
Submitted:
18 February 2024
Posted:
19 February 2024
You are already at the latest version
Abstract
Peptides, both natural and synthetic, are well suited for a wide range of purposes and offer versa-tile applications in different fields such as biocatalysts, injectable hydrogels, tumor treatment, and drug delivery. The rising prominence of therapeutic peptides encompasses anticancer, antiviral, antimicrobial, and anti-neurodegenerative properties. The metals Na, K, Mg, Ca, Fe, Mn, Co, Cu, Zn, and Mo are ten of the twenty elements that are considered essential for life. Crucial for under-standing the biological role of metals is the exploration of metal-bound proteins and peptides. Aside from essential metals, there are other, non-essential metals that also interact biologically, exhibiting either therapeutic or toxic effects. Irregularities in metal binding contribute to diseases like Alzheimer’s, neurodegenerative disorders, Wilson's, and Menkes disease. Certain metal com-plexes have potential applications as radiopharmaceuticals. The examination of these complexes was done by preforming UV-Vis, IR, EPR, NMR spectroscopy, and X-ray analysis.
Keywords:
peptides
; metal complexes
; biological activities
Introduction
Nature has fine-tuned protein sequences, structures, and binding sites over thousands of years. Amino acid building blocks exhibit both chemical and structural versatility, efficiently and specifically binding to other biological macromolecules. Numerous peptides are either naturally occurring or derives from parent proteins through cleavage. As a result, peptides - wheather natural, derived or synthetic - are exceptionally well-suited for various purposes and find applications across a wide range of scenarios [1] (see Figure 1). Among the long list of applications of therapeutic peptides [2] are: combating cancer [3], microbial infections [4] (Figure 2), viral infections [5], and neurodegenerative diseases [6], among others. Peptides have the ability to autonomously organize into various structures, some of them being: and nanostructures [7], microstructures [8], facilitating their use in injectable hydrogels [9], treatment of tumorous growths [10], and drug delivery [11].
Metal ions have the ability to facilitate the self-assembly of peptides, leading to the formation of supra-molecular structures. These structures are able to serve various functions, such as: catalysts [12,13] or forming assemblies with nanocavities that can be modified, potentially enabling enantioselective recognition [14] (Figure 3).
Peptides serve as recognition elements in molecular probes because of their specific binding capabilities [15,16]. Additionally, their capacity to bind to metals is employed for direct detection of those metals [17,18]. Metallopeptides are also utilized as electrical probes to identify bacteria [19,20].
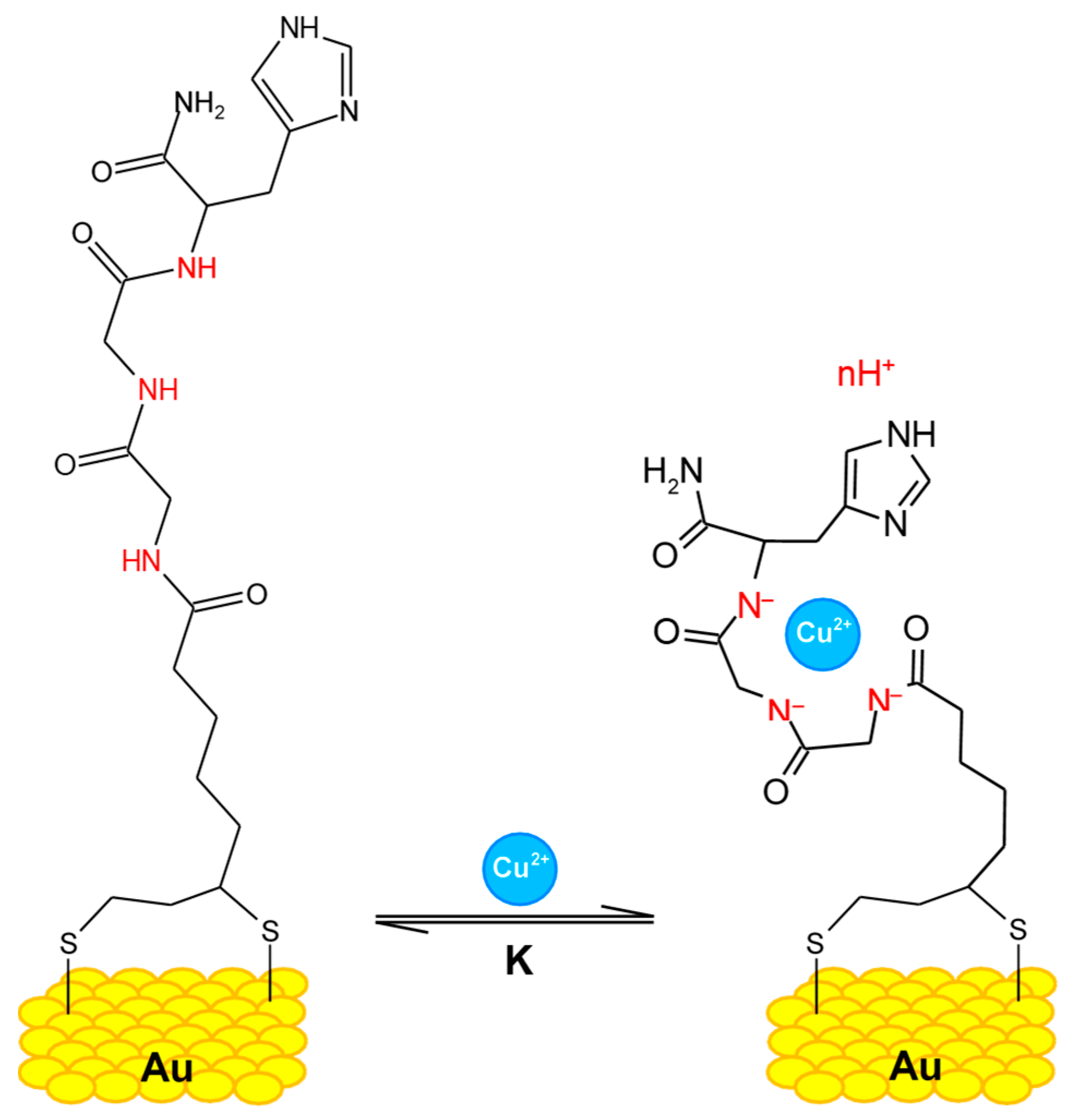
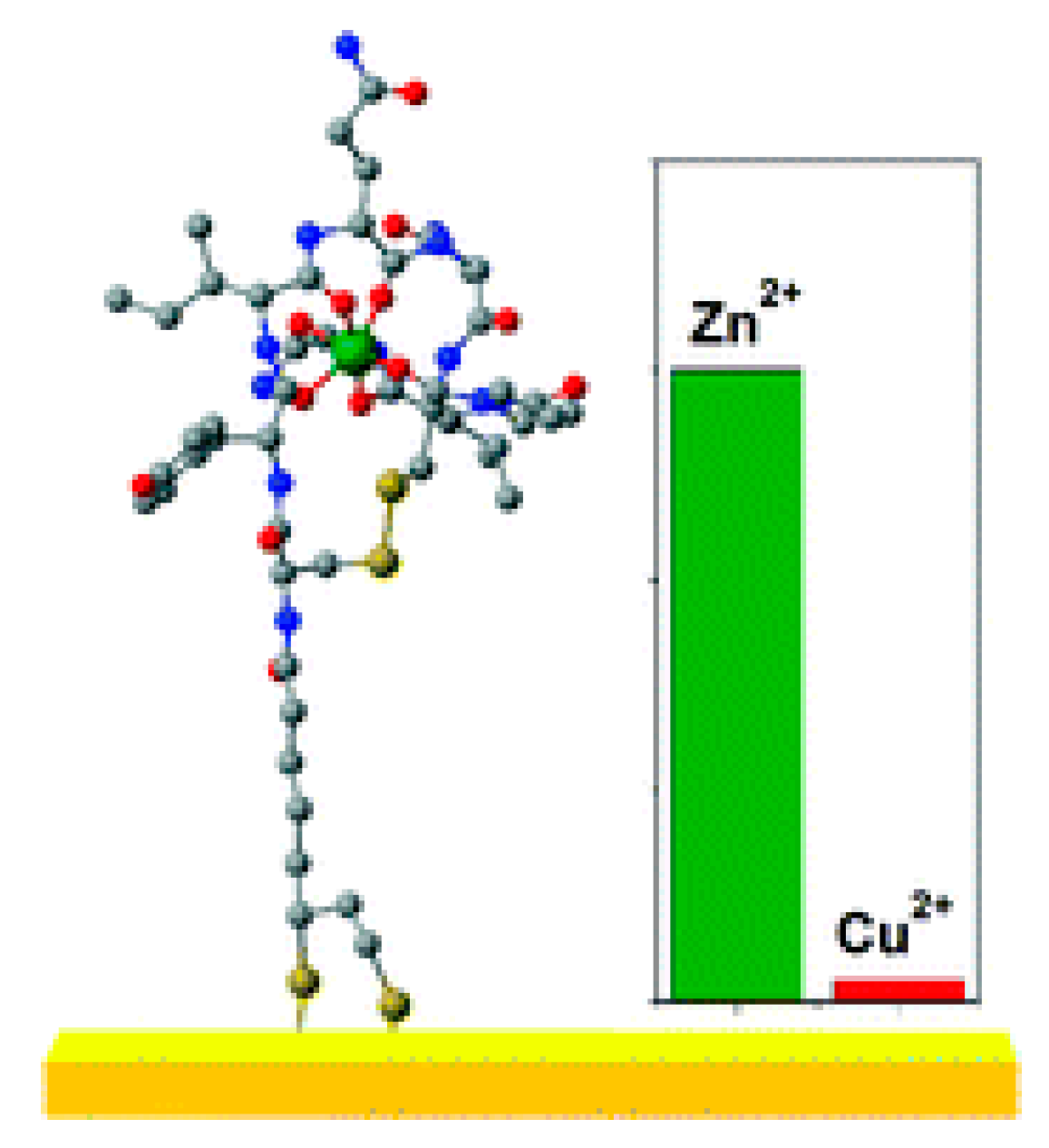
Figure 4.
Glycine–glycine–histidine (Gly–Gly–His, GGH) monolayer on a gold surface and the of Cu2+ ions. Secondary amines, carrying different charges depending on the electrolyte's pH, are indicated in red [17].
Figure 4.
Glycine–glycine–histidine (Gly–Gly–His, GGH) monolayer on a gold surface and the of Cu2+ ions. Secondary amines, carrying different charges depending on the electrolyte's pH, are indicated in red [17].
Figure 5.
A new Zn2+ selective oxytocin-based biosensor that utilizes the terminal amine for surface anchoring, also preventing the response to Cu2+ [18].
Figure 5.
A new Zn2+ selective oxytocin-based biosensor that utilizes the terminal amine for surface anchoring, also preventing the response to Cu2+ [18].
Metal-peptide complexes offer chemical stabilization to the metal through chelation. Within the food industry, peptides function as chelators, enhancing the bioavailability of mineral supplements by safeguarding them against oxidation and altering their solubility properties [21,22].
Additionally, peptides’ metal-binding capacity has been harnessed to enhance the biocompatibility of gold nanoclusters for imaging [23], and bound lanthanide serves as an imaging agent [24]. Moreover, the specific binding between natural peptides and heavy metals can be utilized for metal remediation [25,26].
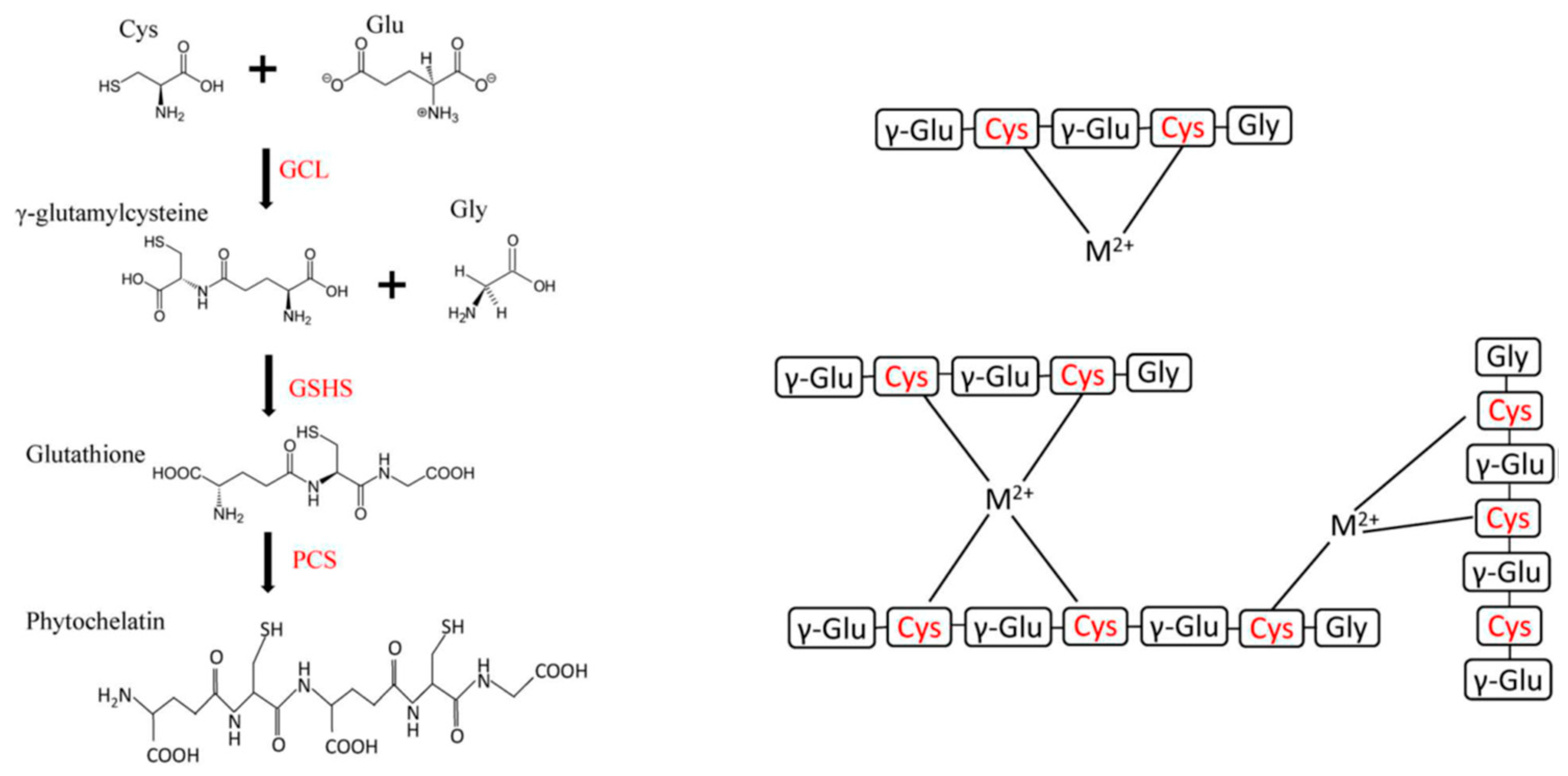
Figure 6.
Structure and biosynthetic pathways of glutathione (GSH) and phytochelatin (PC). M2+ = bivalent metal cations, whereas cysteine residues are indicated in red [25].
Figure 6.
Structure and biosynthetic pathways of glutathione (GSH) and phytochelatin (PC). M2+ = bivalent metal cations, whereas cysteine residues are indicated in red [25].
The literature contains various reviews on the subject [1,22,23,25,27,28,29,30,31,32,33]. Some of metal complexes can be used to as potential therapeutics to treat Alzheimer’s [27,34,35,36], neurodegenerative disorders, Wilson's [37], Parkinson’s disease [38,39] and Menkes disease [40] (see Table 1).
Some essential elements
Copper (I)
Copper holds a crucial status as a metal essential for various biological functions, contributing significantly to both structural integrity and biochemical processes. Facilitating its transportation are specialized copper chaperones, a necessity due to its inherent toxicity, particularly stemming from its capability to transition between its reduced form, Cu(I), and oxidized state, Cu(II) [41]. In the realm of structural NMR (Nuclear Magnetic Resonance), the distinction between these two states is stark, as Cu(I) exhibits diamagnetic properties while Cu(II) manifests paramagnetic behavior. The handling of Cu(I) samples poses a primary challenge, demanding meticulous preparation within a glove box environment and ensuring airtight sealing to prevent oxidation upon exposure to air [42].
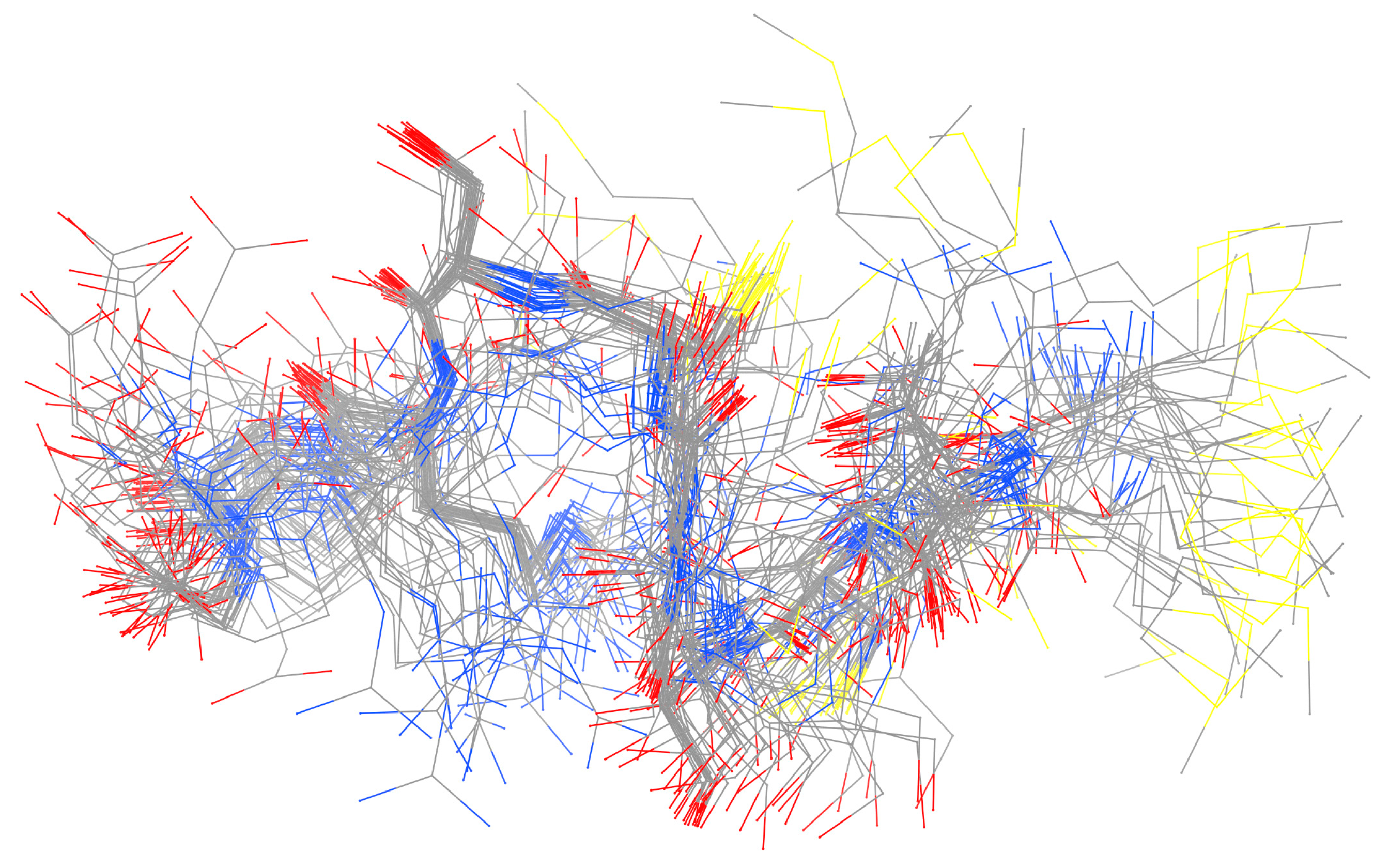
Figure 7.
