
Submitted:
08 February 2024
Posted:
09 February 2024
You are already at the latest version
Abstract
This review article discusses the involvement of the Brain-Derived Neurotrophic Factor (BDNF) in the control of pain. BDNF, a neurotrophin known for its essential role in neuronal survival and plasticity, has garnered significant attention for its potential implications as a modulator of nociception and pain. This comprehensive review aims to provide insights into the multifaceted interactions between BDNF and pain pathways, encompassing both physiological and pathological pain conditions. I delve into the molecular mechanisms underlying BDNF's involvement in pain processing and discuss potential therapeutic applications of BDNF and its mimetics in managing pain. Furthermore, I highlight recent advancements and challenges in translating BDNF-related research into clinical practice.
Keywords:
BDNF
; Nociception
; Pain
; Neurons
; Glia
; Presynaptic mechanisms
; Postsynaptic mechanisms
; LTP
; Neuropeptides
1. Introduction
1.1. A Brief Overview of BDNF Functions
Brain-derived neurotrophic factor (BDNF) is a pivotal molecule in the field of neuroscience and neurobiology, playing a crucial role in the growth, development, maintenance, and plasticity of the nervous system [1]. BDNF is a member of the neurotrophin family of proteins, which includes nerve growth factor (NGF), neurotrophin-3 (NT-3), and neurotrophin-4/5 (NT-4/5). It is primarily synthesized as a precursor protein known as proBDNF (Figure 1). This precursor molecule is then proteolytically cleaved to generate mature BDNF, which is the biologically active form [2], although proBDNF has been recognized to also play a role in regulating neuronal activity [3].
The mature BDNF protein consists of 247 amino acids and forms a homodimer. Each monomer consists of a long pro-domain, followed by a mature BDNF domain. The pro-domain is cleaved during secretion to release the biologically active mature BDNF. This cleavage process is essential for the correct functioning of BDNF. Once released, BDNF can bind to its high-affinity receptor, the tropomyosin receptor kinase B (trkB), and initiate a series of intracellular signaling pathways that play a crucial role in neuronal survival, growth, and plasticity [2]. ProBDNF and mature BDNF activate different receptors to produce different effects on neurons. Specifically, proBDNF activates the low-affinity p75 pan neurotrophin receptor (p75NTR) rather than trkB, and years ago, it was demonstrated neurons release both proBDNF and mature BDNF [3].
BDNF is primarily known for its role in the development, maintenance, and plasticity of the nervous system. However, the role of BDNF extends beyond the realm of neuroscience and influences various aspects of human health and disease. The several functions of BDNF, span from the regulation of neuronal differentiation, survival, and growth, to the intervention in synaptic plasticity [4,5], neuroprotection [6,7], neurodegeneration [1], and the control of mood disorders [4]. Thus, most of the (initially) discovered functions of this neurotrophin play a primary role in shaping the cellular and functional organization of the normal brain, but more recent findings have converged to demonstrate that dysregulation of BDNF is implicated in a range of neurological disorders, including Alzheimer’s disease [6,7], Parkinson’s disease [8], Huntington’s disease [9,10], and amyotrophic lateral sclerosis (ALS) [11,12] and that low BDNF levels associated with depression [13] and anxiety [14].
BDNF also intervenes in the modulation of pain, the subject of this contribution.
1.2. Pain as a Complex and Multifaceted Sensory Experience
Pain is a universal and complex phenomenon in the animal world that has intrigued scientists, clinicians, and philosophers for centuries. It transcends mere sensory perception and encompasses a multidimensional experience that involves not only the discernment of noxious stimuli but also emotional, cognitive, and behavioral responses. This multifaceted nature of pain makes it a subject of great interest and study in various fields, including neuroscience, psychology, and clinical medicine.
Pain can be broadly defined as an unpleasant sensory and emotional experience associated with actual or potential tissue damage [16]. It serves as a vital protective mechanism, alerting individuals to harmful stimuli and prompting them to take action to prevent further injury. However, pain is far from being a simple “alarm system” for the body; instead, it exhibits complexity at multiple levels.
At its core, pain begins with nociception, the detection of noxious or potentially harmful stimuli by specialized receptors known as nociceptors [17]. These receptors lie in the peripheral nervous system (PNS) and are scattered throughout the body, from the skin to the internal organs [18]. When stimulated, nociceptors initiate a cascade of events that lead to the transmission of pain signals to the central nervous system (CNS), via the spinal nerves and the ascending pathways to the brain and some cranial nerves that possess somatic and/or visceral sensory fibers [19]. Beyond nociception, i.e., the encoding of a noxious stimulus and its transduction into electric signals, pain involves emotional and affective components. The brain processes these components of pain in regions associated with emotions, such as the amygdala and anterior cingulate cortex [20]. This emotional aspect of pain gives rise to the subjective experience of suffering, distress, and fear often associated with painful stimuli. Emotional responses to pain can vary greatly among individuals and are influenced by factors like previous experiences, psychological state, and cultural background. Cognition plays a critical role in shaping the perception of pain. Cognitive factors, such as attention, expectation, and belief, can significantly influence how individuals perceive and respond to painful stimuli. Cognitive strategies, such as distraction or cognitive reappraisal, can either exacerbate or mitigate the experience of pain. Pain is also influenced by sociocultural and contextual factors.
According to the International Association for the Study of Pain (IASP), there are several types of pain (https://www.iasp-pain.org/resources/terminology/) [16]. Nociceptive pain is the pain that arises from non-neural tissues and is a consequence of the activation of nociceptors. Neuropathic pain is a type of pain caused by a lesion or disease of the somatosensory nervous system. It can be further classified as central and peripheral according to the division of the somatosensory system that is lesioned. Nociplastic pain arises from altered nociception, despite no clear evidence of actual or threatened tissue damage activating peripheral nociceptors or evidence of disease or lesion of the somatosensory system causing the pain. There are also differences between acute and chronic pain [21]. Acute pain is self-limited, is triggered by a particular illness or injury, and has a protective function. On the other hand, chronic pain might be seen as a medical condition. It is discomfort that, whether connected to a sickness or injury, lasts longer than the typical recovery period. Chronic pain lacks a biological purpose, can have psychological causes, and has no obvious endpoint. Chronic pain conditions, characterized by persistent pain lasting beyond the expected healing period, represent a particularly challenging aspect of pain.
1.3. Overview of the Anatomical Arrangement of Pain Pathways
This section describes the general organization of somatic and visceral pain pathways (Figure 2). More comprehensive accounts can be found e.g., in ref. [19,22].
1.3.1. Somatic Pain Pathways – Subcortical Structures
The somatic pain pathways [23] are responsible for gathering sensory input from several sources, including the skin, muscles, joints, ligaments, and bones. Except for the head and the proximal regions of the neck, which receive sensory innervation from the peripheral projections of the trigeminal primary sensory neurons (PSNs), the transmission of pain signals from all other body parts is initially carried out by the PSNs of the dorsal root ganglia (DRGs). From these two sets of neurons originate the main pathways responsible for transmitting nociceptive signals to the brain: the trigeminothalamic (TTP) and spinothalamic (STP) pathways. Both are composed of a polysynaptic chain including three sensory neurons, commonly known as first, second, and third-order somatic sensory neurons. The sensory discriminative features of pain perception are reliant upon the crucial role of these cells. The first-order sensory neurons, i.e., the PSNs, are situated in the trigeminal ganglion (TG), the proximal ganglia of the glossopharyngeal and vagus nerves (the petrosal – PG and nodose NG ganglia), or the DRGs. The second-order sensory neurons are located within the dorsal horn of the spinal cord or the spinal nucleus of the trigeminal nerve (SNTN). These are projection neurons that give rise to long axons crossing the midline and ascending throughout the spinal cord and/or brainstem, ultimately reaching the thalamus. Third-order sensory neurons are in the ventroposterior lateral nucleus of the thalamus (VPLN) and project their axons to the first-order (S1) and second-order (S2) somatosensory area of the cerebral cortex. Nociceptive signal modulation can take place at various points within this polyneuronal chain. It is primarily observed at the substantia gelatinosa of the dorsal horn (lamina II) of the spinal cord or the SNTN. Additionally, several descending pathways can exert inhibitory or facilitatory effects on the trigeminothalamic or spinothalamic neurons.
Other ascending pathways play a significant role in the overall perception of pain. These pathways transmit stimuli associated with motivational and cognitive aspects (the spinoreticular tract and spinoparabrachial tract), motor responses, and affectivity (the spinomesencephalic tract and spinoparabrachial tract), as well as neuroendocrine and autonomic responses (the spinohypothalamic tract). Remarkably, the spinoparabrachial pathway (SPBP) serves as a significant site of convergence for both somatic and visceral nociceptive stimuli. It is also worth noting that besides nociceptive responses visual and auditory stimuli can generate exteroceptive responses that contribute to pain [24].
These basic circuits may undergo modifications in switching from physiological (acute) to pathological (chronic) pain. Likewise, additional pathways may be recruited in several conditions leading to chronic pain. The polysynaptic dorsal column pathway (DCP) is e.g., involved in the processing of pain following peripheral nerve damage, as a phenotypic switch (see Section 3.1 – Peripheral sensitization) occurs in DRG neurons projecting to the gracile and cuneate nucleus following nerve injury [25–27].
1.3.2. Visceral Pain Pathways – Subcortical Structures
Visceral pain originates from the internal organs, such as the heart, blood vessels, airways, gastrointestinal tract, and urinary and reproductive organs [28]. Nevertheless, it should be noted that visceral discomfort does not arise from all internal organs, nor is it necessarily associated with demonstrable physical harm to these organs. The challenging task of identifying and localizing visceral pain stems from the comparatively sparse distribution of sensory nerves in visceral organs and the vast dispersion of visceral input within the CNS. Input from the viscera has a broader and less precise distribution, primarily targeting lamina I and the deep dorsal horn. In different individuals, pain originating from various visceral organs can exhibit distinct patterns of presentation, such as discomfort radiating from the bladder to the perineal area or from the heart to the left arm and neck. Consequently, visceral pain frequently exhibits localization to anatomical sites that are distant from its actual source, leading to its designation as referred pain. Thus, symptoms often involve the occurrence of referred pain to somatic tissues located in the same metameric field as the damaged viscera. Furthermore, it is worth noting that secondary hyperalgesia can manifest in the superficial or deep body wall tissues because of viscerosomatic convergence. Visceral discomfort is frequently accompanied by notable motor and autonomic reactions, including excessive perspiration, feelings of nausea, vomiting, gastrointestinal disturbances, as well as alterations in body temperature, blood pressure, and heart rate.
The afferent fibers that primarily innervate the viscera transmit signals to the CNS via sympathetic and parasympathetic neurons. The sympathetic innervation is provided by visceral afferent fibers that originate from a small subset of DRG neurons and travel along the hypogastric, lumbar, and splanchnic nerves, which traverse both prevertebral and paravertebral ganglia to connect with their target organs. The parasympathetic innervation is made of the afferent fibers originating from the glossopharyngeal (IX), vagus (X), and pelvic nerves, which end in the brainstem and lumbosacral cord, respectively. While it is true that vagal afferents do not directly transmit pain signals, multiple investigations have shown evidence that the stimulation of the vagus nerve can reduce both somatic and visceral pain. The glossopharyngeal and vagal afferents arise from the PSNs in the PG and NG, respectively, which then transmit signals to the second-order neurons of the nucleus tractus solitarius (NTS) of the medulla oblongata. Sympathetic visceral afferents that converge in the spinal cord synapse with second-order neurons located in the dorsal horn. These second-order neurons then transmit signals to higher centers via the DCP, the SPBP, and a component of the STP projecting to specific midline thalamic nuclei. The projections originating from the superficial dorsal horn, primarily comprising the SPBP, are linked to autonomic and emotional reactions to painful stimuli. In addition to projections from vagal afferents, spinoparabrachial projections are conveyed to higher relay centers associated with limbic and cognitive functions, including brain regions implicated in affectivity, such as the amygdala, hypothalamus, and periaqueductal grey (PAG). The visceral spinothalamic projections originate from the deep dorsal horn traverse to the contralateral side and reach the ventroposterior medial nucleus (VPMN) and VPLN of the thalamus. The medial thalamic nuclei ultimately transmit signals to the cortical regions associated with visceral pain. It is of relevance that also the DCP plays a part in visceral nociception and that the gracile nucleus cells react to painful stimulation of the viscera. This pathway originates from cells in lamina III of the dorsal horn and is located slightly lateral to lamina X [26].
1.3.3. Cortical Structures
Somatic pain stimuli that reach the thalamus relay nuclei (VPLN and ventroposterior inferior nucleus) are primarily conveyed to S1. However, other cortical areas are involved in the processing of these stimuli including S2. A ventrally oriented cortico-limbic somatosensory circuit connects S1 and S2, integrating somatosensory input with learning and memory as well as other sensory modalities such as vision and audition. S1 and S2 also connect with the posterior parietal cortical areas and the insular cortex. The insular cortex is, in turn, linked to the amygdala, perirhinal cortex, and hippocampus.
Visceral pain stimuli that are conveyed to the midline thalamic nuclei are subsequently transferred to limbic cortical areas such as the anterior cingulate (ACC) and insular cortex (INS). Individual neurons often project in more than one of these pathways. Significantly, the ascending spinal connections directly target the limbic and subcortical areas where this system ultimately converges. This twofold convergence could be associated with a process wherein several different brain sources mediate the impact of pain.
The provided diagram (Figure 2) is presented for reference.
2. Role of BDNF in Nociception and Pain: Insights and Mechanisms
Under basal conditions, BDNF is synthesized by various types of neurons and glia within pain pathways. Noxious stimuli can trigger the production and release of BDNF by these cells and upregulate BDNF synthesis and release. Thus, BDNF is involved in the induction of a form of synaptic plasticity, leading to an increase in the responsiveness of peripheral nociceptors to nociceptive stimuli. This sensitization process can take place at the level of peripheral nerve endings (peripheral sensitization) or central neurons (central sensitization) [29,30] – see Section 3. BDNF and neuronal sensitization. Inflammatory processes are often accompanied by an increased release of BDNF as part of a complex network of signaling cascades. Inflammation-induced BDNF release can have both neuroprotective and detrimental effects, depending on the context and duration of the inflammation [31]. Similarly, lesions of the peripheral nerves leading to neuropathic pain may be accompanied by an upregulation of BDNF and the activation of the BDNF/trkB signaling cascade [32].
In the following, I will review the state-of-art information on the localization of BDNF in cells of the pain pathways.
2.1. BDNF in Pain Pathways Neurons
The localization of the BDNF protein in neurons of the somatic and visceral pathways is summarized in Table 1 and Table 2, respectively. It is also shown in graphical form in Figure 1. One very important issue to be considered when dealing with BDNF localization studies is that BDNF, differently from other neurotrophic factors, can be anterogradely transported to targets [33,34] this being fundamental to explaining its messenger role in the modulation of synaptic activity [22]. From a different perspective, the anterograde transport of BDNF explains why in certain areas of the CNS it has been possible to detect the BDNF mRNA in the partial or total absence of protein immunostaining. In addition to BDNF, it is worth noting that pro-BDNF can also be anterogradely transported [35]. Central neurons in the cerebral cortex, parabrachial nucleus, hippocampus, and locus coeruleus have been e.g., discovered to synthesize and transport BDNF anterogradely [33,36,37]. Regarding peripheral neurons, peptidergic small- to medium-sized dark neurons in DRGs produce and transport BDNF to their central terminals in the dorsal horn of the spinal cord [38,39].
The overall frame should be completed considering several intricate regulatory processes contributing to BDNF functional flexibility. These processes comprise controlling the movement of different BDNF transcripts and managing the modification and release of BDNF after protein synthesis. Although they are very important for understanding the context-dependent mechanisms of BDNF signaling [40] they will be mentioned only if directly relevant to the topic of this paper.
2.1.1. Primary Sensory Neurons
As reported in Table 1 and Table 2 and shown in Figure 1, PSNs located in different ganglia of the PNS express BDNF under basal conditions and/or under circumstances of altered pain perception. One of the first studies on the localization of BDNF in PSNs reported that about 25% of small to medium-sized neurons in the rat lumbar DRGs are immunoreactive for the neurotrophin [41]. Subsequently, the same authors have localized BDNF and its mRNA in certain cranial ganglia containing special PSNs (vestibulocochlear and geniculate ganglion that are not relevant to the present discussion), the NG, and the TG [42]. The neurotrophin was then localized to the jugular ganglion (JG) [43] and subsequent studies (reported in Table 1 and Table 2) were confirmative of these observations. Thus, physiologically, BDNF is expressed by small-to-medium-sized PSNs of the DRGs, and cranial ganglia containing general somatic or visceral PSNs except for the superior ganglion of the glossopharyngeal nerve, for which data are not available. The anatomy, function, and neuronal diversity of DRGs have long since been extensively investigated [44,45] and more recently somatosensory neuron types have been defined with single-cell transcriptomics [46]. The neuronal diversification of cranial sensory ganglia is known in much less detail and has been recently reviewed [47]. The TG houses the PSNs of cranial nerve V, which are accountable for sensing and relaying general somatic stimuli from the skin and deep tissues of the head, as well as visceral stimuli from intracranial blood vessels. As a result, the TG is seen as reasonably comparable to the DRGs whose PSNs convey the same types of stimuli from the remaining parts of the body.
DRGs consist of proprioceptive, mechanoceptive, and nociceptive neurons that transmit distinct characteristics of somatosensation. A major difference in the PSN composition between the TG and DRGs is that the former lacks proprioceptive neurons that are instead located within the CNS in the trigeminal mesencephalic nucleus and provide proprioceptive fibers to the masticatory muscles [48] leaving somewhat of an enigma regarding the facial muscles proprioception [49]. DRG proprioceptors and mechanoreceptors are characterized by their large soma and medium to large diameter myelinated axons, specifically Aα and Aβ fibers, respectively, in terms of their morphology. Nevertheless, a certain group of neurons that are responsible for transmitting light touch sensations and expressing emotional components of mild touch, known as C- low-threshold mechanoreceptors (C-LTMRs), gives rise to unmyelinated axons. These neurons also regulate the transmission of nociceptive stimuli [45,50]. They appear to originate from precursor cells differentiating into nociceptors, which are the third type of PSNs in DRGs. The nociceptive lineage consists of thermoreceptors that detect harmless changes in temperature, pruriceptors that detect itch, and nociceptors that detect harmful thermal, mechanical, and chemical stimuli [51]. Most nociceptive neurons are polymodal, meaning they may respond to a wide range of stimuli. Nociceptors constitute the primary neuronal subtype in both the TG and DRGs. Neurons belonging to the nociceptive lineage are present in the JG, NG, and PG as well [52]. Nociceptive neurons, in contrast to proprioceptors and mechanoreceptors, are characterized by their small size and possess axon fibers that are either mildly myelinated (Aδ) or unmyelinated (C) [53,54]. The nociceptors can be further classified into peptidergic and non-peptidergic types. The former class provides sensory innervation to the skin, as well as deep tissues including bones and viscera. On the other hand, the latter specifically innervates the epidermis [51]. Peptidergic nociceptors release neuropeptides such as substance P, somatostatin, or calcitonin gene-related peptide (CGRP) [55,56]. Under basal conditions, expression of BDNF has been reported in peptidergic nociceptors containing substance P and CGRP [57–60] and expressing the transient receptor potential vanilloid receptor 1 (TRPV1) [56,61]. Even, under basal conditions, there is another subpopulation of DRG neurons expressing BDNF that is not peptidergic and remains to be characterized in full [60]. These neurons are larger and likely immunoreactive to the RT97 antibody that recognizes the phosphorylated form of neurofilament protein NF200 [53]. Alterations in BDNF immunoreactivity in the L4 and L5 rat DRGs were observed in experimental models of neuropathic pain [62]. Chronic constriction injury (CCI) damage to the sciatic nerve caused a substantial ipsilateral rise in the proportion of small, medium, and large BDNF-immunoreactive neurons. After spinal nerve ligation (SNL), there was instead a considerable rise in the proportion of BDNF-immunoreactive large neurons and a notable drop in the proportion of small BDNF-immunoreactive neurons in the L5 DRG on the same side. Conversely, there was a marked increase in the proportion of BDNF-immunoreactive neurons of all sizes in the L4 DRG. Injection of complete Freund adjuvant (CFA) into the hind paw of rats caused increased sensitivity to mechanical pain and significant elevations in levels of tumor necrosis α (TNFα) in the inflamed tissues accompanied by an increase in BDNF, trkB receptor, CGRP, and TRPV1 in the DRGs [63]. Although data on the distribution of BDNF immunoreactivity according to neuronal size were not provided by the authors, an inspection of their images suggests that the increase in BDNF signal occurred in all classes of DGR neurons (small, medium, and large). In another paper in which CFA was used to induce peripheral inflammation, BDNF immunoreactivity increased ipsilaterally in L5 DRG neurons of different sizes [64]. In a different model of pathologic pain (bone cancer), up-regulation of BDNF was instead localized to small but not medium or large neurons [65].
Like those in DRGs, PSNs in TG can be classified based on histological, neurochemical, and functional features [66]. As in DRGs, the peptides CGRP and substance P are commonly colocalized in small-size TG nociceptors. BDNF immunoreactivity was detected in a large subpopulation of TG neurons. Positive neurons were mostly small- or medium-sized and substantial fractions of these neurons were also stained for CGRP and TRPV1 [67,68].
The superior ganglia of cranial nerves IX and X (JG) contain specialized somatic sensory neurons (SSN) that primarily supply nerve fibers to the external auditory meatus, the dura mater, and the pharyngeal region. Although the neuronal diversity of these ganglia remains uncharacterized, single-cell RNA sequencing investigations have demonstrated a significant resemblance between JG neurons and DRG neurons [52,69,70]. In the JG, about 50% of neurons were BDNF-immunoreactive, and large percentages of these neurons were also immunoreactive for CGRP or TRPV1 [43].
As mentioned previously, the PG and NG are the distal ganglia of cranial nerves IX and X, respectively. The PG harbors the visceral sensory neurons responsible for innervating some respiratory system organs and transmitting taste information from the posterior portion of the tongue. The NG neurons provide sensory innervation to the pharynx area, thoracic organs, and a large portion of the digestive tract. They transmit information regarding stretch, pressure, and the chemical environment, including inflammatory mediators. The PG and NG PSNs connect centrally with the NTS (Figure 1) neurons in the brainstem, forming the afferent component of reflex circuits responsible for regulating vital functions such as heart rhythm and peristalsis. Recent research unveiled a significant and unanticipated variety of neurons within the NG [71]. There have been numerous subtypes identified, which are responsible for controlling the respiratory, gastrointestinal, and cardiovascular systems. These subtypes include mechanoreceptors that sense stretch and volume, baroreceptors, and receptors that respond to chemicals and nutrients [52,70]. BDNF immunoreactive neurons were detected in both PG and NG and were, respectively, about 50% (PG) or 80% (NG) of the total neurons in these ganglia [43]. BDNF immunoreactivity coexisted with CGRP or TRPV1 immunostaining but there were significant differences between the percentages of double-staining neurons in the two ganglia [43]. In the PG, BDNF-positive neurons were specifically identified as chemoafferent neurons [72].
2.1.2. Second-Order Neurons
Second-order sensory neurons in somatic and visceral sensory pathways receive information from PSNs and relay it to other neurons along these pathways (Figure 1). Somatic (SNTN) and visceral (NTS) second-order sensory neurons express BDNF in a study of the distribution of the protein and its mRNA in the normal adult rat CNS [36]. In this study, the SNTN contained a few lightly stained scattered cells, and the NTS only rare moderately stained cells. Remarkably, in both locations, there was also a moderate (SNTN) or heavy (NTS) presence of BDNF-immunoreactive fibers. In the NTS, BDNF was demonstrated to tonically regulate blood pressure, heart rate, and sympathetic nervous system activity in vivo and modulate glutamatergic excitatory synaptic transmission at postsynaptic sites to inhibit excitatory transmission at glutamatergic primary afferent synapses [73].
2.1.3. Higher-Order Neurons
The localization of BDNF in higher-order neurons along pain-related pathways is not fully characterized. In their seminal paper, Conner and colleagues published a semiquantitative detailed map of the distribution of the BDNF protein, its mRNA, and its trkB receptors in the CNS of normal adult rats [36]. The following description is mainly based on their data.
- Cerebral cortex
BDNF-immunoreactive cells have been localized in several cortical areas related to pain perception (see Figure 1). The insular and cingulate cortices were found to contain low-to-moderate numbers of immunoreactive cells, as well as moderate levels of expression of the BDNF mRNA. There were no BDNF-immunoreactive fibers in cortical areas.
Expression of BDNF was upregulated in certain conditions of pathologic pain in cortical neurons. After CFA injection, BDNF levels in neurons (NeuN+) and glia (see below) were markedly increased in ACC and S1 [74]. In the same study administration of recombinant BDNF by injections into the ACC, or injecting a viral vector engineered to synthesize BDNF into either the ACC or S1, resulted in increased neuronal excitability, as seen by increased long-term potentiation (LTP), and persistent sensitivity to pain. Cyclotraxin-B, a powerful antagonist specifically targeting the BDNF receptor trkB signaling (see Section 4. Implications for pain management), effectively halted neuronal hyperexcitability, the development of cold hypersensitivity, and passive avoidance behavior in the ACC. These findings indicate that the development and persistence of the emotional component of chronic pain are strongly influenced by BDNF-dependent neuronal plasticity in this cortical area. In another study, the BDNF/trkB-mediated signaling pathway in the rostral ACC was shown to be involved in the development of neuropathic spontaneous pain-related aversion upon activation of the N-methyl-d-aspartate (NMDA) NR2B receptors [75]. It has also very recently been reported that neonatal pain induced by intraplantar CFA injection or repetitive needle prick stimuli resulted in impaired hearing in adulthood associated with a decrease of BDNF immunoreactivity in neurons (and glia – see below) of the primary auditory cortex (Au1) in parallel with increased dendritic spine density, whilst no damage or synapse loss was found in the cochlea [76]. Oxycodone, a semi-synthetic opioid, attenuated hearing loss and alterations in Au1, and a trkB agonist reversed neonatal pain-induced hearing impairment and decreased caspase 3 expression. On the other hand, administration of a trkB antagonist in naïve mouse pups damaged hearing development and increased caspase 3 expression. Therefore, these observations altogether confirm the modulatory action of BDNF in central auditory pathways.
Finally, a study in rats with peripheral inflammation demonstrated that continuous and spontaneous pain interferes with the connections between the ventral hippocampal CA1 region and the infralimbic cortex, as well as the ability of the hippocampus to regulate the neuronal activity in the latter [77]. Targeted upregulation of BDNF in this circuit counteracts electrophysiological alterations, alleviates spontaneous pain, and expedites total recuperation from inflammatory pain.
- Amygdala
The complex of the amygdaloid nuclei displayed a variegate pattern of BDNF immunostaining. Remarkably, the central nucleus of the amygdala was devoid of the BDNF protein and mRNA but contained a very dense network of heavily stained BDNF-immunoreactive fibers originating from the parabrachial nucleus (PBN) [36]. The basolateral nucleus contained occasional or moderate numbers of positive cells. BDNF immunoreactivity was shown to increase in neurons of the amygdala after stress-induced visceral hyperalgesia [78].
- Brainstem
Within the brainstem, besides the aforementioned nuclei containing immunoreactive SSNs, the PBN was reported to contain no BDNF-immunoreactive neurons but to be particularly enriched in positive fibers [36]. In this location, there was a moderate number of cells expressing the BDNF mRNA. The lateral PBN sends a strong contingent of BDNF immunoreactive fiber to the central nucleus of the amygdala and has recently been demonstrated to be directly involved in the control of neuropathic pain behaviors [79]. This conclusion was reached after the observation that activation of glutamatergic or inhibition of GABAergic neurons in the lateral PBN by optogenetics induced neuropathic pain-like behavior in mice and that inhibition of glutamatergic neurons diminished both basal nociception and neuropathic pain behaviors. In another study, the localized deletion of BDNF in the PBN showed that the basal thresholds of thermal and mechanical nociceptive responses were not altered, but animals with a deletion of the BDNF gene exhibited a reduction in the pain-relieving properties of morphine. [80]
2.2. Neuronal Mechanisms
The investigations on the neuronal mechanisms by which BDNF could regulate nociception and pain have concentrated on spinal and (to a lesser extent) trigeminal nociceptive pathways at the synapses between PSNs and SSNs. Many of these studies have been carried out in vivo, but ex vivo studies were also used to better explore the intervention of BDNF as a pain modulator. However, much less information is available regarding higher-order neurons along the pain transmission pathways described in Section 1.3. Overview of the anatomical arrangement of pain pathways. Broadly speaking, BDNF plays a pivotal role in synaptic plasticity within the spinal cord and the SNTN, where nociceptive signals are first processed. At PSN/SSN synapses (Figure 2), BDNF enhances glutamate release and modifies the function of α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA) and NMDA receptors, resulting in amplified nociceptive signals [83,84].
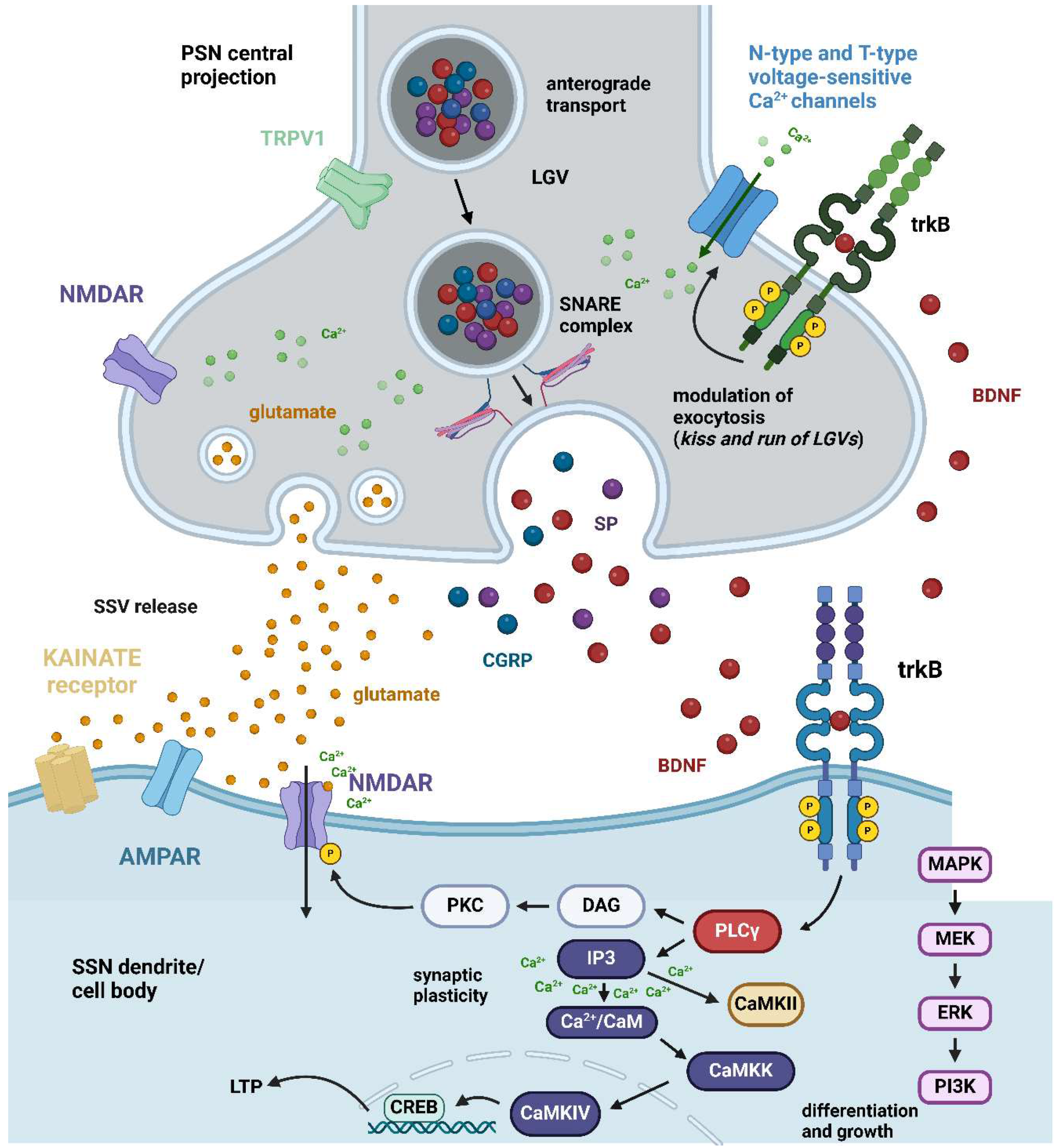
Figure 3.
