
Submitted:
19 February 2024
Posted:
19 February 2024
You are already at the latest version
Abstract
B4GALNT2 is a vital candidate gene that affects the growth traits litter size in and growth traits of sheep, but whether it has the same function in goats remains to be further investigated. In this study, we utilized 348 Nanjiang Yellow goats as experimental subjects, conducted a whole exon, and the conserved the non-coding region SNP scanning of the B4GALNT2 gene. Our results revealed the presence of a synonymous mutation, rs672215506, within the exon of the B4GALNT2 gene in the Nanjiang Yellow goat population. This mutation was found to be associated with reduced mRNA stability of the B4GALNT2 gene. The results of SNPs detection of conserved non-coding region of B4GALNT2 gene showed that there were five potential regulatory SNPs in the Nanjiang Yellow goat population, except for rs66095343, the ~500 bp fragments of four SNPs (rs649127714, rs649573228, rs652899012 and rs639183528) could significantly increase luciferase activity both in goat skeletal muscle cells and 293T cells . Genetic diversity indexes indicated low or intermediate levels for all six SNPs analyzed, each conforming to Hardy-Weinberg equilibrium regarding genotypic frequencies. Association analysis indicated that rs660965343, rs649127714 and rs649573228 were significantly correlated with growth traits in the later stage of growth and development of Nanjiang Yellow goat. The haplotype combinations of H2H3 and H2H2 had higher body weight and greater body size; moreover, H2H2 haplotype combinations were significantly correlated with the litter size of Nanjiang Yellow goat. The results of our study demonstrate the potential utility of the B4GALNT2 gene as a functional genetic marker in breeding programs for Nanjiang Yellow goats.
Keywords:
B4GALNT2
; SNP
; Association analysis
; Goat
; Growth traits
1. Introduction
B4GALNT2 belongs to the family of N-acetyl galactosaminyl transferase (GalNAc-Tases), mainly involved in the formation of O-glycosylation[1]. O-glycosylation is intricately associated with cellular recognition, adhesion, immune response and other vital biological processes[2].The B4GALNT2 gene is currently under extensive investigation for its involvement in animal immunology[1,3,4], gastrointestinal disorders [5,6,7,8,9], and its association with litter size. Additionally, B4GALNT2 has been identified as a potential candidate gene regulating FecL mutations in sheep [10]. Furthermore, the FecL gene and its fertile allele FecLL are among the critical genetic factors influencing ovulation in sheep [11]. Additionally, the B4GALNT2 gene has been implicated in congenital muscular dystrophy [12], and its expression in mouse muscle is dynamic[13]. Manipulation of B4GALNT2 expression can affect the expression of several modifiers associated with muscular dystrophy, and deletion of this gene exacerbates the severity of congenital muscular dystrophy in mice[14]. These findings suggest that B4GALNT2 plays a role in muscle development.
The majority of investigations on the impact of B4GALNT2 polymorphisms on animal production characteristics have primarily focused on sheep reproductive traits [15,16,17]. While B4GALNT2 is located on chromosome 19 in goats, studies describing B4GALNT2 variations associated with the number of lambs in Inner Mongolia White cashmere goats have been reported [18]. However, there is a lack of research investigating changes in goat B4GALNT2 performance, particularly regarding growth parameters.
The B4GALNT2 gene and its non-coding region from 37,020,001 to 37,180,000 on chromosome 19 of Boer goats were subjected to strong selection [19]. In our previous study, this gene and its adjacent regions were strongly selected among Chinese native goat breeds with high reproductive performance and small body size (Meigu goat and Jianchang black goat), which suggested that B4GALNT2 may be a candidate gene affecting the performance and growth traits in goats as well. Consequently, we postulated that genetic variations at the B4GALNT2 gene loci were associated with growth traits and litter size in goats.
In this study, we comprehensively screened variants in the exons and non-coding regions of the B4GALNT2 gene in Nanjiang Yellow goat. Our focus was identifying functional regions containing regulatory elements, which were then subjected to preliminary functional verification. Additionally, we investigated the association between these B4GALNT2 gene variants and growth traits at different stages of development, as well as litter size. We aimed to establish a theoretical basis for selecting and breeding Nanjiang Yellow goats through functional validation at the cellular level and in vivo association analysis.
2. Materials and Methods
2.1. MuSCs isolation and identification
According to previous methods, the LD of the 1D after birth goat (male) was successfully used to isolate the MuSCs for this study [20]. Then, we used the antibody against myogenic marker genes Pax7 (Santa Cruz, USA) and MyHC (myosin heavy chain, Santa Cruz, USA) for immunofluorescence. Store MuSCs in liquid nitrogen tanks. The identification results are shown in Figure S1.
2.2. Animals and samples collection
The Nanjiang Yellow goat population (n = 348) used in the experiment originated exclusively from the Nanjiang Yellow goat stock farm. All goats were subjected to identical management practices and environmental conditions throughout the study. Grazing and appropriate supplementary feeding were employed to raise the goats, ensuring their dietary nutrient levels met their growth requirements. 1.5 mL of whole blood was collected from each test goat via jugular vein puncture, anticoagulated with heparin sodium, and stored at -20 °C for subsequent genomic DNA extraction. Pregnant ewes were randomly selected (n = 3) (unrelated) and humanely killed. Different tissues (LD, lung, heart, spleen, liver, and kidney) were obtained.
2.3. Performance measurement of Nanjiang Yellow goat
The birth weight, body length, body height, and chest circumference of Nanjiang Yellow goats were measured using standard methods at the ages of 6 months, 12 months, and 18 months. The basic data of growth and development and reproductive performance of Nanjiang Yellow sheep are shown in Table S9.
2.4. Plasmid construction
The CDS sequences of the B4GALNT2 (NM_001314262.1) gene were amplified with specific primers, and the full-length of CDS was inserted into the pEGFP-N1 (Promega, WI, USA) vector by the homologous recombinant cloning kit (Vazyme, Nanjing, China) to construct overexpression plasmids. Site-directed mutagenesis kit (Vazyme, Nanjing, China) was used to obtain mutant sequences, and the vector was constructed in the same way.Primers are shown Table S5 and Table S6.
