and their circulating concentrations in the human body [39]. Clinical and experimental studies suggest that obese and overweight individuals are more susceptible to T2DM and that obesity significantly elevates the risk of developing T2DM. Moreover, individuals with T2DM have an increased risk of cardiovascular disease, even with aggressive control of glucose, cholesterol, and blood pressure [40]. In a previous investigation, it was documented that soy isoflavones markedly attenuate body weight gain and fat accumulation in both obese and lean rats [41]. Moreover, a study report suggested that supplementation with chungkookjang enhances lean body mass while diminishing visceral fat areas. Another study also suggested that a diet rich in isoflavones improves lipid metabolism and exerts anti-obesity effects [42]. The isoflavones daidzein and genistein, which are present at high levels in fermented soybean foods, are recognized for their bioactivity, as they regulate the generation of lipids and thermogenesis in vivo [43]. Aglycones and metabolites showcase their anti-obesity effects through mechanisms such as lipogenesis (synthesis of fatty acids and TGs), hyperlipidemia (high levels of fat particles in the blood), hyperglycemia (elevated blood glucose levels), and improved insulin resistance [44]. γ-PGA is a hydrolyzed biopolymer composed of L-glutamic acid and/or D-glutamic acid monomers produced when Bacillus subtilis ferments soybeans [17,18,45]. Studies have reported that γ-PGA has various physiological activities, including potential anti-obesity and anti-inflammatory properties [46,47]. Building on previous studies, isoflavones and soy proteins have been shown to possess anti-obesity properties. In this study, PGA-K treatment demonstrated similar effects.
Additionally, the process of adipocyte differentiation is paralleled by an upregulation in the expression of inflammatory cytokines [10]. Thus, we tested the inflammatory cytokines expression in WATs, and the results showed that PGA-K treatment decreased the expression of TNF-α and IL-6 in serum. Adipose tissue releases adipocytokines, including TNF-α, IL-6, leptin, and adiponectin, and elevated serum leptin and reduced serum adiponectin levels are characteristic features of obesity [48]. In another study, prolonged consumption of an HFD resulted in heightened levels of insulin in the serum, triggered insulin resistance, and promoted increased fat accumulation within the liver [49]. In the present study, we found that PGA-K decreased serum insulin and leptin levels, indicating that its potential in improving insulin clearance and significantly reducing fat accumulation.
Our findings demonstrate that PGA-K administration has beneficial effects on obesity, as demonstrated by enhancements in parameters such as body weight, adipose tissue weight, and histological features. Administration of PGA-K significantly reduced body weight and adipose tissue weight. Furthermore, PGA-K restored insulin and leptin production, indicating its ability to prevent diabetes mellitus. In summary, our findings strongly indicated that PGA-K exerts a preventive effect against obesity.
5. Conclusions
This study provides scientific evidence supporting the anti-obesity function of PGA-K in HFD-fed mice, with a focus on adipose tissue dysfunction. The results of this study demonstrated that PGA-K from B. subtilis chungkookjang, administered daily at 100 and 200 mg/kg BW, significantly reduced HFD-induced body weight gain, especially adipose tissue hypertrophy. In addition, PGA-K ameliorated HFD-induced obesity-related adipose tissue disorders, such as inflammation, indicating its ability to attenuate obesity-related metabolic disorders. Thus, PGA-K is a potential candidate ingredient in functional foods for reducing and managing obesity and obesity-related diseases.
Author Contributions
Conceptualization, J.W.C., K.H.I., and H.J.C.; methodology, S.-H.L., J.W.C., M.K.S., and J.Y.P.; software, S.-H.L. and H.-R.K.; validation, S.-H.L., J.Y.P., and H.-R.K.; formal analysis, S.-H.L. and J.Y.P.; investigation, B.-J.O., S.-Y.K., and M.H.P. T.-G.L.; resources, J.-C.C. and Y.-C.P.; data curation, S.-H.L., H.-R.K., J.Y.P., and M.H.P.; writing—original draft preparation, S.-H.L., T.-G.L., M.-H.P., and S.-Y.K.; writing—review and editing, S.-H.L., M.-H.P., and S.-Y.K.; visualization, M.-H.P. and S.-Y.K.; supervision, S.-Y. K; project administration, H.Y.S., H.S.J., and S.-Y.K.; funding acquisition, H.Y.S., H.S.J., and S.-Y.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research was financially supported by the Ministry of Trade, Industry, and Energy (MOTIE), Korea, under the “Infrastructure program for smart specialization” supervised by the Korea Institute for Advancement of Technology (KIAT) (grant number: P0017238).
Institutional Review Board Statement
The animal study protocol was approved by the Institutional Animal Care and Use Committee of the Jeonju AgroBio-Materials Institute (approval number: JAMI IACUC 2023003).