BDNF pre- and postsynaptic neuronal mechanism. The interface between the central projection of a PSN and an SSN dendrite/cell body is shown with special reference to the spinal cord dorsal horn. In C-type peptidergic primary afferent fibers originating from PSNs, there is a coexistence of glutamate as the main fast-acting neurotransmitter and some high molecular weight slow-acting neurotransmitters of peptide nature. The former is stored in SSV whereas peptides colocalize in LGVs. Specifically, the BDNF peptide has been shown to colocalize with SP and CGRP, two of the most widely investigated sensory neuropeptides [15]. Along the central projections of these fibers, the mature BDNF peptide is anterogradely transported to PSN terminals. A subgroup of these terminals in the spinal cord dorsal horn expresses presynaptic (green) trkB receptors, but another subgroup does not express trkB [56]. For simplicity, an axo-dendritic contact is shown expressing trkB receptors at both pre- and postsynaptic sites. Under basal conditions and nociceptive pain, besides glutamate C-type fibers originating from PSNs also release BDNF. The effects of BDNF in this case are due to the activation of postsynaptic trkB receptors (cobalt). There are three main pathways activated by these receptors within the cell: the MAPK/ERK cascade, which is mainly responsible for differentiation and growth but also intervenes in certain forms of pathological pain (see main text), the PLCγ/DAG/PKC pathway that leads to phosphorylation of postsynaptic NMDA receptors leading increased NMDA Ca2+ currents, and the PLCγ/IP3 cascade that leads to a release of Ca2+ from internal stores and activation of CAMKs, eventually leading to synaptic plasticity and long-lasting LTP (see main text). When excitatory neurotransmission is reinforced, BDNF can also act on presynaptic trkB receptors leading to an increase of Ca2+ in the presynaptic terminal across N-type and T-type voltage-sensitive Ca2+ channels [85] leading to augmented vesicular release of glutamate, as well exocytosis of LGVs. Abbreviations: AMPAR = α-amino-3-hydroxy-5-methyl-4-isoxazole propionic acid receptor; BDNF = brain-derived neurotrophic factor; Ca2+/CaM = calcium/calmodulin; CaMKII = calcium/calmodulin-stimulated protein kinase II; CaMKIV = calcium/calmodulin-stimulated protein kinase IV; CGRP = calcitonin gene-related peptide; CREB = cAMP response element-binding protein; DAG = diacylglycerol; ERK = extracellular signal-regulated kinase; IP3 = inositol 1,4,5-trisphosphate; LGV = large granular vesicles; LTP = long-term potentiation; MAPK = mitogen-activated protein kinase; MEK = mitogen-activated protein kinase kinase; NMDAR = N-methyl-D-aspartate receptor; PI3K = phosphatidylinositol 3-kinase; PKC = protein kinase C; PLCγ = phospholipase C γ; PSN = primary sensory neuron; SNARE = soluble N-ethylmaleimide-sensitive-factor attachment protein receptor; SP = substance P; SSN = secondary sensory neuron; trkB = trkB = tropomyosin receptor kinase B; TRPV1 = transient receptor potential vanilloid. Created with BioRender.com.
Figure 3.
BDNF pre- and postsynaptic neuronal mechanism. The interface between the central projection of a PSN and an SSN dendrite/cell body is shown with special reference to the spinal cord dorsal horn. In C-type peptidergic primary afferent fibers originating from PSNs, there is a coexistence of glutamate as the main fast-acting neurotransmitter and some high molecular weight slow-acting neurotransmitters of peptide nature. The former is stored in SSV whereas peptides colocalize in LGVs. Specifically, the BDNF peptide has been shown to colocalize with SP and CGRP, two of the most widely investigated sensory neuropeptides [15]. Along the central projections of these fibers, the mature BDNF peptide is anterogradely transported to PSN terminals. A subgroup of these terminals in the spinal cord dorsal horn expresses presynaptic (green) trkB receptors, but another subgroup does not express trkB [56]. For simplicity, an axo-dendritic contact is shown expressing trkB receptors at both pre- and postsynaptic sites. Under basal conditions and nociceptive pain, besides glutamate C-type fibers originating from PSNs also release BDNF. The effects of BDNF in this case are due to the activation of postsynaptic trkB receptors (cobalt). There are three main pathways activated by these receptors within the cell: the MAPK/ERK cascade, which is mainly responsible for differentiation and growth but also intervenes in certain forms of pathological pain (see main text), the PLCγ/DAG/PKC pathway that leads to phosphorylation of postsynaptic NMDA receptors leading increased NMDA Ca2+ currents, and the PLCγ/IP3 cascade that leads to a release of Ca2+ from internal stores and activation of CAMKs, eventually leading to synaptic plasticity and long-lasting LTP (see main text). When excitatory neurotransmission is reinforced, BDNF can also act on presynaptic trkB receptors leading to an increase of Ca2+ in the presynaptic terminal across N-type and T-type voltage-sensitive Ca2+ channels [85] leading to augmented vesicular release of glutamate, as well exocytosis of LGVs. Abbreviations: AMPAR = α-amino-3-hydroxy-5-methyl-4-isoxazole propionic acid receptor; BDNF = brain-derived neurotrophic factor; Ca2+/CaM = calcium/calmodulin; CaMKII = calcium/calmodulin-stimulated protein kinase II; CaMKIV = calcium/calmodulin-stimulated protein kinase IV; CGRP = calcitonin gene-related peptide; CREB = cAMP response element-binding protein; DAG = diacylglycerol; ERK = extracellular signal-regulated kinase; IP3 = inositol 1,4,5-trisphosphate; LGV = large granular vesicles; LTP = long-term potentiation; MAPK = mitogen-activated protein kinase; MEK = mitogen-activated protein kinase kinase; NMDAR = N-methyl-D-aspartate receptor; PI3K = phosphatidylinositol 3-kinase; PKC = protein kinase C; PLCγ = phospholipase C γ; PSN = primary sensory neuron; SNARE = soluble N-ethylmaleimide-sensitive-factor attachment protein receptor; SP = substance P; SSN = secondary sensory neuron; trkB = trkB = tropomyosin receptor kinase B; TRPV1 = transient receptor potential vanilloid. Created with BioRender.com.
Remarkably BDNF exerts a neuromodulatory effect under both uninjured (normal) and injured conditions [86]. Multiple lines of evidence suggest that BDNF primarily functions as a pro-nociceptive modulator, playing a key role in the onset and persistence of inflammatory, chronic, and/or neuropathic pain [32]. However, there have been sporadic reports of BDNF having anti-nociceptive effects, indicating that BDNF actions can vary depending on specific cell types, molecular pathways, and pathological circumstances [22,86–89]. Nonetheless, it was also reported that BDNF produced by primary afferents has minimal relevance in the processing of pain and itch [90]. All these issues will be discussed in the following.
2.2.1. Ex Vivo Studies on Nociceptive/Pain Modulation by Mature BDNF
Ex vivo studies allow analyzing the effects of endogenous or pharmacologically administered BDNF on acute slices or slice organotypic cultures [91]. Most of these studies were carried out on spinal cord slices and were focused on the substantia gelatinosa (lamina II of the gray matter). It is worth mentioning here that neurons in the two types of preparations display similar electrophysiological properties [92]. It is also useful to recall that the approach ex vivo allowed the performing of electrophysiological recordings and other functional studies (e.g., Ca2+ imaging) that have been useful in ameliorating our understanding of the circuitry involved in BDNF modulatory functions. I will in this section describe only the results related to slices/organotypic cultures prepared from normal untreated animals. When these preparations were obtained from animals previously treated in vivo they will be discussed in the following sections.
Acute slices of the rat spinal substantia gelatinosa have been used to dissect the role of BDNF as a pain modulator focusing on the synapses between PSNs and SSNs [59]. In this study, two series of experiments were performed. In the first, BDNF was applied to naïve slices at pharmacological concentrations whereas in the second inflammation was mimicked after capsaicin administration to slices. After patch-clamp recordings, calcium imaging, and immunocytochemistry on slices from 8-12 days post-natal rats it was thus demonstrated that BDNF enhanced the release of sensory neurotransmitters (glutamate, substance P, CGRP) in lamina II by acting on pre-synaptic trkB receptors expressed by TRPV1 immunoreactive peptidergic primary afferent fibers. It is worth noting that Ib1-immunoreactive microglial cells did not stain for BDNF in this study, because the BDNF released by microglia has been implicated in the generation of neuropathic pain in other reports in vivo (see Section 2.3.1 Glial cells – Microglia). BDNF also enhances the excitatory responses to NMDA in the rat spinal cord [93]. The underlying mechanism may include enhancing the activity of presynaptic NMDA receptors on primary afferent terminals, leading to an increase in the transmission of excitatory glutamatergic signals [94].
It was also shown that exposing substantia gelatinosa neurons to BDNF in organotypic culture for a period of 5 to 6 days results in an electrical pattern that closely resembles the one induced by CCI in live subjects [92,95]. Both BDNF and CCI enhance the amplitude and frequency of spontaneous excitatory postsynaptic currents (sEPSC) and miniature excitatory postsynaptic currents (mEPSC) in neurons that exhibit delayed firing. Furthermore, a more comprehensive examination reveals that both interventions also disclose the existence of a previously unidentified group of large mEPSC events, which are not present in neurons from rats that underwent sham surgery or in control organotypic cultures [96,97].
As will be discussed in the following sections, peripheral nerve damage stimulates the secretion of BDNF from primary afferent terminals and spinal microglia leading to increased excitability of the dorsal horn, which plays a role in central sensitization and the initiation of neuropathic pain (see Section 3.2 Central sensitization). While it is widely acknowledged that the reduction of GABAergic and/or glycinergic inhibition is responsible for the onset of neuropathic pain [98], some research indicates that BDNF administration increases the K+-induced release of GABA in the dorsal horn [99]. BDNF also increases spontaneous GABA release [as shown by increases in miniature inhibitory postsynaptic current (mIPSC) frequency] yet reduces stimulation-evoked GABA release (i.e., reduces the amplitude of evoked IPSCs) very likely regulating the release of the inhibitory transmitter from the islet cells [100].To shed more light on these conflicting results, rat spinal cord neurons were challenged with BDNF for six days in a defined-medium organotypic culture to replicate the alterations in spinal BDNF levels that occur with peripheral nerve injury [101]. Very interestingly it was demonstrated that BDNF enhanced the amplitude of GABAergic and glycinergic mIPSCs in both inhibitory tonic-islet-central neurons and excitatory delay-radial neurons (for classification and properties of substantia gelatinosa neurons see [56]). Furthermore, the neurotrophin enhanced the amplitude and frequency of sIPSCs in presumed excitatory neurons, whereas it decreased the amplitude of sIPSCs in inhibitory neurons, although the frequency remained unchanged. From these results, it was concluded that this dual effect of BDNF on GABAergic transmission (the increase in inhibitory input to excitatory neurons and the decrease in inhibitory input to inhibitory neurons) only apparently contradicts the general notion that BDNF enhances the overall excitability of the dorsal horn because a possible explanation is that the effect of BDNF in the substantia gelatinosa is primarily influenced by the enhancements in excitatory synaptic transmission, rather than by the inhibition of inhibitory transmission [101]. Along this line of thinking, some time ago I proposed a neuronal circuitry to explain the apparent dual effect of BDNF in lamina II whereby inhibition of inhibitory transmission occurs under conditions of long-lasting pain and central sensitization [56].
In a 2022 paper, both rat and human acute spinal cord slices were treated with recombinant BDNF to investigate the possible occurrence of sexual dimorphism in neuronal hyperexcitability under pathological pain conditions [102]. The administration of ex vivo BDNF did not have any impact on synaptic NMDA receptor responses in female lamina I neurons of rats. Crucially, this disparity in the way spinal pain is processed was maintained across species, from rats to humans. Similar to rats, the application of BDNF to spinal cord slices obtained from human donors resulted in a decrease in markers of reduced inhibition and an increase in markers of enhanced excitation in neurons located in the superficial dorsal horn. This effect was observed in neurons from males but not females. Notably, ovariectomy in female rats replicated the neuronal phenotype of pathological pain observed in males.
2.2.2. In Vivo Studies on Inflammatory Pain Modulation by Mature BDNF
- DRG/Spinal cord
Numerous in vivo studies have concluded that BDNF is an important pro-nociceptive modulator in several forms of inflammatory pain. The release of BDNF by sensory neurons and their primary afferent axons was demonstrated to be essential for the development of inflammatory pain caused by both formalin and carrageenan [103]. The pro-nociceptive impact of BDNF in inflammatory pain is commonly linked to pre- and post-synaptic enhancement of glutamatergic transmission in the spinal dorsal horn through NMDA receptor plasticity. BDNF enhances nociceptive spinal reflex activity and triggers c-fos expression in dorsal horn neurons in an NMDA receptor-dependent manner in rats treated with carrageenan [93]. Furthermore, it has been discovered that the molecule trkB-IgG, which sequesters BDNF, is capable of alleviating hyperalgesic behavior and restoring normal function [93]. In mice, the combination of BDNF and the NMDA receptor antagonist D-2-amino-5-phosphonovaleric acid (D-APV) stops the development of BDNF-induced hyperalgesia [104]. The enhancement of NMDA neurotransmission is facilitated by the activation of protein kinase C (PKC)- and phospholipase C (PLC)-dependent pathways [83,105].
It was long ago observed that BDNF can cause hyperalgesia by acting presynaptically at the synapses between PSNs and SSNs in the spinal dorsal horn. In a rat model of inflammation with CFA subcutaneous injections, it was demonstrated that BDNF enhances the presynaptic release of glutamate in lamina II neurons [106]. The observed impact was linked to a stronger synaptic input from large myelinated Aβ afferent fibers. These fibers are known to normally encode light touch and may therefore be responsible for the development of allodynia in this model of inflammatory pain. In keeping with these observations, CFA- and TNFα-induced peripheral inflammation was observed to increase the in vivo expression of BDNF, trkB, and TRPV1 in DRGs [63]. In the same study, DRG cultures that were treated with TNFα over a long time exhibited a notable increase in the amounts of mRNA and protein for BDNF and the trkB receptor. Additionally, there was an increase in the release of BDNF and the activation of the phospho-extracellular signal-regulated kinases 1 and 2 (ERK1/2) signal pathway induced by trkB. Furthermore, the production of CGRP and substance P was increased following prolonged exposure to TNFα or immediate activation with BDNF [63]. Altogether these observations are fully confirmatory of our data in spinal cord slices subjected to capsaicin challenge to mimic inflammation ex vivo (see above).
There may be several pathways driving neuronal BDNF signaling in response to inflammatory conditions. In one study ATP-gated purinergic receptor 4 (P2RX4) expression was increased in nociceptive neurons during prolonged peripheral inflammation and co-localized with BDNF in these neurons [107]. P2RX4-deficient mice exhibit defective BDNF-dependent signaling pathways, including phosphorylation of ERK 1/2 and the GluN1 subunit of the NMDA receptor, as well as downregulation of the K-Cl co-transporter 2 (KCC2) in the dorsal horn of the spinal cord. It is worth noting that to ascertain if sensory neurons were the primary cells responsible for producing BDNF in the dorsal horn during prolonged inflammation, the authors examined if the paw injection of CFA induced microglial activation and if BDNF was expressed by spinal microglia to conclude that no apparent alterations in the structure or quantity of Iba1+ microglia were found in the dorsal horn when comparing the control and CFA conditions. These findings indicate that P2RX4, which is expressed by PSNs, regulates the release of neuronal BDNF, hence contributing to increased neuronal excitability in the context of chronic inflammatory pain [107].
Recently, the role of peripheral BDNF in inflammatory pain has been reevaluated using a conditional Advilin-CreERT2 knock-out mice model. This mouse selectively lacks BDNF in sensory neurons [32,90]. Both investigations using this model noted a decrease in the nocifensive response during the second phase of the formalin test, indicating a decrease in central sensitization. However, this effect was only detected in males [90]. Interestingly, these authors did not observe any disparity between sexes in the progression of pain hypersensitivity in the CFA model of chronic inflammatory pain [90]. However, another group discovered that the deletion of BDNF has a significant impact on hyperalgesic priming [32]. This is a neuroplastic mechanism that creates a hidden state of reinforced sensitivity in sensory neurons, which is believed to facilitate the shift from acute to chronic pain (see Section 3. BDNF and neuronal sensitization). Induction of hyperalgesic priming was achieved with the intraplantar injection of carrageenan, and the subsequent effect was revealed by the injection of prostaglandin E2 (PGE2) after six days. While control mice that were prepared in advance demonstrated long-lasting reinforced sensitivity to pain following the injection of PGE2, animals devoid of BDNF in their sensory neurons only showed a temporary increase in their ability to sense pain [32].
Another recent study has confirmed the intervention of BDNF in inflammatory (and neuropathic – see below) after assessment of the effects of 1-(1, 1-dimethyl ethyl)-3-(1-naphthalenyl methyl)-1H-pyrazolo [3,4-d] pyrimidine-4-amine (1NMP), a cell-permeable kinase inhibitor blocking trkB receptor autophosphorylation and signaling [108]. Inflammatory pain was induced by formalin in transgenic TrkBF616 mice expressing a mutated trkB that displayed sustained signaling in the absence of the inhibitor and was strongly reduced after 1-NMP treatment.
- Trigeminal system
Similar mechanisms to those described in the DRGs and spinal cord have been identified in the TG neurons and SNTN receiving the peripheral stimuli from the orofacial territories and the dura mater (see Section 2.1 Overview of the anatomical arrangement of pain pathways). The intervention of BDNF (and other neurotrophins) in modulating the trigeminal pain pathways has been very recently reviewed and the authors highlighted that research on the head sensory territories has been minor, likely due to a relative lack of animal models in comparison to those available for spinal pain [109]. Yet one has to keep in mind that the laminar structure of the SNTN is very similar to that of the spinal dorsal horn considering that both derive from the alar plate of the embryonic neural tube developing into the dorsal gray column. The alar plate of the brainstem is located lateral to the sulcus limitans in the floor of the rhomboid fossa and gives rise to the sensory receiving nuclei of the adult brainstem among which the sensory nuclei of the trigeminal nerve. Thus it is not surprising that most mechanisms of BDNF modulation of trigeminal nociceptive pathways are similar to those described for the spinal cord dorsal horn, particularly the substantia gelatinosa [56].
The SNTN is physically organized into three subdivisions: pars oralis, pars interporalis, and pars caudalis [110]. The pars oralis refers to the endpoint of incoming connections from the mouth and nose areas. It includes circuits that play a role in brain stem reflexes. The pars interpolaris gets sensory information from the same side of the face and combines with the pars oralis to create the ascending anterior trigeminothalamic tract. The pars caudalis also receives sensory input from the face, forehead, cheek, and jaw, and is responsible for transmitting temperature sensation from these regions.
Hyperalgesia associated with BDNF/trkB signaling in TG neurons projecting to the cranial/interpolaris transition zone of the SNTN has been studied after masseter muscle inflammation [111]. BDNF/trkB immunoreactive small/medium-diameter TG neurons were substantially more abundant in inflamed animals compared to naïve rats, and most dissociated small-diameter TG neurons displayed a depolarization response to BDNF that was coupled with spike discharge in whole-cell current-clamp registrations. As expected BDNF-induced TG neuron excitability alterations were eliminated by the pan-trk inhibitor K252a. It is worth noting that in this study the neurons responsible for trigeminal inflammatory hyperalgesia expressed both BDNF and trkB suggesting the possibility of an autocrine effect of the neurotrophin as described earlier after ex vivo studies on the spinal substantia gelatinosa (see Section 2.2.1 Modulation of inflammatory pain by mature BDNF - Ex vivo studies). Expression of trkB in mouse lingual sensory afferents (i.e., the peripheral branches of the TG neurons giving rise to the lingual nerves) was demonstrated in a paper where oral squamous carcinoma cells were shown to release BDNF contributing to oral cancer pain via peripheral trkB activation [112]. The cellular mechanism by which BDNF caused TG neuron hyperexcitability was examined in culture-isolated neurons. Exogenously administered BDNF was shown to increase T-type Ca2+ currents by stimulating the trkB-phosphoinositide 3-kinase (PI3K)-p38-protein kinase A (PKA) intracellular pathway [113]. Furthermore, an upregulation of the Bdnf gene in mouse TG neurons was observed in a model of mechanical irritation and localized inflammation in the periodontium mimicking the pain derived from orthodontic forces in most patients [114]. Remarkably, intraganglionic injection of resiniferatoxin (a potent functional analog of capsaicin) before the application of an orthodontic force reduces mice’s orthodontic-induced nocifensive behavior indicating that also in TG neurons TRPV1 receptor expression is pivotal for the genesis of inflammatory pain as observed in DRG neurons [115].
Some other studies have analyzed the expression of BDNF in the SNTN neurons following different experimental forms of inflammatory pain. After subcutaneous capsaicin injection in the upper lip, SSNs in SNTN displayed a reduction in BDNF immunoreactivity [116]. In this study, orexin-A lowered capsaicin-induced trigeminal pain through the modulation of pain effects on BDNF and cyclooxygenase-2 (COX2) expression, but how TG neurons activation by capsaicin influenced the expression of BDNF in SNTN neurons was not investigated. In another paper, inflammatory pain was generated by injecting CFA into the rat temporomandibular joint to study the effects of transcranial direct-current stimulation as a therapeutic tool [117]. Joint inflammation raised the level of brainstem BDNF after ELISA immunodetection and transcranial direct-current stimulation reversed such an effect. A third study evaluated the effects of pituitary adenylate cyclase-activating peptide (PACAP) type I receptor (PAC1R) antagonist, PACAP6-38 in nitroglycerin-induced central sensitization, a chronic migraine model, after stereotaxic injection of the organic nitrate into the SNTN [118]. It was thus demonstrated that the effects of nitroglycerine could be reversed by inhibition of PAC1R and down-regulation of the ERK/cAMP response element-binding protein (CREB)/BDNF signaling pathway.
Whereas the studies above converged to indicate the primary role of neuronal BDNF in the genesis of inflammatory trigeminal pain, in a trigeminal allodynia rat model, which closely mimics chronic migraine, P2X4 receptors, and associated signaling pathways were examined to conclude that it was the SNTN microglial cells expressing BDNF to upregulate these receptors [119]. Blocking the purinergic P2X4 receptor inhibited the expression of BDNF and other molecules (p38, excitatory amino acid transporter 3 - EAAT3, c-Fos, and CGRP), causing anti-nociception.
2.2.3. Modulation of Inflammatory Pain by proBDNF In Vivo
Although the intervention of BDNF in inflammatory pain is widely accepted, the role of proBDNF, which is also expressed in the spinal cord, has been comparatively little investigated.
In rodents, proBDNF acts as an inflammatory mediator in pain regulation [120]. While it is well established that BDNF is produced by PSNs and thereafter released in an activity-dependent manner, the mechanisms of proBNF secretion remain to be understood in full [121]. It was nonetheless shown in vitro that the activity-dependent release of BDNF could be regulated by the interaction of sortilin with proBDNF [122]. In vivo, the expression of BDNF and sortilin was dramatically elevated in the ipsilateral L4/5 DRGs following the injection of CFA into the rat hind paw, and they were found to be co-localized [123]. Knocking down sortilin 1 in the L5 DRG using in vivo an adeno-associated virus effectively reduced the pain-like response. A synthetic peptide, corresponding to the sequence of amino acid residues 89-98 of proBDNF, was discovered to disrupt the interaction between proBDNF and sortilin when compared to a randomly arranged peptide. This synthetic peptide also hindered the release of BDNF in vitro in response to activity and decreased the occurrence of CFA-induced mechanical allodynia and heat hyperalgesia in vivo. The synthesized peptide also disrupted the capsaicin-induced activation of ERK in the spinal cord of rats injected with CFA [123].
Recently, the importance of the proBDNF/ p75NTR signaling pathway in inflammatory and rheumatoid arthritis pain was studied in mice and human patients to conclude that proBDNF/p75NTR boosts rheumatoid arthritis and inflammation by triggering proinflammatory cytokines [124]. In this study, it was observed a higher degree of synovitis in the synovial membrane of patients with rheumatoid arthritis compared to the control group with osteoarthritis. The mRNA levels of P75NTR and sortilin, as well as the protein level of proBDNF, were considerably higher in the peripheral blood mononuclear cells of rheumatoid arthritis patients compared to healthy donors. ELISA consistently demonstrated elevated levels of p75NTR, sortilin, TNFα, interleukin-1β (IL-1β), interleukin-6 (IL-6), and interleukin-10 (IL-10) in the serum of patients with rheumatoid arthritis compared to healthy donors. Expression of the proBDNF/p75NTR signaling pathway molecules and elevated levels of inflammatory cytokines were also observed in the spinal cord utilizing a widely accepted collagen-induced arthritis mouse model. It was reported that after CFA-induced inflammation in male rats, there was a rapid increase of proBDNF in NeuN-positive neurons and GFAP-positive astrocytes in the spinal cord ipsilateral to the CFA injection site [125]. In addition, the intrathecal administration of a cleavage-resistant proBDNF protein decreased thermal and mechanical pain thresholds, whereas a monoclonal anti-proBDNF antibody reduced CFA-triggered pain and inflammation. In the same study, the administration of CFA caused the activation of p75NTR and its downstream signaling pathways, ERK1/2 and nuclear factor-kappa B (NF- κB) p65, in the spinal cord, and the intrathecal administration of p75NTR extracellular domain effectively inhibited pain and neuroinflammation generated by CFA leading to the conclusion that up-regulation of proBDNF/p75NTR signaling also contributes to inflammatory pain generation in the spinal cord [125].
2.2.4. In Vivo Studies on Neuropathic Pain Modulation by Mature BDNF
- DRG/Spinal cord
BDNF is also responsible for the maladaptive changes in neural circuits that occur in neuropathic pain. Also, for this type of pain, most observations refer to the synapses between PSNs and SSNs in the dorsal horn of the spinal cord. Research conducted on rats and mice with a partial sciatic nerve injury long ago revealed that spinal BDNF plays a role in the changes that occur in pain pathways because of nerve damage [126,127,128,129]. These studies showed that the administration of either anti-BDNF or anti-trkB neutralizing antibodies, trk inhibitors, or the BDNF scavenger trkB/Fc through the bloodstream eliminates the behavioral responses to the increased pain sensitivity. Several other reports documented that increased expression of BDNF in PSNs and its subsequent release at their central processes in the spinal cord dorsal horn occurred in several models of nerve damage, such as sciatic nerve transection, CCI, and nerve ligation [130,131,132,133,134,135,136]. More recently, expression changes in TRPV1 receptors, CGRP, and BDNF were observed in rat DRGs after resiniferatoxin (a very strong agonist of TRPV1 receptors)-induced neuropathic pain, with reduction of CGRP and TRPV1 and increase of BDNF with the development of pathologic pain [115].
Under neuropathic pain conditions, BDNF induces hyperexcitability in spinal neurons by enhancing the action of NMDA receptors, as observed in inflammatory pain. When rats with spinal nerve ligation are given BDNF through an injection into the spinal cord, it causes LTP and increased sensitivity to touch by activating the NMDA-NR2B receptors [130,137]. Simultaneous changes in inhibitory and excitatory transmission are anticipated to interact and increase the sensitivity of central sensory neurons, leading to the so-called central sensitization of these neurons - See Section 3.2 Central sensitization. Specifically, it was documented that the decrease in KCC2 caused by BDNF is both enough and essential to facilitate NMDA-NR2B-receptor phosphorylation and the initiation of LTP in lamina I neurons of rats with nerve injuries [138]. STEP61 phosphatase plays a crucial role in the communication between BDNF-dependent Cl dysregulation and NMDA phosphorylation. When STEP61 phosphatase is down-regulated, it leads to a decrease in KCC2 levels and ultimately results in the enhancement of neuronal excitation [139] – See also 2.2.1 Modulation of inflammatory pain by mature BDNF - Ex vivo studies.
While the mechanisms mentioned above primarily rely on post-synaptic changes at PSN-SSN synapses, there have also been reports of BDNDF-mediated presynaptic effects. In rats with CCI, the expression of the Cl- transporter NKCC1 in DRGs was enhanced by BDNF leading to increased intracellular Cl- levels with temporary GABAergic presynaptic inhibition, and enhanced nociception [140]. The presynaptic effects are probably influenced by microglial BDNF, which has been demonstrated to enhance the NMDA-mediated responses by acting on afferent terminals [94] – See also Section 2.3.1 Glial cells - Microglia.
