Submitted:
30 May 2024
Posted:
31 May 2024
You are already at the latest version
Abstract
The allosteric SERCA (Sarcoplasmic/Endoplasmic Reticulum Ca2+-ATPase) activator CDN1163 has been recently added to the group of pharmacological tools for probing SERCA function. We chose to investigate the effects of the compound on T lymphocyte Ca2+ stores, using the well-described Jurkat T lymphocyte as a reliable cell system for Ca2+ signaling pathways. Our study identified the lowest concentrations of the SERCA inhibitors thapsigargin (TG) and 2,5-di-(tert butyl)-1,4-benzohydroquinone (tBHQ) capable of releasing Ca2+, permitting the differentiation of the TG-sensitive SERCA 2b Ca2+ store from the tBHQ-sensitive SERCA 3 Ca2+ store. We proceeded to test the effects of CDN1163 on Ca2+ stores, examining specific actions on the SERCA 2b and SERCA 3 Ca2+ pools using our low dose SERCA blocker regimen. In contrast to previous work, we find CDN1163 exerts complex time-sensitive and SERCA isoform-specific actions on Ca2+ stores. Surprisingly, short-term exposure (0-30 minutes) to CDN1163 perturbs T cell Ca2+ stores by suppressing Ca2+ uptake with diminished Ca2+ release from the SERCA 2b-controlled store. Concomitantly, we find evidence for a SERCA-activating effect of CDN1163 on the SERCA 3 regulated store, given the observation of increased Ca2+ release inducible by low dose tBHQ. Intriguingly, longer-term (>12 hours) CDN1163 exposure reversed this pattern, with increased Ca2+ release from SERCA 2b-regulated pools, yet decreased Ca2+ release responses from tBHQ-sensitive SERCA 3 pool. Our results reveal differential effects of CDN1163 on Ca2+ stores regulated by distinct SERCA isoforms, prompting the need for careful interpretation of the compound’s effects, particularly in cells expressing multiple SERCA isoforms.
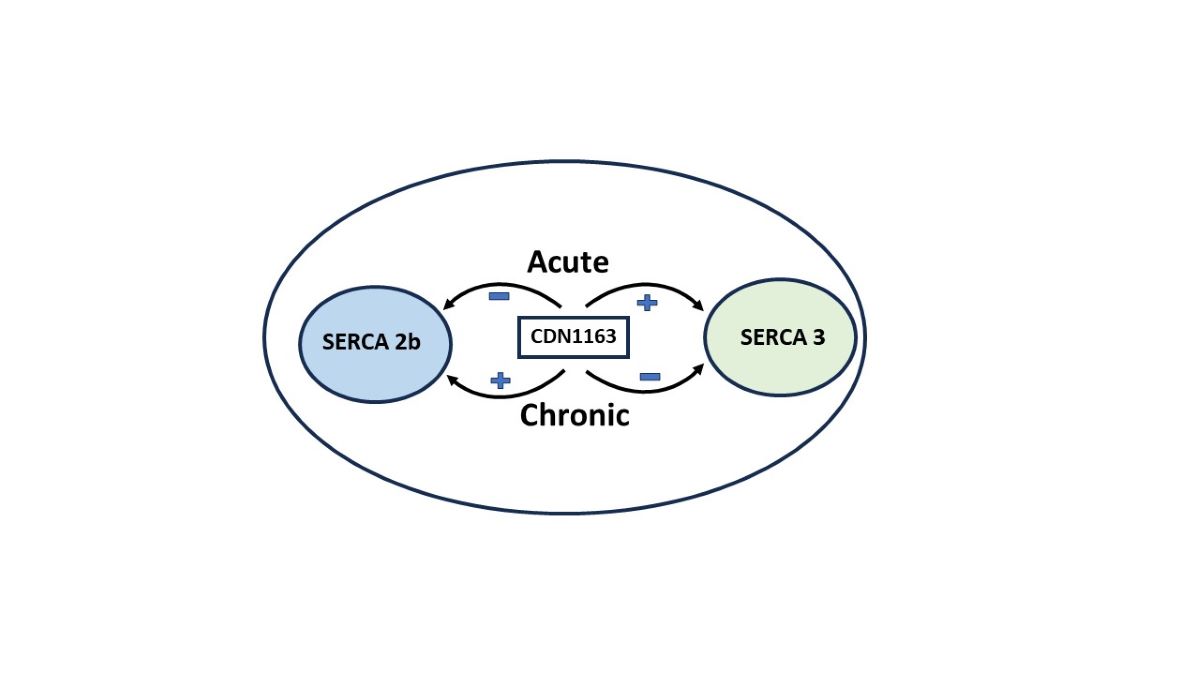
Keywords:
Ca2+ Stores
; SERCA Regulation
; ER Ca2+ Pools
; T Cell Activation
1. Introduction
The T lymphocyte is the chief orchestrator of the adaptive immune system, coordinating and regulating the multi-level interactions and deployment of both cell-mediated immunity and antibody responses [1]. T cells initiate the critical early stages of an immune response via the signaling reactions induced by T cell receptor (TCR) binding to an antigenic stimulus. A major component of the antigen-activated TCR signaling pathway is an early rapid increase in cytosolic Ca2+ levels deriving from intracellular Ca2+ storage sites as well as activation of tightly coupled Ca2+ influx pathways [2,3,4]. T cell activation, thus relies heavily on the functional integrity of intracellular Ca2+ stores, generally thought to reside in the endoplasmic reticulum (ER). Regulation and maintenance of ER Ca2+ levels is therefore essential for activation of the TCR pathway, and the central family of ion-transporting enzymes mediating these functions are the sarcoplasmic/endoplasmic reticulum Ca2+-ATPases (SERCAs) [4,5,6,7]. The SERCA Ca2+-ATPases or Ca2+ pumps have attracted much interest as potential targets for drug modulation in disease states given their prominent role in contributing to Ca2+ release/uptake events integrated within the TCR-induced signaling framework [8,9,10,11,12,13,14,15,16].
In this study we have sought to further characterize the pharmacology of SERCA-regulated Ca2+ stores in T lymphocytes. There is a clear imperative to gain a better understanding of the roles played by Ca2+ stores as essential regulators of the complex spatiotemporal Ca2+ signal underlying critical early signaling events driving T cell activation [17,18,19]. A powerful approach to probing Ca2+ store functions in T cell signaling networks is to modulate SERCA pump function using an array of small molecule pharmacological agents that can potentially provide a means for fine control of the various SERCA pump states as the primary regulators determining ER Ca2+ store levels [9,11,12,16,19].
Much has been learned about Ca2+ stores regulation using the classic thapsigargin, cyclopiazonic acid and 2,5-di-(tert butyl)-1,4-benzohydroquinone trio of SERCA blockers, clearly validating the profitable application of SERCA pharmacology in efforts to dissect Ca2+ signaling mechanisms [20,21,22,23]. Thus to further augment our tools for SERCA pump modulation, there is compelling interest to complement the SERCA inhibitors with compounds that can increase SERCA activity. We can potentially achieve a greater insight into the complex roles and functions of ER Ca2+ stores by utilizing pharmacological agents that can both downregulate as well as upregulate SERCA functional activity. At present, CDN1163 appears to be the best pharmacological agent with the capacity to increase SERCA activity, having been shown to exert a significant boost in SERCA enzymatic action in muscle and nonmuscle SERCA isoforms [8,12,15,19,24,25,26]. We were thus motivated to examine the effects of CDN1163 on Ca2+ stores in T cells, the critical central coordinator of the adaptive immune system with a uniquely pronounced dependency on ER Ca2+ stores and Ca2+ influx pathways underlying T cell activation.
2. Results
2.1. Low Concentrations of TG and tBHQ Induce Ca2+ Release in T Cells and Establish a Pharmacological Regimen for Specific Blockade of SERCA 2b and SERCA 3
Jurkat lymphocytes and rat splenocytes were loaded with Fura 2 to measure intracellular Ca2+ changes in response to SERCA drugs and other agents that mobilize cytoplasmic Ca2+. Experiments were conducted in the absence of extracellular Ca2+, except where indicated, to directly assess Ca2+ release events and Ca2+ store status without the need to consider Ca2+ elevation due to Ca2+ influx. To acquire additional information on Ca2+ release and Ca2+ store content as well as to utilize Ca2+ mobilizing agents impermeable to plasma membrane (PM) we also conducted experiments using Jurkat cells in permeabilized membrane assays.
In order to investigate the effects of the SERCA activator CDN1163 on T cell Ca2+ stores we sought to characterize SERCA regulated Ca2+ store function using Jurkat lymphocytes as the model system for T cell Ca2+ signaling [27,28,29,30,31].
Previous work has revealed that T lymphocytes express both SERCA 2b and SERCA 3 Ca2+ pump isoforms [32,33,34,35], yet it is not known what the specific function of these two SERCA pumps is with respect to regulating Ca2+ signaling events in T cell function.
Figure 1 shows the establishment of the lowest concentrations of the SERCA inhibitors thapsigargin (TG) and 2,5-di-(tert butyl)-1,4-benzohydroquinone (tBHQ) capable of inducing detectable Ca2+ release responses in Jurkat lymphocytes with our methods. Figure 1A shows responses to the sequential application of TG (100 pM), tBHQ (1 μM) and ionomycin (1 μM) in Jurkat cells loaded with Fura 2 and suspended in a Ca2+-free medium. Following TG-induced Ca2+ release (ΔF340/380= 0.16 ± 0.04 ratio units, n=12), the further addition of low concentration tBHQ induces an additional increment of Ca2+ discharge on top of the elevated TG response (ΔF340/380= 0.10 ± 0.007, n=10) from intracellular stores, suggesting that the two SERCA blockers are acting on distinct Ca2+ storage sites. Lastly, the addition of ionomycin (1 μM) induces the release of remaining sequestered Ca2+ (ΔF340/380= 0.28 ± 0.07, n=12), suggesting that the releasable Ca2+ induced by low dose TG and tBHQ is approximately half of the total Ca2+ stored in the Jurkat lymphocyte. Previous studies using a variety of hematopoietic-derived cells including platelets and Jurkat lymphocytes have revealed that low concentrations of TG specifically block SERCA 2b pumps whereas low doses of tBHQ specifically target the SERCA 3 isoform [32,36,37,38].
Figure 1B shows a similar experimental outcome where we reversed the order of application of the SERCA blockers, adding tBHQ first followed by TG addition. This result is consistent with the interpretation that the drugs used at these concentrations are revealing that the SERCA 2b and SERCA 3 regulated Ca2+ stores represent separate units of Ca2+ release in the Jurkat lymphocyte, given Ca2+ release responses are similar in magnitude (tBHQ: 0.18 ± 0.06, TG: 0.08 ± 0.003 ratio units, n=6) and independent of the order of application of the SERCA blockers. We obtained additional evidence that the SERCA blockers are indeed releasing Ca2+ from distinct storage sites by depleting the tBHQ-releasable pool with higher tBHQ levels (2 μM), revealing that the TG-releasable store remains essentially unaffected in this sequence (Figure 1C, n=4). As mentioned, there is a strong imperative to keep TG levels low in these experiments to maintain specificity for SERCA 2b blockade [32,36,37,38]. Thus, in contrast to Figure 1C, we could not effectively deplete the TG-releasable Ca2+ store without affecting tBHQ-induced release given that increasing TG levels inhibits SERCA3 pumps due to the general high potency for TG-induced inhibition on all SERCA isoforms.
Figure 1D and Figure 1E show Ca2+ responses to TG and tBHQ application in rat primary lymphocytes. As above, we used TG (100 pM) and tBHQ (1 μM) at low concentrations determined to work in Jurkat cell Ca2+ release experiments. We find that a similar effect of TG and tBHQ treatment is observed in primary lymphocytes, regardless of the order of drug application and with similar relative proportions of releasable Ca2+ pools. We could not however perform these experiments in Ca2+-free media as the Ca2+ release responses were too small to measure; thus the experiments shown in Figures 1D and 1E reflect Ca2+ responses in cells suspended in Ca2+-containing media, which resulted in larger fluorescence responses compared to Jurkat cell experiments. The Ca2+ release responses induced by TG and tBHQ, albeit larger due to contributions from Ca2+ influx (Figure 1D, TG induced response: ΔF340/380= 2.6 ± 0.62, n=5; tBHQ induced response: ΔF340/380= 5.2 ± 1.1, n=5), suggest that primary lymphocytes contain the same Ca2+ pool profile as we observe in Jurkat lymphocytes with low concentrations of TG and tBHQ inducing release from SERCA 2b and SERCA 3 Ca2+ pools respectively, and thus validate the usage of the Jurkat lymphocyte as a good model system for investigating SERCA regulation of T cell Ca2+ signaling networks.