Ensemble derived from NMR data of copper-bound peptide [42].
Figure 7.
Ensemble derived from NMR data of copper-bound peptide [42].
In exploring the binding sites, both linear and cyclic peptides have served as valuable models, mimicking the conserved sequence essential for copper chaperone interactions. Investigation into their binding modes has been conducted through mutation studies and manipulation of environmental factors. For instance, altering the conserved Met residue within the binding sequence severely hindered copper complexation [38], while adjustments in pH levels led to varying binding modes [43]. These findings imply that pH conditions can influence the coordination environment of the metal, highlighting the utility of peptide-metal models in elucidating potential release mechanisms.
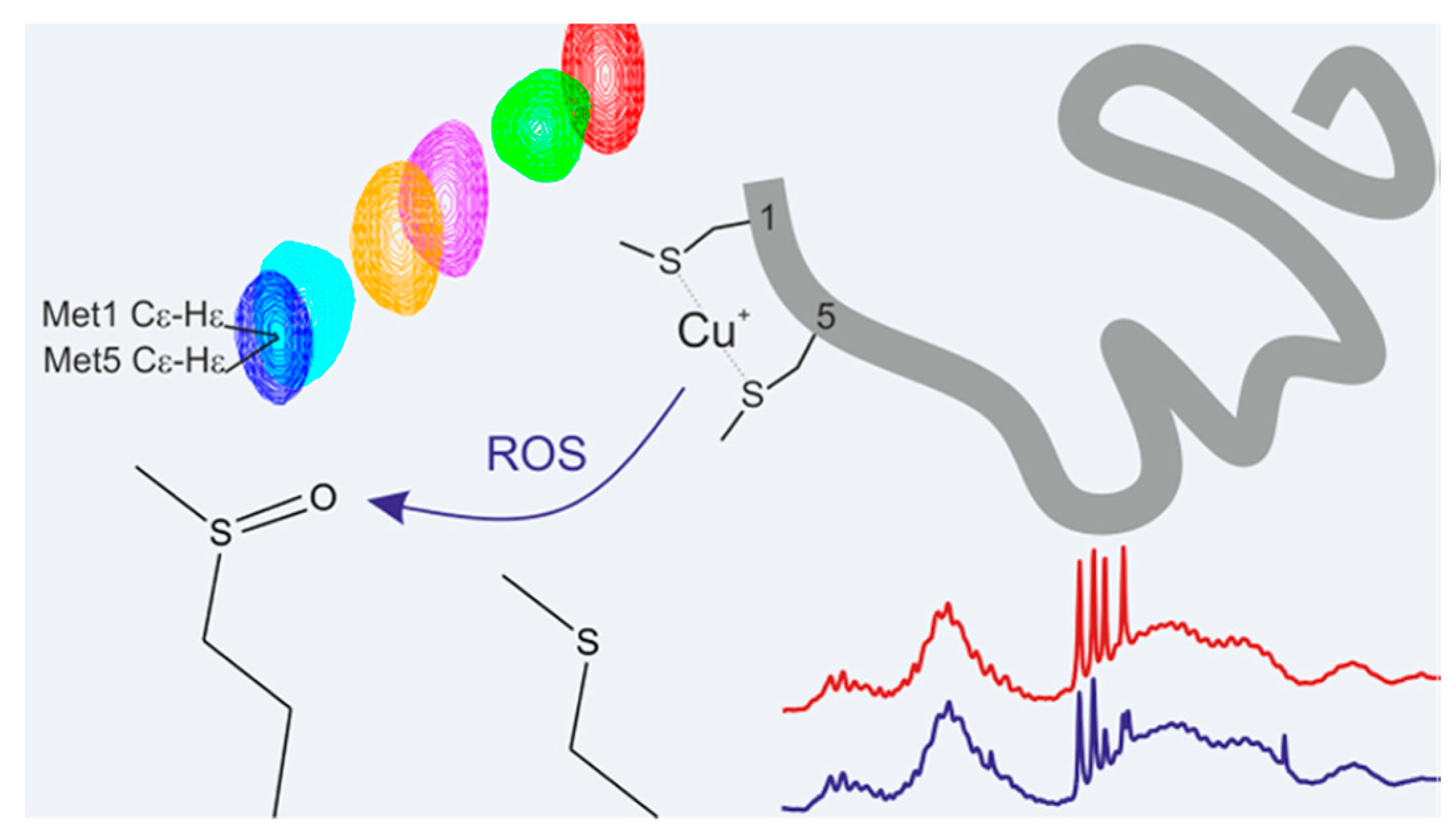
Figure 8.
Possible mechanism of action of copper complex [38].
Figure 8.
Possible mechanism of action of copper complex [38].
Further structural examinations focused on the interaction between Cu(I) and the N-terminal A 16 fragment of amyloid beta (A) sought to ascertain the involvement of all three His residues in metal ion binding [44]. Notably, these studies were conducted on small peptides derived from the binding sites of larger molecules, underscoring their relevance in dissecting intricate molecular interactions.
Copper (II)
Copper(II) stands out as the heaviest among the paramagnetic essential metals, attracting considerable attention in studies involving various metal-binding proteins [45]. Research endeavors have employed signal broadening techniques to pinpoint the residues involved in binding [46], with such phenomena often offering insights into the sequence of binding events [47]. When proton (1H) spectra exhibit broadening effects, complementary information can be extracted from the corresponding carbon-13 (13C) spectra [48]. Typically, these investigations necessitate the initial assignment of the 1H spectrum followed by titration experiments with Cu(II) to identify the specific binding groups. Subsequently, the formation of the bound complex can be confirmed either through experimental means [47] or through computational simulations [49]. This allows for the positioning of the Cu(II) ion within the identified binding site, ultimately yielding a detailed depiction of the bound structure.
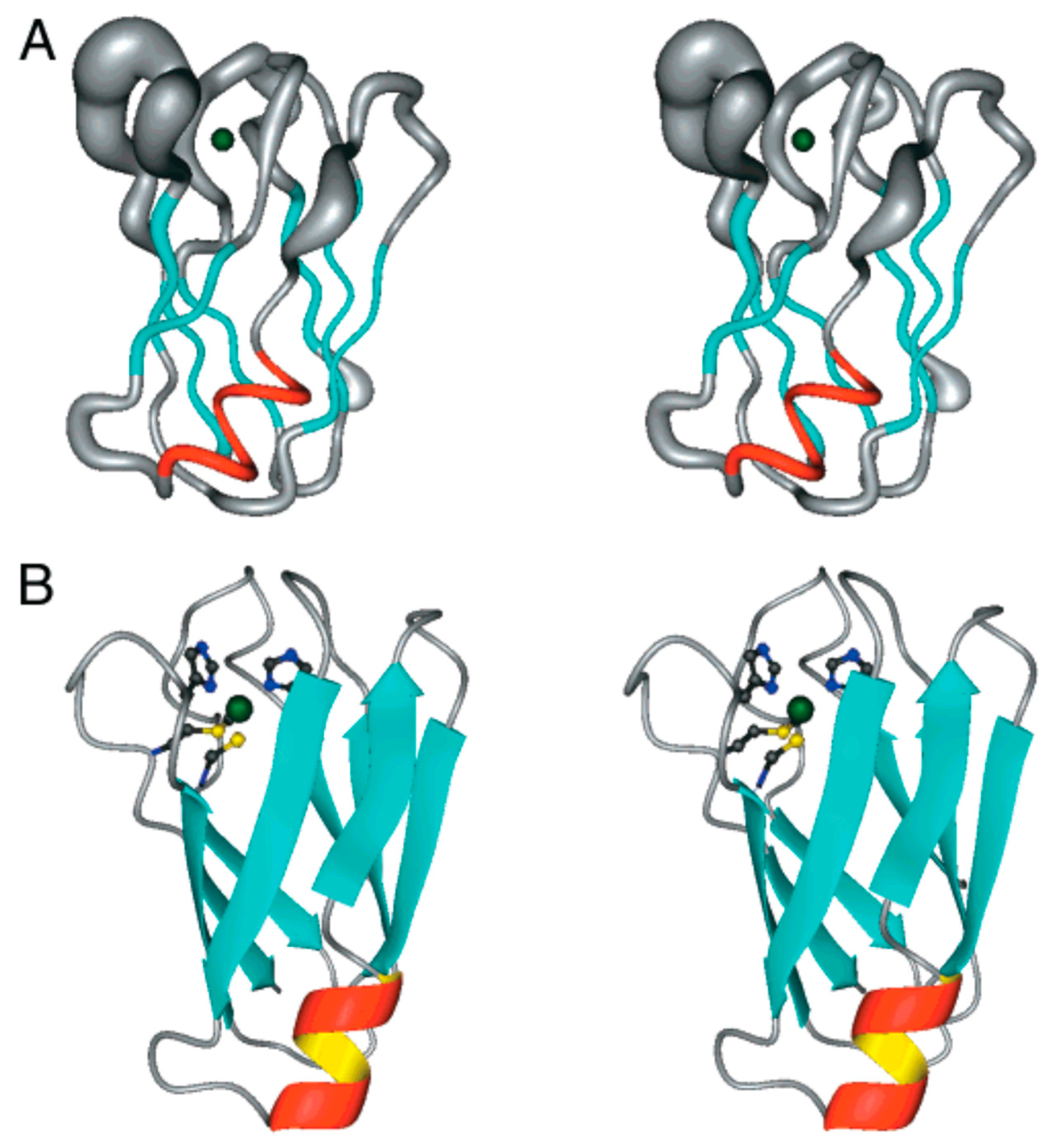
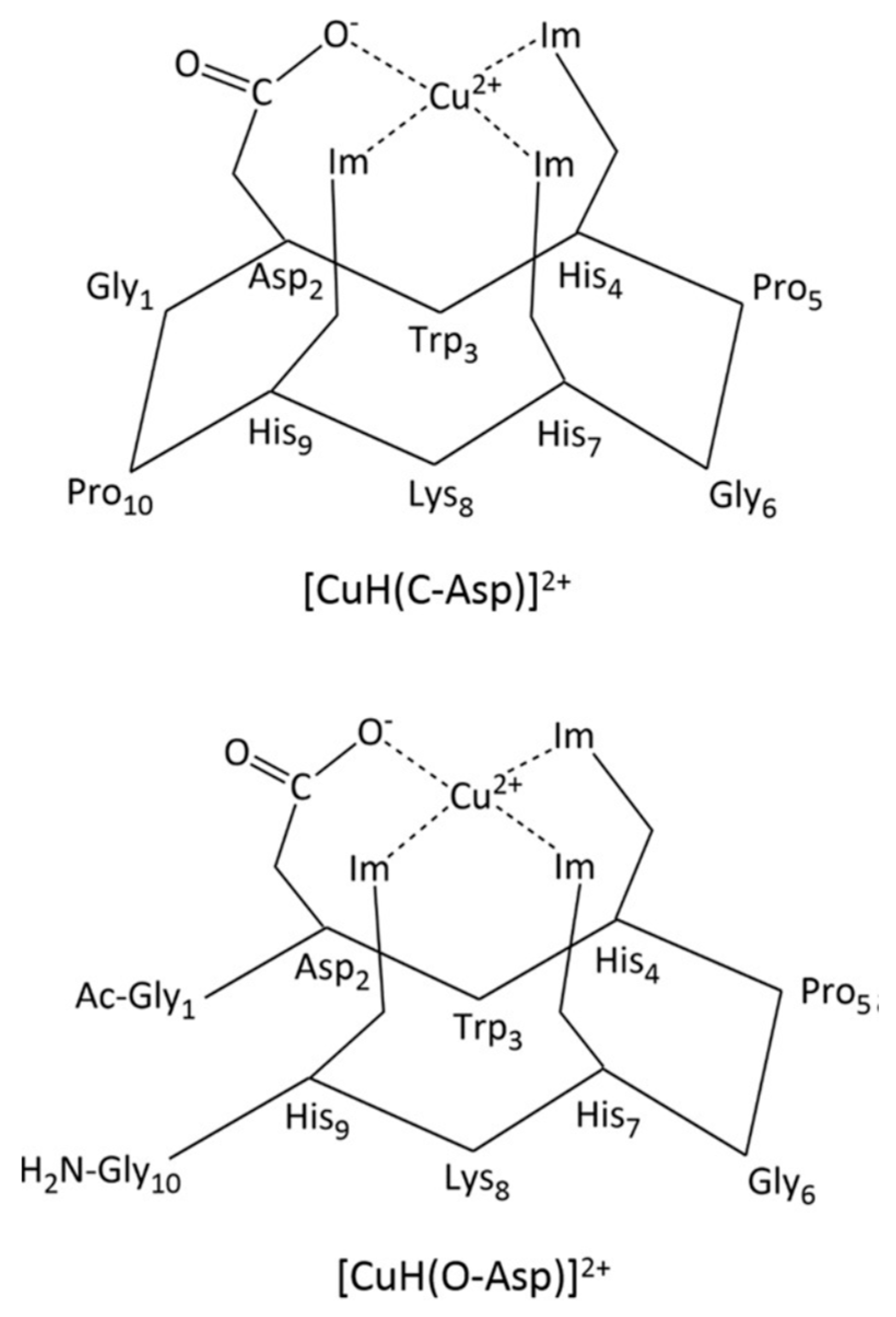
Figure 9.
Metal-binding proteins presented by Bertini et al. [45].
Figure 9.
Metal-binding proteins presented by Bertini et al. [45].
Figure 10.
Schematic representation of the proposed structures (Im=imidazole) [46].
Figure 10.
Schematic representation of the proposed structures (Im=imidazole) [46].
Recently, Georgieva et al. have proposed the formation of complexes between Cu(II) and hemorphin peptide derivatives in an aqueous solution with a pH of 10.98 [50]. Through meticulous examination of spectroscopic data and voltammetric calculations, complete agreement has been reached, confirming the formation of stable copper(II) complexes with peptides in aqueous solutions. These complexes exhibit a 1:2 stoichiometry for Cu-H-V, Cu-AH-V, Cu-NH7C, and Cu-NCH7, while a 1:1 stoichiometry is observed for Cu-AC-V and Cu-C-V. Notably, the virucidal activity against human respiratory syncytial virus and adenovirus (HRSV-2) at 30 and 60 minutes remains weakly affected by these complexes, similar to the starting peptide compounds [50]. Figure 11 illustrates the structures of two of these newly identified complexes.
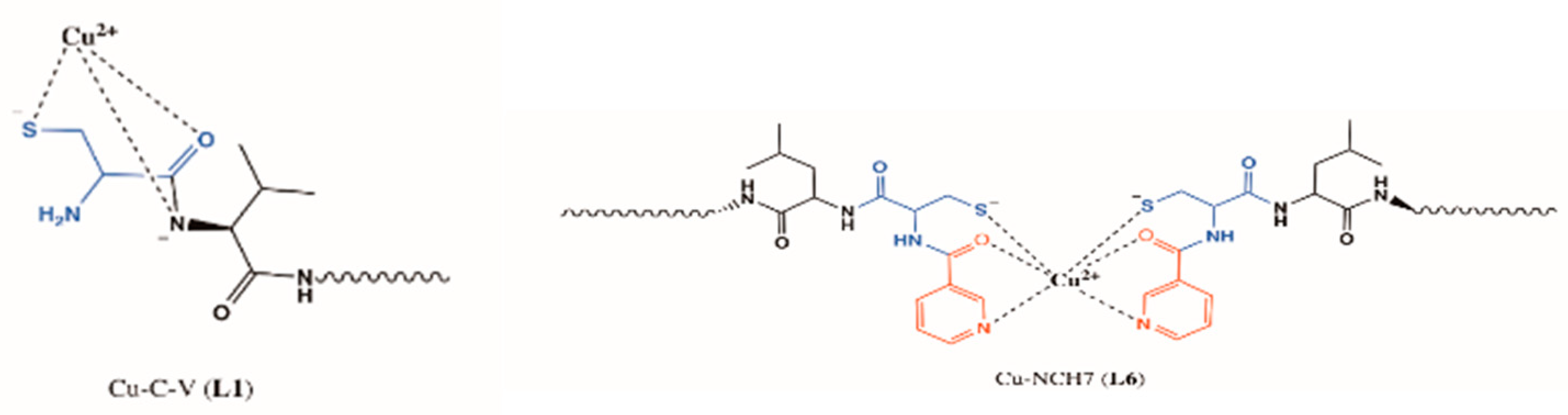
Figure 11.
Schematic structure of Cu(II)-peptide complexes: Cu-C-V(L1) and Cu-NCH7 (L6) [50].
Figure 11.
Schematic structure of Cu(II)-peptide complexes: Cu-C-V(L1) and Cu-NCH7 (L6) [50].
Hickey et al. documented the discovery of a copper complex featuring a styrylpyridine group (depicted in Figure 12), which exhibited an affinity for Aβ plaques within human brain tissue [36].