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available in this article.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Ruze, R.; Liu, T.; Zou, X.; Song, J.; Chen, Y.; Xu, R.; Yin, X.; Xu, Q. Obesity and type 2 diabetes mellitus: connections in epidemiology, pathogenesis, and treatments. Front Endocrinol (Lausanne) 2023, 14, 1161521. [Google Scholar] [CrossRef]
- Park, S.Y.; Kim, J.E.; Kang, H.M.; Song, H.J.; Kang, N.J.; Hwang, D.Y.; Choi, Y.W. Adiposity Reduction by Cucumis melo var. gaettongchamoe Extract in High-Fat Diet-Induced Obese Mice. Nutrients 2023, 15. [Google Scholar] [CrossRef]
- Horwitz, A.; Birk, R. Adipose Tissue Hyperplasia and Hypertrophy in Common and Syndromic Obesity-The Case of BBS Obesity. Nutrients 2023, 15. [Google Scholar] [CrossRef]
- Hildebrandt, X.; Ibrahim, M.; Peltzer, N. Cell death and inflammation during obesity: “Know my methods, WAT(son)”. Cell Death Differ 2023, 30, 279–292. [Google Scholar] [CrossRef]
- Cavaliere, G.; Cimmino, F.; Trinchese, G.; Catapano, A.; Petrella, L.; D’Angelo, M.; Lucchin, L.; Mollica, M.P. From Obesity-Induced Low-Grade Inflammation to Lipotoxicity and Mitochondrial Dysfunction: Altered Multi-Crosstalk between Adipose Tissue and Metabolically Active Organs. Antioxidants (Basel) 2023, 12. [Google Scholar] [CrossRef]
- Chait, A.; den Hartigh, L.J. Adipose Tissue Distribution, Inflammation and Its Metabolic Consequences, Including Diabetes and Cardiovascular Disease. Front Cardiovasc Med 2020, 7, 22. [Google Scholar] [CrossRef] [PubMed]
- Shang, A.; Gan, R.Y.; Xu, X.Y.; Mao, Q.Q.; Zhang, P.Z.; Li, H.B. Effects and mechanisms of edible and medicinal plants on obesity: an updated review. Crit Rev Food Sci Nutr 2021, 61, 2061–2077. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Lee, M.S.; Jung, S.; Son, H.Y.; Park, S.; Kang, B.; Kim, S.Y.; Kim, I.H.; Kim, C.T.; Kim, Y. Ginger Extract Ameliorates Obesity and Inflammation via Regulating MicroRNA-21/132 Expression and AMPK Activation in White Adipose Tissue. Nutrients 2018, 10. [Google Scholar] [CrossRef] [PubMed]
- Pelczyńska, M.; Moszak, M.; Wesołek, A.; Bogdański, P. The Preventive Mechanisms of Bioactive Food Compounds against Obesity-Induced Inflammation. Antioxidants (Basel) 2023, 12. [Google Scholar] [CrossRef] [PubMed]
- Al-Mansoori, L.; Al-Jaber, H.; Prince, M.S.; Elrayess, M.A. Role of Inflammatory Cytokines, Growth Factors and Adipokines in Adipogenesis and Insulin Resistance. Inflammation 2022, 45, 31–44. [Google Scholar] [CrossRef] [PubMed]
- Yao, J.; Wu, D.; Qiu, Y. Adipose tissue macrophage in obesity-associated metabolic diseases. Front Immunol 2022, 13, 977485. [Google Scholar] [CrossRef] [PubMed]
- Lindhorst, A.; Raulien, N.; Wieghofer, P.; Eilers, J.; Rossi, F.M.V.; Bechmann, I.; Gericke, M. Adipocyte death triggers a pro-inflammatory response and induces metabolic activation of resident macrophages. Cell Death Dis 2021, 12, 579. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Ren, Y.; Chang, K.; Wu, W.; Griffiths, H.R.; Lu, S.; Gao, D. Adipose tissue macrophages as potential targets for obesity and metabolic diseases. Front Immunol 2023, 14, 1153915. [Google Scholar] [CrossRef] [PubMed]
- do Prado, F.G.; Pagnoncelli, M.G.B.; de Melo Pereira, G.V.; Karp, S.G.; Soccol, C.R. Fermented Soy Products and Their Potential Health Benefits: A Review. Microorganisms 2022, 10. [Google Scholar] [CrossRef] [PubMed]
- Kim, I.S.; Hwang, C.W.; Yang, W.S.; Kim, C.H. Current Perspectives on the Physiological Activities of Fermented Soybean-Derived Cheonggukjang. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef]
- Gopikrishna, T.; Suresh Kumar, H.K.; Perumal, K.; Elangovan, E. Impact of Bacillus in fermented soybean foods on human health. Ann Microbiol 2021, 71, 30. [Google Scholar] [CrossRef]
- Guo, Y.; Liu, Y.; Yang, Z.; Chen, G.; Liang, Z.; Zeng, W. Enhanced Production of Poly-γ-glutamic Acid by Bacillus subtilis Using Stage-controlled Fermentation and Viscosity Reduction Strategy. Appl Biochem Biotechnol 2023. [Google Scholar] [CrossRef]