2.2. Low Dose SERCA Blockers and Agonist-Induced Ca2+ Release Patterns Suggest a Complex T Cell Ca2+ Pool Profile with up to Five Distinct Ca2+ Store Compartments
In order to investigate the effects of the SERCA activator CDN1163 on T cell Ca2+ stores, we sought to further characterize the properties and relationships of the SERCA 2b and SERCA3-regulated Ca2+ pools in Jurkat lymphocytes as revealed by treatment with low doses of TG and tBHQ.
Previous work has identified at least four distinct Ca2+ stores in Jurkat lymphocytes comprising inositol 1,4,5-trisphosphate (IP3), TG, ryanodine receptor (RyR) and ionomycin releasable Ca2+ pools [27]. The recognition that low tBHQ concentrations release Ca2+ from a SERCA 3-regulated Ca2+ pool suggests one additional Ca2+ store and thus increases to at least five the number of distinct Ca2+ pools in the T lymphocyte Ca2+ signaling paradigm. Indeed, we show in Figure 2A the sequential release of Ca2+ induced by the application of low dose tBHQ (1 μM, 0.13 ± 0.03 ratio units) and TG (100 pM, 0.04 ± 0.008 ratio units) followed by a further Ca2+ release response induced by the addition of the T cell receptor crosslinker phytohemagglutinin A (PHA, 10 μg/ml, 0.07 ± 0.004 ratio units), which is known to mobilize Ca2+ from the IP3 sensitive stores. Along with the ionomycin-releasable Ca2+ pool (1 μM, 0.17 ± 0.04 ratio units), Figure 2A suggests the presence of four intracellular Ca2+ pools in the Jurkat lymphocyte. Figure 2B shows that we can further induce Ca2+ release in this sequential application scheme by the inclusion of 30 μM ryanodine (0.04 ± 0.002 ratio units), revealing RyR, tBHQ, TG, PHA (IP3) and ionomycin releasable stores, thereby accounting for five distinct Ca2+ pools in the Jurkat lymphocyte. The final ionomycin induced release in Figure 2B is approximately 35% reduced compared to Figure 2A (0.17 vs. 0.11 ratio units), consistent with the more extensive discharge of intracellular Ca2+ pools via application of both SERCA blockers and RyR/IP3R activators.
To examine the intracellular Ca2+ store profile in T lympocytes more closely we conducted experiments using permeabilized Jurkat T cells allowing investigation of direct Ca2+ release responses induced by agonists impermeant to the plasma membrane. For these experiments we used NAADP to release Ca2+ from RyR-sensitive stores and the direct application of IP3 to release Ca2+ from TCR (PHA)-coupled IP3R-sensitive stores in addition to the SERCA blockers and ionomycin. Figure 2C shows the detection of Ca2+ release from internal pools by the sequential application of NAADP (peak ΔF: 0.55 ± 0.03), tBHQ (ΔF: 0.59 ± 0.04), IP3 (ΔF: 0.49 ± 0.02), TG (ΔF: 0.46 ± 0.05) and ionomycin (ΔF: 0.31 ± 0.02). Figure 2D shows that regardless of the order of application these Ca2+ release activators can induce release from five distinct intracellular storage sites. Thus, Figure 2C,D confirm the results observed from intact cell experiments indicating the presence of five releasable Ca2+ pools in T lymphocytes.
2.3. Assessment of Inter-Relationships of Agonist Releasable Ca2+ Pools in T Lymphocytes
We explored the nature of the relationships between the SERCA 2b and SERCA 3 regulated Ca2+ stores to gain further insight into the functional roles of these distinct Ca2+ pools in T lymphocytes. Previous studies have reported that different Ca2+ mobilizing agents demonstrate the capacity to specifically induce Ca2+ release from SERCA 2b or SERCA 3 storage compartments, which may reveal recruitment of distinct Ca2+ release pathways corresponding to either SERCA 2b or SERCA 3 gated pools to subserve specific signaling functions in T cells, as has been shown previously for ADP secretion in platelets [39,40,41,42].
We chose to examine further a select group of Ca2+ mobilizing agonists that operate through the TCR, RyR and GPCR Ca2+ release pathways. As above, we utilized both intact and permeabilized Ca2+ assays to determine the effects of SERCA 2b and SERCA 3 modulation on PHA, ryanodine and thrombin responses in intact T lymphocytes and, correspondingly, IP3 and the RyR activator NAADP in permeablized cells.
In agreement with previous experiments using Jurkat lymphocytes [27], we find that the IP3 sensitive Ca2+ pool is a subcompartment of the larger TG releasable Ca2+ store given we still observe TG (1.5 nM)-mediated Ca2+ release following IP3 (0.5 μM)-induced responses in permeabilized cells (Figure 2C,D). However, by adding TG first and by gradually increasing its concentration up to just 15 nM we can abolish the IP3 induced Ca2+ release responses (Figure 3A–C), verifying that with more aggressive SERCA 2b inhibition we deplete the IP3-sensitive Ca2+ pool. Indeed, by increasing TG levels to 15 nM we find that we can abolish tBHQ responses as well, indicating loss of the ability to pharmacologically discriminate between SERCA 2b and SERCA 3- regulated stores at this TG concentration (Figure 3C). These observations provide assurance we can perturb TCR-coupled IP3 sensitive Ca2+ stores via specific pharmacologic modulation of SERCA 2b function with low dose TG.
RyR signaling in T cells has been marked by a relatively high degree of irresolution given mostly low expression levels of the receptor in lymphocytes [43,44,45]. Nonetheless, recent work claims a prominent role for RyR in shaping the earliest Ca2+ signals essential for T cell activation [43,44,46]. Jurkat T lymphocytes as a clonal homogeneous population have been useful in clarifying the roles of RyR in T cell signaling given their expression of RyRs, albeit at low levels [28,47,48]. Studies using Jurkat lymphocytes have revealed that the Ca2+ mobilizing agonist NAADP acts on RyRs to induce Ca2+ release from the ER and not from a unique separate acidic Ca2+ store compartment as has been observed in platelets[49,50]. Moreover, platelet studies have indicated that the SERCA 3 Ca2+ pump and not SERCA 2b isoform controls the NAADP Ca2+ releasable store [39,51]. Thus, we performed experiments to determine whether T cell RyR regulated Ca2+ stores, like platelet NAADP releasable pools, are affiliated specifically with SERCA 3 pumps using our low dose TG and tBHQ SERCA blocker regimen. Figure 4A shows that ryanodine (30 μM) induces a small Ca2+ transient (0.06 ± 0.005 ratio units, n=5) in Jurkat cells suspended in a Ca2+ free medium which rapidly decays in the presence of functional SERCA 2b/3 Ca2+ pump activity. The ryanodine inducible responses have no effect on subsequent Ca2+ release responses induced by tBHQ (0.12 ± 0.008 ratio units) and TG (0.08 ± 0.006 ratio units) suggesting that RyR activation in the Jurkat lymphocyte is inducing release from a smaller subcompartment of the larger tBHQ and/or TG releasable pools. However, when we reverse the order of application and increase the concentration of tBHQ we find that we can abolish the ryanodine inducible response (Figure 4B, p<0.05, n=5) with little effect on the subsequent TG induced Ca2+ release response (Figure 4B). This finding suggests that, similar to platelets, NAADP releasable Ca2+ stores are replenished by SERCA 3 Ca2+ pumps in T cells.
We investigated whether we could alter thrombin responses in T cells using low dose TG and tBHQ to establish specific linkage of thrombin releasable Ca2+ stores with SERCA 2b or SERCA 3 Ca2+ pools. Thrombin acts on G protein-coupled receptors (GPCRs) and has been observed in platelet studies to be coupled to two distinct signaling functions [40,41]: thrombin induced Ca2+ release from SERCA 3-regulated pools comprised an early platelet signal to generate ADP secretion which was amplified by a secondary thrombin-induced Ca2+ signal deriving from SERCA 2b-regulated stores [52]. It was proposed that the thrombin activated GPCR pathway produced two second messengers in the platelet system: IP3 moblized Ca2+ from the SERCA 2b Ca2+ stores whereas NAADP released Ca2+ as the initial early signal from SERCA 3 Ca2+ stores [52]. We find that an apparent similar mechanism may be working in T cells (Figure 4C–E). Figure 4C shows that exposure of Jurkat lymphocytes to tBHQ (2 μM) fails to abolish thrombin responses (0.13 ± 0.03 ratio units, n=5), while also leaving intact subsequent TG responses (0.09 ± 0.006 ratio units) representing the SERCA 2b Ca2+ pool. Similarly, the addition of TG (200 pM) first also does not eradicate the thrombin induced Ca2+ release response (0.11 ± 0.008 ratio units, n=5), while subsequent tBHQ responses are also still inducible albeit significantly reduced (p<0.05, n=5), suggesting that most of the thrombin releasable Ca2+ is contained in the SERCA 2b regulated stores (Figure 4D). Conversely, if T cells are treated first with tBHQ (2 μM) followed by TG (200 pM) thrombin responses are abolished (Figure 4E). This result is consistent with the platelet observations suggesting that thrombin in T lymphocytes appears to release Ca2+ from both SERCA 2b and SERCA 3 Ca2+ stores.
2.4. Despite Differences in SERCA Blocker Sensitivities and Agonist-Mobilizable Ca2+ Responses, the SERCA 2b and SERCA 3-Regulated Ca2+ Stores Exhibit Similar Ca2+ Influx Coupling Actions with Similar Sensitivity to Actin Cytoskeletal Disruption
T cell activation and gene expression pathways require that SERCA regulated Ca2+ stores communicate with PM Orai channels to mediate Ca2+ influx [3,7]. Thus, we sought to investigate whether the TG and tBHQ releasable Ca2+ stores described above exhibited similar features in regulating Ca2+ influx pathways. The platelet Ca2+ signaling system provides a useful framework for understanding the T cell signaling network which also appears to contain multiple distinct SERCA 2b and SERCA 3 regulated intracellular Ca2+ stores. Platelet studies have revealed differences in the SERCA 2b and SERCA 3 regulated Ca2+ stores, with the SERCA 3 Ca2+ stores demonstrating weaker coupling to Ca2+ influx pathways but less sensitivity to cytoskeletal disruption as compared to SERCA 2b Ca2+ stores [53]. Our experiments using T cells reveal differences to the platelet system with respect to Ca2+ store regulation of Ca2+ influx pathways. Figure 5A shows that tBHQ-mediated SERCA 3 blockade and pool depletion activates Ca2+ influx responses (2.3 ± 0.52 ratio units, n=12) similar to those induced by TG treatment (2.4 ± 0.71 ratio units, n=15), suggesting that, unlike platelets, the SERCA 3 and SERCA 2b- regulated stores exhibit similar coupling sensitivities to depletion-induced Ca2+ influx responses in T lymphocytes. However, in contrast to the platelet system, we find that treating Jurkat cells with the actin cytoskeletal disruptor cytochalasin D (cytD,10 μM) increases the ability of both SERCA 2b and SERCA 3 Ca2+ stores to couple to Ca2+ influx responses, as we observed larger Ca2+ influx responses in both TG (6.9 ± 2.5 control vs. 9.0 ± 2.1 cytD ratio units, n=4) and tBHQ (2.1 ± 0.88 control vs. 5.5 ± 1.2 cytD ratio, n=4) treated cells (Figure 5B,C). Thus, in T cells the two Ca2+ stores both appear to be negatively regulated by actin polymerization, suggesting that actin dynamics may be interfering with SERCA 2b and SERCA 3 vesicle trafficking to the plasma membrane Ca2+ channels. In platelets the insensitivity of the SERCA 3 Ca2+ store to cytD induced actin disruption led to speculation that this Ca2+ pool may reside in close junctional apposition to the plasma membrane [53], which our experiments suggest may not be the case in the T cell system. Intriguingly, we find that pre-treating Jurkat lymphocytes with cytD significantly reduces the TG (0.25 ± 0.03 control vs. 0.14 ± 0.05 cytD ratio units, p<0.05, n=4) and tBHQ (0.16 ± 0.008 vs. 0.10 ± 0.006 cytD ratio units, p<0.05, n=4) inducible Ca2+ release responses (Figure 5D,E) suggesting that stable actin filament networks may also be essential for supporting Ca2+ release structures, a feature that was not observed in platelet experiments. Indeed, this effect of cytoskeletal perturbation to attenuate Ca2+ release may reflect a more depleted Ca2+ pool state and thus explain a more robust Ca2+ influx coupling response following cytD exposure (Figure 5B,C).