A recent study has examined the effects of 1NMP treatment in TrkBF616 mice (see also 2.2.1 Modulation of inflammatory pain by mature BDNF – In vivo studies DRG/Spinal Cord) in an experimental model of neuropathic pain [108]. Mice were subjected to spinal cord injury (SCI) and weekly evaluation of tactile sensitivity using the von Frey test revealed that administering 1NMP immediately after SCI prevented the onset of mechanical hypersensitivity for a duration of up to 4 weeks following injury. In contrast, when therapy commenced 2 weeks after injury, 1NMP only partially reduced the hypersensitivity of the hind paw. These findings indicate that the activation of trkB signaling soon after SCI plays a role in the development of maladaptive changes in neural connections, resulting in pain hypersensitivity [108]. Another proof of the importance of BDNF in neuropathic pain came from a novel study of the microRNA-489-3p regulation of the oncoprotein DEK [141]. DEK, a DNA-modifying nuclear protein, is involved in maintaining the overall structure and function of heterochromatin, as well as regulating processes such as transcription, DNA replication, and DNA repair for gene control. DEK is widely expressed in CNS and, among others, has been proposed to have a role in neuropathic allodynia albeit elusive and fascinatingly complex. SNL, another widely used model of neuropathic pain, was reported to induce a decrease in miR-489-3p expression, while increasing the expression of DEK, which recruited ten-eleven translocation methylcytosine dioxygenase (TET1) to the promoter fragments of the Bdnf gene, thereby enhancing its transcription in the dorsal horn [141]. SNL-induced neuropathic pain was also shown to be alleviated by the type-B monoamine oxidase (MAO-B) inhibitor, KDS2010 through competitively blocking the BDNF/trkB/NMDA-NR2B pathway [142], giving further support to the intervention of this pathway in the genesis of neuropathic pain. This concept was further validated in another investigation, which demonstrated a substantial increase in BDNF and trkB immunoreactivity in the spinal dorsal horn after a burn injury that is responsible for the development of both inflammatory and neuropathic pain [143]. This study found that the intrathecal administration of trkB-Fc, which is a scavenger of endogenous BDNF, decreased mechanical allodynia. Additionally, the administration of resolvin D1 (RvD1), which is a natural lipid modulator produced during the end of acute inflammation and has powerful anti-inflammatory and pro-resolution effects, prevented the increase of BDNF and trkB associated with injury.
- Trigeminal system
A mouse model of orofacial neuropathic pain, the inferior alveolar nerve (IAN) transection that causes whisker pad ectopic allodynia can be used to assess trigeminal pain. IAN transection increases the peripheral synthesis of BDNF, but the regulation of NMDA receptors in peripheral and central sensitization following the procedure shows different mechanisms than those described above that were centered upon the activation of the NMDA-NR2B receptors (See 2.2.3 Modulation of neuropathic pain by mature BDNF). Notably, neither high- nor low-dose NMDA increased TG BDNF after IAN transection [144]. In TG neurons, NR2A deletion significantly blocked IAN transection-induced alterations in BDNF and other molecules. However, NR2B knockout did not affect BDNF. Thus, NR2A and NR2B had opposing effects on BDNF and chemokine (C-C motif) ligand 2 (CCL2) production [144]. In CCI of the infraorbital nerve, another model of neuropathic pain that generates facial mechanical hyperalgesia, BDNF was upregulated in TG neurons, but observations at higher level neurons in the brainstem and cerebral cortex gave puzzling results as BDNF levels increased also in sham-operated animals [145]. Other authors used the CCI of the infraorbital nerve to model neuropathic pain in rats. They employed different doses of naltrexone to reverse facial mechanical allodynia but observed that the opioid antagonist was unable to modulate the levels of BDNF in the brainstem [146].
At the supraspinal level, it is notable that the CREB/BDNF pathway in the ACC regulates neuropathic pain and anxiety/depression-like behaviors in rats as demonstrated after the reversal of pain hypersensitivity by central specific knockdown and peripheral inhibition of CREB [147].
As a final comment on the intervention of BDNF in spinal and trigeminal neuropathic pain, it is important to be well aware that while the role of BDNF is well-explained in mechanistic investigations such as those reported above, findings from human studies display inconsistency, and further research is required to assess the methodological difficulties associated with utilizing serum BDNF as a biomarker in neuropathic pain [148] – See 5. Clinical trials.
2.3. BDNF in Glia and Other Cells of Relevance in Nociceptive/Pain Pathways – Localization and Mechanisms
Undoubtedly, the domain of pain research first focused on neurons. However, the current emphasis has also turned towards investigating the contribution of non-neuronal cells, specifically their interaction with neurons, which arises because of the response of these cells to a modified neuronal activity [149,150]. As I will discuss in the following, several types of glia and immune cells can synthesize BDNF (Figure 4) and thereby influence the activity of neurons in nociceptive/pain pathways.
2.3.1. Glial Cells
Schwann cells [151] in the PNS, microglia [152,153,154], astrocytes [155,156], and oligodendrocytes [157] in the CNS, synthesize and release BDNF. Broadly speaking, glial cells contribute to BDNF production during certain developmental processes and in response to inflammatory signals and injury in the mature nervous system [158,159]. This BDNF release from the glia can influence synaptic plasticity and pain sensitization [160].
The localization of glial cells expressing the BDNF protein in pain pathways is summarized in Table 3.
- Schwann cells
Schwann are the primary glial cells found in the PNS. Myelinating Schwann cells produce the myelin sheath, which provides electrical insulation to axons and enables the rapid conduction of action potentials by a process called saltatory conduction. Non-myelinating Schwann cells encircle unmyelinated fibers to create Remak bundles, in which Schwann cells wrap around several small-diameter axons, separating them from each other [166]. In addition to these structural tasks, Schwann cells interact with neurons, particularly with whole axons, to regulate their physiological functions. Schwan cells produce and release several mediators including BDNF, chiefly in response to nerve injury [167]. A recent in vitro study [161] has demonstrated that with the activation of a signaling cascade by which allopregnanolone synthesized by Schwann cells upregulates the synthesis and release of BDNF in an autocrine fashion, the latter eventually regulates protein kinase type Cε (PKCε) in DRG neurons, via trkB activation in a paracrine manner. This finally leads to the sensitization of primary afferent nociceptors [168]. BDNF secretion by Schwann cells has also been postulated to occur in trigeminal neuralgia with a mechanism involving P2RX4 [169] in a similar fashion to that described in microglia (see below).
- Microglia
Microglia are the resident CNS macrophages. However, the exact contribution of these cells to the immune defense of the CNS is uncertain due to the immunological privilege of the brain and spinal cord [170]. Research on microglia and pain is very extensive and has mainly focused on the spinal cord somatosensory pathways [171].
The first demonstration that spinal microglia produce BDNF to induce neuropathic pain dates back to 2005 [152]. It is accepted that microglia regulates neuropathic pain after peripheral nerve injury primarily as a consequence of C-fiber activation [172], but also with the contribution of A fibers [173]. There are different classes of microglial activators involved in the regulation of spinal nociception [171]. A series of activators are at first released by PSNs. As a result, many microglial signal molecules are upregulated and contribute to supporting neuropathic pain. Mediators released by microglia then control synaptic plasticity and central sensitization. Specifically, among these mediators, BDNF was implicated in the disinhibition of spinal cord circuits and neuropathic pain [150,152]. BDNF is secreted by microglia because of nerve injury that leads to ATP release, activation of microglial P2RX4, and neurotrophin production. Microglial BDNF then binds to trkB receptors in lamina I neurons, resulting in the upregulation of intracellular Cl– levels through the downregulation of neuronal KCC2 in male rats [174]. The accumulation of Cl– in lamina I neurons reduces the effectiveness of GABA/glycine ionotropic transmission, leading to greater excitability and heightened sensitivity to pain because of the enhancement of synaptic GluN2B-NMDA receptor currents through the activation of Fyn kinase. This eventually leads to the firing of action potentials in lamina I neurons, ultimately promoting the development of neuropathic pain [152]. Microglial-derived BDNF increases the excitatory drive to excitatory dorsal horn neurons with an intensification of spontaneous action potential discharge and inhibits that to inhibitory neurons by both presynaptic and postsynaptic mechanisms [175,176,177]. The absence of GABAergic inhibition allows non-nociceptive Aβ fiber-mediated excitatory signals to reach the outer layers of the spinal dorsal horn, this mechanism promoting the development of allodynia [178].
Very recently the activation of microglia through the BDNF-trkB signaling pathway was demonstrated to downregulate neuronal KCC2 also in dynamic allodynia (caused by mild stimulation of the skin and mediated by low-threshold mechanoreceptors) that follows a sparing nerve injury [179]. In keeping with these observations, it was observed that minocycline, a partially synthetic derivative of tetracycline that inhibits the activation and replication of microglia without directly affecting astrocytes or neurons, increased the expression of KCC2 and GABA-A/γ2 proteins but decreased BDNF expression in the lumbar spinal cord after nerve injury [180]. In the trigeminal system, neuropathic pain can be provoked by delivering ATP-stimulated microglia or BDNF intrathecally [144,145]. On the other hand, reducing BDNF in microglia, blocking trkB, or deleting P2RX4 expression [117,118,145] can oppose the onset of pain.
Yet, the identification of BDNF in spinal (and trigeminal) microglia poses a challenge, despite the straightforward detection of BDNF in DRG and TG neurons and primary afferents within the spinal cord and SNTN [56] – See Section 2.1.1. Primary sensory neurons (PSNs). Nonetheless, it was recently demonstrated that induction of spinal LTP in the absence of nerve injury triggered long-lasting chronic pain with an increase of CGRP immunoreactive terminals in the dorsal horn and that behavioral and structural changes were prevented by blocking NMDA receptors, ablating spinal microglia, or conditionally deleting microglial BDNF [181]. In keeping with the observations converging to show the intervention of spinal microglia in the onset of neuropathic pain, a study conducted by Zhao et al. in 2006 found that BDNF, produced from nociceptors, plays a role in regulating acute and inflammatory pain, but not neuropathic pain [103]. It was also demonstrated that sensory neuron-derived BDNF is also an important factor in the progression from acute to chronic neuropathic pain [32]. These issues are discussed also in Section 2.2.1 Modulation of inflammatory pain by mature BDNF.
Emerging research has additionally indicated the existence of sexual dimorphism in the effects of microglia on nociception [182]. Although there is comparable proliferation of microglia in the dorsal horn of both males and females, it is noteworthy that females do not exhibit upregulation of P2RX4 under neuropathic pain conditions. Instead, they employ an alternative mechanism that is independent of microglia to mediate pain hypersensitivity. It is reasonable that pain hypersensitivity in female mice could be modulated by adaptive immune cells, theoretically T cells. In females, changes in sensory processing in the dorsal horn involve the invasion of macrophages and T-lymphocytes [182]. Yet as in males, this leads to attenuation of inhibition following the collapse of the Cl− gradient. In females, the downfall of the Cl− gradient is also provoked by CGRP [183] released from primary afferent terminals [184].
The intervention of microglial BDNF has been postulated not only in neuropathic pain but also in other forms of pain such as e.g., inflammatory and nociplastic pain. Two polarized forms of activated microglia, i.e., the proinflammatory M1 classically activated microglia and the anti-inflammatory M2 alternatively activated microglia are currently described (reviewed in [158]). BDNF was demonstrated to be released by M1 microglia leading to enhanced neuroinflammation in nociplastic pain [158]. BDNF was also detected immunocytochemically in microglia (and astrocytes) in a model of inflammatory cystitis [185], in autoimmune prostatitis [163], during the early stage of diabetic neuropathy [162], and in a mouse model of chronic post-ischemic pain [186].
Release of BDNF from microglia is promoted, among others by colony-stimulating factor 1 (CSF-1) [154,186,187], and/or Wnt3a [188] and requires the activation of P2RX4 by ATP.
- Astrocytes
Astrocytes are five times as numerous as neurons. They perform numerous fundamental activities in a healthy CNS. Reactive astrogliosis, a pathological characteristic of CNS structural lesions, occurs when astrocytes respond to CNS stressors. Astrocytes have been categorized into protoplasmic or fibrous subgroups based on their morphologies and anatomical placements [155]. Regulated increases in intracellular Ca2+ concentration [Ca2+]i indicate astrocyte excitability [189]. A vast body of evidence shows that these controlled increases in astrocyte [Ca2+]i are important for astrocyte-astrocyte and astrocyte-neuron communication [190]. Studies in vitro have shown that astrocytes produce BDNF under normal conditions [191] and after injury [192,193]. Other in vitro studies have shown that the release of the neurotrophin is mediated by trkB.T1 [194], a truncated isoform of the trkB receptor that lacks the intracellular kinase domain of the full-length receptor and is up-regulated in numerous CNS-damaged scenarios [195]. In addition, in vitro evidence was obtained that the inhibition of the inwardly rectifying potassium channel 4.1, an astrocytic channel that regulates neuronal activity, induces BDNF expression in these glial cells [196].
Pathological reactive astrocytes cause inflammation and neuropathies, and, together with microglia and neurons interact in spinal cord dorsal horn pain signaling [197], and there is growing evidence that astrocytic BDNF has an important role in nociception [156]. Specifically, BDNF has a detrimental effect on neuropathic pain, primarily because of the activation of trkB.T1 [198]. Deletion of trkB.T1 reduced spinal cord injury-related abnormalities, including pain [199]. Indeed, trkB.T1 controls astrocytic proliferation, migration, and inflammation, and its deletion reduces astrocytic reactivity [200], supporting the idea that astrocytes depend on trkB isoform balance to regulate their physiological signaling output [195]. It is of note that trkB.T1 deletion reduced the behavioral response to capsaicin, a powerful TRPV1 agonist [198]. Astrocytic TRPV1, in turn, mediates astrocyte activation [201].
Other studies have demonstrated that BDNF is upregulated in cortical astrocytes under different pain conditions. Upregulation of the neurotrophin was observed in the ACC and S1 of rats with inflammatory pain [74]. Injections of recombinant BDNF (into the ACC) or a viral vector synthesizing BDNF (into the ACC or S1) triggered an elevation of neuronal LTP and sustained pain hypersensitivity, which were abolished after blocking the trkB receptors. In a different study, CFA injection induced BDNF expression in astrocytes (and neurons but not microglia) in Au1 [76].
- Oligodendroglia
Oligodendrocytes serve as the myelinating cells in the CNS. They are the result of a cell lineage that must undergo an intricate and perfectly timed process of growth, migration, specialization, and formation of the myelin, the protective covering of nerve fibers. Oligodendrocytes are considered to be highly susceptible cells in the CNS because of their unique metabolism and physiology, as well as the intricate development program they undergo [202]. After spinal cord injury, oligodendrocytes produce BDNF [203]. The contribution of oligodendrocytes to chronic pain is not well understood; nonetheless, some investigations have shown their involvement in pain caused by neurodegenerative, viral, or neuropathic conditions [204]. Nonetheless, it was demonstrated that oligodendrocytes by secreting BDNF regulate the release of neurotransmitters at presynaptic brainstem terminals expressing trkB receptors [157]. Therefore, it may be possible that some links between this type of glia, BDNF, and pain will be discovered in the future.
2.3.2. Immune Cells
Immune cells interact with neurons to modify pain sensitivity and facilitate the shift from acute to chronic pain [205]. T cells, B cells, and macrophages can produce circulating BDNF, especially in cases of inflammation and injury [206]. Strong evidence has accumulated that the immune system has a role in neuropathic pain as a consequence of nerve injury as authoritatively reviewed in the recent past [159]. The mediators causing neuropathic pain have been classified into three main groups, primary secondary, and tertiary [207]. Primary mediators are released by the immune cells after injury and act peripherally on DRG neurons, secondary mediators are released in the spinal cord and functionally modulate microglia and astrocytes, tertiary mediators, among which BDNF, are released by the spinal glia and act on the central terminals of PSNs and the dorsal horn neurons. However, data are accumulating to indicate that the BDNF released at the periphery by certain types of immune cells has a role in chronic (neuropathic) and inflammatory pain. When BDNF was measured in parallel with different subpopulations of immune cells in patients with chronic pain and healthy controls it was observed that serum levels of BDNF were lower in pain-suffering patients that displayed a reduction in the number of CD4+ CD8+ T cells, CD3- CD56bright NK cells and CD20+ CD3- cells [208]. Likewise, proBDNF was shown to be upregulated in inflammatory cells during arthrosynovitis [209]. Similarly, upregulation of BDNF in mast cells was observed following back pain associated with intervertebral disk degeneration [210] and intradiscal inflammatory stimulation induces spinal pain behavior and intervertebral disc degeneration in vivo with an increase of macrophage markers and BDNF (as well as other nociceptive markers) [211]. Furthermore, studies have shown that following various forms of severe sciatic nerve damage, resident macrophages expressing Iba1 protein gather around DRG neurons, thereby exacerbating pain hypersensitivity caused by nerve injury [212]. This study also found that by inhibiting the expression of Iba1 and modifying the characteristics of resident macrophages, specifically shifting them from an M1 to an M2 phenotype, analgesic effects can be achieved in neuropathic pain caused by SNL. This is accompanied by an enhanced release of BDNF, which triggers the regeneration of processes in adult DRG neurons. Another report reinforced the intervention of BDNF produced by peripheral macrophages in the genesis of neuropathic pain [213]. In this paper, after putting contact between the nucleus pulposus of the intervertebral disc and the sciatic nerve several neuropathic behaviors were observed in mice of both sexes in the absence of an activation of the spinal microglia and pain hypersensitivity was barred by genetically disrupting BDNF electively in macrophages.
2.4. BDNF Polymorphisms Related to Pain Signaling
Several studies highlighted the importance of genetic polymorphisms in the control of pain signaling [214,215]. The single nucleotide polymorphism Val66Met (rs6265) in the BDNF gene, which involves replacing valine with methionine, is of relevance to the present discussion since it has the potential to disrupt the control of the BDNF/trkB signaling system (see [216] for a recent review).
The BDNF Val66Met polymorphism leads to the inability of BDNF to mature correctly in secretory vesicles following its synthesis (Figure 1). Studies in hippocampal neurons [217] have shown that Met-BDNF is primarily found in the cell body, but Val-BDNF is also present in distal dendritic processes. Furthermore, whereas Val-BDNF is the predominant form found in the medium from cultured neurons, the levels of Met-BDNF were hardly detectable. This indicates a difference in the release of the two forms of BDNF. It is worth remembering here that the Met allele is associated with a decrease in the sorting of pro-BDNF and reduced secretion of mature BDNF when neurons are depolarized [218], likely because of decreased interaction with sortilin [217]. The aberrant interaction between mutant BDNF and the sortilin or sortilin-related VPS10 domain-containing receptor 2 (SorCS2) may not only hinder the release of mature BDNF but also cause deficiencies in neuronal development and plasticity [122].
In their very insightful study, Tian and colleagues found that individuals who possessed the G allele (Val/Met and Val/Val) were at greater susceptibility to chronic postsurgical pain in comparison to individuals with the A allele of rs6265 [219]. These authors also provided evidence that Bdnf Met/Met mice have reduced mechanic allodynia following a plantar incision compared to the Bdnf Val/Val group. In terms of pain perception, the Met allele may have a dual function, acting as an analgesic under normal circumstances and as a promoter of pain sensitivity under chronic pain. Indeed, the Met allele has been shown to increase the sense of discomfort during chronic pain, compared to Val homozygotes [220]. The Val66Met polymorphism may have a direct effect on gene methylation and the resulting expression and data showed that individuals with chronic fatigue syndrome and fibromyalgia have higher amounts of BDNF in their bloodstream. These elevated levels of BDNF are directly related to the severity of pain sensations experienced by these patients, as compared to healthy individuals [221].
It was also hypothesized that the presence of the BDNF Val66Met variant might combine with early exposure to pain-related stress in preterm children, leading to distinct effects on the regulation of the hypothalamic-pituitary-adrenal axis. In an investigation of these issues, a correlation between reduced BDNF availability (indicated by the presence of the Met allele) and susceptibility to newborn pain and stress was observed in boys, but not in girls [222]. In keeping with these observations, elevated stress response was consistently found in mice with the BDNFMet knock-in mutation, in parallel with notable alterations in BDNF levels and dendritic spine density in the prefrontal cortex and amygdala that are involved in the processing of the emotional component of pain [223].
Several very recent clinical studies have found a correlation between BDNF polymorphism and inflammatory pain. Compared to non-carriers, subjects suffering from osteoarthritis with BDNF polymorphisms were less likely to improve pain after rehabilitation [224] and, at EEG, displayed decreased alpha oscillations in the frontal area, likely reflecting the disruption of resting states to also compensate for the increased injury associated with arthritis [225]. Other investigations have disclosed an association between BDNF polymorphism and fibromyalgia (reviewed in [226]), and a systematic review and meta-analysis study demonstrated that the circulating BDNF levels were significantly higher in patients suffering from fibromyalgia than in controls [227]. Other studies examined the role of BDNF genotype and expression in the development of chronic low back pain. The findings of one of these studies suggest that having one or more minor alleles, which are associated with decreased BDNF activity, may lower the risk of transitioning from acute to chronic low back pain, possibly as a consequence of a decrease in BDNF signaling-dependent neuropathic pain mechanisms [228]. However, other authors subsequently failed to find an association between the Val66Met polymorphism of the BDNF gene with clinical and biopsychosocial characteristics of chronic low back pain [229]. It was also reported that BDNF polymorphism is associated with neuropathic pain in female cancer survivors [230] or peripheral neuropathy in type II diabetic patients [231].
3. BDNF and Neuronal Sensitization
Much of the above-described effects of BDNF may be linked to neuronal sensitization. There are two forms of neuronal sensitization, peripheral and central. After an accident or tissue damage, the lesioned tissue becomes sensitized to facilitate recovery by triggering behaviors that would avoid additional injury, such as guarding. Both the CNS and PNS have a role in sensitizing the nociceptive system, resulting in primary hyperalgesia in the periphery secondary hyperalgesia, and allodynia in the CNS.
The presence and duration of the so-called hyperalgesic priming is another significant matter in pain plasticity. Priming occurs as a result of an initial injury and leads to a notable vulnerability to often insignificant painful stimuli, resulting in a prolonged state of pain in animals that have been primed [232]. Hyperalgesic priming can occur at the level of the nociceptors and relies on the activation of PKCε after local translation, and/or in CNS, i.e., the dorsal horn of the spinal cord (and although little investigated the SNTN) where it depends on an atypical isoform of PKC (aPKC) referred to as PKMζ (reviewed in [232]). Interestingly, BDNF also participates in the regulation and maintenance of CNS hyperalgesic priming [233].
3.1. Peripheral Sensitization
Peripheral sensitization is the sensitization of the nociceptive system in the periphery. It is characterized by increased responsiveness and reduced threshold of nociceptive neurons to stimulation of their receptive fields. The increased responsiveness of the nociceptors is linked to an increase in the expression, sensitivity, and receptiveness of receptors such as the TRP channels TRPV1 and transient receptor potential cation channel, subfamily A, member 1 (TRPA1) [234] that participate in the hypersensitivity of PSNs to mediators such as CGRP, substance P, and BDNF [30]. BDNF, together with other neurotrophins and glial-derived neurotrophic factor (GDNF) is an important modulator of intraepidermal nerve fibers and is involved in the regulation of pain and itch [235]. BDNF enhances the sensitivity of peripheral nociceptors, making them more responsive to painful stimuli. This effect is mediated, among others, by the upregulation of ion channels, such as TRP channels and voltage-gated sodium channels, in sensory neurons [81]. Peripheral sensitization of nociceptors can also depend on the BDNF released by the Schwann cells activating the pronociceptive PKCε [161]. Notably, peripheral inflammation leads to an upregulation of BDNF mRNA and protein in trkA+ PSNs (which normally do not produce BDNF) via an NGF-dependent mechanism [236], the so-called phenotypic switch that occurs in both inflammatory [237] and neuropathic [238] pain. Nociceptor sensitization is believed to be sustained by elevated peripheral levels of BDNF and its anterograde transport to dorsal horn neurons. It was shown that peripheral inflammation and the electrical activity in C fibers resulted in an upregulation of BDNF expression in the DRG neurons [239]. This increase also led to increased quantities of trkB receptors in the dorsal horn [127]. The increased production and occurrence of BDNF in PSNs, as well as its transport along the axons to the terminals of their peripheral branches has also been linked to the regulation of acute and inflammatory pain [103,240]. As mentioned, increased production of BDNF in DRG neurons may be influenced by a higher concentration of NGF [151,241,242] and will result in a greater amount of the factor being present at the periphery. Interestingly, a considerably increased release of BDNF was observed when acute nociceptor stimulation was applied to the inflamed human skin [243]. Given that elevated levels of BDNF necessitate an enhanced buildup of peripheral neurotrophins, these findings indicate an increased retrograde axonal transport of BDNF from the skin to the cell bodies of PSNs.
Another effect of nerve injury is the synthesis of macrophage CSF-1 in primary afferents and its release in the dorsal horn [175]. CSF-1 increases the excitatory drive to excitatory dorsal horn neurons via BDNF activation of presynaptic trk, postsynaptic trkB, and p75NTR. In parallel, CSF-1 reduces the stimulation of inhibitory neurons by excitatory signals through mechanisms that do not comprise BDNF.
3.2. Central Sensitization
BDNF contributes to central sensitization, a process that amplifies pain signals in the CNS [244]. In central sensitization, CNS neurons begin generating action potentials even in the absence of stimuli from nociceptors due to changes within the neurons themselves. Central sensitization can arise through the rearrangement of synapses in the spinal cord and terminal sensory nuclei of the cranial nerves following prolonged nociceptor activation. This can result in the inappropriate transmission of pain signals to other CNS areas. Rearrangement of synapses is commonly referred to as synaptic plasticity and BDNF plays an important role in this process [86]. LTP, perhaps the most investigated form of synaptic plasticity, is a well-known process in the CNS where there is a permanent enhancement in the effectiveness of communication between neurons. LTP at C-fiber synapses in the spinal dorsal horn and SNTN is regarded as a synaptic representation of pathological pain [245]. This is because spinal LTP is specifically triggered by nociceptive but not innocuous stimuli. Additionally, stimulation that induces LTP can cause persistent behavioral indications of pathological pain in both humans and animals. The role of BDNF in spinal plasticity has been authoritatively reviewed [86]. The chemical mechanisms underlying LTP at C-fiber synapses in the spinal cord share similarities with LTP in the hippocampus in several respects. The induction of LTP relies on an increase in Ca2+ in the postsynaptic second-order neuron. This increase is caused by the opening of NMDA channels and voltage-gated calcium channels, as well as the release of Ca2+ from intracellular stores. Early-phase LTP requires the activation of intracellular PKA, PKC, calcium/calmodulin-dependent protein kinase II (CaMKII), PLC, and the production of nitric oxide (NO) [245]. It was demonstrated that inhibition of PLCγ-PKC signaling blocks the enhancement of synaptic responses in lamina II caused by BDNF, specifically in response to C fiber stimulation [83]. Activation of the PLCγ pathway leads to increased intracellular Ca2+ levels, which can happen by either releasing calcium from internal stores or across phosphorylated calcium-permeable glutamate receptors. These findings are consistent with multiple studies that suggest the activation of MAPK/ERK and PLCγ-PKC, along with the increase in intracellular calcium levels, play crucial roles in the development of nociceptive plasticity, inflammatory, and neuropathic pain. LTP of C-fiber responses is enhanced by BDNF and BDNF pretreatment prevents subsequent LTP generated by SNL [246]. The significance of these effects was emphasized by the discovery that antibodies targeting CSF-1 can block spinal LTP, microglial activation, and induction of BDNF [175].
LTP that occurs after more than three hours is reliant on the synthesis of new proteins. The induction of late-phase LTP can be achieved by activating either dopamine D1 receptors or PKA, as well as by administering exogenous BDNF or ATP directly. Consequently, drugs that target these molecules may negatively affect the memory function of the hippocampus. The notable distinction between hippocampal LTP and spinal LTP at C-fiber synapses lies in the fact that the activation of glial cells and the excessive production of proinflammatory cytokines, such as TNF-α and IL-1β, impede LTP in the hippocampus, whereas they facilitate LTP in the spinal dorsal horn [245].
LTP at C-fibers causes hyperalgesia and is activity-dependent (reviewed in [247]). It can be induced by high-frequency electrical stimulation or conditioning low-frequency stimulation but also by many of the conditions described in the previous sections to model inflammatory or neuropathic pain, i.e., noxious thermal, chemical (formalin, capsaicin) or mechanical (pinching) stimulation of the skin. Induction of LTP varies among afferent fibers innervating different organs or tissues. LTP is more difficult to induce in nociceptive skin afferents than muscle afferents [248]. This difference has been correlated to the relative lack of BDNF in cutaneous afferents, giving further strength to the importance of the molecule in central sensitization. As a result of the binding of BDNF to the trkB receptor, activation of the PI3K-Akt, PLCγ, and MAPK/ERK pathways, and phosphorylation of the NMDA-R1 subunit occurs in target neurons [249].
It is of relevance that BDNF was demonstrated to play a crucial role in protein synthesis-dependent long-term LTP in the spinal dorsal horn via activation of ERK, p38 MAPK, and NF-kB signal pathways [250]. The NF-kB pathway is also implicated in upregulating BDNF after activation of the proteinase-activated receptor 2 (PAR2) in the pathogenesis of several types of inflammatory or neuropathic pain [251]. In addition, spinal application of BDNF in rats has been demonstrated to contribute to spinal late-phase LTP and mechanical hypersensitivity by activation of microglia, Src-family kinases, and p38 MAPK in the absence of any C-fiber activation [252]. Interestingly, the initial processes of central sensitization were suggested to involve the release of BDNF, most likely originating from neurons. This is then followed by a short period of elevated expression of BDNF and phosphorylated trkB in glial cells, which may trigger the transition from early-phase sensitization to late-phase sensitization [253]. Observations on the transgenic mouse L1-DREAM that displays reduced expression in the spinal cord of several genes related to pain, including prodynorphin and BDNF, are consistent with the notion that neuronal BDNF has a primary role in central sensitization [254]. The downstream regulatory element antagonist modulator (DREAM) modulates the endogenous responses to pain and this study found that peripheral inflammation caused an increase in spinal reflexes and the production of BDNF in wild-type mice, but not in DREAM transgenic mice. In vitro, the amplification of the spinal reflexes was replicated with continuous electrical stimulation of C-fibers in wild-type but not in transgenic mice. Finally, transgenic mice displayed a lasting improvement in their dorsal root-ventral root responses because of exogenous BDNF. Thus, BDNF contributes to the heightened sensitivity of the spinal cord following inflammation. Conversely, its absence in DREAM transgenic mice is the cause of the inability of these animals to develop central sensitization in the spinal cord.
The role of BDNF in SCI is however quite elusive, as it has been suggested that SCI modifies the influence of BDNF on the activity of the GABAergic neurons. After SCI, the administration of capsaicin to a single hind paw results in heightened sensitivity to mechanical stimulation that can be averted by delivering a regular (fixed-spaced) stimulation before the injection of the vanilloid [255]. The protective effect was nullified when rats were pre-treated with neutralizing trkB-IgG antibodies. BDNF decreased ERK and pERK after SCI displaying opposing effects than those observed in non-injured rats. These divergent effects were linked to different impacts on GABAergic neurons. In non-injured rats, BDNF reduced the expression of KCC2 and attenuated the inhibitory effect of the GABAA agonist muscimol, whereas, after SCI, BDNF caused an upregulation of KCC2, which would enhance the recovery of GABAergic inhibition.
