
Submitted:
08 February 2024
Posted:
12 February 2024
You are already at the latest version
Abstract
Liver is structurally organized into zonation where LSECs endothelial cells play a crucial role during chronic liver injury and early stages of fibrosis. Fibrosis can be reversed if diagnosed early at molecular level in zonation before progressing to advanced stages like bridging fibrosis. This study identified zonation marker genes using scRNA-seq and spatial transcriptomics molecular profiling technologies in normal and diseased fibrotic human liver. DGE analysis is performed over LSECs and identified the top 20 expressed genes in periportal, perivenous, and intermediate acinar zones. Multi-omics and scRNA-seq analysis over Visium images and ECs liver cells figured out OIT3, DNASE1L3, CLEC4G, LYVE1, FCN2, CRHBP as commonly expressed mid-lobular zonation-specific genes. Also, this study detected STAB2, F8, AQP1, TEK, TIMP3, TIE1, CTSL genes as expressed in DILI and NASH ECs populations. The connection between LSECs marker genes in zone 2 and liver fibrosis holds significant promise for advancing our understanding in developing new therapeutic strategies for fibrosis reversal and designing computational molecular biomarkers in NASH and DILI fibrotic liver diseases.
Keywords:
NASH
; DILI
; LSEC
; DGE
; liver zonation marker
; bridging fibrosis
; spatial transcriptomics
; liver injury
; computational biomarker
1. Introduction
In a recent report, annually liver disease contributes around 4% of all the deaths worldwide and accounts for over two million deaths. The global burden of liver disease is placing a significant strain on public healthcare systems. Cirrhosis and Hepatocellular Carcinoma (HCC) are major contributors including Nonalcoholic fatty liver disease (NAFLD) and Drug-induced liver injury (DILI) which significantly contributes as common cause of liver related deaths[1].
The liver is the largest organ in the human body responsible for vital functions including metabolism of nutrients and xenobiotics. The liver is structurally made-up of small anatomical units called liver lobules. Every lobule is histologically organized in a unique hexagonal architectural arrangement of hepatocytes (parenchymal cells PCs) into three zones that exhibit a distinct functionality known as liver zonation. Venous blood from the gut mixes at portal vein with oxygenated atrial blood and flows towards central vein through sinusoids. The liver tissue area around portal triad i.e., peri-portal recognized as Zone1 (oxygen enriched) is involved in metabolic functions and specialized in gluconeogenesis whereas tissue area around central vein recognized as zone3 (less oxygen) is involved in bile acid production drug metabolism and glycolysis. The area in between i.e., mid-lobular known as Zone 2 is a transitional region where concentration of oxygenated blood, nutrients, metabolites, and gut-derived toxins varies along portal-central vein axis. Spatially heterogenous hepatocytes are distinguished by the gene expression profiles where 50% of the genes are expressed along lobular zonation axis[2]. Zone 2 is believed to plays an important role in homeostatic renewal of hepatocytes, liver mass regeneration and proliferation upon liver injury [3,4]. Spatial heterogeneity of parenchymal and non- parenchymal cells (NPCs) is zone dependent which modulate the Differentially Expressed Genes (DGE) expression and initiate several liver diseases including Nonalcoholic steatohepatitis (NASH), DILI, HCC and liver regeneration[5].
Other than hepatocytes, Liver Sinusoidal Endothelial Cells (LSECs) contribute around 15–20% of total number of liver cells are highly specialized non parenchymal endothelial cells and act as a physical barrier between blood substrates and hepatocytes. Liver endothelial cells (ECs) includes LSEC, vascular ECs, and lymphatic ECs (LyECs)[6,7]. As chronic liver disease advances, hepatocyte functioning impaired by crosstalk between other liver cells which initiates an important role in regulating fibrosis. LSECs have unique fenestrae (pores) that allow for efficient clearance of pathogens, debris, and toxins from the blood, keeping the liver clean and functional. LSECs act as determinants of hepatic fibrosis where the process of capillarization precedes fibrosis in which LSECs lack fenestration and develop an organized basement membrane[8]. LSECs are the major drivers in fibrosis[9] and differential gene expression is observed between the different zones of the liver lobule during fibrosis[10]. For example, the periportal zone 1 expresses genes involved in the uptake of nutrients, mid-lobular zone2 expresses genes involved in metabolism of nutrients, pericentral zone 3 expresses genes involved in the secretion of bile. Gradient of differentially expressed genes (DGE) is observed in LSECs and hepatocytes following disrupted zonation architecture and liver functionalities in most liver diseases including NAFLD, NASH and DILI. DGE manifests around pericentral zone 3 that later progresses towards advanced stages of fibrosis, bridging fibrosis and cirrhosis [11,12]. Liver bridging fibrosis, a specific advanced NASH feature, is a type of scarring caused by the accumulation of excess collagen around hexagonal portal triad-central vein region in the later stages of liver fibrosis. These collagen bands connect different areas of the liver and block the flow of blood and bile, which leads to liver failure. Early liver fibrosis, a condition where scar tissue builds up, can indeed be reversed if detected early in zonation at molecular level and addressed promptly. However, it’s crucial to remember that this reversal is contingent on preventing the progression to more advanced stages like bridging fibrosis, where the scar tissue becomes more organized, extensive and the damage is irreversible.
It’s challenging to measure zonation-specific structural variations during early stages of fibrosis and later at bridging fibrosis level. Our knowledge is limited and poorly understood on the role of 1) zone 2 LSECs in early liver fibrosis, 2) LSECs-specific marker genes in hepatotoxic DILI conditions, 3) DGE profiles within unclear boundaries of liver zones under normal, NASH and DILI disease fibrotic patterns. In this study, efforts are made to computationally characterize and quantify spatial heterogeneity of hepatocytes at molecular level using single cell RNA seq and spatial molecular imaging techniques to develop our understanding of zonal restructuring, role of LSECs and marker genes in normal and diseased fibrotic conditions.
2. Results
2.1. Histopathology H&E Image Classification Experimental Workflow for Early and Bridging Fibrosis
Image analytics and experimental workflow, illustrated in Figure 1 D, is designed to classify Hematoxylin and Eosin (H&E) histopathology images under five categories i.e., normal, steatosis, early fibrosis, bridging fibrosis and cirrhosis. Machine learning widgets, shown in workflow, are executed to load, process, classify, cluster, and visualize the imaging data. After predictive analysis, images are clustered based on the common histologic disease features learned by the machine learning models, i.e., all the bridging fibrosis images grouped together under one group as shown in t-SNE plot Figure 1E and hierarchical clustering Figure 1B. Other category images are grouped similarly as per their disease morphology features. The Confusion matrix shown in Figure 1 F illustrates correctly predicted images (diagonally highlighted blocks) under respective categories. Closely clustered images share common morphological disease features in early fibrosis and steatosis (NASH specific feature) where white fat droplets are present in both images, shown as an example in Figure 1C. Classification models performance summarized in Table S1.
1.2. DGE Analysis for NASH and Zonation Expression
DGE analysis for NASH and zonation expression is performed over four independent studies GSE89632, GSE126848, GSE83990, GSE105127 using NCBI GEO2R analysis tool. Volcano plots for NASH vs Control are illustrated in Figure 2A, zone1 vs zone 3 expression are illustrated in Figure 2B. Volcano plot displays statistical significance (-log10 P value) versus magnitude of change (log2 fold change) of differentially expressed genes in NASH vs control and zonation-specific expression studies. The most up regulated genes identified after analysis in NASH are TYMS, FMO1, UQCRB; and in zone1 vs zone 3 analysis are HAL, AQP1 KRT19. Details of DGEs and top up/down expressed genes are summarized in Table S2.
2.3. Single Cell Clustering and DGEs Expression Profiles in DILI and NASH ECs
Single-cell transcriptomic analysis is performed over liver cells (n=9 i.e., three patients in each group) in control, DILI(intrinsic) and NASH datasets available at NCBI GEO GSE166178[13] and analyzed for the heterogeneity of inter- and intra-group endothelial cells in healthy and diseased mouse livers. Cell clustering results are illustrated in Figure 3A-B for control vs DILI, and control vs NASH illustrated in Figure 3C-D. DGEs expression is performed for eight commonly expressed genes STAB2, OIT3, F8, AQP1, TEK, TIMP3, TIE1, CTSL in DILI and NASH ECs populations, illustrated in Supplemental Figure S1.