- Afzaal, M.; Saeed, F.; Islam, F.; Ateeq, H.; Asghar, A.; Shah, Y.A.; Ofoedu, C.E.; Chacha, J.S. Nutritional Health Perspective of Natto: A Critical Review. Biochem Res Int 2022, 2022, 5863887. [Google Scholar] [CrossRef]
- Jang, H.H.; Noh, H.; Kim, H.W.; Cho, S.Y.; Kim, H.J.; Lee, S.H.; Lee, S.H.; Gunter, M.J.; Ferrari, P.; Scalbert, A.; et al. Metabolic tracking of isoflavones in soybean products and biosamples from healthy adults after fermented soybean consumption. Food Chem 2020, 330, 127317. [Google Scholar] [CrossRef] [PubMed]
- Kim, I.S.; Yang, W.S.; Kim, C.H. Beneficial Effects of Soybean-Derived Bioactive Peptides. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef] [PubMed]
- Tan, B.L.; Norhaizan, M.E.; Liew, W.P.; Sulaiman Rahman, H. Antioxidant and Oxidative Stress: A Mutual Interplay in Age-Related Diseases. Front Pharmacol 2018, 9, 1162. [Google Scholar] [CrossRef]
- Zhao, X.; Song, J.-L.; Wang, Q.; Qian, Y.; Li, G.-J.; Pang, L. Comparisons of Shuidouchi, Natto, and Cheonggukjang in their physicochemical properties, and antimutagenic and anticancer effects. Food Science and Biotechnology 2013, 22, 1077–1084. [Google Scholar] [CrossRef]
- Fuentes, E.; Fuentes, F.; Vilahur, G.; Badimon, L.; Palomo, I. Mechanisms of chronic state of inflammation as mediators that link obese adipose tissue and metabolic syndrome. Mediators Inflamm 2013, 2013, 136584. [Google Scholar] [CrossRef]
- Kirichenko, T.V.; Markina, Y.V.; Bogatyreva, A.I.; Tolstik, T.V.; Varaeva, Y.R.; Starodubova, A.V. The Role of Adipokines in Inflammatory Mechanisms of Obesity. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef] [PubMed]
- Fantuzzi, G. Adipose tissue, adipokines, and inflammation. Journal of Allergy and Clinical Immunology 2005, 115, 911–919. [Google Scholar] [CrossRef] [PubMed]
- Ellulu, M.; Patimah, I.; Khaza’ai, H.; Rahmat, A.; Abed, Y. Obesity and inflammation: the linking mechanism and the complications. Arch Med Sci. 2017, 13, 851–863, External Resources Crossref (DOI) 2016. [Google Scholar] [CrossRef]
- Wondmkun, Y.T. Obesity, insulin resistance, and type 2 diabetes: associations and therapeutic implications. Diabetes, Metabolic Syndrome and Obesity 2020, 3611–3616. [Google Scholar] [CrossRef]
- Kojta, I.; Chacińska, M.; Błachnio-Zabielska, A. Obesity, Bioactive Lipids, and Adipose Tissue Inflammation in Insulin Resistance. Nutrients 2020, 12. [Google Scholar] [CrossRef]
- Zatterale, F.; Longo, M.; Naderi, J.; Raciti, G.A.; Desiderio, A.; Miele, C.; Beguinot, F. Chronic Adipose Tissue Inflammation Linking Obesity to Insulin Resistance and Type 2 Diabetes. Front Physiol 2019, 10, 1607. [Google Scholar] [CrossRef] [PubMed]
- Choe, S.S.; Huh, J.Y.; Hwang, I.J.; Kim, J.I.; Kim, J.B. Adipose Tissue Remodeling: Its Role in Energy Metabolism and Metabolic Disorders. Front Endocrinol (Lausanne) 2016, 7, 30. [Google Scholar] [CrossRef]
- Stadler, J.T.; Marsche, G. Obesity-Related Changes in High-Density Lipoprotein Metabolism and Function. Int J Mol Sci 2020, 21. [Google Scholar] [CrossRef]
- Chakhtoura, M.; Haber, R.; Ghezzawi, M.; Rhayem, C.; Tcheroyan, R.; Mantzoros, C.S. Pharmacotherapy of obesity: an update on the available medications and drugs under investigation. EClinicalMedicine 2023, 58, 101882. [Google Scholar] [CrossRef]
- Kang, J.G.; Park, C.Y. Anti-Obesity Drugs: A Review about Their Effects and Safety. Diabetes Metab J 2012, 36, 13–25. [Google Scholar] [CrossRef] [PubMed]
- Tak, Y.J.; Lee, S.Y. Anti-Obesity Drugs: Long-Term Efficacy and Safety: An Updated Review. World J Mens Health 2021, 39, 208–221. [Google Scholar] [CrossRef]
- Zhou, M.; Huang, J.; Zhou, J.; Zhi, C.; Bai, Y.; Che, Q.; Cao, H.; Guo, J.; Su, Z. Anti-Obesity Effect and Mechanism of Chitooligosaccharides Were Revealed Based on Lipidomics in Diet-Induced Obese Mice. Molecules 2023, 28. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Kim, H.R.; Noh, E.M.; Park, J.Y.; Kwak, M.S.; Jung, Y.J.; Yang, H.J.; Ryu, M.S.; Seo, H.Y.; Jang, H.; et al. Anti-Inflammatory Effect and Signaling Mechanism of Glycine max Hydrolyzed with Enzymes from Bacillus velezensis KMU01 in a Dextran-Sulfate-Sodium-Induced Colitis Mouse Model. Nutrients 2023, 15. [Google Scholar] [CrossRef]