In another model of neuropathic pain, SNL, it was investigated whether BDNF controlled the activation of GluN2B-NMDA receptors through Src homology-2 domain-containing protein tyrosine phosphatase-2 (SHP2) phosphorylation to produce pain [137]. It was thus established that spinal BDNF is involved in the central sensitization of dorsal horn wide dynamic range (WDR) neurons and the occurrence of pain allodynia in both intact and SNL rats. Furthermore, it was discovered that BDNF triggers LTP in C-fiber synapses by enhancing the activity of GluN2B-NMDA receptors in the spinal dorsal horn. Finally, it was confirmed that the phosphorylation of SHP2 by BDNF is necessary for the subsequent up-regulation of GluN2B-NMDA receptors and the induction of spinal LTP with the development of allodynia.
3.3. Hyperalgesic Priming in the Spinal Cord Dorsal Horn
As mentioned, BDNF interacts with aPKCs in the beginning and maintenance of hyperalgesic priming in the CNS [256]. Activation of nociceptors triggers the release of BDNF in the spinal cord, which, in turn, leads to the translation of aPKC through the activation of the trkB receptor and the involvement of the mammalian target of rapamycin complex 1 (mTORC1). Elevated levels and phosphorylation of aPKCs are believed to have a role in starting priming. After priming is established, the elevated levels of aPKC protein and phosphorylation result in a continuous augmentation of AMPA receptor transportation to the postsynaptic membrane with a persistent augmentation of postsynaptic glutamate-mediated signaling [257]. BDNF signaling through trkB is likely responsible for the regulation of this process. Hyperalgesic priming, once established, can be irreversibly reversed through the inhibition of aPKC using Zrt and Irt-like proteins (ZIP), the disruption of AMPA receptor trafficking with a peptide called pep2M, or the inhibition of trkB/BDNF signaling using ANA-12 or trkB-Fc, respectively.
4. Implications for Pain Management
Understanding the role of BDNF in nociception has significant implications for the management of chronic pain and pain-related conditions. There are three different primary therapeutic strategies derived from preclinical research for pain treatment through inhibition of BDNF/trkB signaling: the sequestration of BDNF, the blocking of the extracellular trkB region, or disruption of the intracellular kinase region of trkB. These approaches have therapeutic potential but are far from being of use in clinical settings.
To scavenge BDNF in living organisms, one can use a trkB-Fc fusion protein derived from the extracellular domain of trkB and the Fc domain of human IgGs [258]. This method has been widely employed to investigate the function of naturally occurring BDNF in various pain models, e.g., [152,259,260]. While the approach is highly targeted, its applicability in clinical settings is limited due to the requirement of intrathecal or local application at the exact central areas where BDNF is released.
To decrease the likelihood of ligand binding, one can target the extracellular domain by using trkB-blocking antibodies [261] or by synthesizing novel receptor antagonists [262]. Monoclonal antibodies that block the function of TrkB receptors have been used successfully in preclinical studies. Specifically, a mouse monoclonal antibody [263] has been shown to effectively block the effects of BDNF on neuronal activity in acute spinal cord slices [59]. Additionally, other neutralizing antibodies have been found to reverse neuropathic [152] and other types of pathological pain [264] in rodents. Like trkB-Fc, anti-trkB blocking antibodies that are delivered systemically do not cross the blood-brain barrier (BBB) and thus cannot penetrate the CNS. This, combined with their relatively limited sensitivity, makes them unsuitable for clinical therapy.
Over the past ten years, the discovery of small new compounds that function as negative allosteric modulators of trkB receptors has led to significant progress in understanding how ligands are processed in the body and their ability to be absorbed into the bloodstream. A peptidomimetic strategy was used to discover cyclotraxin-B, a tiny fragment of BDNF. This fragment can modify the conformation of trkB through an allosteric mechanism [265]. Subsequently, cyclotraxin-B was demonstrated to possess distinct antinociceptive properties in various pain models [74,158,266,267,268,269], to be able to block LTP and central sensitization induced by BDNF [253], as well as the in vitro activation of PKCε in rat Schwann cells that, as mentioned, plays a role in peripheral sensitization [270] (see 3.1 Peripheral sensitization). However, the peptide has little specificity as it affects both BDNF-dependent and -independent trkB activation. In addition, cyclotraxin-B demonstrates efficacy when given intravenously, but not by other tested routes of administration. These shortcomings were bypassed after discovering ANA-12, a non-peptidic small molecule that allosterically inhibits the binding of BDNF to trkB at nanomolar concentrations [262]. ANA-12 has shown great potential in preclinical research, with a growing body of evidence confirming its effectiveness in reducing pain in many experimental settings [94,119,185,188,271,272,273,274,275,276,277,278,279,280,281,282,283,284,285,286]. Unlike cyclotraxin-B, ANA-12 can be administered orally, making it a potential and targeted option for clinical trials.
To inhibit the activity of tyrosine kinases within cells, the primary method is to disrupt the ATP binding site of these enzymes [287,288]. K252a, an indolocarbazole molecule, is among the initial substances employed as a competitive inhibitor of the ATP site, effectively obstructing trk catalytic activity [288,289]. Administration of K252a successfully inhibits the increase of intracellular Ca2+ induced by BDNF in acute slices of the rat superficial dorsal horn [59]. Moreover, in vivo, it diminishes hypersensitivity in several pain models [152,290,291,292,293,294,295,296,297,298]. Based on promising results from preclinical studies, various pharmaceutical companies have allocated significant resources toward developing and ameliorating kinase inhibitors. Many of these inhibitors have been patented and some are (or have been) undergoing clinical trials or investigations for a wide range of diseases and pain [287,299,300,301,302]. While several of these compounds demonstrate significant effectiveness in laboratory tests, the creation of a trkB inhibitor that can be successfully brought to market has not yet been achieved. Regrettably, the task of creating tyrosine kinase antagonists for particular trk receptors is difficult because ATP competitive trk inhibitors, at most, are only selective to trk receptors as a whole (i.e., they are pan-trk inhibitors) and struggle to differentiate between trk subtypes [287]. Hence, a significant drawback associated with the utilization of these compounds is the potential for developing adverse effects on the CNS. To minimize negative consequences, numerous endeavors have been undertaken to create compounds that do not penetrate the BBB but mostly exert their actions at the periphery. Certain compounds, such as ARRY-470 [303] or PF-06273340A [304], demonstrate antinociceptive effects in animal models of chronic pain. The efficacy of the pan-trk inhibitor ONO-4474 as an analgesic in patients with moderate to severe osteoarthritis was demonstrated in a recent randomized, double-blinded clinical trial [305]. However, it is probable that these chemicals also affect peripheral trk receptors in addition to trkB, such as trkA.
As a result of the restrictions mentioned above, there are currently no successful pain therapies that rely on BDNF/trkB. The crucial function performed by BDNF and trkB receptors in the preservation of central neurons imposes further constraints on the utilization of trkB antagonists as a secure treatment strategy. Furthermore, the intricate and diverse nature of the intracellular signaling cascades triggered by tyrosine kinase receptors poses challenges in the development of targeted pharmaceutical interventions. Targeting BDNF/trkB might result in unintended side effects caused by the deactivation of BDNF-dependent pathways crucial for the proper functioning of healthy neurons or by the lack of specificity and interaction with other non-BDNF pathways [306].
To tackle trkB-dependent pathological changes, alternative treatment approaches can be pursued by targeting either the upstream or downstream effectors. Targeting upstream microglia can effectively decrease the BDNF-induced changes in neuropathic pain [174]. Microglial P2RX4, when activated, plays a crucial role in releasing BDNF. Therefore, targeting these receptors could be a promising approach for clinical therapies [307]. As mentioned, one important way that trkB-dependent pain hypersensitivity occurs is through the downregulation of KCC2, which leads to an imbalance between excitatory and inhibitory neurotransmission in downstream signaling pathways [152]. Therefore, KCC2 is considered a potential pharmaceutical target [308] and the making of KCC2 enhancers holds great potential as a therapeutic approach to reinstate inhibition and mitigate the symptoms linked to the BDNF-trkB-KCC2 cascade [309].
5. Clinical Trials
In this section, I will briefly consider the clinical trials of the last ten years related to BDNF in the context of pain modulation. It is worth noting that these trials have focused on measuring BDNF as a biomarker in several clinical pain conditions of a heterogeneous nature, rather than trying to translate into the clinic the results of the preclinical studies that have used the approaches described in the previous section to inhibit BDNF/trkB signaling.
Among the pathologies, one of the most intensely investigated is fibromyalgia, a persistent and recurring disorder characterized by widespread pain that is accompanied by intense emotional discomfort and functional impairment. Several clinical trials were devoted to the study of BDNF levels in fibromyalgia because persons diagnosed with fibromyalgia exhibit elevated levels of serum BDNF compared to healthy persons, indicating a significant involvement of BDNF in the pathophysiology of fibromyalgia [310]. One of the first trials using serum BDNF as a biomarker dates back to 2014 and aimed, among others, at assessing the association with conditional pain modulation (CPM) [311]. In this randomized, double-dummy, placebo-controlled study the cold-heat task (CPM-TASK) was used as an experimental pain stimulus to activate the diffuse noxious inhibitory control-like effect to evaluate the effect of melatonin analgesia. It was thus suggested that BDNF could be used as a biomarker of central sensitization and correlated with pain reduction in the CPM-TASK although with some caveat. In another study, plasma samples were analyzed after a 15-week progressive resistance exercise, and it was shown that the levels of NGF remained unchanged while the levels of BDNF increased suggesting that BDNF may affect nociception/pain in fibromyalgia [312]. In addition, serum BDNF was proposed to be a valuable predictor of the tDCS effect on pain score decreases across the treatment for the condition [313,314]. In keeping with these observations are those in patients not only suffering from fibromyalgia but also other forms of central sensitivity syndrome such as osteoarthritis and endometriosis, leading to somatic and visceral pain, respectively [315]. That serum BDNF inversely correlated with different forms of postoperative pain was also demonstrated using a new predictive tool for postoperative pain, the brief measure of emotional preoperative stress (B-MEPS) to find that blood BDNF was inversely correlated with morphine consumption and length of stay after surgery [316]. Likewise, serum levels of BDNF and trkB were inversely associated with depressive symptoms and sleep quality in patients undergoing adjuvant chemotherapy for breast cancer [317].
The effect of repetitive transcranial magnetic stimulation (rTMS) was investigated in chronic myofascial pain syndrome in a double-blinded, randomized, sham-controlled trial [318]. They observed that rTMS reduced pain scores in parallel with an increase in serum BDNF. The increase in serum BDNF in the rTMS-treated patients was interpreted as an indication of the neuroplasticity that underlies the therapeutic effect of rTMS and serum BDNF was considered as a surrogate marker that could be used to monitor the therapeutic effects of rTMS rather than a marker of the pain severity. These authors have also investigated the effect of electroacupuncture in chronic tension-type headache (CTTH) in a randomized, sham-controlled, crossover trial and concluded that electroacupuncture analgesia in CTTH was related to neuroplasticity that could be monitored by serum BDNF [319]. Partly in agreement with these observations, another group observed a decrease in urinary NGF but not BDNF in patients with interstitial cystitis/bladder pain syndrome treated with hyaluronic acid [320]. Moreover, in a randomized, double-blind, factorial design, and controlled placebo-sham clinical trial using rTMS and deep intramuscular stimulation therapy in chronic myofascial pain syndrome no variations in blood BDNF were observed although both treatments were effective in relieving pain [321]. The variation of serum BDNF levels following integrated multimodal intervention in postherpetic neuralgia was studied in a randomized, double-blind controlled study to conclude that minimally invasive pulsed radiofrequency and pregabalin were effective in early pain reduction that was accompanied by elevated serum BDNF levels [322]. In another randomized double-blind, controlled study to test the effectiveness of intraoperative ketamine on postoperative depressed mood after elective orthopedic surgery, there was an increase in blood BDNF after surgery [323]. Similar results were obtained in a randomized sham-controlled study on patients with persistent chronic pain after hallux valgus surgery where an increase in the level of BDNF in cerebrospinal fluid was observed after anodal transcranial direct current stimulation (tDCS) and was again interpreted as a result of an indirect measurement of neuroplasticity changes induced by tDCS [324]. Similarly, another group observed that serum BDNF increased after three weeks of high-frequency rTMS in patients with SCI [325]. These authors speculated that frequency of rTMS may play an important role in the BDNF secretion processes and in keeping with most of the findings reported above showed a negative correlation between serum BDNF levels and pain scores. Transcutaneous electrical nerve stimulation (TENS) has been used to relief osteoarthritic knee pain and a clinical trial has explored the effects of genotype on TENS efficacy without finding any correlation with expression of the BDNF gene [326].
Altogether, a large part of these and other studies [327,328] converged to the conclusion that BDNF could be a good serum biomarker of neuroplasticity state under different clinical settings related to a pain experience with an increase of serum BDNF in parallel with the reduction of pain. If this interpretation is correct, one should consider that the elevation of BDNF is related to its function as a trophic factor rather than a pain modulator. On the other hand, in a study on fibromyalgia, attachment-based compassion therapy seemed to reduce serum BDNF and appeared to be correlated to anti-inflammatory effects on patients, leading to speculation that reduction in BDNF could be a mechanism of functional status improvement [329].
At the beginning of this year, a study protocol for a randomized clinical trial was proposed aiming to test whether a targeted biobehavioral therapeutic education program could induce alterations in pain perception and biomarkers of brain plasticity (among which BDNF) in individuals suffering from chronic pain [330]. Notably, the authors emphasized that it is crucial to acknowledge that measuring BDNF levels can be challenging because of the inherent unpredictability of collecting, storing, and analyzing samples. Their study will focus on measuring BDNF levels in plasma samples since they have been determined to be more resistant to changes during analysis compared to serum samples [331] which were the most used in the studies quoted above. The investigation will be performed with standardized ELISA kits, incorporating regular calibration and quality control checks to mitigate potential sources of variation. For future studies, implementing these steps is essential to guarantee the precision and authenticity of the BDNF measurements and maintain the integrity and dependability of the study’s results.
6. Conclusion
BDNF plays a significant role in the modulation of nociception and pain signaling. Its effects on peripheral nociceptors, synaptic transmission, and central sensitization are key aspects of pain processing. Understanding the molecular mechanisms behind BDNF’s role in nociception provides valuable insights into potential targets for pain management therapies, offering hope for improved treatments for pain-related conditions. Future research should also be directed at understanding the interactions of BDNF with other pain modulators. GDNF, for example, has antinociceptive opposite effects than BDNF and is present with BDNF in the spinal cord dorsal horn. The circuitry of the interactions of the two molecules in the substantia gelatinosa (lamina II) is starting to be unveiled [15,56,332,333,334] and could offer new opportunities to develop more targeted pain therapies.
Abbreviations
1NMP = 1-(1, 1-dimethyl ethyl)-3-(1-naphthalenyl methyl)-1H-pyrazolo [3,4-d] pyrimidine-4-amine
ACC = anterior cingulate cortex
ALS = amyotrophic lateral sclerosis
AMPA = α-amino-3-hydroxy-5-methyl-4-isoxazole propionic acid
AMPAR = α-amino-3-hydroxy-5-methyl-4-isoxazole propionic acid receptor
AMYG = amygdala
aPKC = atypical isoform of PKC
Au1 =primary auditory area of the cerebral cortex
BBB = blood-brain barrier
BDNF = brain-derived neurotrophic factor
B-MEPS = brief measure of emotional preoperative stress
[Ca++]i = intracellular calcium concentration
CaMKII = calcium/calmodulin-dependent protein kinase II
CCI = chronic constriction injury
CCL2 = chemokine (C-C motif) ligand 2
CCR2 = C-C chemokine receptor type 2
CFA = complete Freund adjuvant
CGRP = calcitonin gene-related peptide
C-LTMRs = C- low-threshold mechanoreceptors
CN = cuneiform nucleus
CNS = central nervous system
CPM = conditional pain modulation
COX2 = cyclooxygenase-2
CREB = cAMP response element-binding protein
CSF-1 = colony-stimulating factor 1
CTTH = chronic tension-type headache
D-APV= D-2-amino-5-phosphonovaleric acid
DCP = dorsal column pathway
DREAM = downstream regulatory element antagonist modulator
DRGs = dorsal root ganglia
EAAT3 = excitatory amino acid transporter 3
ER = endoplasmic reticulum
ERK = extracellular signal-regulated kinase
GABA = γ-amino-butyric acid
GABAAR= γ-amino-butyric acid receptor A
GDNF = glial-derived neurotrophic factor
GFAP = glial fibrillary acidic protein
GLUN2B-NMDA = NMDA receptor GluN2B
GlyR = glycine receptor
HYPO = hypothalamus
IAN = inferior alveolar nerve
Iba1 = ionized calcium-binding adaptor molecule 1
IL-10 = interleukin-10
IL-1β = interleukin-1β
IL-6 = interleukin-6
INS = insular cortex
JG = jugular ganglion
KCC2 =K-Cl co-transporter 2
LTD = long-term depression
LTP = long-term potentiation
MAO-B = type-B monoamine oxidase
mEPSC = miniature excitatory postsynaptic currents
mIPSC = miniature inhibitory postsynaptic currents
mTORC1 = mammalian target of rapamycin complex 1
NaV1.8 = voltage-gated sodium channel 1.8
NF-κB = nuclear factor-kappa B
NGF = nerve growth factor
NMDA = N-methyl-d-aspartate
NMDAR = N-methyl-d-aspartate receptor
NO = nitric oxide
NT-3 = neurotrophin 3
NT-4/5 = neurotrophin-4/5
NTS = nucleus tractus solitarius
P2RX4 = ATP-gated purinergic receptor 4
p75NTR = low-affinity p75 neurotrophin receptor
PAC1R = PACAP type I receptor
PACAP = pituitary adenylate cyclase-activating peptide
PAG = periaqueductal gray
PAR2 = proteinase-activated receptor 2
PBN = parabrachial nucleus
PGE2 = prostaglandin E2
PI3K = phosphoinositide 3-kinase
PKA = protein kinase A
PKC = protein kinase C
PKCε = protein kinase type Cε
PLC = phospholipase C
PNS = peripheral nervous system
PSNs = primary sensory neurons
rTMS = repetitive transcranial magnetic stimulation
RvD1 = resolvin D1
S1 = first-order somatosensory area of the cerebral cortex
S2 = second-order somatosensory area of the cerebral cortex
SCI = spinal cord injury
sEPSC = spontaneous excitatory postsynaptic currents
SHP2 = Src homology-2 domain-containing protein tyrosine phosphatase-2
sIPSC = spontaneous inhibitory postsynaptic currents
SNL = spinal nerve ligation
SNTN = spinal nucleus of the trigeminal nerve
SorCS2 = sortilin-related VPS10 domain-containing receptor 2
SPBP = spinoparabrachial pathway
SSN = somatic sensory neurons
STP = spinothalamic pathway
tDCS = transcranial direct current stimulation
TENS = transcutaneous electrical nerve stimulation
TET1 = ten-eleven translocation methylcytosine dioxygenase
TG = trigeminal ganglion
TNFα = tumor necrosis α
trkB = tropomyosin receptor kinase B
trkB.T1 = truncated isoform 1 of trkB
TRP = transient receptor potential
TRPA1 = transient receptor potential cation channel, subfamily A, member 1
TRPV1 = transient receptor potential vanilloid receptor 1
TTP = trigeminothalamic pathway
VPLN = ventroposterior lateral nucleus of the thalamus
VPMN = ventroposterior medial nucleus of the thalamus
WDR = wide dynamic range
ZIP = Zrt and Irt-like proteins
References
- Ateaque, S.; Merkouris, S.; Barde, Y.A. Neurotrophin signalling in the human nervous system. Front Mol Neurosci 2023, 16, 1225373. [Google Scholar] [CrossRef]
- Arévalo, J.C.; Deogracias, R. Mechanisms Controlling the Expression and Secretion of BDNF. Biomolecules 2023, 13. [Google Scholar] [CrossRef]
- Yang, J.; Siao, C.J.; Nagappan, G.; Marinic, T.; Jing, D.; McGrath, K.; Chen, Z.Y.; Mark, W.; Tessarollo, L.; Lee, F.S.; et al. Neuronal release of proBDNF. Nat.Neurosci. 2009, 12, 113–115. [Google Scholar] [CrossRef]
- Wang, C.S.; Kavalali, E.T.; Monteggia, L.M. BDNF signaling in context: From synaptic regulation to psychiatric disorders. Cell 2022, 185, 62–76. [Google Scholar] [CrossRef]
- Lu, B.; Nagappan, G.; Lu, Y. BDNF and Synaptic Plasticity, Cognitive Function, and Dysfunction. In Neurotrophic Factors; Lewin, G.R., Carter, B.D., Eds.; Springer Berlin Heidelberg: Berlin, Heidelberg, 2014; pp. 223–250. [Google Scholar]
- Miranda, M.; Morici, J.F.; Zanoni, M.B.; Bekinschtein, P. Brain-Derived Neurotrophic Factor: A Key Molecule for Memory in the Healthy and the Pathological Brain. Front Cell Neurosci 2019, 13. [Google Scholar] [CrossRef] [PubMed]
- Nagahara, A.H.; Merrill, D.A.; Coppola, G.; Tsukada, S.; Schroeder, B.E.; Shaked, G.M.; Wang, L.; Blesch, A.; Kim, A.; Conner, J.M.; et al. Neuroprotective effects of brain-derived neurotrophic factor in rodent and primate models of Alzheimer’s disease. Nat Med 2009, 15, 331–337. [Google Scholar] [CrossRef] [PubMed]
- Palasz, E.; Wysocka, A.; Gasiorowska, A.; Chalimoniuk, M.; Niewiadomski, W.; Niewiadomska, G. BDNF as a Promising Therapeutic Agent in Parkinson’s Disease. Int J Mol Sci 2020, 21. [Google Scholar] [CrossRef] [PubMed]
- Zuccato, C.; Ciammola, A.; Rigamonti, D.; Leavitt, B.R.; Goffredo, D.; Conti, L.; MacDonald, M.E.; Friedlander, R.M.; Silani, V.; Hayden, M.R.; et al. Loss of huntingtin-mediated BDNF gene transcription in Huntington’s disease. Science 2001, 293, 493–498. [Google Scholar] [CrossRef] [PubMed]
- Ou, Z.-Y.A.; Byrne, L.M.; Rodrigues, F.B.; Tortelli, R.; Johnson, E.B.; Foiani, M.S.; Arridge, M.; De Vita, E.; Scahill, R.I.; Heslegrave, A.; et al. Brain-derived neurotrophic factor in cerebrospinal fluid and plasma is not a biomarker for Huntington’s disease. Scientific Reports 2021, 11, 3481. [Google Scholar] [CrossRef] [PubMed]
- Pradhan, J.; Noakes, P.G.; Bellingham, M.C. The Role of Altered BDNF/TrkB Signaling in Amyotrophic Lateral Sclerosis. Front Cell Neurosci 2019, 13. [Google Scholar] [CrossRef]
- Riolo, G.; Ricci, C.; De Angelis, N.; Marzocchi, C.; Guerrera, G.; Borsellino, G.; Giannini, F.; Battistini, S. BDNF and Pro-BDNF in Amyotrophic Lateral Sclerosis: A New Perspective for Biomarkers of Neurodegeneration. Brain Sci 2022, 12. [Google Scholar] [CrossRef] [PubMed]
- Duman, R.S.; Monteggia, L.M. A neurotrophic model for stress-related mood disorders. Biol Psychiatry 2006, 59, 1116–1127. [Google Scholar] [CrossRef] [PubMed]
- Martinowich, K.; Manji, H.; Lu, B. New insights into BDNF function in depression and anxiety. Nat Neurosci 2007, 10, 1089–1093. [Google Scholar] [CrossRef] [PubMed]
- Merighi, A. Neuropeptides and Coexistence. In Reference Module in Neuroscience and Biobehavioral Psychology; Elsevier, 2017. [Google Scholar]
- Raja, S.N.; Carr, D.B.; Cohen, M.; Finnerup, N.B.; Flor, H.; Gibson, S.; Keefe, F.J.; Mogil, J.S.; Ringkamp, M.; Sluka, K.A.; et al. The revised International Association for the Study of Pain definition of pain: concepts, challenges, and compromises. Pain 2020, 161, 1976–1982. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, S.A.; Herr, M.J. Physiology, Nociception. In StatPearls; StatPearls Publishing LLC.: Treasure Island (FL), 2023. [Google Scholar]
- Dubin, A.E.; Patapoutian, A. Nociceptors: the sensors of the pain pathway. J Clin Invest 2010, 120, 3760–3772. [Google Scholar] [CrossRef] [PubMed]
- Frias, B.; Merighi, A. Capsaicin, Nociception and Pain. Molecules 2016, 21, 797. [Google Scholar] [CrossRef]
- Vogt, B.A. Pain and emotion interactions in subregions of the cingulate gyrus. Nat Rev Neurosci 2005, 6, 533–544. [Google Scholar] [CrossRef]
- Grichnik, K.P.; Ferrante, F.M. The difference between acute and chronic pain. Mt Sinai J Med 1991, 58, 217–220. [Google Scholar]
- Merighi, A.; Salio, C.; Ghirri, A.; Lossi, L.; Ferrini, F.; Betelli, C.; Bardoni, R. BDNF as a pain modulator. Prog.Neurobiol. 2008, 85, 297–317. [Google Scholar] [CrossRef]
- Murphy, P.M. Somatic Pain. In Encyclopedia of Pain; Schmidt, R.F., Willis, W.D., Eds.; Springer Berlin Heidelberg: Berlin, Heidelberg, 2007; pp. 2190–2192. [Google Scholar]
- Price, D.D. Psychological and neural mechanisms of the affective dimension of pain. Science 2000, 288, 1769–1772. [Google Scholar] [CrossRef]
- Michael, G.J.; Averill, S.; Shortland, P.J.; Yan, Q.; Priestley, J.V. Axotomy results in major changes in BDNF expression by dorsal root ganglion cells: BDNF expression in large trkB and trkC cells, in pericellular baskets, and in projections to deep dorsal horn and dorsal column nuclei. Eur.J.Neurosci. 1999, 11, 3539. [Google Scholar] [CrossRef]
- Hudspith, M.J.; Siddall, P.J.; Munglani, R. Chapter 23—Physiology of pain. In Foundations of Anesthesia, 2nd ed.; Hemmings, H.C., Hopkins, P.M., Eds.; Mosby: Edinburgh, 2006; pp. 267–285. [Google Scholar]
- Kim, H.Y.; Wang, J.; Gwak, Y.S. Gracile neurons contribute to the maintenance of neuropathic pain in peripheral and central neuropathic models. J Neurotrauma 2012, 29, 2587–2592. [Google Scholar] [CrossRef] [PubMed]
- Gebhart, G.F. Visceral Pain. In Encyclopedia of Neuroscience; Squire, L.R., Ed.; Academic Press: Oxford, 2009; pp. 189–194. [Google Scholar]