2.4. Spatial Transcriptomics Data Analysis for Zonation Marker Genes
2.4.1. 10x Genomics Visium Image Analysis for Spatial Distribution of Zonation Expression Markers
Spatial transcriptomics (ST) image analysis is performed on human liver H&E stained healthy images using 10x genomic`s Visium platform to identify zonation patterns based on known markers[14]. 10x genomics uses space ranger tool for automated image detection to determine the position of fiducial markers and to identify tissue boundaries. Expressed dots(red) are the 55-micron Visium spots from where transcriptomes (RNA) are extracted for studying spatial cellular populations present and gene expressions at that specific tissue location. Figure 4B-4D illustrates zonation-specific expressed genes such as FGG, SAA1, and GLUL overlaid Visium liver tissue image[15]. Expression profiles unraveled zonation expressions in peri-portal, mid-lobular and peri-central zones.
2.4.2. Spatial Molecular Imaging to Demonstrate Liver Zonation Architecture
Nanostring`s CosMxTM Spatial Molecular Imager (SMI) platform, illustrated in Figure 5A is a high-plex spatial multiomics image generated from a 5 µm thick human formalin-fixed paraffin-embedded (FFPE) liver section stained for both protein and RNA analytes to demonstrate zone1, zone2 and zone 3 architecture. Figure 5B-5D illustrates differential gene expression with three target markers i.e., FGG, SAA1 & GLUL to highlight the zonation boundaries in a healthy liver tissue at molecular resolution. Similar equivalent patterns demonstrated in 10x genomics Visium images for the same three zonation-specific genes which highlights zonation boundaries in a normal human liver. However, in the diseased and chronic liver injury conditions, this zonation disruption exhibits varying DGE expression.
2.5. Liver Cell Clustering and LSECs Markers
Endothelial cell (ECs) clustering data analysis is executed here to demonstrate LSECs gene expression markers on an integrated scRNA-seq data of 28 healthy human liver samples. Single-cell scRNA-seq data used here was acquired from separate five independent studies reported elsewhere in [16]. Figure 6A illustrates UMAP clustering of all the normal liver cells. LSECs cluster shown in Figure 6B are picked as subset of clustered ECs cells shown in Figure 6A. Vascual Central Venous ECs (VCVEC) and Vascual Portal Venous ECs (VPEC) are clustered along-with LSECs, shown in Figure 6C which further separated as subsets and highlighted using ECs and LSECs separately with CLEC4G, FCN2, OIT3, LYVE1 expression marker genes illustrated in Figure 6D-6E. VCVEC and VPEC clusters further separated and highlighted using MGP, VWF, CD34 expression marker genes, illustrated in Figure 6F. VCVEC and VPEC shown in Figure 6 C-D are identified based on the list of endothelial cell markers selected from the FindAllMarker() list and plotted at National Institute of Health`s (NIH), Human BioMolecular Atlas Program (HuBMAP) Azimuth app which is a Seurat based web application tool, that provides clustering of all human normal liver cells (including VCVEC and VPEC).
2.6. DGE Expression Profiles in Zonated LSECs
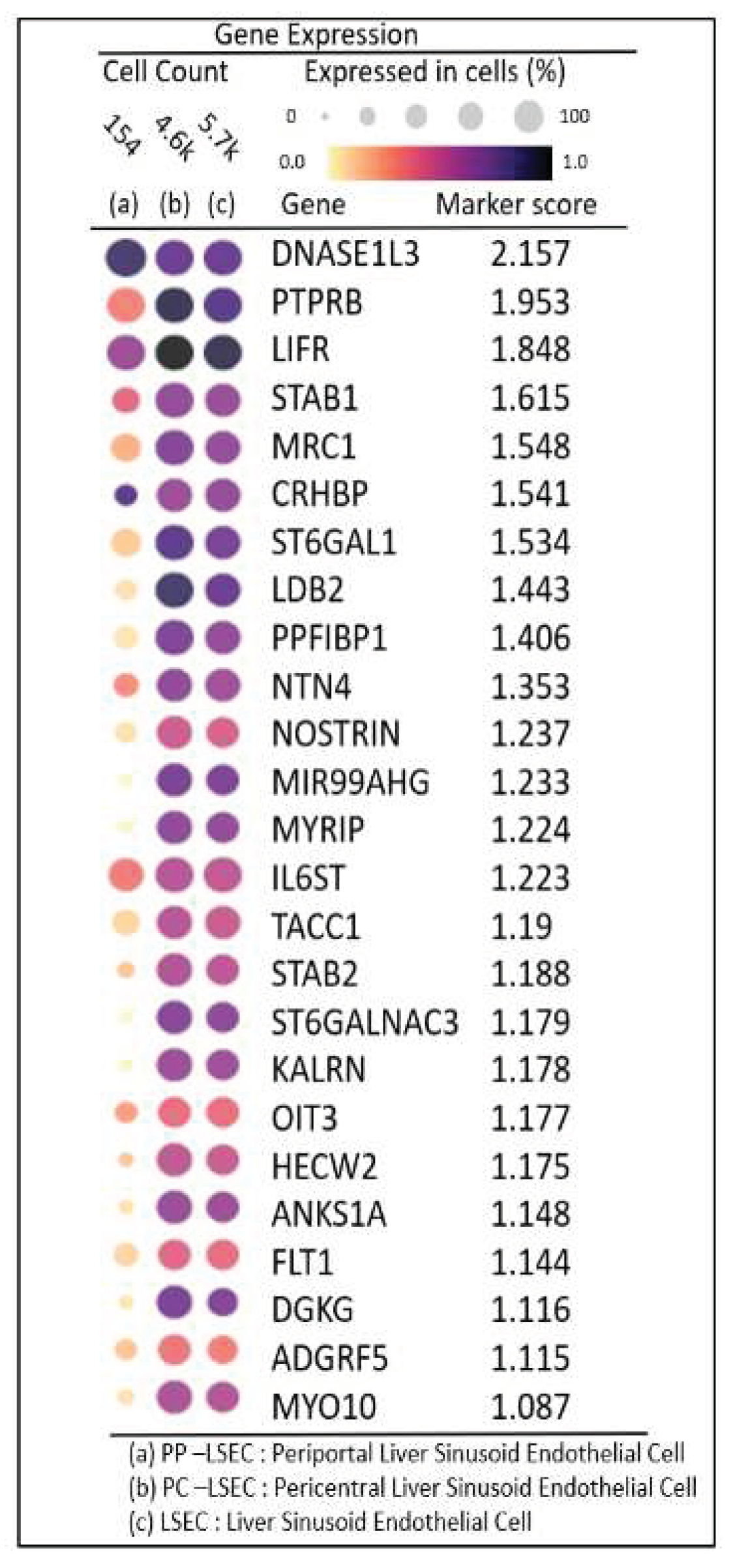
DGE expression profiles identified the top 25 highly expressed genes with their marker score in PP–LSEC: Periportal Liver Sinusoid Endothelial Cells, PC–LSEC: Pericentral Liver Sinusoid Endothelial Cells and LSECs cells illustrated in Figure 7. The combination of two metrics represented in gene expression dot plots i.e., gene expression and percentage of expressing cells, enables us to assess gene expression within a grid of genes by cell types. The genes that are expressed in a small percentage of cells may be difficult to visually identify in a dot plot. This is particularly important for certain marker genes which are specifically but lowly expressed in their target cell types, for example transcription factors and cell-surface receptors.
A marker score is the 10th percentile of the effect sizes across all comparisons for each gene. DGE analysis is performed at cellxgene web-tool, a well curated, standardized, wide collection of interoperable single-cell transcriptomic data platform available at cellxgene.cziscience.com[17]. Five most commonly expressed genes DNASE1L3,LIFR,STAB1,MRC1,CRHBP are detected using cellxgene analysis and confirmed in another datasets[14]. Also, identified top 20 expressed genes in each zone are summarized in Table S3. The gene list is selected by the single cell analysis algorithm based on the markers detected through “FindAllMarkers()” function in Seurat single cell analysis R- package. This function identifies differentially expressed genes for each of the identity classes (clusters) in the supplied dataset and builds a list of expressed genes in the given dataset. From this list, 60 genes are picked (tabulated in Table S3), expressed in endothelial cells (20 in each zone). Their zonation expression is further validated in zonation browser tool and plotted, shown in supplementary Figure S2. Commonly expressed genes in this group are MRC1, HAL, TIMP1 (zone1), DNASE1L3, CRHBP, C9 (zone2) and SELE, APOB, GLUL (zone3). Their zonation-specific expression is confirmed using the human hepatocyte zonation browser tool, another open-source web platform publicly available web tool [18], illustrated in supplementary Figure S2.