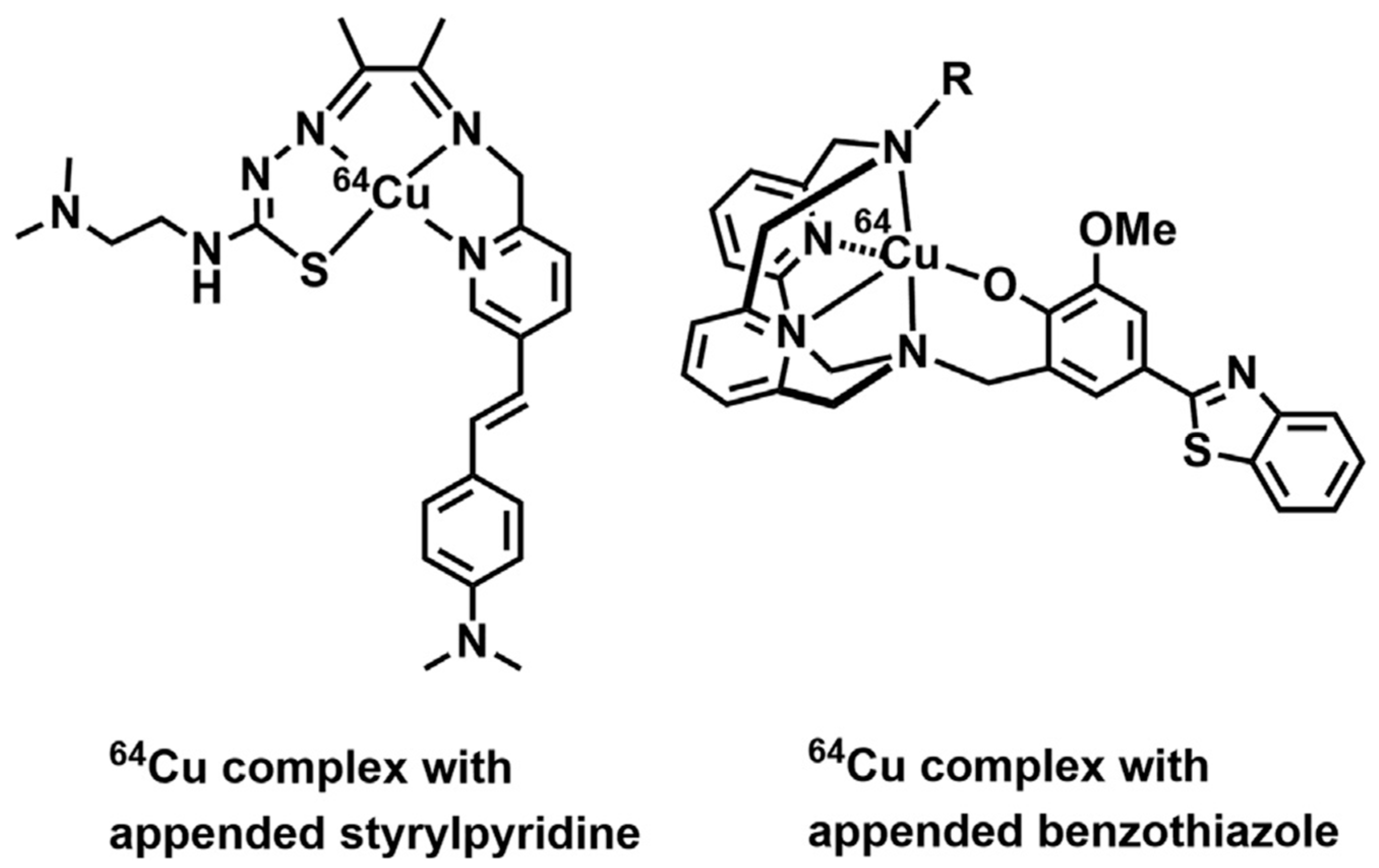
Figure 12.
64Cu complexes used as plaque imaging agents in Alzheimer’s disease [36].
Figure 12.
64Cu complexes used as plaque imaging agents in Alzheimer’s disease [36].
Given the challenges in directly measuring copper(II)-bound complexes, diamagnetic metals with square planar binding geometry often serve as substitutes for deriving the bound structure. Through line-broadening analysis, the position of copper binding can be inferred. If there exists experimental evidence indicating similarity in binding modes, Cu(II) can be introduced into structure calculations, enabling the determination of copper(II)-bound structures. This methodology has been successfully applied to peptide-metal complexes utilizing palladium(II) [51], silver(I) [52], and nickel(II) [53,54].
Line-broadening analysis of proton (1H) spectra has been instrumental in identifying residues involved in binding Cu(II) to model copper(II)-binding sites [55], as well as in copper(II)-mediated aggregation processes [56]. Additionally, line-broadening effects in carbon-13 (13C) spectra can be utilized [57], while chemical shift deviations can be monitored through various experiments including 1H-1D and 2D 15N-HSQC, 13C-HSQC, and 1H-1H TOCSY experiments. Furthermore, precise determination of the binding nitrogen in histidine residues has been achieved through proton spin-lattice relaxation rate studies [58].
Zinc (II)
Zinc stands out as a crucial ion involved in structural, catalytic, and regulatory functions within proteins, being present in approximately 10% of all known proteins [29,59]. Notably, zinc finger proteins represent a prominent category among these, showcasing their flexibility and stability through metal-binding interactions. To unravel the intricacies of zinc finger binding, both short and longer peptides have been extensively utilized. These peptides serve as invaluable tools for modeling various aspects of zinc finger behavior, including metal coordination, folding dynamics, and actual binding processes [60,61,62].
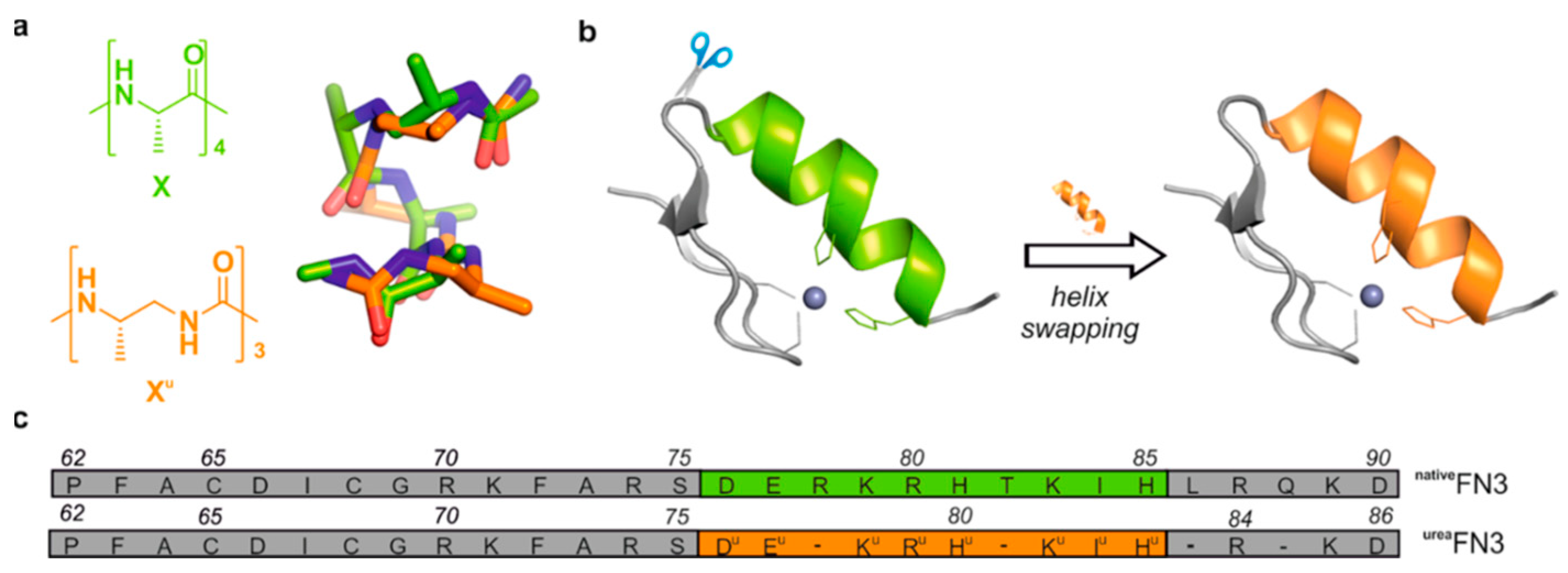
Figure 13.
Design guided by the structure of composite proteins containing artificial helical foldamer region. (a) Comparison of α-peptide (green) and oligourea (orange) backbones [62].
Figure 13.
Design guided by the structure of composite proteins containing artificial helical foldamer region. (a) Comparison of α-peptide (green) and oligourea (orange) backbones [62].
In the realm of peptide modeling, cyclic peptides featuring linear tails have emerged as particularly promising candidates. These structures exhibit notable conformational and thermodynamic stability when compared to their linear counterparts, effectively mimicking the zinc-ribbon fold characteristic of zinc fingers. Moreover, cyclic peptides demonstrate enhanced binding affinity towards Zn(II) ions, underscoring their utility in zinc finger modeling [63]. Through peptide-based investigations, insights into secondary structural elements common in zinc fingers have been gleaned, shedding light on their pivotal role in both folding and zinc binding processes. Notably, peptide engineering efforts have led to the development of modified peptides that retain tertiary folds and stability comparable to their natural zinc finger protein counterparts, even with substantial alterations to their native residues [64].
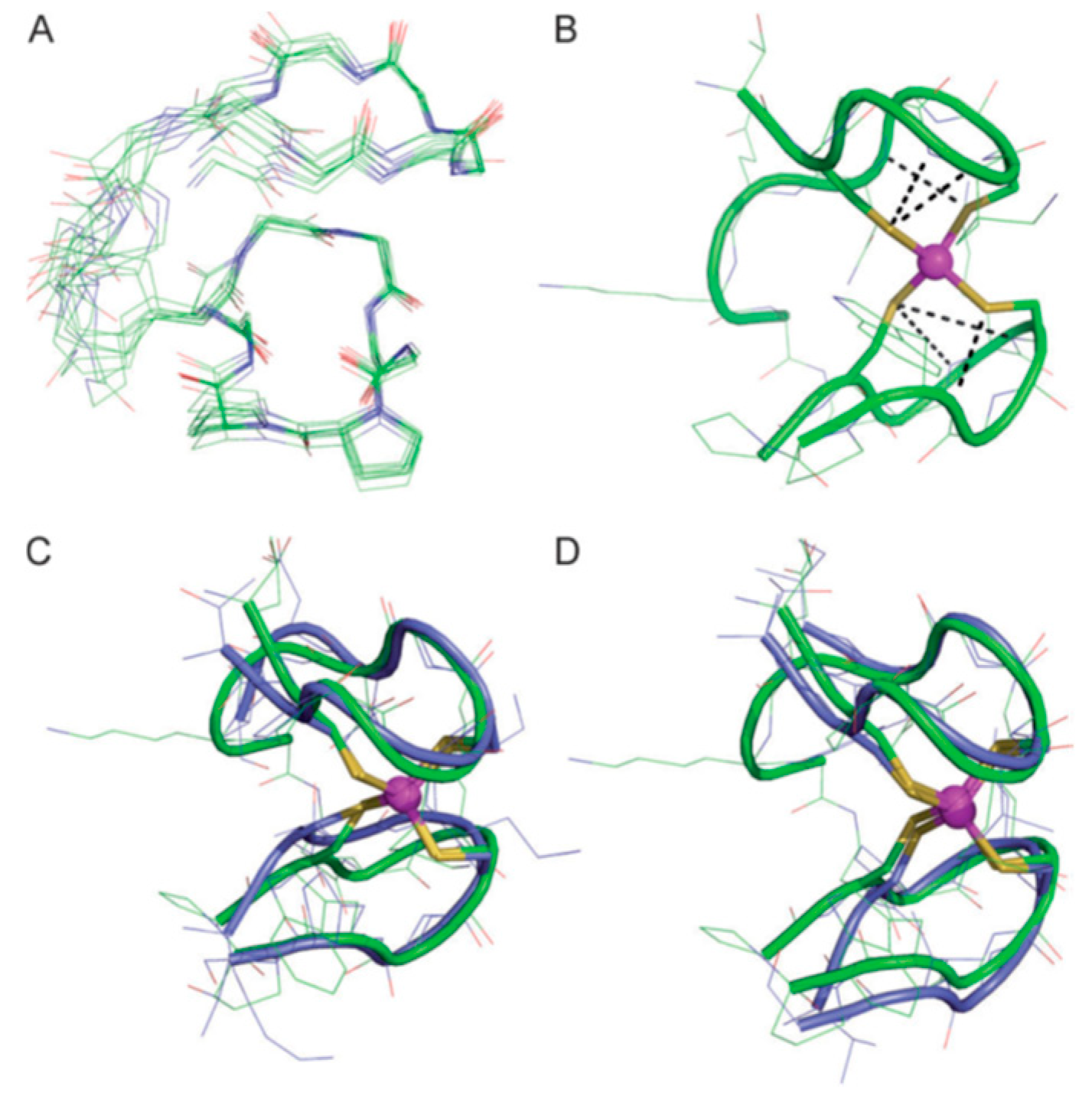
Figure 14.
Solution structure of ZnLZR (pH 6.3, 298 K) deduced from NMR studies [63].
Figure 14.
Solution structure of ZnLZR (pH 6.3, 298 K) deduced from NMR studies [63].
The binding of zinc ions can induce significant conformational changes in peptides [65], a phenomenon exemplified by its ability to trigger the oligomerization of amyloid beta, a 42-amino acid polypeptide associated with Alzheimer's disease. To study such processes effectively, truncated peptides preserving the beta-sheet formation region while minimizing complete oligomerization have been employed. For instance, investigations into the familial Taiwanese mutation D7H region of amyloid beta, which impacts zinc-induced oligomerization, have been facilitated through the use of stable homodimers formed via zinc-binding [34,35].
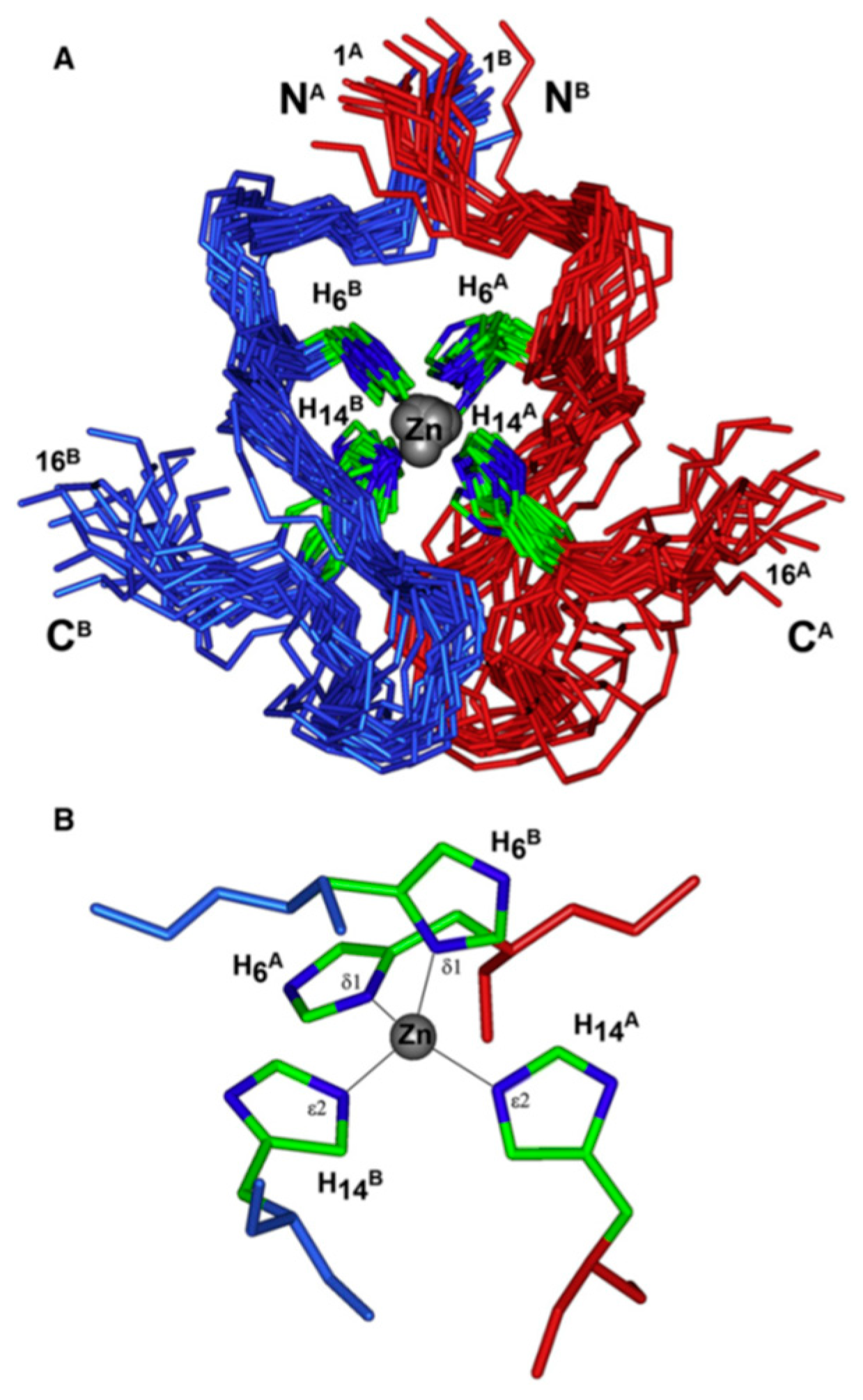
Figure 15.
Structure of the rat Aβ(1–16) dimer complexes with Zn ions in solution [34].
Figure 15.
Structure of the rat Aβ(1–16) dimer complexes with Zn ions in solution [34].
Furthermore, exploring the resistance of rats to Alzheimer's disease has provided valuable insights into zinc-induced dimerization within amyloid beta proteins. Utilizing truncated peptides, researchers have elucidated the interface involved in zinc-induced dimerization, showcasing a potential avenue for the rational design of drug compounds aimed at disrupting plaque-forming processes.
Cobalt (II) and (III)
Cobalt, existing in both its (II) and (III) oxidation states, though present in trace amounts within the body, holds vital significance as an essential element. However, when present in excess, cobalt poses toxicity risks attributed to its capacity for generating reactive oxygen species and displacing iron in metalloenzymes, thereby rendering them inactive [31]. Predominantly paramagnetic, cobalt manifests distinct characteristics in its (III) state, where it can adopt both high- and low-spin configurations, the latter being diamagnetic [31]. For instance, the elucidation of a diamagnetic Co(III) complex with a peptide-porphyrin conjugate was achieved through conventional methodologies [66]. Notably, the large paramagnetic chemical shifts (PCSs) and minimal paramagnetic relaxation enhancements (PRE) exhibited by high-spin Co(II) render it conducive to structure determination [30].
