Submitted:
07 February 2024
Posted:
08 February 2024
You are already at the latest version
Abstract
Clusterin (CLU) protein is involved in various pathophysiological processes including carcinogenesis and tumor progression. In recent years, the role of the secretory isoform has been demonstrated in tumor cells, where it inhibits apoptosis and favors the acquisition of resistance to conventional treatments used to treat cancer. To determine the possible therapeutic potential of inhibiting this protein, numerous studies have been carried out in this field. In this paper, we show the existing knowledge on the inhibition of this protein in different types of cancer and discuss the importance it could have in the development of new therapies directed against this pathology.
Keywords:
clusterin
; human cancer
; targeted treatment
; resistance
; therapeutic potential.
1. Introduction
Clusterin (CLU), also known by various names such as APOJ, CLI, Complement Lysis Inhibitor, SP-40,40, Sulfated Glycoprotein 2, Testosterone-Repressed Prostate Message 2, Apolipoprotein J, Ku70-Binding Protein 1, KUB1, SGP-2, SP-40, or TRPM2, is a multifunctional protein encoded by the CLU gene [1], widely expressed in various human tissues [2,3]. Over the years, CLU has been identified as a key molecule in a variety of physiological processes [3], including cell differentiation, morphogenesis [4], sperm maturation, lipid transport, complement inhibition, tissue remodeling, membrane recycling, cell-cell and cell-substrate interactions, stabilization of stressed proteins, cell proliferation, survival, and apoptosis [5,6].
Alterations in CLU expression has been associated with serious physiological diseases, such as spongiform encephalopathies, hippocampal and heart ischemic injuries, atherosclerosis [7], schizophrenia [8], cardiovascular diseases [9], cancer, vascular damage, diabetes, osteoarthritis [10], and degenerative conditions such as age-related macular degeneration, retinitis pigmentosa [11], Parkinson's disease [12], and Alzheimer's [13,14]. These conditions are often more prominent in advanced aging [13,14], where clusterin overexpression has been observed in phenomena such as enhanced cell migration [15], chemotherapy resistance [16,17], and more aggressive biological behaviors [18] in malignant cells. CLU has been proposed as a cancer biomarker [19] and cellular senescence [20], generating interest in research to better understand its implications and explore its utility in the diagnosis, prevention, and treatment of these pathologies.
2. Isoforms and Regulation of CLU Expression Gene
The human CLU gene, located on chromosome 8p21-p12, is a highly conserved gene with at least 63 orthologs in different species [21]. It has a structure consisting of 11 exons and spans approximately 16 kilobases. This gene produces at least 17 splice variants, both coding and non-coding [21,22]. However, it generates three main transcription products identified as Variant 1 (NM_001831), Variant 2 (NR_038335), and Variant 3 (NR_045494). These variants share exons 2-11 but differ in exon 1 (1a, 1b, 1c) and in the untranslated terminal regions [23,24,25], indicating the existence of distinct transcription start sites for each variant, although the biological relevance of the different transcripts generated is still unclear.
The sequence initially encodes a translocation signal to direct the preprotein to the endoplasmic reticulum (ER), where the signal sequence is removed, and N-glycosylation and disulfide bond formation occur [26]. Subsequently, the protein is translocated to the Golgi apparatus, undergoes more complex glycosylation, and reaches a weight of 70 to 80 kDa. The mature form of the protein, known as the secreted isoform (sCLU), is generated by cleavage and disulfide bond formation between residues 227 and 228 (Figure 1). The 449-amino acid sCLU is secreted as a heterodimeric glycoprotein complex and functions as an extracellular chaperone, inhibiting the aggregation of partially unfolded proteins, enhancing phagocytosis and cellular degradation, and exhibiting anti-apoptotic activity in cancer cells [27,28,29]. This isoform can stabilize the synthesis of the p53 protein, involved in the activation of genes that halt the cell cycle, thus inhibiting cell proliferation [30,31].
In situations of alternative splicing between exons 1 and 3, exon 2 is deleted, generating a non-functional prenuclear isoform of 49 kDa (pnCLU). Under stress, this isoform transforms into a functional nuclear isoform of 55 kDa (nCLU), which, upon translocating to the nucleus and binding to the Ku70 protein, promotes cellular apoptosis [32,33,34,35] (Figure 1).
Additionally, there is an almost mature presecreted isoform of 53 kDa (psCLU) that, under stress, binds to the chaperone GRP78 (Bip) in the ER and translocates to the mitochondria (Figure 1). In the mitochondria, it binds to the activated form of the Bax protein, modulating its ability to form homodimers and inhibiting the formation of Bax-Bak complexes [36]. This anti-apoptotic activity is also evident in the cytoplasm, where psCLU can stabilize the Ku70-Bax complex, preventing Bax from reaching the mitochondria and promoting the proteasomal degradation of cytotoxic substances [37,38].
The expression of CLU is meticulously regulated in both healthy and pathological contexts, where various pathways exert influence on its basal expression in healthy tissues as well as its upregulation in conditions such as apoptosis, pathologies, and aging. In this scenario, two forms of CLU with antagonistic functions are distinguished: sCLU, playing a cytoprotective role, and nCLU, capable of promoting cell death [39,40]. Each CLU variant is subject to different signaling pathways, depending not only on its molecular configuration but also on the cell type in which it manifests. The sophisticated regulation of both CLU isoforms encompasses a broad spectrum of factors, including growth factors such as TGF-β, NGF, EGF, cytokines like TNF-α and IL-1, IL-2, and IL-6, transcription factors such as TCF1, Jak/STAT1, and IGF1, as well as the Wnt signaling pathway [41,42,43,44,45] and epigenetic modifications. Furthermore, expression can be induced by various agents such as oxidative stress, chemotherapeutic agents, ionizing and ultraviolet radiation, estrogen, and androgen deprivation in hormone-sensitive tumors [46,47].Gene expression control in vertebrates is an intricate process orchestrated by numerous regulatory proteins that collaborate to influence the expression of a specific gene or transcript within a cell. Typically, vertebrate gene expression is regulated by the basal promoter, a DNA region spanning from -200bp to the transcription start site (TSS) (+1), facilitating the binding of regulatory proteins, including transcription factors [48]. The variation in CLU expression across different tissues suggests its finely tuned and tissue-specific regulation at transcriptional, translational, and post-translational levels [49]. The human CLU promoter, initially identified as a unique element, contains conserved DNA motifs, including a palindrome (-73 to -87) and a TATAA box (-26), crucial for decoding the genetic sequence. Furthermore, a GCATT box (-93) plays a key role in regulating transcription frequency [50]. Additional studies have uncovered a second promoter region (P2) within the first CLU intron, potentially controlling CLU2 expression [23].
Epigenetics, the study of heritable changes in gene expression without altering the DNA sequence, emerges as a pivotal player in regulating clusterin (CLU) expression during aging and cancer progression [51]. The intricate interplay of DNA methylation and histone acetylation [52,53], influenced by aging and cancer, suggests epigenetic regulation's significant role in controlling CLU [23,51,52,54].
The Histone Code Hypothesis outlines the diverse histone modifications governing chromatin structure and transcriptional status [55]. Histone modifications impact gene silencing or activation by modulating DNA accessibility. Notably, histone 3 lysine 9 trimethylation (H3K9me3) and histone 3 lysine 27 trimethylation (H3K27me) contribute to nCLU down-regulation [51] and cell survival. Conversely, histone 3 lysine 4 trimethylation (H3K4me3) or histone 3 lysine 9 acetylation and serine 10 phosphorylation (H3K9AcS10P) lead to nCLU activation [51] and cell death. Epigenetic drug treatments alter histone modifications, impacting CLU1 and CLU2 transcription [23].
Consistent findings across diverse cell types, including cancer cells [23,52], retinal pigment epithelial cells [52], hepatocellular carcinoma cell lines [56], and endothelial tumor cells [53], emphasize the role of histone hyperacetylation induced by histone deacetylase inhibition in promoting CLU expression.
In differentiated mammalian cells, gene repression often involves DNA methylation, co-coordinated with histone modifications. While colon cancer studies indicate predominant regulation of CLU by histone modifications [51], the presence of a G+C-rich region and a CpG mini-island within the CLU promoter suggests epigenetic control via methylation [50]. Promoter methylation is associated with decreased protein expression, observed in breast [54], ovarian epithelial cancer [18], and hormone-refractory prostate carcinoma samples [57]. Induced demethylation of CLU promoters significantly increases CLU1 and CLU2 transcripts, enhancing CLU expression across various colon cell lines [51] and tissues [58].
Furthermore, micro-RNAs (mi-RNAs) contribute to post-transcriptional regulation of CLU expression [59]. In head and neck squamous cell carcinoma, oncogenic miRNA-21 specifically targets the CLU1 isoform, down-regulating its growth-suppressive variant [60]. Elevated levels of miRNA-378 inhibit lung adenocarcinoma cell growth and sCLU expression [61].
While this field's studies are recent, the emerging evidence underscores the intricate epigenetic control mechanisms orchestrating CLU expression in diverse cellular contexts. These insights provide a foundation for understanding the molecular intricacies of CLU regulation, offering potential avenues for therapeutic interventions in aging and cancer-related conditions.
3. Clusterin and Its Involvement in Cancer
Despite initially being believed to be linked to biological processes such as scar formation, membrane remodeling, or sperm maturation [50], early studies revealed a correlation between the overexpression of mRNA-CLU and cell death after different toxic signals [62,63,64]. However, later research presented contradictory results, establishing a relationship between CLU overexpression and cell protection [65,66].
The identification of two sets of CLU isoforms, with opposing roles and cellular locations in terms of cell survival, has contributed to clarifying these contradictions [67,68]. While the expression of the nuclear isoform (nCLU) is associated with healthy tissues, the overexpression of the secretory isoform (sCLU) is related to most malignancies [19]. In tumor cells, a pro-survival environment is observed with a higher sCLU/nCLU ratio [69,70].
Post-translational modifications appear to regulate the distinctive roles of CLU, determining the expression of specific isoforms based on the presence or absence of a leader sequence in the CLU peptide [71]. Inhibition of exon 2 of mRNA-CLU, leading to the silencing of the sCLU isoform, has been crucial for the design of cancer-targeted therapies that are currently under study [72].
Early studies suggest that CLU overexpression persists after tissue degeneration, suggesting that this gene is probably not induced as part of apoptosis that may lead to a degenerative disorder but as a secondary consequence of the disease phenotype [50]. nCLU-mediated apoptosis is associated with the accumulation of mature nCLU protein, while the rapid degradation of CLU and the regulation of its nuclear export/import can explain why highly malignant tumors avoid accumulating nCLU levels [72].The lethality of nCLU is linked to members of the BCL-2 protein family, with nCLU-mediated apoptosis depending on Bax [69]. Additionally, nCLU can sequester the antiapoptotic BCL-XL, promoting apoptosis by forming pores in mitochondrial membranes [73].