3. Discussion
In this study, efforts are made to develop our understanding towards complex interplay of LSECs in mid-lobular zone2 and their contributions in initiation of early fibrosis and regeneration. LSECs presence in zone 2 is influential and play a critical role in determining whether injured hepatocytes will regenerate or susceptible to fibrosis. Their anti-fibrotic ability to support regeneration is crucial for a successful liver repair process. LSECs secrete extracellular matrix (ECM) components like laminins, providing a supporting microenvironment for regeneration and ensuring proper tissue restructuring [19].
It’s important to understand the zonation LSECs functioning at cellular and molecular level and to quantify those architectural changes spatially in normal healthy and diseased liver conditions. Scarring tissue in NASH and DILI disease models exhibit common features of liver fibrosis. In this study, OIT3 is computationally identified as commonly expressed marker gene in mid-lobular zone2 by LSECs cells population both in NASH and DILI datasets, which is also reported elsewhere in a study as a hallmark gene expressed in ECs [20]. The second most prominent gene identified is DNASE1L3. Other zonation-specific marker genes identified in our study are MRC1, HAL (zone1), CRHBP, LYVE1(zone2) and SELE, GLUL (zone3). Protein F8 secreted by LSECs[21] plays important role in blood clotting is another marker gene detected in our experiment on GSE166178 dataset as highly expressed zone 2 marker gene and confirmed by zonation browser tool. Also found in this series STAB2,CLEC4G, LYVE1 genes as mid-lobular zone 2 markers and confirmed as expressed healthy human liver[6,7]. Gene AQP1 ( Aquaporin-1 ) promotes angiogenesis, fibrosis and portal hypertension and reported as over expressed in endothelial cells in advanced fibrotic stages such as cirrhosis[22], which is exhibiting similar trends of over expression in our dataset , illustrated in supplementary Figure S1. Also, LYVE1 was found among the genes with increased expression suggests several possibilities such as a) Lymphangiogenesis: Might promote the formation of new lymphatic vessels, potentially contributing to inflammation and fibrosis progression, b) Endothelial cell activation: LYVE1 upregulation could indicate a shift in the function of sinusoidal endothelial cells towards a more pro-fibrotic phenotype, c) Scar tissue remodeling: Lymphatic vessels may play a role in clearing collagen debris and remodeling scar tissue during the later stages of fibrosis.
The liver displays unique spatial heterogeneity and functioning of hepatocytes by exhibiting distinct metabolic and functional profiles across different acinar zones (periportal, perivenous, and intermediate). Specific enzymes, transporters, and other proteins act as zonation markers, reflecting the specialized functions of each zone. 10x genomic`s Visium platform and Nanostring’s CosMx technologies used here to demonstrate these changes at molecular level and confirmed by zonation browser tool as the zonation-specific i.e., GLUL as zone3, FGG as zone2 and SAA1 as zone 1 expressed marker genes.
H&E histopathology images are still considered as gold standard in pathology for the analysis and detection of early fibrotic lesions. Also, 10x genomics Visium platform uses tissues for extracting transcriptome & histopathology H&E-stained images to align Visium spots (extracted transcriptome locations) for studying cellular heterogeneity and spatial analysis. Fibrosis, which is a formation of scar tissue in response to liver injury or disease. Machine learning based histopathology image classification model is designed here to understand these fibrosis development stages on H&E-stained images i.e., starting from normal lobular liver architecture, and then how zonal pattern of fibrosis (collagen fibers) develops in early fibrosis, bridging fibrosis and cirrhosis. This approach can be used further for the prediction of fibrosis in other datasets used in this study.
DGE analysis of liver zonation can provide valuable insights into the mechanisms and identification of potential therapeutic targets for fibrosis in NASH and DILI. In Figure 3D, single cell analysis exhibits reduction in endothelial cell numbers which is mostly observed in the NASH patients. Potential mechanisms may explain the observed reduction in endothelial cell numbers in NASH liver such as 1) Oxidative stress: NASH is characterized by increased oxidative stress and endoplasmic reticulum (ER) stress, which can trigger endothelial cell apoptosis and necrosis, 2) Inflammatory response: Chronic inflammation in NASH liver can release pro-inflammatory cytokines and chemokines, further promoting endothelial cell damage and apoptosis. Single-cell data has revealed interactions between endothelial cells and macrophages in NASH, suggesting these interactions might contribute to endothelial cell loss in NASH. Furthermore, macrophages in DILI are not a homogenous population. Depending on the microenvironment and stimuli, they can adopt different phenotypes with distinct functions. DILI attracts pro-inflammatory cells like macrophages, which can further damage endothelial cells and impair their normal function. Single cell analysis in DILI illustrated in Figure 3B, shows a growing trend of macrophages, that could be due to their activated role often associated as pro-inflammatory in DILI disease model[23].
In NASH related fibrosis, DGEs in Zone 1 might be related to lipid metabolism and oxidative stress, while Zone 3 DGEs could be involved in inflammation and bile acid signaling. Similarly in a specific DILI drug, DGEs might be related to mitochondrial dysfunction, immune response, or direct cell injury in specific zones. Fibrosis is caused by long-term chronic liver injury and is considered a hallmark disease feature of both NASH and DILI progression. This disrupts liver architecture and function, leading to potential organ failure. Different genes might be driving fibrosis in each zone, suggesting targeted therapies for each zonated area.
DILI is a challenging disease to diagnose, a leading cause of acute liver failure, and responsible for drug withdrawal from the market. Computational biomarkers could be helpful as emerging new technologies in diagnosis of DILI and NASH related fibrotic patterns[24]. DGEs could serve as early diagnostic or prognostic biomarkers for fibrosis progression in NASH and DILI. DGE analysis in NCBI GEO GSE126848 study identified UQCRB as the most up regulated gene in NASH and confirmed as a zone 2 marker, which is reported in a study elsewhere as molecular prognostic biomarker in human colorectal cancer[25]. DGE expression profiles identified five other commonly expressed genes such as DNASE1L3, LIFR, STAB1, MRC1, CRHBP. Table summarized the top 25 highly expressed genes in PP–LSEC and PC-LSEC, such as MRC1, HAL, TIMP1 (zone1), DNASE1L3, CRHBP, C9 (zone2) and SELE, APOB, GLUL (zone3).
4. Methods and Materials
4.1. H&E Histopathology Image Classifications
Open-source visual programming data mining framework orange toolbox is used for classification of H&E histopathology image tiles. Orange is a machine learning data mining platform (http://orange.biolab.si) for image analysis and data visualization[26,27]. H&E stained Whole Slide (WSI) histopathology images were acquired from open-source National Cancer Institute (NCI) USA Biorepositories and Biospecimen Research Branch’s (BBRB) Genotype-Tissue Expression (GTEx)Tissue Image library of annotated WSI with clinical data publicly available at https://brd.nci.nih.gov/brd/specimen/GTEX-117XS-0926 and downloaded under five categories as normal, steatosis, early fibrosis, bridging fibrosis & cirrhosis. Each WSI were cropped into tiles for training machine learning models and classified in orange tool classification layout. Experimental workflow is shown in Figure 1D.
4.2. Spatial Transcriptomics Data Analysis
4.2.1. Visium Data Analysis and Visualization
The Visium dataset used here is available at the Gene Expression Omnibus NCBI GEO public database under accession number GSE192742. Visium 10x genomics loupe browser image files downloaded from liver cell atlas available at https://www.livercellatlas.org/download.php The Visium ST images of normal human liver samples were processed using Loupe Browser 7.0.1 (10x Genomics Inc.) downloaded from here https://www.10xgenomics.com/support/software/loupe-browser/downloads. The gene expression data for the k-mean clusters generated by the Space Ranger software for up regulated genes. The data consists of the median-normalized average of gene expression, log2 fold changes, and statistical significance (p values) computed for genes with a p value < 0.05.