2.5. Cell culture and transfection
MuSCs were cultured at 5 % CO2 and 37℃ in GM (growth medium) containing 89 % Dulbecco’s modified eagle medium(DMEM), 10 % Fetal Bovine Serum (FBS; Gibco, NY, USA), and 1 % penicillin-streptomycin (Invitrogen, NY, USA). Plasmids were transfected into MuSCs using Lipofectamine 3000 (Life Technologies, USA).
2.6. RNA stability assays
Actinomycin D (AcTD, A1410, Sigma-Aldrich) was used on B4GALNT2-G or B4GALNT2-G MuSCs for 0 h, 1 h, 2 h,4,h and 6 h to inhibit global mRNA transcription [21].
2.7. Total RNA isolation and qPCR
Total RNA was isolated from tissues and MuSCs using RNAiso Plus (Takara, Dalian, China). The cDNAs were obtained by the PrimeScript™ RT kit (Takara). In addition, SYBR Premix Ex TaqTM II (Takara) was used for qPCR. Using the GAPDH and 2-ΔΔCt method to normalize relative RNA expression. Primers are shown in Table S3.
2.8. Luciferase reporter assays
Five SNPs (rs660965343, rs649127714, rs639183528, rs652899012 and rs649573228) were inserted into the pGL3-Promoter vector. WT and MUT plasmids were transfected into H293T and MuSCs, respectively. Dual-luciferase reporter kit (Transgen, Beijing, China) was used to detect luciferase activity. Primers for restriction enzyme digestion Table S7.
2.9. Extraction of genomic DNA and detection of DNA quality
Goat genomic DNA was extracted by routine blood genome extraction kit (Tiangen, Beijing, China) and then subjected to 1.5% agarose gel electrophoresis and ultraviolet imaging in gel image analyzer BIO-RAD ChemDOC XRS. The images were analyzed by Quality One 4.6.2 software to determine DNA integrity. The purity and concentration of DNA were determined using a nucleic acid protein detector (BIO-RAD, USA). The samples that met the requirements were stored at -20℃ for later use. Gel electrophoresis is shown in Figure S2.
2.10. PCR amplification and sequencing
Based on the SNP position, each SNP and its flanking sequences were retrieved from the Ensembl database, and primers were designed using the sequence as a template using Primer Premier 5.0 software and synthesized by Sangon (Shanghai, China). The birth record table of each sheep in Nanjiang Yellow goat breeding farm was consulted, and 20 DNA samples were selected and diluted to a concentration of 20 ng/μL. From each sample, 2 μL of DNA was extracted and thoroughly mixed. The resulting mixed pool of DNA served as the template for PCR amplification. The PCR products were sent to Shanghai Sangon (Shanghai, China) for bidirectional Sanger sequencing. SnapGene software was used to verify the SNPS in the samples by comparing the sequencing results with the reference genome sequence and SNP sites. Primers are shown Table S4.
2.11. Mass-array genotyping
In this study, the Sequenom MassArray system genotyping technology was used to genotype 348 Nanjiang Yellow goat experimental population. According to the information of 6 SNPs in 348 DNA samples of Nanjiang Yellow goat, the SNP loci and the upstream and downstream 100 bp sequence information were obtained by Ensembl database. The blood genomic DNA of all samples were submitted to Fuyu Biotechnology (Beijing, China) for genotyping.
2.12. Data Analysis
Haploview4.2 was used to calculate Hardy-Weinberg equilibrium and analyze linkage disequilibrium (LD) among SNPs. PHASE 2.1.1 software was used to construct haplotypes. SAS 9.4 software was used to analyze the association between genotypes of each locus and growth traits of Nanjiang Yellow sheep, and the GLM model in SAS 9.4 was used to establish the model. Y = a+G+p+i+j+fm+e, Y represents the data for each growth trait, a represents the population mean, G is the SNP genotype effect, p is the field effect, i is the year of birth, j is the month of birth, fm represents female (m = male) and e represents the random residual.
2.13. Bioinformatics Analysis
The binding sites of transcription factors at the location of the mutant site and the transcription factors bound were predicted using Jaspar (http://jaspar.genereg.net/).
3. Results
3.1. The synonymous mutation rs672215506 affected the mRNA stability of B4GALNT2 gene
We performed a qPCR to explore the expression profile of B4GALNT2 in goat tissues. (Figure 1A). Notably, a significantly higher expression level was observed in the uterus compared to other tissues (P < 0.01). This result suggests a potential correlation between this gene and reproductive traits in goats. Then, we screened its all exonic SNPs through the utilization of the Ensembl online database and previously collected resequencing data from Nanjiang Yellow goat in our laboratory, resulting in a total discovery of 10 SNPs. Those SNPs were validated through mixed-pool PCR and sequencing in the Nanjiang Yellow goat population, resulting in the identification of only rs672215506, a synonymous mutation, in exonic region (Figure 1B).
To validate the function of rs672215506 mutation, firstly, we predicted the secondary structure of B4GALNT2 mRNA. It revealed that there was only a slight change between the wild type and mutantation type, which was showed in the red box (Figure 1C). Secondly, we calculated the free energy in both types, it was observed that after the mutation, there was a slight increase in the minimum free energy from -2087.40 kJ/mol to -2078.20 kJ/mol. Lastly, we tested its mRNA stability between wild type and mutant type. After successfully transfected B4GALNT2 gene, either contained wild type G or mutant type A, in goat MuSCs, (Figure 1D), which was confirmed by RT-qPCR method (Figure 1E), we found that the expression of B4GALNT2 mRNA was significantly reduced after addition of ACTD 2h and 4h compared with the wild type (Figure 1F) (P < 0.05 or P < 0.01).