2.5. The SERCA Activator CDN1163 Exerts Complex Short and Long-Term Effects on T Cell Ca2+ Stores Revealing a Differential Regulatory Action on SERCA 2b Versus SERCA 3 Ca2+ Pools.
We next focused on exploring the effects of the recently identified SERCA activator molecule CDN1163 on T cell Ca2+ stores. As mentioned, there is much interest in identifying and characterizing a small molecule complement to the group of SERCA blockers that can achieve SERCA activation. Our foregoing experiments have further characterized some of the additional complexity in T cell Ca2+ stores in revealing distinct SERCA 2b and SERCA 3 Ca2+ pools that appear to be recruited to produce Ca2+ responses tailored to distinct signaling triggers, including TCR activation, thrombin and RyR-mediated signals. Thus, given previous work elucidating the salutary effects of CDN1163 attributable to increased activity of SERCA function[8,12,16,26], we anticipated that the compound would produce an augmented Ca2+ store condition with perhaps greater Ca2+ release responses in the Jurkat T cell system. However when we measured intact Jurkat cell Ca2+ responses treated with varying concentrations of CDN1163 we did not uniformly observe this effect.
Indeed, Figure 6A shows that a 20 minute pre-incubation of T cells with CDN1163 (10 μM) significantly reduced TCR-mediated Ca2+ release as measured by PHA treatment in Ca2+-free media (Peak ratio units: 0.29 ± 0.06 untreated vs. 0.15 ± 0.04 CDN treated, p< 0.05, n=7), which represents the IP3 releasable component of the larger TG sensitive Ca2+ stores. We observed the same effect of short-term (<30 minutes) CDN1163 exposure when cells were treated with low dose TG (100 pM), revealing reduced Ca2+ release responses from this pharmacologically defined SERCA 2b Ca2+ pool (Figure 6B). Surprisingly, this effect was not observed in the tBHQ-sensitive SERCA 3 Ca2+ store. Figure 6C shows that Jurkat lymphocytes exposed to CDN1163 (10 μM) for 20 minutes exhibit significantly increased Ca2+ release responses when treated with low dose tBHQ (peak ratio units: 0.17 ± 0.008 untreated vs. 0.35 ± 0.06 CDN treated, p<0.05, n=8). And thus our low dose TG/tBHQ treatment regimen reveals reciprocal effects of short-term CDN1163 exposure on T cell SERCA 2b versus SERCA 3-regulated Ca2+ stores. These findings suggest the intriguing possibility that CDN1163 can exert opposing differential regulatory influences on the SERCA pumps, perturbations induced by short-term exposure to the compound that produce diminished Ca2+ loading in the SERCA 2b pool with concurrent augmentation of the SERCA 3 Ca2+ pool.
Our experiments with CDN1163 in T cells thus yielded somewhat paradoxical effects, given we appear to observe diminished levels of stored Ca2+ in the major SERCA 2b-regulated pool after treatment with a putative SERCA pump activator. To further explore CDN1163’s effects on T cell Ca2+ stores, we used saponin-permeabilization assays to measure Ca2+ uptake responses directly. We incubated Jurkat lymphocytes with CDN1163 (25 μM) over a short time interval (≤ 30 min) and then proceeded to induce PM permeabilization using low concentrations of saponin. Calcium uptake into ER stores was then initiated by the addition of ATP in a cuvette-based assay, tracking Ca2+ uptake into stores by the decline in Fluo-3 fluorescence. Indeed, Figure 6D shows that CDN1163-treated permeabilized Jurkat cells revealed pronounced inhibition of Ca2+ uptake as determined by linear initial rates of fluorescence decay (ΔF/sec: 5.2 × 10−3 untreated vs. 1.5 x 10−3 30 min CDN treated, n=8), suggesting a possible mechanism to explain the observed attenuated Ca2+ release responses from the SERCA 2b-regulated Ca2+ stores. This result however does not explain the apparent augmentation of Ca2+ stored in the tBHQ-sensitive SERCA 3-regulated Ca2+ pool. It is worth noting that previous studies using Jurkat cells and platelets have revealed that the SERCA 2b-regulated Ca2+ store is likely to be larger than the SERCA 3 Ca2+ store [27,51,53]. Thus, it is possible that CDN1163 short-term exposure is specifically perturbing the SERCA 2b-mediated Ca2+ uptake which would comprise the dominant effect in our permeabilized cell assays masking a smaller contribution from increased SERCA 3-mediated Ca2+ uptake. We also observed that application of CDN1163 directly to Fura 2 loaded Jurkat lymphocytes induced a gradual small increase in cytoplasmic Ca2+ levels (data not shown) further suggesting that the compound over this time interval blocks aggregate SERCA activity and thereby induces Ca2+ leakage from the ER Ca2+ stores leading to a relatively depleted state in the SERCA 2b Ca2+ store. As above, this result may be explained by the larger effect on the SERCA 2b Ca2+ pool experiencing a CDN1163-induced downregulatory action even while the compound exerts a modest increase in Ca2+ loading into the SERCA 3 pool. Notably, this ability of CDN1163 to induce gradual increases in cytosolic Ca2+ levels with short-term exposure was also recently reported in experiments measuring Ca2+ changes in A549 lung epithelial cells [54,55].
Given CDN1163’s well-described action as an allosteric SERCA pump activator [8], we tested whether the compound may require longer periods of exposure to T lymphocytes to increase SERCA activity globally and augment Ca2+ store levels in SERCA 2b and SERCA 3 regulated stores. Indeed, we observed that longer exposure to CDN1163 (72 hours, Figure 6E) increased Ca2+ store levels in the SERCA 2b pool, as revealed by the application of low dose TG (peak ratio units: 0.33 ± 0.06 untreated vs. 0.49 ± 0.07 72 hr CDN treated, p<0.05, n=7). Yet, surprisingly, Ca2+ store levels in the SERCA 3-regulated stores moved in the opposite direction exhibiting reduced levels with long-term CDN1163 treatment (Figure 6F), as the low dose tBHQ-induced Ca2+ release responses revealed (peak ratio units: 0.54 ± 0.05 untreated vs. 0.39 ± 0.04 72 hr CDN treated, n=9). Thus, as with the short-term incubation experiments, we also find in the longer-term exposures a remarkable asymmetry in CDN1163’s effects on the SERCA 2b and SERCA 3-regulated Ca2+ stores, with an augmented SERCA 2b and a concomitant reduced SERCA 3 pool with longer CDN1163 treatment. We conducted additional experiments using the permeabilized Jurkat T cell assay to determine if this reversal effect due to longer-term CDN1163 exposure could be observed in direct Ca2+ release assays and on Ca2+ uptake responses. Indeed, we observed the same pattern in these experiments with permeabilized T lymphocytes as was found using intact cells, detecting increased Ca2+ release directly with TG application (Figure 6G) and, conversely, diminished Ca2+ release inducible by tBHQ application (Figure 6H) in cells incubated with CDN1163 (10 μM) for 72 hours. Intriguingly, it appears that, like TG and tBHQ, CDN1163 exerts differential effects on SERCA 2b and SERCA 3 Ca2+ pump isoforms. And thus, our findings suggest that the stimulatory effect of CDN1163 on T cell Ca2+ stores is multiplex, requiring prolonged incubation to reverse an initial inhibitory action and augment the SERCA 2b Ca2+ store, yet over the extended incubation period the compound exerts a gradual downregulation of Ca2+ replenishment of the SERCA 3 regulated store. Using the Ca2+ uptake assay in permeabilized cells over a range of different CDN1163 exposure periods, we found that by approximately 12 hours Ca2+ uptake activity had been restored (Figure 6I), as well as perhaps slightly elevated (ΔF/sec: 7.2 x 10−3 untreated vs. 8.8 x 10−3 CDN 12 hr treated, n=8). This time frame for the restoration of ATP-activated Ca2+ uptake responses in the presence of CDN1163 aligns well with the general time frame required for the shift in Ca2+ release responses to occur in the intact cell experiments.
3. Discussion
To better assess novel actions of potential SERCA activators within the Ca2+ signaling landscape of T cell functions, we were motivated to extend seminal previous work characterizing the major intracellular Ca2+ stores. We have employed the strategy of using low concentrations of TG and tBHQ in our experiments, an approach successfully applied to probe the functions SERCA 2b and SERCA 3- regulated Ca2+ pools in human platelets. We applied this strategy using the Jurkat T lymphocyte model which has been used less extensively than platelets to characterize properties of the intracellular Ca2+ stores. We identified the lowest concentrations of SERCA blockers that elicited measurable Ca2+ release responses in cells incubated in Ca2+-free media, thus providing greater assurances of using these agents to specifically modulate the SERCA 2b and SERCA 3 pump isoforms. We validated the use of the Jurkat T cell line by verifying that these same effects could be produced in primary lymphocytes isolated from rat splenocytes, confirming that Jurkat cells share the same pharmacological phenotype as primary T lymphocytes when treated with low dose TG and tBHQ. Indeed, use of the Jurkat T lymphocyte provides a significant experimental advantage given their clonal homogeneous responses and the ability to cultivate large numbers of cells, features which greatly assist analysis of measuring relatively weak signals due to modest SERCA perturbations. Moreover, Jurkat lymphocytes continue to be used as powerful T cell model systems given the strong validation and close overlap with primary T cells in the signaling representation of the TCR pathway as the primary upstream activator of ER Ca2+ release along with the tightly coupled Ca2+ influx pathway [28,30,31,46].
We have examined the Ca2+ pool profile in Jurkat lymphocytes with the added discriminatory refinement of employing low concentrations of TG and tBHQ, referencing previous work in human platelets in which this approach has been productively used to gain insight into SERCA 2b and SERCA 3- regulated Ca2+ stores [36,39,40,41,51,53]. Using low dose TG and tBHQ in Ca2+-free cell suspensions we have determined Ca2+ release specifically from SERCA 2b versus SERCA 3-regulated Ca2+ stores in T lymphocytes. Indeed, our experiments extend earlier investigations to describe at least five distinct Ca2+-releasable storage sites in T cells: a TG-sensitive SERCA 2b pool, a subcompartment of the TG-sensitive pool releasable by IP3, a tBHQ-sensitive SERCA 3 pool, a pool dischargeable by agonists of RyRs, a pool releasable by GPCR agonists and finally the remaining Ca2+ storage pool releasable by ionomycin application. These are clearly approximate estimations given agonists, such as thrombin, can release Ca2+ from multiple SERCA-controlled stores; and, moreover, Ca2+ stores in T cells are likely to contain built-in interconnectivity with Ca2+ release from one compartment being captured by a neighboring SERCA-regulated compartment, as has been shown for the RyR expressing Ca2+ pools in T lymphocytes [27,28,43,56]. These observations suggest the intriguing scenario of a complex and dynamic interrelationship among the various intracellular Ca2+ stores whereby rapid exchange and flow of Ca2+ ions through discrete regional space of the larger ER organelle, whether phsyically separated or not, may establish de novo spatially localized gradients adapted to accommodate specific T cell signaling functions. These experiments have further characterized the complex array of Ca2+ storage compartments and functions in T lymphocytes and have provided a useful foundation of SERCA-specific actions (inducible via low dose TG and tBHQ) to examine the effects of the novel SERCA-activating compound CDN1163.
In contrast to most CDN1163 studies, we did not observe a clear and unambiguous stimulatory effect of the compound on T cell SERCA activity. Indeed, our experiments revealed a surprising short and long-term acting dichotomy in which an initial period of apparent SERCA inhibition and Ca2+ stores depletion gradually shifts to SERCA activation and Ca2+ stores repletion. This discrepancy with previous reports may be due to a more complex SERCA pump expression profile in T cells, given that these cells rely on a minimum of at least two distinct SERCA pump isoforms to manage intracellular Ca2+ signaling dynamics. Jurkat T lymphocytes, an often used surrogate for T cell function, are well known for their expression of multiple SERCA isoforms with the predominant pump species being the SERCA 2b and SERCA 3 isoforms [32,57]. And although the specific protein functions are unknown, Jurkat T cells appear to tap into an extensive diversity in SERCA gene expression with earlier studies revealing the expression of all six SERCA 3 pump isoforms (SERCA 3a-f) along with the SERCA 2b isoform [35,58], which suggests a high degree of precisioned and specialized control built in for regulation of T cell Ca2+ store functions.