4.2.2. Spatial Molecular Imaging
Spatial molecular imaging highlights DGE in human liver zonation. Imaging data were acquired from an open-source publicly available multiplex dataset at http://nanostring.com/CosMx-dataset CosMxTM Spatial Molecular Imager which includes staining for a panel of morphological features.
4.3. Liver Cell Clustering and Analysis Tool
For liver cell clustering and LSECs marker analysis, National Institute of Health`s (NIH) USA, Human BioMolecular Atlas Program (HuBMAP) Azimuth app is used. Azimuth is a Seurat based web application tool that uses an annotated reference dataset to automate the processing, analysis, and interpretation of single-cell RNA-seq data. Azimuth utilizes ‘reference-based mapping’ pipeline that accepts counts matrix file in multiple formats as input and performs normalization, visualization, cell annotation and DGE expression without any coding requirement on web cloud. All results can be visualized within the app, and can be downloadable for additional downstream analysis[28,29,30,31,32]. The development of Azimuth is led by the New York Genome Center Mapping Component as part of the NIH HuBMAP consortium.
4.4. LSECs Markers
Cellxgene Differential Expressed Gene (DGE) tool is used to run expression analysis for the cell types of PP-LSEC, PC-LSEC and LSECs cells. CellxGene is an open-source public database available at https://cellxgene.cziscience.com/ ; a suite of tools to run analysis of single-cell transcriptomic dataset. List of ECs expressed exclusively for LSECs cell populations are acquired from liver single cell expression atlas of 28 human livers[16] available at http://liveratlas-vilarinholab.med.yale.edu/ and plotted on LSECs HuBMAP Azimuth reference-based single-cell analysis tool for expression analysis. The analyzed results then compared with zonation browser tool to see the zonation profiles of same expressed genes in mid-lobular zone2. All three plots are illustrated in supplemental Figure S3. Zonation browser tool is available at https://itzkovitzwebapps.weizmann.ac.il/webapps/home/session.html?app=HumanandMouseHepatocyteZonation.
5. Conclusions
Mammalian healthy liver lobule is spatially well zonated and recognized with known markers based on distribution of metabolic functions that disrupts in diseased pathological conditions and liver injury. However, due to hepatocyte heterogeneity these zonation markers, such as enzymes, metabolites, and gene expression patterns vary in their expression and activity even within the same lobule. These molecular markers are not 1) zonation-specific; 2) disease -specific, for example, the expression and distribution of specific molecules within the liver changes during fibrogenesis in NASH and DILI. LSECs plays an important and intricated role in the early stages of liver fibrosis and regeneration during mid-lobular zonation restructuring. Their strategic position allows them to act as critical conductors in influencing the delicate balance between tissue repair and scarring. Emerging trends in new technologies such as scRNA-seq, spatial transcriptomics and multiomics can be helpful to determine the role of LSECs in identifying the zonation marker DGEs at cellular and molecular level. LSECs can provide valuable insights into how zonation marker variations and fibrosis are interconnected. The goal of this study is mostly to establish a baseline to understand which normal liver zones might experience changes, specifically the mid-lobular zone 2 [3] during early fibrosis, liver injury conditions or at the stages of advanced fibrosis.
This study identified zonation-specific mid-lobular markers of LSECs. STAB2 family genes are the most highly expressed genes in liver LSECs[33] related to fibrosis detected by this study, illustrated in supplementary Figure S1. Liver fibrosis in early stages can be reversible if detected in zonation at molecular level before moving to advanced stages like bridging fibrosis. Current diagnostic methods often lack accuracy at early stages of liver fibrosis. Diagnosis of DILI in early stages is important but a complex and challenging task. Accurate evaluation and causality assessment are the key aspects of early detection. This is where computational biomarkers derived from integrated multi-omics, cellular and molecular data hold immense promise for providing a deeper understanding and improved diagnosis of NASH and DILI related liver fibrosis.
NASH and DILI are complex diseases and unraveling its secrets requires a powerful orchestra of technologies. Efforts are made here to leverage the latest advancements in single-cell analysis, histopathology, spatial transcriptomics, and multi-omics playing their part in deciphering cellular diversity and unveiling the cell state transitions involved in fibrosis developments. Integration of other technologies with histopathology which serves as a valuable platform for spatially correlating single-cell data with tissue morphology. This allows researchers to link gene expression patterns to specific cell locations and structures within the liver, adding a crucial spatial dimension to develop their understanding towards fibrosis developmental stages. By combining these powerful tools, researchers can create a rich, multidimensional picture of liver fibrosis in NASH and DILI. This holistic understanding can lead to Identification of novel therapeutic targets, improved diagnosis, prognosis, and development of personalized medicine by understanding the unique molecular signatures of individual patients.
Studying liver zonation fibrosis in NASH and DILI using new technologies offers significant novelty and holds great promise for advancing our understanding of these complex diseases. These new technologies offer a revolutionary approach to studying liver zonation fibrosis in NASH and DILI. Their ability to reveal previously hidden cellular and spatial heterogeneity, pinpoint molecular drivers, and elucidate disease mechanisms will significantly improve our understanding of fibrosis progression and ultimately lead to the development of more effective and personalized therapeutic strategies.
The limitation of this study is that DGE analysis alone is not sufficient to understand early stages of fibrosis in complex diseases like NASH and DILI. Integrating it with other data, such as protein expression and metabolic profiling is required to analyze in normal and diseased conditions. More research is needed to validate DGE as reliable zonation markers and translate them into effective diagnostic and prognostic biomarkers. Investigating LSECs in the context of both zonation and fibrosis holds significant promise for developing novel therapeutic strategies against liver diseases. This may help to protect zonation patterns, prevent fibrosis development, and ultimately improve liver health.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Data availability
the tool used for building machine learning image classification model is freely available (link provided in the text) open-source fully user friendly (no python coding required) GUI (Graphical User Interface) which works on drag-n-drop widgets (building blocks) method. Classifiers can be designed based on the workflow developed here, shown in Figure, ID. Information can be shared on trained classifier, designing methodologies, and building experimental workflow upon request.
Conflicts of Interest
No conflict of interest.