The proapoptotic role of nCLU is also related to Ku70, which binds to CLU in response to DNA damage. The formation of a trimeric protein complex (nCLU/Ku70/Ku80) with reduced DNA end-binding activity and interaction with other proteins like Ku70 in the C-terminal coiled-coil domain of nCLU is essential for inducing apoptosis [74].
Although the constant expression of nCLU is related to cell maintenance and apoptosis induction, cells can also express cytoplasmic clusterin (cCLU) or sCLU isoforms in response to different cellular triggers. This results in increased cell survival associated with tumorigenesis, with CLU promoting apoptosis under low or moderate stress conditions but favoring the overexpression of the antiapoptotic isoform [19] under extreme cellular conditions, resulting in increased cell proliferation, viability, and invasiveness [75].
In addition, CLU has garnered attention from the scientific community due to its essential role in various biological processes, such as cell proliferation, apoptosis, epithelial-mesenchymal transition (EMT), metastasis, and resistance to chemotherapy.
3.1. Tumorigenesis
Under normal conditions, CLU expression is low and significantly increases in response to stress, activating the transcription of clusterin mRNA [76,77]. The cytoprotective isoform sCLU, when faced with stress, protects cells through its extracellular chaperone activity, thereby preventing apoptosis and conferring resistance to cytotoxic agents. sCLU plays a crucial role by binding to the Ku70-bax complex, acting as a bax retention factor in the cytosol and inhibiting its proapoptotic function. Inhibition of CLU weakens this complex, increases bax translocation to the mitochondria, triggers cytochrome c release, and activates caspase 9, initiating apoptosis [78]. On the other hand, the proapoptotic isoform nCLU induces apoptosis in cancers such as breast and prostate by interacting with proteins like Ku70 and Bcl-XL [79,80].
CLU is also involved in various signaling pathways, such as the transcription factor B-MYB, Akt phosphorylation, and the PI3K/Akt pathway, regulating its apoptotic and antiapoptotic functions [81,82]. The production ratio of both isoforms is crucial for regulating these functions, and CLU overexpression has been associated with the promotion of tumorigenesis and resistance to chemotherapy in cancers such as breast and prostate [83].
3.2. Cell Proliferation
Cell proliferation, a fundamental biological process, involves the increase in the number of cells through repeated cell divisions to maintain a dynamic balance between cell death and promote metabolism in biology. On the other hand, cell growth refers to the physical expansion of cell volume or size as the cell matures [84]. These processes are orderly and strictly controlled by various growth factors and cytokines in normal cells [85]. However, in cancer cells, cell proliferation is uncontrolled and exhibits a sustained proliferation property due to the hyperactivation of proliferative signaling pathways and evasion of growth suppressors [85].
The acquisition of sustained proliferative signaling pathways becomes a distinctive feature of cancer cells, and recent evidence has highlighted the regulatory role of the CLU protein in cell proliferation and growth in various types of cancer [86]. Relevant studies have revealed that c-Myc, a transcription factor encoded by the oncogene MYC involved in tumorigenesis, inhibits the expression of nCLU by upregulating the microRNA cluster miRNA-17 ~ 92 and attenuating the TGF-β axis, thus promoting angiogenesis and tumor growth in colon cancer [87].
Both nCLU and sCLU play a prominent role in regulating the canonical ERK, NF-κB, and AKT pathways, mediating cell proliferation and growth in various types of cancer. Inhibition of sCLU by apocynin, an NADPH oxidase inhibitor, inactivates the MEK-ERK1/2 pathway and suppresses cell proliferation in prostate cancer [88]. Meanwhile, induction of sCLU by DDP promotes the phosphorylation of ERK1/2, thereby driving cell growth and increasing resistance to DDP in OS [89].
The NF-κB axis plays a crucial role in cell proliferation, where activated NF-κB induces the expression of Bcl-2 to promote cell proliferation [90]. Melittin inhibits sCLU, inactivating both the cholesterol/NF-κB/Bcl-2 axis and the cholesterol/p-ERK axis to suppress tumor growth in pancreatic cancer [90]. Interestingly, sCLU exhibits an anti-proliferative property by inactivating the TAK1/NF-κB axis, preventing the transforming growth factor beta receptor 1 (TGFBR1) from recruiting the TNF receptor-activating factor 6 (TRAF6)/TAK1-binding protein 2 (TAB2)/TGF-β-activated kinase 1 (TAK1) complex to inhibit tumor proliferation and growth in human non-small cell lung cancer (NSCLC) [91].
In addition to the ERK and NF-κB pathways, the PI3K/AKT axis is also involved in regulating cell proliferation and growth. Notably, sCLU activates AKT by downregulating the expression of the protein phosphatase 2A catalytic subunit C (PP2AC) to promote the proliferation of PC-3 cells in prostate cancer [92]. This aligns with the finding that silencing CLU gene transcription decreases the phosphorylation level of AKT and GSK-3β, subsequently inhibiting the proliferation of HCCLM3 cells in hepatocellular carcinoma (HCC) [93].
Various external factors also influence CLU expression and, consequently, cell proliferation. Insulin-like growth factor 1 (IGF-1), a crucial component of insulin receptor signaling involved in sustained cell proliferation [94], has been shown in several studies to promote sCLU expression. For example, IGF-1 upregulates sCLU expression and subsequently activates the PI3K/AKT axis to promote the proliferation of A549 cells in NSCLC [95]. Meanwhile, IGF-1 upregulates the transcriptional activity of the CLU gene by activating the STAT-3/Twist-1 axis in prostate cancer [96]. Interestingly, pituitary tumor transforming gene (PTTG) and forkhead box protein L2 (FOXL2), respectively, stimulate sCLU expression and CLU gene transcription by activating the ATM/IGF-1/p38MAPK/CLU axis and directly binding to the CLU promoter. Meanwhile, sCLU exhibits an anti-proliferative property that, in turn, attenuates PTTG expression to restrain cell proliferation in pituitary carcinoma [97].
Metformin, a classic drug for treating type II diabetes, shows anti-tumor properties, as sCLU suppressed by metformin inhibits tumor growth by inactivating the SREBP-1c/fatty acid synthase (FASN) axis in bladder cancer [98]. Additionally, sCLU promotes cell growth and proliferation by upregulating the expression of the calcium-binding protein S100A4 in renal cancer [99]. Along with metformin and melittin, epigallocatechin-3-gallate (EGCG) and green tea extract (GTE) also induce sCLU expression by suppressing the transcriptional activity of β-catenin, promoting the proliferation of COLO 205 cells in colon cancer [100].
3.3. Epithelial-Mesenchymal Transition and Metastasis
Epithelial-mesenchymal transition (EMT) is a biological process involving significant changes in cell structure and function, such as loss of polarity, cytoskeletal remodeling, and acquisition of invasive properties. EMT facilitates invasion, metastasis, and tumor progression. Clusterin (CLU) has emerged as a key regulator in these events, influencing matrix metalloproteinase (MMP) activity and the ERK1/2 and PI3K/Akt signaling pathways [101]. NF-κB activation by CLU also plays a crucial role in increasing MMP-2 and MMP-9 expression, thus promoting metastasis [102].
In different cancer types, CLU overexpression has been associated with metastasis. In nasopharyngeal carcinoma, CLU is positively regulated by N, N′-dinitrosopiperazine (DNP), inducing MMP-9 and VEGF expression, contributing to metastasis [103]. In breast cancer, CLU collaborates with eHsp90α to activate key signaling pathways, promoting EMT, migration, and tumor metastasis [104]. In colon cancer, CLU interaction with platelets activates the p38MAPK pathway and positively regulates MMP-9, facilitating invasion [95]. Additionally, studies on prostate cancer have shown that miRNA-217-5p regulates invasion and migration by targeting CLU [105].
In patients with pancreatic ductal adenocarcinoma (PDAC), positive regulation of CLU by hepatocyte nuclear factor 1b (HNF1B) correlates with worse survival. Decreased CLU levels, along with reduced HNF1B, contribute to increased cell proliferation, EMT, and chemotherapy resistance, accelerating PDAC progression. These findings suggest that the HNF1B/CLU pathway is crucial for restraining pancreatic cancer progression [106].
3.4. Chemoresistance and Chemosensitivity with Clusterin
Chemoresistance, involving the ability of cancer cells to resist the effects of chemotherapy, is a significant challenge in cancer treatment. This phenomenon can manifest through various mechanisms, including genetic changes, alterations in drug metabolism, and suppression of apoptosis [107]. Chaperone proteins, like clusterin (CLU), play a crucial role in cancer therapy resistance, facilitating the growth of malignant tumors and protecting drug-resistant cells [108].
The stress response induced by treatments such as radiotherapy and chemotherapy lead to the overexpression of sCLU, a cytoprotective chaperone, which, by interacting with activated Bax, blocks the release of cytochrome c and prevents apoptosis [109]. Negative regulation of CLU in certain types of cancer, such as testicular seminoma, has been shown to increase sensitivity to radiotherapy and chemotherapy [110].
In hepatocellular carcinoma (HCC), sCLU is strongly overexpressed, contributing to oxaliplatin resistance by negatively regulating Gadd45a expression and activating the PI3K/Akt pathway. Inhibition of CLU in HepG2/ADM HCC cells restores chemosensitivity to various drugs [111].
4. Clusterin as a Biomarker and Therapeutic Target in Cancer
In the field of medicine, biomarkers have become essential tools to provide valuable information about patient health and the progression of various diseases. From measuring simple protein levels to identifying genetic mutations, these markers play a crucial role in the diagnosis, prognosis, monitoring, and treatment of diseases. In this context, clusterin has been studied as a potential biomarker in various types of cancer due to its tendency to be overexpressed in stressful situations (Table 1).
However, further research is needed in this field to determine the exact value of CLU levels and their association with each type of cancer. Currently, it can only be used as a tool to assess risks [150].
As a therapeutic target, the inhibition of CLU has demonstrated excellent therapeutic effects in various cancers both in vitro and in vivo, prolonging the survival of patients. Custirsen (OGX-011), a second-generation antisense oligonucleotide, has proven effective by interacting with the ATG sequence in exon 2 of the secretory isoform of CLU (sCLU), inhibiting its translation and suppressing cancer progression such as prostate cancer [151], renal cell carcinoma [152], bladder [153], liver [154], lung [155], prostate [156], breast [157], lung adenocarcinoma [158], melanoma [159], osteosarcoma [160] and ovary [161].