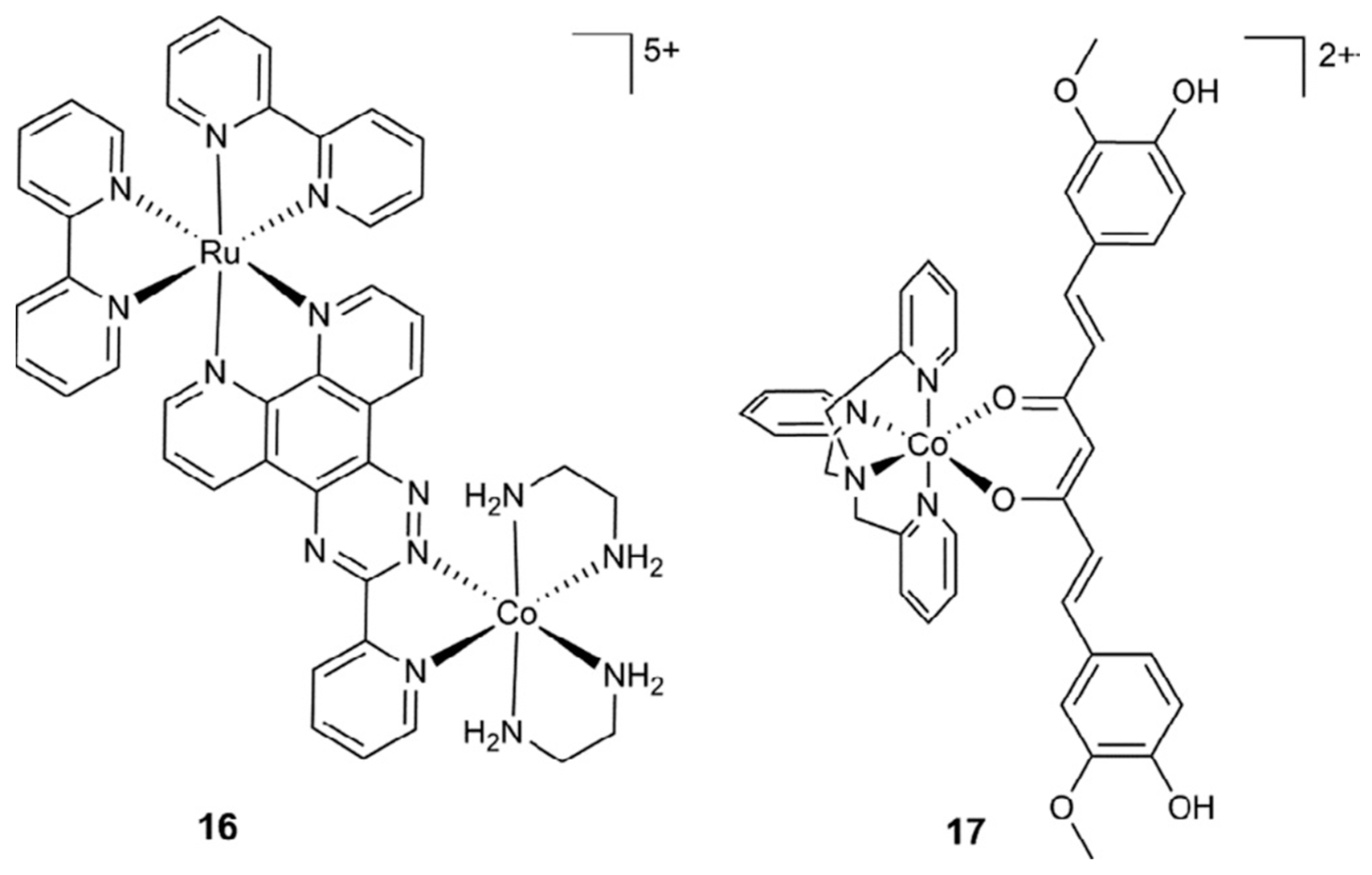
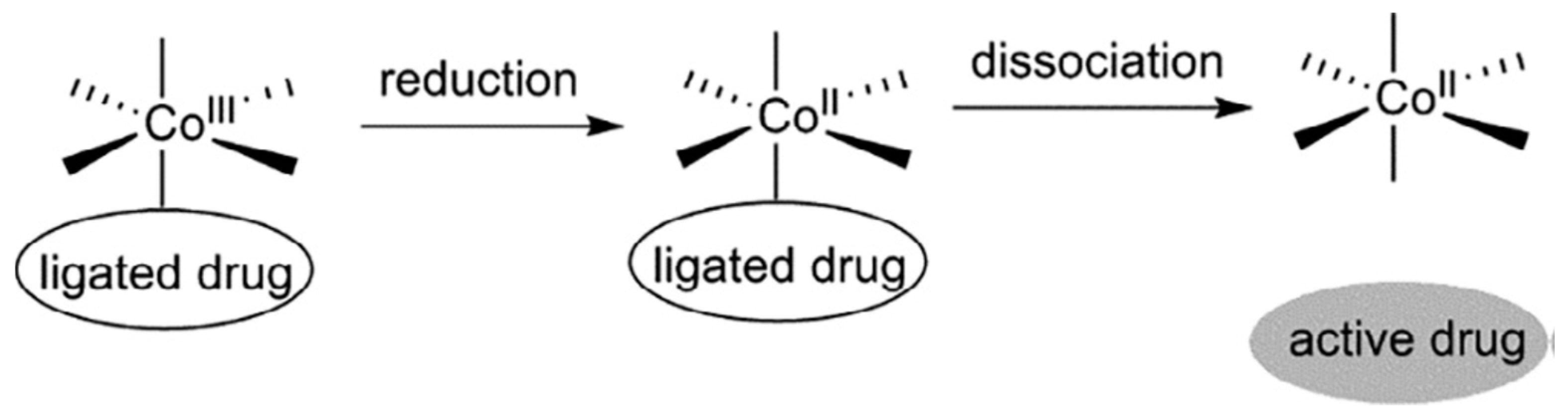
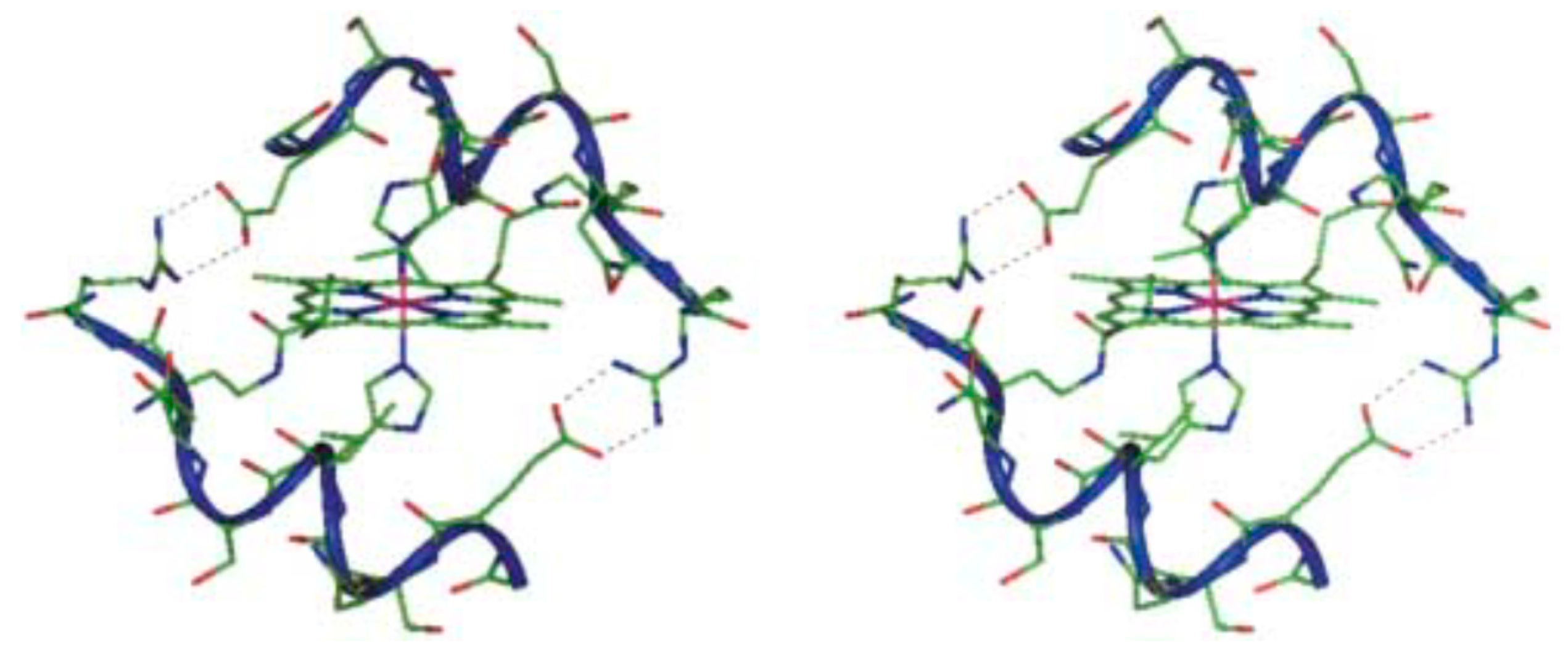
Figure 16.
The structure of some Co(III) complexes - light-activated Co(III) prodrugs [31].
Figure 16.
The structure of some Co(III) complexes - light-activated Co(III) prodrugs [31].
Figure 17.
Generalised mechanism of action of cobalt complexes for reductionactivated drug delivery [31].
Figure 17.
Generalised mechanism of action of cobalt complexes for reductionactivated drug delivery [31].
Figure 18.
Molecular structure of Co(III)-mimochrome IV. Stereo view of the average structure, as obtained from NMR experimental data and RMD calculations [66].
Figure 18.
Molecular structure of Co(III)-mimochrome IV. Stereo view of the average structure, as obtained from NMR experimental data and RMD calculations [66].
Similarly, the binding site of a cobalt(III)-Schiff base complex to another amyloid beta protein fragment was elucidated using 1H-NMR, with binding histidines identified through line-broadening effects [67].
Figure 19.
Transition metal complexes proposed by Heffern et al. [67].
Figure 19.
Transition metal complexes proposed by Heffern et al. [67].
Further insights into cobalt binding were gleaned from an investigation into cobalt(II) binding to fibrinopeptide B, a factor implicated in thrombosis. Although the peptide structures were determined in the presence of cobalt(II) and gadolinium(III), the metal ions were not visualized within the structures [68]. Moreover, a 30-amino acid peptide initially recognized for its manganese-binding capabilities, derived from a decapeptide repeat in the calcium protein Cap43, was also found to exhibit affinity for Co(II). Line-broadening observed during the interaction enabled the identification of binding histidine residues, while the pH range conducive to binding was elucidated as part of the binding mechanism. Subsequent modeling efforts generated and minimized postulated coordination spheres of Co(II) based on NMR data, utilizing software like HyperChem™ [69].
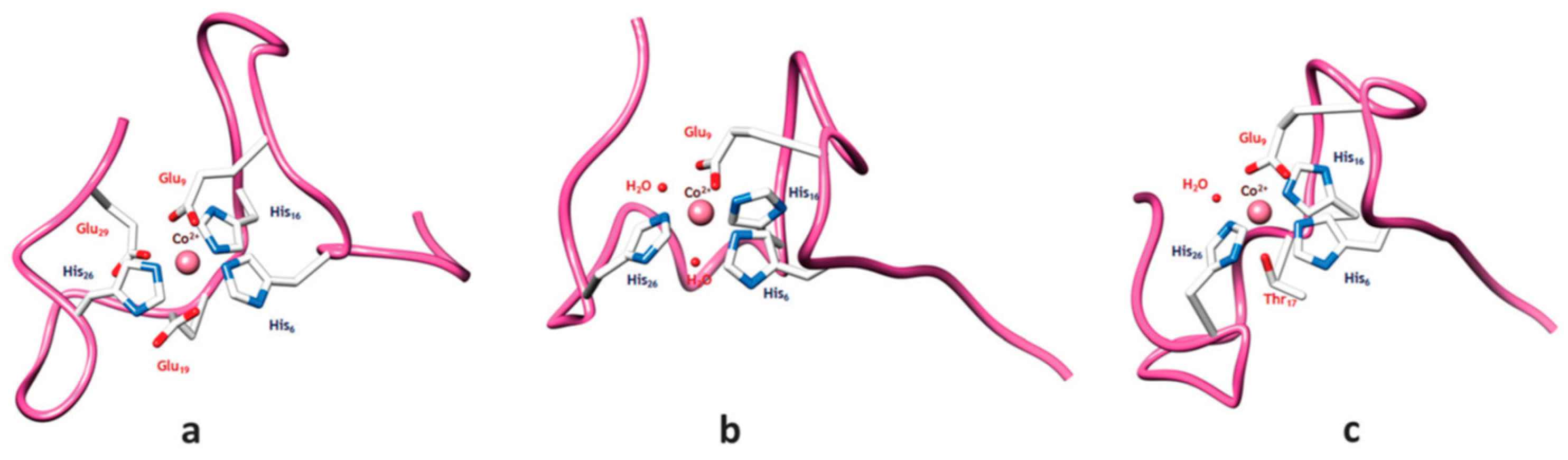
Figure 20.
Model of the most likely coordination spheres of Co(II) with the multi-histidine peptide fragment; (a) Co-{3Nε(Im)-3O(Glu-COO−)}, (b) Co-{3Nε(Im)-3O(1Glu-COO−, 2H2O)}, (c) Co-{3Nε(Im)-3O(1 Glu-COO−, 1 Thr-O, 1 H2O)} [69].
Figure 20.
Model of the most likely coordination spheres of Co(II) with the multi-histidine peptide fragment; (a) Co-{3Nε(Im)-3O(Glu-COO−)}, (b) Co-{3Nε(Im)-3O(1Glu-COO−, 2H2O)}, (c) Co-{3Nε(Im)-3O(1 Glu-COO−, 1 Thr-O, 1 H2O)} [69].
Additionally, the conformational alterations of peptides upon binding to chiral cobalt oxide nanoparticles were investigated to explore chiral evolution phenomena. Tripeptide ligands exhibited distinct peak sets attributed to the high PCS of the nanoparticles, facilitating structure determination owing to their minimal PRE effects [70].
Iron (II) and (III)
Iron stands as an indispensable element crucial for oxygen transport within the body, with both excess and deficiency leading to various disorders [32,33]. Existing in both ferric and ferrous forms, iron exhibits paramagnetic properties, and the interconversion between these forms plays a central role in its functionality. Given the facile oxygenation of ferrous to ferric iron ions, meticulous sample preparation within an oxygen-free glove box environment is imperative to mitigate undesired reactions. The following studies underscore innovative strategies for investigating iron, highlighting the efficacy of a multidisciplinary approach.
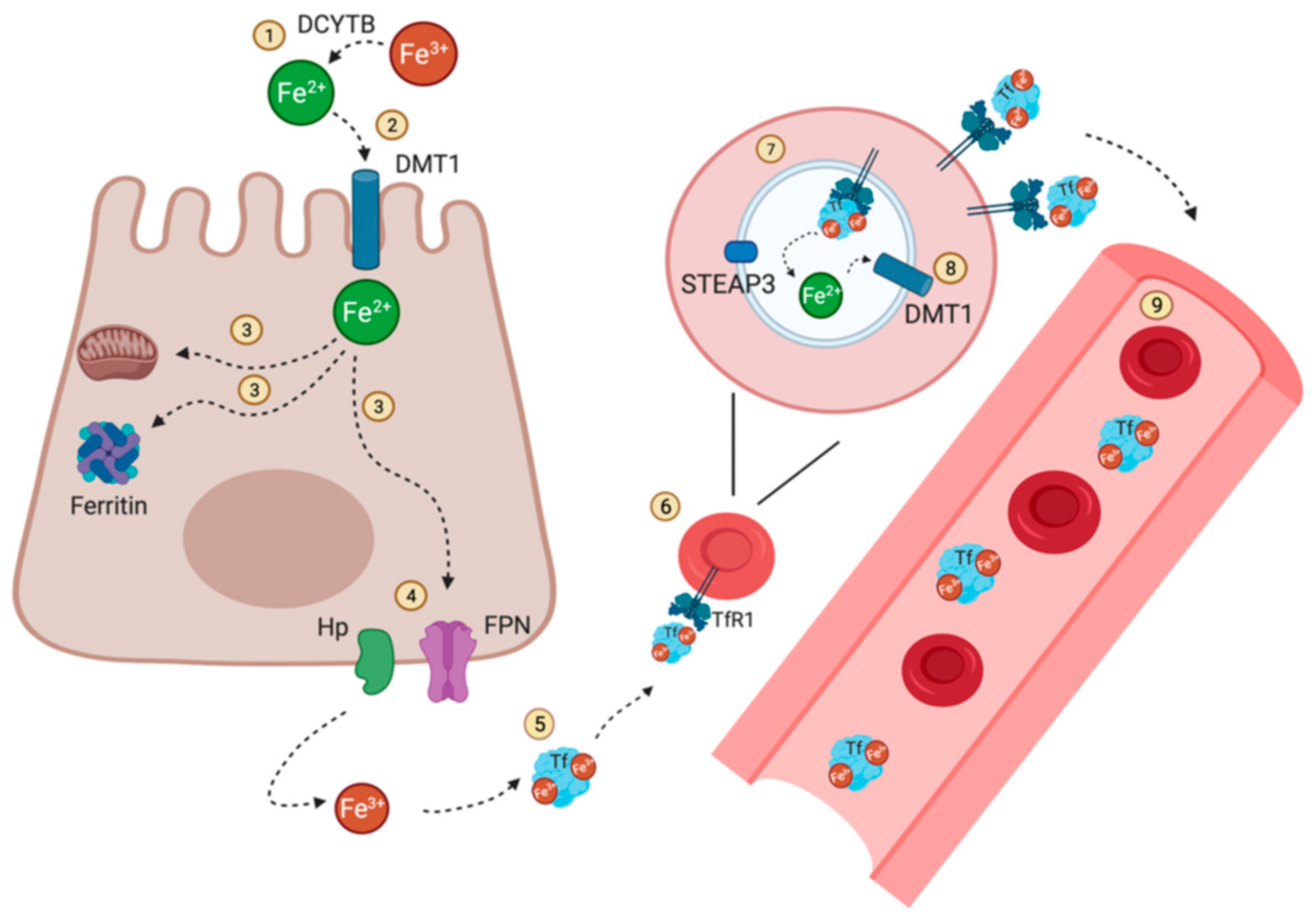
Figure 21.
Iron distribution and circulation [33].
Figure 21.
Iron distribution and circulation [33].