- Weggemans, R.M.; Trautwein, E.A. Relation between soy-associated isoflavones and LDL and HDL cholesterol concentrations in humans: a meta-analysis. Eur J Clin Nutr 2003, 57, 940–946. [Google Scholar] [CrossRef] [PubMed]
- Song, D.K.; Hong, Y.S.; Sung, Y.A.; Lee, H. The effect of menopause on cardiovascular risk factors according to body mass index in middle-aged Korean women. PLoS ONE 2023, 18, e0283393. [Google Scholar] [CrossRef] [PubMed]
- Khan, J.; Deb, P.K.; Priya, S.; Medina, K.D.; Devi, R.; Walode, S.G.; Rudrapal, M. Dietary Flavonoids: Cardioprotective Potential with Antioxidant Effects and Their Pharmacokinetic, Toxicological and Therapeutic Concerns. Molecules 2021, 26. [Google Scholar] [CrossRef]
- Einarson, T.R.; Acs, A.; Ludwig, C.; Panton, U.H. Prevalence of cardiovascular disease in type 2 diabetes: a systematic literature review of scientific evidence from across the world in 2007-2017. Cardiovasc Diabetol 2018, 17, 83. [Google Scholar] [CrossRef]
- Akhlaghi, M.; Zare, M.; Nouripour, F. Effect of Soy and Soy Isoflavones on Obesity-Related Anthropometric Measures: A Systematic Review and Meta-analysis of Randomized Controlled Clinical Trials. Adv Nutr 2017, 8, 705–717. [Google Scholar] [CrossRef] [PubMed]
- Byun, M.S.; Yu, O.K.; Cha, Y.S.; Park, T.S. Korean traditional Chungkookjang improves body composition, lipid profiles and atherogenic indices in overweight/obese subjects: a double-blind, randomized, crossover, placebo-controlled clinical trial. Eur J Clin Nutr 2016, 70, 1116–1122. [Google Scholar] [CrossRef] [PubMed]
- Langa, S.; Peirotén, Á.; Curiel, J.A.; de la Bastida, A.R.; Landete, J.M. Isoflavone Metabolism by Lactic Acid Bacteria and Its Application in the Development of Fermented Soy Food with Beneficial Effects on Human Health. Foods 2023, 12. [Google Scholar] [CrossRef] [PubMed]
- On, J.Y.; Kim, S.H.; Kim, J.M.; Park, S.; Kim, K.H.; Lee, C.H.; Kim, S.K. Effects of Fermented Artemisia annua L. and Salicornia herbacea L. on Inhibition of Obesity In Vitro and In Mice. Nutrients 2023, 15. [Google Scholar] [CrossRef]
- Ogunleye, A.; Bhat, A.; Irorere, V.U.; Hill, D.; Williams, C.; Radecka, I. Poly-γ-glutamic acid: production, properties and applications. Microbiology 2015, 161, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Balogun-Agbaje, O.A.; Odeniyi, O.A.; Odeniyi, M.A. Drug delivery applications of poly-γ-glutamic acid. Futur J Pharm Sci 2 0217, 125. [Google Scholar] [CrossRef]
- Liang, S.; Qian, W.; Rui, W.; Ke, S.; Sha, L.; Gui, L.; Peng, L.; Hong, X. Effect of dietary poly-γ-glutamic acid on growth, digestive enzyme activity, antioxidant capacity, and TOR pathway gene expression of gibel carp (Carassius auratus gibelio). Aquaculture Reports 2022, 27, 101412. [Google Scholar] [CrossRef]
- Zorena, K.; Jachimowicz-Duda, O.; Ślęzak, D.; Robakowska, M.; Mrugacz, M. Adipokines and Obesity. Potential Link to Metabolic Disorders and Chronic Complications. Int J Mol Sci 2020, 21. [Google Scholar] [CrossRef]
- Qi, Y.; Wang, X. The Role of Gut Microbiota in High-Fat-Diet-Induced Diabetes: Lessons from Animal Models and Humans. Nutrients 2023, 15. [Google Scholar] [CrossRef]
Figure 1.
Effects of PGA-K on body weight and eWAT weight in HFD-induced obesity mice. (A) PGA-K effects on body weight gain, (B) food intake, (C) final body weight, (D) eWAT weight, and (E) the ratio (%) of eWAT to body weight for 12 weeks of obesity induction by HFD. All values represent the mean ± SD. Duncan’s multiple comparison test was utilized for data analysis. ***p < 0.001, versus the N group; #p < 0.05, ##p < 0.01, and ###p < 0.001, versus the HFD group. PGA-K, poly-γ-glutamate potassium; HFD, high-fat diet; eWAT, epididymal white adipose tissue; N, normal; PC, positive control.
Figure 1.