When combined with other cancer therapies in clinical trials, OGX 011 enhances antitumor activity. In the case of renal cell carcinoma, CLU inhibition by custirsen improves sorafenib cytotoxicity [162]. In ovarian cancer, it has shown to improve the survival of patients treated with paclitaxel by enhancing its response [163], like what happens in castration-resistant prostate cancer with mitoxantrone and docetaxel by reducing sCLU expression [164,165]. Clinical studies have also explored the efficacy of custirsen in metastatic castration-resistant prostate cancer, showing promising results in early-phase trials, although the standard treatment remains prednisone and cabazitaxel [166].
In metastatic breast cancer, similar effects have been shown with combinations of OGX 011 and docetaxel [167], while in lung cancer, combinations with gemcitabine or cisplatin have extended survival [168].
In addition to OGX-011, RNA interference therapies, such as microRNA (miRNA), short hairpin RNA (shRNA), and small interfering RNA (siRNA), have proven effective in inhibiting CLU expression in various cancers. Both miRNA-217-5p and miRNA-195 have negatively regulated CLU expression, inhibiting cell migration and invasion and promoting apoptosis in prostate cancer in vitro [169]. Other therapeutic agents, such as melittin, green tea extract (GTE), apocynin, vitamin D, metformin, and verteporfin, have also shown antitumor activity by blocking CLU protein expression and, therefore, inhibiting cancer progression, as evidenced earlier [170,171,172,173,174,175].
In the realm of emerging therapies, the drug AB-16B5, a monoclonal antibody targeting sCLU, is being evaluated in a phase II clinical trial in combination with docetaxel in patients with metastatic non-small cell lung cancer [176].
5. Conclusions
CLU plays a crucial role in cancer progression by regulating processes such as programmed cell death, epithelial-mesenchymal transition (EMT), metastasis, and cell proliferation and growth through various signaling pathways. The complexity of its biological role lies in the presence of two alternatively spliced isoforms and the variable localization of their protein products inside and outside the cell. Despite the complexity associated with the functions of these isoforms, it has been observed that the coexistence of nCLU and sCLU in cells, along with the precise regulation of their balance, confers pro or anti-apoptotic properties.
Several potent clusterin inhibitors, such as OGX-011 and RNAi, have been developed for use in cancer therapies and have demonstrated positive therapeutic effects in patients. Given the various regulatory pathways of clusterin in cancer progression and the proven survival benefits of clusterin inhibition in different types of cancer, clusterin emerges as a promising therapeutic target for developing more effective agents and targeted therapies. Considering its carcinogenic properties, the sCLU isoform could be a blood biomarker for cancer diagnosis. However, despite numerous studies on the function of clusterin isoforms, it is not yet fully understood which of them contributes to specific biological effects. Therefore, it is crucial to explore in more detail the specific functional role of the clusterin isoform, especially in the case of sCLU, to address clusterin as a therapeutic target more precisely.
Author Contributions
D.M.-G., conception and design and wrote the paper with input from all authors; M.G.-A., review and editing and contribution to the final version of the manuscript; M.R., conception, critical revision, and contribution to the final version of the manuscript. All authors have read and agreed to the published version of the manuscripts.
Funding
The researcher Marilina García-Aranda is the benefactor of a postdoctoral contract financed by the European Social Fund—Operational Program of Andalusia 2014–2020 for the “Incorporation of Research Personnel with a PhD degree in the field of Health Sciences and Technologies in R&D and Innovation Centers of the Public Health System of Andalusia” (RH-0055-2020). This work was partially supported by grants from the University of Malaga—Consejería de Transformación Económica, Industria, Conocimiento y Universidades—Junta de Andalucia (UMA20-FEDERJA-161) and from Instituto de Salud Carlos III (PI18/01181, PI21/00252) and were cofunded by the European Regional Development fund. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hogg, S.D., Embery, G. The isolation and partial characterization of a sulphated glycoprotein from human whole saliva which aggregates strains of Streptococcus sanguis but not Streptococcus mutans. Arch. Oral Biol. 1979, 24, 791–797. [CrossRef]
- Jones, S.E., & C. Jomary. Clusterin. Int J Biochem Cell Biol. 2002, 34(5), 427-31.
- Trougakos, I.P., & E.S. Gonos. Regulation of clusterin/apolipoprotein J, a functional homologue to the small heat shock proteins, by oxidative stress in ageing and age-related diseases. Free Radic Res. 2006, 40(12), 1324-34. [CrossRef]
- Itahana, Y., Piens, M., Sumida, T., Fong, S., Muschler, J., & Desprez, P. Y. Regulation of clusterin expression in mammary epithelial cells. Exp Cell Res. 2007, 313(5), 943-51. [CrossRef]
- Trougakos, I.P. & E.S. Gonos. Clusterin/apolipoprotein J in human aging and cancer. Int J Biochem Cell Biol. 2002, 34(11), 1430-48. [CrossRef]
- Bettuzzi, S. Chapter 1: Introduction. Adv Cancer Res. 2009, 104, 1-8.
- Prochnow, H., Gollan, R., Rohne, P., Hassemer, M., Koch-Brandt, C., & Baiersdörfer, M. Non-secreted clusterin isoforms are translated in rare amounts from distinct human mRNA variants and do not affect Bax-mediated apoptosis or the NF-kappaB signaling pathway. PLoS One. 2013, 8(9), e75303.
- Athanas, K.M., S.L. Mauney, & T.U. Woo, Increased extracellular clusterin in the prefrontal cortex in schizophrenia. Schizophr Res. 2015, 169(1-3), 381-5. [CrossRef]
- Yu, B., Yang, Y., Liu, H., Gong, M., Millard, R. W., Wang, Y. G., Ashraf, M., & Xu, M. Clusterin/Akt Up-Regulation Is Critical for GATA-4 Mediated Cytoprotection of Mesenchymal Stem Cells against Ischemia Injury. PLoS One. 2016, 11(3), e0151542. [CrossRef]
- Fandridis, E., Apergis, G., Korres, D. S., Nikolopoulos, K., Zoubos, A. B., Papassideri, I., & Trougakos, I. P. Increased expression levels of apolipoprotein J/clusterin during primary osteoarthritis. In Vivo. 2011, 25(5), 745-9.
- Suuronen, T., Nuutinen, T., Ryhänen, T., Kaarniranta, K., & Salminen, A. Epigenetic regulation of clusterin/apolipoprotein J expression in retinal pigment epithelial cells. Biochem Biophys Res Commun. 2007, 357(2), 397-401. [CrossRef]
- Prikrylova Vranova, H., Mareš, J., Nevrlý, M., Stejskal, D., Zapletalová, J., Hluštík, P., & Kaňovský, P. CSF markers of neurodegeneration in Parkinson's disease. J Neural Transm (Vienna). 2010, 117(10), 1177-81. [CrossRef]
- Weinstein, G., Beiser, A. S., Preis, S. R., Courchesne, P., Chouraki, V., Levy, D., & Seshadri, S. Plasma clusterin levels and risk of dementia, Alzheimer's disease, and stroke. Alzheimers Dement (Amst). 2016, 3, 103-9. [CrossRef]
- Schrijvers, E.M., Koudstaal, P. J., Hofman, A., & Breteler, M. M. Plasma clusterin and the risk of Alzheimer disease. JAMA. 2011, 305(13), 1322-6. [CrossRef]
- Chen, D., et al. Antisense oligonucleotide against clusterin regulates human hepatocellular carcinoma invasion through transcriptional regulation of matrix metalloproteinase-2 and E-cadherin. Int J Mol Sci. 2012, 13(8), 10594-607. [CrossRef]
- Cheng, C.Y., Wang, Y., Zhang, K., Jiao, X., Yan, B., & Liang, J. Regulation of chemosensitivity and migration by clusterin in non-small cell lung cancer cells. Cancer Chemother Pharmacol. 2012, 69(1), 145-54.
- Lourda, M., I.P., Trougakos, & E.S. Gonos. Development of resistance to chemotherapeutic drugs in human osteosarcoma cell lines largely depends on up-regulation of Clusterin/Apolipoprotein J. Int J Cancer. 2007, 120(3), 611-22. [CrossRef]
- Yang, G., Zhang, H., Liu, Y., Zhou, J., He, W., Quick, C. M., Xie, D., Smoller, B. R., & Fan, C. Y. Epigenetic and immunohistochemical characterization of the Clusterin gene in ovarian tumors. Arch Gynecol Obstet. 2013, 287(5), 989-95. [CrossRef]
- Tellez, T., García-Aranda, M., & Redondo, M. The role of clusterin in carcinogenesis and its potential utility as a therapeutic target. Curr Med Chem. 2016, 23, 1-12 (In Press). [CrossRef]
- Trougakos, I.P., Pawelec, G., Tzavelas, C., Ntouroupi, T., & Gonos, E. S. Clusterin/Apolipoprotein J up-regulation after zinc exposure, replicative senescence, or differentiation of human hematopoietic cells. Biogerontolog. 2006, 7(5-6), 375-82. [CrossRef]
- http://www.ensembl.org. Available at: http://www.ensembl.org/Homo_sapiens/Gene/Summary?g=ENSG00000120885,r=8:27596917-27615031.
- http://vega.sanger.ac.uk. Available at: http://vega.sanger.ac.uk/Homo_sapiens/Gene/Summary?g=OTTHUMG00000102114,r=8:27596917-27615031.
- Bonacini, M., Coletta, M., Ramazzina, I., Naponelli, V., Modernelli, A., Davalli, P., Bettuzzi, S., & Rizzi, F. Distinct promoters, subjected to epigenetic regulation, drive the expression of two clusterin mRNAs in prostate cancer cells. Biochim Biophys Acta. 2015, 1849(1), 44-54. [CrossRef]
- Rizzi, F., M. Coletta, & S. Bettuzzi. Chapter 2: Clusterin (CLU): From one gene and two transcripts to many proteins. Adv Cancer Res. 2009, 104, 9-23.
- Yu, J.T. & L. Tan. The role of clusterin in Alzheimer's disease: pathways, pathogenesis, and therapy. Mol Neurobiol. 2012, 45(2), 314-26.
- Foster, E.M., Dangla-Valls, A., Lovestone, S., Ribe, E.M., Buckley, N.J. Clusterin in Alzheimer's Disease: Mechanisms, Genetics, and Lessons from Other Pathologies. Front. Neurosci. 2019, 13, 164.
