
Submitted:
16 February 2024
Posted:
18 February 2024
You are already at the latest version
Abstract
Plant-microbe interactions play a crucial role in shaping plant health and survival. In recent years, the role of extracellular vesicles (EVs) in mediating intercellular communication between plants and microbes has emerged as an intriguing area of research. EVs, serve as important carriers of bioactive molecules and genetic information, facilitating communication between cells and even between different organisms. Pathogenic bacteria leverage extracellular vesicles (EVs) to amplify their virulence, exploiting their cargo rich in toxins and virulence factors. Conversely, beneficial microbes initiate EV secretion to stimulate plant immune responses and nurture symbiotic relationships. The transfer of EV-packed small RNAs (sRNAs) has been demonstrated to facilitate the modulation of immune responses. Furthermore, harnessing the potential of EVs holds promise for the development of innovative diagnostic tools and sustainable crop protection strategies. This review highlights the biogenesis and functions of EVs in bacteria, and their importance in plant defense and paves the way for future research in this exciting field.
Keywords:
extracellular vesicles
; plant-microbe interactions
; plant defense mechanisms
; interkingdom communication
1. Introduction
In the natural environment, plants coexist with a diverse array of microorganisms, including bacteria, oomycetes, fungi, archaea, protists, and viruses, collectively forming the plant holobiont [1]. These organisms interact with the abiotic environment and each other, influencing the composition of the plant microbiota and its impact on the host [2,3]. Such interactions can range from mutualistic to commensal or even pathogenic, ultimately shaping the plant’s ability to cope with biotic and abiotic stresses [4]. While plants possess their own adaptive mechanisms, they also rely on their microbial partners to survive environmental challenges and defend against pathogens [5]. Root exudates play an important role in the interaction with soil microorganisms [6]. Notably, the communication between plants and microorganisms involves a complex web of interactions, with root exudates playing a crucial role in shaping the microbial community [7,8].
Cell-to-cell communication is fundamental across all living organisms, facilitating species interactions and the exchange of molecular signals. Extracellular vesicles (EVs) have emerged as key mediators of this communication, enabling cells to transport cargo over long distances and deliver it in higher concentrations [9]. EVs are evolutionary conserved structures found in both prokaryotes and eukaryotes. EVs are a heterogeneous population composed of nano to microscale size (20–1,000 nm) and derive from various sources [10]. EVs serve as vehicles for transporting proteins, lipids, secondary metabolites, and nucleic acids, including small RNAs (sRNAs). The communication mediated by EVs can occur through either uptake of EVs by recipient cells via membrane fusion or endocytosis, [11,12], or through the interaction of EV surface receptors with receptors on recipient cells [13].
While EVs have been extensively studied in human cells and human-pathogenic bacteria, their role in plant-microbe interactions remains largely unexplored. Understanding the significance of microbial EVs, represents a rapidly advancing field. In this review, we aim to shed light on the importance of EVs in host-microbe interactions and inter-species communication, with a specific focus on their role in plant-microbe interactions.
EVs in Plant-Microbe Interactions: Biogenesis and Functional Insights
Bacterial EVs
Bacterial EVs are produced by both Gram-negative and Gram-positive bacteria [14]. Their diameter ranges from 20 to 250 nm. A recent review has shed light on the dual origins of vesicle formation in both Gram-positive and Gram-negative bacteria, as this process can arise from the budding of outer membrane blebs or result from explosive cell lysis triggered by endolysins [15]. These vesicles are categorized into two types: outer membrane vesicles (OMVs), which encapsulate periplasmic components, and outer-inner membrane vesicles (OIMVs), as well as inner membrane vesicles (IMV) formed during explosive cell lysis and containing cytoplasmic contents, including DNA. To maintain simplicity and clarity, we will collectively refer to both OMVs, IMVs and OIMVs as "EVs." Bacterial EVs contain various cargo such as cell wall components, outer membrane proteins, lipopolysaccharides, phospholipids, proteins, nucleic acids and secondary metabolites [16]. The contents of EVs may be delivered into animal, plant and bacterial cells by membrane fusion and/or internalization, and its delivery can be targeted by molecules attached to the outside of vesicles. The production of EVs depends on many factors such as growth stage and stress, peaking at the end of the log phase and beginning of the stationary phase, and increasing in response to stress [17,18].
EVs of Gram-negative bacteria pathogenic to humans are well characterized, but little is known about microbes interacting with plants. Since the 1980s, EVs have been observed by electron microscopy in cultures of the pathogenic bacteria Erwinia amylovora and Erwinia carotovora [19]. Several studies have reported the production of EVs by plant pathogens in culture and during plant infection [20,21,22,23]. During infection, EVs have been detected in Gram-negative bacteria interacting with plants and within the plant itself [20], suggesting their involvement in cross-kingdom communication between bacteria and plant cells.
Biogenesis, Secretion and Uptake of EVs
The biogenesis of EVs in bacteria is still not fully understood, and different models have been proposed to explain their formation [24]. Gram-negative bacteria and Gram-positive bacteria have distinct mechanisms for vesiculation [25,26]. Some proposed models for EV biogenesis include [27]:
- Cell wall turnover: During routine cell wall recycling, lipoproteins between the outer membrane and the peptidoglycans dissociate, leading to membrane protrusion and the release of vesicles into the extracellular space [28].
- Conformational changes in outer membrane proteins (OMPs): Changes in the conformation of OMPs can promote vesicle formation. Specific proteins and lipids are locally enriched in areas with high vesicle abundance, while other proteins inhibiting vesiculation such as lipoproteins are reduced [29].
- Explosive cell lysis: A newly proposed mechanism suggests that vesiculation is a result of explosive cell lysis or bubbling cell death, [31], which involves the release of DNA-containing lytic EVs.
A recent review of Juodeikis and Simon in 2022 [32] proposes other EVs biogenesis pathways like the weakening of the outer membrane-peptidoglycan linkage or the flagellar rotation. It reminds the difficulty in distinguishing between the functions of lytic and nonlytic EVs.
The rate of vesicle production and the protein content of EVs can vary under different environmental conditions, indicating regulated biogenesis and cargo-sorting processes [17,33]. The determinants, machinery, and rules governing EV formation and the incorporation/exclusion of specific proteins into EVs are still areas of active investigation [34].
Functions of Bacterial EVs
A variety of EVs functions have been reported, related to interbacterial as well as host-bacteria interactions. A recent review by Toyofuku et al. [15] well describes the functions of bacterial EVs. In various aspects, the formation of bacterial EVs is in favor of the bacteria and their host partner. They are associated with several crucial functions such as (i) cell-cell communication, (ii) formation of biofilms and horizontal gene transfer to survive environmental stressors [35], (iii) transport and delivery (iv) stress response [17,18] (Figure 1).
Cell-Cell Communications and Quorum Sensing (QS)
EVs play a crucial role in cell-to-cell communication, particularly in the distribution of quorum sensing (QS) signals among bacterial populations [36]. QS signals play a pivotal role in the regulation of virulence factors in numerous pathogenic bacteria. They also have a significant impact on the modulation of beneficial traits in the microbiota associated with plants. QS molecules are often hydrophobic, like Pseudomonas Quinolone Signal (PQS) of Pseudomonas aeruginosa [36], C16-HSL of Paracoccus denitrificans and CAI-1 of Vibrio harveyi. By packaging hydrophobic QS molecules into vesicles, EVs solubilize and stabilize these signals, allowing their passage through the lipopolysaccharide layer of producing and receiving cell [37,38]. QS signals can also influence EVs production in other bacteria. PQS produced by P. aeruginosa can induce EVs formation in other species such as Escherichia coli, Burkholderia cepacia, and Bacillus subtilis [35,39]. In a recent study, Fan et al. [40] illustrated that the α-hydroxy-ketone compound Legionella autoinducer-1 (LAI-1) is secreted via EVs. This secretion mechanism facilitates interbacterial communication and interactions with eukaryotic host cells.
Biofilm Formation
Bacterial EVs play a significant role in biofilm formation, an essential survival strategy for bacteria. EVs contribute to biofilm formation and maintenance by mediating adhesive interactions, facilitating nutrient delivery to cells within the biofilm matrix, and enabling the long-range transport of molecules involved in virulence and antibiotic resistance [45,46,47].