- Vardeh, D.; Naranjo, J.F. Peripheral and Central Sensitization. In Pain Medicine: An Essential Review; Yong, R.J., Nguyen, M., Nelson, E., Urman, R.D., Eds.; Springer International Publishing: Cham, 2017; pp. 15–17. [Google Scholar]
- Li, C.; Kim, H.J.; Back, S.K.; Na, H.S. Common and discrete mechanisms underlying chronic pain and itch: peripheral and central sensitization. Pflügers Archiv—European Journal of Physiology 2021, 473, 1603–1615. [Google Scholar] [CrossRef]
- Seidel, M.F.; Hügle, T.; Morlion, B.; Koltzenburg, M.; Chapman, V.; MaassenVanDenBrink, A.; Lane, N.E.; Perrot, S.; Zieglgänsberger, W. Neurogenic inflammation as a novel treatment target for chronic pain syndromes. Exp Neurol 2022, 356, 114108. [Google Scholar] [CrossRef] [PubMed]
- Sikandar, S.; Minett, M.S.; Millet, Q.; Santana-Varela, S.; Lau, J.; Wood, J.N.; Zhao, J. Brain-derived neurotrophic factor derived from sensory neurons plays a critical role in chronic pain. Brain 2018, 141, 1028–1039. [Google Scholar] [CrossRef]
- Altar, C.A.; Cai, N.; Bliven, T.; Juhasz, M.; Conner, J.M.; Acheson, A.L.; Lindsay, R.M.; Wiegand, S.J. Anterograde transport of brain-derived neurotrophic factor and its role in the brain. Nature 1997, 389, 856–860. [Google Scholar] [CrossRef]
- von Bartheld, C.S.; Wang, X.; Butowt, R. Anterograde axonal transport, transcytosis, and recycling of neurotrophic factors: the concept of trophic currencies in neural networks. Mol.Neurobiol. 2001, 24, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.F.; Song, X.Y.; Zhong, J.H.; Barati, S.; Zhou, F.H.; Johnson, S.M. Distribution and localization of pro-brain-derived neurotrophic factor-like immunoreactivity in the peripheral and central nervous system of the adult rat. J.Neurochem. 2004, 91, 704–715. [Google Scholar] [CrossRef] [PubMed]
- Conner, J.M.; Lauterborn, J.C.; Yan, Q.; Gall, C.M.; Varon, S. Distribution of brain-derived neurotrophic factor (BDNF) protein and mRNA in the normal adult rat CNS: evidence for anterograde axonal transport. J.Neurosci. 1997, 17, 2295–2313. [Google Scholar] [CrossRef]
- Kohara, K.; Kitamura, A.; Morishima, M.; Tsumoto, T. Activity-dependent transfer of brain-derived neurotrophic factor to postsynaptic neurons. Science 2001, 291, 2419–2423. [Google Scholar] [CrossRef]
- Kerr, B.J.; Bradbury, E.J.; Bennett, D.L.; Trivedi, P.M.; Dassan, P.; French, J.; Shelton, D.B.; McMahon, S.B.; Thompson, S.W. Brain-derived neurotrophic factor modulates nociceptive sensory inputs and NMDA-evoked responses in the rat spinal cord. J.Neurosci. 1999, 19, 5138–5148. [Google Scholar] [CrossRef] [PubMed]
- Michael, G.J.; Averill, S.; Nitkunan, A.; Rattray, M.; Bennett, D.L.H.; Yan, Q.; Priestley, J.V. Nerve growth factor treatment increases brain-derived neurotrophic factor selectively in trk-A expressing dorsal root ganglion cells and in their central terminations within the spinal cord. J.Neurosci. 1997, 17, 8476–8490. [Google Scholar] [CrossRef] [PubMed]
- Bothwell, M. Recent advances in understanding context-dependent mechanisms controlling neurotrophin signaling and function. F1000Research 2019, 8. [Google Scholar] [CrossRef]
- Zhou, X.-F.; Rush, R.A. Endogenous brain-derived neurothrophic factor is anterogradely transported in primary sensory neurons. Neuroscience 1996, 74, 945–951. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.F.; Chie, E.T.; Rush, R.A. Distribution of brain-derived neurotrophic factor in cranial and spinal ganglia. Exp Neurol 1998, 149, 237–242. [Google Scholar] [CrossRef] [PubMed]
- Ichikawa, H.; Terayama, R.; Yamaai, T.; Yan, Z.; Sugimoto, T. Brain-derived neurotrophic factor-immunoreactive neurons in the rat vagal and glossopharyngeal sensory ganglia; co-expression with other neurochemical substances. Brain Res. 2007, 1155, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Price, J. An immunohistochemical and quantitative examination of dorsal root ganglion neuronal subpopulations. J Neurosci 1985, 5, 2051–2059. [Google Scholar] [CrossRef]
- Lallemend, F.; Ernfors, P. Molecular interactions underlying the specification of sensory neurons. Trends Neurosci 2012, 35, 373–381. [Google Scholar] [CrossRef]
- Wang, K.; Cai, B.; Song, Y.; Chen, Y.; Zhang, X. Somatosensory neuron types and their neural networks as revealed via single-cell transcriptomics. Trends Neurosci 2023, 46, 654–666. [Google Scholar] [CrossRef]
- Vermeiren, S.; Bellefroid, E.J.; Desiderio, S. Vertebrate Sensory Ganglia: Common and Divergent Features of the Transcriptional Programs Generating Their Functional Specialization. Frontiers in Cell and Developmental Biology 2020, 8. [Google Scholar] [CrossRef]
- Capra, N.F.; Wax, T.D. Distribution and central projections of primary afferent neurons that innervate the masseter muscle and mandibular periodontium: a double-label study. J Comp Neurol 1989, 279, 341–352. [Google Scholar] [CrossRef]
- Cobo, J.L.; Solé-Magdalena, A.; Menéndez, I.; de Vicente, J.C.; Vega, J.A. Connections between the facial and trigeminal nerves: Anatomical basis for facial muscle proprioception. JPRAS Open 2017, 12, 9–18. [Google Scholar] [CrossRef]
- Delfini, M.C.; Mantilleri, A.; Gaillard, S.; Hao, J.; Reynders, A.; Malapert, P.; Alonso, S.; Francois, A.; Barrere, C.; Seal, R.; et al. TAFA4, a Chemokine-like Protein, Modulates Injury-Induced Mechanical and Chemical Pain Hypersensitivity in Mice. Cell Reports 2013, 5, 378–388. [Google Scholar] [CrossRef]
- Emery, E.C.; Ernfors, P. Dorsal Root Ganglion Neuron Types and Their Functional Specialization. In The Oxford Handbook of the Neurobiology of Pain; Wood, J.N., Ed.; Oxford University Press, 2020. [Google Scholar]
- Kupari, J.; Häring, M.; Agirre, E.; Castelo-Branco, G.; Ernfors, P. An Atlas of Vagal Sensory Neurons and Their Molecular Specialization. Cell Rep 2019, 27, 2508–2523.e2504. [Google Scholar] [CrossRef] [PubMed]
- Averill, S.; McMahon, S.B.; Clary, D.O.; Reichardt, L.F.; Priestley, J.V. Immunocytochemical localization of trkA receptors in chemically identified subgroups of adult rat sensory neurons. Eur.J.Neurosci. 1995, 7, 1484–1494. [Google Scholar] [CrossRef] [PubMed]
- Lawson, S.N. Phenotype and Function of Somatic Primary Afferent Nociceptive Neurones with C-, Aδ- or Aα/β-Fibres. Exp Physiol 2002, 87, 239–244. [Google Scholar] [CrossRef] [PubMed]
- Merighi, A.; Polak, J.M.; Gibson, S.J.; Gulbenkian, S.; Valentino, K.L.; Peirone, S.M. Ultrastructural studies on calcitonin gene-related peptide-, tachykinins- and somatostatin-immunoreactive neurones in rat dorsal root ganglia: Evidence for the colocalisation of different peptides in single secretory granules. Cell Tiss.Res. 1988, 254, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Merighi, A. The histology, physiology, neurochemistry and circuitry of the substantia gelatinosa Rolandi (lamina II) in mammalian spinal cord. Prog.Neurobiol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Salio, C.; Lossi, L.; Ferrini, F.; Merighi, A. Ultrastructural evidence for a pre- and post-synaptic localization of full length trkB receptors in substantia gelatinosa (lamina II) of rat and mouse spinal cord. Eur.J.Neurosci. 2005, 22, 1951–1966. [Google Scholar] [CrossRef] [PubMed]
- Salio, C.; Averill, S.; Priestley, J.V.; Merighi, A. Costorage of BDNF and neuropeptides within individual dense-core vesicles in central and peripheral neurons. Dev.Neurobiol. 2007, 67, 326–338. [Google Scholar] [CrossRef] [PubMed]
- Merighi, A.; Bardoni, R.; Salio, C.; Lossi, L.; Ferrini, F.; Prandini, M.; Zonta, M.; Gustincich, S.; Carmignoto, G. Presynaptic functional trkB receptors mediate the release of excitatory neurotransmitters from primary afferent terminals in lamina II (substantia gelatinosa) of postnatal rat spinal cord. Dev.Neurobiol. 2008, 68, 457–475. [Google Scholar] [CrossRef] [PubMed]
- Salio, C.; Ferrini, F. BDNF and GDNF expression in discrete populations of nociceptors. Ann.Anat. 2016, 207, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Xia, C.M.; Gulick, M.A.; Yu, S.J.; Grider, J.R.; Murthy, K.S.; Kuemmerle, J.F.; Akbarali, H.I.; Qiao, L.Y. Up-regulation of brain-derived neurotrophic factor in primary afferent pathway regulates colon-to-bladder cross-sensitization in rat. J Neuroinflammation 2012, 9, 30. [Google Scholar] [CrossRef] [PubMed]
- Ha, S.O.; Kim, J.K.; Hong, H.S.; Kim, D.S.; Cho, H.J. Expression of brain-derived neurotrophic factor in rat dorsal root ganglia, spinal cord and gracile nuclei in experimental models of neuropathic pain. Neuroscience 2001, 107, 301–309. [Google Scholar] [CrossRef]
- Lin, Y.-T.; Ro, L.-S.; Wang, H.-L.; Chen, J.-C. Up-regulation of dorsal root ganglia BDNF and trkB receptor in inflammatory pain: an in vivo and in vitrostudy. Journal of Neuroinflammation 2011, 8, 126. [Google Scholar] [CrossRef]
- Cho, H.J.; Kim, J.K.; Zhou, X.F.; Rush, R.A. Increased brain-derived neurotrophic factor immunoreactivity in rat dorsal root ganglia and spinal cord following peripheral inflammation. Brain Res. 1997, 764, 269–272. [Google Scholar] [CrossRef]
- Tomotsuka, N.; Kaku, R.; Obata, N.; Matsuoka, Y.; Kanzaki, H.; Taniguchi, A.; Muto, N.; Omiya, H.; Itano, Y.; Sato, T.; et al. Up-regulation of brain-derived neurotrophic factor in the dorsal root ganglion of the rat bone cancer pain model. Journal of pain research 2014, 7, 415–423. [Google Scholar] [CrossRef]
- Waite, P.M.E. CHAPTER 26—Trigeminal Sensory System. In The Rat Nervous System, 3rd ed.; Paxinos, G., Ed.; Academic Press: Burlington, 2004; pp. 817–851. [Google Scholar]
- Ichikawa, H.; Yabuuchi, T.; Jin, H.W.; Terayama, R.; Yamaai, T.; Deguchi, T.; Kamioka, H.; Takano-Yamamoto, T.; Sugimoto, T. Brain-derived neurotrophic factor-immunoreactive primary sensory neurons in the rat trigeminal ganglion and trigeminal sensory nuclei. Brain Res. 2006, 1081, 113–118. [Google Scholar] [CrossRef]
- Buldyrev, I.; Tanner, N.M.; Hsieh, H.y.; Dodd, E.G.; Nguyen, L.T.; Balkowiec, A. Calcitonin gene-related peptide enhances release of native brain-derived neurotrophic factor from trigeminal ganglion neurons. Journal of Neurochemistry 2006, 99, 1338–1350. [Google Scholar] [CrossRef]
- Usoskin, D.; Furlan, A.; Islam, S.; Abdo, H.; Lonnerberg, P.; Lou, D.; Hjerling-Leffler, J.; Haeggstrom, J.; Kharchenko, O.; Kharchenko, P.V.; et al. Unbiased classification of sensory neuron types by large-scale single-cell RNA sequencing. Nat.Neurosci. 2015, 18, 145–153. [Google Scholar] [CrossRef]
- Wang, J.; Kollarik, M.; Ru, F.; Sun, H.; McNeil, B.; Dong, X.; Stephens, G.; Korolevich, S.; Brohawn, P.; Kolbeck, R.; et al. Distinct and common expression of receptors for inflammatory mediators in vagal nodose versus jugular capsaicin-sensitive/TRPV1-positive neurons detected by low input RNA sequencing. PLoS One 2017, 12, e0185985. [Google Scholar] [CrossRef]
- Huerta, T.S.; Haider, B.; Adamovich-Zeitlin, R.; Chen, A.C.; Chaudhry, S.; Zanos, T.P. Calcium imaging and analysis of the jugular-nodose ganglia enables identification of distinct vagal sensory neuron subsets. 2023, 20. [Google Scholar] [CrossRef]
- Brady, R.; Zaidi, S.I.; Mayer, C.; Katz, D.M. BDNF is a target-derived survival factor for arterial baroreceptor and chemoafferent primary sensory neurons. J Neurosci 1999, 19, 2131–2142. [Google Scholar] [CrossRef] [PubMed]
- Clark, C.G.; Hasser, E.M.; Kunze, D.L.; Katz, D.M.; Kline, D.D. Endogenous brain-derived neurotrophic factor in the nucleus tractus solitarius tonically regulates synaptic and autonomic function. J Neurosci 2011, 31, 12318–12329. [Google Scholar] [CrossRef]
- Thibault, K.; Lin, W.K.; Rancillac, A.; Fan, M.; Snollaerts, T.; Sordoillet, V.; Hamon, M.; Smith, G.M.; Lenkei, Z.; Pezet, S. BDNF-dependent plasticity induced by peripheral inflammation in the primary sensory and the cingulate cortex triggers cold allodynia and reveals a major role for endogenous BDNF as a tuner of the affective aspect of pain. J Neurosci 2014, 34, 14739–14751. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, G.; Ma, J.; Liu, C.; Liu, X.; Zhan, Y.; Zhang, M. Brain-derived neurotrophic factor (BDNF) in the rostral anterior cingulate cortex (rACC) contributes to neuropathic spontaneous pain-related aversion via NR2B receptors. Brain Res Bull 2016, 127, 56–65. [Google Scholar] [CrossRef]
- Li, N.; Chen, B.; Jia, G.; Xu, R.; Xia, Y.; Lai, C.; Li, G.; Li, W.; Han, Y. Reduced BDNF expression in the auditory cortex contributed to neonatal pain-induced hearing impairment and dendritic pruning deficiency in mice. Reg Anesth Pain Med 2023, 48, 85–92. [Google Scholar] [CrossRef]
- Ma, L.; Yue, L.; Zhang, Y.; Wang, Y.; Han, B.; Cui, S.; Liu, F.Y.; Wan, Y.; Yi, M. Spontaneous Pain Disrupts Ventral Hippocampal CA1-Infralimbic Cortex Connectivity and Modulates Pain Progression in Rats with Peripheral Inflammation. Cell Rep 2019, 29, 1579–1593. [Google Scholar] [CrossRef] [PubMed]
- Chung, E.K.; Bian, Z.X.; Xu, H.X.; Sung, J.J. Neonatal maternal separation increases brain-derived neurotrophic factor and tyrosine kinase receptor B expression in the descending pain modulatory system. Neurosignals 2009, 17, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Liu, R.; Guo, F.; Wen, M.Q.; Ma, X.L.; Li, K.Y.; Sun, H.; Xu, C.L. Parabrachial nucleus circuit governs neuropathic pain-like behavior. 2020, 11, 5974. [CrossRef]
- Sarhan, M.; Pawlowski, S.A.; Barthas, F.; Yalcin, I.; Kaufling, J.; Dardente, H.; Zachariou, V.; Dileone, R.J.; Barrot, M.; Veinante, P. BDNF parabrachio-amygdaloid pathway in morphine-induced analgesia. Int.J.Neuropsychopharmacol. 2013, 16, 1649–1660. [Google Scholar] [CrossRef]
- Obata, K.; Yamanaka, H.; Dai, Y.; Tachibana, T.; Fukuoka, T. Differential activation of extracellular signal-regulated protein kinase in primary afferent neurons regulates brain-derived neurotrophic factor expression after peripheral inflammation and nerve injury. J.Neurosci. 2003, 23, 4117. [Google Scholar] [CrossRef] [PubMed]
- Qiao, L.-y.; Gulick, M.A.; Bowers, J.; Kuemmerle, J.F.; Grider, J.R. Differential changes in brain-derived neurotrophic factor and extracellular signal-regulated kinase in rat primary afferent pathways with colitis. Neurogastroenterology & Motility 2008, 20, 928–938. [Google Scholar]
- Garraway, S.M.; Petruska, J.C.; Mendell, L.M. BDNF sensitizes the response of lamina II neurons to high threshold primary afferent inputs. Eur.J Neurosci. 2003, 18, 2467–2476. [Google Scholar] [CrossRef] [PubMed]
- Garraway, S.M.; Anderson, A.J.; Mendell, L.M. BDNF-induced facilitation of afferent-evoked responses in lamina II neurons is reduced after neonatal spinal cord contusion injury. J.Neurophysiol. 2005, 94, 1798–1804. [Google Scholar] [CrossRef]
- Yaksh, T.L. Calcium Channels As Therapeutic Targets in Neuropathic Pain. The Journal of Pain 2006, 7, S13–S30. [Google Scholar] [CrossRef]
- Garraway, S.M.; Huie, J.R. Spinal Plasticity and Behavior: BDNF-Induced Neuromodulation in Uninjured and Injured Spinal Cord. Neural Plast 2016, 2016, 9857201. [Google Scholar] [CrossRef] [PubMed]
- Pezet, S.; Malcangio, M.; McMahon, S.B. BDNF: a neuromodulator in nociceptive pathways? Brain Res.Brain Res.Rev. 2002, 40, 240–249. [Google Scholar] [CrossRef] [PubMed]
- Merighi, A.; Carmignoto, G.; Gobbo, S.; Lossi, L.; Salio, C.; Vergnano, A.M.; Zonta, M. Neurotrophins in spinal cord nociceptive pathways. Prog.Brain Res. 2004, 146, 291–321. [Google Scholar]
- Pezet, S.; McMahon, S.B. NEUROTROPHINS: Mediators and Modulators of Pain. Annual Review of Neuroscience 2006, 29, 507–538. [Google Scholar] [CrossRef]
- Dembo, T.; Braz, J.M. Primary Afferent-Derived BDNF Contributes Minimally to the Processing of Pain and Itch. 2018, 5. [CrossRef]
- Lossi, L.; Merighi, A. The Use of ex Vivo Rodent Platforms in Neuroscience Translational Research With Attention to the 3Rs Philosophy. Frontiers in Veterinary Science 2018, 5, 164. [Google Scholar] [CrossRef]
- Lu, V.B.; Moran, T.D.; Balasubramanyan, S.; Alier, K.A.; Dryden, W.F.; Colmers, W.F.; Smith, P.A. Substantia Gelatinosa neurons in defined-medium organotypic slice culture are similar to those in acute slices from young adult rats. Pain. 2006, 121, 261–275. [Google Scholar] [CrossRef]
- Kerr, B.J.; Bradbury, E.J.; Bennett, D.L.; Trivedi, P.M.; Dassan, P.; French, J.; Shelton, D.B.; McMahon, S.B.; Thompson, S.W. Brain-derived neurotrophic factor modulates nociceptive sensory inputs and NMDA-evoked responses in the rat spinal cord. J Neurosci. 1999, 19, 5138–5148. [Google Scholar] [CrossRef]
- Chen, W.; Walwyn, W.; Ennes, H.S.; Kim, H.; McRoberts, J.A.; Marvizón, J.C. BDNF released during neuropathic pain potentiates NMDA receptors in primary afferent terminals. Eur J Neurosci 2014, 39, 1439–1454. [Google Scholar] [CrossRef] [PubMed]
- Biggs, J.E.; Lu, V.B.; Kim, H.J.; Lai, A.; Todd, K.G.; Ballanyi, K.; Colmers, W.F.; Smith, P.A. Defined Medium Organotypic Cultures of Spinal Cord Put ‘Pain in a Dish’. In Isolated Central Nervous System Circuits; Ballanyi, K., Ed.; Humana Press: Totowa, NJ, 2012; pp. 405–436. [Google Scholar]
- Biggs, J.E.; Lu, V.B.; Stebbing, M.J.; Balasubramanyan, S.; Smith, P.A. Is BDNF sufficient for information transfer between microglia and dorsal horn neurons during the onset of central sensitization? Mol.Pain 2010, 6, 44. [Google Scholar] [CrossRef] [PubMed]
- Smith, P.A. BDNF: No gain without pain? Neuroscience 2014, 283, 107–123. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Lei, Y.; Tian, Y.; Xu, S.; Shen, X.; Wu, H.; Bao, S.; Wang, F. The etiological contribution of GABAergic plasticity to the pathogenesis of neuropathic pain. Mol Pain 2019, 15, 1744806919847366. [Google Scholar] [CrossRef] [PubMed]
- Pezet, S.; Cunningham, J.; Patel, J.; Grist, J.; Gavazzi, I.; Lever, I.J.; Malcangio, M. BDNF modulates sensory neuron synaptic activity by a facilitation of GABA transmission in the dorsal horn. Mol.Cell Neurosci. 2002, 21, 51–62. [Google Scholar] [CrossRef] [PubMed]
- Bardoni, R.; Ghirri, A.; Salio, C.; Prandini, M.; Merighi, A. BDNF-mediated modulation of GABA and glycine release in dorsal horn lamina II from postnatal rats. Dev.Neurobiol. 2007, 67, 960–975. [Google Scholar] [CrossRef] [PubMed]
- Lu, V.B.; Colmers, W.F.; Smith, P.A. Long-term actions of BDNF on inhibitory synaptic transmission in identified neurons of the rat substantia gelatinosa. Journal of Neurophysiology 2012, 108, 441. [Google Scholar] [CrossRef] [PubMed]
- Dedek, A.; Xu, J.; Lorenzo, L.; Godin, A.G.; Kandegedara, C.M.; Glavina, G.; Landrigan, J.A.; Lombroso, P.J.; De Koninck, Y.; Tsai, E.C.; et al. Sexual dimorphism in a neuronal mechanism of spinal hyperexcitability across rodent and human models of pathological pain. Brain 2022, 145, 1124–1138. [Google Scholar] [CrossRef]
- Zhao, J.; Seereeram, A.; Nassar, M.A.; Levato, A.; Pezet, S. Nociceptor-derived brain-derived neurotrophic factor regulates acute and inflammatory but not neuropathic pain. Mol.Cell Neurosci. 2006. [Google Scholar] [CrossRef]
- Groth, R.; Aanonsen, L. Spinal brain-derived neurotrophic factor (BDNF) produces hyperalgesia in normal mice while antisense directed against either BDNF or trkB, prevent inflammation-induced hyperalgesia. Pain 2002, 100, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Slack, S.E.; Thompson, S.W. Brain-derived neurotrophic factor induces NMDA receptor 1 phosphorylation in rat spinal cord. Neuroreport 2002, 13, 1967–1970. [Google Scholar] [CrossRef] [PubMed]
- Matayoshi, S.; Jiang, N.; Katafuchi, T.; Koga, K.; Furue, H.; Yasaka, T.; Nakatsuka, T.; Zhou, X.F.; Kawasaki, Y.; Tanaka, N.; et al. Actions of brain-derived neurotrophic factor on spinal nociceptive transmission during inflammation in the rat. J.Physiol. 2005, 569, 685–695. [Google Scholar] [CrossRef] [PubMed]
- Lalisse, S.; Hua, J.; Lenoir, M.; Linck, N.; Rassendren, F. Sensory neuronal P2RX4 receptors controls BDNF signaling in inflammatory pain. 2018, 8, 964. [CrossRef]
- Martin, K.K.; Noble, D.J.; Parvin, S.; Jang, K.; Garraway, S.M. Pharmacogenetic inhibition of TrkB signaling in adult mice attenuates mechanical hypersensitivity and improves locomotor function after spinal cord injury. Front Cell Neurosci 2022, 16, 987236. [Google Scholar] [CrossRef] [PubMed]
- Bonomini, F.; Favero, G.; Castrezzati, S.; Borsani, E. Role of Neurotrophins in Orofacial Pain Modulation: A Review of the Latest Discoveries. International journal of molecular sciences 2023, 24, 12438. [Google Scholar] [CrossRef] [PubMed]
- Bradnam, L.; Barry, C. The role of the trigeminal sensory nuclear complex in the pathophysiology of craniocervical dystonia. J Neurosci 2013, 33, 18358–18367. [Google Scholar] [CrossRef] [PubMed]
- Takeda, M.; Takahashi, M.; Kitagawa, J.; Kanazawa, T.; Nasu, M.; Matsumoto, S. Brain-derived neurotrophic factor enhances the excitability of small-diameter trigeminal ganglion neurons projecting to the trigeminal nucleus interpolaris/caudalis transition zone following masseter muscle inflammation. Mol Pain 2013, 9, 49. [Google Scholar] [CrossRef]
- Grayson, M.; Arris, D. Oral squamous cell carcinoma-released brain-derived neurotrophic factor contributes to oral cancer pain by peripheral tropomyosin receptor kinase B activation. 2022, 163, 496–507. [CrossRef]
- Wang, H.; Wei, Y.; Pu, Y.; Jiang, D.; Jiang, X.; Zhang, Y.; Tao, J. Brain-derived neurotrophic factor stimulation of T-type Ca<sup>2+</sup> channels in sensory neurons contributes to increased peripheral pain sensitivity. Science signaling 2019, 12, eaaw2300. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Chung, M.-K. Orthodontic force induces nerve injury-like transcriptomic changes driven by TRPV1-expressing afferents in mouse trigeminal ganglia. Molecular Pain 2020, 16, 1744806920973141. [Google Scholar] [CrossRef]
- Koshida, T.; Maruta, T.; Tanaka, N.; Hidaka, K.; Kurogi, M.; Nemoto, T.; Yanagita, T.; Takeya, R.; Tsuneyoshi, I. Changes in TRPV1 Receptor, CGRP, and BDNF Expression in Rat Dorsal Root Ganglion with Resiniferatoxin-Induced Neuropathic Pain: Modulation by Pulsed Radiofrequency Applied to the Sciatic Nerve. Acta Med Okayama 2023, 77, 359–364. [Google Scholar] [CrossRef]
- Kooshki, R.; Abbasnejad, M.; Mahani, S.E.; Raoof, M.; Aghtaei, M.M.M.; Dabiri, S. Orexin-A inhibits capsaicin-induced changes in cyclooxygenase-2 and brain-derived neurotrophic factor expression in trigeminal nucleus caudalis of rats. Korean J Pain 2018, 31, 174–182. [Google Scholar] [CrossRef]
- Scarabelot, V.L.; de Oliveira, C.; Medeiros, L.F.; de Macedo, I.C.; Cioato, S.G.; Adachi, L.N.S.; Paz, A.H.; de Souza, A.; Caumo, W.; Torres, I.L.S. Transcranial direct-current stimulation reduces nociceptive behaviour in an orofacial pain model. J Oral Rehabil 2019, 46, 40–50. [Google Scholar] [CrossRef]
- Zhang, L.; Zhou, Y.; Yang, L.; Wang, Y.; Xiao, Z. PACAP6-38 improves nitroglycerin-induced central sensitization by modulating synaptic plasticity at the trigeminal nucleus caudalis in a male rat model of chronic migraine. The Journal of Headache and Pain 2023, 24, 66. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Zhang, Y.; Liu, Q.; Jiang, L.; Li, M.; Wang, S.; Long, T.; He, W.; Kong, X.; Qin, G.; et al. P2X4-receptor participates in EAAT3 regulation via BDNF-TrkB signaling in a model of trigeminal allodynia. Mol Pain 2018, 14, 1744806918795930. [Google Scholar] [CrossRef] [PubMed]
- Luo, C.; Zhong, X.L.; Zhou, F.H.; Li, J.Y.; Zhou, P.; Xu, J.M.; Song, B.; Li, C.Q.; Zhou, X.F.; Dai, R.P. Peripheral Brain Derived Neurotrophic Factor Precursor Regulates Pain as an Inflammatory Mediator. Sci Rep 2016, 6, 27171. [Google Scholar] [CrossRef] [PubMed]
- Balkowiec, A.; Katz, D.M. Activity-dependent release of endogenous brain-derived neurotrophic factor from primary sensory neurons detected by ELISA in situ. J Neurosci. 2000, 20, 7417–7423. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.Y.; Ieraci, A.; Teng, H.; Dall, H.; Meng, C.X.; Herrera, D.G.; Nykjaer, A.; Hempstead, B.L.; Lee, F.S. Sortilin controls intracellular sorting of brain-derived neurotrophic factor to the regulated secretory pathway. J Neurosci 2005, 25, 6156–6166. [Google Scholar] [CrossRef] [PubMed]
- Ho, I.H.T.; Liu, X.; Zou, Y.; Liu, T.; Hu, W.; Chan, H.; Tian, Y.; Zhang, Y.; Li, Q.; Kou, S.; et al. A Novel Peptide Interfering with proBDNF-Sortilin Interaction Alleviates Chronic Inflammatory Pain. Theranostics 2019, 9, 1651–1665. [Google Scholar] [CrossRef]
- Yang, C.R.; Ding, H.J.; Yu, M.; Zhou, F.H.; Han, C.Y.; Liang, R.; Zhang, X.Y.; Zhang, X.L.; Meng, F.J.; Wang, S.; et al. proBDNF/p75NTR promotes rheumatoid arthritis and inflammatory response by activating proinflammatory cytokines. Faseb j 2022, 36, e22180. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Liu, T.; Sun, J.; Zhao, S. Up-Regulation of ProBDNF/p75(NTR) Signaling in Spinal Cord Drives Inflammatory Pain in Male Rats. 2023, 16, 95–107. [CrossRef]
- Theodosiou, M.; Rush, A.R.; Zhou, F.X.; Hu, D.; Walker, S.J.; Tracey, J.D. Hyperalgesia due to nerve damage: role of nerve growth factor. Pain 1999, 81, 245–255. [Google Scholar] [CrossRef] [PubMed]
- Yajima, Y.; Narita, M.; Narita, M.; Matsumoto, N.; Suzuki, T. Involvement of a spinal brain-derived neurotrophic factor/full-length TrkB pathway in the development of nerve injury-induced thermal hyperalgesia in mice. Brain Res. 2002, 958, 338–346. [Google Scholar] [CrossRef] [PubMed]
- Yajima, Y.; Narita, M.; Usui, A.; Kaneko, C.; Miyatake, M. Direct evidence for the involvement of brain-derived neurotrophic factor in the development of a neuropathic pain-like state in mice. J.Neurochem. 2005, 93, 584. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.F.; Deng, Y.S.; Xian, C.J.; Zhong, J.H. Neurotrophins from dorsal root ganglia trigger allodynia after spinal nerve injury in rats. Eur.J.Neurosci. 2000, 12, 100. [Google Scholar] [CrossRef] [PubMed]
- Geng, S.J.; Liao, F.F.; Dang, W.H.; Ding, X.; Liu, X.D.; Cai, J.; Han, J.S.; Wan, Y.; Xing, G.G. Contribution of the spinal cord BDNF to the development of neuropathic pain by activation of the NR2B-containing NMDA receptors in rats with spinal nerve ligation. Exp Neurol 2010, 222, 256–266. [Google Scholar] [CrossRef] [PubMed]
- Miletic, G.; Miletic, V. Increases in the concentration of brain derived neurotrophic factor in the lumbar spinal dorsal horn are associated with pain behavior following chronic constriction injury in rats. Neurosci.Lett. 2002, 319, 137–140. [Google Scholar] [CrossRef] [PubMed]
- Obata, K.; Yamanaka, H.; Kobayashi, K.; Dai, Y.; Mizushima, T.; Katsura, H.; Fukuoka, T.; Tokunaga, A.; Noguchi, K. Role of mitogen-activated protein kinase activation in injured and intact primary afferent neurons for mechanical and heat hypersensitivity after spinal nerve ligation. J Neurosci 2004, 24, 10211–10222. [Google Scholar] [CrossRef]
- Obata, K.; Noguchi, K. BDNF in sensory neurons and chronic pain. Neurosci.Res. 2006, 55, 1–10. [Google Scholar] [CrossRef]
- Obata, K.; Yamanaka, H.; Kobayashi, K.; Dai, Y.; Mizushima, T.; Katsura, H.; Fukuoka, T.; Tokunaga, A.; Noguchi, K. The effect of site and type of nerve injury on the expression of brain-derived neurotrophic factor in the dorsal root ganglion and on neuropathic pain behavior. Neuroscience 2006, 137, 961–970. [Google Scholar] [CrossRef]
- Vanelderen, P.; Rouwette, T.; Kozicz, T.; Roubos, E.; Van Zundert, J.; Heylen, R.; Vissers, K. The role of brain-derived neurotrophic factor in different animal models of neuropathic pain. Eur J Pain 2010, 14, 473.e471–479. [Google Scholar] [CrossRef]
- Walker, S.M.; Mitchell, V.A.; White, D.M.; Rush, R.A.; Duggan, A.W. Release of immunoreactive brain-derived neurotrophic factor in the spinal cord of the rat following sciatic nerve transection. Brain Res 2001, 899, 240–247. [Google Scholar] [CrossRef]