- Rohne, P., H. Prochnow, & C. Koch-Brandt. The CLU-files: disentanglement of a mystery. Biomol Concepts. 2016, 7(1), 1-15. [CrossRef]
- Zhang, Q., Zhou, W., Kundu, S., Jang, T. L., Yang, X., Pins, M., Smith, N., Jovanovic, B., Xin, D., Liang, L., Guo, Y., & Lee, C. The leader sequence triggers and enhances several functions of clusterin and is instrumental in the progression of human prostate cancer in vivo and in vitro. BJU Int. 2006, 98(2), 452-60. [CrossRef]
- Poon, S., Treweek, T. M., Wilson, M. R., Easterbrook-Smith, S. B., & Carver, J. A. Clusterin is an extracellular chaperone that specifically interacts with slowly aggregating proteins on their off-folding pathway. FEBS Lett. 2002, 513(2-3), 259-66. [CrossRef]
- Trougakos, I.P., Gonos, E.S. Regulation of clusterin/apolipoprotein J, a functional homologue to the small heat shock proteins, by oxidative stress in aging and age-related diseases. Free. Radic. Res. 2006, 40, 1324–1334. [CrossRef]
- Bettuzzi, S., Scorcioni, F., Astancolle, S., Davalli, P., Scaltriti, M., Corti, A. Clusterin (SGP-2) transient overexpression decreases proliferation rate of SV40-immortalized human prostate epithelial cells by slowing down cell cycle progression. Oncogene 2002, 21, 4328–4334. [CrossRef]
- Pucci, S., Bonanno, E., Pichiorri, F., Angeloni, C., Spagnoli, L. G. Modulation of different clusterin isoforms in human colon tumorigenesis. Oncogene 2004, 23, 2298–2304. [CrossRef]
- Caccamo, A. E., Scaltriti, M., Caporali, A., D'Arca, D., Corti, A., Corvetta, D., Sala, A., Bettuzzi, S. Ca2+ depletion induces nuclear clusterin, a novel effector of apoptosis in immortalized human prostate cells. Cell Death Differ. 2005, 12, 101–104. [CrossRef]
- Caccamo, A.E., Scaltriti, M., Caporali, A., D'Arca, D., Scorcioni, F., Astancolle, S., Mangiola, M., Bettuzzi, S. Cell detachment and apoptosis induction of immortalized human prostate epithelial cells are associated with early accumulation of a 45 kDa nuclear isoform of clusterin. Biochem. J. 2004, 382, 157–168. [CrossRef]
- Leskov, K.S., Klokov, D.Y., Li, J., Kinsella, T.J., Boothman, D.A. Synthesis and functional analyses of nuclear clusterin, a cell death protein. J. Biol. Chem. 2003, 278, 11590–11600. [CrossRef]
- Li, N., Zoubeidi, A., Beraldi, E., Gleave, M. E. GRP78 regulates clusterin stability, retrotranslocation and mitochondrial localization under ER stress in prostate cancer. Oncogene 2013, 32, 1933–1942. [CrossRef]
- Trougakos, I.P., Gonos, E.S. Regulation of clusterin/apolipoprotein J, a functional homologue to the small heat shock proteins, by oxidative stress in aging and age-related diseases. Free. Radic. Res. 2006, 40, 1324–1334. [CrossRef]
- Nizard, P., Tetley, S., Le Dréan, Y., Watrin, T., Le Goff, P., Wilson, M.R., and Michel, D. Stress-Induced Retrotranslocation of Clusterin/ApoJ into the Cytosol. Traffic 2007, 8, 554–565. [CrossRef]
- Scaltriti, M., Bettuzzi, S., Sharrard, R.M., Caporali, A., Caccamo, A.E., Maitland, N.J. Clusterin overexpression in both malignant and nonmalignant prostate epithelial cells induces cell cycle arrest and apoptosis. Br. J. Cancer 2004, 91, 1842–1850. [CrossRef]
- Scaltriti, M., Santamaria, A., Paciucci, R., Bettuzzi, S. Intracellular clusterin induces G2-M phase arrest and cell death in PC-3 prostate cancer cells1. Cancer Res. 2004, 64, 6174–6182. [CrossRef]
- Park, S., Mathis, K.W., Lee, I.K. The physiological roles of apolipoprotein J/clusterin in metabolic and cardiovascular diseases. Rev. Endocr. Metab. Disord. 2014, 15, 45–53. [CrossRef]
- Zoubeidi, A., Chi, K., Gleave, M. Targeting the cytoprotective chaperone, clusterin, for treatment of advanced cancer. Clin. Cancer Res. 2010, 16, 1088–1093. [CrossRef]
- Goetz, E. M., Shankar, B., Zou, Y., Morales, J. C., Luo, X., Araki, S., Bachoo, R., Mayo, L. D., Boothman, D. A. ATM dependent IGF-1 induction regulates secretory clusterin expression after DNA damage and in genetic instability. Oncogene. Nat. Publ. Group 2011, 30, 3745–3754. [CrossRef]
- Prochnow, H., Gollan, R., Rohne, P., Hassemer, M., Koch-Brandt, C., Baiersdörfer, M. Non-Secreted Clusterin Isoforms Are Translated in Rare Amounts from Distinct Human mRNA Variants and Do Not Affect BaxMediated Apoptosis or the NF-κB Signaling Pathway. PLoS ONE 2013, 8, e75303.
- Michel, D., Chatelain, G., North, S., Brun, G. Stress-induced transcription of the clusterin/apoJ gene. Biochem. J. 1997, 50, 45–50. [CrossRef]
- Trougakos, I.P. The Molecular Chaperone Apolipoprotein J/Clusterin as a Sensor of Oxidative Stress: Implications in Therapeutic Approaches—A Mini-Review. Gerontology 2013, 59, 514–523. [CrossRef]
- July, L.V., Akbari, M., Zellweger, T., Jones, E.C., Goldenberg, S.L., Gleave, M.E. Clusterin expression is significantly enhanced in prostate cancer cells following androgen withdrawal therapy. Prostate 2002, 50, 179–188. [CrossRef]
- FitzGerald, P. C., Shlyakhtenko, A., Mir, A. A., & Vinson, C. Clustering of DNA sequences in human promoters. Genome research. 2004, 14(8), 1562–1574. [CrossRef]
- Kim, N., & Choi, W. S. Proapoptotic role of nuclear clusterin in brain. Anatomy & cell biology. 2011, 44(3), 169–175. [CrossRef]
- Wong, P., Taillefer, D., Lakins, J., Pineault, J., Chader, G., & Tenniswood, M. Molecular characterization of human TRPM-2/clusterin, a gene associated with sperm maturation, apoptosis and neurodegeneration. Eur J Biochem. 1994, 221(3), 917-25.
- Deb, M., Sengupta, D., Rath, S.K., Kar, S., Parbin, S., Shilpi, A., et al. Clusterin gene is predominantly regulated by histone modifications in human colon cancer and ectopic expression of the nuclear isoform induces cell death. Biochim. Biophys. Acta. Elsevier B.V. 2015, 1852, 1630–1645. [CrossRef]
- Suuronen, T., Nuutinen, T., Ryhänen, T., Kaarniranta, K., & Salminen, A. Epigenetic regulation of clusterin/apolipoprotein J expression in retinal pigment epithelial cells. Biochem Biophys Res Commun. 2007, 357(2), 397-401. [CrossRef]
- Hellebrekers, D. M., Melotte, V., Viré, E., Langenkamp, E., Molema, G., Fuks, F., Herman, J. G., Van Criekinge, W., Griffioen, A. W., & van Engeland, M. Identification of epigenetically silenced genes in tumor endothelial cells. Cancer research. 2007, 67(9), 4138–4148. [CrossRef]
- Serrano, A., Redondo, M., Tellez, T., Castro-Vega, I., Roldan, M.J., Mendez, R., Rueda, A., Jiménez, E. Regulation of clusterin expression in human cancer via DNA methylation. Tumour. Biol. 2009, 30, 286–291. [CrossRef]
- Handy, D.E., R. Castro, & J. Loscalzo, Epigenetic modifications: basic mechanisms and role in cardiovascular disease. Circulation. 2011, 123(19), 2145-56.
- Liao, F. T., Lee, Y. J., Ko, J. L., Tsai, C. C., Tseng, C. J., & Sheu, G. T. Hepatitis delta virus epigenetically enhances clusterin expression via histone acetylation in human hepatocellular carcinoma cells. The Journal of general virology. 2009, 90(Pt 5), 1124–1134. [CrossRef]
- Rauhala, H. E., Porkka, K. P., Saramäki, O. R., Tammela, T. L., & Visakorpi, T. Clusterin is epigenetically regulated in prostate cancer. International journal of cancer. 2008, 123(7), 1601–1609. [CrossRef]
- Rosemblit, N., Chen, C.L. Regulators for the rat clusterin gene: DNA methylation and cis-acting regulatory elements. J. Mol. Endocrinol. 1994, 13, 69–76. [CrossRef]
- Ambros V. The functions of animal microRNAs. Nature. 2004, 431(7006), 350–355. [CrossRef]
- Mydlarz, W., Uemura, M., Ahn, S., Hennessey, P., Chang, S., Demokan, S., Sun, W., Shao, C., Bishop, J., Krosting, J., Mambo, E., Westra, W., Ha, P., Sidransky, D., & Califano, J. Clusterin is a gene-specific target of microRNA-21 in head and neck squamous cell carcinoma. Clinical cancer research : an official journal of the American Association for Cancer Research. 2014, 20(4), 868–877.
- Chen, X., Jiang, Y., Huang, Z., Li, D., Chen, X., Cao, M., Meng, Q., Pang, H., Sun, L., Zhao, Y., & Cai, L. miRNA-378 reverses chemoresistance to cisplatin in lung adenocarcinoma cells by targeting secreted clusterin. Scientific reports. 2016, 6, 19455. [CrossRef]
- Buttyan, R., Olsson, C. A., Pintar, J., Chang, C., Bandyk, M., Ng, P. Y., & Sawczuk, I. S. Induction of the TRPM-2 gene in cells undergoing programmed death. Mol Cell Biol. 1989, 9(8), 3473-81. [CrossRef]
- Leger, J.G., M.L. Montpetit, & M.P. Tenniswood, Characterization and cloning of androgen-repressed mRNAs from rat ventral prostate. Biochem Biophys Res Commun, 1987, 147(1), 196-203. [CrossRef]
- Leger, J.G., R. Le Guellec, & M.P. Tenniswood, Treatment with antiandrogens induces an androgen-repressed gene in the rat ventral prostate. Prostate. 1988, 13(2), 131-42. [CrossRef]
- Sensibar, J.A., Sutkowski, D. M., Raffo, A., Buttyan, R., Griswold, M. D., Sylvester, S. R., Kozlowski, J. M., & Lee, C. Prevention of cell death induced by tumor necrosis factor alpha in LNCaP cells by overexpression of sulfated glycoprotein-2 (clusterin). Cancer Res. 1995, 55(11), 2431-7.
- Schwochau, G.B., K.A. Nath, & M.E. Rosenberg. Clusterin protects against oxidative stress in vitro through aggregative and nonaggregative properties. Kidney Int. 1998, 53(6), 1647-53. [CrossRef]
- Leskov, K.S., Klokov, D. Y., Li, J., Kinsella, T. J., & Boothman, D. A. Synthesis and functional analyses of nuclear clusterin, a cell death protein. J Biol Chem. 2003, 278(13), 11590-600. [CrossRef]
- Yang, C.R., L.K., Hosley-Eberlein, K., Criswell, T., Pink, J., Kinsella, T., Boothman, D. Nuclear Clusterin/XIP8, an x-ray-induced Ku70-binding protein that signals cell death. PNAS. 2000, 97 (11), 5907-12.