Furthermore, EVs can act as offensive tools by lysing competing bacteria, thereby enhancing the competitive advantage of the producing bacteria [48]. In the context of the plant microbiota, EVs may influence the ability of competing microbes to adapt to the host environment by promoting cell lysis [27]. This inter-microbial competition within the root microbiota can involve EVs, which exhibit direct antifungal activities and contribute to the establishment of bacterial commensals in roots, protecting plants from harmful filamentous eukaryotes [49,50].
Additionally, EVs derived from Xylella fastidiosa ssp. pauca contain XfYgiT, a component of the toxin-antitoxin system known to regulate biofilm formation and contribute to the survival of X. fastidiosa ssp. Fastidiosa in planta [23].
These findings highlight the significant role of bacterial EVs in biofilm formation and their potential as mediators of inter-bacterial interactions and plant-microbe relationships within the complex microbial communities.
Transport and Delivery
For pathogenic bacteria, toxins and virulence factors are transferred to eukaryotic target cells by EVs [51]. Extracellular vesicles produced by Campylobacter jejuni, carrying cytolethal distending toxin, interact with host cell glycans, triggering cell cycle arrest in the host cells [52].
Proteomic analysis of EVs produced by various strains of Xanthomoas campestris and Pseudomonas syringae revealed the presence of virulence determinants as cellulase and xylosidase), type-II or III components (AvrA1 and HopI1) produced by P. syringae, which are known to suppress plant immune responses, and secreted proteins [20,53]. However, it remains unclear whether microbial EVs release their cargo into the extracellular space or have the ability to interact with plant cells to deliver their cargo into the cytosol.
Biller et al., [54] analyzed the cargo of Prochlorococcus, a marine cyanobacterium, using omics techniques. EVs produced by this marine bacterium contain lipids (polar lipids IPLs), and pigments (carotenoids and plastoquinone), proteins implicated in transport (porins and transporters), peptidase, hydrolase and chaperones, suggesting various functions for vesicles, such as dissipation of oxidative stress, transport of damaged or hydrophobic molecules. The presence of sugars like triose and tetrose suggests that the vesicles may be a carbon source for marine organisms [54]. These functions may enhance the bacterial fitness in the rhizosphere, a dynamic environment where plants and other microorganisms can produce antimicrobial compounds, and where potential exposure to toxic substances, such as pesticides, is not uncommon. Moreover, the nutritional content within the EVs could be a valuable resource for bacterial survival in the oligotrophic soil environment, especially in the absence of the host plant.
Moreover, EV-associated DNA has been implicated in facilitating horizontal gene transfer (HGT) among bacteria, which encompasses the dissemination of antibiotic resistance genes within and across bacterial species. The study by Johnston et al. [55] indicates that P. aeruginosa EVs have the capability to promote the spread of antibiotic resistance genes. This mechanism facilitates the survival of susceptible bacteria during antibiotic treatment.
Research conducted by Li et al. [56] has revealed that avian pathogenic Escherichia coli EVs facilitate the horizontal transfer of blaCTX-M-55. This discovery sheds light on how resistance is potentially proliferated within the poultry industry, underscoring the necessity for strict limitations on antibiotic use in poultry farming. The phenomenon of EVs-mediated gene transfer has also been observed in Klebsiella pneumoniae, where plasmids carrying resistance genes are transferred via EVs, as demonstrated by Dell’Annunziata et al. [57].
Stress Response
Bacterial EVs are a crucial strategy employed by bacteria to survive environmental stresses. They sequester and export misfolded toxic proteins generated during heat stress ([17], providing protection against complement components [58], long-chain alcohols, metal chelators [45] and antimicrobial peptides [46,59].
Membrane vesicles production is highly influenced by various stressors such as oxidative stress, UV-radiation, nutrient deprivation, pH, temperature, hydration, antibiotics [60]. These stressors can directly induce export of misfolded proteins or induce outer membrane blebbing following changes in outer membrane composition.
Lima et al. [61] present compelling evidence suggesting that bacteria from diverse ecological niches release abundant EVs in response to copper exposure. Their findings, particularly with the cyanobacterium Synechocystis, indicate that EV release in bacteria offers a novel mechanism for copper secretion, providing insights into alternative pathways for bacterial metal resistance.
EVs contribute to the management of iron scarcity, a crucial factor, particularly in the case of pathogens such as Mycobacterium tuberculosis. These EVs carry mycobactin, a hydrophobic siderophore, which helps sequester iron from the environment and enhances pathogen survival during infection [62]. In the extracellular milieu of P. aeruginosa, the formation of the PQS-Fe3+ complex occurs, which is efficiently transported by EVs. These EVs serve as carriers, facilitated by the T6SS effector protein TseF, allowing the PQS-Fe3+ complex to hitchhike onto them. Subsequently, the complex gains access to the bacterial periplasm through outer membrane receptors OprF and FptA, as detailed by Zhang et al., [63].
Furthermore, EVs produced by Corynebacterium glutamicum also function as extracellular iron carriers, enabling iron uptake through a mechanism that operates independently of membrane-associated proteins or siderophores, as demonstrated by Kawashima et al., [64]. These intricate processes hold significant implications in environments such as the rhizosphere, where iron availability is often constrained. The investigation into the acquisition of extracellular vesicle-associated iron by EV-producing bacteria, other microbial entities, or the plant host represents a promising avenue for further exploration.
Role of EVs and sRNA in Plant-Microbe Interaction
Plant-Microbe Symbiosis
Research has documented the presence of EVs in rhizobia, with their isolation achieved under living conditions [65]. Moreover, characterization efforts have utilized proteomics to compare the expression profiles of periplasmic space proteins with those found in EVs of Rhizobium etli [66], along with investigating the protein profiles of EVs from Rhizobium etli both before and after naringenin induction [67]. Role of EVs in symbiosis establishment, such as EVs produced by Sinorhizobium fredii HH103, play a role in the establishment of rhizobium-soybean symbiosis. Treatment of soybean with EVs induced the expression of nodulation genes, suppressed plant defense genes, and affected root development [68].
Overall, EVs contribute to interorganismic signal and nutrient exchange, enhancing symbiotic interactions between plants and microbes (Figure 2).
EV-Packed sRNA and Pathogenicity
EVs produced by pathogenic bacteria are enriched in toxins and virulence factors, which are internalized into host cells to promote pathogenicity [9,17,18,29,69]. Proteomic studies have revealed that EVs from plant pathogenic bacteria contain plant cell wall-degrading enzymes, protein secretion machinery components, effectors, nucleic acids inducing plant immune responses, and various virulence factors [20,53,69,70]. Some of these nucleic acids are small RNAs (sRNAs). Imploying a blend of infrared and circular dichroism spectroscopies, Turnbant et al. [71] showcased the translocation of Hfq from the inner membrane into the periplasm, followed by its exportation within EVs, potentially binding to sRNAs. sRNAs hold the capability to directly enter host cells, thereby silencing host defense genes and attenuating host immunity. Numerous studies have affirmed that the host immune responses, provoked by sRNAs encapsulated within EVs, facilitate bacterial growth and host infection in vivo [72,73,74,75]. Recent evidence from studies on Xanthomonas oryzae pv. oryzicola has demonstrated the presence of sRNAs packaged within EVs, specifically transferred into the mechanical tissues of rice leaves. Among these, the most abundant sRNA in EVs, Xosr001, plays a crucial role in regulating OsJMT1, which exploits the stomatal immune response of the host plant for virulence [76]. Xosr001 acts as an inhibitor to attenuate OsJMT1 transcripts in vivo, consequently reducing the endogenous MeJA content in rice leaves [76]. Furthermore, recent studies have shown that EV-packed sRNAs of P. aeruginosa can enter host cells and regulate immune responses [77].
Bacterial EVs: Activating Signal for Plant Immune Response
EVs produced by pathogenic bacteria can act as activating signals for plant immune responses [81,82]. Pathogenic bacteria often induce expression of the isochorismate synthase 1 (ICS1) gene in the host, which encodes an enzyme that catalyzes the production of salicylic acid (SA), a plant immune signal for systemic acquired resistance [83,84]. EVs from X. campestris pv. vesicatoria, X. campestris pv. campestris, and X. oryzae pv. oryzae and virulence factors purified from these EVs have been shown to trigger immune responses in plants, including callose deposition, alkalinization of the medium, increased transcription of pattern recognition receptors, activation of Mitogen-Activated Protein Kinase (MAPK) and release of reactive oxygen species [20,21,85,86]. They activate the salicylic acid (SA) pathway, leading to systemic acquired resistance and protection against infection by P. syringae ([87] and X. campestris pv. campestris [85].