References
- Devarbhavi, H.; Asrani, S.K.; Arab, J.P.; Nartey, Y.A.; Pose, E.; Kamath, P.S. Global Burden of Liver Disease: 2023 Update. J. Hepatol. 2023, 79, 516–537. [Google Scholar] [CrossRef] [PubMed]
- Ben-Moshe, S.; Itzkovitz, S. Spatial Heterogeneity in the Mammalian Liver. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 395–410. [Google Scholar] [CrossRef] [PubMed]
- Itoh, T. The Truth Lies Somewhere in the Middle: The Cells Responsible for Liver Tissue Maintenance Finally Identified. Cell Regen. Lond. Engl. 2021, 10, 28. [Google Scholar] [CrossRef]
- Wei, Y.; Wang, Y.G.; Jia, Y.; Li, L.; Yoon, J.; Zhang, S.; Wang, Z.; Zhang, Y.; Zhu, M.; Sharma, T.; et al. Liver Homeostasis Is Maintained by Midlobular Zone 2 Hepatocytes. Science 2021, 371, eabb1625. [Google Scholar] [CrossRef]
- Panday, R.; Monckton, C.P.; Khetani, S.R. The Role of Liver Zonation in Physiology, Regeneration, and Disease. Semin. Liver Dis. 2022, 42, 001–016. [Google Scholar] [CrossRef] [PubMed]
- Verhulst, S.; van Os, E.A.; De Smet, V.; Eysackers, N.; Mannaerts, I.; van Grunsven, L.A. Gene Signatures Detect Damaged Liver Sinusoidal Endothelial Cells in Chronic Liver Diseases. Front. Med. 2021, 8, 750044. [Google Scholar] [CrossRef]
- Su, T.; Yang, Y.; Lai, S.; Jeong, J.; Jung, Y.; McConnell, M.; Utsumi, T.; Iwakiri, Y. Single-Cell Transcriptomics Reveals Zone-Specific Alterations of Liver Sinusoidal Endothelial Cells in Cirrhosis. Cell. Mol. Gastroenterol. Hepatol. 2021, 11, 1139–1161. [Google Scholar] [CrossRef]
- DeLeve, L.D. Liver Sinusoidal Endothelial Cells in Hepatic Fibrosis. Hepatol. Baltim. Md 2015, 61, 1740–1746. [Google Scholar] [CrossRef]
- Lafoz, E.; Ruart, M.; Anton, A.; Oncins, A.; Hernández-Gea, V. The Endothelium as a Driver of Liver Fibrosis and Regeneration. Cells 2020, 9, 929. [Google Scholar] [CrossRef]
- Ghallab, A.; Myllys, M.; Holland, C.H.; Zaza, A.; Murad, W.; Hassan, R.; A. Ahmed, Y.; Abbas, T.; A. Abdelrahim, E.; Schneider, K.M.; et al. Influence of Liver Fibrosis on Lobular Zonation. Cells 2019, 8, 1556. [Google Scholar] [CrossRef]
- Nagy, D.; Maude, H.; Birdsey, G.M.; Randi, A.M.; Cebola, I. RISING STARS: Liver Sinusoidal Endothelial Transcription Factors in Metabolic Homeostasis and Disease. J. Mol. Endocrinol. 2023, 71. [Google Scholar] [CrossRef] [PubMed]
- Ben-Moshe, S.; Veg, T.; Manco, R.; Dan, S.; Papinutti, D.; Lifshitz, A.; Kolodziejczyk, A.A.; Halpern, K.B.; Elinav, E.; Itzkovitz, S. The Spatiotemporal Program of Zonal Liver Regeneration Following Acute Injury. Cell Stem Cell 2022, 29, 973–989.e10. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Qian, J.; Lu, X.; Zhang, P.; Guo, R.; Lou, H.; Zhang, S.; Yang, J.; Fan, X. A Single-Cell Transcriptomic Atlas Characterizes Liver Non-Parenchymal Cells in Healthy and Diseased Mice; Genomics, 2021;
- Halpern, K.B.; Shenhav, R.; Matcovitch-Natan, O.; Tóth, B.; Lemze, D.; Golan, M.; Massasa, E.E.; Baydatch, S.; Landen, S.; Moor, A.E.; et al. Single-Cell Spatial Reconstruction Reveals Global Division of Labour in the Mammalian Liver. Nature 2017, 542, 352–356. [Google Scholar] [CrossRef] [PubMed]
- Hildebrandt, F.; Andersson, A.; Saarenpää, S.; Larsson, L.; Van Hul, N.; Kanatani, S.; Masek, J.; Ellis, E.; Barragan, A.; Mollbrink, A.; et al. Spatial Transcriptomics to Define Transcriptional Patterns of Zonation and Structural Components in the Mouse Liver. Nat. Commun. 2021, 12, 7046. [Google Scholar] [CrossRef]
- Brancale, J.; Vilarinho, S. A Single Cell Gene Expression Atlas of 28 Human Livers. J. Hepatol. 2021, 75, 219–220. [Google Scholar] [CrossRef]
- Program, C.S.-C.B.; Abdulla, S.; Aevermann, B.; Assis, P.; Badajoz, S.; Bell, S.M.; Bezzi, E.; Cakir, B.; Chaffer, J.; Chambers, S.; et al. CZ CELL×GENE Discover: A Single-Cell Data Platform for Scalable Exploration, Analysis and Modeling of Aggregated Data 2023, 2023.10.30.563174.
- Massalha, H.; Bahar Halpern, K.; Abu-Gazala, S.; Jana, T.; Massasa, E.E.; Moor, A.E.; Buchauer, L.; Rozenberg, M.; Pikarsky, E.; Amit, I.; et al. A Single Cell Atlas of the Human Liver Tumor Microenvironment. Mol. Syst. Biol. 2020, 16, e9682. [Google Scholar] [CrossRef]
- Natarajan, V.; Harris, E.N.; Kidambi, S. SECs (Sinusoidal Endothelial Cells), Liver Microenvironment, and Fibrosis. BioMed Res. Int. 2017, 2017, 4097205. [Google Scholar] [CrossRef]
- Li, Z.-W.; Ruan, B.; Yang, P.-J.; Liu, J.-J.; Song, P.; Duan, J.-L.; Wang, L. Oit3, a Promising Hallmark Gene for Targeting Liver Sinusoidal Endothelial Cells. Signal Transduct. Target. Ther. 2023, 8, 1–10. [Google Scholar] [CrossRef]
- Jamil, M.A.; Singer, H.; Al-Rifai, R.; Nüsgen, N.; Rath, M.; Strauss, S.; Andreou, I.; Oldenburg, J.; El-Maarri, O. Molecular Analysis of Fetal and Adult Primary Human Liver Sinusoidal Endothelial Cells: A Comparison to Other Endothelial Cells. Int. J. Mol. Sci. 2020, 21, 7776. [Google Scholar] [CrossRef]
- Huebert, R.C.; Jagavelu, K.; Hendrickson, H.I.; Vasdev, M.M.; Arab, J.P.; Splinter, P.L.; Trussoni, C.E.; LaRusso, N.F.; Shah, V.H. Aquaporin-1 Promotes Angiogenesis, Fibrosis, and Portal Hypertension Through Mechanisms Dependent on Osmotically Sensitive MicroRNAs. Am. J. Pathol. 2011, 179, 1851–1860. [Google Scholar] [CrossRef]
- Gerussi, A.; Natalini, A.; Antonangeli, F.; Mancuso, C.; Agostinetto, E.; Barisani, D.; Di Rosa, F.; Andrade, R.; Invernizzi, P. Immune-Mediated Drug-Induced Liver Injury: Immunogenetics and Experimental Models. Int. J. Mol. Sci. 2021, 22, 4557. [Google Scholar] [CrossRef]
- Puri, M. Automated Machine Learning Diagnostic Support System as a Computational Biomarker for Detecting Drug-Induced Liver Injury Patterns in Whole Slide Liver Pathology Images. Assay Drug Dev. Technol. 2020, 18, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.-C.; Chang, J.; Lee, H.S.; Kwon, H.J. Mitochondrial UQCRB as a New Molecular Prognostic Biomarker of Human Colorectal Cancer. Exp. Mol. Med. 2017, 49, e391. [Google Scholar] [CrossRef] [PubMed]
- Demšar, J.; Curk, T.; Erjavec, A.; Gorup, Č.; Hočevar, T.; Milutinovič, M.; Možina, M.; Polajnar, M.; Toplak, M.; Starič, A.; et al. Orange: Data Mining Toolbox in Python. J. Mach. Learn. Res. 2013, 14, 2349–2353. [Google Scholar]
- Godec, P.; Pančur, M.; Ilenič, N.; Čopar, A.; Stražar, M.; Erjavec, A.; Pretnar, A.; Demšar, J.; Starič, A.; Toplak, M.; et al. Democratized Image Analytics by Visual Programming through Integration of Deep Models and Small-Scale Machine Learning. Nat. Commun. 2019, 10, 4551. [Google Scholar] [CrossRef]
- Hao, Y.; Hao, S.; Andersen-Nissen, E.; Mauck, W.M.; Zheng, S.; Butler, A.; Lee, M.J.; Wilk, A.J.; Darby, C.; Zager, M.; et al. Integrated Analysis of Multimodal Single-Cell Data. Cell 2021, 184, 3573–3587.e29. [Google Scholar] [CrossRef]
- Aizarani, N.; Saviano, A.; Sagar; Mailly, L.; Durand, S.; Herman, J.S.; Pessaux, P.; Baumert, T.F.; Grün, D. A Human Liver Cell Atlas Reveals Heterogeneity and Epithelial Progenitors. Nature 2019, 572, 199–204. [Google Scholar] [CrossRef] [PubMed]
- Ramachandran, P.; Dobie, R.; Wilson-Kanamori, J.R.; Dora, E.F.; Henderson, B.E.P.; Luu, N.T.; Portman, J.R.; Matchett, K.P.; Brice, M.; Marwick, J.A.; et al. Resolving the Fibrotic Niche of Human Liver Cirrhosis at Single-Cell Level. Nature 2019, 575, 512–518. [Google Scholar] [CrossRef]
- Zhang, M.; Yang, H.; Wan, L.; Wang, Z.; Wang, H.; Ge, C.; Liu, Y.; Hao, Y.; Zhang, D.; Shi, G.; et al. Single-Cell Transcriptomic Architecture and Intercellular Crosstalk of Human Intrahepatic Cholangiocarcinoma. J. Hepatol. 2020, 73, 1118–1130. [Google Scholar] [CrossRef]
- MacParland, S.A.; Liu, J.C.; Ma, X.-Z.; Innes, B.T.; Bartczak, A.M.; Gage, B.K.; Manuel, J.; Khuu, N.; Echeverri, J.; Linares, I.; et al. Single Cell RNA Sequencing of Human Liver Reveals Distinct Intrahepatic Macrophage Populations. Nat. Commun. 2018, 9, 4383. [Google Scholar] [CrossRef]
- Olsavszky, V.; Sticht, C.; Schmid, C.D.; Winkler, M.; Wohlfeil, S.A.; Olsavszky, A.; Schledzewski, K.; Géraud, C.; Goerdt, S.; Leibing, T.; et al. Exploring the Transcriptomic Network of Multi-Ligand Scavenger Receptor Stabilin-1- and Stabilin-2-Deficient Liver Sinusoidal Endothelial Cells. Gene 2021, 768, 145284. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Experimental workflow, histopathology image classification & analysis; (A) H&E image classification for Cirrhosis; (B) Bridging Fibrosis; (C) Early Fibrosis & Steatosis; (D) Workflow of Classification experiment study design; (E) t-SNE plot for clustered images; (F) Confusion matrix of classifier performance.