Indeed, some of the apparent incongruous effects we observe with CDN1163 may be due to this complex SERCA environment in T cells, with distinct pump isoforms working in diverse groups of interacting protein partners within heterogeneous ER/PM locales. We find, for example, that when Jurkat lymphocytes are exposed to the putative SERCA activator CDN1163 for short durations (≤ 30 minutes) Ca2+ release induced by the IP3 pathway or by low dose TG treatment are significantly reduced. We observed this effect in Ca2+ responses measured in both intact and permeabilized cells, a result which paradoxically suggests that CDN1163 may be acting to perturb SERCA function, initiate ER Ca2+ leak pathways and promote loss of ER Ca2+ levels. This interpretation is consistent with our experiments using permeabilized cells in which we observed that short duration CDN1163 exposure suppressed Ca2+ uptake. These actions of CDN1163 to impair ER Ca2+ uptake produced a gradual increase in Ca2+ release observable in our permeabilized cell assays which was the likely cause of reduced IP3 and TG- inducible Ca2+ release.
Our approach in this study to use low concentration TG and tBHQ to specifically target SERCA 2b and SERCA 3 has provided insight into the novel actions of CDN1163. We report here intriguing differences using these two SERCA blockers in the sensitivity of SERCA 2b and SERCA 3 to the effects of the SERCA activator CDN1163. In our experiments CDN1163 appears to perturb SERCA 2b regulated Ca2+ stores to a greater extent than the low dose tBHQ sensitive SERCA 3 Ca2+ store. It is worth noting that CDN1163 has been shown in previous studies to bind to and modulate SERCA 2 isoforms in various cells and tissues, but no clear evidence has emerged for the compound’s effects on the SERCA 3 isoform [8,12,26]. Indeed, we find that CDN1163 attenuates the low dose tBHQ releasable Ca2+ store albeit with a less pronounced effect as compared to the TG sensitive pool. Furthermore, we have reported that long-term (>24 hours) CDN1163 exposure fails to produce a stimulatory effect with improved Ca2+ release inducible by tBHQ unlike what is observed in the long-term incubation experiments with the TG-releasable Ca2+ pool. These findings suggest that there are likely not uniform stimulatory effects induced by CDN1163 across all SERCA pump isoforms; and indeed, these experiments suggest that clear unambiguous stimulation of SERCA function may be difficult to achieve in T cells and other cells that express multiple SERCA pump isoforms.
Our work does appear to align with previous investigations characterizing CDN1163 as a SERCA activator, albeit acting on an enigmatically slower timeframe in our T lymphocyte model. As mentioned, this effect may be attributable to differential actions of the compound on the different SERCA pump isoforms expressed in the T lymphocyte, with the global cellular SERCA activity being the sum of complex stimulatory and inhibitory effects on SERCA 2b and SERCA 3. Intriguingly, however, CDN1163’s time-dependent augmentation of SERCA 2b Ca2+ stores may be hinting at the compound’s ability to react to and promote differential SERCA states and/or SERCA-pumping environments. Indeed, CDN1163 was initially identified in a chemical library screen for its ability to interfere with SERCA binding interactions with phospholamban [8], the cardiac protein regulator of the SERCA 2a pump isoform. Perhaps CDN1163 is targeting a similar site of regulatory control in the SERCA 2b pump explaining an initial early period of perturbation on Ca2+ transport activity with accompanying Ca2+ leak expression; yet with prolonged incubation, regulatory control possibly arising from time-dependent SERCA-associated protein partners re-configures ER systems to enhance Ca2+ uptake.
It has been reported that hematopoietic and other cell types can recruit opposing SERCA actions with downregulation of SERCA 3 activity linked to time-dependent upregulation of SERCA 2b expression/function, an observation clearly identified during T cell activation [57,59,60]. Indeed, other studies have identified STIM1 as a candidate potential SERCA regulator, such that when ER Ca2+ stores experience relative depletion STIM1, as an ER Ca2+ sensor, may participate in SERCA activating functions to replenish ER Ca2+ levels [61]. This may also explain the time delay we observe in our T cell model given the initial CDN1163-mediated ER Ca2+ leak as a relatively weak, gradual depletion-activating signal may utlimately couple to STIM1 or other protein regulators to promote greater SERCA activity with increased ER Ca2+ transport. Thus, continued interrogation and characterization of CDN1163 may offer an additional SERCA pharmacological tool to probe novel SERCA regulatory networks that appear to play multi-layered Ca2+ signaling roles in T lymphocyte signaling.
4. Materials and Methods
4.1. Materials
Fura 2/AM (fura 2 acetoxymethylester), Fluo-3 pentapotassium salt, pluronic acid, RPMI-160, fetal bovine serum (FBS), streptomycin, and penicillin were obtained from Thermo Fisher. Ryanodine, oligomycin, thapsigargin, cytochalasin D were obtained from Santa Cruz Biotechnology, Inc. (Dallas, Texas). D-myo-Inositol 1,4,5 trisphosphate K salt (IP3), nicotinic acid adenine dinucleotide phosphate sodium salt (NAADP), phytohemagglutinin (PHA), 2,5-di-(tert butyl)-1,4-benzohydroquinone (tBHQ), thrombin, creatine phosphokinase (CPK), phosphocreatine disodium salt hydrate, adenosine 5′-triphosphate disodium salt hydrate (ATP), DTT, and saponin were obtained from Sigma. Sterile Cell Strainers (100 μm), 50 ml syringe tubes and 60 mm cell culture dishes were from Fisher Scientific. CDN1163 was from Bio-Techne (Minneapolis, USA).
4.2. Cell Culture
Jurkat cells (Clone E6-1, ATCC TIB-152) were maintained in RPMI-1640 medium supplemented with 10% fetal bovine serum, 2 mM L-glutamine, penicillin (100 IU/ml), and streptomycin (100 μg/ml) and grown at 37 °C in a humidified atmosphere (95% air,5% CO2). Cells were maintained and expanded in either 25 cm2 (T25) or 75 cm2 (T75) tissue culture flasks (Fisher Scientific). Cell density was not allowed to exceed 3 X 106 cells/mL and cultures were generally maintained at a cell concentration between 1 x 105 and 1 X 106 viable cells/mL. Fresh medium was added every 2 to 3 days depending on cell density.
4.3. Splenocyte Isolation
The use of animals for these experiments was conducted in accord with protocols approved by the institutional animal care and use committees at the University of the Pacific (IACUC #22R05). Spleen and lymph nodes were aseptically isolated from adult Sprague-Dawley (SD) rats (8 weeks old, N=4). Briefly, rats were anesthetized and spleens were aseptically removed and placed into 60-mm cell culture dishes containing ice-cold HBSS (Hanks Balanced Salt Solution) and minced into small pieces with a scissor. Tissue fragments were dissected and passed through a 100-μm cell strainer using a 10-ml syringe plunger and ice-cold HBSS into a 50-ml conical tube and then centrifuged (200g at 4°C) for 10 min. The supernatant was discarded, and the pellet was resuspended in 5 ml of a red blood cell (RBC) lysis buffer containing 155 mM NH4Cl (9 parts) in 130 mM Tris base pH 7.65 (1 part) and incubated at 37°C for 5 min. RBC lysis was halted by the addition of 10 ml ice-cold complete cell culture medium and cells were then centrifuged (200g at 4°C) for 10 min. The supernatant was discarded, and the pellet was resuspended in 10 ml complete cell culture medium and maintained in a humidified atmosphere (37°C, 95% air and 5% CO2).
4.4. Cell Calcium Assays
Cells (approximately 1×106 cells/ml) were washed in Ca2+-containing (1.8 mM) HBSS (Hanks Balanced Salt Solution) and loaded with 1.5 μM fura-2/AM in 20% (w/v) Pluronic F-127 and incubated for one hour at 37°C. After loading, the cells were washed twice with HBSS and incubated at 37°C for an additional 30 min to allow for de-esterification of the dye. Cells loaded with fura 2/AM were kept in the dark at room temperature throughout the experiments. Changes in cytosolic Ca2+ were measured in cell population experiments using a fluorescence spectrophotometer equipped with a thermostatically controlled sample compartment, permitting continuous stirring of samples in the cuvette. All measurements were carried out at room temperature (25°C). To achieve Ca2+-free conditions, EGTA (2 mM) was added to chelate extracellular Ca2+ just before the addition of Ca2+ mobilizing agonists (1-2 min). Ca2+ changes in Jurkat cells and rat splenocytes loaded with fura 2/AM were measured via rapid alternation of the excitation monochromator between 340 and 380 nm, with fluorescence emission measured at 510 nm using a ratiometric spectrofluorimiter (PTI). Cytosolic Ca2+ responses are presented as the changes in the fluorescence ratio values measured at 340/380 nm for Fura-2, or as non-ratiometric Fluo-3 fluorescence changes for the excitation/emission (503/530 nm) wavelength pair. The data are reported as either peak amplitude changes in fluorescence values or as initial rates of fluorescence changes and presented as the means ± S.E.M., with the number of experimental repetitions indicated in parentheses.
4.5. Permeabilized Cell Assays
For preparation of permeabilized cells, 4×107 cells were washed twice and resuspended in 2 ml of an intracellular-like medium (110 mM-KCl, 10 mM-NaCl, 2 mM-MgCl2, 20 mM-Hepes, 5 mM-KH2PO4, pH 7.5) in the presence of 1 mM DTT. Saponin (20 μg/ml) was added, and the cell suspension was incubated for 5 min at 37°C to complete permeabilization. An ATP-regenerating system consisting of creatine kinase (40 units/ml) and phosphocreatine (20 mM) was added. Oligomycin (10 μg/ml) was also included to inhibit the mitochondrial ATPase. Following cell permeabilization, Fluo-3 (0.5 μM), was added to the cuvette. Subsequent addition of ATP to a final concentration of 1 mM resulted in a decrease in the fluorescence, indicating Ca2+ uptake by the intracellular stores. After baseline stabilization, drugs were added according to the experimental plan. Ca2+ release from intracellular stores was measured from cells suspended in cuvettes using a fluorescence spectrophotometer equipped with a thermostatically controlled sample compartment maintained at 37°C with continuous stirring. Fluorescence changes with Fluo-3 in permeabilized cell suspensions were measured with excitation wavelength settings of 503 nm and 530 nm for the emission wavelength.
4.6. Statistical Analysis
Analysis of statistical significance was performed using the Student’s T test. P values ≤ 0.05 were considered to represent significant differences in the results.
5. Conclusions
We have used a well-documented approach in platelets and Jurkat cells of low TG and tBHQ in an effort to specifically modulate SERCA 2b and SERCA 3 regulated T cell Ca2+ store functions. Using this approach we have added new insight into complex T cell Ca2+ stores: we have added new detail on the heterogenous profile on intracellular Ca2+ stores, noting evidence for at least five distinct Ca2+ pools. We observed that a key group of Ca2+ mobilizing agonists (TCR, RyR and GPCR) appear to access SERCA 2b and SERCA 3 in a differential fashion suggesting SERCA regulation of T cell signaling pathways. We also observe that unlike the platelet system, T cell SERCA 2b and SERCA 3 Ca2+ stores appear to be equally sensitive to cytoskeletal disruption and we examine actions of the novel SERCA activator CDN1163 on T cell Ca2+ store functions. We add new detail to this important new pharmacological tool as the currently best described SERCA activator, showing that the compound appears to regulate T cell SERCA 2b and SERCA 3 Ca2+ stores in a complex differential manner alternately reducing or augmenting Ca2+ release functions depending on the length of exposure to the agent. In contrast to previous studies we suggest that CDN1163 can exert complex effects in cells expressing both SERCA 2b and SERCA 3 pump isoforms that are not unfailingly consistent with pump activation, yet still may provide useful insight into SERCA regulated Ca2+ store functions.
Author Contributions
Conceptualization, D.W.T and Md.N.U; methodology, D.W.T. and Md.N.U; validation, D.W.T. and Md.N.U; formal analysis, D.W.T. and Md.N.U; investigation, D.W.T and Md.N.U; resources, D.W.T; data curation, D.W.T and Md.N.U; writing—original draft preparation, D.W.T; writing—review and editing, D.W.T and Md.N.U; visualization, D.W.T and Md.N.U; supervision, D.W.T; project administration, D.W.T; funding acquisition, D.W.T. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by a seed grant from the Jie Du Center for Innovation and Excellence for Drug Discovery, University of the Pacific.
Institutional Review Board Statement
The animal study protocol was approved by the Institutional Animal Care and Use Committee of the University of the Pacific.