Even, non-pathogenic bacteria such as Pseudomonas fluorescens can also activate plant immune responses at low levels [88]. The immune activation is triggered by Microbe-Associated Molecular Patterns (MAMPs) and induces a systemic immune response via SA-independent pathways [88]. Notably, pathogens such as P. syringae can overcome plant defenses using the type III secretion system (T3SS) [89], while other bacterial secretion pathways also play a role in plant-microbe interactions [90]. EVs of P. fluorescens can induce Induced Systemic Resistance (ISR) in A. thaliana and reduce the development of P. syringae [87].
Plants have developed two forms of innate immunity in response to infection: pathogen-associated molecular pattern (PAMP)-triggered immunity (PTI) and effector-triggered immunity (ETI), also known as R-gene-based immunity [91]. PTI is initiated by the recognition of conserved microbe-associated molecular patterns (MAMPs) by receptor-like kinases (RLKs) and receptor-like proteins (RLPs) on the cell surface, leading to immune responses [92]. The response includes the production of reactive oxygen species (ROS), activation of defense genes, increase in calcium concentration, callose deposition for cell wall thickening, MAPK activation, and expression of pathogenesis-related genes [93,94,95]. ETI, on the other hand, relies on the specific recognition of defense molecules produced by plant resistance genes or pathogen effectors. Pathogens deliver effector molecules to inhibit the PTI response, prompting plants to activate the intracellular receptor proteins containing a nucleotide binding domain (NBD) and a leucine-rich repeat (LRR) domain (NLR) for ETI [91]. During microbial infection, the host immune responses require a specific reprogramming of gene expression and communication between hosts and microbes.
Lipopolysaccharide (LPS) and elongation factor Tu (EF-Tu) are prevalent components of bacterial EVs, and they function as MAMPs activating pattern-triggered immunity (PTI) upon recognition by plant-encoded immune receptors [21,53]. EVs carrying these MAMPs, such as EVs from X. campestris pv. campestris, induce ROS production, ion release, and defense gene expression in A. thaliana through the EF-Tu receptor (EFR) [21,85]. EVs from other Gram-negative bacteria, including X. oryzae pv. oryzae, P. syringae pv. tomato DC3000, and Acidovorax citrulli M6, also induce defense gene expression [21]. Proteins involved in immune signaling and polarized immune response, such as BIR2, GRP7, RIN4, SOBIR1, PEN1, SYP122, and SYP132, have been identified as cargoes of A. thaliana EVs ([96], suggesting their involvement in plant immune responses and the regulation of EV release.
The ability of bacterial EVs to induce plant immune responses demonstrates their immunogenic potential, and their interaction with cell-surface-localized receptors EFR, BAK1, and SOBIR1, suggests direct interaction with plant cells [97].
Plant-Microbiota Vesicle Interactions
In recent years, groundbreaking research has established a connection between bacterial EVs and inflammatory disease processes, primarily through their involvement in mediating cell death [98]. Furthermore, Wei et al., [99], have demonstrated that EVs extracted from the microbiota, rather than a single strain, play a critical role in inducing significant pathological changes in AD (Alzheimer’s Disease) patients [99].
This emerging area of research holds tremendous potential for understanding their impact on plant growth and health, particularly in the rhizosphere. By delving deeper into the composition and effects of EVs in the context of the rhizosphere, we may uncover valuable insights into the intricate dynamics that govern microbiota assembly, microbe-microbe, and plant-microbe interactions (Figure 3).
2. Perspectives
By delving deeper into the cargo of EVs released by various microbial species engaged in plant-microbe interactions, such as beneficial microbes or pathogens, we can gain valuable insights into their strategies for manipulating host plants and influencing the plant immune system.
Either plant, fungi and bacteria are known to produce EVs. One intriguing area of research lies in the crosstalk between plant-derived EVs and microbial EVs. Exploring how these EVs communicate and exchange cargo with one another could shed light on the underlying mechanisms of symbiotic relationships, pathogen recognition, and modulation of plant immune responses.
By manipulating the cargo of EVs or engineering EVs to optimize plant-microbe interactions, by selectively loading specific molecules into EVs, we may unlock the potential for novel strategies in plant protection and improved crop production.
It is important to explore the contribution of microbial EVs in microbe-microbe interactions and the intricate dynamics of microbiota-plant interaction, under natural conditions.
Continued exploration and a deeper understanding of the intricate interplay between plants and their associated microbiota through EV-mediated communication will contribute significantly to advancements in agriculture, crop protection, and ecological sustainability.
Author Contributions
SP and WA conceptualized the manuscript. SP, AB drafted the manuscript. WA revised the manuscript and the figures. The authors have read and agreed to the final version of the review. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by a grant DST (DST/INSPIRE/04/2016/000768, INSPIRE Faculty Award), New Delhi, India, and by a Ph.D. program grant from CEA (France). This work is part of COMBAT project (FR. ECCOREV/ITEM).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Hassani, M.A.; Durán, P.; Hacquard, S. Microbial interactions within the plant holobiont. Microbiome 2018, 6, 1–17. [Google Scholar] [CrossRef]
- Benidire, L., El Khalloufi, F., Oufdou, K., Barakat, M., Tulumello, J., Ortet, P., Heulin, T., Achouak, W. Phytobeneficial bacteria improve saline stress tolerance in Viciafaba and modulate microbial interaction network. Sci. Total Environ. 2020, 729, 139020. [CrossRef]
- Tulumello, J.; Chabert, N.; Rodriguez, J.; Long, J.; Nalin, R.; Achouak, W.; Heulin, T. Rhizobium alamii improves water stress tolerance in a non-legume. Sci. Total. Environ. 2021, 797, 148895. [Google Scholar] [CrossRef]
- Thrall, P.H.; Hochberg, M.E.; Burdon, J.J.; Bever, J.D. Coevolution of symbiotic mutualists and parasites in a community context. Trends Ecol. Evol. 2007, 22, 120–126. [Google Scholar] [CrossRef]
- Turner, T. R., James, E. K., and Poole, P. S. The plant microbiome. Genome Biol 2013, 14:209. [CrossRef]
- Olanrewaju, O.S.; Ayangbenro, A.S.; Glick, B.R.; Babalola, O.O. Plant health: feedback effect of root exudates-rhizobiome interactions. Appl. Microbiol. Biotechnol. 2018, 103, 1155–1166. [Google Scholar] [CrossRef] [PubMed]
- Haichar, F.e.Z.; Santaella, C.; Heulin, T.; Achouak, W. Root exudates mediated interactions belowground. Soil Biol. Biochem. 2014, 77, 69–80. [Google Scholar] [CrossRef]