- Leskov, K.S., Araki, S., Lavik, J. P., Gomez, J. A., Gama, V., Gonos, E. S., Trougakos, I. P., Matsuyama, S., & Boothman, D. A. CRM1 protein-mediated regulation of nuclear clusterin (nCLU), an ionizing radiation-stimulated, Bax-dependent pro-death factor. J Biol Chem. 2011, 286(46), 40083-90. [CrossRef]
- Essabbani, A., Garcia, L., Zonetti, M. J., Fisco, T., Pucci, S., & Chiocchia, G. Exon-skipping strategy by ratio modulation between cytoprotective versus pro-apoptotic clusterin forms increased sensitivity of LNCaP to cell death. PLoS One. 2013, 8(2), e54920. [CrossRef]
- Kim, N. & W.S. Choi. Proapoptotic role of nuclear clusterin in brain. Anat Cell Biol. 2011, 44(3), 169-75. [CrossRef]
- García-Aranda, M., Serrano, A., & Redondo, M. Regulation of Clusterin Gene Expression. Current protein & peptide science. 2017, 19. [CrossRef]
- Kim, N., Yoo, J. C., Han, J. Y., Hwang, E. M., Kim, Y. S., Jeong, E. Y., Sun, C. H., Yi, G. S., Roh, G. S., Kim, H. J., Kang, S. S., Cho, G. J., Park, J. Y., & Choi, W. S. Human nuclear clusterin mediates apoptosis by interacting with Bcl-XL through C-terminal coiled coil domain. J Cell Physiol. 2012, 227(3), 1157-67. [CrossRef]
- Shannan B, S.M., Leskov K, Willis J, Boothman D, Tilgen W, Reichrath J. Challenge and promise: roles for clusterin in pathogenesis, progression and therapy of cancer. Cell Death Differ. 2006, 13, 12-19.
- Prochnow, H., Gollan, R., Rohne, P., Hassemer, M., Koch-Brandt, C., & Baiersdörfer, M. Non-secreted clusterin isoforms are translated in rare amounts from distinct human mRNA variants and do not affect Bax-mediated apoptosis or the NF-kappaB signaling pathway. PLoS One. 2013, 8(9), e75303.
- Viard, I., Wehrli, P., Jornot, L., Bullani, R., Vechietti, J.-L., French, L.E., Schifferli, J.A., Tschopp, J. Clusterin gene expression mediates resistance to apoptotic cell death induced by heat shock and oxidative stress. J. Invest. Dermatol. 1999, 112, 290–296. [CrossRef]
- Nicholson, G., Lawrence, A., Ala, F.A., Bird, G.W. Semi-quantitative assay of antigen site density by flow cytometry analysis. Transfusion. Medicine. 1991, 1, 87–90.
- Trougakos, I.P., Lourda, M., Antonelou, M.H., Kletsas, D., Gorgoulis, V.G., Papassideri, I.S., Zou, Y., Margaritis, L.H., Boothman, D.A., Gonos, E.S. Intracellular clusterin inhibits mitochondrial apoptosis by suppressing stress signals activating p53 and stabilizing the cytosolic Ku70-Bax protein complex. Clin. Cancer Res. 2009, 15, 48–59. [CrossRef]
- Blume, A.J., Foster, C.J. Adenylate cyclase of mouse neuroblastoma cells: regulation by 2-chloroadenosine, prostaglandin E1, and the cations Mg2+, Ca2+, and Mn2+. J. Neurochem. 1976, 26, 305–311.
- Scaltriti, M., Bettuzzi, S., Sharrard, R.M., Caporali, A., Caccamo, A.E., Maitland, N.J. Overexpression of clusterin in both malignant and nonmalignant prostate epithelial cells induces cell cycle arrest and apoptosis. Br. J. Cancer. 2004, 91, 1842–1850. [CrossRef]
- Cervellera, M., Raschella, G., Santilli, G., Tanno, B., Ventura, A., Mancini, C., Sevignani, C., Calabretta, B., Sala, A. Direct transactivation of the antiapoptotic gene apolipoprotein J (Clusterin) by B-MYB. J. Biol. Chem. 2000, 275, 21055–21060. [CrossRef]
- Ma, X., Bai, Y. IGF-1 activates the P13K/AKT signaling pathway by positively regulating secretory clusterin. Mol. Med. Rep. 2012, 6, 1433-1437.
- Flanagan, L., Whyte, L., Chatterjee, N., Tenniswood, M. Effects of clusterin overexpression on metastatic progression and therapy in breast cancer. BMC Cancer. 2010, 10, 107. [CrossRef]
- Meunier, A., Cornet, F., Campos, M. Bacterial cell proliferation: from molecules to cells. FEMS Microbiol Rev. 2020. [CrossRef]
- Wang, Z. Regulation of cell cycle progression by growth factor-induced cell signaling. Cells. 2021, 10, 3327. [CrossRef]
- Zhang, Y., Lv, X., Chen, L., & Liu, Y. The role and function of CLU in cancer biology and therapy. Clinical and experimental medicine. 2023, 23(5), 1375–1391. [CrossRef]
- Dews, M., Fox, J. L., Hultine, S., Sundaram, P., Wang, W., Liu, Y. Y., Furth, E., Enders, G. H., El-Deiry, W., Schelter, J. M., Cleary, M. A., & Thomas-Tikhonenko, A. The myc-miR-17~92 axis blunts TGF{beta} signaling and production of multiple TGF{beta}-dependent antiangiogenic factors. Cancer research. 2010, 70(20), 8233–8246.
- Suzuki, S., Shiraga, K., Sato, S., Punfa, W., Naiki-Ito, A., Yamashita, Y., Shirai, T., & Takahashi, S. Apocynin, an NADPH oxidase inhibitor, suppresses rat prostate carcinogenesis. Cancer science. 2013, 104(12), 1711–1717. [CrossRef]
- Huang, H., Wang, L., Li, M., Wang, X., & Zhang, L. Secreted clusterin (sCLU) regulates cell proliferation and chemosensitivity to cisplatin by modulating ERK1/2 signals in human osteosarcoma cells. World journal of surgical oncology. 2014, 12, 255.
- Wang, X., Xie, J., Lu, X., Li, H., Wen, C., Huo, Z., Xie, J., Shi, M., Tang, X., Chen, H., Peng, C., Fang, Y., Deng, X., & Shen, B. Melittin inhibits tumor growth and decreases resistance to gemcitabine by downregulating cholesterol pathway gene CLU in pancreatic ductal adenocarcinoma. Cancer Lett. 2017, 399, 1–9. [CrossRef]
- Chen, Z., Fan, Z., Dou, X., Zhou, Q., Zeng, G., Liu, L., Chen, W., Lan, R., Liu, W., Ru, G., Yu, L., He, Q. Y., & Chen, L. Inactivation of tumor suppressor gene Clusterin leads to hyperactivation of TAK1-NF-kappaB signaling axis in lung cancer cells and denotes a therapeutic opportunity. Theranostics. 2020, 10(25), 11520–34.
- Chun, Y.J. Knockdown of clusterin expression increases the in vitro sensitivity of human prostate cancer cells to paclitaxel. J Toxicol Environ Health A. 2014, 77(22–24), 1443–50. [CrossRef]
- Zheng, W., Yao, M., Qian, Q., Sai, W., Qiu, L., Yang, J., Wu, W., Dong, Z., & Yao, D. Oncogenic secretory clusterin in hepatocellular carcinoma: Expression at early staging and emerging molecular target. Oncotarget. 2016, 8(32), 52321–32. [CrossRef]
- Bailes, J., Soloviev, M. Insulin-Like Growth Factor-1 (IGF-1) and Its Monitoring in Medical Diagnostic and in Sports. Biomolecules. 2021, 11(2), 217. [CrossRef]
- Ma, X., Bai, Y. IGF-1 activates the P13K/AKT signaling pathway via upregulation of secretory clusterin. Mol Med Rep. 2012, 6(6), 1433–7. [CrossRef]
- Takeuchi, A., Shiota, M., Beraldi, E., Thaper, D., Takahara, K., Ibuki, N., Pollak, M., Cox, M. E., Naito, S., Gleave, M. E., & Zoubeidi, A. Insulin-like growth factor-I induces CLU expression through Twist1 to promote prostate cancer growth. Mol Cell Endocrinol. 2014, 384(1–2), 117–25. [CrossRef]
- Chesnokova, V., Zonis, S., Wawrowsky, K., Tani, Y., Ben-Shlomo, A., Ljubimov, V., Mamelak, A., Bannykh, S., & Melmed, S. Clusterin and FOXL2 act concordantly to regulate pituitary gonadotroph adenoma growth. Mol Endocrinol. 2012, 26(12), 2092–103. [CrossRef]
- Deng, J., Peng, M., Zhou, S., Xiao, D., Hu, X., Xu, S., Wu, J., & Yang, X. Metformin targets Clusterin to control lipogenesis and inhibit the growth of bladder cancer cells through SREBP-1c/FASN axis. Signal Transduct Target Ther. 2021, 6(1), 98. [CrossRef]
- Liu, Y., Men, C., Xu, Y., Zhao, K., Luo, L., Dong, D., & Yu, Q. Clusterin promotes growth and invasion of clear cell renal carcinoma cell by upregulation of S100A4 expression. Cancer Biomark. 2018, 21(4), 915–23. [CrossRef]
- Pajak, B., Kania, E., Gajkowska, B., & Orzechowski, A. Lipid rafts mediate epigallocatechin-3-gallate- and green tea extract-dependent viability of human colon adenocarcinoma COLO 205 cells; clusterin affects lipid rafts-associated signaling pathways. J Physiol Pharmacol Official J Polish Physiol Soc. 2011, 62(4), 449–59.
- Tang, Z., Wang, W., Liu, Z., Sun, X., Liao, Z., Chen, F., Jiang, G., Huo, G. Blocking ERK signaling pathway reduces MMP-9 expression to alleviate cerebral edema after traumatic brain injury in rats. Nan Fang Yi Ke Da Xue Xue Bao. 2020, 40, 1018-1022.
- Shim, Y.-J., Kang, B.-H., Jeon, H.-S., Park, I.-S., Lee, K.-U., Lee, I.-K., Park, G.-H., Lee K.-M., Schedin P., Min B.-H. Clusterin induces matrix metalloproteinase-9 expression via ERK1/2 and PI3K/Akt/NF-κB pathways in monocytes/macrophages. J. Leukoc. Biol. 2011, 90, 761–769.