Data Availability Statement
Dataset available on request from the authors.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Janeway: C.A., Jr. and K. Bottomly, Signals and signs for lymphocyte responses. Cell, 1994. 76(2): p. 275-85. [CrossRef] [PubMed]
- Lewis, R.S. and M.D. Cahalan, Ion channels and signal transduction in lymphocytes. Annu Rev Physiol, 1990. 52: p. 415-30. [CrossRef] [PubMed]
- Lewis, R.S., Calcium signaling mechanisms in T lymphocytes. Annu Rev Immunol, 2001. 19: p. 497-521. [CrossRef] [PubMed]
- Gil, D., et al., Three-Dimensional Model of Sub-Plasmalemmal Ca(2+) Microdomains Evoked by T Cell Receptor/CD3 Complex Stimulation. Front Mol Biosci, 2022. 9: p. 811145. [CrossRef] [PubMed]
- Berridge, M.J., P. Lipp, and M.D. Bootman, The versatility and universality of calcium signalling. Nat Rev Mol Cell Biol, 2000. 1(1): p. 11-21. [CrossRef] [PubMed]
- Berridge, M.J., Lymphocyte Activation in Health and Disease. Crit Rev Immunol, 2017. 37(2-6): p. 439-462. [CrossRef]
- Trebak, M. and J.P. Kinet, Calcium signalling in T cells. Nat Rev Immunol, 2019. 19(3): p. 154-169. [CrossRef] [PubMed]
- Cornea, R.L., et al., High-throughput FRET assay yields allosteric SERCA activators. J Biomol Screen, 2013. 18(1): p. 97-107. [CrossRef] [PubMed]
- Bidwell, P.A., et al., A Large-Scale High-Throughput Screen for Modulators of SERCA Activity. Biomolecules, 2022. 12(12). [CrossRef]
- Zhang, C., D.D. Bose, and D.W. Thomas, Paradoxical effects of sarco/endoplasmic reticulum Ca(2+)-ATPase (SERCA) activator gingerol on NG115-401L neuronal cells: failure to augment ER Ca(2+) uptake and protect against ER stress-induced cell death. Eur J Pharmacol, 2015. 762: p. 165-73. [CrossRef]
- Rahate, K., L.K. Bhatt, and K.S. Prabhavalkar, SERCA stimulation: A potential approach in therapeutics. Chem Biol Drug Des, 2020. 95(1): p. 5-15. [CrossRef] [PubMed]
- Kang, S., et al., Small Molecular Allosteric Activator of the Sarco/Endoplasmic Reticulum Ca2+-ATPase (SERCA) Attenuates Diabetes and Metabolic Disorders. J Biol Chem, 2016. 291(10): p. 5185-98. [CrossRef]
- Tadini-Buoninsegni, F., et al., Drug Interactions With the Ca(2+)-ATPase From Sarco(Endo)Plasmic Reticulum (SERCA). Front Mol Biosci, 2018. 5: p. 36. [CrossRef] [PubMed]
- Rakovskaya, A., et al., Positive Allosteric Modulators of SERCA Pump Restore Dendritic Spines and Rescue Long-Term Potentiation Defects in Alzheimer's Disease Mouse Model. Int J Mol Sci, 2023. 24(18). [CrossRef] [PubMed]
- Krajnak, K. and R. Dahl, A new target for Alzheimer's disease: A small molecule SERCA activator is neuroprotective in vitro and improves memory and cognition in APP/PS1 mice. Bioorg Med Chem Lett, 2018. 28(9): p. 1591-1594. [CrossRef] [PubMed]
- Sordi, G., et al., Stimulation of Ca(2+) -ATPase Transport Activity by a Small-Molecule Drug. ChemMedChem, 2021. 16(21): p. 3293-3299.
- Berridge, M.J., The endoplasmic reticulum: a multifunctional signaling organelle. Cell Calcium, 2002. 32(5-6): p. 235-49. [CrossRef]
- Berridge, M.J., M.D. Bootman, and H.L. Roderick, Calcium signalling: dynamics, homeostasis and remodelling. Nat Rev Mol Cell Biol, 2003. 4(7): p. 517-29. [CrossRef]
- Chemaly, E.R., L. Troncone, and D. Lebeche, SERCA control of cell death and survival. Cell Calcium, 2018. 69: p. 46-61. [CrossRef] [PubMed]
- Inesi, G. and Y. Sagara, Specific inhibitors of intracellular Ca2+ transport ATPases. J Membr Biol, 1994. 141(1): p. 1-6. [CrossRef]
- Inesi, G., et al., Studies of Ca2+ ATPase (SERCA) inhibition. J Bioenerg Biomembr, 2005. 37(6): p. 365-8. [CrossRef] [PubMed]
- Lytton, J., M. Westlin, and M.R. Hanley, Thapsigargin inhibits the sarcoplasmic or endoplasmic reticulum Ca-ATPase family of calcium pumps. J Biol Chem, 1991. 266(26): p. 17067-71. [CrossRef]
- Thomas, D. and M.R. Hanley, Pharmacological tools for perturbing intracellular calcium storage. Methods Cell Biol, 1994. 40: p. 65-89. [CrossRef]
- Mengeste, A.M., et al., The small molecule SERCA activator CDN1163 increases energy metabolism in human skeletal muscle cells. Curr Res Pharmacol Drug Discov, 2021. 2: p. 100060. [CrossRef] [PubMed]
- Britzolaki, A., et al., Chronic but not acute pharmacological activation of SERCA induces behavioral and neurochemical effects in male and female mice. Behav Brain Res, 2021. 399: p. 112984. [CrossRef] [PubMed]
- Nguyen, H.T., et al., CDN1163, an activator of sarco/endoplasmic reticulum Ca(2+) ATPase, up-regulates mitochondrial functions and protects against lipotoxicity in pancreatic beta-cells. Br J Pharmacol, 2023. 180(21): p. 2762-2776. [CrossRef]
- Guse, A.H., E. Roth, and F. Emmrich, Intracellular Ca2+ pools in Jurkat T-lymphocytes. Biochem J, 1993. 291 ( Pt 2)(Pt 2): p. 447-51. [CrossRef] [PubMed]
- Dadsetan, S., et al., Store-operated Ca2+ influx causes Ca2+ release from the intracellular Ca2+ channels that is required for T cell activation. J Biol Chem, 2008. 283(18): p. 12512-9. [CrossRef]
- Astoul, E., et al., PI 3-K and T-cell activation: limitations of T-leukemic cell lines as signaling models. Trends Immunol, 2001. 22(9): p. 490-6. [CrossRef]
- Benson, J.C., et al., A multiple-oscillator mechanism underlies antigen-induced Ca(2+) oscillations in Jurkat T-cells. J Biol Chem, 2023. 299(11): p. 105310. [CrossRef] [PubMed]
- Zani, F., et al., The dietary sweetener sucralose is a negative modulator of T cell-mediated responses. Nature, 2023. 615(7953): p. 705-711. [CrossRef] [PubMed]
- Wuytack, F., et al., A sarco/endoplasmic reticulum Ca(2+)-ATPase 3-type Ca2+ pump is expressed in platelets, in lymphoid cells, and in mast cells. J Biol Chem, 1994. 269(2): p. 1410-6. [CrossRef] [PubMed]
- Wu, K.D., et al., Localization and quantification of endoplasmic reticulum Ca(2+)-ATPase isoform transcripts. Am J Physiol, 1995. 269(3 Pt 1): p. C775-84. [CrossRef] [PubMed]
- Poch, E., et al., Functional characterization of alternatively spliced human SERCA3 transcripts. Am J Physiol, 1998. 275(6): p. C1449-58. [CrossRef] [PubMed]
- Kovacs, T., et al., All three splice variants of the human sarco/endoplasmic reticulum Ca2+-ATPase 3 gene are translated to proteins: a study of their co-expression in platelets and lymphoid cells. Biochem J, 2001. 358(Pt 3): p. 559-68. [CrossRef]
- Cavallini, L., M. Coassin, and A. Alexandre, Two classes of agonist-sensitive Ca2+ stores in platelets, as identified by their differential sensitivity to 2,5-di-(tert-butyl)-1,4-benzohydroquinone and thapsigargin. Biochem J, 1995. 310 ( Pt 2)(Pt 2): p. 449-52. [CrossRef]
- Papp, B., et al., Simultaneous presence of two distinct endoplasmic-reticulum-type calcium-pump isoforms in human cells. Characterization by radio-immunoblotting and inhibition by 2,5-di-(t-butyl)-1,4-benzohydroquinone. Biochem J, 1992. 288 ( Pt 1)(Pt 1): p. 297-302. [CrossRef]
- Lopez, J.J., et al., Ca2+ accumulation into acidic organelles mediated by Ca2+- and vacuolar H+-ATPases in human platelets. Biochem J, 2005. 390(Pt 1): p. 243-52. [CrossRef] [PubMed]
- Jardin, I., et al., Intracellular calcium release from human platelets: different messengers for multiple stores. Trends Cardiovasc Med, 2008. 18(2): p. 57-61. [CrossRef] [PubMed]
- Jardin, I., et al., Differential involvement of thrombin receptors in Ca2+ release from two different intracellular stores in human platelets. Biochem J, 2007. 401(1): p. 167-74. [CrossRef] [PubMed]
- Lopez, J.J., et al., Two distinct Ca2+ compartments show differential sensitivity to thrombin, ADP and vasopressin in human platelets. Cell Signal, 2006. 18(3): p. 373-81. [CrossRef] [PubMed]
- Elaib, Z., et al., Full activation of mouse platelets requires ADP secretion regulated by SERCA3 ATPase-dependent calcium stores. Blood, 2016. 128(8): p. 1129-38. [CrossRef] [PubMed]
- Fomina, A.F., Neglected wardens: T lymphocyte ryanodine receptors. J Physiol, 2021. 599(19): p. 4415-4426. [CrossRef] [PubMed]
- Thakur, P., S. Dadsetan, and A.F. Fomina, Bidirectional coupling between ryanodine receptors and Ca2+ release-activated Ca2+ (CRAC) channel machinery sustains store-operated Ca2+ entry in human T lymphocytes. J Biol Chem, 2012. 287(44): p. 37233-44. [CrossRef] [PubMed]
- Guse, A.H. and B.P. Diercks, Integration of nicotinic acid adenine dinucleotide phosphate (NAADP)-dependent calcium signalling. J Physiol, 2018. 596(14): p. 2735-2743. [CrossRef] [PubMed]
- Wolf, I.M.A. and A.H. Guse, Ca(2+) Microdomains in T-Lymphocytes. Front Oncol, 2017. 7: p. 73. [CrossRef]
- Schwarzmann, N., et al., Knock-down of the type 3 ryanodine receptor impairs sustained Ca2+ signaling via the T cell receptor/CD3 complex. J Biol Chem, 2002. 277(52): p. 50636-42. [CrossRef]
- Hohenegger, M., et al., Pharmacological activation of the ryanodine receptor in Jurkat T-lymphocytes. Br J Pharmacol, 1999. 128(6): p. 1235-40. [CrossRef]
- Dammermann, W. and A.H. Guse, Functional ryanodine receptor expression is required for NAADP-mediated local Ca2+ signaling in T-lymphocytes. J Biol Chem, 2005. 280(22): p. 21394-9. [CrossRef] [PubMed]
- Steen, M., T. Kirchberger, and A.H. Guse, NAADP mobilizes calcium from the endoplasmic reticular Ca(2+) store in T-lymphocytes. J Biol Chem, 2007. 282(26): p. 18864-71. [CrossRef]
- Rosado, J.A., Acidic Ca(2+) stores in platelets. Cell Calcium, 2011. 50(2): p. 168-74. [CrossRef] [PubMed]
- Feng, M., et al., NAADP/SERCA3-Dependent Ca(2+) Stores Pathway Specifically Controls Early Autocrine ADP Secretion Potentiating Platelet Activation. Circ Res, 2020. 127(7): p. e166-e183. [CrossRef] [PubMed]
- Rosado, J.A., et al., Two pathways for store-mediated calcium entry differentially dependent on the actin cytoskeleton in human platelets. J Biol Chem, 2004. 279(28): p. 29231-5. [CrossRef]
- Huang, Y.P., et al., CDN1163, a SERCA activator, causes intracellular Ca(2+) leak, mitochondrial hyperpolarization and cell cycle arrest in mouse neuronal N2A cells. Neurotoxicology, 2023. 98: p. 9-15. [CrossRef] [PubMed]
- Wu, K.C., et al., Suppression of Ca(2+) oscillations by SERCA inhibition in human alveolar type 2 A549 cells: rescue by ochratoxin A but not CDN1163. Life Sci, 2022. 308: p. 120913. [CrossRef] [PubMed]
- Kunerth, S., et al., Analysis of subcellular calcium signals in T-lymphocytes. Cell Signal, 2003. 15(8): p. 783-92. [CrossRef] [PubMed]
- Launay, S., et al., Modulation of endoplasmic reticulum calcium pump expression during T lymphocyte activation. J Biol Chem, 1997. 272(16): p. 10746-50. [CrossRef]
- Bobe, R., et al., Identification, expression, function, and localization of a novel (sixth) isoform of the human sarco/endoplasmic reticulum Ca2+ATPase 3 gene. J Biol Chem, 2004. 279(23): p. 24297-306. [CrossRef]
- Arbabian, A., et al., Endoplasmic reticulum calcium pumps and cancer. Biofactors, 2011. 37(3): p. 139-49. [CrossRef] [PubMed]
- Papp, B., et al., Endoplasmic reticulum calcium pumps and cancer cell differentiation. Biomolecules, 2012. 2(1): p. 165-86. [CrossRef]
- Lopez, J.J., et al., STIM1 regulates acidic Ca2+ store refilling by interaction with SERCA3 in human platelets. Biochem Pharmacol, 2008. 75(11): p. 2157-64. [CrossRef]
Figure 1.