Detection of iron binding is notably achieved through the observation of line-broadening effects proximal to the binding site. For instance, a study involving the grafting of a six-residue iron-binding motif onto a 29-residue peptide utilized NMR spectroscopy to discern line-broadening phenomena indicative of specific interactions between the peptide and Fe(III) ions. Complementary techniques such as circular dichroism, isothermal titration calorimetry, capillary zone electrophoresis, thermal denaturation, and computational modeling were employed to elucidate the binding mode and structure of the peptide model system [71]. In magnetotactic bacteria, biomineralization processes are governed by magnetite-associated proteins featuring short sequences capable of binding iron. Peptides derived from these iron-binding regions were subjected to reactions with Fe(II), Fe(III), Ni(II), and Zn(II) ions, enabling the determination of specificity, binding coefficients, and key binding residues through NMR spectroscopy. Subsequent coprecipitation assays were employed to ascertain the significance of each binding residue [72]. Additionally, computational methodologies were integrated with NMR spectroscopy to elucidate the structures of artificial peptides forming chiral helicate complexes with Fe(II) and Co(II) ions [73]. Furthermore, the utility of 1D and 2D NMR techniques capable of directly detecting paramagnetic complexes was demonstrated using the 8-amino acid microperoxidase-8 bound with heme iron as a model peptide for the cytochrome C binding site. Excitation sculpting with gradients was employed to suppress water signals in Fe(II)-bound samples for acquiring 1D spectra, while a superWEFT pulse sequence was utilized for measuring Fe(III)-bound samples [74,75].
Some Non-Essential Elements
A variety of non-essential metals have been implicated in contributing to the pathogenesis of Alzheimer's disease, prompting investigations into their interactions with amyloid beta-derived peptides to unravel potential mechanisms of action. For instance, studies have explored the interactions of aluminum and palladium with these peptides [51,76].
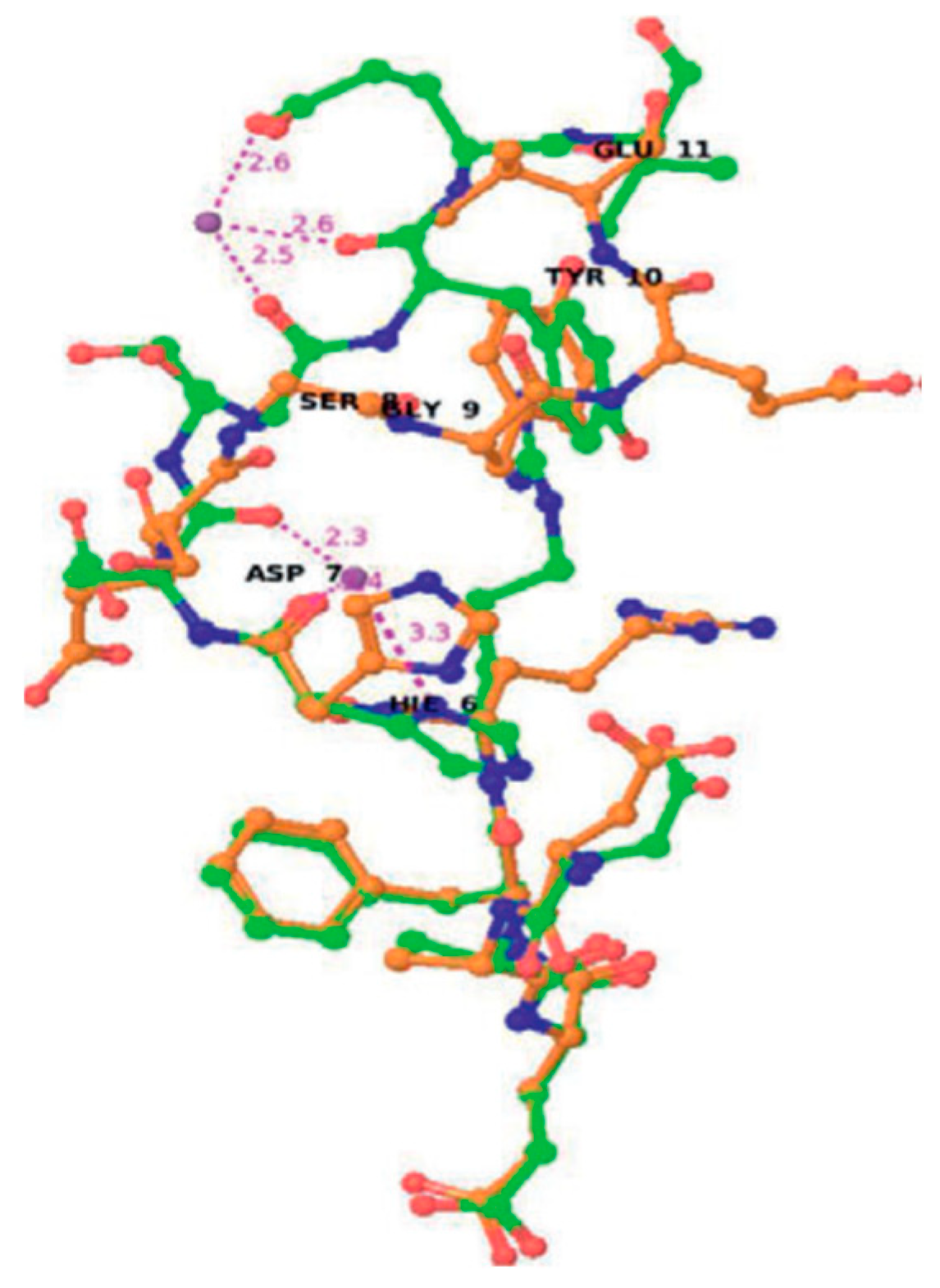
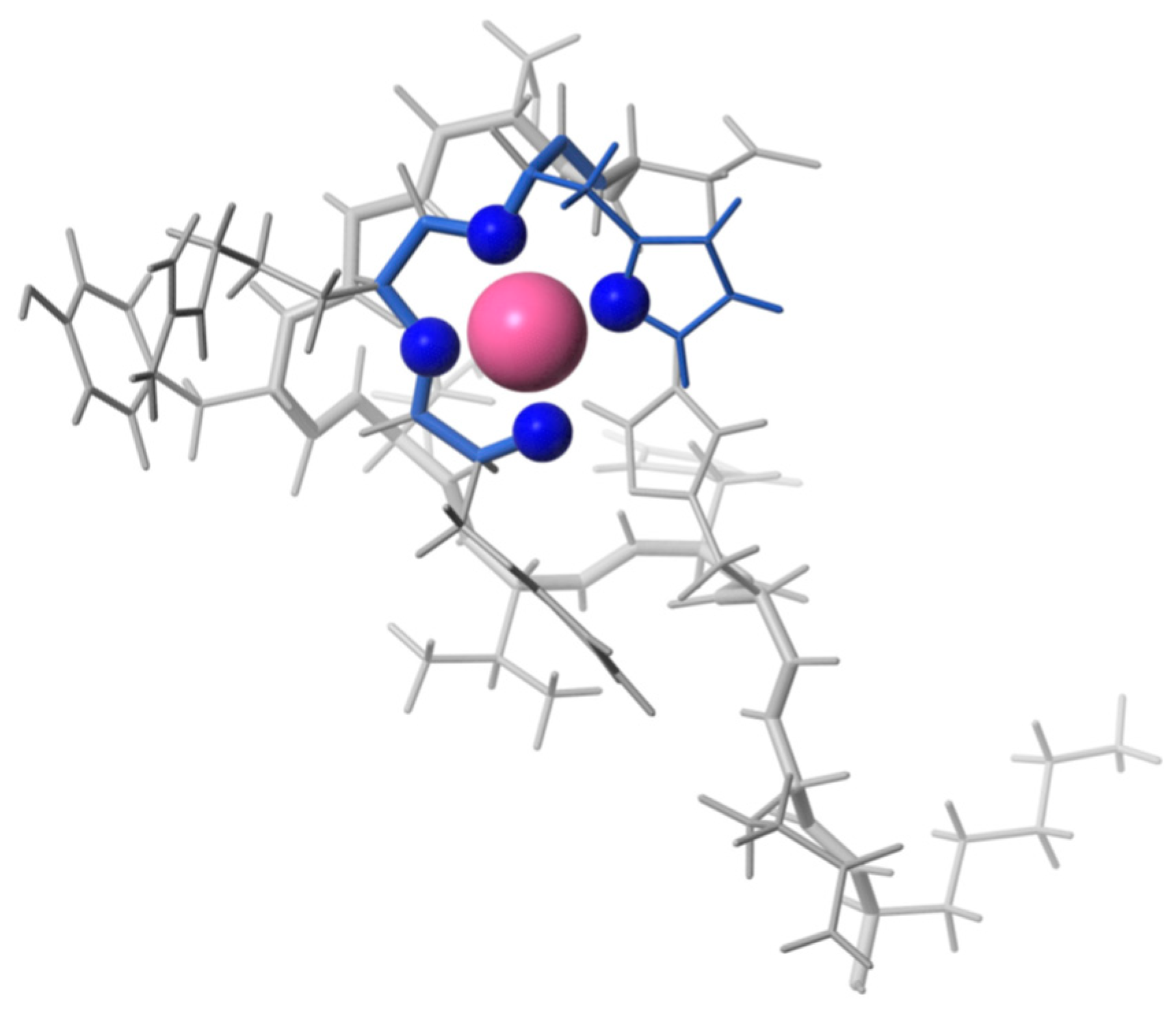
Figure 22.
Overlap of Aβ12 structures with (green) and without metal (orange) M=Al(III [76].
Figure 22.
Overlap of Aβ12 structures with (green) and without metal (orange) M=Al(III [76].
Figure 23.
The 3D structure of Pd(Aβ4-16) complex included the N-terminal ATCUN/NTS motif binding the Pd(II) ion [51].
Figure 23.
The 3D structure of Pd(Aβ4-16) complex included the N-terminal ATCUN/NTS motif binding the Pd(II) ion [51].
Nickel NMR measurements serve as a valuable tool for detecting the binding of nickel ions in diamagnetic form, thereby serving as a model for understanding the behavior of paramagnetic ions that may otherwise pose challenges for structural characterization [77]. Studies comparing different forms of nickel-binding peptides offer insights into the role of nickel complexation in peptide structure and hint at the potential roles of paramagnetic ions. The amino-terminal copper- and nickel-binding (ATCUN) motif, found in numerous proteins, has been extensively investigated using linear and cyclic peptide models with various divalent ions, including Co(II), Ni(II), and others [51,78]. Nickel itself has been implicated in metal-induced toxicity and carcinogenesis, prompting studies on Ni(II)-peptide models derived from the C-terminal of histone H2B to elucidate potential roles in carcinogenesis [79]. Additionally, interactions of Ni(II) with peptides derived from the human Toll-like receptor (hTLR4) have been studied due to nickel's ability to elicit allergic responses [80].
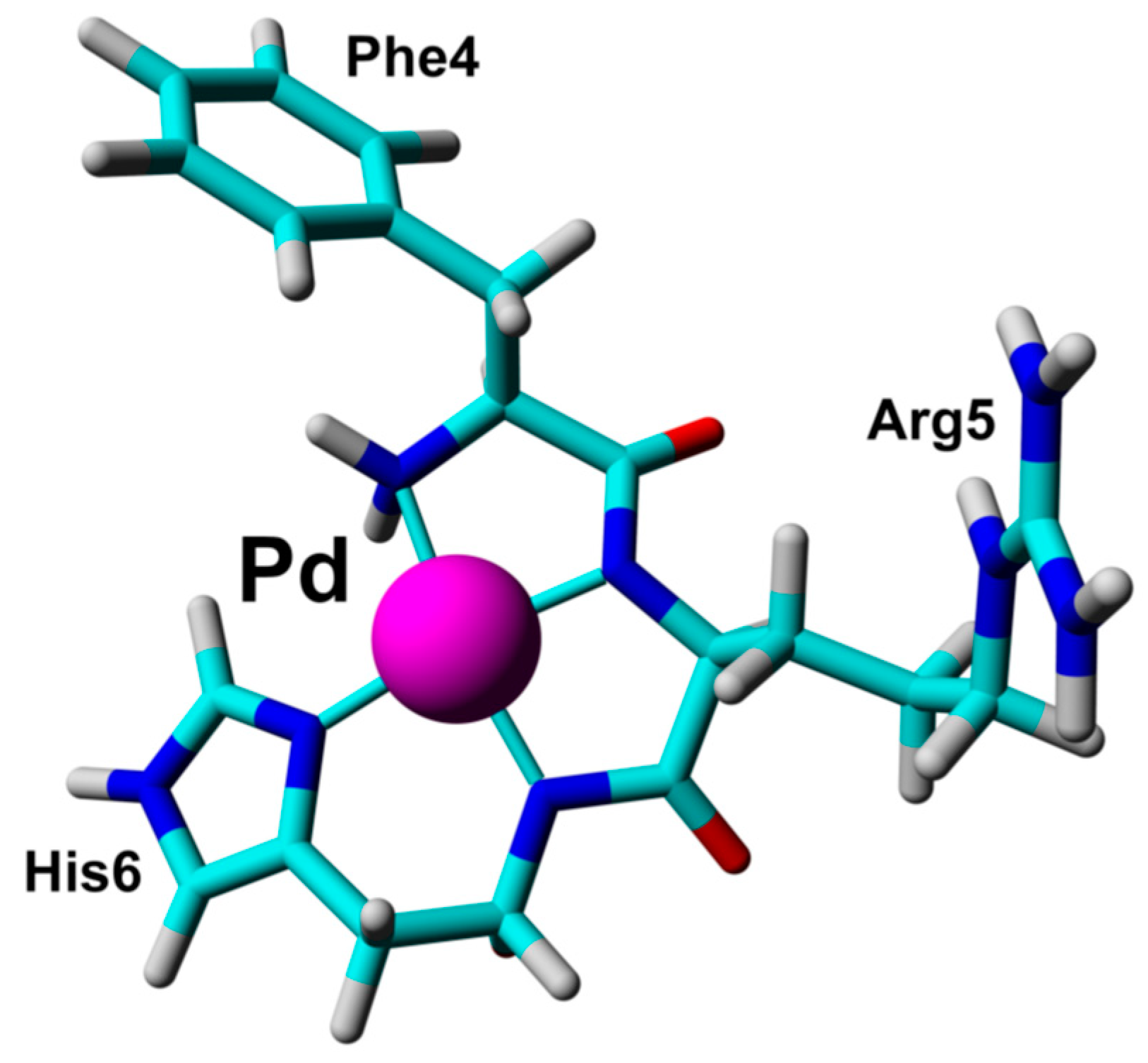
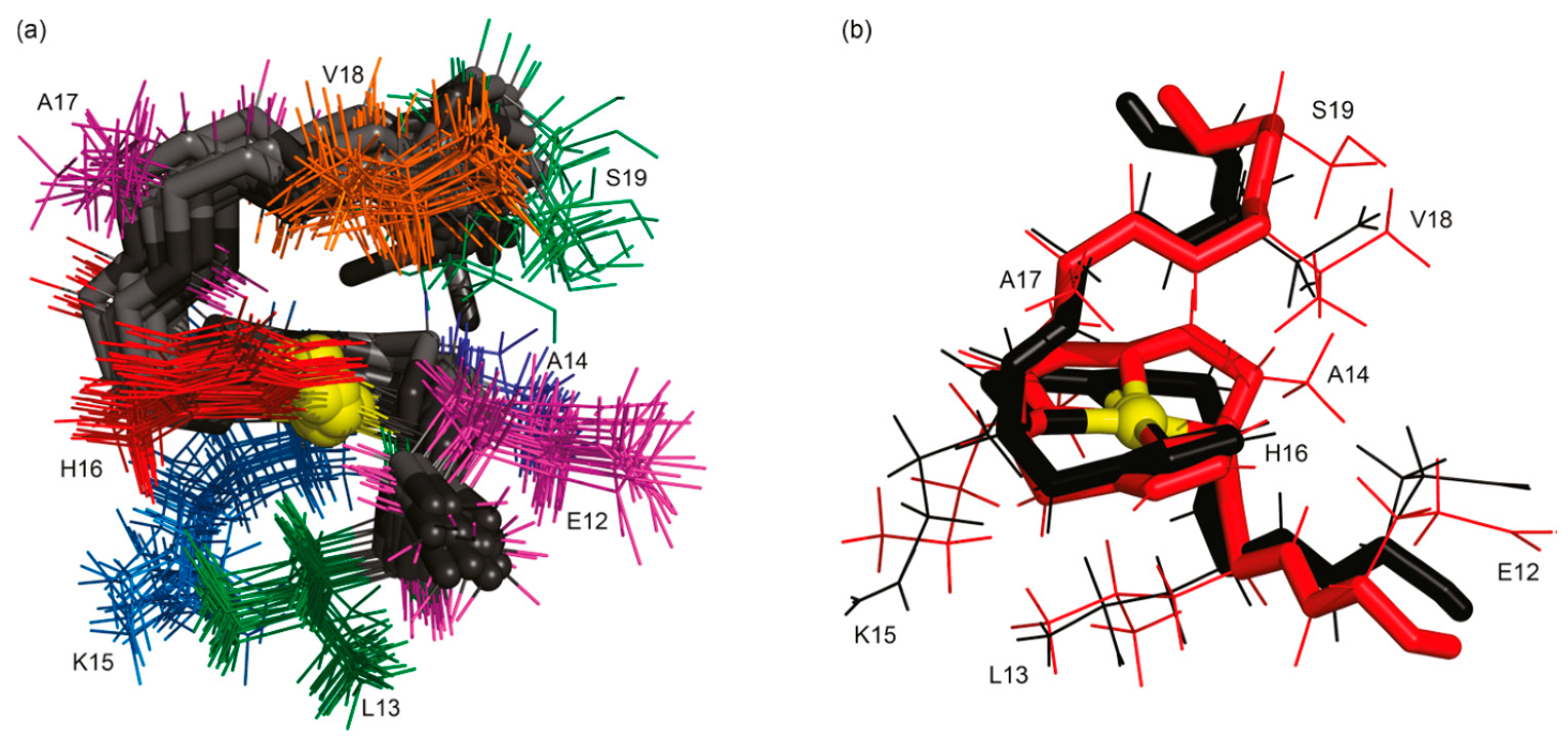
Figure 24.
The structure of Pd(II) complex reported by Mital et al. [51].
Figure 24.
The structure of Pd(II) complex reported by Mital et al. [51].
Figure 25.
(a) Stereoview of superposition of the 20 lowest energy structures of Ni2+-H2B105-112 obtained from NMR data [79].
Figure 25.
(a) Stereoview of superposition of the 20 lowest energy structures of Ni2+-H2B105-112 obtained from NMR data [79].