Figure 1.
Experimental workflow, histopathology image classification & analysis; (A) H&E image classification for Cirrhosis; (B) Bridging Fibrosis; (C) Early Fibrosis & Steatosis; (D) Workflow of Classification experiment study design; (E) t-SNE plot for clustered images; (F) Confusion matrix of classifier performance.

Figure 2.
Volcano plots of Differential Gene Expression in; (A) NASH vs Control. (B) Zone 1 vs Zone 3.
Figure 2.
Volcano plots of Differential Gene Expression in; (A) NASH vs Control. (B) Zone 1 vs Zone 3.
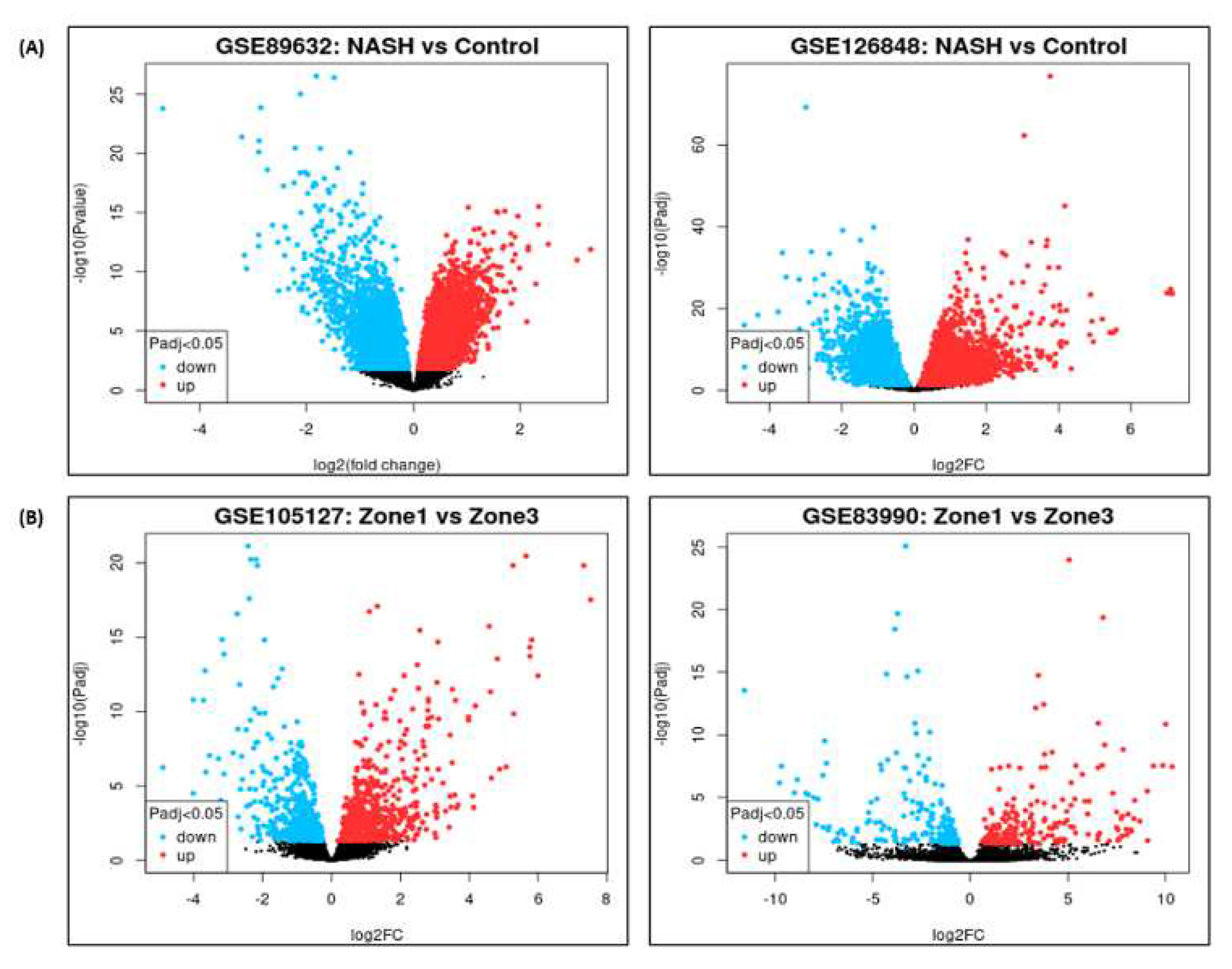
Figure 3.
Single Cell clustering in DILI and NASH ECs (study GSE166178) (A) cell clustering for control & DILI samples (combined); (B) cell annotations and clustering(separately); (C) cell clustering for control & NASH samples(combined); (D) cell annotations and clustering(separately). Abbreviations: Endo-Endothelial cells, DC- Dendritic cells, Macro-Macrophages, IM- Immune cells, MESN-Mesenchymal cells, NK-Tcell-Natural killer Tcells, Neutro-Neutrophils.
Figure 3.
Single Cell clustering in DILI and NASH ECs (study GSE166178) (A) cell clustering for control & DILI samples (combined); (B) cell annotations and clustering(separately); (C) cell clustering for control & NASH samples(combined); (D) cell annotations and clustering(separately). Abbreviations: Endo-Endothelial cells, DC- Dendritic cells, Macro-Macrophages, IM- Immune cells, MESN-Mesenchymal cells, NK-Tcell-Natural killer Tcells, Neutro-Neutrophils.