- Alahmad, A.; Harir, M.; Fochesato, S.; Tulumello, J.; Walker, A.; Barakat, M.; Ndour, P.M.S.; Schmitt-Kopplin, P.; Cournac, L.; Laplaze, L.; et al. Unraveling the interplay between root exudates, microbiota, and rhizosheath formation in pearl millet. Microbiome 2024, 12, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Kulp, A.; Kuehn, M.J. Biological Functions and Biogenesis of Secreted Bacterial Outer Membrane Vesicles. Annu. Rev. Microbiol. 2010, 64, 163–184. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.-W.; Um, J.-H.; Cho, J.-H.; Lee, H.-J. Tiny RNAs and their voyage via extracellular vesicles: Secretion of bacterial small RNA and eukaryotic microRNA. Exp. Biol. Med. 2017, 242, 1475–1481. [Google Scholar] [CrossRef] [PubMed]
- Abels, E.R.; Breakefield, X.O. Introduction to Extracellular Vesicles: Biogenesis, RNA Cargo Selection, Content, Release, and Uptake. Cell. Mol. Neurobiol. 2016, 36, 301–312. [Google Scholar] [CrossRef] [PubMed]
- O’Donoghue, E.J.; Krachler, A.M. Mechanisms of outer membrane vesicle entry into host cells. Cell. Microbiol. 2016, 18, 1508–1517. [Google Scholar] [CrossRef]
- Colombo, M.; Raposo, G.; Théry, C. Biogenesis, secretion, and intercellular interactions of exosomes and other extracellular vesicles. Annu. Rev. Cell Dev. Biol. 2014, 30, 255–289. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.-Y., Choi, D.-Y., Kim, D.-K., Kim, J.-W., Park, J. O., Kim, S., Kim, S.-H., Desiderio, D. M., Kim, Y.-K., Kim, K.-P., Gho, Y. S. Gram-positive bacteria produce membrane vesicles: Proteomics-based characterization of Staphylococcus aureus-derived membrane vesicles. Proteomics 2009, 9:5425–5436. [CrossRef]
- Toyofuku, M.; Schild, S.; Kaparakis-Liaskos, M.; Eberl, L. Composition and functions of bacterial membrane vesicles. Nat. Rev. Microbiol. 2023, 21, 415–430. [Google Scholar] [CrossRef]
- Schulz, E.; Goes, A.; Garcia, R.; Panter, F.; Koch, M.; Müller, R.; Fuhrmann, K.; Fuhrmann, G. Biocompatible bacteria-derived vesicles show inherent antimicrobial activity. J. Control. Release 2018, 290, 46–55. [Google Scholar] [CrossRef] [PubMed]
- McBroom, A. J., and Kuehn, M. J. Release of outer membrane vesicles by Gram-negative bacteria is a novel envelope stress response: Outer membrane vesicles relieve envelope stress. Molecular Microbiology 2007, 63:545–558. [CrossRef]
- Schwechheimer, C.; Kuehn, M.J. Outer-membrane vesicles from Gram-negative bacteria: biogenesis and functions. Nat. Rev. Microbiol. 2015, 13, 605–619. [Google Scholar] [CrossRef] [PubMed]
- Yaganza, E.-S., Rioux, D., Simard, M., Arul, J., and Tweddell, R. J. Ultrastructural Alterations of Erwinia carotovora subsp. atroseptica Caused by Treatment with Aluminum Chloride and Sodium Metabisulfite. Appl Environ Microbiol 2004, 70:6800–6808. [CrossRef]
- Solé, M.; Scheibner, F.; Hoffmeister, A.-K.; Hartmann, N.; Hause, G.; Rother, A.; Jordan, M.; Lautier, M.; Arlat, M.; Büttner, D. Xanthomonas campestris pv. vesicatoria Secretes Proteases and Xylanases via the Xps Type II Secretion System and Outer Membrane Vesicles. J. Bacteriol. 2015, 197, 2879–2893. [Google Scholar] [CrossRef]
- Bahar, O.; Mordukhovich, G.; Luu, D.D.; Schwessinger, B.; Daudi, A.; Jehle, A.K.; Felix, G.; Ronald, P.C. Bacterial Outer Membrane Vesicles Induce Plant Immune Responses. Mol. Plant-Microbe Interactions® 2016, 29, 374–384. [Google Scholar] [CrossRef] [PubMed]
- Ionescu, M.; Zaini, P.A.; Baccari, C.; Tran, S.; da Silva, A.M.; Lindow, S.E. Xylella fastidiosa outer membrane vesicles modulate plant colonization by blocking attachment to surfaces. Proc. Natl. Acad. Sci. 2014, 111, E3910–E3918. [Google Scholar] [CrossRef]
- Santiago, A.d.S.; Mendes, J.S.; dos Santos, C.A.; de Toledo, M.A.S.; Beloti, L.L.; Crucello, A.; Horta, M.A.C.; Favaro, M.T.d.P.; Munar, D.M.M.; de Souza, A.A.; et al. The Antitoxin Protein of a Toxin-Antitoxin System from Xylella fastidiosa Is Secreted via Outer Membrane Vesicles. Front. Microbiol. 2016, 7, 2030. [Google Scholar] [CrossRef]
- Roier, S., Leitner, D. R., Iwashkiw, J., Schild-Prüfert, K., Feldman, M. F., Krohne, G., Reidl, J., and Schild, S. Intranasal Immunization with Nontypeable Haemophilus influenzae Outer Membrane Vesicles Induces Cross-Protective Immunity in Mice. PLoS ONE 2012, 7:e42664. [CrossRef]
- Brown, L.; Wolf, J.M.; Prados-Rosales, R.; Casadevall, A. Through the wall: extracellular vesicles in Gram-positive bacteria, mycobacteria and fungi. Nat. Rev. Microbiol. 2015, 13, 620–630. [Google Scholar] [CrossRef]
- Resch, U.; Tsatsaronis, J.A.; Le Rhun, A.; Stübiger, G.; Rohde, M.; Kasvandik, S.; Holzmeister, S.; Tinnefeld, P.; Wai, S.N.; Charpentier, E. A Two-Component Regulatory System Impacts Extracellular Membrane-Derived Vesicle Production in Group A Streptococcus. mBio 2016, 7, e00207-16. [Google Scholar] [CrossRef]
- MacDonald, I.A.; Kuehn, M.J. Offense and defense: microbial membrane vesicles play both ways. Res. Microbiol. 2012, 163, 607–618. [Google Scholar] [CrossRef]
- Yeh, Y.-C.; Comolli, L.R.; Downing, K.H.; Shapiro, L.; McAdams, H.H. The Caulobacter Tol-Pal Complex Is Essential for Outer Membrane Integrity and the Positioning of a Polar Localization Factor. J. Bacteriol. 2010, 192, 4847–4858. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, H.M.; Jagannadham, M.V. Biogenesis and multifaceted roles of outer membrane vesicles from Gram-negative bacteria. Microbiology 2014, 160, 2109–2121. [Google Scholar] [CrossRef] [PubMed]
- Elhenawy, W.; Bording-Jorgensen, M.; Valguarnera, E.; Haurat, M.F.; Wine, E.; Feldman, M.F. LPS Remodeling Triggers Formation of Outer Membrane Vesicles in Salmonella. mBio 2016, 7, e00940–16. [Google Scholar] [CrossRef] [PubMed]
- Toyofuku, M. Bacterial communication through membrane vesicles. Biosci. Biotechnol. Biochem. 2019, 83, 1599–1605. [Google Scholar] [CrossRef] [PubMed]
- Juodeikis, R., Simon, R. C. Outer Membrane Vesicles: Biogenesis, Functions, and Issues. Microbiology and Molecular Biology reviews 2022, 86(4). [CrossRef]
- Altindis, E.; Fu, Y.; Mekalanos, J.J. Proteomic analysis of Vibrio cholerae outer membrane vesicles. Proc. Natl. Acad. Sci. 2014, 111, E1548-56. [Google Scholar] [CrossRef] [PubMed]
- Kanno, M.; Shiota, T.; Ueno, S.; Takahara, M.; Haneda, K.; Tahara, Y.O.; Shintani, M.; Nakao, R.; Miyata, M.; Kimbara, K.; et al. Identification of genes involved in enhanced membrane vesicle formation in Pseudomonas aeruginosa biofilms: surface sensing facilitates vesiculation. Front. Microbiol. 2023, 14, 1252155. [Google Scholar] [CrossRef] [PubMed]
- Mashburn, L.M.; Whiteley, M. Membrane vesicles traffic signals and facilitate group activities in a prokaryote. Nature 2005, 437, 422–425. [Google Scholar] [CrossRef]
- Lin, J.; Cheng, J.; Wang, Y.; Shen, X. The Pseudomonas Quinolone Signal (PQS): Not Just for Quorum Sensing Anymore. Front. Cell. Infect. Microbiol. 2018, 8, 230. [Google Scholar] [CrossRef]