Gallium, a diamagnetic ion, has been shown to effectively substitute Fe(III) binding in Fe2S2 clusters while preserving the overall structure [81]. Palladium, also diamagnetic, has been utilized as a model for Cu(II) binding in amyloid beta-derived peptides and prion protein peptide derivatives due to its similar square-planar geometry [51,82]. Investigations into the pH dependence of palladium coordination using Pd(II)-peptide complexes have further expanded our understanding [83]. Tsiveriotis et al. presented the interaction of histidyl containing peptides with Pt(II) and Pd(II) [84].
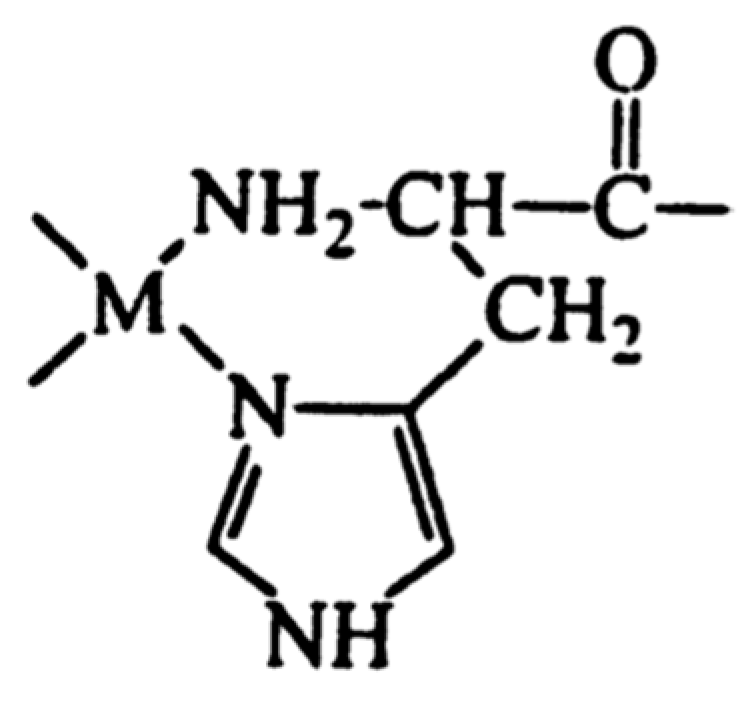
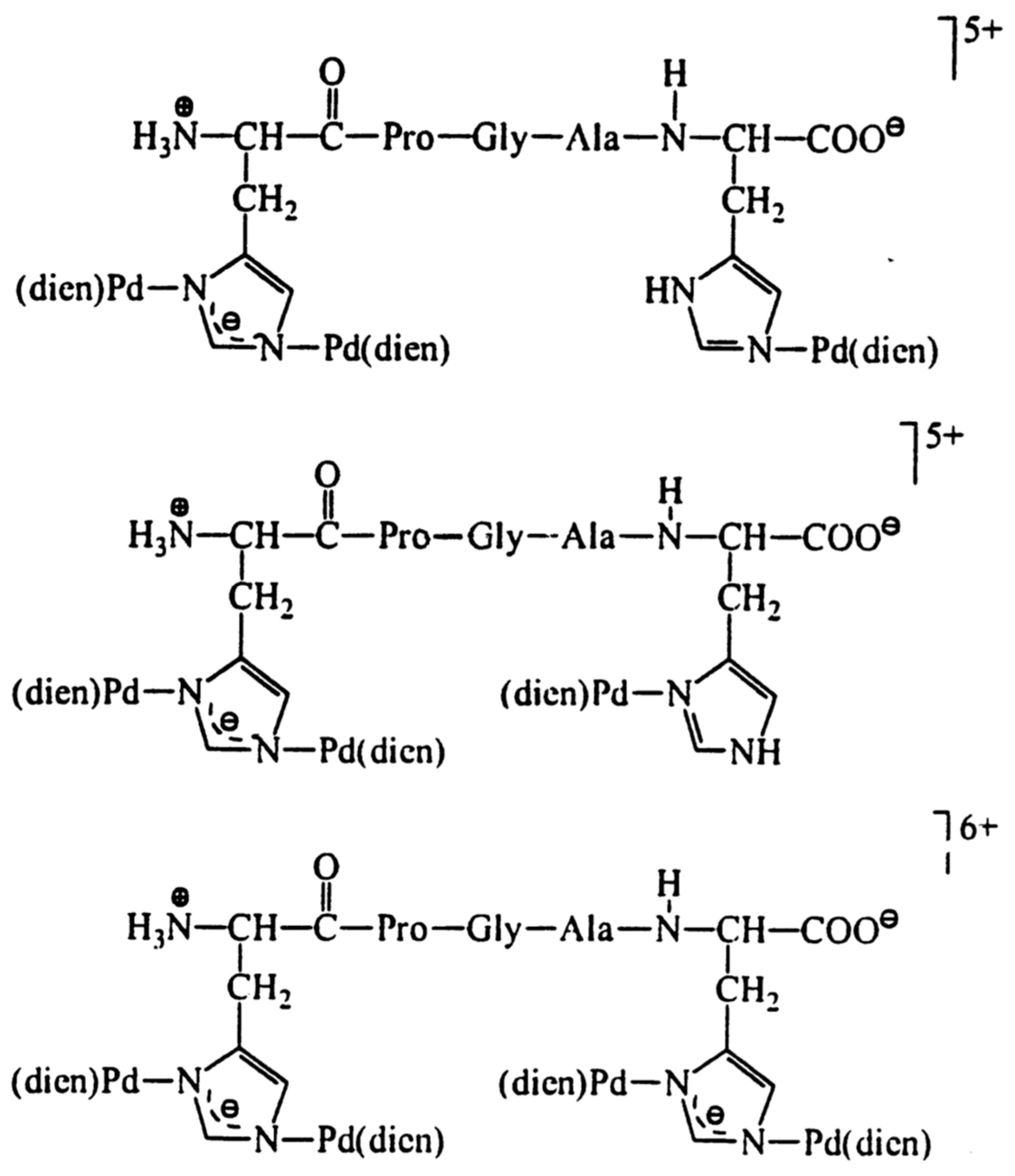
Figure 26.
The chelate structure of some metal complexes with Pt(II) and Pd(II) reported by Tsiveriotis et al. [84].
Figure 26.
The chelate structure of some metal complexes with Pt(II) and Pd(II) reported by Tsiveriotis et al. [84].
Figure 27.
The structure of some metal complexes with Pd(II) [84].
Figure 27.
The structure of some metal complexes with Pd(II) [84].
Silver, a toxic diamagnetic ion, interacts with the human copper transporter 1 (hCtr1). Structural studies involving an Ag(I) complex with a micelle-bound peptide derived from hCtr1 have suggested that the membrane surface may influence the structure of the extracellular domain of the protein and its binding to Ag(I) [85]. Similarly, studies on peptides derived from the human copper transporter 2 (hCtr2) have shown that Ag(I) binding occurs when the peptide is in its trimeric form [52].
Figure 28.
Ensemble of backbone atoms of 20 structures with the lowest target functions for the WT peptide in SDS micellar solution [85].
Figure 28.
Ensemble of backbone atoms of 20 structures with the lowest target functions for the WT peptide in SDS micellar solution [85].
Platinum, another diamagnetic metal with potent anticancer properties, has been studied for its binding with a peptide comprising the receptor binding sequence of transferrin, employing titration followed by NMR to determine the binding [86].
Lanthanides, mostly paramagnetic, are frequently used as shift reagents to monitor significant changes in chemical shift that occur in their proximity. Studies involving lanthanides and peptides have aimed at conjugating lanthanide-peptide complexes to proteins to leverage the shift reagent properties on the protein structure. For example, NMR-derived structures of fibrinopeptide B in the presence of salts, including Ga(III), have been elucidated [68]. Additionally, investigations into the binding properties of small peptides derived from Ca(II)-binding sites with toxic lanthanides such as La(III), Eu(III), and Tb(III) have been conducted to evaluate their potential for diagnostic use as contrast agents [87].
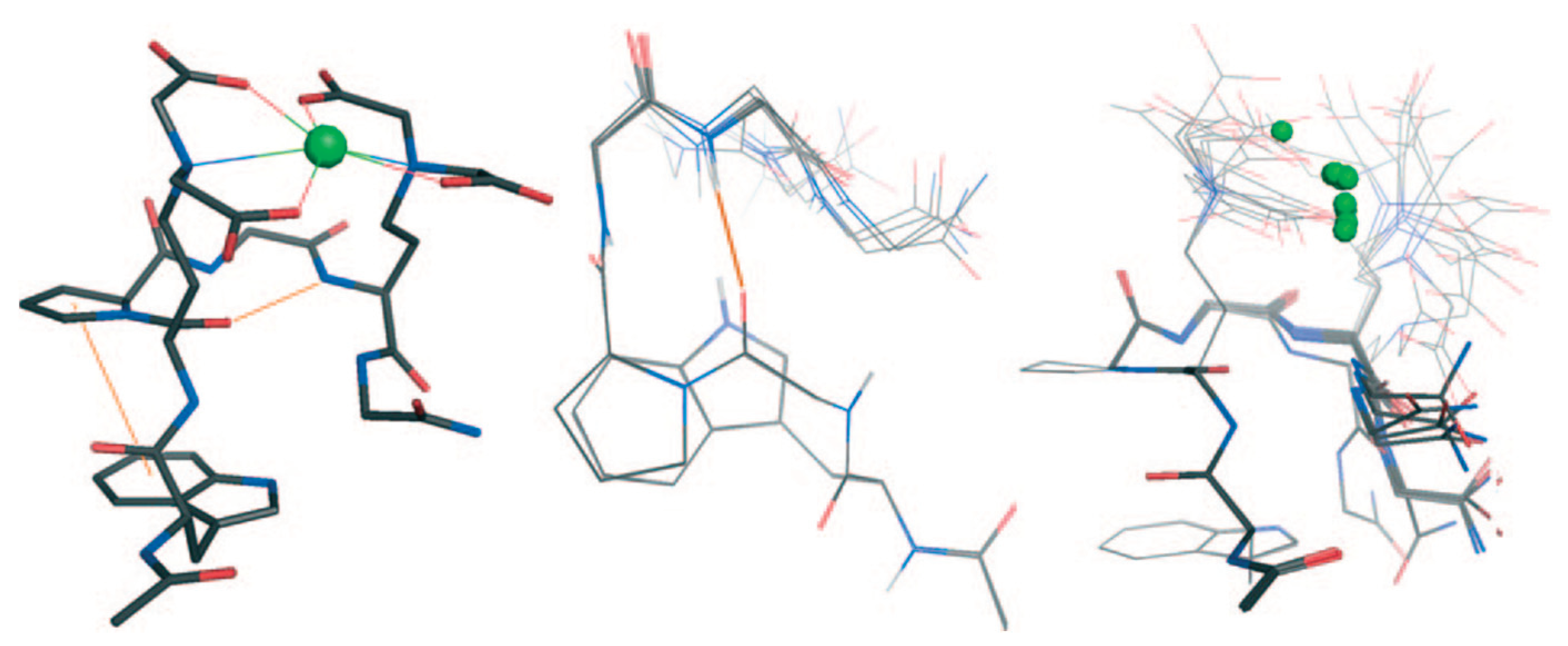
Figure 29.
The structure of La(III) complexes [87].
Figure 29.
The structure of La(III) complexes [87].

Table 2.
Summary table with some essential and non-essential elements.
| Essential elements | References | Non-essential elements | References |
|---|---|---|---|
| Cu(I) | 38, 41- 44 | Ni(II) | 77, 79, 80 |
| Cu(II) | 36, 45-50 | Pd(II) | 51, 82, 83, 84 |
| Zn(II) | 29, 34, 35, 59-65 | Pt(II) | 84 |
| Co(II) and Co(III) | 30, 31, 66-70 | Ag(I) | 52, 85 |
| Fe(II) and Fe(III) | 32, 33, 71-75 | Ga(III) | 68, 81 |
| La(III), Eu(III), Tb(III) | 87 | ||
Table 3.
Summary data on sturucture of the complexes and the donor atoms involved in the coordination.
Table 3.
Summary data on sturucture of the complexes and the donor atoms involved in the coordination.
| Technique | Donor atom | Metal | Structure | References |
|---|---|---|---|---|
| 1D 1H and 2D 1H−1H TOCSY NMR | S | Cu(I) | 38 | |
| COSY, TOCSY and NOESY, or ROESY | S | Cu(I) | 42 | |
| X-ray crystallography, elemental analysis, UV-Vis, 1H-, 13C-NMR, LC/MS | N, S | Cu(II) | 36 | |
| 1D 1H and 2D 1H−1H TOCSY and NOESY, HMQC, HSQC, 1H-15N 2D NMR | S | Cu(II) | 45 | |
| UV-Vis, ESI-MS, EPR, COSY, ROESY and TOCSY NMR | O, N | Cu(II) | square-planar or square-pyramidal geometries | 46 |
| UV-Vis, CD, ESR, NMR spectroscopic and MS methods | N | Cu(II) | 48 | |
| voltammetric (cyclic) and spectral (UV-Vis and fluorimetric) analytical techniques, IR, EPR | O, N, S for L1, L3, L6 and O, N for L2, L4 | Cu(II) | 50 | |
| UV-Vis, EPR, 1H–1H TOCSY, 1H–13C HSQC | N, O | Co(II) and Mn(II) | octahedral | 69 |
| 1H NMR, 1H-1H TOCSY | N, O | Co(III) | 67 | |
| 1D 1H and 2D NMR, CD and computational methods | N | Fe(II) and Co(II) | 73 | |
| UV-Vis, 1D 1H NMR spectra or 2D soft-COSY experiments | S | Zn(II) | 63 | |
| ESI-MS, TOCSY, NOESY, ROESY, 1H-13C HSQC and 1H-15N HSQC NMR | N | Pd(II) | square-planar | 51 |
| ESI–MS, TOCSY and ROESY, 1H-15N HSQC NMR | N | Al(III) | 76 | |
| UV-Vis, CD, ROESY, TOCSY, NOESY NMR | N | Ni(II) and Cu(II) | square-planar | 77 |
| 1H-, 13C- and 195Pt-NMR | N | Pd(II) | square-planar | 84 |
Conclusions
In summary, the essential metals play pivotal roles in various biological processes, ranging from oxygen transport to enzymatic catalysis. Their involvement in structural integrity, regulation, and signaling underscores their indispensability for life. While their deficiency can lead to debilitating disorders, excess levels can result in toxicity and disease. Through meticulous research spanning diverse methodologies, scientists have elucidated the intricate mechanisms underlying the interactions of essential metals with peptides and proteins, shedding light on their physiological functions and potential implications in diseases such as Alzheimer's. Furthermore, ongoing exploration of metal-peptide interactions holds promise for the development of novel therapeutic strategies and diagnostic tools. Thus, understanding the roles of essential metals in biological systems remains a cornerstone in advancing our knowledge of human health and disease.
In conclusion, the intricate interplay between non-essential metals and peptides underscores the multifaceted nature of their interactions and their potential implications in various biological processes and diseases, such as Alzheimer's disease and carcinogenesis. Through diverse methodologies ranging from NMR spectroscopy to computational modeling, researchers have elucidated the binding modes, structural changes, and functional consequences of these interactions. Investigations into metals like nickel, palladium, silver, and platinum have provided valuable insights into their roles in toxicity, carcinogenesis, and therapeutic potential. Furthermore, studies involving lanthanides have offered novel approaches for utilizing their paramagnetic properties as shift reagents for structural analysis. Continued exploration of these metal-peptide interactions promises to deepen our understanding of their biological significance and may pave the way for the development of novel diagnostic and therapeutic strategies in various fields of medicine and biotechnology.
Author Contributions
Conceptualization, P.E.M. and K.D.T.; writing—original draft preparation, P.E.M. and K.D.T.; writing—review and editing, P.E.M. and K.D.T.; project administration, S.T.; funding acquisition, P.E.M.