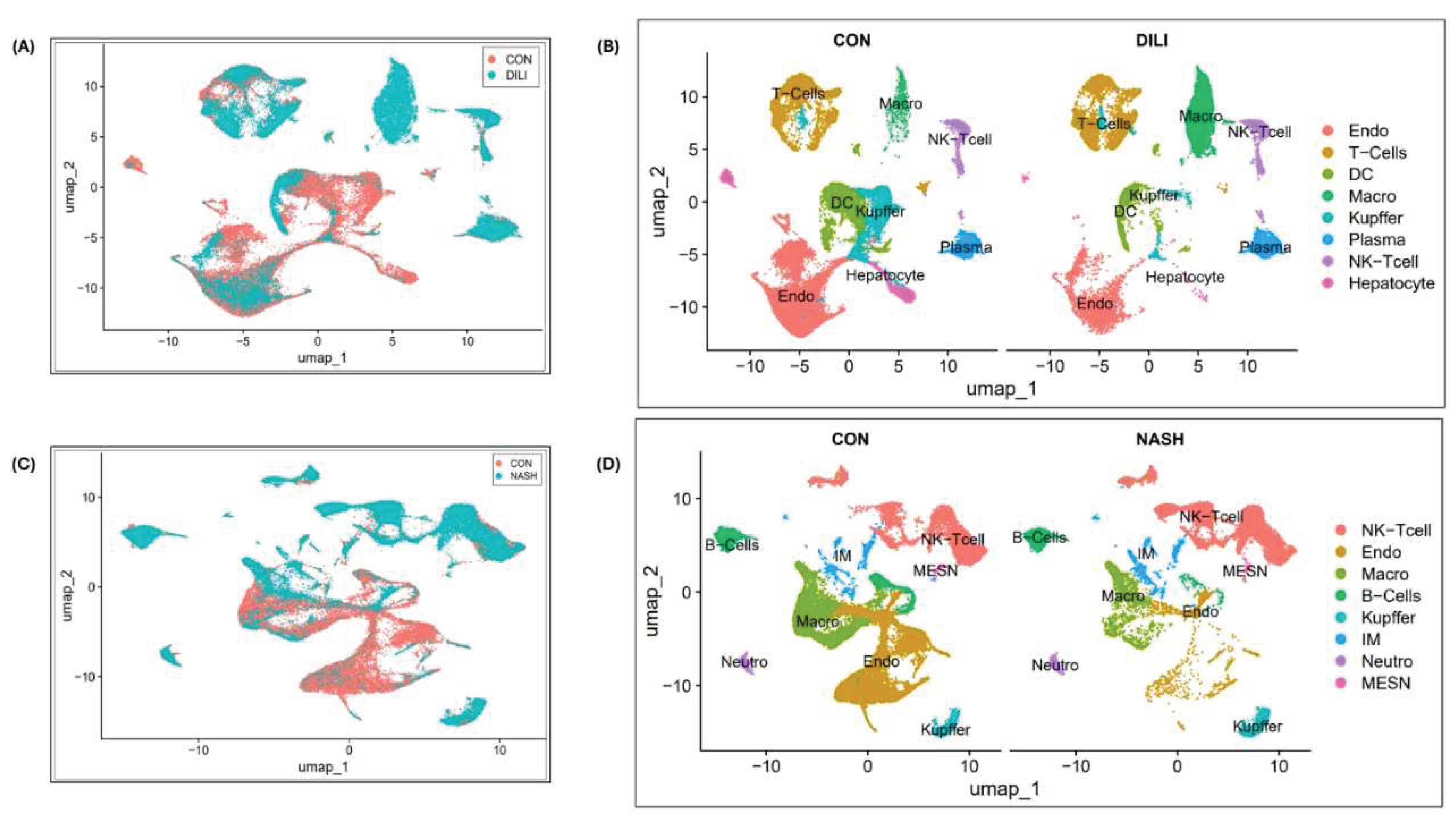
Figure 4.
x genomic`s Visium image analysis for zonation marker;(A) fiducial image of H&E-stained normal liver, (inset) highlighting zonation areas of zone1, zone2 & zone3, (B) gene expression for FGG in zone1, (C) expression for SAA1 in zone2, (D) expression for GLUL in zone3.
Figure 4.
x genomic`s Visium image analysis for zonation marker;(A) fiducial image of H&E-stained normal liver, (inset) highlighting zonation areas of zone1, zone2 & zone3, (B) gene expression for FGG in zone1, (C) expression for SAA1 in zone2, (D) expression for GLUL in zone3.
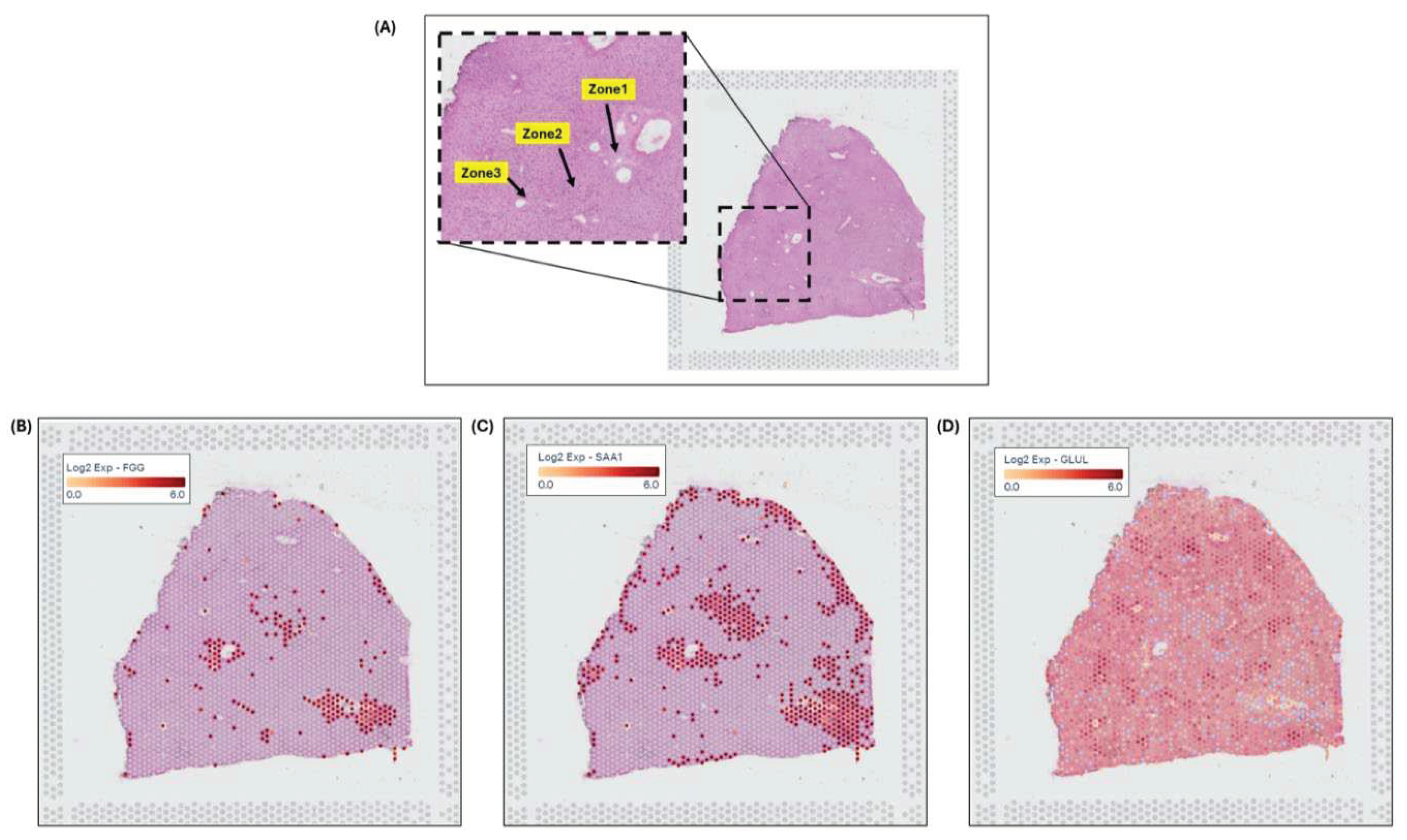
Figure 5.
Nanostring`s CosMx Spatial Molecular Imager (SMI) of normal human liver demonstrates zonation architecture: (A) Zonation boundaries for zone1,zone2,zone 3 and portal & central veins, (B) Zonation-specific gene expression, FGG (pink marker) expresses as in zone1(spreads across zone2), (C) SAA1 (aqua marker) expresses as in zone 2 ,(D) GLUL (white marker) expresses as in zone 3.
Figure 5.
Nanostring`s CosMx Spatial Molecular Imager (SMI) of normal human liver demonstrates zonation architecture: (A) Zonation boundaries for zone1,zone2,zone 3 and portal & central veins, (B) Zonation-specific gene expression, FGG (pink marker) expresses as in zone1(spreads across zone2), (C) SAA1 (aqua marker) expresses as in zone 2 ,(D) GLUL (white marker) expresses as in zone 3.