3.2. The detection and functional verification of non-coding SNPs in B4GALNT2.
The conservation analysis of the upstream and downstream non-coding region of the B4GALNT2 gene led to the preliminary selection of five candidate SNPs, which were subsequently identified in the Nanjiang Yellow goat population through Sanger sequencing (Figure 2A-E). Then, the analysis of transcription factor binding sites associated with these SNPs was conducted by Jaspar, numerous transcription factors was found to significantly impact the binding affinity (Table S1). To further investigate the functionality of these sites, wild-type and mutant dual luciferase reporter vectors were constructed for each site and subsequently transfected into 293T cells. The results showed that the dual luciferase activity of the wild-type group at all five sites was extremely significantly higher than control group, indicating that the fragment contained these SNPs had enhancer-like activity (Figure 2F-J). Specifically, both rs660965343 and rs649573228 mutations led to a significant reduction in luciferase activity (P < 0.05 or P < 0.01) (Figure 2F&H), whereas rs649127714 and rs652899012 mutations resulted in a significant increase (P < 0.05 or P < 0.01) (Figure 2G&I), while rs639183528 had no significant change (P > 0.05) (Figure 2J). Furthermore, dual-fluorescent vectors exhibiting significant disparities in 293T cells were selected and transfected into MuSCs to investigate whether these motifs possess identical functionality in muscle cells. The results showed that the luciferase activity of the wild-type and mutant vectors showed a similar trend in both cell types (Figure 2K-N). It is interesting that the luciferase activity of the rs660965343 mutant in muscle cells exhibited a significant reduction, even surpassing that of the PGL3-promoter (control) group (P < 0.05), the implication of this finding is that this mutation may play a more significant role in the functionality of muscle cells.
3.3. Population genetic diversity statistics of 6 SNPs in Nanjiang Yellow goat population
From results above, we identified one SNP located at exonic region and 5 SNPs in non-coding region, totally 6 SNPs in B4GALNT2 genes. Then, we are asking whether they are existed in goat population. Mass spectrometry method was applied to analyze genetic distribution of these SNPs in 348 Nanjiang Yellow goats, results showed that 6 SNPs all existed in the goat population (Figure 3A). Among them, rs652062749 had the highest homozygosity, and rs672215506 had the lowest homozygosity. The analysis of effective allele numbers for different SNPs reveals that the overall effective allele number is approximately 2, except for rs652062749, which shows relatively low values, indicating an even distribution of effective alleles within the population. Population genetic statistics showed that all mutations were moderately or weakly polymorphic in Nanjiang Yellow goat (0.25 < PIC ≤ 0.5 or PIC ≤ 0.25). Chi-square fitness test showed that the SNP distribution was in Hardy-Weinberg equilibrium (p > 0.05, Table 1).
3.4. Analysis of linkage disequilibrium and construction of haplotypes
The presence of SNPs in linkage disequilibrium (LD) can provide additional insights into genetic information, and it was conducted based on the SNPs and genotyping information (Figure 3B). A strong LD was observed between rs672215506 and rs660965343 (D’ = 1.000). Subsequently, two highly linked SNPs were used to construct three haplotypes, with the GG haplotype having the highest frequency (0.487). Additionally, six different combinations of these haplotypes were identified. Except for H1H1, all other haplotype combinations had frequencies higher than 5%(Table 2).
3.5. Association of SNPs and haplotype combinations with growth traits
To validate these SNPs’ function in vivo, we analyzed the its asscciation with growth traits both in a SNP locus and haplotype combination. For a single SNPs part, we found that the birth weight of those with the AA genotype was significantly higher than that of goats with the GG genotype in rs672215506 site, (P < 0.05) (Table 3). However, the BH-6, BL-12, and BH-12 measurements in goats with the AA genotype were significantly lower compared to those with the GG genotype (P < 0.05 or P < 0.01). Individuals with the TT genotype at rs660965343 exhibited a significant advantage in terms of birth weight (P < 0.05), but demonstrated inferiority during subsequent periods (P < 0.05 or P < 0.01). The goats with the AA genotype of rs649127714 exhibited significantly larger body size or heavier body mass compared to goats with other genotypes at all periods (P < 0.05 or P < 0.01). The individuals with the CC genotype of rs639183528 exhibited significantly higher BL-6 levels compared to the TT genotype (P < 0.05). The rs649573228 locus did not exhibit any GG homozygous genotype. In goats with the AA genotype, BL, BH-6, CC-12, BL-12, BH-12, BL-18, BH-18 and CC-18 showed significantly better results compared to the GA genotype (P < 0.05 or P < 0.01). The rs652899012 AA genotype exhibited a significantly lower frequency compared to the AG genotype in BL-6, BH-6, CC-6, BL-12, and BH-12 (P < 0.05 or P < 0.01) (Table 4). For the haplotype combinations part, overall, H3H3 exhibited inferior performance in terms of body weight and body size compared to other haplotype combinations (P < 0.05 or P < 0.01, STable 5)
3.6. Association of SNPs and haplotype combinations with lambing number
B4GALNT2 gene was well known for affecting lamb number in sheep, from our best knowledge, which was never tested in goat population. Then, we investigated the association between six SNPs and lambing performance in primiparous and multiparous Nanjiang Yellow goat populations (Table S2). Evnthough, results showed no significant variation in lambing performance among different genotypes of each SNP in primiparous and multiparous ewes (P > 0.05), and no significant correlation between individual haplotype combinations and the number of primiparous lambs (P > 0.05), interertingly, the H2H2 haplotype combination exhibited a significantly higher multiparous lamb count than other haplotype combinations (P < 0.05, Table 6).
4. Discussion
Variations in the B4GALNT2 gene in goats, especially those associated with growth traits, have not been extensively studied. In this study, we identified six SNPs in the B4GALNT2 gene located in the conserved non-coding region and exons of the gene in 348 Nanjiang Yellow Goat (known for its fast growth and high reproductive efficiency in China). We found significant associations between these SNPs and production traits and their number of lambs in Nanjiang Yellow Goat, which provide insights for further characterization of the production performance of livestock.