- Toyofuku, M., Morinaga, K., Hashimoto, Y. Uhl, J., Shimamura, H., Inaba, H., Schmitt-Kopplin, P., Eberl, L., Nomura, N. Membrane vesicle-mediated bacterial communication. ISME J 2017, 11:1504-1509. [CrossRef]
- Brameyer, S.; Plener, L.; Müller, A.; Klingl, A.; Wanner, G.; Jung, K. Outer Membrane Vesicles Facilitate Trafficking of the Hydrophobic Signaling Molecule CAI-1 between Vibrio harveyi Cells. J. Bacteriol. 2018, 200, e00740–17. [Google Scholar] [CrossRef]
- Tashiro, Y.; Ichikawa, S.; Nakajima-Kambe, T.; Uchiyama, H.; Nomura, N. Pseudomonas Quinolone Signal Affects Membrane Vesicle Production in not only Gram-Negative but also Gram-Positive Bacteria. Microbes Environ. 2010, 25, 120–125. [Google Scholar] [CrossRef]
- Fan, M.; Kiefer, P.; Charki, P.; Hedberg, C.; Seibel, J.; Vorholt, J.A.; Hilbi, H. The Legionella autoinducer LAI-1 is delivered by outer membrane vesicles to promote interbacterial and interkingdom signaling. J. Biol. Chem. 2023, 299, 105376. [Google Scholar] [CrossRef]
- Ciofu, O., Beveridge, T. J., Kadurugamuwa, J., Walther-Rasmussen, J., and Høiby, N. Chromosomal β-lactamase is packaged into membrane vesicles and secreted from Pseudomonas aeruginosa. Journal of Antimicrobial Chemotherapy 2000, 45:9–13. [CrossRef]
- Zhao, M.; He, S.; Wen, R.; Li, C.; Chen, X.; Lin, X.; Wang, H.; Tang, Y. Membrane vesicles derived from Enterococcus faecalis promote the co-transfer of important antibiotic resistance genes located on both plasmids and chromosomes. J. Antimicrob. Chemother. 2023, 79, 320–326. [Google Scholar] [CrossRef] [PubMed]
- Schaar, V.; Uddbäck, I.; Nordström, T.; Riesbeck, K. Group A streptococci are protected from amoxicillin-mediated killing by vesicles containing -lactamase derived from Haemophilus influenzae. J. Antimicrob. Chemother. 2013, 69, 117–120. [Google Scholar] [CrossRef] [PubMed]
- Rumbo, C.; Fernández-Moreira, E.; Merino, M.; Poza, M.; Mendez, J.A.; Soares, N.C.; Mosquera, A.; Chaves, F.; Bou, G. Horizontal Transfer of the OXA-24 Carbapenemase Gene via Outer Membrane Vesicles: a New Mechanism of Dissemination of Carbapenem Resistance Genes in Acinetobacter baumannii. Antimicrob. Agents Chemother. 2011, 55, 3084–3090. [Google Scholar] [CrossRef]
- Baumgarten, T.; Sperling, S.; Seifert, J.; von Bergen, M.; Steiniger, F.; Wick, L.Y.; Heipieper, H.J. Membrane Vesicle Formation as a Multiple-Stress Response Mechanism Enhances Pseudomonas putida DOT-T1E Cell Surface Hydrophobicity and Biofilm Formation. Appl. Environ. Microbiol. 2012, 78, 6217–6224. [Google Scholar] [CrossRef]
- Manning, A.J.; Kuehn, M.J. Contribution of bacterial outer membrane vesicles to innate bacterial defense. BMC Microbiol. 2011, 11, 258–258. [Google Scholar] [CrossRef] [PubMed]
- Huang, B.; Liu, C.; Yang, J.; Xiang, E.; Ivanovski, S.; Han, P. Saliva biofilm-derived outer membrane vesicles regulate biofilm formation and immune response of oral epithelial cells on titanium surfaces. Clin. Oral Investig. 2024, 28, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Clarke, A.J.; Beveridge, T.J. Gram-Negative Bacteria Produce Membrane Vesicles Which Are Capable of Killing Other Bacteria. J. Bacteriol. 1998, 180, 5478–83. [Google Scholar] [CrossRef] [PubMed]
- Durán, P., Thiergart, T., Garrido-Oter, R., Agler, M., Kemen, E., Schulze-Lefert, P., and Hacquard, S. Microbial Interkingdom Interactions in Roots Promote Arabidopsis Survival. Cell 2018, 175:973-983.e14. [CrossRef]
- Rossoni, S.; Beard, S.; Segura-Bidermann, M.I.; Duarte-Ramírez, J.; Osorio, F.K.; Varas-Godoy, M.; Martínez-Bellange, P.; Vera, M.; Quatrini, R.; Castro, M. Membrane vesicles in Acidithiobacillia class extreme acidophiles: influence on collective behaviors of ‘Fervidacidithiobacillus caldus’. Front. Microbiol. 2024, 14, 1331363. [Google Scholar] [CrossRef]
- Rivera, J.; Cordero, R.J.B.; Nakouzi, A.S.; Frases, S.; Nicola, A.; Casadevall, A. Bacillus anthracis produces membrane-derived vesicles containing biologically active toxins. Proc. Natl. Acad. Sci. USA 2010, 107, 19002–19007. [Google Scholar] [CrossRef]
- Le, L.H.M.; Elgamoudi, B.; Colon, N.; Cramond, A.; Poly, F.; Ying, L.; Korolik, V.; Ferrero, R.L. Campylobacter jejuni extracellular vesicles harboring cytolethal distending toxin bind host cell glycans and induce cell cycle arrest in host cells. Microbiol. Spectr. 2024, 12, e0323223. [Google Scholar] [CrossRef]
- Chowdhury, C.; Jagannadham, M.V. Virulence factors are released in association with outer membrane vesicles of Pseudomonas syringae pv. tomato T1 during normal growth. Biochim. et Biophys. Acta (BBA) - Proteins Proteom. 2013, 1834, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Biller, S.J.; Lundeen, R.A.; Hmelo, L.R.; Becker, K.W.; Arellano, A.A.; Dooley, K.; Heal, K.R.; Carlson, L.T.; Van Mooy, B.A.S.; Ingalls, A.E.; et al. Prochlorococcus extracellular vesicles: molecular composition and adsorption to diverse microbes. Environ. Microbiol. 2021, 24, 420–435. [Google Scholar] [CrossRef] [PubMed]
- Johnston, E.L.; Zavan, L.; Bitto, N.J.; Petrovski, S.; Hill, A.F.; Kaparakis-Liaskos, M. Planktonic and Biofilm-Derived Pseudomonas aeruginosa Outer Membrane Vesicles Facilitate Horizontal Gene Transfer of Plasmid DNA. Microbiol. Spectr. 2023, 11, e0517922. [Google Scholar] [CrossRef]
- Li, C.; Wen, R.; Mu, R.; Chen, X.; Ma, P.; Gu, K.; Huang, Z.; Ju, Z.; Lei, C.; Tang, Y.; et al. Outer Membrane Vesicles of Avian PathogenicEscherichia coli Mediate the Horizontal Transmission of blaCTX-M-55. Pathogens 2022, 11, 481. [Google Scholar] [CrossRef] [PubMed]
- Dell’annunziata, F.; Dell’aversana, C.; Doti, N.; Donadio, G.; Piaz, F.D.; Izzo, V.; De Filippis, A.; Galdiero, M.; Altucci, L.; Boccia, G.; et al. Outer Membrane Vesicles Derived from Klebsiella pneumoniae Are a Driving Force for Horizontal Gene Transfer. Int. J. Mol. Sci. 2021, 22, 8732. [Google Scholar] [CrossRef] [PubMed]
- Grenier, D.; Bélanger, M. Protective effect of Porphyromonas gingivalis outer membrane vesicles against bactericidal activity of human serum. Infect. Immun. 1991, 59, 3004–3008. [Google Scholar] [CrossRef]
- Meyer, K.J.; Nodwell, J.R. Streptomyces extracellular vesicles are a broad and permissive antimicrobial packaging and delivery system. J. Bacteriol. 2024, e0032523. [Google Scholar] [CrossRef]
- Mozaheb, N.; Mingeot-Leclercq, M.-P. Membrane Vesicle Production as a Bacterial Defense Against Stress. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef]
- Lima, S.; Matinha-Cardoso, J.; Giner-Lamia, J.; Couto, N.; Pacheco, C.C.; Florencio, F.J.; Wright, P.C.; Tamagnini, P.; Oliveira, P. Extracellular vesicles as an alternative copper-secretion mechanism in bacteria. J. Hazard. Mater. 2022, 431, 128594. [Google Scholar] [CrossRef]
- Prados-Rosales, R.; Weinrick, B.C.; Piqué, D.G.; Jr., W.R.J.; Casadevall, A.; Rodriguez, G.M. Role for Mycobacterium tuberculosis Membrane Vesicles in Iron Acquisition. J. Bacteriol. 2014, 196, 1250–1256. [CrossRef]
- Zhang, H, Yang, J., Cheng, J., Zeng, J., Ma, X., Lin, J. PQS and pyochelin in Pseudomonas aeruginosa share inner membrane transporters to mediate iron uptake. Microbiol Spectr. 2024, 12:e0325623. [CrossRef]