- Li, Y., Lu, J., Zhou, S., Wang, W., Tan, G., Zhang, Z., Dong, Z., Kang, T., Tang, F. N, N′-dinitrosopiperazine-induced clusterin participates in nasopharyngeal carcinoma metastasis. Oncotarget. 2016, 7, 5548–5563.
- Tian, Y., Wang, C., Chen, S., Liu, J., Fu, Y., Luo, Y. Extracellular Hsp90alpha and clusterin synergistically promote breast cancer epithelial-to-mesenchymal transition and metastasis through LRP1. J. Cell Sci. 2019, 132, jcs228213.
- Yang, P., Yang, Z., Dong, Y., Yang, L., Peng, S., Yuan, L., Hu, X., Chen, S., Tang, H., Yang, X., et al. Clusterin is a prognostic biomarker of breast cancer and correlates with the immune microenvironment. Transl Cancer Res. 2023, 12, 31–45.
- Yang, S., Tang, W., Azizian, A., Gaedcke, J., Strobel, P., Wang, L., Cawley, H., Ohara, Y., Valenzuela, P., Zhang L., et al. Dysregulation of the HNF1B/Clusterin axis enhances disease progression in a highly aggressive subset of pancreatic cancer patients. Carcinogenesis. 2022, 43, 1198-1210. [CrossRef]
- Mansoori, B., Mohammadi, A., Davudian, S., Shirjang, S., Baradaran, B. Different mechanisms of resistance to cancer drugs: a brief review. Adv. Pharm. Bull. 2017, 7, 339–348.
- Whitesell, L., Lindquist, S.L. HSP90 and the chaperoning of cancer. Nat. Rev. Cancer. 2005, 5, 761–772. [CrossRef]
- Zhang, H., Kim, J.K., Edwards, C.A., Xu, Z., Taichman, R., Wang, C.-Y. Clusterin inhibits apoptosis by interacting with activated Bax. Nat. Cell Biol. 2005, 7, 909–915. [CrossRef]
- Tang, M., Li, J., Liu, B., Song, N., Wang, Z., Yin, C. Clusterin expression in human testicular seminoma. Med. Hypotheses. 2013, 81, 635–637. [CrossRef]
- Wang, X., Zou, F., Zhong, J., Yue, L., Wang, F., Wei, H., Yang, G., Jin, T., Dong, X., Li J., et al. Secretory clusterin mediates resistance to oxaliplatin through the Gadd45a/PI3K/Akt signaling pathway in hepatocellular carcinoma. J. Cancer. 2018, 9, 1403-1413.
- Panico, F., Rizzi, F., Fabbri, L. M., Bettuzzi, S., & Luppi, F. Clusterin (CLU) and lung cancer. Advances in cancer research. 2009, 105, 63–76.
- July, L. V., Beraldi, E., So, A., Fazli, L., Evans, K., English, J. C., Gleave, M. E. Nucleotide-based therapies targeting clusterin chemosensitize human lung adenocarcinoma cells both in vitro and in vivo. Mol. Cancer Ther. 2004, 3, 223–232.
- Bi, J., Guo, A. L., Lai, Y. R., Li, B., Zhong, J. M., Wu, H. Q., Xie, Z., He, Y. L., Lv, Z. L., Lau, S. H., et al. Overexpression of clusterin correlates with tumor progression, metastasis in gastric cancer: A study on tissue microarrays. Neoplasm. 2010, 57, 191. [CrossRef]
- Xie, D., Lau, S.H., Sham, J.S., Wu, Q.L., Fang, Y., Liang, L.Z., Che, L.H., Zeng, Y.X., Guan, X.Y. Up-regulated expression of cytoplasmic clusterin in human ovarian carcinoma. Cancer. 2005, 103, 277–283. [CrossRef]
- Yang, G. F., Li, X. M., & Xie, D. Overexpression of clusterin in ovarian cancer is correlated with impaired survival. International journal of gynecological cancer: official journal of the International Gynecological Cancer Society. 2009, 19(8), 1342–1346.
- Al-Maghrabi, J. A., Butt, N. S., Anfinan, N., Sait, K., Sait, H., Bajouh, O., & Khabaz, M. N. Clusterin immunoexpression is associated with early stage endometrial carcinomas. Acta histochemical. 2016, 118(4), 430–434. [CrossRef]
- Fuzio, P., Valletti, A., Napoli, A., Napoli, G., Cormio, G., Selvaggi, L., Liuni, S., Pesole, G., Maiorano, E., & Perlino, E. Regulation of the expression of CLU isoforms in endometrial proliferative diseases. International journal of oncology. 2013, 42(6), 1929–1944. [CrossRef]
- Won, Y. S., Lee, S. J., Yeo, S. G., & Park, D. C. Effects of female sex hormones on clusterin expression and paclitaxel resistance in endometrial cancer cell lines. International journal of medical sciences. 2012, 9(1), 86–92. [CrossRef]
- Park, D. C., Yeo, S. G., Shin, E. Y., Mok, S. C., & Kim, D. H. Clusterin confers paclitaxel resistance in cervical cancer. Gynecologic oncology. 2006, 103(3), 996–1000. [CrossRef]
- Niu, Z., Li, X., Hu, B., Li, R., Wang, L., Wu, L., & Wang, X. Small interfering RNA targeted to secretory clusterin blocks tumor growth, motility, and invasion in breast cancer. Acta biochimica et biophysica Sinica. 2012, 44(12), 991–998. [CrossRef]
- Li, J., Jia, L., Zhao, P., Jiang, Y., Zhong, S., & Chen, D. Stable knockdown of clusterin by vectorbased RNA interference in a human breast cancer cell line inhibits tumour cell invasion and metastasis. The Journal of international medical research. 2012, 40(2), 545–555. [CrossRef]
- Redondo, M., Villar, E., Torres-Muñoz, J., Tellez, T., Morell, M., Petito, C. K. Overexpression of clusterin in human breast carcinoma. Am. J. Pathol. 2000, 157, 393–39. [CrossRef]
- Zhang, D., Sun, B., Zhao, X., Cui, Y., Xu, S., Dong, X., Zhao, J., Meng, J., Jia, X., & Chi, J. Secreted CLU is associated with the initiation of triple-negative breast cancer. Cancer biology & therapy. 2012, 13(5), 321–329. [CrossRef]
- Niu, Z. H., Wang, Y., Chun, B., Li, C. X., & Wu, L. Secretory clusterin (sCLU) overexpression is associated with resistance to preoperative neoadjuvant chemotherapy in primary breast cancer. European review for medical and pharmacological sciences. 2013, 17(10), 1337–1344.
- Redondo, M., Tellez, T., and Roldan, M.J. The role of clusterin (CLU) in malignant transformation and drug resistance in breast carcinomas. Adv. Cancer Res. 2009, 105, 21–43.
- Mazzarelli, P., Pucci, S., Spagnoli, L.G. CLU and colon cancer. The dual face of CLU: From the normal to the malignant phenotype. Adv. Cancer Res. 2009, 105, 45–61.
- Pucci, S., Bonanno, E., Pichiorri, F., Angeloni, C., & Spagnoli, L. G. Modulation of different clusterin isoforms in human colon tumorigenesis. Oncogene. 2004, 23(13), 2298–2304. [CrossRef]
- Redondo, M.; Rodrigo, I.; Alcaide, J.; Tellez, T.; Roldan, M.J.; Funez, R.; Diaz-Martin, A.; Rueda, A.; Jiménez, E. Clusterin expression is associated with decreased disease-free survival of patients with colorectal carcinomas. Histopathology. 2010, 56, 932–936. [CrossRef]
- Sun, B., Moibi, J. A., Mak, A., Xiao, Z., Roa, W., & Moore, R. B. Response of bladder carcinoma cells to TRAIL and antisense oligonucleotide, Bcl-2 or clusterin treatments. The Journal of urology. 2009, 181(3), 1361–1371. [CrossRef]
- Miyake, H., Gleave, M., Kamidono, S., Hara, I. Overexpression of clusterin in transitional cell carcinoma of the bladder is related to disease progression and recurrence. Urology. 2002, 59, 150–154. [CrossRef]
- Hazzaa, S. M., Elashry, O. M., & Afifi, I. K. Clusterin as a diagnostic and prognostic marker for transitional cell carcinoma of the bladder. Pathology oncology research:POR. 2010, 16(1), 101–109. [CrossRef]
- Lau, S. H., Sham, J. S., Xie, D., Tzang, C. H., Tang, D., Ma, N., Hu, L., Wang, Y., Wen, J. M., Xiao, G., Zhang, W. M., Lau, G. K., Yang, M., & Guan, X. Y. Clusterin plays an important role in hepatocellular carcinoma metastasis. Oncogene. 2006, 25(8), 1242–1250. [CrossRef]
- Chen, D., Wang, Y., Zhang, K., Jiao, X., Yan, B., & Liang, J. Antisense oligonucleotide against clusterin regulates human hepatocellular carcinoma invasion through transcriptional regulation of matrix metalloproteinase-2 and E-cadherin. International journal of molecular sciences. 2012, 13(8), 10594–10607. [CrossRef]
- Xiu, P., Dong, X., Dong, X., Xu, Z., Zhu, H., Liu, F., Wei, Z., Zhai, B., Kanwar, J. R., Jiang, H., Li, J., & Sun, X. Secretory clusterin contributes to oxaliplatin resistance by activating Akt pathway in hepatocellular carcinoma. Cancer science. 2013, 104(3), 375–382. [CrossRef]
- Patarat, R.; Riku, S.; Kunadirek, P.; Chuaypen, N.; Tangkijvanich, P.; Mutirangura, A.; Puttipanyalears, C. The expression of FLNA and CLU in PBMCs as a novel screening marker for hepatocellular carcinoma. Sci. Rep. 2021, 11, 14838. [CrossRef]
- Gao, G., Luan, X. Diagnostic performance of clusterin in hepatocellular carcinoma: A meta-analysis. Int. J. Biol. Markers. 2022, 37, 404–411. [CrossRef]
- Rasmy, H. S., Mohammed, H. A., Mohammed, E. S., Ahmed, A. S. M., & Isaac, A. Serum clusterin as a promising diagnostic and prognostic marker for hepatocellular carcinoma after locoregional treatment. The Egyptian journal of immunology. 2022, 29(2), 26–40. [CrossRef]
- Li, Y., Wang, X., Chen, Y. H., Tan, Q. Q., Liu, X. B., & Tan, C. Clusterin is upregulated by erastin, a ferroptosis inducer and exerts cytoprotective effects in pancreatic adenocarcinoma cells. Anti-cancer drugs. 2023, 10.1097/CAD.0000000000001561. Advance online publication.1. [CrossRef]
- Xie, M.J., Motoo, Y., Su, S.B., Mouri, H., Ohtsubo, K., Matsubara, F., Sawabu, N. Expression of clusterin in human pancreatic cancer. Pancreas. 2002, 25, 234–238. [CrossRef]
- Hoeller, C., Pratscher, B., Thallinger, C., Winter, D., Fink, D., Kovacic, B., Sexl, V., Wacheck, V., Gleave, M.E., Pehamberger, H., et al. Clusterin regulates drug-resistance in melanoma cells. J. Investig. Dermatol. 2005, 124, 1300–1307. [CrossRef]
- He, L.R., Liu, M.Z., Li, B.K., Rao, H.L., Liao, Y.J., Zhang, L.J., Guan, X.Y., Zeng, Y.X., Xie, D. Clusterin as a predictor for chemoradiotherapy sensitivity and patient survival in esophageal squamous cell carcinoma. Cancer Sci. 2009, 100, 2354–2360. [CrossRef]
- Bijian, K., Mlynarek, A. M., Balys, R. L., Jie, S., Xu, Y., Hier, M. P., Black, M. J., Di Falco, M. R., LaBoissiere, S., & Alaoui-Jamali, M. A. Serum proteomic approach for the identification of serum biomarkers contributed by oral squamous cell carcinoma and host tissue microenvironment. Journal of proteome research. 2009, 8(5), 2173–2185. [CrossRef]
- Redondo, M., Fùnez, R., Esteban, F. Apoptosis in the Development and Treatment of Laryngeal Cancer: Role of p53, Bcl-2 and Clusterin. 2009. In: Chen, G.G., Lai, P.B. (eds) Apoptosis in Carcinogenesis and Chemotherapy. Springer, Dordrecht.