Synonymous mutations occurring in exons do not alter amino acid or protein sequences. However, they may regulate gene function by affecting codon bias during protein translation [22,23,24], and studies have shown that synonymous mutations can affect animal production performance by influencing the efficiency of transcription and translation of genes [25,26,27]. In this study, a synonymous mutation rs672215506 was identified in the B4GALNT2 gene of goats. This locus may influence the birth weight of goats by affecting the stability of B4GALNT2 gene mRNA.
SNPs in non-coding regions can indirectly regulate gene expression processes, thereby affecting animal phenotypes or reproductive performance [28,29]. The dual luciferase reporter vector assay is a precise and dependable method for validating non-coding SNPs in research [30,31,32]. In this study, we carried out dual luciferase assay in two cell types, 293T and MuSCs. Interestingly, the rs660965343 mutant vector showed lower luciferase activity in MuSCs. This locus may have unique effects on muscle development, but further studies are needed.
The level of genetic variation within a population is the most direct expression of genetic diversity, and the level of genetic variation can directly affect the evolutionary potential of the population [33,34]. Population heterozygosity is an essential indicator for judging the genetic diversity of a certain population, which can reflect the degree of genetic diversity of a population [35]. All the six SNPS in this study were in Hardy-Weinberg equilibrium, and there was rich genetic diversity within the Nanjiang Yellow Goat population, which had good purification and selection potential [36]. In addition, all loci had low (PIC ≤ 0.25) or moderate polymorphism (0.25 < PIC ≤ 0.5), with some genetic variation potential [37,38].
The influence of genes on traits may be influenced by the linkage effect of multiple SNP loci [39,40]. Linkage disequilibrium SNPs can provide more comprehensive genetic information and enhance selection efficiency [41,42]. In this study, the haplotype combination of H2H3 and H2H2 was found to be beneficial for increasing body weight and size, while H2H2 showed advantages for increasing the number of multiparous lambs.In addition, the mean value of the multiparous lambing number was significantly higher than the number of primiparous lambing numbers, which was in line with the results of the previous study [43].
5. Conclusions
This study focused on the SNPs in the exons and conserved non-coding regions of the B4GALNT2 gene in goats, validated their cellular functions, and described their associations with growth traits and lamb size in Nanjiang Yellow goat. The synonymous mutation rs672215506 was found to impact the mRNA stability of the gene. Both rs649127714 and rs652899012 were observed to enhance luciferase activity in MuSCs and 293T cells. Additionally, significant associations were identified between rs660965343, rs649127714, and rs649573228 with growth traits in Nanjiang Yellow goat. The haplotype combination of H2H3 and H2H2 was found to be beneficial for increasing body weight and size, while H2H2 showed advantages for increasing the number of multiparous lambs. These findings provide a theoretical foundation for genetic selection and breeding strategies for Nanjiang Yellow goat.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1. Transcription factors prediction. Table S2. The association between six SNPs and lambing performance in primiparous and multiparous Nanjiang Yellow goat populations. Table S3. RT-qPCR primers information. Table S4. Primers for exon validation. Table S5. Primers Information of homologous recombination. Table S6. Information of site-directed mutagenesis primers. Table S7. Primers information of non-coding region SNPs of B4GALNT2. Table S8. The primers information of enzyme digestion sites. Table S9. Basic data of growth, development and reproductive performance of Nanjiang Yellow goat. Figure S1. The identification results. Pax7 (red) and MyHC (red) immunofluorescence staining were performed in MuSCs. Figure S2. Gel electrophoresis of genomic DNA samples. M. DL2000 DNA maker; (1-6) Genomic DNA of Nanjiang Yellow goat.
Author Contributions
Conceptualization, L.X., H.Z. and J.C.; methodology, S.C.; software, Z.C.; validation, J.C.; formal analysis, Y.C. and J.G.; investigation, T.Z. and L.W.; resources, T.Z. and L.W.; data curation, L.X.; writing—original draft preparation, L.X.; writing—review and editing, J.C., S.Z. and L.L.; visualization, S.C. and Z.C.; supervision, J.C.; project administration, J.C.; funding acquisition, H.Z. and J.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Science and Technology Support for Returned Personnel of Sichuan Province (2021-22), the Young Scientists Fund of the National Natural Science Foundation of China, grant No.31802048, and the Natural Science Foundation of Sichuan Province, grant No. 2022NSFSC1611.
Institutional Review Board Statement
The Institutional Animal Care and Utilization Committee of Sichuan Agricultural University approved all experimental schemes of this study. Moreover, the animals were sacrificed in accordance with the Regulations for the Administration of Affairs Concerning Experimental Animals, under permit NO DKY-2020202033.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data from the current study are exhibited in the manuscript and supplementary materials.