Low concentrations of the SERCA blockers thapsigargin (TG) and 2,5, di-(tert-butyl) 1,4-benzohydroquinone (tBHQ) can specifically induce Ca2+ release from distinct Ca2+ stores in Jurkat and rat T lymphocytes. For A and B, Jurkat T lymphocytes were loaded with Fura-2 and suspended in Ca2+-free media (balanced salt solution plus 2 mM EGTA). A, Jurkat cell Ca2+ release responses induced by the sequential application (arrows) of TG (100 pM), tBHQ (Q, 1 μM) and ionomycin (I, 1 μM) as determined by the ratio of fluorescence changes at 340 and 380 nm (F340/380). B, the same experiment as in A but with the reverse application to Jurkat cells of tBHQ (Q, 1 μM), TG (100 pM) and ionomycin (I, μM). C, the same experiment as in B but with consecutive tBHQ applications (Q, 2 μM) to deplete the tBHQ pool, followed by TG application (100 pM). D, Rat spleen T cells loaded with Fura 2 and stimulated with TG (100 pM), tBHQ (Q, μM) and ionomycin (I, μM) in a balanced salt solution containing Ca2+. E, Rat spleen T cells stimulated with tBHQ (Q, μM), TG (100 pM) and ionomycin (I, μM) in Ca2+-containing media. Fluorescence traces shown are representative of four to ten separate experiments.
Figure 1.
Low concentrations of the SERCA blockers thapsigargin (TG) and 2,5, di-(tert-butyl) 1,4-benzohydroquinone (tBHQ) can specifically induce Ca2+ release from distinct Ca2+ stores in Jurkat and rat T lymphocytes. For A and B, Jurkat T lymphocytes were loaded with Fura-2 and suspended in Ca2+-free media (balanced salt solution plus 2 mM EGTA). A, Jurkat cell Ca2+ release responses induced by the sequential application (arrows) of TG (100 pM), tBHQ (Q, 1 μM) and ionomycin (I, 1 μM) as determined by the ratio of fluorescence changes at 340 and 380 nm (F340/380). B, the same experiment as in A but with the reverse application to Jurkat cells of tBHQ (Q, 1 μM), TG (100 pM) and ionomycin (I, μM). C, the same experiment as in B but with consecutive tBHQ applications (Q, 2 μM) to deplete the tBHQ pool, followed by TG application (100 pM). D, Rat spleen T cells loaded with Fura 2 and stimulated with TG (100 pM), tBHQ (Q, μM) and ionomycin (I, μM) in a balanced salt solution containing Ca2+. E, Rat spleen T cells stimulated with tBHQ (Q, μM), TG (100 pM) and ionomycin (I, μM) in Ca2+-containing media. Fluorescence traces shown are representative of four to ten separate experiments.
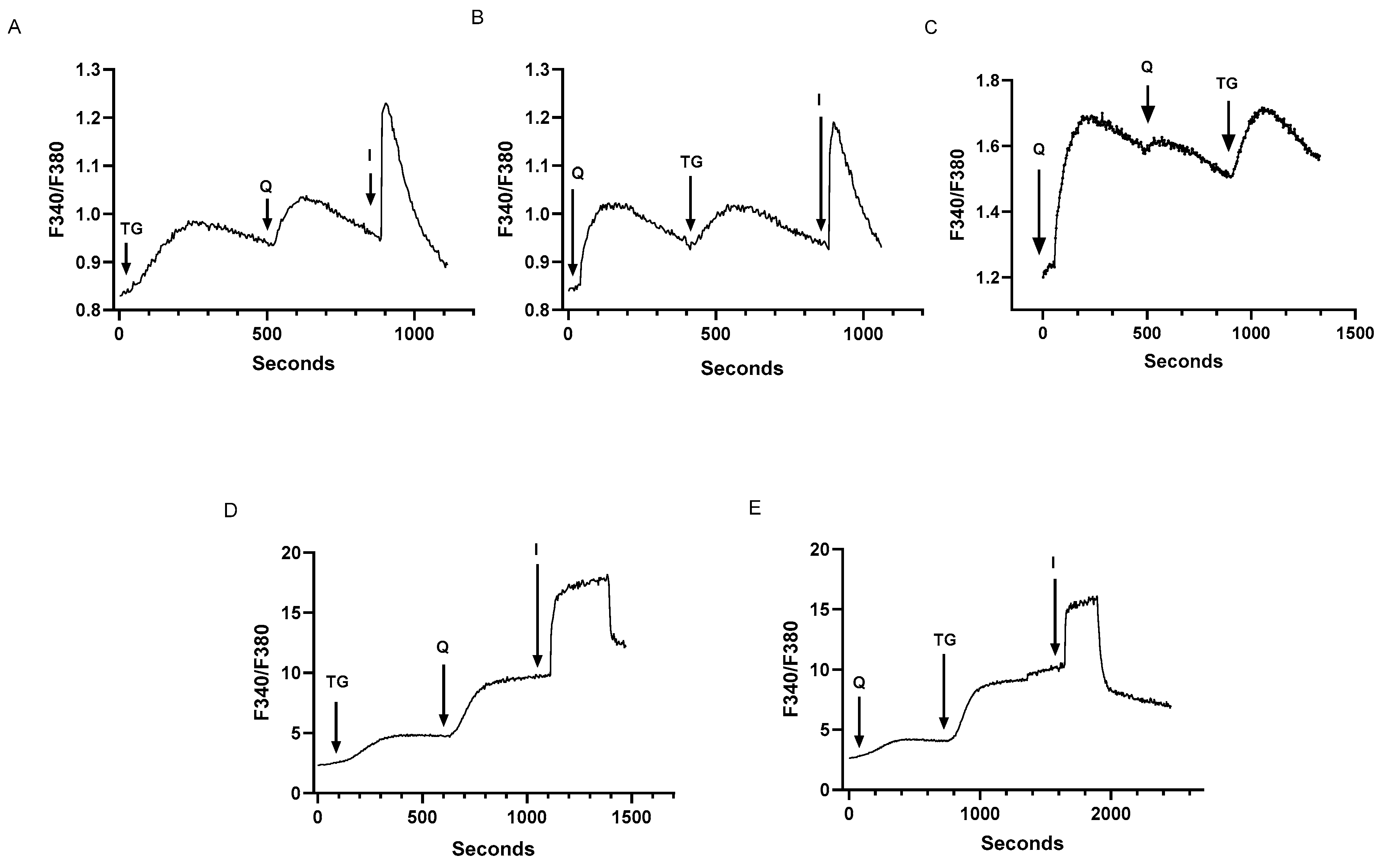
Figure 2.
Low dose SERCA blockers and Ca2+ release agonists reveal at least five distinct Ca2+ pools in intact and membrane permeabilized Jurkat T lymphocytes. For A and B, Jurkat T lymphocytes were loaded with Fura-2 and suspended in Ca2+-free media (balanced salt solution plus 2 mM EGTA). A, Jurkat cell Ca2+ release responses induced by the sequential application (arrows) of tBHQ (Q, 1 μM), TG (100 pM), PHA (10 μg/ml) and ionomycin (I, 1 μM) as determined by the ratio of fluorescence changes at 340 and 380 nm (F340/380). B, the same experiment as in A but with the sequential application to Jurkat cells of ryanodine (Ry, 30 μM), tBHQ (Q, 1 μM), TG (100 pM), PHA (10 μg/ml) and ionomycin (I, μM). C and D show experiments using saponin permeabilized Jurkat cells, depicting Ca2+ release responses as detected by Fuo-3 fluorescence changes. C, permeabilized cell responses to the sequential addition of NAADP (N, 400 μM), tBHQ (Q, 1 μM), IP3 (IP, 0.5 μM), TG (1.5 nM) and ionomycin (I, 1 μM). D. same experiment as shown in C but with the sequential addition of IP3 (IP, 0.5 μM), TG (1.5 nM), NAADP (N, 400 μM), tBHQ (Q, 1 μM) and ionomycin (I, 1 μM). Fluorescence traces shown are representative of six to ten individual experiments.
Figure 2.
Low dose SERCA blockers and Ca2+ release agonists reveal at least five distinct Ca2+ pools in intact and membrane permeabilized Jurkat T lymphocytes. For A and B, Jurkat T lymphocytes were loaded with Fura-2 and suspended in Ca2+-free media (balanced salt solution plus 2 mM EGTA). A, Jurkat cell Ca2+ release responses induced by the sequential application (arrows) of tBHQ (Q, 1 μM), TG (100 pM), PHA (10 μg/ml) and ionomycin (I, 1 μM) as determined by the ratio of fluorescence changes at 340 and 380 nm (F340/380). B, the same experiment as in A but with the sequential application to Jurkat cells of ryanodine (Ry, 30 μM), tBHQ (Q, 1 μM), TG (100 pM), PHA (10 μg/ml) and ionomycin (I, μM). C and D show experiments using saponin permeabilized Jurkat cells, depicting Ca2+ release responses as detected by Fuo-3 fluorescence changes. C, permeabilized cell responses to the sequential addition of NAADP (N, 400 μM), tBHQ (Q, 1 μM), IP3 (IP, 0.5 μM), TG (1.5 nM) and ionomycin (I, 1 μM). D. same experiment as shown in C but with the sequential addition of IP3 (IP, 0.5 μM), TG (1.5 nM), NAADP (N, 400 μM), tBHQ (Q, 1 μM) and ionomycin (I, 1 μM). Fluorescence traces shown are representative of six to ten individual experiments.