- Wellmann, A., Thieblemont, C., Pittaluga, S., Sakai, A., Jaffe, E. S., Siebert, P., Raffeld, M. Detection of differentially expressed genes in lymphomas using cDNA arrays: Identification of clusterin as a new diagnostic marker for anaplastic large-cell lymphomas. Blood. 2000, 96, 398–404.
- Ma, J., Gao, W., Gao, J. sCLU as prognostic biomarker and therapeutic target in osteosarcoma. Bioengineered. 2019, 10, 229–239. [CrossRef]
- Rizzi, F., Bettuzzi, S. Clusterin (CLU) and prostate cancer. Adv. Cancer Res. 2009, 105, 1–19.
- Miyake, H., Gleave, M. E., Arakawa, S., Kamidono, S., & Hara, I. Introducing the clusterin gene into human renal cell carcinoma cells enhances their metastatic potential. The Journal of urology. 2002, 167(5), 2203–2208.
- Shi, H., Deng, J. H., Wang, Z., Cao, K. Y., Zhou, L., & Wan, H. Knockdown of clusterin inhibits the growth and migration of renal carcinoma cells and leads to differential gene expression. Molecular medicine reports. 2013, 8(1), 35–40. [CrossRef]
- Beheshti Namdar, A., Kabiri, M., Mosanan Mozaffari, H., Aminifar, E., Mehrad-Majd, H. Circulating clusterin levels and cancer risk: A systematic review and meta-analysis. Cancer Control. 2022, 29, 10732748211038437. [CrossRef]
- Chen, D., Wang, Y., Zhang, K., Jiao, X., Yan, B., & Liang, J. Antisense oligonucleotide against clusterin regulates human hepatocellular carcinoma invasion through transcriptional regulation of matrix metalloproteinase-2 and E-cadherin. Int J Mol Sci. 2012, 13(8), 10594–607. [CrossRef]
- Nishikawa, M., Miyake, H., Gleave, M., & Fujisawa, M. Effect of targeting clusterin using OGX-011 on antitumor activity of temsirolimus in a human renal cell carcinoma model. Target Oncol. 2016, 12(1), 1–11. [CrossRef]
- Yamanaka, K., Gleave, M., Muramaki, M., Hara, I., & Miyake, H. Enhanced radiosensitivity by inhibition of the anti-apoptotic gene clusterin using antisense oligodeoxynucleotide in a human bladder cancer model. Oncol Rep. 2005, 13(5), 885. [CrossRef]
- Chen, Q., Wang, Z., Zhang, K., Liu, X., Cao, W., Zhang, L., Zhang, S., Yan, B., Wang, Y., & Xia, C. Clusterin confers gemcitabine resistance in pancreatic cancer. World J Surgical Oncol. 2011, 9, 1–8. [CrossRef]
- Cao, C., Shinohara, E. T., Li, H., Niermann, K. J., Kim, K. W., Sekhar, K. R., Gleave, M., Freeman, M., & Lu, B. Clusterin as a therapeutic target for radiation sensitization in a lung cancer model. Int J Radiat Oncol Biol Phys. 2005, 63(4), 1228–36. [CrossRef]
- Zellweger, T., et al. Enhanced radiation sensitivity in prostate cancer by inhibition of the cell survival protein clusterin. Clin Cancer Res. 2002, 8(10), 3276–84.
- So, A., Chi, K., Miyake, H., Adomat, H., Kiyama, S., Skov, K., & Gleave, M. E. Knockdown of the cytoprotective chaperone, clusterin, chemosensitizes human breast cancer cells both in vitro and in vivo. Mol Cancer Ther. 2005, 4(12), 1837–49.
- July, L. V., Beraldi, E., So, A., Fazli, L., Evans, K., English, J. C., Gleave, M. E. Nucleotide-based therapies targeting clusterin chemosensitize human lung adenocarcinoma cells both in vitro and in vivo. Mol. Cancer Ther. 2004, 3, 223–232.
- Hoeller, C., Pratscher, B., Thallinger, C., Winter, D., Fink, D., Kovacic, B., Sexl, V., Wacheck, V., Gleave, M.E., Pehamberger, H., et al. Clusterin regulates drug-resistance in melanoma cells. J. Investig. Dermatol. 2005, 124, 1300–1307. [CrossRef]
- Ma, J., Gao, W., Gao, J. sCLU as prognostic biomarker and therapeutic target in osteosarcoma. Bioengineered. 2019, 10, 229–239. [CrossRef]
- Zwain, I., & Amato, P. Clusterin protects granulosa cells from apoptotic cell death during follicular atresia. Experimental cell research. 2000, 257(1), 101–110. [CrossRef]
- Kususda, Y., Miyake, H., Gleave, M. E., & Fujisawa, M. Clusterin inhibition using OGX-011 synergistically enhances antitumour activity of sorafenib in a human renal cell carcinoma model. British journal of cancer. 2012, 106(12), 1945–1952. [CrossRef]
- Hassan, M. K., Watari, H., Han, Y., Mitamura, T., Hosaka, M., Wang, L., Tanaka, S., & Sakuragi, N. Clusterin is a potential molecular predictor for ovarian cancer patient's survival: targeting clusterin improves response to paclitaxel. Journal of experimental & clinical cancer research: CR. 2011, 30(1), 113.
- Blumenstein, B., Saad, F., Hotte, S., Chi, K. N., Eigl, B., Gleave, M., & Jacobs, C. Reduction in serum clusterin is a potential therapeutic biomarker in patients with castration-resistant prostate cancer treated with custirsen. Cancer Med. 2013, 2(4), 468–77. [CrossRef]
- Higano, C. S. Potential use of custirsen to treat prostate cancer. OncoTargets and therapy. 2013, 6, 785–797. [CrossRef]
- Chi, K.N., Hotte, S.J., Yu, E.Y., Tu, D., Eigl, B.J., Tannock, I., Saad, F., North, S., Powers, J., Gleave, M.E., et al. Randomized phase II study of docetaxel and prednisone with or without OGX-011 in patients with metastatic castration-resistant prostate cancer. J. Clin. Oncol. 2010, 28, 4247–4254. [CrossRef]
- Chia, S., Dent, S., Ellard, S., Ellis, P. M., Vandenberg, T., Gelmon, K., Powers, J., Walsh, W., Seymour, L., & Eisenhauer, E. A. Phase II trial of OGX-011 in combination with docetaxel in metastatic breast cancer. Clin Cancer Res. 2009, 15(2), 708–13. [CrossRef]
- Laskin, J.J., Nicholas, G., Lee, C., Gitlitz, B., Vincent, M., Cormier, Y., Stephenson, J., Ung, Y., Sanborn, R., Pressnail B., et al. Phase I/II trial of custirsen (OGX-011), an inhibitor of clusterin, in combination with a gemcitabine and platinum regimen in patients with previously untreated advanced non-small cell lung cancer. J. Thorac. Oncol. 2012, 7, 579–586. [CrossRef]
- Ma, X., Zou, L., Li, X., Chen, Z., Lin, Q., & Wu, X. MicroRNA-195 regulates docetaxel resistance by targeting clusterin in prostate cancer. Biomed Pharmacother. 2018, 99, 445–50; Zhao, W., Wang, X., Jiang, Y., Jia, X., & Guo, Y. miR-217-5p inhibits invasion and metastasis of prostate cancer by targeting clusterin. Mamm Genome. 2021, 32(5), 371–80.
- Davalli, P., Rizzi, F., Caldara, G. F., Davoli, S., Corti, A., Silva, A., Astancolle, S., Vitale, M., Bettuzzi, S., Arcari, M., & Azzali, G. Chronic administration of green tea extract to TRAMP mice induces the collapse of Golgi apparatus in prostate secretory cells and results in alterations of protein post-translational processing. Int J Oncol. 2011, 39(6), 1521–7. [CrossRef]
- Zhu, Y., Chen, P., Gao, Y., Ta, N., Zhang, Y., Cai, J., Zhao, Y., Liu, S., & Zheng, J. MEG3 activated by Vitamin D inhibits colorectal cancer cells proliferation and migration via regulating clusterin. EBioMedicine. 2018, 30, 148–57. [CrossRef]
- Suzuki, S., Shiraga, K., Sato, S., Punfa, W., Naiki-Ito, A., Yamashita, Y., Shirai, T., & Takahashi, S. Apocynin, an NADPH oxidase inhibitor, suppresses rat prostate carcinogenesis. Cancer Sci. 2013, 104(12), 1711–7. [CrossRef]
- Wang, X., Xie, J., Lu, X., Li, H., Wen, C., Huo, Z., Xie, J., Shi, M., Tang, X., Chen, H., Peng, C., Fang, Y., Deng, X., & Shen, B. Melit-tin inhibits tumor growth and decreases resistance to gemcitabine by downregulating cholesterol pathway gene CLU in pancreatic ductal adenocarcinoma. Cancer Lett. 2017, 399, 1–9.