Acknowledgments
Data related to the production of Nanjiang Yellow goat were obtained from the breeding farm of Nanjiang Yellow goat in Sichuan Province. Thanks to researcher Yu Chen for her support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Byrne, G.; Ahmad-Villiers, S.; Du, Z.; McGregor, C. B4GALNT2 and xenotransplantation: A newly appreciated xenogeneic antigen. Xenotransplantation 2018, 25, e12394. [CrossRef]
- Stwora-Wojczyk, M.M.; Kissinger, J.C.; Spitalnik, S.L.; Wojczyk, B.S. O-glycosylation in Toxoplasma gondii: identification and analysis of a family of UDP-GalNAc:polypeptide N-acetylgalactosaminyltransferases. International journal for parasitology 2004, 34, 309-322. [CrossRef]
- Zhao, C.; Cooper, D.K.C.; Dai, Y.; Hara, H.; Cai, Z.; Mou, L. The Sda and Cad glycan antigens and their glycosyltransferase, β1,4GalNAcT-II, in xenotransplantation. Xenotransplantation 2018, 25. [CrossRef]
- Duca, M.; Malagolini, N.; Dall’Olio, F. The story of the Sd(a) antigen and of its cognate enzyme B4GALNT2: What is new? Glycoconjugate journal 2023, 40, 123-133. [CrossRef]
- Groux-Degroote, S.; Vicogne, D.; Cogez, V.; Schulz, C.; Harduin-Lepers, A. B4GALNT2 Controls Sd(a) and SLe(x) Antigen Biosynthesis in Healthy and Cancer Human Colon. Chembiochem : a European journal of chemical biology 2021, 22, 3381-3390. [CrossRef]
- Galeev, A.; Suwandi, A.; Cepic, A.; Basu, M.; Baines, J.F.; Grassl, G.A. The role of the blood group-related glycosyltransferases FUT2 and B4GALNT2 in susceptibility to infectious disease. International journal of medical microbiology : IJMM 2021, 311, 151487. [CrossRef]
- Groux-Degroote, S.; Wavelet, C.; Krzewinski-Recchi, M.A.; Portier, L.; Mortuaire, M.; Mihalache, A.; Trinchera, M.; Delannoy, P.; Malagolini, N.; Chiricolo, M.; et al. B4GALNT2 gene expression controls the biosynthesis of Sda and sialyl Lewis X antigens in healthy and cancer human gastrointestinal tract. The international journal of biochemistry & cell biology 2014, 53, 442-449. [CrossRef]
- Rausch, P.; Steck, N.; Suwandi, A.; Seidel, J.A.; Künzel, S.; Bhullar, K.; Basic, M.; Bleich, A.; Johnsen, J.M.; Vallance, B.A.; et al. Expression of the Blood-Group-Related Gene B4galnt2 Alters Susceptibility to Salmonella Infection. PLoS pathogens 2015, 11, e1005008. [CrossRef]
- Pucci, M.; Gomes Ferreira, I.; Malagolini, N.; Ferracin, M.; Dall’Olio, F. The Sd(a) Synthase B4GALNT2 Reduces Malignancy and Stemness in Colon Cancer Cell Lines Independently of Sialyl Lewis X Inhibition. International journal of molecular sciences 2020, 21. [CrossRef]
- Drouilhet, L.; Mansanet, C.; Sarry, J.; Tabet, K.; Bardou, P.; Woloszyn, F.; Lluch, J.; Harichaux, G.; Viguié, C.; Monniaux, D.; et al. The highly prolific phenotype of Lacaune sheep is associated with an ectopic expression of the B4GALNT2 gene within the ovary. PLoS genetics 2013, 9, e1003809. [CrossRef]
- Drouilhet, L.; Lecerf, F.; Bodin, L.; Fabre, S.; Mulsant, P. Fine mapping of the FecL locus influencing prolificacy in Lacaune sheep. Animal genetics 2009, 40, 804-812. [CrossRef]
- Thomas, P.J.; Xu, R.; Martin, P.T. B4GALNT2 (GALGT2) Gene Therapy Reduces Skeletal Muscle Pathology in the FKRP P448L Mouse Model of Limb Girdle Muscular Dystrophy 2I. The American journal of pathology 2016, 186, 2429-2448. [CrossRef]
- Xu, R.; Singhal, N.; Serinagaoglu, Y.; Chandrasekharan, K.; Joshi, M.; Bauer, J.A.; Janssen, P.M.; Martin, P.T. Deletion of Galgt2 (B4Galnt2) reduces muscle growth in response to acute injury and increases muscle inflammation and pathology in dystrophin-deficient mice. The American journal of pathology 2015, 185, 2668-2684. [CrossRef]
- Cramer, M.L.; Xu, R.; Martin, P.T. Soluble Heparin Binding Epidermal Growth Factor-Like Growth Factor Is a Regulator of GALGT2 Expression and GALGT2-Dependent Muscle and Neuromuscular Phenotypes. Molecular and cellular biology 2019, 39. [CrossRef]
- Gao, Y.; Wang, J.; Chao, L.; Kuan, M.; Yu, H.; Wang, J.; Bao, S.; Liu, Y.; Zhang, W.; Ma, Q.; et al. Association analysis of B4GALNT2 gene polymorphism with lambing number in Mongolian and Uzhumqin sheep. Journal of Agricultural Biotechnology 2022, 30, 1510-1523.
- Rong, X.; Shao, S.; Liang, P.; Zhang, T.; Zou, S.; Meng, K.; Qiang, H.; Feng, D. Analysis of polymorphisms in B4GALNT2 and ESR1 genes and their association with lambing number in sheep. China Animal Husbandry & Veterinary Medicine 2021, 48, 3332-3342. [CrossRef]
- Guo, X.; Wang, X.; Liang, B.; Di, R.; Liu, Q.; Hu, W.; He, X.; Zhang, J.; Zhang, X.; Chu, M. Molecular Cloning of the B4GALNT2 Gene and Its Single Nucleotide Polymorphisms Association with Litter Size in Small Tail Han Sheep. Animals : an open access journal from MDPI 2018, 8. [CrossRef]
- Sa, C.; Wu, T.; Ma, Y.; He, Y.; Zhu, L.; He, T.; Wu, Y.; Liu, B. Analysis of polymorphisms in four candidate genes for multiparous traits and their association with lambing number in cashmere goats. China Animal Husbandry & Veterinary Medicine 2023, 50, 1037-1047. [CrossRef]
- Yang, B.G.; Yuan, Y.; Zhou, D.K.; Ma, Y.H.; Mahrous, K.F.; Wang, S.Z.; He, Y.M.; Duan, X.H.; Zhang, W.Y.; E, G. Genome-wide selection signal analysis of Australian Boer goat reveals artificial selection imprinting on candidate genes related to muscle development. Animal genetics 2021, 52, 550-555. [CrossRef]
- Zhao, W.; Chen, L.; Zhong, T.; Wang, L.; Guo, J.; Dong, Y.; Feng, J.; Song, T.; Li, L.; Zhang, H. The differential proliferation and differentiation ability of skeletal muscle satellite cell in Boer and Nanjiang brown goats. Small Ruminant Research 2018, 169, 99-107. [CrossRef]
- Chen, L.; Zhang, C.; Ma, W.; Huang, J.; Zhao, Y.; Liu, H. METTL3-mediated m6A modification stabilizes TERRA and maintains telomere stability. Nucleic Acids Research 2022, 50, 11619-11634. [CrossRef]
- Chaney, J.L.; Clark, P.L. Roles for Synonymous Codon Usage in Protein Biogenesis. Annual review of biophysics 2015, 44, 143-166. [CrossRef]
- Otsuka, H.; Sasai, H.; Nakama, M.; Aoyama, Y.; Abdelkreem, E.; Ohnishi, H.; Konstantopoulou, V.; Sass, J.O.; Fukao, T. Exon 10 skipping in ACAT1 caused by a novel c.949G>A mutation located at an exonic splice enhancer site. Molecular medicine reports 2016, 14, 4906-4910. [CrossRef]
- Plotkin, J.B.; Kudla, G. Synonymous but not the same: the causes and consequences of codon bias. Nature reviews. Genetics 2011, 12, 32-42. [CrossRef]
- Szewczuk, M.; Zych, S.; Wojcik, J.; Czerniawska-Piatkowska, E. Association of two SNPs in the coding region of the insulin-like growth factor 1 receptor (IGF1R) gene with growth-related traits in Angus cattle. J Appl Genet 2013, 54, 305-308. [CrossRef]
- Cui, C.; Jiang, H.; Liang, Y.; Xiao, C.; Liu, Y.; Jin, H.; Cao, Y. Analysis of HLF gene polymorphism and its association with muscle fatty acid and amino acid contents in sheep. Heilongjiang Animal Husbandry and Veterinary Medicine 2022.