Effects of PGA-K on body weight and eWAT weight in HFD-induced obesity mice. (A) PGA-K effects on body weight gain, (B) food intake, (C) final body weight, (D) eWAT weight, and (E) the ratio (%) of eWAT to body weight for 12 weeks of obesity induction by HFD. All values represent the mean ± SD. Duncan’s multiple comparison test was utilized for data analysis. ***p < 0.001, versus the N group; #p < 0.05, ##p < 0.01, and ###p < 0.001, versus the HFD group. PGA-K, poly-γ-glutamate potassium; HFD, high-fat diet; eWAT, epididymal white adipose tissue; N, normal; PC, positive control.
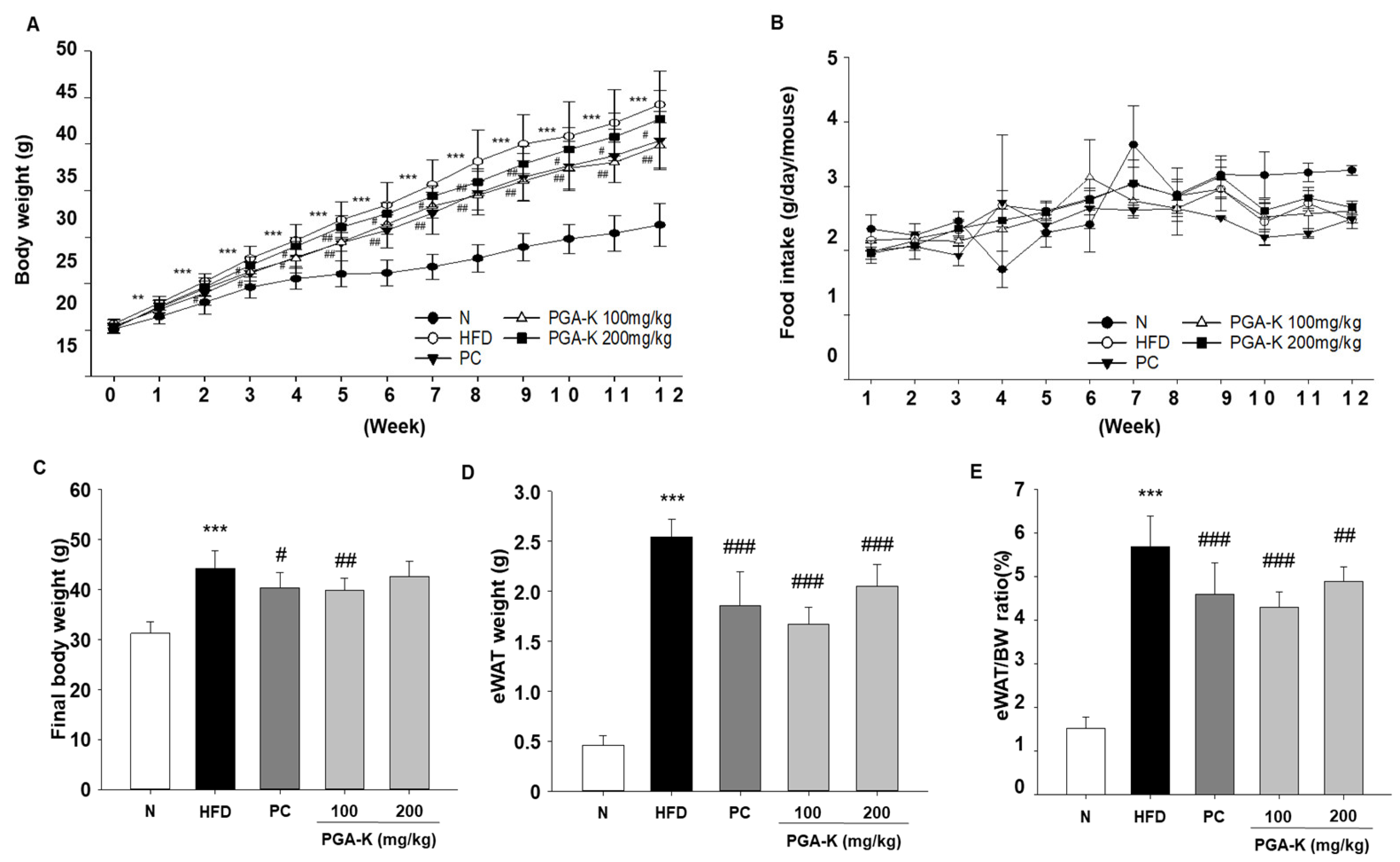
Figure 2.
Histopathological assessment of the effects of PGA-K on the liver and eWAT of HFD-induced obesity mice. (A) Representative image of eWAT tissue stained with H&E. Magnification, 100×; scale bar, 100 μm. (B) Representative image of liver tissue stained with Oil Red O. Magnification, 200×; scale bar, 60 μm. PGA-K, poly-γ-glutamate potassium; HFD, high-fat diet; eWAT, epididymal white adipose tissue; PC, positive control; N, normal; H&E, hematoxylin and eosin.
Figure 2.