- Ding, X.; Cai, J.; Li, S.; Liu, X.D.; Wan, Y.; Xing, G.G. BDNF contributes to the development of neuropathic pain by induction of spinal long-term potentiation via SHP2 associated GluN2B-containing NMDA receptors activation in rats with spinal nerve ligation. Neurobiol Dis 2015, 73, 428–451. [Google Scholar] [CrossRef]
- Hildebrand, M.E.; Xu, J.; Dedek, A.; Li, Y.; Sengar, A.S.; Beggs, S.; Lombroso, P.J.; Salter, M.W. Potentiation of Synaptic GluN2B NMDAR Currents by Fyn Kinase Is Gated through BDNF-Mediated Disinhibition in Spinal Pain Processing. Cell Reports 2016, 17, 2753–2765. [Google Scholar] [CrossRef]
- Dedek, A.; Xu, J.; Kandegedara, C.M.; Lorenzo, L.; Godin, A.G.; De Koninck, Y.; Lombroso, P.J.; Tsai, E.C.; Hildebrand, M.E. Loss of STEP61 couples disinhibition to N-methyl-d-aspartate receptor potentiation in rodent and human spinal pain processing. Brain 2019, 142, 1535–1546. [Google Scholar] [CrossRef]
- Chen, J.T.; Guo, D.; Campanelli, D.; Frattini, F.; Mayer, F.; Zhou, L.; Kuner, R.; Heppenstall, P.A.; Knipper, M.; Hu, J. Presynaptic GABAergic inhibition regulated by BDNF contributes to neuropathic pain induction. Nat Commun 2014, 5, 5331. [Google Scholar] [CrossRef] [PubMed]
- Lai, C.Y.; Hsieh, M.C.; Yeh, C.M.; Yang, P.S.; Cheng, J.K.; Wang, H.H.; Lin, K.H.; Nie, S.T.; Lin, T.B.; Peng, H.Y. MicroRNA-489-3p attenuates neuropathic allodynia by regulating oncoprotein DEK/TET1-dependent epigenetic modification in the dorsal horn. Neuropharmacology 2022, 210, 109028. [Google Scholar] [CrossRef] [PubMed]
- Phạm, T.L.; Noh, C.; Neupane, C.; Sharma, R.; Shin, H.J.; Park, K.D.; Lee, C.J.; Kim, H.W.; Lee, S.Y.; Park, J.B. MAO-B Inhibitor, KDS2010, Alleviates Spinal Nerve Ligation-induced Neuropathic Pain in Rats Through Competitively Blocking the BDNF/TrkB/NR2B Signaling. J Pain 2022, 23, 2092–2109. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Li, X.; Guo, H.; Liu, P.; Ma, M.; Wang, Y. Resolvin D1 attenuates mechanical allodynia after burn injury: Involvement of spinal glia, p38 mitogen-activated protein kinase, and brain-derived neurotrophic factor/tropomyosin-related kinase B signaling. Mol Pain 2023, 19, 17448069231159970. [Google Scholar] [CrossRef]
- Zhang, Y.Y.; Liu, F.; Fang, Z.H.; Li, Y.L.; Liao, H.L.; Song, Q.X.; Zhou, C.; Shen, J.F. Differential roles of NMDAR subunits 2A and 2B in mediating peripheral and central sensitization contributing to orofacial neuropathic pain. Brain, behavior, and immunity 2022, 106, 129–146. [Google Scholar] [CrossRef]
- Finamor, F.; Scarabelot, V.L.; Medeiros, L.F.; Stein, D.J.; da Silva, M.D.; Callai, E.; Caumo, W.; de Souza, A.; Torres, I.L.S. Involvement of GABAergic, glutamatergic, opioidergic, and brain-derived neurotrophic factor systems in the trigeminal neuropathic pain process. Neuroscience Letters 2023, 793, 136970. [Google Scholar] [CrossRef]
- de Oliveira, C.L.; Medeiros, L.F.; de Souza, V.S.; Lopes, B.C.; de Oliveira, F.F.; Marques, L.X.; da Silva Torres, I.L.; de Souza, A. LOW-DOSE NALTREXONE REVERSES FACIAL MECHANICAL ALLODYNIA IN A RAT MODEL OF TRIGEMINAL NEURALGIA. Neurosci Lett 2020, 736, 135248. [Google Scholar] [CrossRef] [PubMed]
- Wen, J.; Xu, Y.; Yu, Z.; Zhou, Y.; Wang, W.; Yang, J.; Wang, Y.; Bai, Q.; Li, Z. The cAMP Response Element- Binding Protein/Brain-Derived Neurotrophic Factor Pathway in Anterior Cingulate Cortex Regulates Neuropathic Pain and Anxiodepression Like Behaviors in Rats. Front Mol Neurosci 2022, 15, 831151. [Google Scholar] [CrossRef] [PubMed]
- Thakkar, B.; Acevedo, E.O. BDNF as a biomarker for neuropathic pain: Consideration of mechanisms of action and associated measurement challenges. Brain Behav 2023, 13, e2903. [Google Scholar] [CrossRef] [PubMed]
- Malcangio, M. Microglia and chronic pain. PAIN 2016, 157, 1002–1003. [Google Scholar] [CrossRef] [PubMed]
- Ji, R.-R.; Berta, T.; Nedergaard, M. Glia and pain: Is chronic pain a gliopathy? PAIN 2013, 154, S10–S28. [Google Scholar] [CrossRef] [PubMed]
- Apfel, S.C.; Wright, D.E.; Wiideman, A.M.; Dormia, C.; Snider, W.D.; Kessler, J.A. Nerve growth factor regulates the expression of brain-derived neurotrophic factor mRNA in the peripheral nervous system. Mol.Cell Neurosci. 1996, 7, 134–142. [Google Scholar] [CrossRef]
- Coull, J.A.; Beggs, S.; Boudreau, D.; Boivin, D.; Tsuda, M. BDNF from microglia causes the shift in neuronal anion gradient underlying neuropathic pain. Nature 2005, 438, 1017. [Google Scholar] [CrossRef]
- Ulmann, L.; Hatcher, J.P.; Hughes, J.P.; Chaumont, S.; Green, P.J.; Conquet, F.; Buell, G.N.; Reeve, A.J.; Chessell, I.P.; Rassendren, F. Up-regulation of P2X4 receptors in spinal microglia after peripheral nerve injury mediates BDNF release and neuropathic pain. J Neurosci 2008, 28, 11263–11268. [Google Scholar] [CrossRef]
- Boakye, P.A.; Tang, S.J.; Smith, P.A. Mediators of Neuropathic Pain; Focus on Spinal Microglia, CSF-1, BDNF, CCL21, TNF-α, Wnt Ligands, and Interleukin 1β. Front Pain Res (Lausanne) 2021, 2, 698157. [Google Scholar] [CrossRef]
- Sofroniew, M.V.; Vinters, H.V. Astrocytes: biology and pathology. Acta Neuropathol 2010, 119, 7–35. [Google Scholar] [CrossRef]
- Albini, M.; Krawczun-Rygmaczewska, A.; Cesca, F. Astrocytes and brain-derived neurotrophic factor (BDNF). Neuroscience Research 2023, 197, 42–51. [Google Scholar] [CrossRef]
- Jang, M.; Gould, E.; Xu, J.; Kim, E.J.; Kim, J.H. Oligodendrocytes regulate presynaptic properties and neurotransmission through BDNF signaling in the mouse brainstem. 2019, 8. [CrossRef]
- Atta, A.A.; Ibrahim, W.W.; Mohamed, A.F.; Abdelkader, N.F. Microglia polarization in nociplastic pain: mechanisms and perspectives. Inflammopharmacology 2023, 31, 1053–1067. [Google Scholar] [CrossRef]
- Malcangio, M. Role of the immune system in neuropathic pain. Scandinavian Journal of Pain 2020, 20, 33–37. [Google Scholar] [CrossRef]
- Trang, T.; Beggs, S.; Wan, X.; Salter, M.W. P2X4-receptor-mediated synthesis and release of brain-derived neurotrophic factor in microglia is dependent on calcium and p38-mitogen-activated protein kinase activation. J Neurosci 2009, 29, 3518–3528. [Google Scholar] [CrossRef] [PubMed]
- Bonalume, V.; Caffino, L.; Castelnovo, L.F.; Faroni, A.; Giavarini, F.; Liu, S.; Caruso, D.; Schmelz, M.; Fumagalli, F.; Carr, R.W.; et al. Schwann Cell Autocrine and Paracrine Regulatory Mechanisms, Mediated by Allopregnanolone and BDNF, Modulate PKCε in Peripheral Sensory Neurons. Cells 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Ismail, C.A.N.; Suppian, R.; Ab Aziz, C.B.; Long, I. Ifenprodil Reduced Expression of Activated Microglia, BDNF and DREAM Proteins in the Spinal Cord Following Formalin Injection During the Early Stage of Painful Diabetic Neuropathy in Rats. Journal of Molecular Neuroscience 2021, 71, 379–393. [Google Scholar] [CrossRef] [PubMed]
- Wong, L.; Done, J.D.; Schaeffer, A.J.; Thumbikat, P. Experimental autoimmune prostatitis induces microglial activation in the spinal cord. The Prostate 2015, 75, 50–59. [Google Scholar] [CrossRef]
- Zhou, T.-t.; Wu, J.-r.; Chen, Z.-y.; Liu, Z.-x.; Miao, B. Effects of dexmedetomidine on P2X4Rs, p38-MAPK and BDNF in spinal microglia in rats with spared nerve injury. Brain Research 2014, 1568, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Jin, J.; Chen, K.; You, S.; Zhang, H.; Sideris, A.; Norcini, M.; Recio-Pinto, E.; Wang, J.; Gan, W.B.; et al. BDNF produced by cerebral microglia promotes cortical plasticity and pain hypersensitivity after peripheral nerve injury. PLoS Biol 2021, 19, e3001337. [Google Scholar] [CrossRef] [PubMed]
- Harty, B.L.; Monk, K.R. Unwrapping the unappreciated: recent progress in Remak Schwann cell biology. Curr Opin Neurobiol 2017, 47, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Su, W.F.; Wu, F.; Jin, Z.H.; Gu, Y.; Chen, Y.T.; Fei, Y.; Chen, H.; Wang, Y.X.; Xing, L.Y.; Zhao, Y.Y.; et al. Overexpression of P2X4 receptor in Schwann cells promotes motor and sensory functional recovery and remyelination via BDNF secretion after nerve injury. Glia 2019, 67, 78–90. [Google Scholar] [CrossRef]
- Velázquez, K.T.; Mohammad, H.; Sweitzer, S.M. Protein kinase C in pain: involvement of multiple isoforms. Pharmacol Res 2007, 55, 578–589. [Google Scholar] [CrossRef] [PubMed]
- Liao, J.-Y.; Zhou, T.-H.; Chen, B.-K.; Liu, Z.-X. Schwann cells and trigeminal neuralgia. Molecular Pain 2020, 16, 1744806920963809. [Google Scholar] [CrossRef] [PubMed]
- Crotti, A.; Ransohoff, R.M. Microglial Physiology and Pathophysiology: Insights from Genome-wide Transcriptional Profiling. Immunity 2016, 44, 505–515. [Google Scholar] [CrossRef]
- Chen, G.; Zhang, Y.Q.; Qadri, Y.J.; Serhan, C.N.; Ji, R.R. Microglia in Pain: Detrimental and Protective Roles in Pathogenesis and Resolution of Pain. Neuron 2018, 100, 1292–1311. [Google Scholar] [CrossRef] [PubMed]
- Gruber-Schoffnegger, D.; Drdla-Schutting, R.; Hönigsperger, C.; Wunderbaldinger, G.; Gassner, M.; Sandkühler, J. Induction of thermal hyperalgesia and synaptic long-term potentiation in the spinal cord lamina I by TNF-α and IL-1β is mediated by glial cells. J Neurosci 2013, 33, 6540–6551. [Google Scholar] [CrossRef] [PubMed]
- Decosterd, I.; Ji, R.-R.; Suter, M.R.; Wen, Y.-R. Do glial cells control pain? Neuron Glia Biology 2007, 3, 255–268. [Google Scholar] [CrossRef]
- Ferrini, F.; De Koninck, Y. Microglia control neuronal network excitability via BDNF signalling. Neural Plast 2013, 2013, 429815. [Google Scholar] [CrossRef] [PubMed]
- Boakye, P.A.; Rancic, V.; Whitlock, K.H.; Simmons, D.; Longo, F.M.; Ballanyi, K.; Smith, P.A. Receptor dependence of BDNF actions in superficial dorsal horn: relation to central sensitization and actions of macrophage colony stimulating factor 1. J Neurophysiol 2019, 121, 2308–2322. [Google Scholar] [CrossRef]
- Lu, V.B.; Ballanyi, K.; Colmers, W.F.; Smith, P.A. Neuron type-specific effects of brain-derived neurotrophic factor in rat superficial dorsal horn and their relevance to ‘central sensitization’. J Physiol 2007, 584, 543–563. [Google Scholar] [CrossRef]
- Lu, V.B.; Biggs, J.E.; Stebbing, M.J.; Balasubramanyan, S.; Todd, K.G.; Lai, A.Y.; Colmers, W.F.; Dawbarn, D.; Ballanyi, K.; Smith, P.A. Brain-derived neurotrophic factor drives the changes in excitatory synaptic transmission in the rat superficial dorsal horn that follow sciatic nerve injury. J Physiol 2009, 587, 1013–1032. [Google Scholar] [CrossRef] [PubMed]
- Baba, H.; Ji, R.R.; Kohno, T.; Moore, K.A.; Ataka, T.; Wakai, A.; Okamoto, M.; Woolf, C.J. Removal of GABAergic inhibition facilitates polysynaptic A fiber-mediated excitatory transmission to the superficial spinal dorsal horn. Molecular and Cellular Neuroscience 2003, 24, 818–830. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Yu, X.; Chen, P.; Jin, K.; Zhou, J.; Wang, G.; Yu, J.; Wu, T.; Wang, Y.; Lin, F.; et al. BDNF-TrkB signaling pathway-mediated microglial activation induces neuronal KCC2 downregulation contributing to dynamic allodynia following spared nerve injury. Mol Pain 2023, 19, 17448069231185439. [Google Scholar] [CrossRef] [PubMed]
- Zarei, M.; Sabetkasaei, M.; Moini Zanjani, T.; Sahebi Vaighan, N. The effect of microglial inhibition on the expression of BDNF, KCC2, and GABAA receptor before and after the establishment of CCI-induced neuropathic pain model. Fundamental & Clinical Pharmacology 2022, 36, 277–285. [Google Scholar]
- Zhou, L.J.; Peng, J.; Xu, Y.N.; Zeng, W.J.; Zhang, J.; Wei, X.; Mai, C.L.; Lin, Z.J.; Liu, Y.; Murugan, M.; et al. Microglia Are Indispensable for Synaptic Plasticity in the Spinal Dorsal Horn and Chronic Pain. Cell Rep 2019, 27, 3844–3859.e3846. [Google Scholar] [CrossRef]
- Mapplebeck, J.C.S.; Beggs, S.; Salter, M.W. Sex differences in pain: a tale of two immune cells. PAIN 2016, 157, S2–S6. [Google Scholar] [CrossRef]
- Paige, C.; Plasencia-Fernandez, I.; Kume, M.; Papalampropoulou-Tsiridou, M.; Lorenzo, L.E.; David, E.T.; He, L.; Mejia, G.L.; Driskill, C.; Ferrini, F.; et al. A Female-Specific Role for Calcitonin Gene-Related Peptide (CGRP) in Rodent Pain Models. J Neurosci 2022, 42, 1930–1944. [Google Scholar] [CrossRef]
- Gardell, L.R.; Vanderah, T.W.; Gardell, S.E.; Wang, R.; Ossipov, M.H.; Lai, J.; Porreca, F. Enhanced Evoked Excitatory Transmitter Release in Experimental Neuropathy Requires Descending Facilitation. The Journal of Neuroscience 2003, 23, 8370–8379. [Google Scholar] [CrossRef]
- Ding, H.; Chen, J.; Su, M.; Lin, Z.; Zhan, H.; Yang, F.; Li, W.; Xie, J.; Huang, Y.; Liu, X.; et al. BDNF promotes activation of astrocytes and microglia contributing to neuroinflammation and mechanical allodynia in cyclophosphamide-induced cystitis. J Neuroinflammation 2020, 17, 19. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Liu, L.; Xu, D.; Zhang, W.; Zhang, Y.; Zhou, J.; Huang, W. Interaction between astrocytic colony stimulating factor and its receptor on microglia mediates central sensitization and behavioral hypersensitivity in chronic post ischemic pain model. Brain, behavior, and immunity 2018, 68, 248–260. [Google Scholar] [CrossRef] [PubMed]
- Guan, Z.; Kuhn, J.A.; Wang, X.; Colquitt, B.; Solorzano, C.; Vaman, S.; Guan, A.K.; Evans-Reinsch, Z.; Braz, J.; Devor, M.; et al. Injured sensory neuron-derived CSF1 induces microglial proliferation and DAP12-dependent pain. 2016, 19, 94–101. [CrossRef]
- Zhang, W.; Shi, Y.; Peng, Y.; Zhong, L.; Zhu, S.; Zhang, W.; Tang, S.J. Neuron activity-induced Wnt signaling up-regulates expression of brain-derived neurotrophic factor in the pain neural circuit. 2018, 293, 15641–15651. [CrossRef]
- Charles, A.C.; Merrill, J.E.; Dirksen, E.R.; Sanderson, M.J. Intercellular signaling in glial cells: calcium waves and oscillations in response to mechanical stimulation and glutamate. Neuron 1991, 6, 983–992. [Google Scholar] [CrossRef]
- Nedergaard, M.; Ransom, B.; Goldman, S.A. New roles for astrocytes: redefining the functional architecture of the brain. Trends Neurosci 2003, 26, 523–530. [Google Scholar] [CrossRef]
- Zafra, F.; Lindholm, D.; Castren, E.; Hartikka, J.; Thoenen, H. Regulation of brain-derived neurotrophic factor and nerve growth factor mRNA in primary cultures of hippocampal neurons and astrocytes. The Journal of Neuroscience 1992, 12, 4793–4799. [Google Scholar] [CrossRef]
- Schwartz, J.P.; Sheng, J.G.; Mitsuo, K.; Shirabe, S.; Nishiyama, N. Trophic factor production by reactive astrocytes in injured brain. Ann.NY Acad.Sci. 1993, 679, 226–234. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, J.P.; Nishiyama, N. Neurotrophic factor gene expression in astrocytes during development and following injury. Brain Research Bulletin 1994, 35, 403–407. [Google Scholar] [CrossRef]
- Alderson, R.F.; Curtis, R.; Alterman, A.L.; Lindsay, R.M.; DiStefano, P.S. Truncated TrkB mediates the endocytosis and release of BDNF and neurotrophin-4/5 by rat astrocytes and Schwann cells in vitro. Brain Research 2000, 871, 210–222. [Google Scholar] [CrossRef]
- Cao, T.; Matyas, J.J.; Renn, C.L. Function and Mechanisms of Truncated BDNF Receptor TrkB. T1 in Neuropathic Pain. 2020, 9. [Google Scholar] [CrossRef]
- Kinboshi, M.; Mukai, T.; Nagao, Y.; Matsuba, Y.; Tsuji, Y.; Tanaka, S.; Tokudome, K.; Shimizu, S.; Ito, H.; Ikeda, A.; et al. Inhibition of Inwardly Rectifying Potassium (Kir) 4.1 Channels Facilitates Brain-Derived Neurotrophic Factor (BDNF) Expression in Astrocytes. Frontiers in molecular neuroscience 2017, 10. [Google Scholar] [CrossRef] [PubMed]
- Milligan, E.D.; Watkins, L.R. Pathological and protective roles of glia in chronic pain. Nat Rev Neurosci 2009, 10, 23–36. [Google Scholar] [CrossRef]
- Renn, C.L.; Leitch, C.C.; Dorsey, S.G. In Vivo Evidence that Truncated Trkb.T1 Participates in Nociception. Molecular Pain 2009, 5, 1744–8069–1745-1761. [Google Scholar] [CrossRef]
- Wu, J.; Renn, C.L.; Faden, A.I.; Dorsey, S.G. TrkB.T1 contributes to neuropathic pain after spinal cord injury through regulation of cell cycle pathways. J Neurosci 2013, 33, 12447–12463. [Google Scholar] [CrossRef]
- Matyas, J.J.; O’Driscoll, C.M.; Yu, L. Truncated TrkB.T1-Mediated Astrocyte Dysfunction Contributes to Impaired Motor Function and Neuropathic Pain after Spinal Cord Injury. 2017, 37, 3956–3971. [Google Scholar] [CrossRef]
- Yang, X.-L.; Wang, X.; Shao, L.; Jiang, G.-T.; Min, J.-W.; Mei, X.-Y.; He, X.-H.; Liu, W.-H.; Huang, W.-X.; Peng, B.-W. TRPV1 mediates astrocyte activation and interleukin-1β release induced by hypoxic ischemia (HI). Journal of Neuroinflammation 2019, 16, 114. [Google Scholar] [CrossRef]
- Bradl, M.; Lassmann, H. Oligodendrocytes: biology and pathology. Acta Neuropathol 2010, 119, 37–53. [Google Scholar] [CrossRef]
- Dougherty, K.D.; Dreyfus, C.F.; Black, I.B. Brain-derived neurotrophic factor in astrocytes, oligodendrocytes, and microglia/macrophages after spinal cord injury. Neurobiol Dis 2000, 7, 574–585. [Google Scholar] [CrossRef]
- Malta, I.; Moraes, T.; Rodrigues, G.; Franco, P.; Galdino, G. The role of oligodendrocytes in chronic pain: cellular and molecular mechanisms. Journal of physiology and pharmacology: An official journal of the Polish Physiological Society 2019, 70. [Google Scholar] [CrossRef]
- Ren, K.; Dubner, R. Interactions between the immune and nervous systems in pain. Nat Med 2010, 16, 1267–1276. [Google Scholar] [CrossRef]
- Nakahashi, T.; Fujimura, H.; Altar, C.A.; Li, J.; Kambayashi, J.; Tandon, N.N.; Sun, B. Vascular endothelial cells synthesize and secrete brain-derived neurotrophic factor. FEBS Lett 2000, 470, 113–117. [Google Scholar] [CrossRef] [PubMed]
- Smith, P.A. Neuropathic pain; what we know and what we should do about it. Frontiers in pain research (Lausanne, Switzerland) 2023, 4, 1220034. [CrossRef]
- Dimmek, D.J.; Korallus, C.; Buyny, S.; Christoph, G.; Lichtinghagen, R.; Jacobs, R. Brain-Derived Neurotrophic Factor and Immune Cells in Osteoarthritis, Chronic Low Back Pain, and Chronic Widespread Pain Patients: Association with Anxiety and Depression. 2021, 57. [CrossRef]
- Yang, C.-R.; Ding, H.-J.; Yu, M.; Zhou, F.-H.; Han, C.-Y.; Liang, R.; Zhang, X.-Y.; Zhang, X.-L.; Meng, F.-J.; Wang, S.; et al. proBDNF/p75NTR promotes rheumatoid arthritis and inflammatory response by activating proinflammatory cytokines. The FASEB Journal 2022, 36, e22180. [Google Scholar] [CrossRef] [PubMed]
- Staszkiewicz, R.; Gładysz, D.; Gralewski, M.; Garczarek, M.; Gadzieliński, M.; Grabarek, B.O. Pathomechanism of the IVDs Degeneration and the Role of Neurotrophic Factors and Concentration of Selected Elements in Genesis of Low Back Pain. Curr Pharm Biotechnol 2023, 24, 1164–1177. [Google Scholar] [CrossRef] [PubMed]
- Lisiewski, L.E.; Jacobsen, H.E.; Viola, D.C.M.; Kenawy, H.M.; Kiridly, D.N.; Chahine, N.O. Intradiscal inflammatory stimulation induces spinal pain behavior and intervertebral disc degeneration in vivo. The FASEB Journal 2024, 38, e23364. [Google Scholar] [CrossRef] [PubMed]
- Gheorghe, R.O.; Grosu, A.V.; Magercu, M.; Ghenghea, M.S.; Zbarcea, C.E.; Tanase, A.; Negres, S.; Filippi, A.; Chiritoiu, G.; Gherghiceanu, M.; et al. Switching Rat Resident Macrophages from M1 to M2 Phenotype by Iba1 Silencing Has Analgesic Effects in SNL-Induced Neuropathic Pain. Int J Mol Sci 2023, 24. [Google Scholar] [CrossRef] [PubMed]
- Tu, Y.; Muley, M.M.; Beggs, S.; Salter, M.W. Microglia-independent peripheral neuropathic pain in male and female mice. Pain 2022, 163, e1129–e1144. [Google Scholar] [CrossRef]
- Buskila, D. Genetics of chronic pain states. Best practice & research clinical rheumatology 2007, 21, 535–547. [Google Scholar]
- Fillingim, R.; Wallace, M.; Herbstman, D.; Ribeiro-Dasilva, M.; Staud, R. Genetic contributions to pain: a review of findings in humans. Oral Diseases 2008, 14, 673–682. [Google Scholar] [CrossRef] [PubMed]
- Cappoli, N.; Tabolacci, E.; Aceto, P.; Dello Russo, C. The emerging role of the BDNF-TrkB signaling pathway in the modulation of pain perception. Journal of neuroimmunology 2020, 349, 577406. [Google Scholar] [CrossRef] [PubMed]
- Egan, M.F.; Kojima, M.; Callicott, J.H.; Goldberg, T.E.; Kolachana, B.S.; Bertolino, A.; Zaitsev, E.; Gold, B.; Goldman, D.; Dean, M.; et al. The BDNF val66met polymorphism affects activity-dependent secretion of BDNF and human memory and hippocampal function. Cell 2003, 112, 257–269. [Google Scholar] [CrossRef]
- Hempstead, B.L. Brain-Derived Neurotrophic Factor: Three Ligands, Many Actions. Transactions of the American Clinical and Climatological Association 2015, 126, 9–19. [Google Scholar]
- Tian, Y.; Liu, X.; Jia, M.; Yu, H.; Lichtner, P.; Shi, Y.; Meng, Z.; Kou, S.; Ho, I.H.T.; Jia, B.; et al. Targeted Genotyping Identifies Susceptibility Locus in Brain-derived Neurotrophic Factor Gene for Chronic Postsurgical Pain. Anesthesiology 2018, 128, 587–597. [Google Scholar] [CrossRef]
- Vossen, H.; Kenis, G.; Rutten, B.; van Os, J.; Hermens, H.; Lousberg, R. The Genetic Influence on the Cortical Processing of Experimental Pain and the Moderating Effect of Pain Status. PLOS ONE 2010, 5, e13641. [Google Scholar] [CrossRef]
- Polli, A.; Ghosh, M.; Bakusic, J.; Ickmans, K.; Monteyne, D.; Velkeniers, B.; Bekaert, B.; Godderis, L.; Nijs, J. DNA Methylation and Brain-Derived Neurotrophic Factor Expression Account for Symptoms and Widespread Hyperalgesia in Patients With Chronic Fatigue Syndrome and Comorbid Fibromyalgia. Arthritis & Rheumatology 2020, 72, 1936–1944. [Google Scholar]
- Chau, C.M.; Cepeda, I.L.; Devlin, A.M.; Weinberg, J.; Grunau, R.E. The Val66Met brain-derived neurotrophic factor gene variant interacts with early pain exposure to predict cortisol dysregulation in 7-year-old children born very preterm: Implications for cognition. Neuroscience 2017, 342, 188–199. [Google Scholar] [CrossRef]
- Yu, H.; Wang, D.D.; Wang, Y.; Liu, T.; Lee, F.S.; Chen, Z.Y. Variant brain-derived neurotrophic factor Val66Met polymorphism alters vulnerability to stress and response to antidepressants. J.Neurosci. 2012, 32, 4092–4101. [Google Scholar] [CrossRef]
- Gonçalves, F.T.; Pacheco-Barrios, K.; Rebello-Sanchez, I.; Castelo-Branco, L.; de Melo, P.S.; Parente, J.; Cardenas-Rojas, A.; Firigato, I.; Pessotto, A.V.; Imamura, M.; et al. Association of Mu opioid receptor (A118G) and BDNF (G196A) polymorphisms with rehabilitation-induced cortical inhibition and analgesic response in chronic osteoarthritis pain. International journal of clinical and health psychology: IJCHP 2023, 23, 100330. [Google Scholar] [CrossRef]
- Gonçalves, F.T.; Marques, L.M.; Pessotto, A.V.; Barbosa, S.P.; Imamura, M.; Simis, M.; Fregni, F.; Battistella, L. OPRM1 and BDNF polymorphisms associated with a compensatory neurophysiologic signature in knee osteoarthritis patients. Neurophysiologie clinique = Clinical neurophysiology 2023, 53, 102917. [Google Scholar] [CrossRef] [PubMed]
- Janssen, L.P.; Medeiros, L.F. Fibromyalgia: A Review of Related Polymorphisms and Clinical Relevance. 2021, 93, e20210618. [CrossRef]
- Behnoush, A.H.; Khalaji, A.; Khanmohammadi, S.; Alehossein, P.; Saeedian, B.; Shobeiri, P.; Teixeira, A.L.; Rezaei, N. Brain-derived neurotrophic factor in fibromyalgia: A systematic review and meta-analysis of its role as a potential biomarker. PloS One 2023, 18, e0296103. [Google Scholar] [CrossRef] [PubMed]
- Baumbauer, K.M.; Ramesh, D.; Perry, M.; Carney, K.B.; Julian, T.; Glidden, N.; Dorsey, S.G.; Starkweather, A.R.; Young, E.E. Contribution of COMT and BDNF Genotype and Expression to the Risk of Transition From Acute to Chronic Low Back Pain. The Clinical journal of pain 2020, 36, 430–439. [Google Scholar] [CrossRef] [PubMed]
- Yamada, A.S.; Antunes, F.T.T.; Ferraz, C.; de Souza, A.H.; Simon, D. The genetic influence of the brain-derived neurotrophic factor Val66Met polymorphism in chronic low back pain. Advances in rheumatology (London, England) 2021, 61, 24. [Google Scholar] [CrossRef] [PubMed]
- Goto, T.; Von Ah, D.; Li, X.; Xiang, L.; Kwiat, C.; Nguyen, C.; Hsiao, C.P.; Saligan, L.N. Brain-Derived Neurotrophic Factor rs6265 polymorphism is associated with severe cancer-related fatigue and neuropathic pain in female cancer survivors. Journal of cancer survivorship: Research and practice 2023. [CrossRef]
- Azoulay, D.; Abed, S.; Sfadi, A.; Sheleg, O.; Shaoul, E.; Shehadeh, M.; Kaykov, E.; Nodelman, M.; Bashkin, A. Low brain-derived neurotrophic factor protein levels and single-nucleotide polymorphism Val66Met are associated with peripheral neuropathy in type II diabetic patients. Acta diabetologica 2020, 57, 891–898. [Google Scholar] [CrossRef] [PubMed]
- Kandasamy, R.; Price, T.J. The pharmacology of nociceptor priming. Handbook of experimental pharmacology 2015, 227, 15–37. [Google Scholar] [CrossRef] [PubMed]
- Melemedjian, O.K.; Tillu, D.V.; Asiedu, M.N.; Mandell, E.K.; Moy, J.K.; Blute, V.M.; Taylor, C.J.; Ghosh, S.; Price, T.J. BDNF regulates atypical PKC at spinal synapses to initiate and maintain a centralized chronic pain state. Mol Pain 2013, 9, 12. [Google Scholar] [CrossRef] [PubMed]
- Gouin, O.; L’Herondelle, K.; Lebonvallet, N.; Le Gall-Ianotto, C.; Sakka, M.; Buhé, V.; Plée-Gautier, E.; Carré, J.-L.; Lefeuvre, L.; Misery, L.; et al. TRPV1 and TRPA1 in cutaneous neurogenic and chronic inflammation: pro-inflammatory response induced by their activation and their sensitization. Protein & Cell 2017, 8, 644–661. [Google Scholar] [CrossRef]
- Schmelz, M. Itch and pain. Neuroscience & Biobehavioral Reviews 2010, 34, 171–176. [Google Scholar]
- Pezet, S.; McMahon, S.B. Neurotrophins: mediators and modulators of pain. Annu. Rev. Neurosci. 2006, 29, 507–538. [Google Scholar] [CrossRef] [PubMed]
- Ohtori, S.; Takahashi, K.; Moriya, H. Inflammatory pain mediated by a phenotypic switch in brain-derived neurotrophic factor-immunoreactive dorsal root ganglion neurons innervating the lumbar facet joints in rats. Neurosci Lett 2002, 323, 129–132. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.F.; Chie, E.T.; Deng, Y.S.; Zhong, J.H.; Xue, Q.; Rush, R.A.; Xian, C.J. Injured primary sensory neurons switch phenotype for brain-derived neurotrophic factor in the rat. Neuroscience 1999, 92, 841–853. [Google Scholar] [CrossRef] [PubMed]
- Mannion, R.J.; Costigan, M.; Decosterd, I.; Amaya, F.; Ma, Q.P. Neurotrophins: peripherally and centrally acting modulators of tactile stimulus-induced inflammatory pain hypersensitivity. Proc.Natl.Acad.Sci.USA 1999, 96, 9385. [Google Scholar] [CrossRef] [PubMed]
- Distefano, P.S.; Friedman, B.; Radziejewski, C.; Alexander, C.; Boland, P.; Schick, C.M.; Lindsay, R.M.; Wiegand, S.J. The neurotrophins BDNF, NT-3, and NGF display distinct patterns of retrograde axonal transport in peripheral and central neurons. Neuron 1992, 8, 983–993. [Google Scholar] [CrossRef]
- Heppenstall, P.A.; Lewin, G.R. Neurotrophins, nociceptors and pain. Curr Opin Anaesthesiol 2000, 13, 573–576. [Google Scholar] [CrossRef]
- Park, S.Y.; Lee, J.Y.; Choi, J.Y.; Park, M.J.; Kim, D.S. Nerve growth factor activates brain-derived neurotrophic factor promoter IV via extracellular signal-regulated protein kinase 1/2 in PC12 cells. Mol Cells 2006, 21, 237–243. [Google Scholar] [CrossRef]
- Paterson, S.; Schmelz, M.; McGlone, F.; Turner, G.; Rukwied, R. Facilitated neurotrophin release in sensitized human skin. European Journal of Pain 2009, 13, 399–405. [Google Scholar] [CrossRef] [PubMed]
- Ren, K.; Dubner, R. Pain Facilitation and Activity-Dependent Plasticity in Pain Modulatory Circuitry: Role of BDNF-TrkB Signaling and NMDA Receptors. Mol.Neurobiol. 2007, 35, 224–235. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.-G.; Zhou, L.-J. Long-term Potentiation at Spinal C-fiber Synapses: A Target for Pathological Pain. Current Pharmaceutical Design 2015, 21, 895–905. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Cai, J.; Feng, Z.B.; Jin, Z.R.; Liu, B.H.; Zhao, H.Y.; Jing, H.B.; Wei, T.J.; Yang, G.N.; Liu, L.Y.; et al. BDNF Contributes to Spinal Long-Term Potentiation and Mechanical Hypersensitivity Via Fyn-Mediated Phosphorylation of NMDA Receptor GluN2B Subunit at Tyrosine 1472 in Rats Following Spinal Nerve Ligation. Neurochem Res 2017, 42, 2712–2729. [Google Scholar] [CrossRef] [PubMed]