- Cheng, Y.; Liu, S.; Wang, G.; Wei, W.; Huang, S.; Yang, R.; Geng, H.; Li, H.; Song, J.; Sun, L.; et al. Porcine IGF1 synonymous mutation alter gene expression and protein binding affinity with IGF1R. Int J Biol Macromol 2018, 116, 23-30. [CrossRef]
- Jolma, A.; Yan, J.; Whitington, T.; Toivonen, J.; Nitta, K.R.; Rastas, P.; Morgunova, E.; Enge, M.; Taipale, M.; Wei, G.; et al. DNA-binding specificities of human transcription factors. Cell 2013, 152, 327-339. [CrossRef]
- Hoogendoorn, B.; Coleman, S.L.; Guy, C.A.; Smith, S.K.; O’Donovan, M.C.; Buckland, P.R. Functional analysis of polymorphisms in the promoter regions of genes on 22q11. Hum Mutat 2004, 24, 35-42. [CrossRef]
- Zhang, Z.; Liu, C.; Hao, W.; Yin, W.; Ai, S.; Zhao, Y.; Duan, Z. Novel Single Nucleotide Polymorphisms and Haplotype of MYF5 Gene Are Associated with Body Measurements and Ultrasound Traits in Grassland Short-Tailed Sheep. Genes (Basel) 2022, 13. [CrossRef]
- Chen, W.; Xu, H.; Chen, X.; Liu, Z.; Zhang, W.; Xia, D. Functional and Activity Analysis of Cattle UCP3 Promoter with MRFs-Related Factors. Int J Mol Sci 2016, 17. [CrossRef]
- Wang, D. Analysis of transcriptional regulation of non-coding region of FSHR gene in lake sheep. Nanjing Agricultural University.
- Kamarudin, N.J.; Wang, V.C.; Tan, X.T.; Ramesh, A.; Ling, M.H. A Simulation Study on the Effects of Founding Population Size and Number of Alleles Per Locus on the Observed Population Genetic Profile: Implications to Broodstock Management. 2020.
- Barrandeguy, M.E.; García, M. The Sensitiveness of Expected Heterozygosity and Allelic Richness Estimates for Analyzing Population Genetic Diversity; Genetic Diversity [Working Title]: 2021.
- Silió, L.; Rodríguez, M.C.; Fernández, A.; Barragán, C.; Benítez, R.; Óvilo, C.; Fernández, A.I. Measuring inbreeding and inbreeding depression on pig growth from pedigree or SNP-derived metrics. Journal of Animal Breeding and Genetics 2013, 130, 349-360. [CrossRef]
- Penedo, M.; Weisenberger, M.E.; Boyce, W.M.; Johnson, C.K.J.A.o.O.; Environmental Medicine, 1. Wildlife translocation: the conservation implications of pathogen exposure and genetic heterozygosity. 2011, 11, 1-7.
- Botstein, D.; White, R.L.; Skolnick, M.; Davis, R.W.J.A.j.o.h.g. Construction of a genetic linkage map in man using restriction fragment length polymorphisms. 1980, 32, 314.
- Jiang, Y.; Wang, S.; Zhu, L.; Yang, H.; Hong, Q. Analysis of muscle growth inhibitor gene polymorphism in black goats on Yunnan. Chinese Herbivore Science 2020, 5-7.
- Ma, X.; Du, L.; Zhang, L.; Xuan, J.; Wang, H.; Yuan, Z.; Wu, M.; Zhu, C.; Liu, R.; Wei, C. Association of RIPK2 gene polymorphisms with growth traits in Uzhumqin sheep. Chinese Agricultural Science 2016, 49, 17.
- Peng, Y.; Liu, J.; Zhao, S.; Xu, Z.; Zuo, B. Detection of SNPs in porcine RXRB gene and their association analysis with growth and fattening and reproductive traits. Journal of Animal Husbandry and Veterinary Science 2021, 52, 03.
- Meng, K.; Zhang, T.; Liang, P.; Shao, S.; Zou, S.; Rong, X.; Qiang, H.; Feng, D. Analysis of polymorphisms in sheep MYF5 gene and their association with growth traits. Journal of Agricultural Biotechnology 2022, 030.
- Wu, S.-B.; Franks, T.K.; Hunt, P.; Wirthensohn, M.G.; Gibson, J.P.; Sedgley, M. Discrimination of SNP genotypes associated with complex haplotypes by high resolution melting analysis in almond: implications for improved marker efficiencies. Molecular Breeding 2009, 25, 351-357. [CrossRef]
- Li, L.; Wu, D.; et al. Correlation analysis between LHβ gene polymorphism and reproductive performance of Nanjiang yellow sheep. Livestock and Veterinary Medicine 2006, 3-5.
Figure 1.