- Kawashima, K.; Nagakubo, T.; Nomura, N.; Toyofuku, M. Iron Delivery through Membrane Vesicles in Corynebacterium glutamicum. Microbiol. Spectr. 2023, 11, e0122223. [Google Scholar] [CrossRef]
- Ayala-García, P., Moreno-de Castro, N., Jiménez-Guerrero, I., Müsken, M., Arce-Rodríguez, A., Pérez-Montaño, F., Borrero-de Acuña, J. M. Isolation, Quantification, and Visualization of Extracellular Membrane Vesicles in Rhizobia Under Free-Living Conditions. Methods Mol Biol. 2024, 2751:219-228. [CrossRef]
- Taboada, H.; Meneses, N.; Dunn, M.F.; Vargas-Lagunas, C.; Buchs, N.; Castro-Mondragón, J.A.; Heller, M.; Encarnación, S. Proteins in the periplasmic space and outer membrane vesicles of Rhizobium etli CE3 grown in minimal medium are largely distinct and change with growth phase. Microbiology 2019, 165, 638–650. [Google Scholar] [CrossRef] [PubMed]
- Taboada, H.; Dunn, M.F.; Meneses, N.; Vargas-Lagunas, C.; Buchs, N.; Andrade-Domínguez, A.; Encarnación, S. Qualitative changes in proteins contained in outer membrane vesicles produced by Rhizobium etli grown in the presence of the nod gene inducer naringenin. Arch. Microbiol. 2019, 201, 1173–1194. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Li, Z.; Wu, J.; Tang, Z.; Xie, F.; Chen, D.; Lin, H.; Li, Y. Analysis of Outer Membrane Vesicles Indicates That Glycerophospholipid Metabolism Contributes to Early Symbiosis Between Sinorhizobium fredii HH103 and Soybean. Mol. Plant-Microbe Interactions® 2022, 35, 311–322. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, H.; Swamy, C.V.; Jagannadham, M. The proteome of the outer membrane vesicles of an Antarctic bacterium Pseudomonas syringae Lz4W. Data Brief 2015, 4, 406–409. [Google Scholar] [CrossRef] [PubMed]
- McMillan, H.M.; Kuehn, M.J. Proteomic Profiling Reveals Distinct Bacterial Extracellular Vesicle Subpopulations with Possibly Unique Functionality. Appl. Environ. Microbiol. 2023, 89, e0168622. [Google Scholar] [CrossRef]
- Turbant, F.; Waeytens, J.; Blache, A.; Esnouf, E.; Raussens, V.; Węgrzyn, G.; Achouak, W.; Wien, F.; Arluison, V. Interactions and Insertion of Escherichia coli Hfq into Outer Membrane Vesicles as Revealed by Infrared and Orientated Circular Dichroism Spectroscopies. Int. J. Mol. Sci. 2023, 24, 11424. [Google Scholar] [CrossRef] [PubMed]
- Koeppen., Hampton, T. H., Jarek, M., Scharfe, M., Gerber, S.A., Mielcarz, D. W., Demers, E. G., Dolben, E. L., Hammond, J. H., Hogan, D. A., Stanton, B. A. A Novel Mechanism of Host-Pathogen Interaction through sRNA in Bacterial Outer Membrane Vesicles. PLoS Pathog. 2016, 112:e1005672. [CrossRef]
- Lee, H.-J. Microbe-Host Communication by Small RNAs in Extracellular Vesicles: Vehicles for Transkingdom RNA Transportation. Int. J. Mol. Sci. 2019, 20, 1487. [Google Scholar] [CrossRef]
- Moriano-Gutierrez, S.; Bongrand, C.; Essock-Burns, T.; Wu, L.; McFall-Ngai, M.J.; Ruby, E.G. The noncoding small RNA SsrA is released by Vibrio fischeri and modulates critical host responses. PLOS Biol. 2020, 18, e3000934. [Google Scholar] [CrossRef]
- Liu, D., Liu, S., Liu, J., Miao, L., Zhang, S., Pan, Y. sRNA23392 packaged by Porphyromonas gingivalis outer membrane vesicles promotes oral squamous cell carcinomas migration and invasion by targeting desmocollin-2. Mol Oral Microbiol. 2021, 36:182-191. [CrossRef]
- Wu, Y.; Wang, S.; Wang, P.; Nie, W.; Ahmad, I.; Sarris, P.F.; Chen, G.; Zhu, B. Suppression of host plant defense by bacterial small RNAs packaged in outer membrane vesicles. Plant Commun. 2024, 100817. [Google Scholar] [CrossRef]
- Xie, Z.; Wang, X.; Huang, Y.; Chen, S.; Liu, M.; Zhang, F.; Li, M.; Gu, Y.; Shen, X.; Wang, Y.; et al. Pseudomonas aeruginosa outer membrane vesicle-packed sRNAs can enter host cells and regulate innate immune responses. Microb. Pathog. 2024, 106562. [Google Scholar] [CrossRef]
- He, M.; Yin, S.; Huang, X.; Li, Y.; Li, B.; Gong, T.; Liu, Q. Insights into the regulatory role of bacterial sncRNA and its extracellular delivery via OMVs. Appl. Microbiol. Biotechnol. 2023, 108, 1–13. [Google Scholar] [CrossRef]
- Chowdhury, R., Massé, M. E. New Perspectives on Crosstalks Between Bacterial Regulatory RNAs from Outer Membrane Vesicles and Eukaryotic Cells. Methods Mol Biol. 2024, 2741:183-194. [CrossRef]
- Blache. A, Achouak., W. Extraction and Purification of Outer Membrane Vesicles and Their Associated RNAs. Methods Mol Biol. 2024, 2741:11-24. [CrossRef]
- Acevedo, R.; Fernã¡Ndez, S.; Zayas, C.; Acosta, A.; Sarmiento, M.E.; Ferro, V.A.; Rosenqvist, E.; Campa, C.; Cardoso, D.; Garcia, L.; et al. Bacterial Outer Membrane Vesicles and Vaccine Applications. Front. Immunol. 2014, 5, 121. [Google Scholar] [CrossRef]
- Caruana, J.C.; Walper, S.A. Bacterial Membrane Vesicles as Mediators of Microbe – Microbe and Microbe – Host Community Interactions. Front. Microbiol. 2020, 11, 432. [Google Scholar] [CrossRef]
- Wildermuth, M.C.; Dewdney, J.; Wu, G.; Ausubel, F.M. Isochorismate synthase is required to synthesize salicylic acid for plant defence. Nature 2001, 414, 562–565. [Google Scholar] [CrossRef]
- Glazebrook, J. Contrasting Mechanisms of Defense Against Biotrophic and Necrotrophic Pathogens. Annu. Rev. Phytopathol. 2005, 43, 205–227. [Google Scholar] [CrossRef]
- Chalupowicz, L.; Mordukhovich, G.; Assoline, N.; Katsir, L.; Sela, N.; Bahar, O. Bacterial outer membrane vesicles induce a transcriptional shift in arabidopsis towards immune system activation leading to suppression of pathogen growth in planta. J. Extracell. Vesicles 2023, 12, e12285. [Google Scholar] [CrossRef]
- Tran, T.M.; Chng, C.-P.; Pu, X.; Ma, Z.; Han, X.; Liu, X.; Yang, L.; Huang, C.; Miao, Y. Potentiation of plant defense by bacterial outer membrane vesicles is mediated by membrane nanodomains. Plant Cell 2021, 34, 395–417. [Google Scholar] [CrossRef] [PubMed]
- McMillan, H.M.; Zebell, S.G.; Ristaino, J.B.; Dong, X.; Kuehn, M.J. Protective plant immune responses are elicited by bacterial outer membrane vesicles. Cell Rep. 2021, 34, 108645–108645. [Google Scholar] [CrossRef] [PubMed]
- Pieterse, C.M.J.; Zamioudis, C.; Berendsen, R.L.; Weller, D.M.; Van Wees, S.C.M.; Bakker, P.A.H.M. Induced Systemic Resistance by Beneficial Microbes. Annu. Rev. Phytopathol. 2014, 52, 347–375. [Google Scholar] [CrossRef]
- Feng, F.; Zhou, J.-M. Plant–bacterial pathogen interactions mediated by type III effectors. Curr. Opin. Plant Biol. 2012, 15, 469–476. [Google Scholar] [CrossRef]
- Lomovatskaya, L.A.; Romanenko, A.S. Secretion Systems of Bacterial Phytopathogens and Mutualists (Review). Appl. Biochem. Microbiol. 2020, 56, 115–129. [Google Scholar] [CrossRef]
- Choi, H.W.; Klessig, D.F. DAMPs, MAMPs, and NAMPs in plant innate immunity. BMC Plant Biol. 2016, 16, 1–10. [Google Scholar] [CrossRef]
- Ron, M., and Adi A. The receptor for the fungal elicitor ethylene-inducing xylanase is a member of a resistance-like gene family in tomato. The Plant Cell 2004, 16:1604-1615.