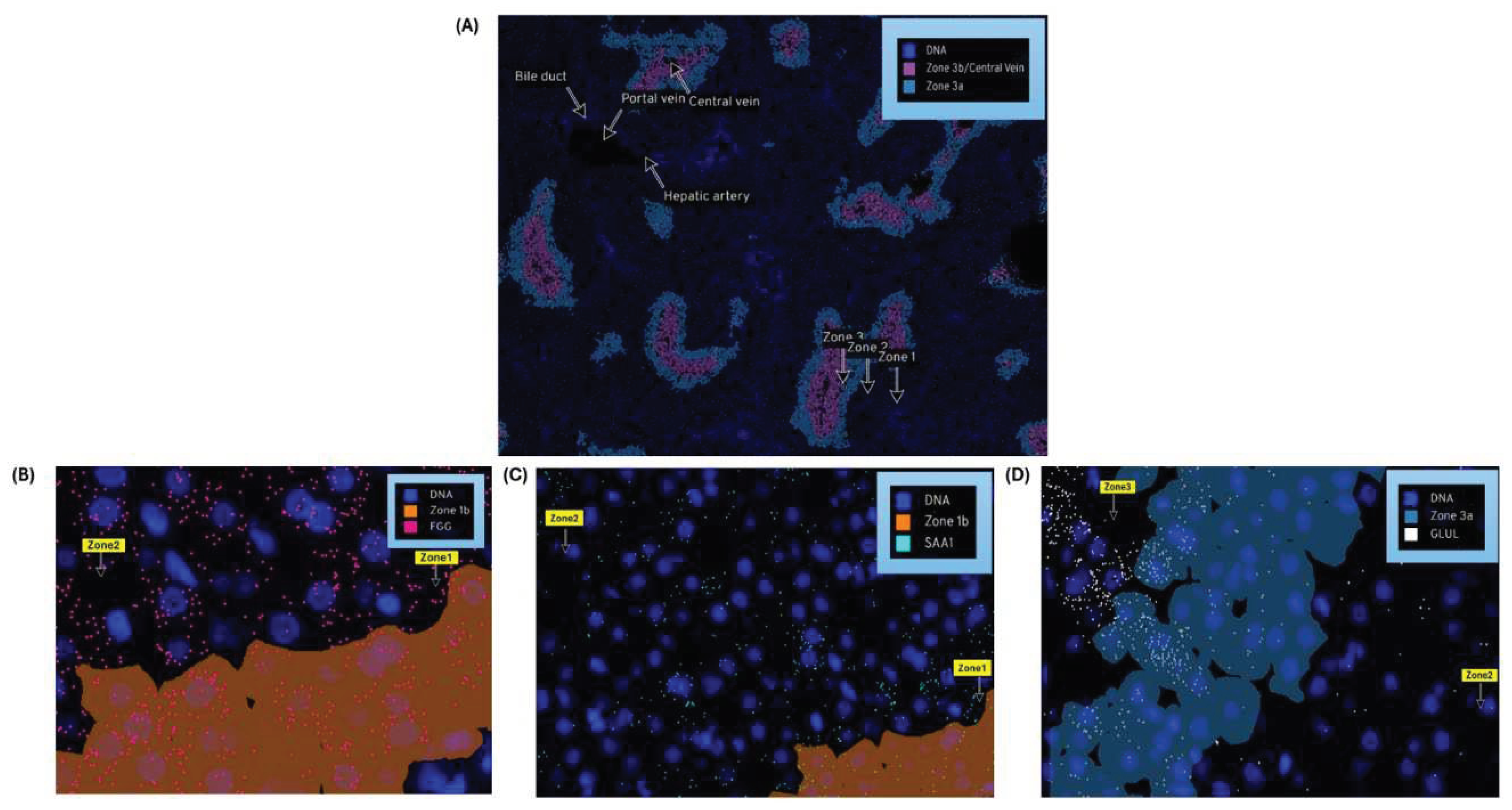
Figure 6.
LSECs cell clustering and markers (A) clustering of all the normal liver cells; (B)LSECs cluster; (C) VCVEC, VPEC, LSECs subset under ECs cluster (D)-(E) LSECs marker gene expressions; (F) VCVEC and VPEC marker gene expression clusters.
Figure 6.
LSECs cell clustering and markers (A) clustering of all the normal liver cells; (B)LSECs cluster; (C) VCVEC, VPEC, LSECs subset under ECs cluster (D)-(E) LSECs marker gene expressions; (F) VCVEC and VPEC marker gene expression clusters.
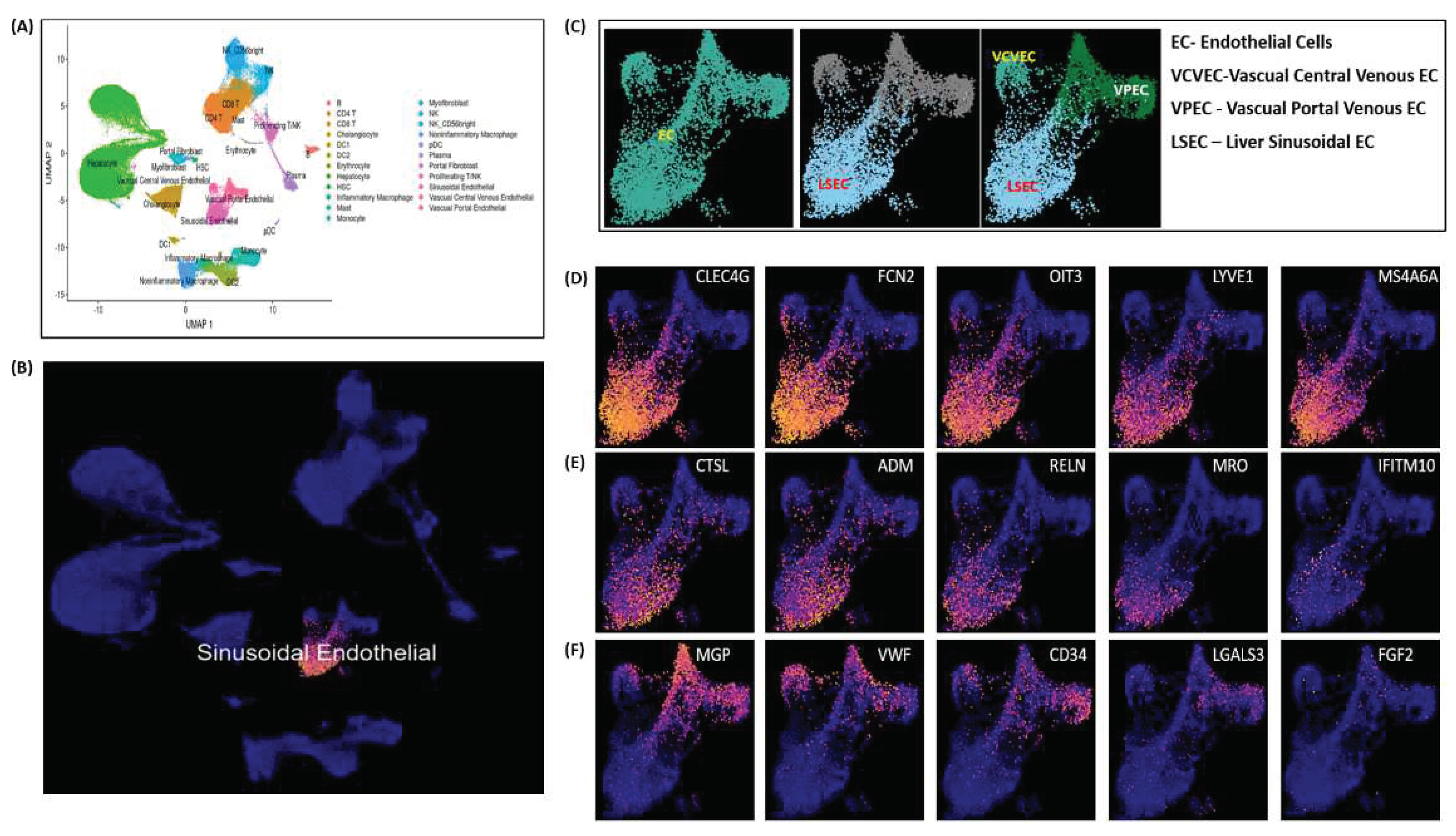
Figure 7.
Gene expression dot plots: Grid of genes by cell types enabling to access DGE expression profiles and percentage of expressing cells in PP–LSEC: Periportal Liver Sinusoid Endothelial Cells, PC–LSEC: Pericentral Liver Sinusoid Endothelial Cells and LSECs cells.
Figure 7.
Gene expression dot plots: Grid of genes by cell types enabling to access DGE expression profiles and percentage of expressing cells in PP–LSEC: Periportal Liver Sinusoid Endothelial Cells, PC–LSEC: Pericentral Liver Sinusoid Endothelial Cells and LSECs cells.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