The expression pattern of the B4GALNT2 gene in various tissues and the effect of B4GALNT2 synonymous mutation on mRNA stability. (A) B4GALNT2 gene expression profile in different tissues of goat. (B) SNPs sequencing peak of B4GALNT2 exons. The base indicated by the arrow in the figure rep-resents the genomic locus of the corresponding SNP. A double peak indicates the presence of this SNP within the Nanjiang Yellow goat population. (C) Prediction of the secondary structure of B4GALNT2 mRNA. After mutation, red box mRNA secondary structure has changed. (D)The transfection efficiency of wild type and mutant type overexpression vectors in goat MuSCs was assessed. Cells carrying green fluorescence represent successful transfection. (E) The expression levels of the wild type and mutant type of the B4GALNT2 gene were detected in goat MuSCs. (F)The relative expression levels of the wild type and mutant type of the B4GALNT2 gene in goat MuSCs were evaluated after treatment with actinomycin D for different times. Results are represented as the mean ± SEM, * p < 0.05, ** p < 0.01.
Figure 1.
The expression pattern of the B4GALNT2 gene in various tissues and the effect of B4GALNT2 synonymous mutation on mRNA stability. (A) B4GALNT2 gene expression profile in different tissues of goat. (B) SNPs sequencing peak of B4GALNT2 exons. The base indicated by the arrow in the figure rep-resents the genomic locus of the corresponding SNP. A double peak indicates the presence of this SNP within the Nanjiang Yellow goat population. (C) Prediction of the secondary structure of B4GALNT2 mRNA. After mutation, red box mRNA secondary structure has changed. (D)The transfection efficiency of wild type and mutant type overexpression vectors in goat MuSCs was assessed. Cells carrying green fluorescence represent successful transfection. (E) The expression levels of the wild type and mutant type of the B4GALNT2 gene were detected in goat MuSCs. (F)The relative expression levels of the wild type and mutant type of the B4GALNT2 gene in goat MuSCs were evaluated after treatment with actinomycin D for different times. Results are represented as the mean ± SEM, * p < 0.05, ** p < 0.01.
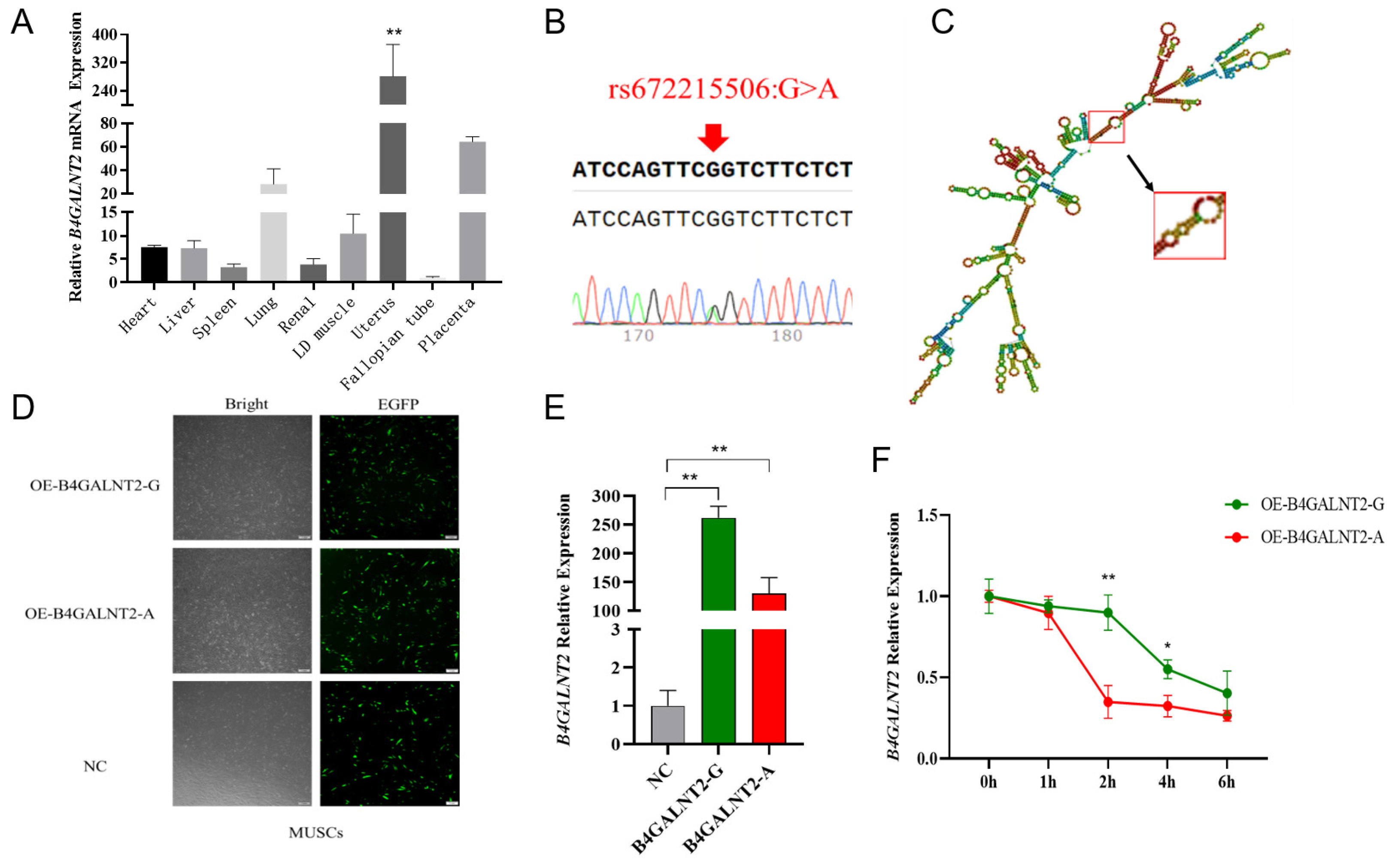
Figure 2.