Acknowledgments
We acknowledge the financial support from the Fund for Scientific Research of the Plovdiv University, project СП 23-ХФ-006.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Akbarian, M.; Khani, A.; Eghbalpour, S.; Uversky, V.N. Bioactive Peptides: Synthesis, Sources, Applications, and Proposed Mechanisms of Action. Int. J. Mol. Sci. 2022, 23, 1445. [Google Scholar] [CrossRef]
- Lau, J.L.; Dunn, M.K. Therapeutic Peptides: Historical Perspectives, Current Development Trends, and Future Directions. Bioorganic Med. Chem. 2018, 26, 2700–2707. [Google Scholar] [CrossRef]
- Xie, M.; Liu, D.; Yang, Y. Anti-Cancer Peptides: Classification, Mechanism of Action, Reconstruction and Modification. Open Biol. 2020, 10, 200004. [Google Scholar] [CrossRef] [PubMed]
- Huan, Y.; Kong, Q.; Mou, H.; Yi, H. Antimicrobial Peptides: Classification, Design, Application and Research Progress in Multiple Fields. Front. Microbiol. 2020, 11, 582779. [Google Scholar] [CrossRef]
- Gao, B.; Zhao, D.; Li, L.; Cheng, Z.; Guo, Y. Antiviral Peptides with in Vivo Activity: Development and Modes of Action. Chem Plus Chem 2021, 86, 1547–1558. [Google Scholar] [CrossRef] [PubMed]
- Baig, M.H.; Ahmad, K.; Saeed, M.; Alharbi, A.M.; Barreto, G.E.; Ashraf, G.M.; Choi, I. Peptide Based Therapeutics and Their Use for the Treatment of Neurodegenerative and Other Diseases. Biomed. Pharmacother. 2018, 103, 574–581. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Fu, L.; Hu, Z.; Zhong, Y. A Mini-Review on Peptide-Based Self-Assemblies and Their Biological Applications. Nanotechnology 2021, 33, 062004. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wang, Y.; Rodriguez, B.J.; Yang, R.; Yu, B.; Mei, D.; Li, J.; Tao, K.; Gazit, E. Microfabrication of Peptide Self-Assemblies:Inspired by Nature towards Applications. Chem. Soc. Rev. 2022, 51, 6936–6947. [Google Scholar] [CrossRef] [PubMed]
- Fu, K.; Wu, H.; Su, Z. Self-Assembling Peptide-Based Hydrogels: Fabrication, Properties, and Applications. Biotechnol. Adv. 2021, 49, 107752. [Google Scholar] [CrossRef]
- Zeng, X.-Z.; An, H.-W.; Wang, H. Chemical Reactions Trigger Peptide Self-Assembly in Vivo for Tumor Therapy. Chem. Med.Chem. 2021, 16, 2452–2458. [Google Scholar] [CrossRef]
- Zhang, Z.; Ai, S.; Yang, Z.; Li, X. Peptide-Based Supramolecular Hydrogels for Local Drug Delivery. Adv. Drug Deliv. Rev. 2021, 174, 482–503. [Google Scholar] [CrossRef] [PubMed]
- Curtis, R.W.; Chmielewski, J. A Comparison of the Collagen Triple Helix and Coiled-Coil Peptide Building Blocks on Metal Ion-Mediated Supramolecular Assembly. Pept. Sci. 2021, 113, e24190. [Google Scholar] [CrossRef]
- Sontz, P.A.; Song, W.J.; Tezcan, F.A. Interfacial Metal Coordination in Engineered Protein and Peptide Assemblies. Curr. Opin. Chem. Biol. 2014, 19, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.; Liu, Y.; Cui, Y. Artificial Metal–Peptide Assemblies: Bioinspired Assembly of Peptides and Metals through Space and across Length Scales. J. Am. Chem. Soc. 2021, 143, 17316–17336. [Google Scholar] [CrossRef] [PubMed]
- Puiu, M.; Bala, C. Peptide-Based Biosensors: From Self-Assembled Interfaces to Molecular Probes in Electrochemical Assays. Bioelectrochemistry 2018, 120, 66–75. [Google Scholar] [CrossRef] [PubMed]
- Joshi, P.N.; Mervinetsky, E.; Solomon, O.; Chen, Y.-J.; Yitzchaik, S.; Friedler, A. Electrochemical Biosensors Based on Peptide-Kinase Interactions at the Kinase Docking Site. Biosens. Bioelectron. 2022, 207, 114177. [Google Scholar] [CrossRef] [PubMed]
- Synhaivska, O.; Mermoud, Y.; Baghernejad, M.; Alshanski, I.; Hurevich, M.; Yitzchaik, S.; Wipf, M.; Calame, M. Detection of Cu2+ Ions with GGH Peptide Realized with Si-Nanoribbon ISFET. Sensors 2019, 19, 4022. [Google Scholar] [CrossRef] [PubMed]
- Mervinetsky, E.; Alshanski, I.; Tadi, K.K.; Dianat, A.; Buchwald, J.; Gutierrez, R.; Cuniberti, G.; Hurevich, M.; Yitzchaik, S. A Zinc Selective Oxytocin Based Biosensor. J. Mater. Chem. B 2019, 8, 155–160. [Google Scholar] [CrossRef]
- Mannoor, M.S.; Zhang, S.; Link, A.J.; McAlpine, M.C. Electrical Detection of Pathogenic Bacteria via Immobilized Antimicrobial Peptides. Proc. Natl. Acad. Sci. USA 2010, 107, 19207–19212. [Google Scholar] [CrossRef]
- Lillehoj, P.B.; Kaplan, C.W.; He, J.; Shi, W.; Ho, C.-M. Rapid, Electrical Impedance Detection of Bacterial Pathogens Using Immobilized Antimicrobial Peptides. SLAS Technol. 2014, 19, 42–49. [Google Scholar] [CrossRef]
- Caetano-Silva, M.E.; Netto, F.M.; Bertoldo-Pacheco, M.T.; Alegría, A.; Cilla, A. Peptide-Metal Complexes: Obtention and Role in Increasing Bioavailability and Decreasing the pro-Oxidant Effect of Minerals. Crit. Rev. Food Sci. Nutr. 2021, 61, 1470–1489. [Google Scholar] [CrossRef] [PubMed]
- Tian, Q.; Fan, Y.; Hao, L.; Wang, J.; Xia, C.; Wang, J.; Hou, H. A Comprehensive Review of Calcium and Ferrous Ions Chelating Peptides: Preparation, Structure and Transport Pathways. Crit. Rev. Food Sci. Nutr. 2021, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Van de Looij, S.M.; Hebels, E.R.; Viola, M.; Hembury, M.; Oliveira, S.; Vermonden, T. Gold Nanoclusters: Imaging, Therapy, and Theranostic Roles in Biomedical Applications. Bioconjugate Chem. 2022, 33, 4–23. [Google Scholar] [CrossRef] [PubMed]
- Webster, A.M.; Peacock, A.F.A. De Novo Designed Coiled Coils as Scaffolds for Lanthanides, Including Novel Imaging Agents with a Twist. Chem. Commun. 2021, 57, 6851–6862. [Google Scholar] [CrossRef] [PubMed]
- Balzano, S.; Sardo, A.; Blasio, M.; Chahine, T.B.; Dell’Anno, F.; Sansone, C.; Brunet, C. Microalgal Metallothioneins and Phytochelatins and Their Potential Use in Bioremediation. Front. Microbiol. 2020, 11, 517. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Selvamani, V.; Yoo, I.-K.; Kim, T.W.; Hong, S.H. A Novel Strategy for the Microbial Removal of Heavy Metals: Cell-Surface Display of Peptides. Biotechnol. Bioprocess Eng. 2021, 26, 1–9. [Google Scholar] [CrossRef]
- Gomes, L.M.F.; Bataglioli, J.C.; Storr, T. Metal Complexes That Bind to the Amyloid-_ Peptide of Relevance to Alzheimer’s Disease. Coord. Chem. Rev. 2020, 412, 213255. [Google Scholar] [CrossRef]
- Shalev, D.E. Studying Peptide-Metal Ion Complex Structures by Solution-State NMR. Int. J. Mol. Sci. 2022, 23, 15957. [Google Scholar] [CrossRef] [PubMed]
- Maret, W. Zinc in Cellular Regulation: The Nature and Significance of “Zinc Signals”. Int. J. Mol. Sci. 2017, 18, 2285. [Google Scholar] [CrossRef]
- Nitsche, C.; Otting, G. Pseudocontact Shifts in Biomolecular NMR Using Paramagnetic Metal Tags. Prog. Nucl. Magn. Reson. Spectrosc. 2017, 98–99, 20–49. [Google Scholar] [CrossRef]
- Renfrew, A.K.; O’Neill, E.S.; Hambley, T.W.; New, E.J. Harnessing the Properties of Cobalt Coordination Complexes for Biological Application. Coord. Chem. Rev. 2018, 375, 221–233. [Google Scholar] [CrossRef]
- Aisen, P.; Enns, C.; Wessling-Resnick, M. Chemistry and Biology of Eukaryotic Iron Metabolism. Int. J. Biochem. Cell Biol. 2001, 33, 940–959. [Google Scholar] [CrossRef]
- Vogt, A.-C.S.; Arsiwala, T.; Mohsen, M.; Vogel, M.; Manolova, V.; Bachmann, M.F. On Iron Metabolism and Its Regulation. Int. J. Mol. Sci. 2021, 22, 4591. [Google Scholar] [CrossRef]
- Istrate, A.N.; Tsvetkov, P.O.; Mantsyzov, A.B.; Kulikova, A.A.; Kozin, S.A.; Makarov, A.A.; Polshakov, V.I. NMR Solution Structure of Rat A Beta(1-16): Toward Understanding the Mechanism of Rats’ Resistance to Alzheimer’s Disease. Biophys. J. 2012, 102, 136–143. [Google Scholar] [CrossRef] [PubMed]
- Polshakov, V.I.; Mantsyzov, A.B.; Kozin, S.A.; Adzhubei, A.A.; Zhokhov, S.S.; van Beek, W.; Kulikova, A.A.; Indeykina, M.I.; Mitkevich, V.A.; Makarov, A.A. A Binuclear Zinc Interaction Fold Discovered in the Homodimer of Alzheimer’s Amyloid-β Fragment with Taiwanese Mutation D7H. Angew. Chem. Int. Ed. 2017, 56, 11734–11739. [Google Scholar] [CrossRef] [PubMed]
- Hickey, J.L.; Lim, S.; Hayne, D.J.; Paterson, B.M.; White, J.M.; Villemagne, V.L.; Roselt, P.; Binns, D.; Cullinane, C.; Jeffery, C.M.; Price, R.I.; Barnham, K.J.; Donnelly, P.S. Diag-nostic Imaging Agents for Alzheimer’s Disease: Copper Radiopharmaceuticals that Target Aβ Plaques. J. Am. Chem. Soc. 2013, 135, 16120–16132. [Google Scholar] [CrossRef]
- Lo, C.; Bandmann, O. Chapter 2—Epidemiology and Introduction to the Clinical Presentation ofWilson Disease. In Handbook of Clinical Neurology; Członkowska, A., Schilsky, M.L., Eds.; Wilson Disease; Elsevier: Amsterdam, The Netherlands, 2017; Volume 142, pp. 7–17. [Google Scholar]
- Miotto, M.C.; Rodriguez, E.E.; Valiente-Gabioud, A.A.; Torres-Monserrat, V.; Binolfi, A.; Quintanar, L.; Zweckstetter, M.; Griesinger, C.; Fernández, C.O. Site-Specific Copper-Catalyzed Oxidation of α-Synuclein: Tightening the Link between Metal Binding and Protein Oxidative Damage in Parkinson’s Disease. Inorg. Chem. 2014, 53, 4350–4358. [Google Scholar] [CrossRef] [PubMed]
- Remelli, M.; Peana, M.; Medici, S.; Ostrowska, M.; Gumienna-Kontecka, E.; Zoroddu, M.A. Manganism and Parkinson’s Disease: Mn(II) and Zn(II) Interaction with a 30-Amino Acid Fragment. Dalton Trans. 2016, 45, 5151–5161. [Google Scholar] [CrossRef] [PubMed]
- Kaler, S.G. Menkes Disease. In Encyclopedia of the Neurological Sciences, 2nd ed.; Aminoff, M.J., Daroff, R.B., Eds.; Academic Press: Oxford, UK, 2014; pp. 1082–1089. ISBN 978-0-12-385158-1. [Google Scholar]
- Festa, R.A.; Thiele, D.J. Copper: An Essential Metal in Biology. Curr Biol 2011, 21, R877–R883. [Google Scholar] [CrossRef]
- Shoshan, M.S.; Tshuva, E.Y.; Shalev, D.E. Structure and Coordination Determination of Peptide-Metal Complexes Using 1D and 2D H-1 NMR. J. Vis. Exp. 2013, 82, e50747. [Google Scholar] [CrossRef]
- Shoshan, M.S.; Dekel, N.; Goch, W.; Shalev, D.E.; Danieli, T.; Lebendiker, M.; Bal, W.; Tshuva, E.Y. Unbound Position II in MXCXXC Metallochaperone Model Peptides Impacts Metal Binding Mode and Reactivity: Distinct Similarities to Whole Proteins. J. Inorg. Biochem. 2016, 159, 29–36. [Google Scholar] [CrossRef] [PubMed]
- De Gregorio, G.; Biasotto, F.; Hecel, A.; Luczkowski, M.; Kozlowski, H.; Valensin, D. Structural Analysis of Copper(I) Interaction with Amyloid _ Peptide. J. Inorg. Biochem. 2019, 195, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Bertini, I.; Ciurli, S.; Dikiy, A.; Fernàndez, C.O.; Luchinat, C.; Safarov, N.; Shumilin, S.; Vila, A.J. The First Solution Structure of a Paramagnetic Copper(II) Protein: The Case of Oxidized Plastocyanin from the Cyanobacterium Synechocystis PCC6803. J. Am. Chem. Soc. 2001, 123, 2405–2413. [Google Scholar] [CrossRef] [PubMed]
- Fragoso, A.; Carvalho, T.; Rousselot-Pailley, P.; Correia dos Santos, M.M.; Delgado, R.; Iranzo, O. Effect of the Peptidic Scaffold in Copper(II) Coordination and the Redox Properties of Short Histidine-Containing Peptides. Chem.—Eur. J. 2015, 21, 13100–13111. [Google Scholar] [CrossRef] [PubMed]