- Monaghan, J.; Zipfel, C. Plant pattern recognition receptor complexes at the plasma membrane. Curr. Opin. Plant Biol. 2012, 15, 349–357. [Google Scholar] [CrossRef]
- Tang, D.; Wang, G.; Zhou, J.-M. Receptor Kinases in Plant-Pathogen Interactions: More Than Pattern Recognition. Plant Cell 2017, 29, 618–637. [Google Scholar] [CrossRef]
- Wang, W.; Feng, B.; Zhou, J.-M.; Tang, D. Plant immune signaling: Advancing on two frontiers. J. Integr. Plant Biol. 2020, 62, 2–24. [Google Scholar] [CrossRef] [PubMed]
- Rutter, B.D.; Innes, R.W. Extracellular Vesicles Isolated from the Leaf Apoplast Carry Stress-Response Proteins. Plant Physiol. 2016, 173, 728–741. [Google Scholar] [CrossRef] [PubMed]
- Boutrot, F.; Zipfel, C. Function, Discovery, and Exploitation of Plant Pattern Recognition Receptors for Broad-Spectrum Disease Resistance. Annu. Rev. Phytopathol. 2017, 55, 257–286. [Google Scholar] [CrossRef]
- Dhital, S.; Deo, P.; Stuart, I.; Naderer, T. Bacterial outer membrane vesicles and host cell death signaling. Trends Microbiol. 2021, 29, 1106–1116. [Google Scholar] [CrossRef]
- Wei, S.; Peng, W.; Mai, Y.; Li, K.; Wei, W.; Hu, L.; Zhu, S.; Zhou, H.; Jie, W.; Wei, Z.; et al. Outer membrane vesicles enhance tau phosphorylation and contribute to cognitive impairment. J. Cell. Physiol. 2019, 235, 4843–4855. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Bacterial Vesicles (EVs) Production and Mediated Functions. A. Production of EVs: EVs are produced through outer membrane blebbing, wherein portions of the outer membrane (OMV), outer-inner membrane (OIMV) bud off, or inner membrane (IMV) resulting in the formation of vesicles containing a cargo of proteins, RNA, and metabolites, EPS stand for exopolysaccharides or even DNA for OIMVs and IMVs. B. EVs-Mediated Functions: EVs play diverse roles facilitated by their unique cargo, including competition, stress response, biofilm formation, host interaction, cell-cell communication and nutrient recovery.
Figure 1.
Bacterial Vesicles (EVs) Production and Mediated Functions. A. Production of EVs: EVs are produced through outer membrane blebbing, wherein portions of the outer membrane (OMV), outer-inner membrane (OIMV) bud off, or inner membrane (IMV) resulting in the formation of vesicles containing a cargo of proteins, RNA, and metabolites, EPS stand for exopolysaccharides or even DNA for OIMVs and IMVs. B. EVs-Mediated Functions: EVs play diverse roles facilitated by their unique cargo, including competition, stress response, biofilm formation, host interaction, cell-cell communication and nutrient recovery.
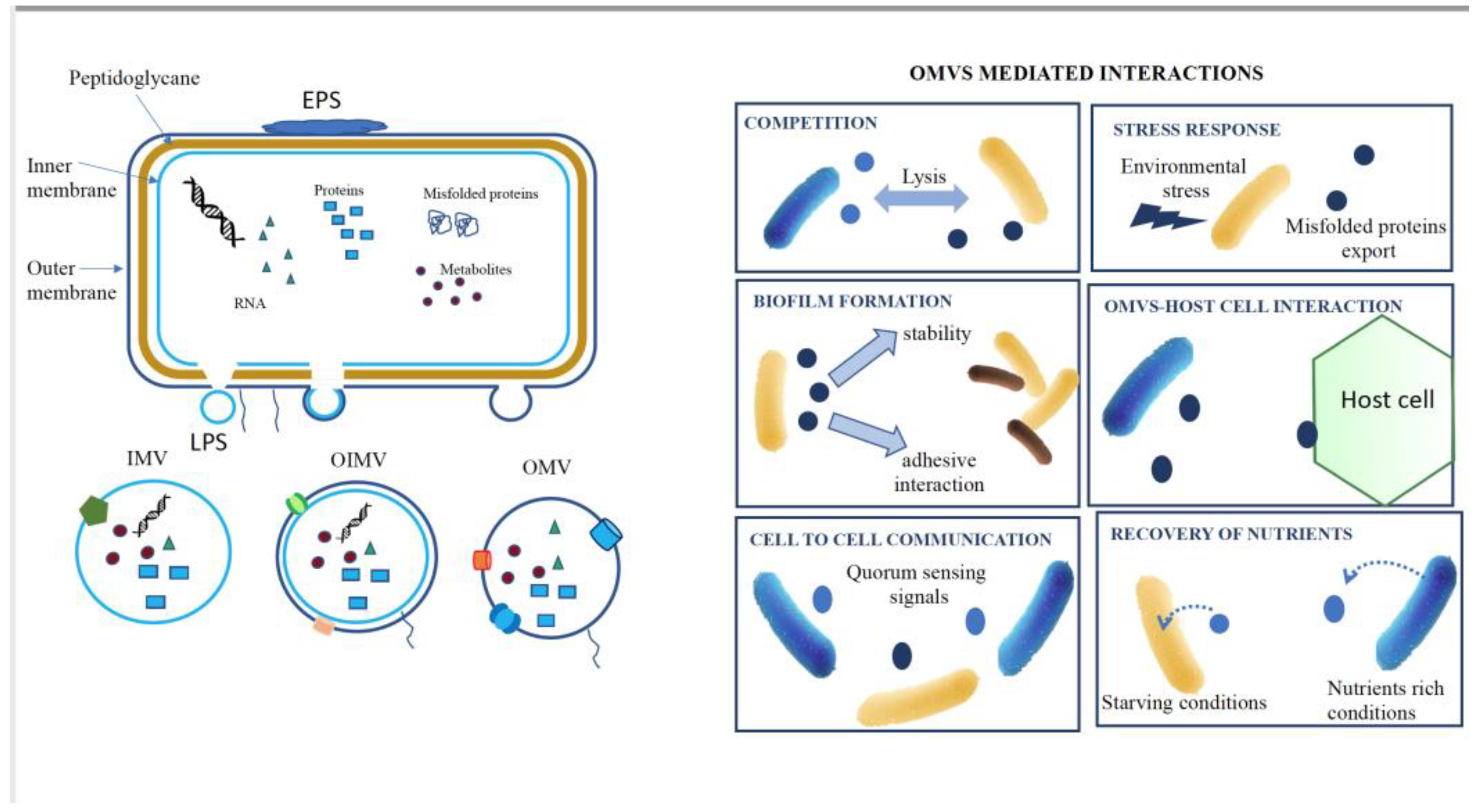
Figure 2.
Extracellular Vesicle-Mediated Interkingdom Communication in the Rhizosphere. This figure illustrates the activation of systemic acquired resistance in plants by pathogenic bacteria, as well as the induction of induced systemic resistance (ISR) in plants by beneficial bacterial EVs.
Figure 2.
Extracellular Vesicle-Mediated Interkingdom Communication in the Rhizosphere. This figure illustrates the activation of systemic acquired resistance in plants by pathogenic bacteria, as well as the induction of induced systemic resistance (ISR) in plants by beneficial bacterial EVs.