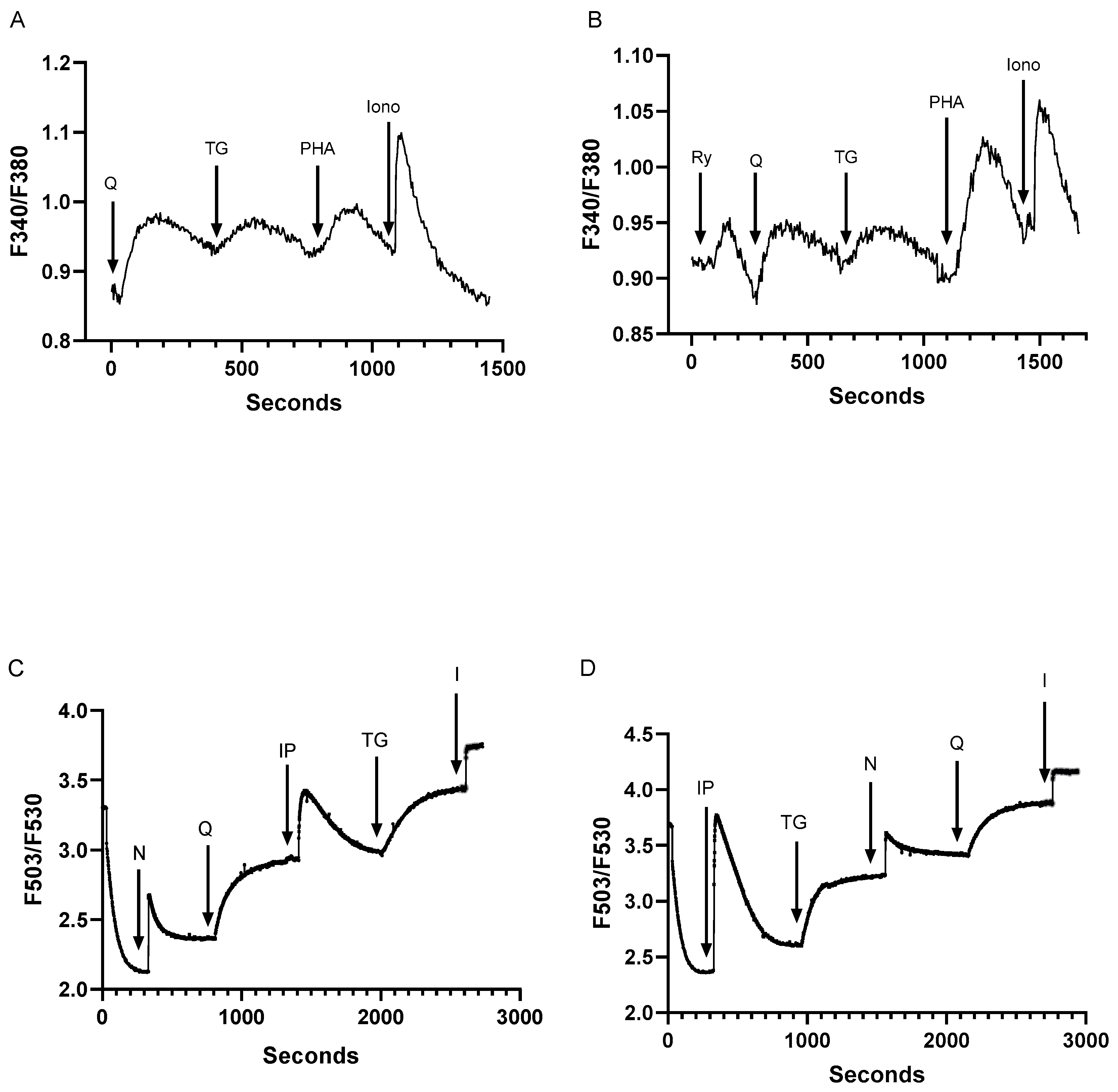
Figure 3.
Increasing TG-induced SERCA inhibition depletes the IP3 and tBHQ- releasable Ca2+ pools establishing TG concentration range permitting SERCA pool specific modulation in Jurkat T lymphocytes. A-C show experiments using saponin permeabilized Jurkat cells, depicting Ca2+ release responses as detected by Fuo-3 fluorescence changes. A, permeabilized cell responses to the sequential addition of TG (2 nM), IP3 (0.5 μM), tBHQ (Q, 1 μM) and ionomycin (I, 1 μM). B, Ca2+ release responses induced in permeabilized Jurkat lymphocytes by the sequential application of TG (10 nM), IP3 (0.5 μM), tBHQ (Q, 1 μM), and ionomycin (I, 1 μM). C, Ca2+ release responses induced in permeabilized Jurkat lymphocytes by the sequential application of TG (15 nM), IP3 (0.5 μM), tBHQ (Q, 1 μM), and ionomycin (I, 1 μM). Fluorescence traces shown are representative of four to eight individual experiments.
Figure 3.
Increasing TG-induced SERCA inhibition depletes the IP3 and tBHQ- releasable Ca2+ pools establishing TG concentration range permitting SERCA pool specific modulation in Jurkat T lymphocytes. A-C show experiments using saponin permeabilized Jurkat cells, depicting Ca2+ release responses as detected by Fuo-3 fluorescence changes. A, permeabilized cell responses to the sequential addition of TG (2 nM), IP3 (0.5 μM), tBHQ (Q, 1 μM) and ionomycin (I, 1 μM). B, Ca2+ release responses induced in permeabilized Jurkat lymphocytes by the sequential application of TG (10 nM), IP3 (0.5 μM), tBHQ (Q, 1 μM), and ionomycin (I, 1 μM). C, Ca2+ release responses induced in permeabilized Jurkat lymphocytes by the sequential application of TG (15 nM), IP3 (0.5 μM), tBHQ (Q, 1 μM), and ionomycin (I, 1 μM). Fluorescence traces shown are representative of four to eight individual experiments.
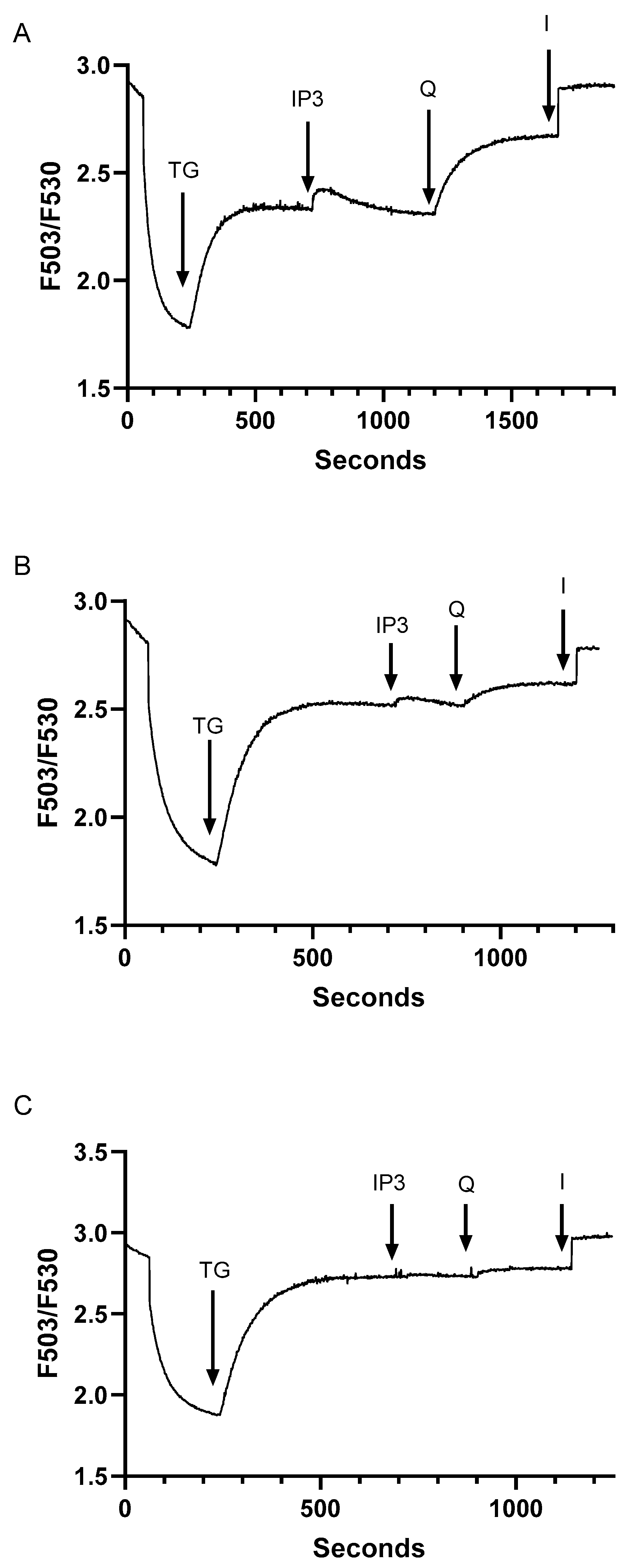
Figure 4.
Relationships of the Ryanodine and Thrombin releasable Ca2+ pools to the low dose TG SERCA 2b and low dose tBHQ SERCA 3 regulated Ca2+ stores. For A-E, Jurkat T lymphocytes were loaded with Fura-2 and suspended in Ca2+-free media (balanced salt solution plus 2 mM EGTA). A, Jurkat cell Ca2+ release responses induced by the sequential application (arrows) of ryanodine (Ry, 30 μM) tBHQ (Q, 2 μM), TG (200 pM), and ionomycin (I, 1 μM) as determined by the ratio of fluorescence changes at 340 and 380 nm (F340/380). B, the same experiment as in A but with the sequential application to Jurkat cells of ryanodine tBHQ (Q, 2 μM), ryanodine (Ry, 30 μM), TG (200 pM), and ionomycin (I, μM). C, Ca2+ release responses to the sequential addition of tBHQ (Q, 2 μM), thrombin (Thr, 0.1 U/ml), TG (200 pM) and ionomycin (1 μM). D, Ca2+ release responses to the sequential addition of TG (200 pM), thrombin (Thr, 0.1 U/ml), tBHQ (Q, 2 μM), and ionomycin (1 μM). E, Ca2+ release responses to the sequential addition of tBHQ (Q, 2 μM), TG (200 pM), thrombin (Thr, 0.1 U/ml), and ionomycin (1 μM). Fluorescence traces shown are representative of four to seven individual experiments.
Figure 4.
Relationships of the Ryanodine and Thrombin releasable Ca2+ pools to the low dose TG SERCA 2b and low dose tBHQ SERCA 3 regulated Ca2+ stores. For A-E, Jurkat T lymphocytes were loaded with Fura-2 and suspended in Ca2+-free media (balanced salt solution plus 2 mM EGTA). A, Jurkat cell Ca2+ release responses induced by the sequential application (arrows) of ryanodine (Ry, 30 μM) tBHQ (Q, 2 μM), TG (200 pM), and ionomycin (I, 1 μM) as determined by the ratio of fluorescence changes at 340 and 380 nm (F340/380). B, the same experiment as in A but with the sequential application to Jurkat cells of ryanodine tBHQ (Q, 2 μM), ryanodine (Ry, 30 μM), TG (200 pM), and ionomycin (I, μM). C, Ca2+ release responses to the sequential addition of tBHQ (Q, 2 μM), thrombin (Thr, 0.1 U/ml), TG (200 pM) and ionomycin (1 μM). D, Ca2+ release responses to the sequential addition of TG (200 pM), thrombin (Thr, 0.1 U/ml), tBHQ (Q, 2 μM), and ionomycin (1 μM). E, Ca2+ release responses to the sequential addition of tBHQ (Q, 2 μM), TG (200 pM), thrombin (Thr, 0.1 U/ml), and ionomycin (1 μM). Fluorescence traces shown are representative of four to seven individual experiments.
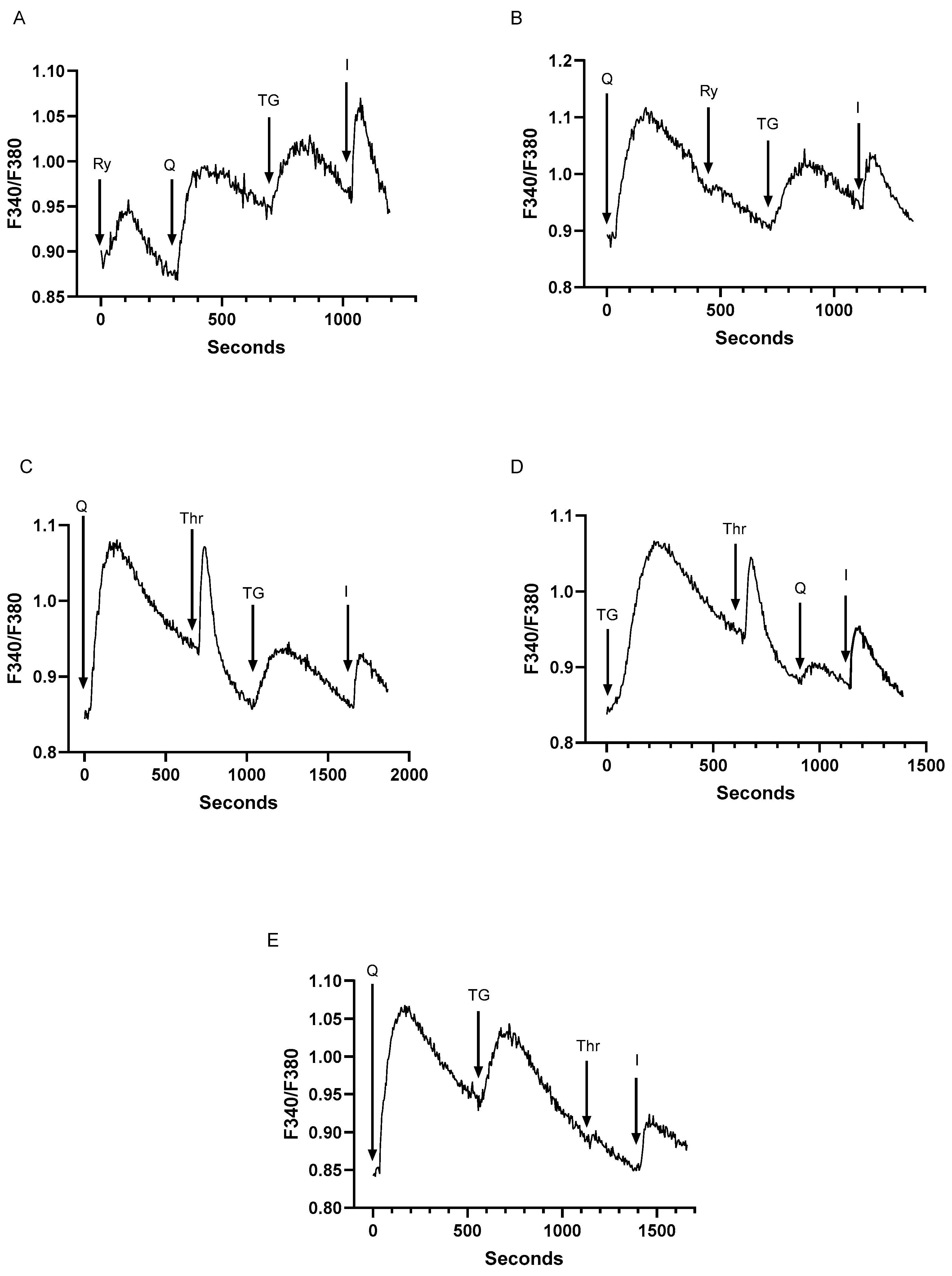
Figure 5.