Histopathological assessment of the effects of PGA-K on the liver and eWAT of HFD-induced obesity mice. (A) Representative image of eWAT tissue stained with H&E. Magnification, 100×; scale bar, 100 μm. (B) Representative image of liver tissue stained with Oil Red O. Magnification, 200×; scale bar, 60 μm. PGA-K, poly-γ-glutamate potassium; HFD, high-fat diet; eWAT, epididymal white adipose tissue; PC, positive control; N, normal; H&E, hematoxylin and eosin.
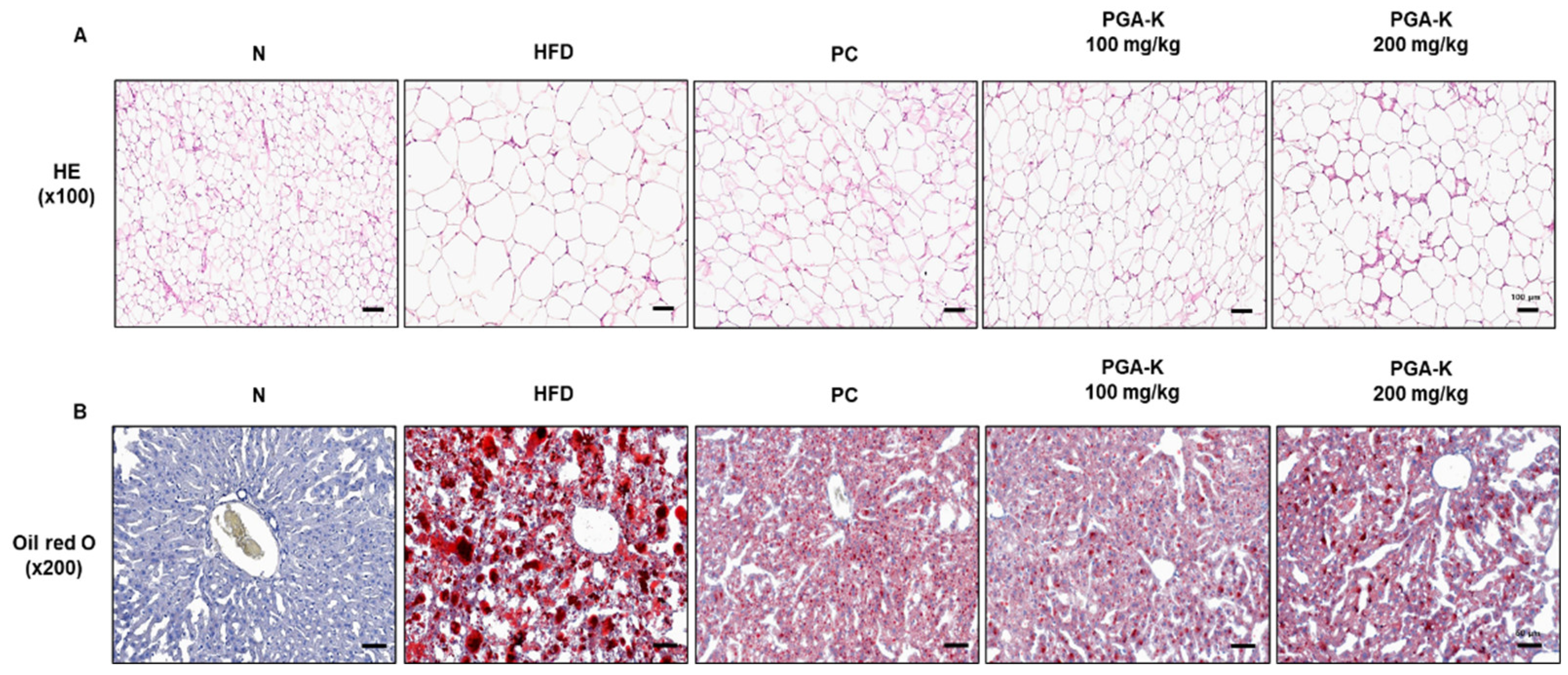
Figure 3.
Effects of PGA-K on serum lipid profiles of HFD-induced obesity mice. The levels of (A) TG, (B) TC, (C) HDL, and (D) LDL. All values represent the mean ± SD. Duncan’s multiple comparison test was utilized for data analysis. ***p < 0.001, versus the N group; ##p < 0.01 and ###p < 0.001, versus the HFD groups. PGA-K, poly-γ-glutamate potassium; HFD, high-fat diet; TG, triglyceride; TC, total cholesterol; HDL, high-density lipoprotein; LDL, low-density lipoprotein; N, normal; PC, positive control.
Figure 3.
Effects of PGA-K on serum lipid profiles of HFD-induced obesity mice. The levels of (A) TG, (B) TC, (C) HDL, and (D) LDL. All values represent the mean ± SD. Duncan’s multiple comparison test was utilized for data analysis. ***p < 0.001, versus the N group; ##p < 0.01 and ###p < 0.001, versus the HFD groups. PGA-K, poly-γ-glutamate potassium; HFD, high-fat diet; TG, triglyceride; TC, total cholesterol; HDL, high-density lipoprotein; LDL, low-density lipoprotein; N, normal; PC, positive control.