- Alshanski, I.; Shalev, D.E.; Yitzchaik, S.; Hurevich, M. Determining the Structure and Binding Mechanism of Oxytocin-Cu2+ Complex Using Paramagnetic Relaxation Enhancement NMR Analysis. J. Biol. Inorg. Chem. 2021, 26, 809–815. [Google Scholar] [CrossRef] [PubMed]
- Dancs, Á.; Selmeczi, K.; May, N.V.; Gajda, T. On the Copper(II) Binding of Asymmetrically Functionalized Tripodal Peptides: Solution Equilibrium, Structure, and Enzyme Mimicking. New J. Chem. 2018, 42, 7746–7757. [Google Scholar] [CrossRef]
- Kamysz, E.; Kotynia, A.; Czyznikowska, ˙ Z.; Jaremko, M.; Jaremko, Ł.; Nowakowski, M.; Brasun, J. Sialorphin and Its Analog as ˙ Ligands for Copper(II) Ions. Polyhedron 2013, 55, 216–224. [Google Scholar] [CrossRef]
- Georgieva, S.; Todorov, P.; Staneva, D.; Grozdanov, P.; Nikolova, I.; Grabchev, I. Met-al–Peptide Complexes with Antimicrobial Potential for Cotton Fiber Protection, J. Funct. Biomater. 2023, 14, 106. [Google Scholar] [CrossRef]
- Mital, M.; Szutkowski, K.; Bossak-Ahmad, K.; Skrobecki, P.; Drew, S.C.; Poznanski, J.; Zhukov, I.; Fraczyk, T.; Bal, W. The Palladium(II) Complex of A Beta(4-16) as Suitable Model for Structural Studies of Biorelevant Copper(II) Complexes of NTruncated Beta-Amyloids. Int. J. Mol. Sci. 2020, 21, 9200. [Google Scholar] [CrossRef]
- Dong, Z.; Guan, L.; Wang, C.; Xu, H.; Li, Z.; Li, F. Reconstruction of a Helical Trimer by the Second Transmembrane Domain of Human Copper Transporter 2 in Micelles and the Binding of the Trimer to Silver. RSC Adv. 2016, 6, 4335–4342. [Google Scholar] [CrossRef]
- Remelli, M.; Valensin, D.; Toso, L.; Gralka, E.; Guerrini, R.; Marzola, E.; Kozłowski, H. Thermodynamic and Spectroscopic Investigation on the Role of Met Residues in CuII Binding to the Non-Octarepeat Site of the Human Prion Protein. Metallomics 2012, 4, 794–806. [Google Scholar] [CrossRef] [PubMed]
- Abbas, I.M.; Vranic, M.; Hoffmann, H.; El-Khatib, A.H.; Montes-Bayón, M.; Möller, H.M.; Weller, M.G. Investigations of the Copper Peptide Hepcidin-25 by LC-MS/MS and NMR. Int. J. Mol. Sci. 2018, 19, 2271. [Google Scholar] [CrossRef] [PubMed]
- Mena, S.; Mirats, A.; Caballero, A.B.; Guirado, G.; Barrios, L.A.; Teat, S.J.; Rodriguez-Santiago, L.; Sodupe, M.; Gamez, P. Drastic Effect of the Peptide Sequence on the Copper-Binding Properties of Tripeptides and the Electrochemical Behaviour of Their Copper(II) Complexes. Chem.—Eur. J. 2018, 24, 5153–5162. [Google Scholar] [CrossRef] [PubMed]
- Klose, D.; Vemulapalli, S.P.B.; Richman, M.; Rudnick, S.; Aisha, V.; Abayev, M.; Chemerovski, M.; Shviro, M.; Zitoun, D.; Majer, K.; et al. Cu2+-Induced Self-Assembly and Amyloid Formation of a Cyclic D,L-α-Peptide: Structure and Function. Phys. Chem. Chem. Phys. 2022, 24, 6699–6715. [Google Scholar] [CrossRef] [PubMed]
- Felten, A.-S.; Pellegrini-Moïse, N.; Selmeczi, K.; Henry, B.; Chapleur, Y. Synthesis and Copper(II)-Complexation Properties of an Unusual Macrocyclic Structure Containing α/β-Amino Acids and Anomeric Sugar β-Amino Acid. Eur. J. Org. Chem. 2013, 2013, 5645–5654. [Google Scholar] [CrossRef]
- Sánchez-López, C.; Cortés-Mejía, R.; Miotto, M.C.; Binolfi, A.; Fernández, C.O.; del Campo, J.M.; Quintanar, L. Copper Coordination Features of Human Islet Amyloid Polypeptide: The Type 2 Diabetes Peptide. Inorg. Chem. 2016, 55, 10727–10740. [Google Scholar] [CrossRef] [PubMed]
- Andreini, C.; Banci, L.; Bertini, I.; Rosato, A. Counting the Zinc-Proteins Encoded in the Human Genome. J. Proteome Res. 2006, 5, 196–201. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, K.; Nakatani, A.; Saito, K. The Unique N-Terminal Zinc Finger of Synaptotagmin-like Protein 4 Reveals FYVE Structure. Protein Sci. 2017, 26, 2451–2457. [Google Scholar] [CrossRef]
- Miyamoto, K.; Uechi, A.; Saito, K. The Zinc Finger Domain of RING Finger Protein 141 Reveals a Unique RING Fold. Protein Sci. 2017, 26, 1681–1686. [Google Scholar] [CrossRef]
- Lombardo, C.M.; Kumar, M.V.V.; Douat, C.; Rosu, F.; Mergny, J.-L.; Salgado, G.F.; Guichard, G. Design and Structure Determination of a Composite Zinc Finger Containing a Nonpeptide Foldamer Helical Domain. J. Am. Chem. Soc. 2019, 141, 2516–2525. [Google Scholar] [CrossRef]
- Jacques, A.; Mettra, B.; Lebrun, V.; Latour, J.-M.; Sénèque, O. On the Design of Zinc-Finger Models with Cyclic Peptides Bearing a Linear Tail. Chem.—Eur. J. 2013, 19, 3921–3931. [Google Scholar] [CrossRef]
- Rao, S.R.; Horne, W.S. Proteomimetic Zinc Finger Domains with Modified Metal-Binding Beta-Turns. Pept. Sci. 2020, 112, e24177. [Google Scholar] [CrossRef]
- Pham, T.L.; Kovermann, M.; Thomas, F. Switchable Zinc(II)-Responsive Globular β-Sheet Peptide. ACS Synth. Biol. 2022, 11, 254–264. [Google Scholar] [CrossRef] [PubMed]
- Lombardi, A.; Nastri, F.; Marasco, D.; Maglio, O.; De Sanctis, G.; Sinibaldi, F.; Santucci, R.; Coletta, M.; Pavone, V. Design of a New Mimochrome with Unique Topology. Chem.—Eur. J. 2003, 9, 5643–5654. [Google Scholar] [CrossRef] [PubMed]
- Heffern, M.C.; Velasco, P.T.; Matosziuk, L.M.; Coomes, J.L.; Karras, C.; Ratner, M.A.; Klein, W.L.; Eckermann, A.L.; Meade, T.J. Modulation of Amyloid-β Aggregation by Histidine-Coordinating Cobalt(III) Schiff Base Complexes. Chem.Bio.Chem. 2014, 15, 1584–1589. [Google Scholar] [CrossRef]
- Faizullina, A.R.; Blokhin, D.S.; Kusova, A.M.; Klochkov, V.V. Investigation of the Effect of Transition Metals (MN, CO, GD) on the Spatial Structure of Fibrinopeptide B by NMR Spectroscopy. J. Mol. Struct. 2020, 1204, 127484. [Google Scholar] [CrossRef]
- Peana, M.; Medici, S.; Nurchi, V.M.; Crisponi, G.; Lachowicz, J.I.; Zoroddu, M.A. Manganese and Cobalt Binding in a MultiHistidinic Fragment. Dalton Trans. 2013, 42, 16293–16301. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Bang, K.-M.; Ha, H.; Cho, N.H.; Namgung, S.D.; Im, S.W.; Cho, K.H.; Kim, R.M.; Choi, W.I.; Lim, Y.-C.; et al. Tyrosyltyrosylcysteine-Directed Synthesis of Chiral Cobalt Oxide Nanoparticles and Peptide Conformation Analysis. ACS Nano 2021, 15, 979–988. [Google Scholar] [CrossRef] [PubMed]
- Vazquez, D.S.; Agudelo, W.A.; Yone, A.; Vizioli, N.; Arán, M.; Flecha, F.L.G.; Lebrero, M.C.G.; Santos, J. A Helix–Coil Transition Induced by the Metal Ion Interaction with a Grafted Iron-Binding Site of the CyaY Protein Family. Dalton Trans. 2015, 44, 2370–2379. [Google Scholar] [CrossRef]
- Nudelman, H.; Lee, Y.-Z.; Hung, Y.-L.; Kolusheva, S.; Upcher, A.; Chen, Y.-C.; Chen, J.-Y.; Sue, S.-C.; Zarivach, R. Understanding the Biomineralization Role of Magnetite-Interacting Components (MICs) From Magnetotactic Bacteria. Front. Microbiol. 2018, 9, 2480. [Google Scholar] [CrossRef]
- Gómez-González, J.; Pérez, Y.; Sciortino, G.; Roldan-Martín, L.; Martínez-Costas, J.; Maréchal, J.-D.; Alfonso, I.; Vázquez López, M.; Vázquez, M.E. Dynamic Stereoselection of Peptide Helicates and Their Selective Labeling of DNA Replication Foci in Cells. Angew. Chem. Int. Ed. 2021, 60, 8859–8866. [Google Scholar] [CrossRef] [PubMed]
- Inubushi, T.; Becker, E.D. Efficient Detection of Paramagnetically Shifted NMR Resonances by Optimizing the WEFT Pulse Sequence. J. Magn. Reson. (1969) 1983, 51, 128–133. [Google Scholar] [CrossRef]
- Zhong, F.; Pletneva, E.V. Ligation and Reactivity of Methionine-Oxidized Cytochrome c. Inorg. Chem. 2018, 57, 5754–5766. [Google Scholar] [CrossRef] [PubMed]
- Narayan, P.; Krishnarjuna, B.; Vishwanathan, V.; Jagadeesh Kumar, D.; Babu, S.; Ramanathan, K.V.; Easwaran, K.R.K.; Nagendra, H.G.; Raghothama, S. Does Aluminium Bind to Histidine? An NMR Investigation of Amyloid B12 and Amyloid B16 Fragments. Chem. Biol. Drug Des. 2013, 82, 48–59. [Google Scholar] [CrossRef] [PubMed]
- Jones, C.E.; Klewpatinond, M.; Abdelraheim, S.R.; Brown, D.R.; Viles, J.H. Probing Copper2+ Binding to the Prion Protein Using Diamagnetic Nickel2+ and 1H NMR: The Unstructured N Terminus Facilitates the Coordination of Six Copper2+ Ions at Physiological Concentrations. J. Mol. Biol. 2005, 346, 1393–1407. [Google Scholar] [CrossRef] [PubMed]
- Gasmi, G.; Singer, A.; Forman-Kay, J.; Sarkar, B. NMR Structure of Neuromedin C, a Neurotransmitter with an Amino Terminal CuII-, NiII-Binding (ATCUN) Motif. J. Pept. Res. 1997, 49, 500–509. [Google Scholar] [CrossRef] [PubMed]
- Nunes, A.M.; Zavitsanos, K.; Del Conte, R.; Malandrinos, G.; Hadjiliadis, N. The Possible Role of 94−125 Peptide Fragment of Histone H2B in Nickel-Induced Carcinogenesis. Inorg. Chem. 2010, 49, 5658–5668. [Google Scholar] [CrossRef] [PubMed]
- Zoroddu, M.A.; Peana, M.; Medici, S.; Potocki, S.; Kozlowski, H. Ni(II) Binding to the 429–460 Peptide Fragment from Human Toll like Receptor (HTLR4): A Crucial Role for Nickel-Induced Contact Allergy? Dalton Trans. 2014, 43, 2764–2771. [Google Scholar] [CrossRef] [PubMed]
- Pochapsky, T.C.; Kuti, M.; Kazanis, S. The Solution Structure of a Gallium-Substituted Putidaredoxin Mutant: GaPdx C85S. J. Biomol. NMR 1998, 12, 407–415. [Google Scholar] [CrossRef]
- Garnett, A.P.; Jones, C.E.; Viles, J.H. A Survey of Diamagnetic Probes for Copper2+ Binding to the Prion Protein. 1H NMR Solution Structure of the Palladium2+ Bound Single Octarepeat. Dalton Trans. 2006, 509–518. [Google Scholar] [CrossRef]
- Monger, L.J.; Runarsdottir, G.R.; Suman, S.G. Directed Coordination Study of [Pd(En)(H2O)2 ]2+ with Hetero-Tripeptides Containing C-Terminus Methyl Esters Employing NMR Spectroscopy. J. Biol. Inorg. Chem. 2020, 25, 811–825. [Google Scholar] [CrossRef] [PubMed]
- Tsiveriotis, P.; Hadjiliadis, N. Studies on the Interaction of Histidyl Containing Peptides with Palladium(II) and Platinum(II) Complex Ions. Coord. Chem. Rev. 1999, 190–192, 171–184. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, L.; Li, F. Micelle-Bound Structure of an Extracellular Met-Rich Domain of HCtr1 and Its Binding with Silver. RSC Adv. 2013, 3, 15245–15253. [Google Scholar] [CrossRef]
- Teles, C.M.; Antunes, V.U.; Cardoso, R.S.; Candido, T.Z.; Lima, C.S.P.; Ruiz, A.L.T.G.; Juliano, M.A.; Favaro, D.C.; Abbehausen, C. Functionalization of New Anticancer Pt(II) Complex with Transferrin Receptor Binding Peptide. Inorg. Chim. Acta 2020, 511, 119811. [Google Scholar] [CrossRef]
- Cisnetti, F.; Gateau, C.; Lebrun, C.; Delangle, P. Lanthanide(III) Complexes with Two Hexapeptides Incorporating Unnatural Chelating Amino Acids: Secondary Structure and Stability. Chem.—Eur. J. 2009, 15, 7456–7469. [Google Scholar] [CrossRef]
Figure 1.
Different applications of bioactive peptides for humans.
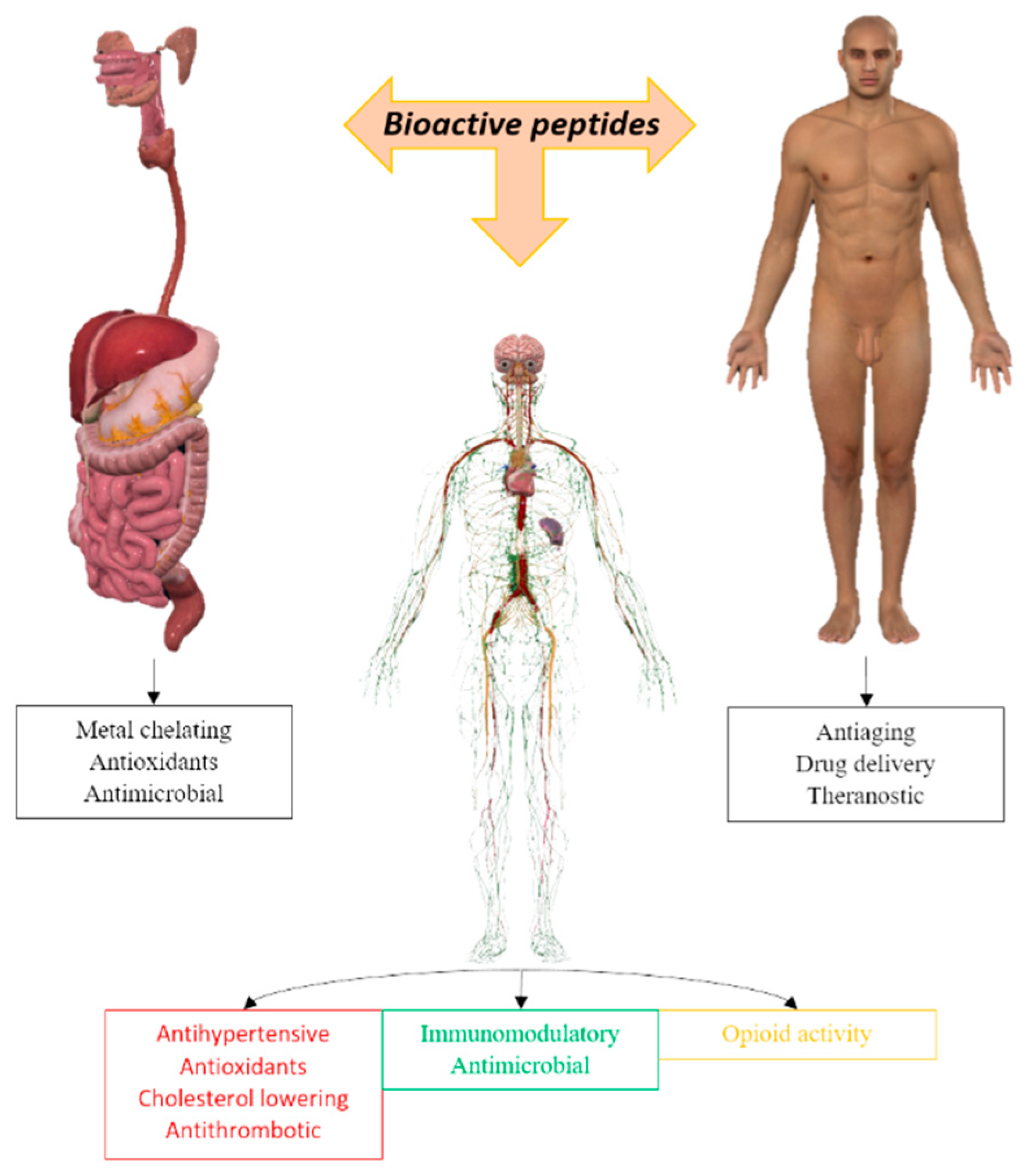
Figure 2.
Classification of antimicrobial peptides.
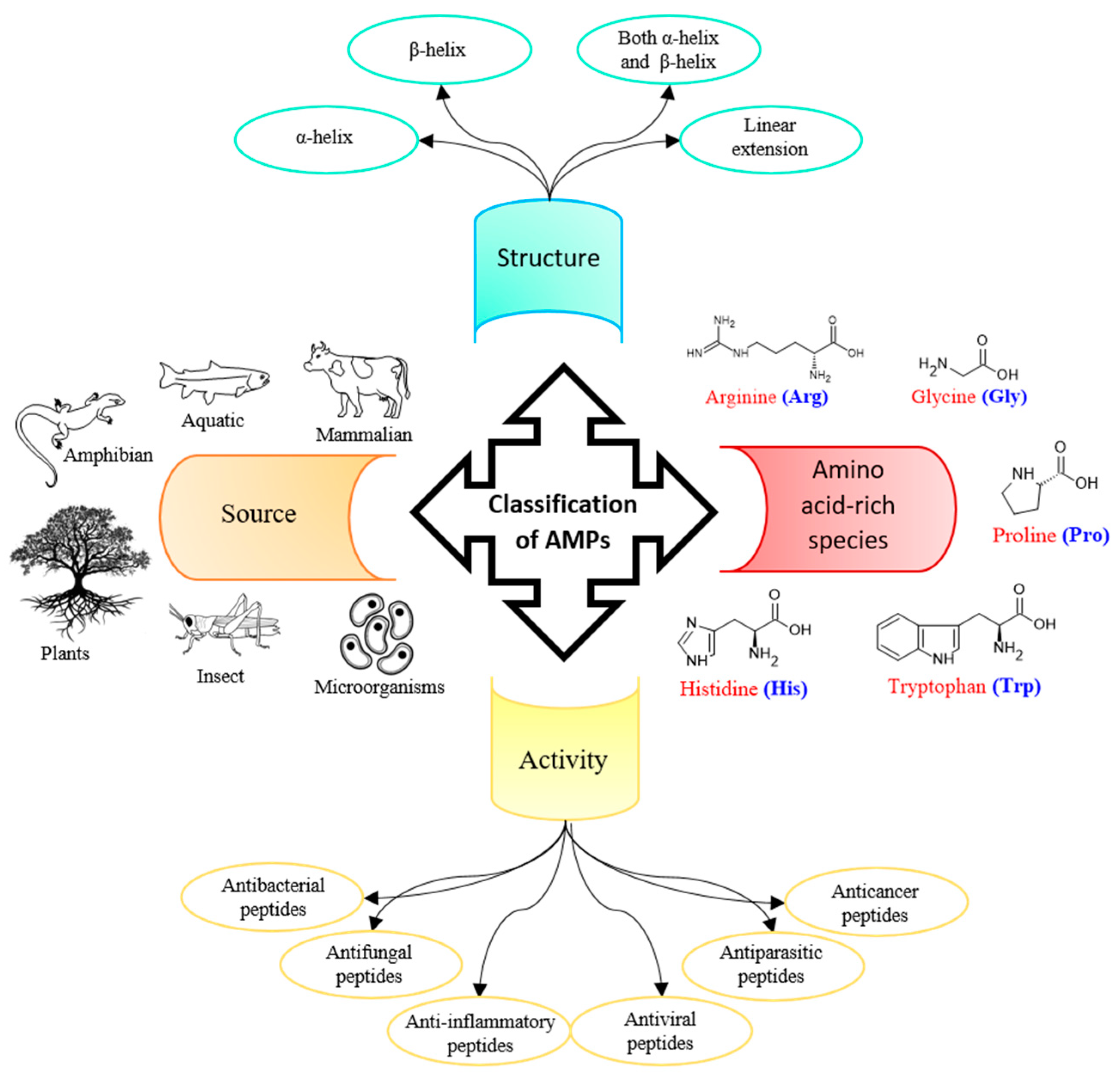
Figure 3.
Metal complexes with peptides.
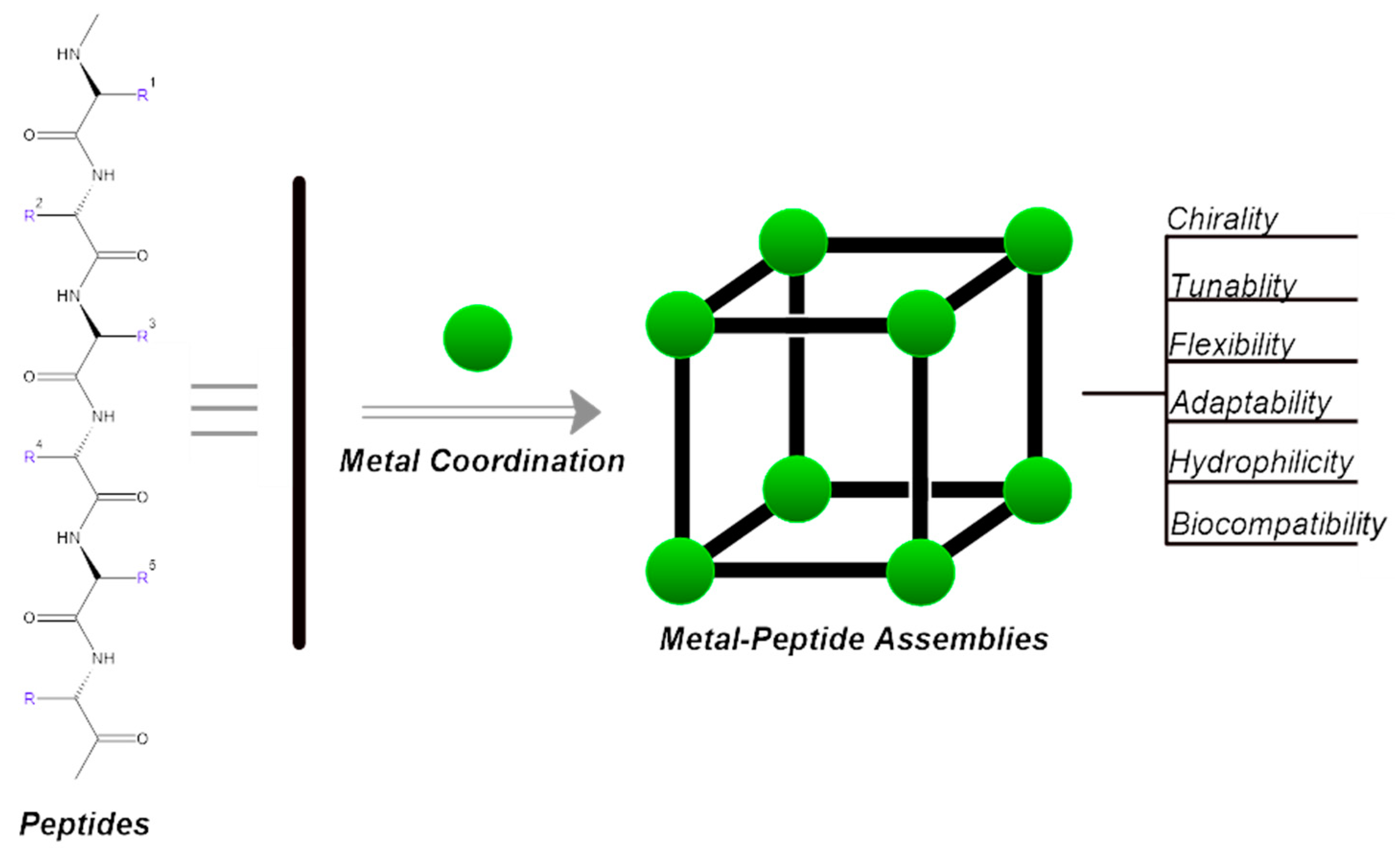
Table 1.
Some diseases and articles related to them.
| Disease | References |
|---|---|
| Alzheimer’s | 27, 34, 35, 36 |
| Wilson's | 37 |
| Parkinson’s | 38, 39 |
| Menkes | 40 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