- Deng, J., Peng, M., Zhou, S., Xiao, D., Hu, X., Xu, S., Wu, J., & Yang, X. Metformin targets Clusterin to control lipogenesis and inhibit the growth of bladder cancer cells through SREBP-1c/FASN axis. Signal Transduct Target Ther. 2021, 6(1), 98. [CrossRef]
- Xiong, J., Wang, S., Chen, T., Shu, X., Mo, X., Chang, G., Chen, J. J., Li, C., Luo, H., & Lee, J. D. Verteporfin blocks Clusterin which is required for survival of gastric cancer stem cell by modulating HSP90 function. Int J Biol Sci. 2019, 15(2), 312–24. [CrossRef]
- https://clinicaltrials.gov/study/NCT02412462?intr=clusterin&page=2&rank=11, accessed on January 10, 2024.
Figure 1.
Generation of nCLU and sCLU. psCLU is transported to the ER and by the leader signaling peptide and then cleaved and glycosylated while being transported to the Golgi apparatus. The result is an 80 kDa protein consisting of the alpha and beta subunit by disulfide bonds which is secreted outside the cell. pnCLU does not undergo any excision or glycosylation process, it is localized in the cytoplasm of unstressed cells. pnCLU is converted into a mature form inside the nucleus, nCLU.
Figure 1.
Generation of nCLU and sCLU. psCLU is transported to the ER and by the leader signaling peptide and then cleaved and glycosylated while being transported to the Golgi apparatus. The result is an 80 kDa protein consisting of the alpha and beta subunit by disulfide bonds which is secreted outside the cell. pnCLU does not undergo any excision or glycosylation process, it is localized in the cytoplasm of unstressed cells. pnCLU is converted into a mature form inside the nucleus, nCLU.
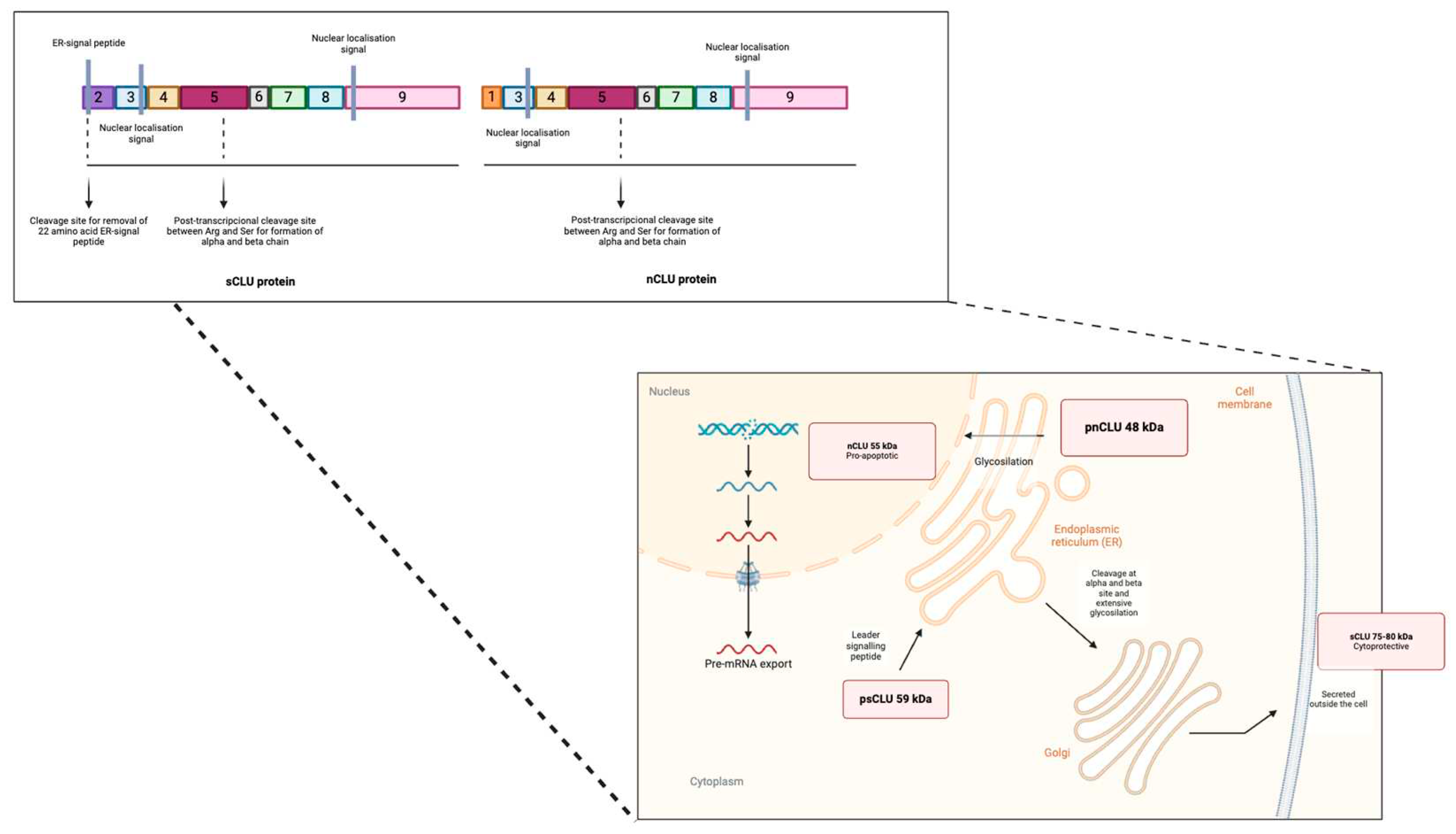

Table 1.
Clusterin as a potential biomarker in various types of cancer.
| Types of cancer | Expression of CLU in vitro | Expression of CLU in vivo |
|---|---|---|
| Non-small cell lung | Non-small cell lung cancer cell lines show overexpression upon treatment with chemotherapy or radiotherapy. ASO therapy sensitizes cells to these treatments and decreases their metastatic potential [112] | In patients with positive CLU expression there is a better overall disease-free survival than in negative patients [113]. |
| More than 80% of the tumors are inmunoreactive for clusterin [113]. | ||
| Gastric | Overexpression of sCLU correlates significantly with metastasis, tumor invasion and TNM stage. In addition, of with unfavorable survival for advanced-stage gastric cancers [114]. | |
| Ovarian | Overexpression of sCLU is inversely correlated with tumor apoptotic index and is more frequently detected in metastatic lesions than in primary tumors [115]. | |
| Increased sCLU expression is associated with increased biological aggressiveness and decreased survival [116]. | ||
| Endometrial | When clusterin is expressed in endometrial tumors, it is associated with a lower stage, supporting its role in the diagnosis of endometrial carcinoma [117]. | |
| There has been detected higher mRNA expression in both neoplastic and hyperplastic tissues compared to normal endometrium. In this regard, an increase in mRNA expression of the specific sCLU isoform has been observed in neoplastic and hyperplastic endometrial diseases, but an increase in CLU protein was not detected. Furthermore, specific CLU immunoreactivity has been observed in all glandular cells of the endometrium compared to other cellular compartments where CLU immunoreactivity was lower or absent [118]. | ||
| Increased CLU expression enhances paclitaxel resistance in endometrial cancer [119,120]. | ||
| Breast | Studies with the MDA-MB-231 cell line show how sCLU silencing significantly inhibits cell proliferation and drastically reduces cell invasion, cell progression and metastatic potential [121,122]. | Atypical hyperplasias, intraductal carcinomas, and invasive carcinomas are characterized by clusterin overexpression, unlike benign lesions [123]. |
| Overexpression of sCLU is observed in a higher percentage of triple-negative breast cancer [124] and is associated with a negative estrogen and progesterone receptor status [123]. | ||
| Likewise, overexpression in the stroma tends to directly correlate with resistance to preoperative neoadjuvant chemotherapy in the primary tumor and inversely with the apoptosis rate, indicating that gene expression may not be necessary for apoptotic cell death [123,125,126]. | ||
| Colon | sCLU is overexpressed, while nCLU is downregulated [127]. Similarly, sCLU overexpression was mainly shown in the cytoplasm of highly infiltrative tumors and metastatic lymph nodes [128], suggesting that clusterin expression could help identify patients with more aggressive tumors who may benefit from targeted therapies [129]. | |
| Bladder | Treatment with the antisense oligonucleotide (ASO) targeting negative regulation of Bcl-2 and clusterin increases the sensitivity of partially resistant bladder carcinoma cells to the tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) [130]. | The recurrence-free survival time of patients with overexpression of clusterin was shorter than that of patients with normal clusterin expression [131,132]. |
| Hepatocellular | High levels of sCLU are associated with migration, invasion, and metastasis [133] due to increased MMP-2 expression and decreased E-cadherin expression [134]. | |
| Furthermore, sCLU overexpression contributes to oxaliplatin resistance [135]. | ||
| In peripheral blood mononuclear cells (PBMC) from hepatocellular carcinoma patients, it has been proposed as a prospective detection biomarker along with other genes for its sensitivity and specificity [136]. The combination of CLU and AFP further improves diagnostic performance [137]. | ||
| The initial levels of clusterin are initially higher for patients with progressive disease than for those with partial or complete response, respectively [138]. | ||
| Pancreatic | The inducer of ferroptosis, a type of cell death characterized by the accumulation of reactive oxygen species (ROS), interferes with apoptotic cell death by regulating clusterin. [139]. | Clusterin expression in stages I and II is not significantly associated with apoptosis. Moreover, patients with positive clusterin expression present better survival rates [140]. |
| Melanomas | Overexpression is associated with increased drug resistance and prolonged survival of tumor cells, while silencing reduces resistance and reduced survival of melanoma cells both in vitro and in vivo [141]. | |
| Esophageal squamous cells | High CLU expression correlates with poor locoregional, overall, and distant progression-free survival. Moreover, patients with CLU overexpression in both epithelium and stroma have shorter survival times [142]. | |
| Head and neck | Overexpression of CLU has been observed, but its implications have not yet been determined [143]. | Although is detected in a low proportion of laryngeal carcinomas, it seems to exert a significant role in local invasiveness [144]. |
| Anaplastic large cell lymphomas | The role of CLU is unknown, but its expression within this lymphoma type provides an additional marker for diagnosis [145]. | |
| Clusterin expression is not related to anaplastic lymphoma kinase-1 (ALK-1) expression, and in reactive lymphoid tissues, only fibroblastic reticular cells and follicular dendritic cells show positive expression [145]. | ||
| Osteosarcoma | sCLU overexpression is associated with metastasis and chemotherapy resistance [146]. | |
| Prostate | The expression of CLU increases in advanced stages of cancer, and its suppression sensitizes cells to chemotherapeutic drugs [147]. | It has been observed that clusterin expression decreases considerably compared to benign tissues [147]. |
| Renal | The introduction of the CLU gene enhances the metastatic potential of renal cell cancer [148], while the removal of the CLU gene inhibits growth and migration [149]. | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