- Sandkühler, J.; Gruber-Schoffnegger, D. Hyperalgesia by synaptic long-term potentiation (LTP): an update. Curr Opin Pharmacol 2012, 12, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.J.; Ren, W.J.; Zhong, Y.; Yang, T.; Wei, X.H.; Xin, W.J.; Liu, C.C.; Zhou, L.H.; Li, Y.Y.; Liu, X.G. Limited BDNF contributes to the failure of injury to skin afferents to produce a neuropathic pain condition. Pain 2010, 148, 148–157. [Google Scholar] [CrossRef]
- Slack, S.E.; Pezet, S.; McMahon, S.B.; Thompson, S.W.; Malcangio, M. Brain-derived neurotrophic factor induces NMDA receptor subunit one phosphorylation via ERK and PKC in the rat spinal cord. Eur.J Neurosci. 2004, 20, 1769–1778. [Google Scholar] [CrossRef]
- Zhou, L.J.; Zhong, Y.; Ren, W.J.; Li, Y.Y.; Zhang, T.; Liu, X.G. BDNF induces late-phase LTP of C-fiber evoked field potentials in rat spinal dorsal horn. Exp Neurol 2008, 212, 507–514. [Google Scholar] [CrossRef]
- Bao, Y.; Hou, W.; Liu, R.; Gao, Y.; Kong, X.; Yang, L.; Shi, Z.; Li, W.; Zheng, H.; Jiang, S.; et al. PAR2-mediated upregulation of BDNF contributes to central sensitization in bone cancer pain. Mol Pain 2014, 10, 28. [Google Scholar] [CrossRef]
- Zhou, L.J.; Yang, T.; Wei, X.; Liu, Y.; Xin, W.J.; Chen, Y.; Pang, R.P.; Zang, Y.; Li, Y.Y.; Liu, X.G. Brain-derived neurotrophic factor contributes to spinal long-term potentiation and mechanical hypersensitivity by activation of spinal microglia in rat. Brain Behav.Immun. 2011, 25, 322–334. [Google Scholar] [CrossRef]
- Retamal, J.; Reyes, A.; Ramirez, P.; Bravo, D.; Hernandez, A.; Pelissier, T.; Villanueva, L.; Constandil, L. Burst-Like Subcutaneous Electrical Stimulation Induces BDNF-Mediated, Cyclotraxin B-Sensitive Central Sensitization in Rat Spinal Cord. Front Pharmacol 2018, 9, 1143. [Google Scholar] [CrossRef]
- Rivera-Arconada, I.; Benedet, T.; Roza, C.; Torres, B.; Barrio, J.; Krzyzanowska, A.; Avendaño, C.; Mellström, B.; Lopez-Garcia, J.A.; Naranjo, J.R. DREAM regulates BDNF-dependent spinal sensitization. Mol Pain 2010, 6, 95. [Google Scholar] [CrossRef]
- Huang, Y.J.; Lee, K.H.; Grau, J.W. Complete spinal cord injury (SCI) transforms how brain derived neurotrophic factor (BDNF) affects nociceptive sensitization. Exp Neurol 2017, 288, 38–50. [Google Scholar] [CrossRef] [PubMed]
- Price, T.J.; Ghosh, S. ZIPping to pain relief: the role (or not) of PKMζ in chronic pain. Mol Pain 2013, 9, 6. [Google Scholar] [CrossRef] [PubMed]
- Sacktor, T.C. How does PKMζ maintain long-term memory? Nat Rev Neurosci 2011, 12, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Croll, S.D.; Chesnutt, C.R.; Rudge, J.S.; Acheson, A.; Ryan, T.E.; Siuciak, J.A.; DiStefano, P.S.; Wiegand, S.J.; Lindsay, R.M. Co-infusion with a TrkB-Fc receptor body carrier enhances BDNF distribution in the adult rat brain. Exp Neurol 1998, 152, 20–33. [Google Scholar] [CrossRef] [PubMed]
- Morgado, C.; Pereira-Terra, P.; Cruz, C.D.; Tavares, I. Minocycline completely reverses mechanical hyperalgesia in diabetic rats through microglia-induced changes in the expression of the potassium chloride co-transporter 2 (KCC2) at the spinal cord. Diabetes, obesity & metabolism 2011, 13, 150–159. [Google Scholar] [CrossRef]
- Moy, J.K.; Szabo-Pardi, T.; Tillu, D.V.; Megat, S.; Pradhan, G.; Kume, M.; Asiedu, M.N.; Burton, M.D.; Dussor, G.; Price, T.J. Temporal and sex differences in the role of BDNF/TrkB signaling in hyperalgesic priming in mice and rats. Neurobiology of pain (Cambridge, Mass.) 2019, 5, 100024. [Google Scholar] [CrossRef] [PubMed]
- Rosenthal, A.; Lin, J.C. Modulation of neurotrophin signaling by monoclonal antibodies. Handbook of experimental pharmacology 2014, 220, 497–512. [Google Scholar] [CrossRef]
- Cazorla, M.; Prémont, J.; Mann, A.; Girard, N.; Kellendonk, C.; Rognan, D. Identification of a low-molecular weight TrkB antagonist with anxiolytic and antidepressant activity in mice. J Clin Invest 2011, 121, 1846–1857. [Google Scholar] [CrossRef]
- Cazorla, M.; Arrang, J.M.; Prémont, J. Pharmacological characterization of six trkB antibodies reveals a novel class of functional agents for the study of the BDNF receptor. Br J Pharmacol 2011, 162, 947–960. [Google Scholar] [CrossRef] [PubMed]
- Ferrini, F.; Trang, T.; Mattioli, T.A.; Laffray, S.; Del’Guidice, T.; Lorenzo, L.E.; Castonguay, A.; Doyon, N.; Zhang, W.; Godin, A.G.; et al. Morphine hyperalgesia gated through microglia-mediated disruption of neuronal Cl⁻ homeostasis. Nat Neurosci 2013, 16, 183–192. [Google Scholar] [CrossRef] [PubMed]
- Cazorla, M.; Jouvenceau, A.; Rose, C.; Guilloux, J.P.; Pilon, C.; Dranovsky, A.; Prémont, J. Cyclotraxin-B, the first highly potent and selective TrkB inhibitor, has anxiolytic properties in mice. PLoS One 2010, 5, e9777. [Google Scholar] [CrossRef] [PubMed]
- Shih, H.C.; Kuan, Y.H.; Shyu, B.C. Targeting brain-derived neurotrophic factor in the medial thalamus for the treatment of central poststroke pain in a rodent model. Pain 2017, 158, 1302–1313. [Google Scholar] [CrossRef] [PubMed]
- Constandil, L.; Goich, M.; Hernández, A.; Bourgeais, L.; Cazorla, M.; Hamon, M.; Villanueva, L.; Pelissier, T. Cyclotraxin-B, a new TrkB antagonist, and glial blockade by propentofylline, equally prevent and reverse cold allodynia induced by BDNF or partial infraorbital nerve constriction in mice. J Pain 2012, 13, 579–589. [Google Scholar] [CrossRef] [PubMed]
- M’Dahoma, S.; Barthélemy, S.; Tromilin, C.; Jeanson, T.; Viguier, F.; Michot, B.; Pezet, S.; Hamon, M.; Bourgoin, S. Respective pharmacological features of neuropathic-like pain evoked by intrathecal BDNF versus sciatic nerve ligation in rats. European neuropsychopharmacology: The journal of the European College of Neuropsychopharmacology 2015, 25, 2118–2130. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.; Zhang, Y.W.; Gu, Q.; Yuan, T.J.; Fan, B.Q. Alteration of neural activity and neuroinflammatory factors in the insular cortex of mice with corneal neuropathic pain. Genes, brain, and behavior 2023, 22, e12842. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, T.; Colciago, A.; Montagnani Marelli, M.; Moretti, R.M.; Magnaghi, V. Protein kinase C epsilon activation regulates proliferation, migration, and epithelial to mesenchymal-like transition in rat Schwann cells. Front Cell Neurosci 2023, 17, 1237479. [Google Scholar] [CrossRef]
- Fan, F.; Tang, Y.; Dai, H.; Cao, Y.; Sun, P.; Chen, Y.; Chen, A.; Lin, C. Blockade of BDNF signalling attenuates chronic visceral hypersensitivity in an IBS-like rat model. European journal of pain (London, England) 2020, 24, 839–850. [Google Scholar] [CrossRef]
- Ma, Y.; Zhao, W.; Chen, D.; Zhou, D.; Gao, Y.; Bian, Y.; Xu, Y.; Xia, S.H. Disinhibition of Mesolimbic Dopamine Circuit by the Lateral Hypothalamus Regulates Pain Sensation. J Neurosci 2023, 43, 4525–4540. [Google Scholar] [CrossRef]
- Ye, Y.; Yan, X.; Wang, L.; Xu, J.; Li, T. Transcranial direct current stimulation attenuates chronic pain in knee osteoarthritis by modulating BDNF/TrkB signaling in the descending pain modulation system. Neurosci Lett 2023, 810, 137320. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Yang, Y.; Wang, C.; Huang, J.; Wang, X.; Liu, Y.; Wang, H. Exploring the role and mechanisms of diallyl trisulfide and diallyl disulfide in chronic constriction-induced neuropathic pain in rats. Korean J Pain 2020, 33, 216–225. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, K.; Lesnak, J.B.; Plumb, A.N.; Janowski, A.J.; Smith, A.F.; Hill, J.K.; Sluka, K.A. Brain-derived neurotrophic factor contributes to activity-induced muscle pain in male but not female mice. bioRxiv 2023. [Google Scholar] [CrossRef]
- Tang, Y.; Chen, L.; Liu, B.; Sun, P.; Chen, Z.; Huang, Y.; Ai-Qin, C.; Chen, Y.; Lin, C. Spinal P2X4 Receptors Involved in Visceral Hypersensitivity of Neonatal Maternal Separation Rats. 2023, 19, 113–122. [CrossRef]
- Hsiang, H.W.; Girard, B.M.; Ratkovits, L.; Campbell, S.E.; Vizzard, M.A. Effects of pharmacological neurotrophin receptor inhibition on bladder function in female mice with cyclophosphamide-induced cystitis. Frontiers in urology 2022, 2. [Google Scholar] [CrossRef] [PubMed]
- Long, T.; He, W.; Pan, Q.; Zhang, S.; Zhang, D.; Qin, G.; Chen, L.; Zhou, J. Microglia P2X4R-BDNF signalling contributes to central sensitization in a recurrent nitroglycerin-induced chronic migraine model. J Headache Pain 2020, 21, 4. [Google Scholar] [CrossRef] [PubMed]
- Chodroff, L.; Bendele, M.; Valenzuela, V.; Henry, M.; Ruparel, S. EXPRESS: BDNF Signaling Contributes to Oral Cancer Pain in a Preclinical Orthotopic Rodent Model. Mol Pain 2016, 12. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.M.; Pan, W.; Xu, N.; Zhou, Z.Q.; Zhang, G.F.; Shen, J.C. Environmental enrichment improves long-term memory impairment and aberrant synaptic plasticity by BDNF/TrkB signaling in nerve-injured mice. Neurosci Lett 2019, 694, 93–98. [Google Scholar] [CrossRef]
- Liang, D.Y.; Sun, Y.; Shi, X.Y.; Sahbaie, P.; Clark, J.D. Epigenetic regulation of spinal cord gene expression controls opioid-induced hyperalgesia. Mol Pain 2014, 10, 59. [Google Scholar] [CrossRef]
- Fang, X.; Yang, C.; Li, S.; Zhan, G.; Zhang, J.; Huang, N.; Du, X.; Xu, H.; Hashimoto, K.; Luo, A. Brain-derived neurotrophic factor-TrkB signaling in the medial prefrontal cortex plays a role in the anhedonia-like phenotype after spared nerve injury. European archives of psychiatry and clinical neuroscience 2020, 270, 195–205. [Google Scholar] [CrossRef]
- Sahbaie, P.; Liang, D.Y.; Shi, X.Y.; Sun, Y.; Clark, J.D. Epigenetic regulation of spinal cord gene expression contributes to enhanced postoperative pain and analgesic tolerance subsequent to continuous opioid exposure. Mol Pain 2016, 12. [Google Scholar] [CrossRef]
- Fu, Y.; Lin, Y.M.; Winston, J.H.; Radhakrishnan, R.; Huang, L.-H.M.; Shi, X.Z. Role of brain-derived neurotrophic factor in the pathogenesis of distention-associated abdominal pain in bowel obstruction. Neurogastroenterology and motility 2018, 30, e13373. [Google Scholar] [CrossRef]
- Tillu, D.V.; Hassler, S.N.; Burgos-Vega, C.C.; Quinn, T.L.; Sorge, R.E.; Dussor, G.; Boitano, S.; Vagner, J.; Price, T.J. Protease-activated receptor 2 activation is sufficient to induce the transition to a chronic pain state. Pain 2015, 156, 859–867. [Google Scholar] [CrossRef]
- Liu, Z.; Liu, F.; Liu, X.; Ma, C.; Zhao, J. Surgical incision induces learning impairment in mice partially through inhibition of the brain-derived neurotrophic factor signaling pathway in the hippocampus and amygdala. Mol Pain 2018, 14, 1744806918805902. [Google Scholar] [CrossRef]
- Yan, W.; Lakkaniga, N.R.; Carlomagno, F.; Santoro, M.; McDonald, N.Q.; Lv, F.; Gunaganti, N.; Frett, B.; Li, H.Y. Insights into Current Tropomyosin Receptor Kinase (TRK) Inhibitors: Development and Clinical Application. J Med Chem 2019, 62, 1731–1760. [Google Scholar] [CrossRef] [PubMed]
- Stachel, S.J.; Sanders, J.M.; Henze, D.A.; Rudd, M.T.; Su, H.P.; Li, Y.; Nanda, K.K.; Egbertson, M.S.; Manley, P.J.; Jones, K.L.; et al. Maximizing diversity from a kinase screen: identification of novel and selective pan-Trk inhibitors for chronic pain. J Med Chem 2014, 57, 5800–5816. [Google Scholar] [CrossRef]
- Angeles, T.S.; Yang, S.X.; Steffler, C.; Dionne, C.A. Kinetics of trkA tyrosine kinase activity and inhibition by K-252a. Arch Biochem Biophys 1998, 349, 267–274. [Google Scholar] [CrossRef]
- Winston, J.H.; Toma, H.; Shenoy, M.; He, Z.J.; Zou, L.; Xiao, S.Y.; Micci, M.A.; Pasricha, P.J. Acute pancreatitis results in referred mechanical hypersensitivity and neuropeptide up-regulation that can be suppressed by the protein kinase inhibitor k252a. J Pain 2003, 4, 329–337. [Google Scholar] [CrossRef] [PubMed]
- Frias, B.; Allen, S.; Dawbarn, D.; Charrua, A.; Cruz, F.; Cruz, C.D. Brain-derived neurotrophic factor, acting at the spinal cord level, participates in bladder hyperactivity and referred pain during chronic bladder inflammation. Neuroscience 2013, 234, 88–102. [Google Scholar] [CrossRef]
- Miletic, G.; Miletic, V. Loose ligation of the sciatic nerve is associated with TrkB receptor-dependent decreases in KCC2 protein levels in the ipsilateral spinal dorsal horn. Pain 2008, 137, 532–539. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.H.; Zhang, X.Q.; Xue, Q.S.; Yan, L.; Huang, J.L.; Zhang, S.; Shao, H.J.; Lu, H.; Wang, W.Y.; Yu, B.W. The BDNF/TrkB signaling pathway is involved in heat hyperalgesia mediated by Cdk5 in rats. PLoS One 2014, 9, e85536. [Google Scholar] [CrossRef]
- Yasui, M.; Shiraishi, Y.; Ozaki, N.; Hayashi, K.; Hori, K.; Ichiyanagi, M.; Sugiura, Y. Nerve growth factor and associated nerve sprouting contribute to local mechanical hyperalgesia in a rat model of bone injury. Eur J Pain 2012, 16, 953–965. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Wang, F.; Wang, L.; Xu, Y.; Lv, L.; Duan, W.; Bai, R.; Meng, Z.; Shao, X. Involvement of the BDNF-TrkB-KCC2 pathway in neuropathic pain after brachial plexus avulsion. Brain Behav 2022, 12, e2464. [Google Scholar] [CrossRef]
- Hiroki, T.; Suto, T.; Ohta, J.; Saito, S.; Obata, H. Spinal γ-Aminobutyric Acid Interneuron Plasticity Is Involved in the Reduced Analgesic Effects of Morphine on Neuropathic Pain. J Pain 2022, 23, 547–557. [Google Scholar] [CrossRef]
- Dos Reis, R.C.; Kopruszinski, C.M.; Nones, C.F.M.; Aguiar, D.A.; Chichorro, J.G. The opposing contribution of neurotrophin-3 and nerve growth factor to orofacial heat hyperalgesia in rats. Behav Pharmacol 2020, 31, 27–33. [Google Scholar] [CrossRef]
- Miao, B.; Yin, Y.; Mao, G.; Zhao, B.; Wu, J.; Shi, H.; Fei, S. The implication of transient receptor potential canonical 6 in BDNF-induced mechanical allodynia in rat model of diabetic neuropathic pain. Life Sci 2021, 273, 119308. [Google Scholar] [CrossRef] [PubMed]
- Bailey, J.J.; Jaworski, C.; Tung, D.; Wängler, C.; Wängler, B.; Schirrmacher, R. Tropomyosin receptor kinase inhibitors: an updated patent review for 2016-2019. Expert Opin Ther Pat 2020, 30, 325–339. [Google Scholar] [CrossRef] [PubMed]
- Bailey, J.J.; Schirrmacher, R.; Farrell, K.; Bernard-Gauthier, V. Tropomyosin receptor kinase inhibitors: an updated patent review for 2010-2016—Part II. Expert Opin Ther Pat 2017, 27, 831–849. [Google Scholar] [CrossRef]
- Bailey, J.J.; Schirrmacher, R.; Farrell, K.; Bernard-Gauthier, V. Tropomyosin receptor kinase inhibitors: an updated patent review for 2010-2016—Part I. Expert Opin Ther Pat 2017, 27, 733–751. [Google Scholar] [CrossRef]
- Wang, T.; Yu, D.; Lamb, M.L. Trk kinase inhibitors as new treatments for cancer and pain. Expert Opin Ther Pat 2009, 19, 305–319. [Google Scholar] [CrossRef]
- Ghilardi, J.R.; Freeman, K.T.; Jimenez-Andrade, J.M.; Mantyh, W.G.; Bloom, A.P.; Bouhana, K.S.; Trollinger, D.; Winkler, J.; Lee, P.; Andrews, S.W.; et al. Sustained blockade of neurotrophin receptors TrkA, TrkB and TrkC reduces non-malignant skeletal pain but not the maintenance of sensory and sympathetic nerve fibers. Bone 2011, 48, 389–398. [Google Scholar] [CrossRef]
- Skerratt, S.E.; Andrews, M.; Bagal, S.K.; Bilsland, J.; Brown, D.; Bungay, P.J.; Cole, S.; Gibson, K.R.; Jones, R.; Morao, I.; et al. The Discovery of a Potent, Selective, and Peripherally Restricted Pan-Trk Inhibitor (PF-06273340) for the Treatment of Pain. J Med Chem 2016, 59, 10084–10099. [Google Scholar] [CrossRef] [PubMed]
- Ishiguro, N.; Oyama, S.; Higashi, R.; Yanagida, K. Efficacy, Safety, and Tolerability of ONO-4474, an Orally Available Pan-Tropomyosin Receptor Kinase Inhibitor, in Japanese Patients With Moderate to Severe Osteoarthritis of the Knee: A Randomized, Placebo-Controlled, Double-Blind, Parallel-Group Comparative Study. J Clin Pharmacol 2020, 60, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Boulle, F.; Kenis, G.; Cazorla, M.; Hamon, M.; Steinbusch, H.W.; Lanfumey, L.; van den Hove, D.L. TrkB inhibition as a therapeutic target for CNS-related disorders. Prog Neurobiol 2012, 98, 197–206. [Google Scholar] [CrossRef]
- Beggs, S.; Trang, T.; Salter, M.W. P2X4R+ microglia drive neuropathic pain. Nat Neurosci 2012, 15, 1068–1073. [Google Scholar] [CrossRef] [PubMed]
- Doyon, N.; Ferrini, F.; Gagnon, M.; De Koninck, Y. Treating pathological pain: is KCC2 the key to the gate? Expert Rev Neurother 2013, 13, 469–471. [Google Scholar] [CrossRef] [PubMed]
- Gagnon, M.; Bergeron, M.J.; Lavertu, G.; Castonguay, A.; Tripathy, S.; Bonin, R.P.; Perez-Sanchez, J.; Boudreau, D.; Wang, B.; Dumas, L.; et al. Chloride extrusion enhancers as novel therapeutics for neurological diseases. Nat Med 2013, 19, 1524–1528. [Google Scholar] [CrossRef] [PubMed]
- Laske, C.; Stransky, E.; Eschweiler, G.W.; Klein, R.; Wittorf, A.; Leyhe, T.; Richartz, E.; Köhler, N.; Bartels, M.; Buchkremer, G.; et al. Increased BDNF serum concentration in fibromyalgia with or without depression or antidepressants. Journal of Psychiatric Research 2007, 41, 600–605. [Google Scholar] [CrossRef]
- de Zanette, S.A.; Vercelino, R.; Laste, G.; Rozisky, J.R.; Schwertner, A.; Machado, C.B.; Xavier, F.; de Souza, I.C.C.; Deitos, A.; Torres, I.L.S.; et al. Melatonin analgesia is associated with improvement of the descending endogenous pain-modulating system in fibromyalgia: a phase II, randomized, double-dummy, controlled trial. BMC Pharmacology and Toxicology 2014, 15, 40. [Google Scholar] [CrossRef]
- Jablochkova, A.; Bäckryd, E.; Kosek, E.; Mannerkorpi, K.; Ernberg, M.; Gerdle, B.; Ghafouri, B. Unaltered low nerve growth factor and high brain-derived neurotrophic factor levels in plasma from patients with fibromyalgia after a 15-week progressive resistance exercise. Journal of Rehabilitation Medicine 2019, 51, 779–787. [Google Scholar] [CrossRef]
- Brietzke, A.P.; Zortea, M.; Carvalho, F.; Sanches, P.R.S.; Silva, D.P.J.; Torres, I.; Fregni, F.; Caumo, W. Large Treatment Effect With Extended Home-Based Transcranial Direct Current Stimulation Over Dorsolateral Prefrontal Cortex in Fibromyalgia: A Proof of Concept Sham-Randomized Clinical Study. J Pain 2020, 21, 212–224. [Google Scholar] [CrossRef]
- Caumo, W.; Alves, R.L.; Vicuña, P.; Alves, C.F.d.S.; Ramalho, L.; Sanches, P.R.S.; Silva, D.P.; da Silva Torres, I.L.; Fregni, F. Impact of Bifrontal Home-Based Transcranial Direct Current Stimulation in Pain Catastrophizing and Disability due to Pain in Fibromyalgia: A Randomized, Double-Blind Sham-Controlled Study. The Journal of Pain 2022, 23, 641–656. [Google Scholar] [CrossRef]
- Stefani, L.C.; Leite, F.M.; da Graça, L.; Tarragó, M.; Zanette, S.A.; de Souza, A.; Castro, S.M.; Caumo, W. BDNF and serum S100B levels according the spectrum of structural pathology in chronic pain patients. Neuroscience Letters 2019, 706, 105–109. [Google Scholar] [CrossRef]
- Wolmeister, A.S.; Schiavo, C.L.; Nazário, K.C.K.; Castro, S.M.d.J.; de Souza, A.; Caetani, R.P.; Caumo, W.; Stefani, L.C. The Brief Measure of Emotional Preoperative Stress (B-MEPS) as a new predictive tool for postoperative pain: A prospective observational cohort study. PLOS ONE 2020, 15, e0227441. [Google Scholar] [CrossRef]
- Palmer, A.C.S.; Zortea, M.; Souza, A.; Santos, V.; Biazús, J.V.; Torres, I.L.S.; Fregni, F.; Caumo, W. Clinical impact of melatonin on breast cancer patients undergoing chemotherapy; effects on cognition, sleep and depressive symptoms: A randomized, double-blind, placebo-controlled trial. PLOS ONE 2020, 15, e0231379. [Google Scholar] [CrossRef]
- Dall’Agnol, L.; Medeiros, L.F.; Torres, I.L.S.; Deitos, A.; Brietzke, A.; Laste, G.; de Souza, A.; Vieira, J.L.; Fregni, F.; Caumo, W. Repetitive Transcranial Magnetic Stimulation Increases the Corticospinal Inhibition and the Brain-Derived Neurotrophic Factor in Chronic Myofascial Pain Syndrome: An Explanatory Double-Blinded, Randomized, Sham-Controlled Trial. The Journal of Pain 2014, 15, 845–855. [Google Scholar] [CrossRef]
- Chassot, M.; Dussan-Sarria, J.A.; Sehn, F.C.; Deitos, A.; de Souza, A.; Vercelino, R.; Torres, I.L.S.; Fregni, F.; Caumo, W. Electroacupuncture analgesia is associated with increased serum brain-derived neurotrophic factor in chronic tension-type headache: a randomized, sham controlled, crossover trial. BMC Complementary and Alternative Medicine 2015, 15, 144. [Google Scholar] [CrossRef]
- Jiang, Y.-H.; Liu, H.-T.; Kuo, H.-C. Decrease of Urinary Nerve Growth Factor but Not Brain-Derived Neurotrophic Factor in Patients with Interstitial Cystitis/Bladder Pain Syndrome Treated with Hyaluronic Acid. PLOS ONE 2014, 9, e91609. [Google Scholar] [CrossRef] [PubMed]
- Medeiros, L.F.; Caumo, W.; Dussán-Sarria, J.; Deitos, A.; Brietzke, A.; Laste, G.; Campos-Carraro, C.; de Souza, A.; Scarabelot, V.L.; Cioato, S.G.; et al. Effect of Deep Intramuscular Stimulation and Transcranial Magnetic Stimulation on Neurophysiological Biomarkers in Chronic Myofascial Pain Syndrome. Pain Medicine 2016, 17, 122–135. [Google Scholar] [CrossRef] [PubMed]
- Saxena, A.K.; Lakshman, K.; Sharma, T.; Gupta, N.; Banerjee, B.D.; Singal, A. Modulation of serum BDNF levels in postherpetic neuralgia following pulsed radiofrequency of intercostal nerve and pregabalin. Pain Management 2016, 6, 217–227. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Wang, M.-H.; Wang, X.-B.; Liu, L.; Wu, J.-L.; Yang, X.-L.; Liu, X.-R.; Zhang, C.-X. Effect of intraoperative application of ketamine on postoperative depressed mood in patients undergoing elective orthopedic surgery. Journal of Anesthesia 2016, 30, 232–237. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, H.; Sesterhenn, R.B.; Souza, A.d.; Souza, A.C.d.; Alves, M.; Machado, J.C.; Burger, N.B.; Torres, I.L.d.S.; Stefani, L.C.; Fregni, F.; et al. Preoperative transcranial direct current stimulation: Exploration of a novel strategy to enhance neuroplasticity before surgery to control postoperative pain. A randomized sham-controlled study. PLOS ONE 2017, 12, e0187013. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.-G.; Sun, W.; Ju, F.; Wang, H.; Sun, X.-L.; Mou, X.; Yuan, H. Analgesic Effects of Directed Repetitive Transcranial Magnetic Stimulation in Acute Neuropathic Pain After Spinal Cord Injury. Pain Medicine 2019, 21, 1216–1223. [Google Scholar] [CrossRef] [PubMed]
- Govil, M.; Mukhopadhyay, N.; Holwerda, T.; Sluka, K.; Rakel, B.; Schutte, D.L. Effects of genotype on TENS effectiveness in controlling knee pain in persons with mild to moderate osteoarthritis. European Journal of Pain 2020, 24, 398–412. [Google Scholar] [CrossRef] [PubMed]
- Beltran Serrano, G.; Pooch Rodrigues, L.; Schein, B.; Zortea, M.; Torres, I.L.S.; Fregni, F.; Caumo, W. The Hypnotic Analgesia Suggestion Mitigated the Effect of the Transcranial Direct Current Stimulation on the Descending Pain Modulatory System: A Proof of Concept Study. J Pain Res 2020, 13, 2297–2311. [Google Scholar] [CrossRef] [PubMed]
- Paula, T.M.H.d.; Castro, M.S.; Medeiros, L.F.; Paludo, R.H.; Couto, F.F.; Costa, T.R.d.; Fortes, J.P.; Salbego, M.d.O.; Behnck, G.S.; Moura, T.A.M.d.; et al. Association of low-dose naltrexone and transcranial direct current stimulation in fibromyalgia: a randomized, double-blinded, parallel clinical trial. Brazilian Journal of Anesthesiology (English Edition) 2023, 73, 409–417. [Google Scholar] [CrossRef]
- Montero-Marin, J.; Andrés-Rodríguez, L.; Tops, M.; Luciano, J.V.; Navarro-Gil, M.; Feliu-Soler, A.; López-del-Hoyo, Y.; Garcia-Campayo, J. Effects of attachment-based compassion therapy (ABCT) on brain-derived neurotrophic factor and low-grade inflammation among fibromyalgia patients: A randomized controlled trial. Scientific Reports 2019, 9, 15639. [Google Scholar] [CrossRef] [PubMed]
- Di Bonaventura, S.; Fernández Carnero, J.; Ferrer-Peña, R. Can a specific biobehavioral-based therapeutic education program lead to changes in pain perception and brain plasticity biomarkers in chronic pain patients? A study protocol for a randomized clinical trial. PLOS ONE 2024, 19, e0289430. [Google Scholar] [CrossRef]
- Polacchini, A.; Metelli, G.; Francavilla, R.; Baj, G.; Florean, M.; Mascaretti, L.G.; Tongiorgi, E. A method for reproducible measurements of serum BDNF: comparison of the performance of six commercial assays. Sci Rep 2015, 5, 17989. [Google Scholar] [CrossRef]
- Salio, C.; Ferrini, F.; Muthuraju, S.; Merighi, A. Presynaptic modulation of spinal nociceptive transmission by glial cell line-derived neurotrophic factor (GDNF). J.Neurosci. 2014, 34, 13819–13833. [Google Scholar] [CrossRef]
- Ferrini, F.; Salio, C.; Boggio, E.M.; Merighi, A. Interplay of BDNF and GDNF in the Mature Spinal Somatosensory System and Its Potential Therapeutic Relevance. Curr Neuropharmacol 2021, 19, 1225–1245. [Google Scholar] [CrossRef]
- Merighi, A. Cross Talk of BDNF and GDNF in Spinal Substantia Gelatinosa (Lamina II): Focus on Circuitry. Adv Exp Med Biol 2021, 1331, 215–229. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Synthesis, secretion, and effects of BDNF and its pro-peptide (proBDNF) on plasticity at the PSN/SSN synapse. PSNs use glutamate as a fast-acting neurotransmitter and some slow-acting neurotransmitters among which is BDNF [15]. BDNF is first produced as a pre-pro-protein precursor (BDNF pre-pro-protein), which consists of a signal sequence (light blue), a pro-domain (magenta), and the mature BDNF peptide (cobalt). Upon entering the endoplasmic reticulum (ER), the signal sequence is promptly removed to produce the BDNF pro-protein (proBDNF), which is then transported to the trans-Golgi network (TGN). In the TGN pre-proBDNF binds to sortilin or sorCS2 to be sorted in dense-core large granular vesicles (LGVs). In mature LGVs, proBDNF detaches from sortilin and becomes ready for release. The Val66Met BDNF polymorphism, represented by a thick red line, influences the release of BDNF in response to activity. The mature BDNF peptide is produced through proteolytic cleavage of proBDNF within presynaptic LGVs and then released by either the constitutive (not represented) or regulated activity-dependent pathway. BDNF and its precursor protein promote the strengthening of synaptic connections in LTP and the weakening of LTD. The receptors trkB and p75NTR mediate signaling in response to their specific ligands. Inhibition of trkB blocks BDNF signaling, whereas p75NTR prevents the signaling of proBDNF. Additional receptor components (indicated by the question mark) may be necessary to transduce the numerous biological effects of the two molecules. The facilitation of LTD generated by proBDNF requires the activation of a pathway that involves NMDAR and AMPAR, as illustrated. See the main text for a description of the precise mechanism by which ligands operate in both healthy and dysfunctional brains. Abbreviations: AMPAR = α-amino-3-hydroxy-5-methyl-4-isoxazole propionic acid receptor; BDNF = brain-derived neurotrophic factor; CaMKII = calcium/calmodulin-stimulated protein kinase II; ER = endoplasmic reticulum; LGV = large granular vesicles; LTD = long-term depression; LTP = long-term potentiation; p75NTR = low-affinity p75 neurotrophin receptor; proBDNF = BDNF pro-protein; PSN = primary sensory neuron; sorCS2 = = sortilin-related VPS10 domain-containing receptor 2; SSN = secondary sensory neuron; TGN = trans-Golgi network; trkB = trkB = tropomyosin receptor kinase B; ↑ = increase; ↓ = reduction. Created with BioRender.com.