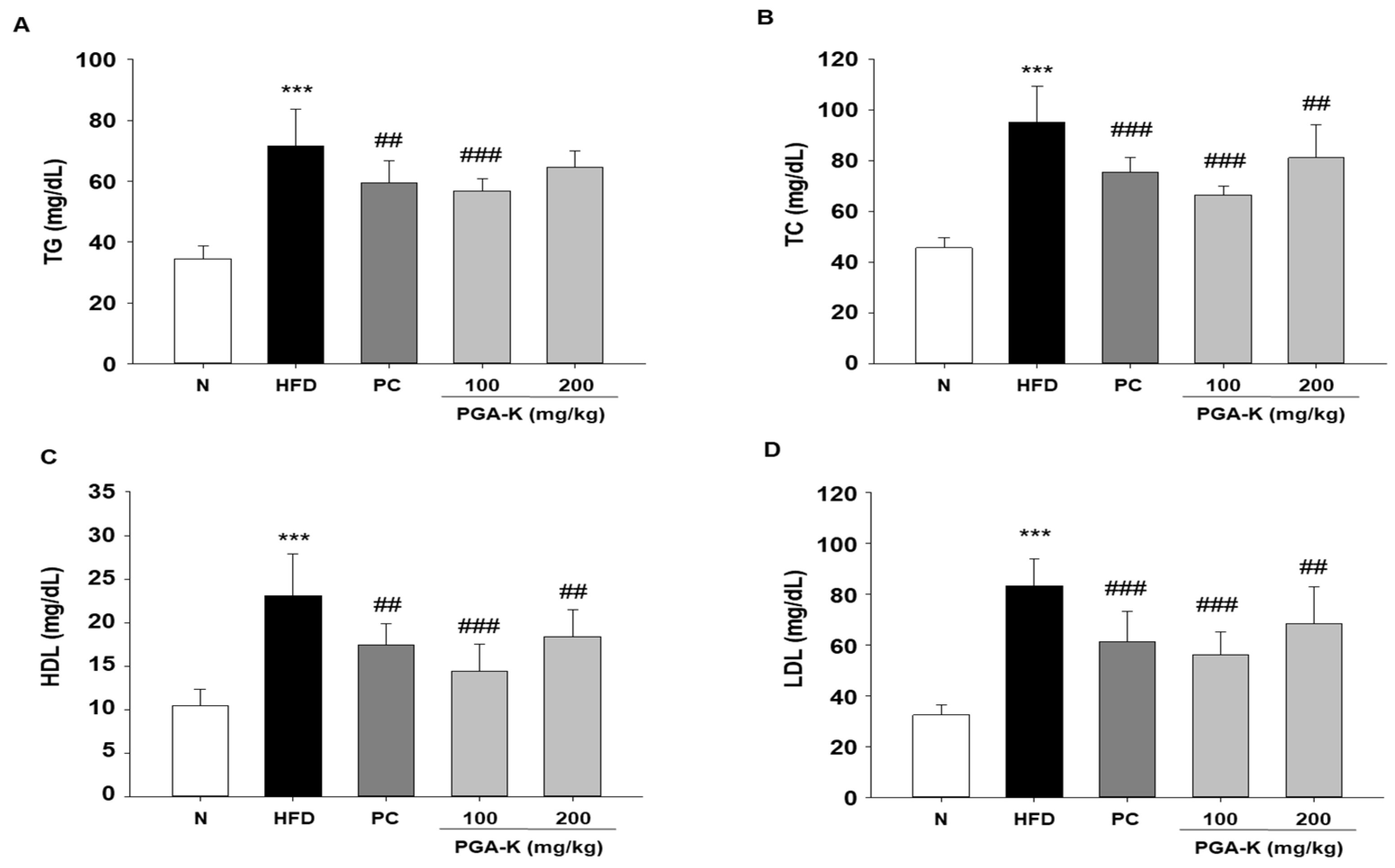
Figure 4.
Effects of PGA-K on liver toxicity in HFD-induced obesity mice. (A) Liver weight and (B) serum ALT and (C) AST levels in HFD-induced obesity mice. All values are presented as the mean ± SD. Duncan’s multiple comparison test was utilized for data analysis. ***p < 0.001, versus the N group; ##p < 0.01 and ###p < 0.001, versus the HFD group. PGA-K, potassium poly-γ-glutamate; HFD, high-fat diet; ALT, alanine aminotransferase; AST, aspartate aminotransferase; N, normal; PC, positive control.
Figure 4.
Effects of PGA-K on liver toxicity in HFD-induced obesity mice. (A) Liver weight and (B) serum ALT and (C) AST levels in HFD-induced obesity mice. All values are presented as the mean ± SD. Duncan’s multiple comparison test was utilized for data analysis. ***p < 0.001, versus the N group; ##p < 0.01 and ###p < 0.001, versus the HFD group. PGA-K, potassium poly-γ-glutamate; HFD, high-fat diet; ALT, alanine aminotransferase; AST, aspartate aminotransferase; N, normal; PC, positive control.
Figure 5.