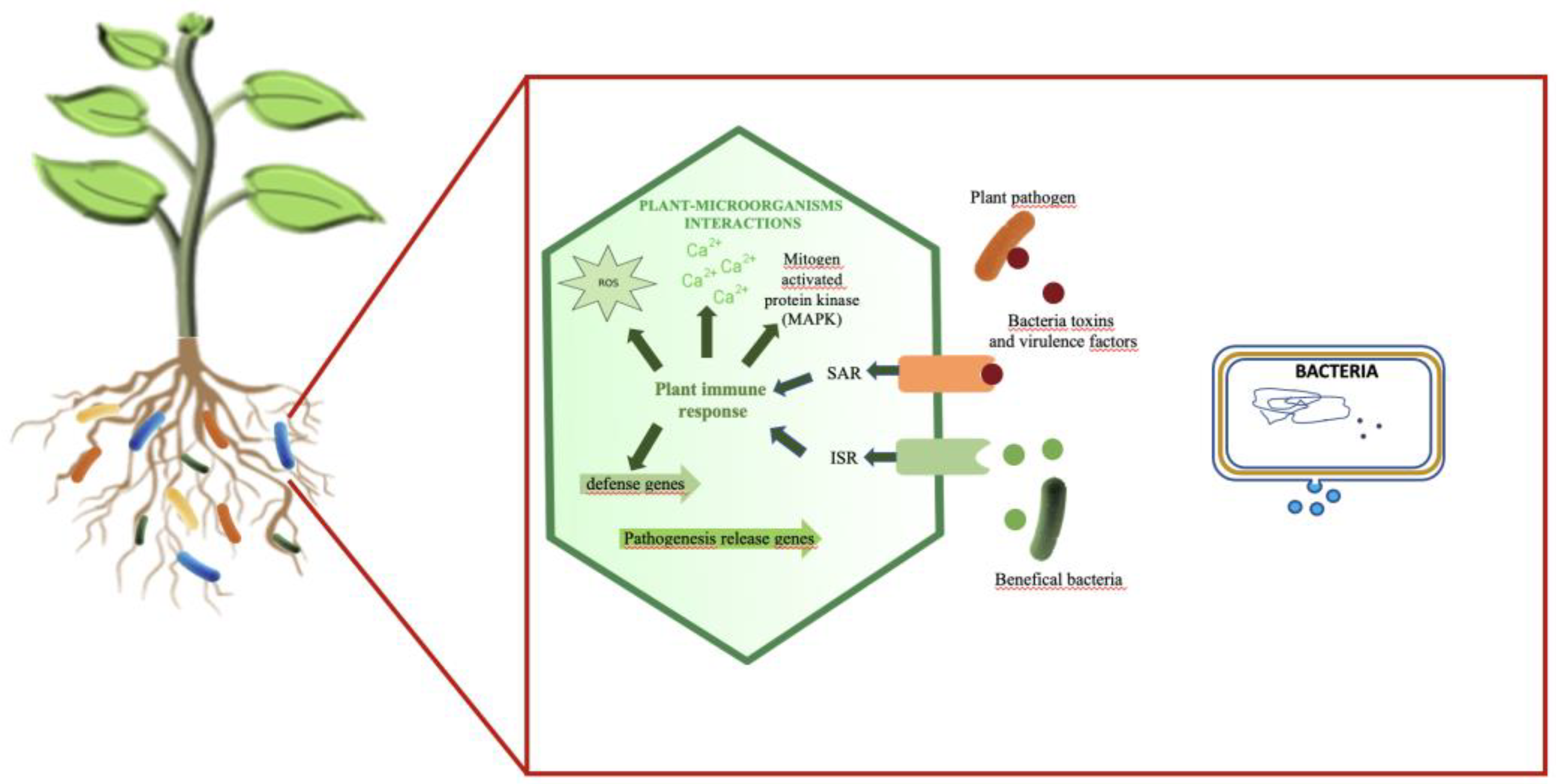
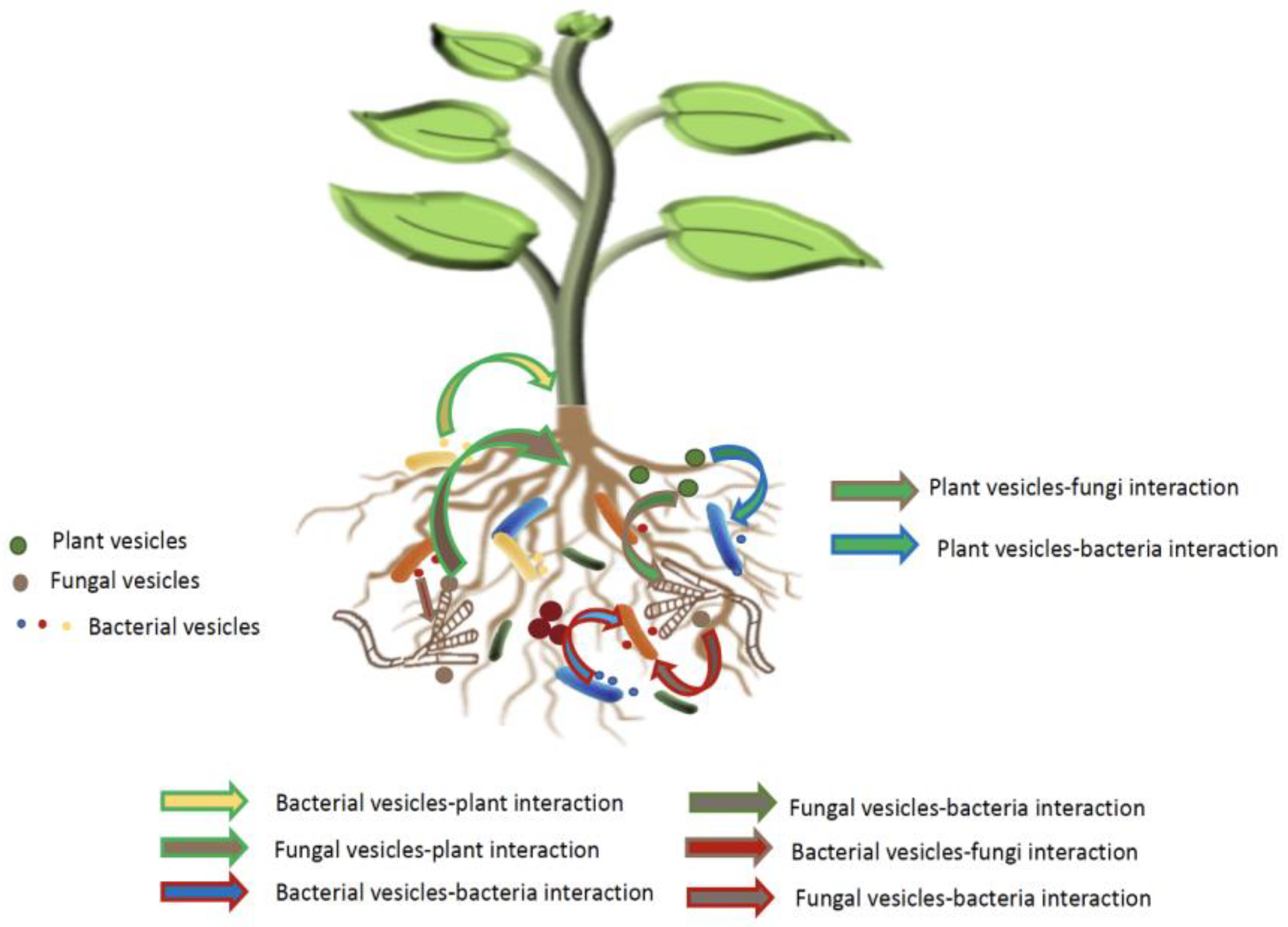
Figure 3.
EVs-Mediated Interaction Network in the Rhizosphere. This figure depicts the intricate interaction network involving EVs in the rhizosphere: Plant EVs and Recruitment of Root-Associated Microbiota; Microbe EVs and Microbiota Assembly; Synergic and Antagonistic Interactions.
Figure 3.
EVs-Mediated Interaction Network in the Rhizosphere. This figure depicts the intricate interaction network involving EVs in the rhizosphere: Plant EVs and Recruitment of Root-Associated Microbiota; Microbe EVs and Microbiota Assembly; Synergic and Antagonistic Interactions.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