Figure 1.
Synthesis, secretion, and effects of BDNF and its pro-peptide (proBDNF) on plasticity at the PSN/SSN synapse. PSNs use glutamate as a fast-acting neurotransmitter and some slow-acting neurotransmitters among which is BDNF [15]. BDNF is first produced as a pre-pro-protein precursor (BDNF pre-pro-protein), which consists of a signal sequence (light blue), a pro-domain (magenta), and the mature BDNF peptide (cobalt). Upon entering the endoplasmic reticulum (ER), the signal sequence is promptly removed to produce the BDNF pro-protein (proBDNF), which is then transported to the trans-Golgi network (TGN). In the TGN pre-proBDNF binds to sortilin or sorCS2 to be sorted in dense-core large granular vesicles (LGVs). In mature LGVs, proBDNF detaches from sortilin and becomes ready for release. The Val66Met BDNF polymorphism, represented by a thick red line, influences the release of BDNF in response to activity. The mature BDNF peptide is produced through proteolytic cleavage of proBDNF within presynaptic LGVs and then released by either the constitutive (not represented) or regulated activity-dependent pathway. BDNF and its precursor protein promote the strengthening of synaptic connections in LTP and the weakening of LTD. The receptors trkB and p75NTR mediate signaling in response to their specific ligands. Inhibition of trkB blocks BDNF signaling, whereas p75NTR prevents the signaling of proBDNF. Additional receptor components (indicated by the question mark) may be necessary to transduce the numerous biological effects of the two molecules. The facilitation of LTD generated by proBDNF requires the activation of a pathway that involves NMDAR and AMPAR, as illustrated. See the main text for a description of the precise mechanism by which ligands operate in both healthy and dysfunctional brains. Abbreviations: AMPAR = α-amino-3-hydroxy-5-methyl-4-isoxazole propionic acid receptor; BDNF = brain-derived neurotrophic factor; CaMKII = calcium/calmodulin-stimulated protein kinase II; ER = endoplasmic reticulum; LGV = large granular vesicles; LTD = long-term depression; LTP = long-term potentiation; p75NTR = low-affinity p75 neurotrophin receptor; proBDNF = BDNF pro-protein; PSN = primary sensory neuron; sorCS2 = = sortilin-related VPS10 domain-containing receptor 2; SSN = secondary sensory neuron; TGN = trans-Golgi network; trkB = trkB = tropomyosin receptor kinase B; ↑ = increase; ↓ = reduction. Created with BioRender.com.
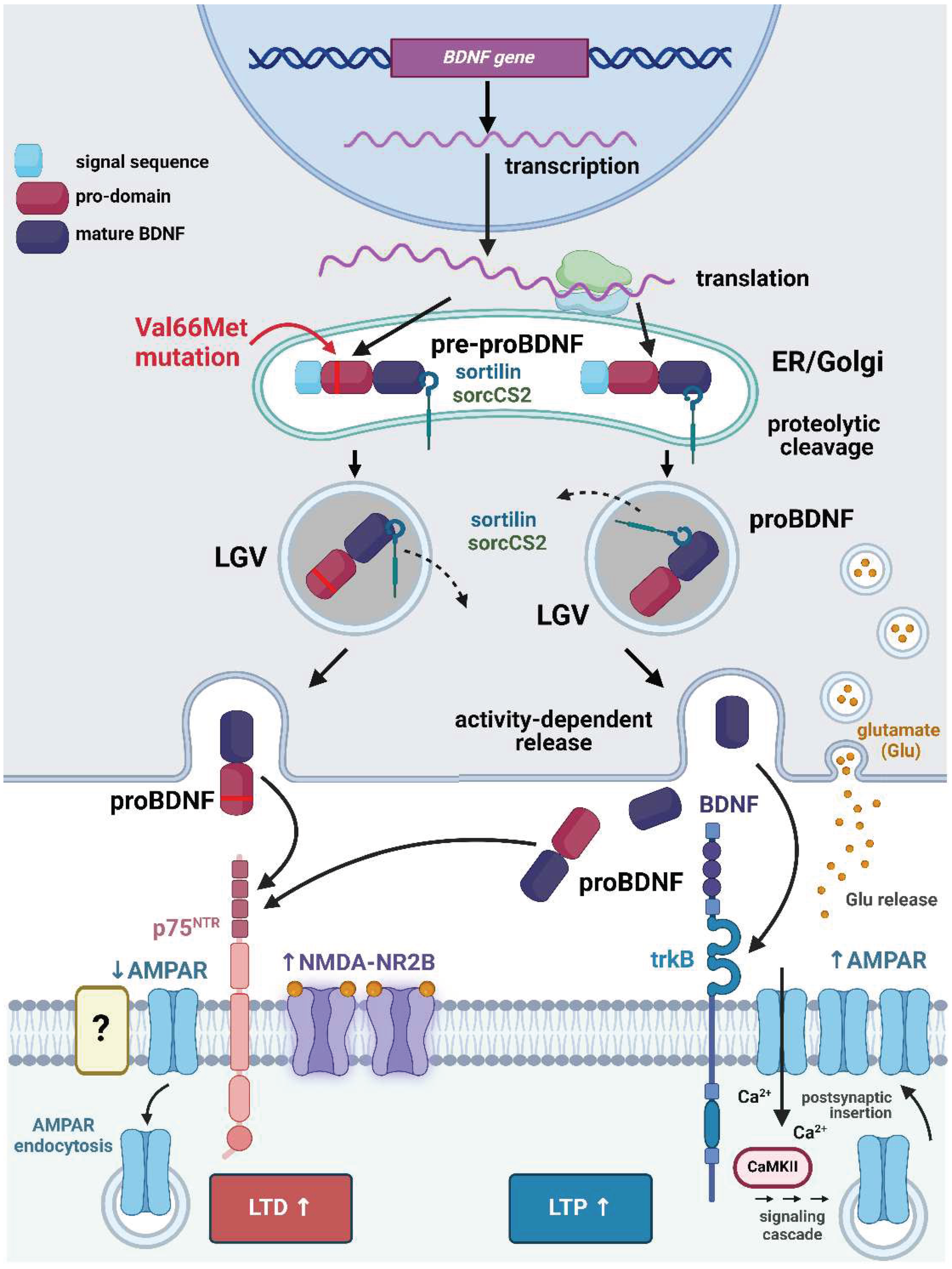
Figure 2.
Schematic representation of the main pathways involved in nociception/pain. Somatic afferents are in light blue, and visceral afferents are in green. BDNF-expressing neurons under basal conditions are in red. Neurons expressing BDNF under inflammation/neuropathy or chronic pain are in fuchsia. Abbreviations: ACC = anterior cingulate cortex; AMYG = amygdala; BDNF = brain-derived neurotrophic factor (protein); CNS = central nervous system; DCP = dorsal column pathway; DCN = dorsal column nuclei; DRGs = dorsal root ganglia; HYPO = hypothalamus; INS = insula; M1 = primary motor cortex; NG = nodose ganglion; NTS = nucleus tractus solitarius; PBN = parabrachial nucleus; PCC = posterior cingulate cortex; PFC = prefrontal cortex; PPC = posterior parietal complex; PG = petrosal ganglion; PNS = peripheral nervous system; PNTN = pontine nucleus of trigeminal nerve; S1 = primary somatosensory cortex; S2 = secondary somatosensory cortex; SMA = supplementary motor area; SNTN = spinal nucleus of trigeminal nerve; SPBN = spinal parabrachial neurons; SPBP = spinoparabrachial pathway; sSTTN = somatic spinothalamic neurons; STP = spinothalamic pathway; TG = trigeminal ganglion; TTP = trigeminothalamic pathway; VPLN = ventroposterior lateral nucleus of the thalamus; VMLN = ventroposterior medial nucleus of the thalamus; vSTTN = visceral spinothalamic neurons.
Figure 2.
Schematic representation of the main pathways involved in nociception/pain. Somatic afferents are in light blue, and visceral afferents are in green. BDNF-expressing neurons under basal conditions are in red. Neurons expressing BDNF under inflammation/neuropathy or chronic pain are in fuchsia. Abbreviations: ACC = anterior cingulate cortex; AMYG = amygdala; BDNF = brain-derived neurotrophic factor (protein); CNS = central nervous system; DCP = dorsal column pathway; DCN = dorsal column nuclei; DRGs = dorsal root ganglia; HYPO = hypothalamus; INS = insula; M1 = primary motor cortex; NG = nodose ganglion; NTS = nucleus tractus solitarius; PBN = parabrachial nucleus; PCC = posterior cingulate cortex; PFC = prefrontal cortex; PPC = posterior parietal complex; PG = petrosal ganglion; PNS = peripheral nervous system; PNTN = pontine nucleus of trigeminal nerve; S1 = primary somatosensory cortex; S2 = secondary somatosensory cortex; SMA = supplementary motor area; SNTN = spinal nucleus of trigeminal nerve; SPBN = spinal parabrachial neurons; SPBP = spinoparabrachial pathway; sSTTN = somatic spinothalamic neurons; STP = spinothalamic pathway; TG = trigeminal ganglion; TTP = trigeminothalamic pathway; VPLN = ventroposterior lateral nucleus of the thalamus; VMLN = ventroposterior medial nucleus of the thalamus; vSTTN = visceral spinothalamic neurons.
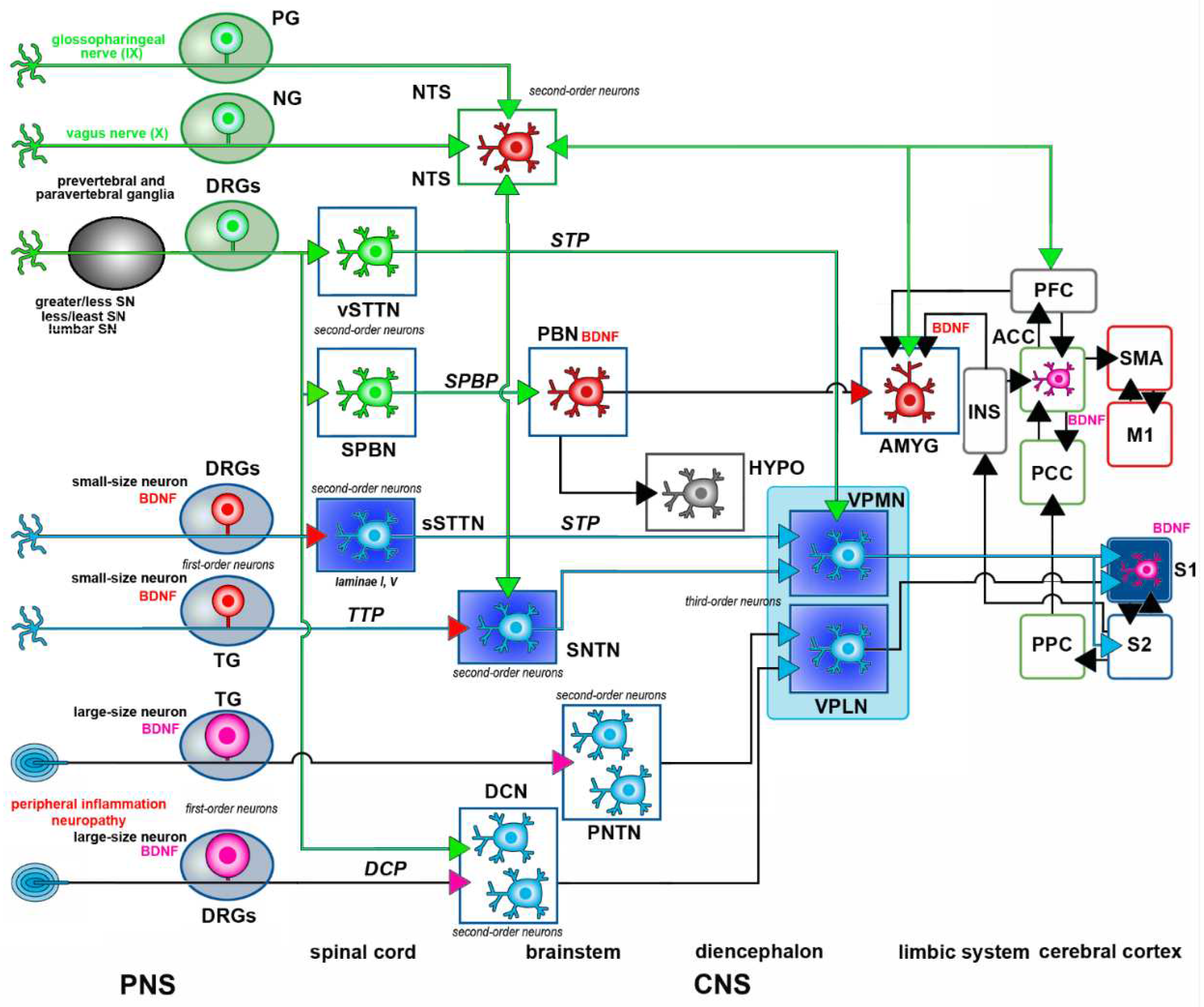
Figure 4.
Glial and immune cell BDNF synthesis related to nociceptive/pain pathways. Top: a small-size pseudounipolar PSN giving rise to unmyelinated/poorly myelinated fibers of the Aδ-/C-type. These neurons have a single process giving rise to a peripheral branch to tissues (e.g., skin) and a central branch reaching the CNS gray matter. At the periphery, the PSN terminals expressing trkB can bind BDNF released by immune cells. Areas delimited by the red squares are shown at higher magnification at the bottom. A: Schwann cell released BDNF binds to trkB to phosphorylate PKCε in the peripheral branch. PKCε in turn activates several ionotropic receptors in the PSN including glutamate receptors, TRVP1, and NaV1.8. B = BDNF released from activated microglia promotes neuropathic pain. Following PNI, spinal microglia undergo activation and exhibit excessive expression of P2RX4. Under pathological circumstances, touch spikes elicited in Aβ fibers (not shown) lead to the release of GABA and ATP from inhibitory interneurons located in the dorsal horn. ATP that has been released activates P2X4Rs on microglial cells, leading to the release of BDNF. BDNF then acts on the trkB receptors of SSNs, resulting in the downregulation of KCC2 and an increase in [Cl⁻]i. Following these events, the released GABA impacts the Cl⁻ channels of SSNs, causing an elevation in Cl⁻ outflow, which in turn depolarizes these neurons and triggers action potentials. C: Release of BDNF from astrocytes is dependent on trkB.T1. Under inflammation, astrocytes release glutamate and several bioactive molecules through CX43 channels, among which IL-1β, and CCL2. CCL2 acts on CCR2 to increase p ERK in Aδ- or C-fiber terminals of PSNs and SSNs. Inhibition of Kir 4.1 induces BDNF expression in astrocytes. D = Oligodendrocytes release BDNF after injury and the neurotrophin acts on presynaptic trkB to promote neurotransmitter release. Abbreviations: BDNF = brain-derived neurotrophic factor; CCL2 = chemokine (C-C motif) ligand 2; [Cl⁻]i = intracellular chloride concentration; CCR2 = C-C chemokine receptor type 2; ERK = extracellular signal-regulated kinase; GABAAR = γ amino-butyric acid receptor A; GLUN2B-NMDA = NMDA receptor GluN2B; GlyR = glycine receptor; KCC2 =K-Cl co-transporter 2; Kir 4.1 = inwardly rectifying potassium channel 4.; NaV1.8 = voltage-gated sodium channel 1.8; PKCε = protein kinase Cε; PNI = peripheral nerve injury; P2RX4 = ATP-gated purinergic receptor 4; PSN = primary sensory neuron; SSN = secondary sensory neuron; trkB = trkB = tropomyosin receptor kinase B; trkB.T1 = truncated isoform 1 of trkB; TRPV1 = transient receptor potential vanilloid 1. Created with BioRender.com.
Figure 4.
Glial and immune cell BDNF synthesis related to nociceptive/pain pathways. Top: a small-size pseudounipolar PSN giving rise to unmyelinated/poorly myelinated fibers of the Aδ-/C-type. These neurons have a single process giving rise to a peripheral branch to tissues (e.g., skin) and a central branch reaching the CNS gray matter. At the periphery, the PSN terminals expressing trkB can bind BDNF released by immune cells. Areas delimited by the red squares are shown at higher magnification at the bottom. A: Schwann cell released BDNF binds to trkB to phosphorylate PKCε in the peripheral branch. PKCε in turn activates several ionotropic receptors in the PSN including glutamate receptors, TRVP1, and NaV1.8. B = BDNF released from activated microglia promotes neuropathic pain. Following PNI, spinal microglia undergo activation and exhibit excessive expression of P2RX4. Under pathological circumstances, touch spikes elicited in Aβ fibers (not shown) lead to the release of GABA and ATP from inhibitory interneurons located in the dorsal horn. ATP that has been released activates P2X4Rs on microglial cells, leading to the release of BDNF. BDNF then acts on the trkB receptors of SSNs, resulting in the downregulation of KCC2 and an increase in [Cl⁻]i. Following these events, the released GABA impacts the Cl⁻ channels of SSNs, causing an elevation in Cl⁻ outflow, which in turn depolarizes these neurons and triggers action potentials. C: Release of BDNF from astrocytes is dependent on trkB.T1. Under inflammation, astrocytes release glutamate and several bioactive molecules through CX43 channels, among which IL-1β, and CCL2. CCL2 acts on CCR2 to increase p ERK in Aδ- or C-fiber terminals of PSNs and SSNs. Inhibition of Kir 4.1 induces BDNF expression in astrocytes. D = Oligodendrocytes release BDNF after injury and the neurotrophin acts on presynaptic trkB to promote neurotransmitter release. Abbreviations: BDNF = brain-derived neurotrophic factor; CCL2 = chemokine (C-C motif) ligand 2; [Cl⁻]i = intracellular chloride concentration; CCR2 = C-C chemokine receptor type 2; ERK = extracellular signal-regulated kinase; GABAAR = γ amino-butyric acid receptor A; GLUN2B-NMDA = NMDA receptor GluN2B; GlyR = glycine receptor; KCC2 =K-Cl co-transporter 2; Kir 4.1 = inwardly rectifying potassium channel 4.; NaV1.8 = voltage-gated sodium channel 1.8; PKCε = protein kinase Cε; PNI = peripheral nerve injury; P2RX4 = ATP-gated purinergic receptor 4; PSN = primary sensory neuron; SSN = secondary sensory neuron; trkB = trkB = tropomyosin receptor kinase B; trkB.T1 = truncated isoform 1 of trkB; TRPV1 = transient receptor potential vanilloid 1. Created with BioRender.com.
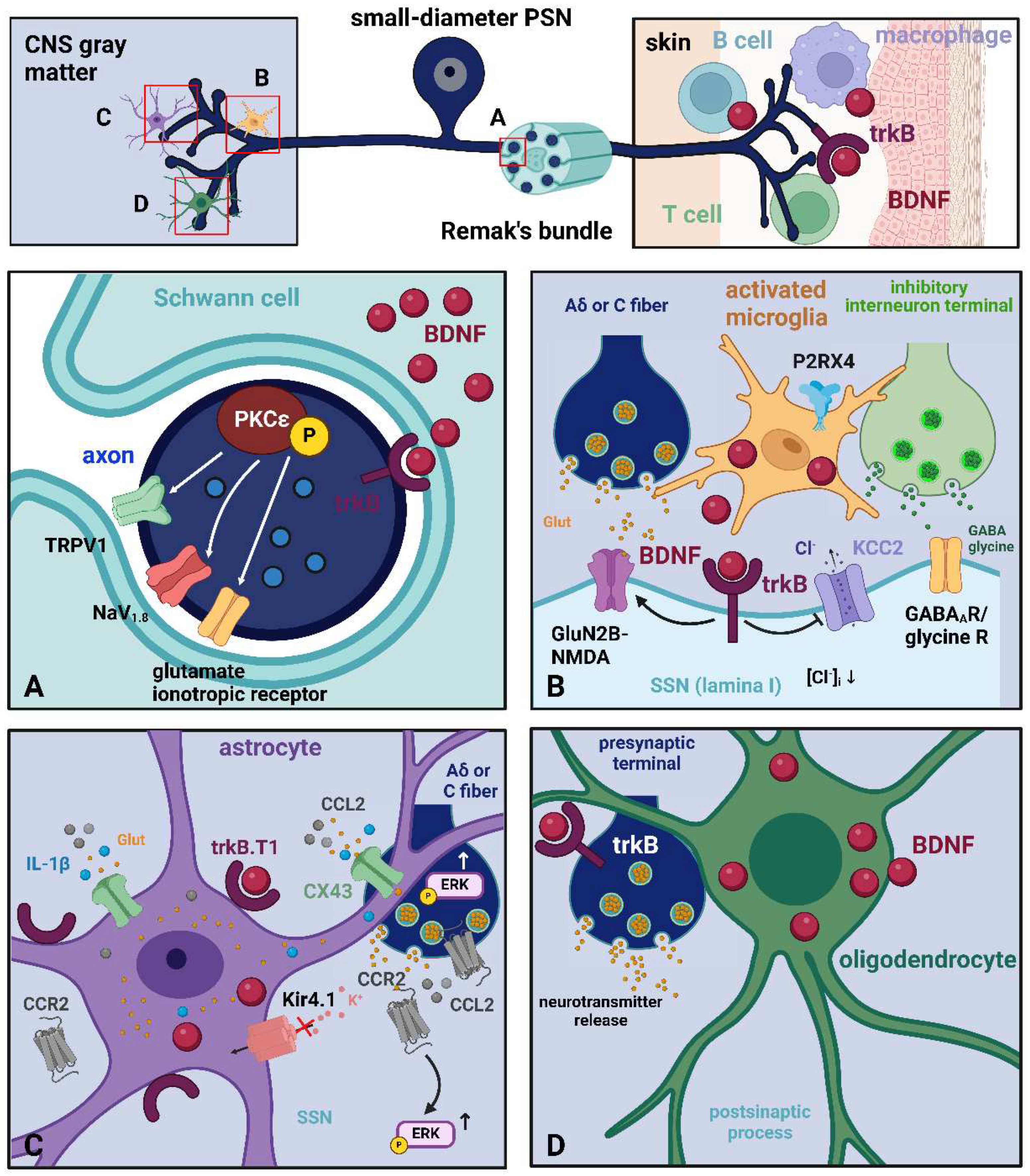

Table 1.
Distribution of BDNF in somatic pain pathway neurons after immunocytochemical staining.
| Location | Type of neurons | Function | Ref |
|---|---|---|---|
| DRGs | Small-to-medium-size PSNs | Nociception | [57–59] |
| DRGs | Small-to-medium-size PSNs | Inflammatory nociception | [81] |
| JG (X) | Small-to-medium-size PSNs | Nociception | [43] |
| TG (V) | Small-to-medium-size PSNs | Nociception | [67,68] |
| DRGs | Large-size PSNs | Nociception (after injury) | [62] |
| SNTN | SSNs | Localization study | [36] |
| CN | SSNs | Localization study | [36] |
| PBN | Projection neurons | Nociception | [36] |
| S1 | Cortical neurons | Inflammatory/neuropathic pain | [74] |
| Au1 | Cortical neurons | Chronic pain | [76] |
| ACC | Cortical neurons | Inflammatory/neuropathic pain | [74,75] |
Abbreviations: ACC = anterior cingulate cortex; Au1= primary auditory cortex; CN = cuneiform nucleus; DRGs = dorsal root ganglia; JG = jugular ganglion; PSNs = primary sensory neurons; S1 = primary somatosensory cortex; SNTN = spinal nucleus of the trigeminal nerve; SSNs = second-order sensory neurons. TG = trigeminal ganglion. Roman numerals indicate the cranial nerves.
Table 2.
Distribution of BDNF in visceral pain pathway neurons after immunocytochemical staining.
| Location | Type of neurons | Function | Ref |
|---|---|---|---|
| DRGs | Small-to-medium-size PSNs | Nociception | [82] |
| NG (X) | Small-to-medium-size PSNs | Chemoafferent neurons | [43,72] |
| PG (IX) | Small-to-medium-size PSNs | Chemoafferent neurons | [43,72] |
| NTS | Small-to-medium-size SSNs | Modulation of cardiovascular afferents | [36,73] |
| AMY | Projection neurons to PBN | Hyperalgesia | [78] |
Abbreviations: AMY = amygdala; DRGs = dorsal root ganglia; NG = nodose ganglion; NTS = nucleus tractus solitarius; PBN = parabrachial nucleus; PG = petrosal ganglion; PSNs = primary sensory neurons; SSNs = second order sensory neurons. Roman numerals indicate the cranial nerves.
Table 3.
Distribution of BDNF in pain pathways glia.
| Location | Type of cells | Function | Ref |
|---|---|---|---|
| PNS | Schwann cells | Modulation of nociception | [161] |
| DH | Activated microglia (OX-42+/ Iba1+) | Inflammatory/neuropathic pain | [162,163,164] |
| SNTN | Microglia (Iba1+) | Nociception (allodynia) | [119] |
| Au1 | Astrocytes (GFAP+) | Chronic pain | [76] |
| ACC | Astrocytes (GFAP+) | ||
| S1 | Astrocytes (GFAP+) Microglia (Iba1+) |
Inflammatory/neuropathic pain | [74,165] |
Abbreviations: ACC = anterior cingulate cortex; Au1= primary auditory cortex; DH = dorsal horn of the spinal cord; GFAP = glial fibrillary acidic protein; Iba1 = ionized calcium-binding adaptor molecule 1; OX42 = antibody against integrin αM; S1 = primary somatosensory cortex; SNTN = spinal nucleus of the trigeminal nerve.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