T lymphocyte SERCA 2b and SERCA 3-regulated Ca2+ stores reveal similar sensitivities to cytoskeletal disruption with enhanced Ca2+ influx responses and reduced Ca2+ release activity. For A-E, Jurkat T lymphocytes were loaded with Fura-2 and suspended in Ca2+-free media (balanced salt solution plus 2 mM EGTA). A, Cells were suspended in Ca2+-free conditions (open bar) and stimulated (arrow) with TG (100 pM) or tBHQ (Q, 1 μM). After approximately 400 seconds of Ca2+ release activity, Ca2+ levels were increased to 2 mM to elicit Ca2+ influx responses (hatched bar). B and C, Ca2+ influx responses were induced as in A (hatched bars) in cell populations pre-incubated for 60 minutes in the presence (solid lines) or absence (dashed lines) of cytochalasin D (10 μM). D and E, Jurkat lymphocytes suspended in Ca2+-free media were stimulated with TG (D, 200 pM) and tBHQ (E, 2 μM) in the presence (CytD) or absence (Con) of cytochalasin D (60 min, 10 μM). Fluorescence traces shown are representative of four to six individual experiments.
Figure 5.
T lymphocyte SERCA 2b and SERCA 3-regulated Ca2+ stores reveal similar sensitivities to cytoskeletal disruption with enhanced Ca2+ influx responses and reduced Ca2+ release activity. For A-E, Jurkat T lymphocytes were loaded with Fura-2 and suspended in Ca2+-free media (balanced salt solution plus 2 mM EGTA). A, Cells were suspended in Ca2+-free conditions (open bar) and stimulated (arrow) with TG (100 pM) or tBHQ (Q, 1 μM). After approximately 400 seconds of Ca2+ release activity, Ca2+ levels were increased to 2 mM to elicit Ca2+ influx responses (hatched bar). B and C, Ca2+ influx responses were induced as in A (hatched bars) in cell populations pre-incubated for 60 minutes in the presence (solid lines) or absence (dashed lines) of cytochalasin D (10 μM). D and E, Jurkat lymphocytes suspended in Ca2+-free media were stimulated with TG (D, 200 pM) and tBHQ (E, 2 μM) in the presence (CytD) or absence (Con) of cytochalasin D (60 min, 10 μM). Fluorescence traces shown are representative of four to six individual experiments.
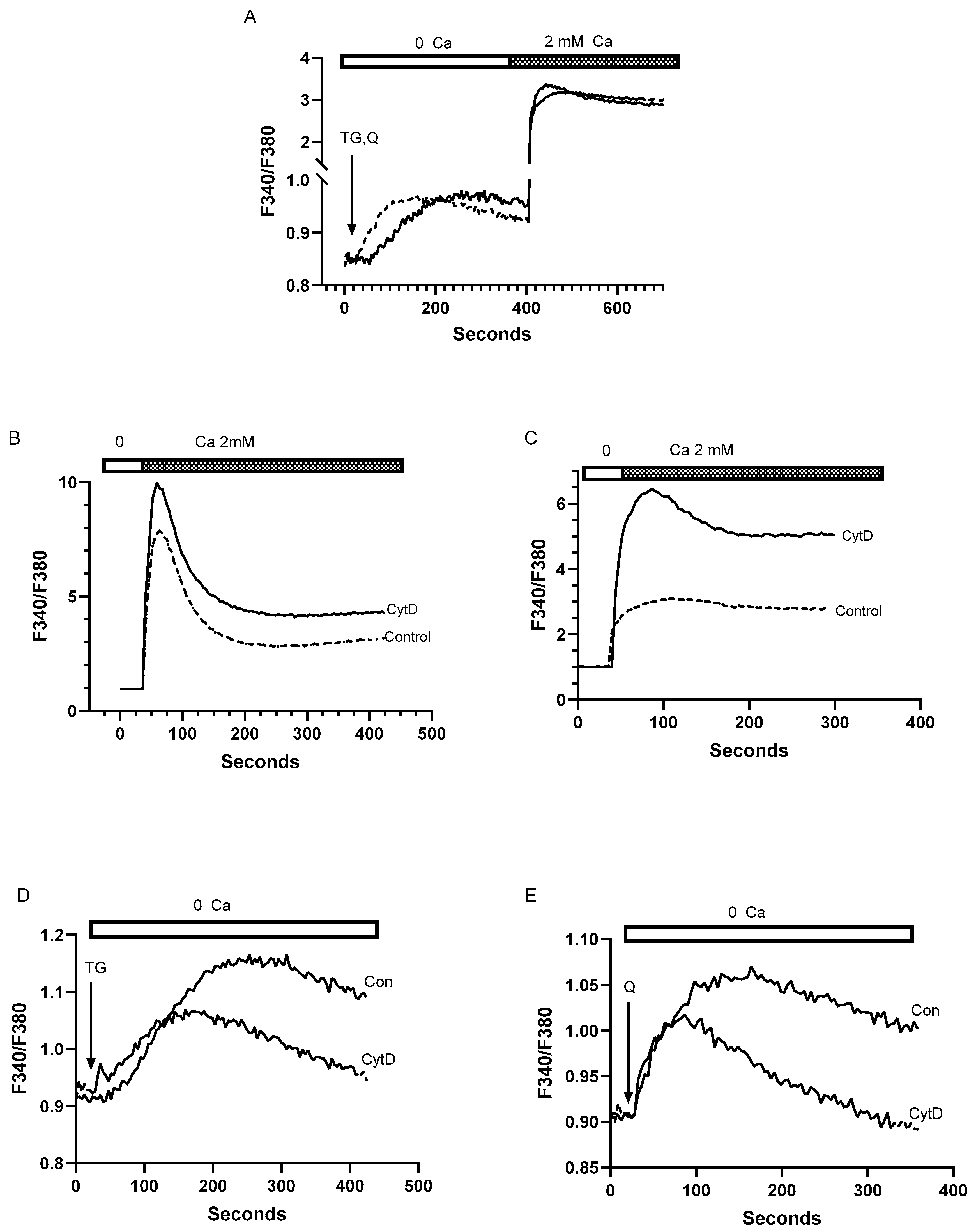
Figure 6.
The SERCA activator CDN1163 exerts differential time-dependent effects on T cell SERCA 2b and SERCA 3 Ca2+ stores. For A-C Jurkat T lymphocytes were loaded with Fura-2 and suspended in Ca2+-free media (balanced salt solution plus 2 mM EGTA). A, Ca2+ release responses induced by treatment with PHA (10 μg/ml) in the presence (dashed trace) or absence (solid trace) of CDN1163 (10 μM, 20 minutes). B, TG (100 pM) induced Ca2+ release in the presence (dashed traces, 0 and 20 min) or absence (solid trace) of CDN1163 (10 μM). C, tBHQ (1 μM) induced Ca2+ release in the presence (dashed trace, 20 min) or absence (solid trace) of CDN1163 (10 μM). D, Fluo-3 fluorescence Ca2+ uptake assay. Ca2+ uptake in ER stores was initiated by the addition of ATP (see Materials & Methods section) in saponin permeabilized Jurkat lymphocytes incubated in the presence (dashed curves, 5 and 30 minutes) or absence (solid curve) of CDN1163 (25 μM). Rate of Ca2+ uptake was estimated based on the linear intial rate of Fluo-3 fluorescene decay. E and F, Ca2+ release responses induced by TG and tBHQ in Jurkat lymphocytes incubated for longer durations with CDN1163. E, Ca2+ release induced by TG (100 pM) in the presence (dashed trace, 72 hours) or absence (solid trace) of CDN1163 (10 μM). F, Ca2+ release induced by tBHQ (1 μM) in the presence (dashed trace, 72 hours) or absence (solid trace) of CDN1163 (10 μM). G and H show experiments using saponin permeabilized Jurkat cells, depicting Ca2+ release responses as detected by Fuo-3 fluorescence changes. G, Ca2+ release induced in permeabilized cells treated with TG (1 nM) in the presence (dashed trace, 72 hours) or absence (solid trace) of CDN1163 (10 μM). H, Ca2+ release induced in permeabilized cells treated with tBHQ (1 μM) in the presence (dashed trace, 72 hours) or absence (solid trace) of CDN1163 (10 μM). I, Ca2+ uptake in ER stores was initiated by the addition of ATP (see Materials & Methods section) in saponin permeabilized Jurkat lymphocytes incubated in the presence (dashed curve, 12 hours) or absence (solid curve) of CDN1163 (25 μM). Rate of Ca2+ uptake was estimated based on the linear intial rate of Fluo-3 fluorescene decay. Fluorescence traces shown are representative of six to eight individual experiments for both permeabilized and intact cell assays.
Figure 6.
The SERCA activator CDN1163 exerts differential time-dependent effects on T cell SERCA 2b and SERCA 3 Ca2+ stores. For A-C Jurkat T lymphocytes were loaded with Fura-2 and suspended in Ca2+-free media (balanced salt solution plus 2 mM EGTA). A, Ca2+ release responses induced by treatment with PHA (10 μg/ml) in the presence (dashed trace) or absence (solid trace) of CDN1163 (10 μM, 20 minutes). B, TG (100 pM) induced Ca2+ release in the presence (dashed traces, 0 and 20 min) or absence (solid trace) of CDN1163 (10 μM). C, tBHQ (1 μM) induced Ca2+ release in the presence (dashed trace, 20 min) or absence (solid trace) of CDN1163 (10 μM). D, Fluo-3 fluorescence Ca2+ uptake assay. Ca2+ uptake in ER stores was initiated by the addition of ATP (see Materials & Methods section) in saponin permeabilized Jurkat lymphocytes incubated in the presence (dashed curves, 5 and 30 minutes) or absence (solid curve) of CDN1163 (25 μM). Rate of Ca2+ uptake was estimated based on the linear intial rate of Fluo-3 fluorescene decay. E and F, Ca2+ release responses induced by TG and tBHQ in Jurkat lymphocytes incubated for longer durations with CDN1163. E, Ca2+ release induced by TG (100 pM) in the presence (dashed trace, 72 hours) or absence (solid trace) of CDN1163 (10 μM). F, Ca2+ release induced by tBHQ (1 μM) in the presence (dashed trace, 72 hours) or absence (solid trace) of CDN1163 (10 μM). G and H show experiments using saponin permeabilized Jurkat cells, depicting Ca2+ release responses as detected by Fuo-3 fluorescence changes. G, Ca2+ release induced in permeabilized cells treated with TG (1 nM) in the presence (dashed trace, 72 hours) or absence (solid trace) of CDN1163 (10 μM). H, Ca2+ release induced in permeabilized cells treated with tBHQ (1 μM) in the presence (dashed trace, 72 hours) or absence (solid trace) of CDN1163 (10 μM). I, Ca2+ uptake in ER stores was initiated by the addition of ATP (see Materials & Methods section) in saponin permeabilized Jurkat lymphocytes incubated in the presence (dashed curve, 12 hours) or absence (solid curve) of CDN1163 (25 μM). Rate of Ca2+ uptake was estimated based on the linear intial rate of Fluo-3 fluorescene decay. Fluorescence traces shown are representative of six to eight individual experiments for both permeabilized and intact cell assays.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