Effects of PGA-K on serum pro-inflammatory cytokine levels in HFD-induced obesity mice. The levels of (A) TNF-α and (B) IL-6 in HFD-induced obese mice serum. All values represent the mean ± SD. Duncan’s multiple comparison test was utilized for data analysis. ***p < 0.001, versus the N group; #p < 0.05, ##p < 0.01, and ###p < 0.001, versus the HFD group. PGA-K, poly-γ-glutamate potassium; HFD, high-fat diet; TNF-α, tumor necrosis factor α; IL-6, interleukin; N, normal; PC, positive control.
Figure 5.
Effects of PGA-K on serum pro-inflammatory cytokine levels in HFD-induced obesity mice. The levels of (A) TNF-α and (B) IL-6 in HFD-induced obese mice serum. All values represent the mean ± SD. Duncan’s multiple comparison test was utilized for data analysis. ***p < 0.001, versus the N group; #p < 0.05, ##p < 0.01, and ###p < 0.001, versus the HFD group. PGA-K, poly-γ-glutamate potassium; HFD, high-fat diet; TNF-α, tumor necrosis factor α; IL-6, interleukin; N, normal; PC, positive control.
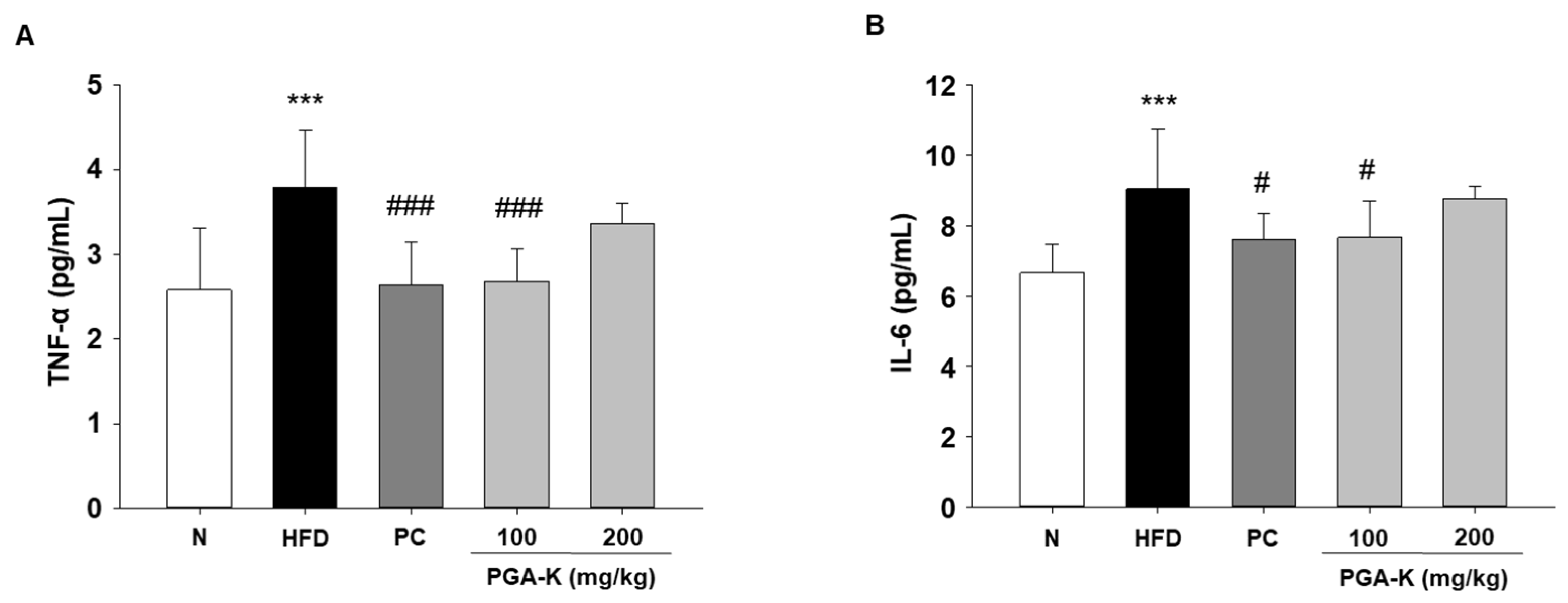
Figure 6.
Effects of PGA-K on serum insulin resistance markers of HFD-induced obesity mice. (A) Leptin and (B) insulin. All values represent the mean ± SD. Duncan’s multiple comparison test was utilized for data analysis. ***p < 0.001, versus the N group; #p < 0.05, ##p < 0.01, and ###p < 0.001, versus the HFD group. PGA-K, poly-γ-glutamate potassium; HFD, high-fat diet; TNF-α, tumor necrosis factor α; IL-6, interleukin; N, normal; PC, positive control.
Figure 6.
Effects of PGA-K on serum insulin resistance markers of HFD-induced obesity mice. (A) Leptin and (B) insulin. All values represent the mean ± SD. Duncan’s multiple comparison test was utilized for data analysis. ***p < 0.001, versus the N group; #p < 0.05, ##p < 0.01, and ###p < 0.001, versus the HFD group. PGA-K, poly-γ-glutamate potassium; HFD, high-fat diet; TNF-α, tumor necrosis factor α; IL-6, interleukin; N, normal; PC, positive control.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