Five SNPs sequencing results and the effect of mutations on enhancer activity. (A-E) Five SNPs sequencing results of mixed pool samples, which located at the non-coding region of B4GALNT2 gene. The base indicated by the arrow in the figure represents the genomic locus of the corresponding SNP. A double peak indicates the presence of this SNP within the Nanjiang Yellow goat population. (F-J) Effect of mutations on enhancer activity. Blue represents the mutant type, red represents the wild type, and green represents control. They were transfected into 293T cells. (K-N) Effect of mutations on enhancer activity. They were transfected into MuSCs cells. The results are expressed as the mean ± SEM (n = 3 or 4) in arbitrary units based on firefly luciferase activity normalized against Renilla luciferase activity. A t-test was conducted using SPSS 25.0 to detect the differences. Bars represent the mean ± SEM of at least three repeats. *p < 0.05; **p < 0.01.
Figure 2.
Five SNPs sequencing results and the effect of mutations on enhancer activity. (A-E) Five SNPs sequencing results of mixed pool samples, which located at the non-coding region of B4GALNT2 gene. The base indicated by the arrow in the figure represents the genomic locus of the corresponding SNP. A double peak indicates the presence of this SNP within the Nanjiang Yellow goat population. (F-J) Effect of mutations on enhancer activity. Blue represents the mutant type, red represents the wild type, and green represents control. They were transfected into 293T cells. (K-N) Effect of mutations on enhancer activity. They were transfected into MuSCs cells. The results are expressed as the mean ± SEM (n = 3 or 4) in arbitrary units based on firefly luciferase activity normalized against Renilla luciferase activity. A t-test was conducted using SPSS 25.0 to detect the differences. Bars represent the mean ± SEM of at least three repeats. *p < 0.05; **p < 0.01.
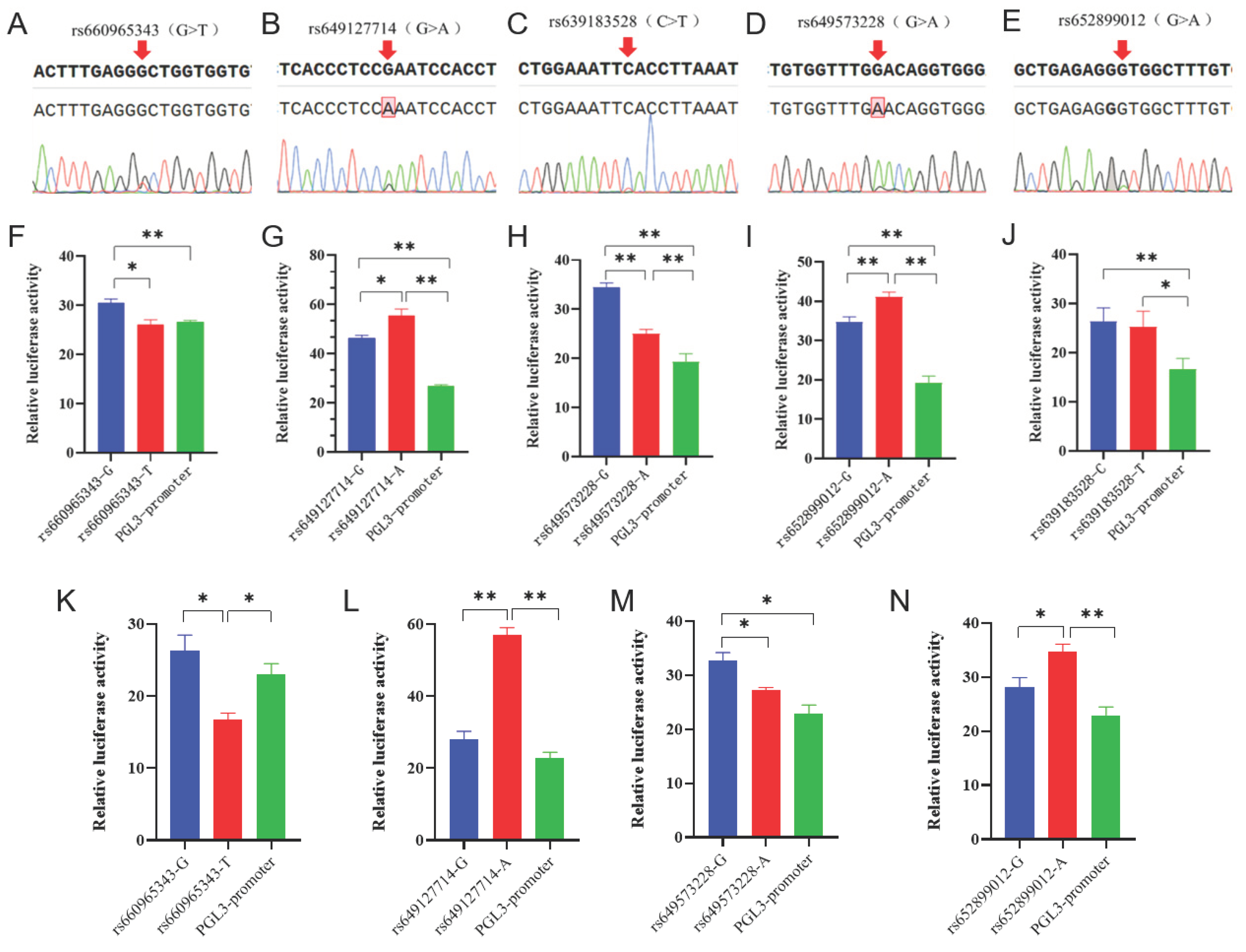
Figure 3.
Mass spectrometry results and linkage disequilibrium analysis. (A) SNP genotyping results are shown as scatter plots, different colors dots representing different genotypes. (B) Linkage disequilibrium analysis of 6 SNPs in Nanjiang Yellow goat population.
Figure 3.
Mass spectrometry results and linkage disequilibrium analysis. (A) SNP genotyping results are shown as scatter plots, different colors dots representing different genotypes. (B) Linkage disequilibrium analysis of 6 SNPs in Nanjiang Yellow goat population.
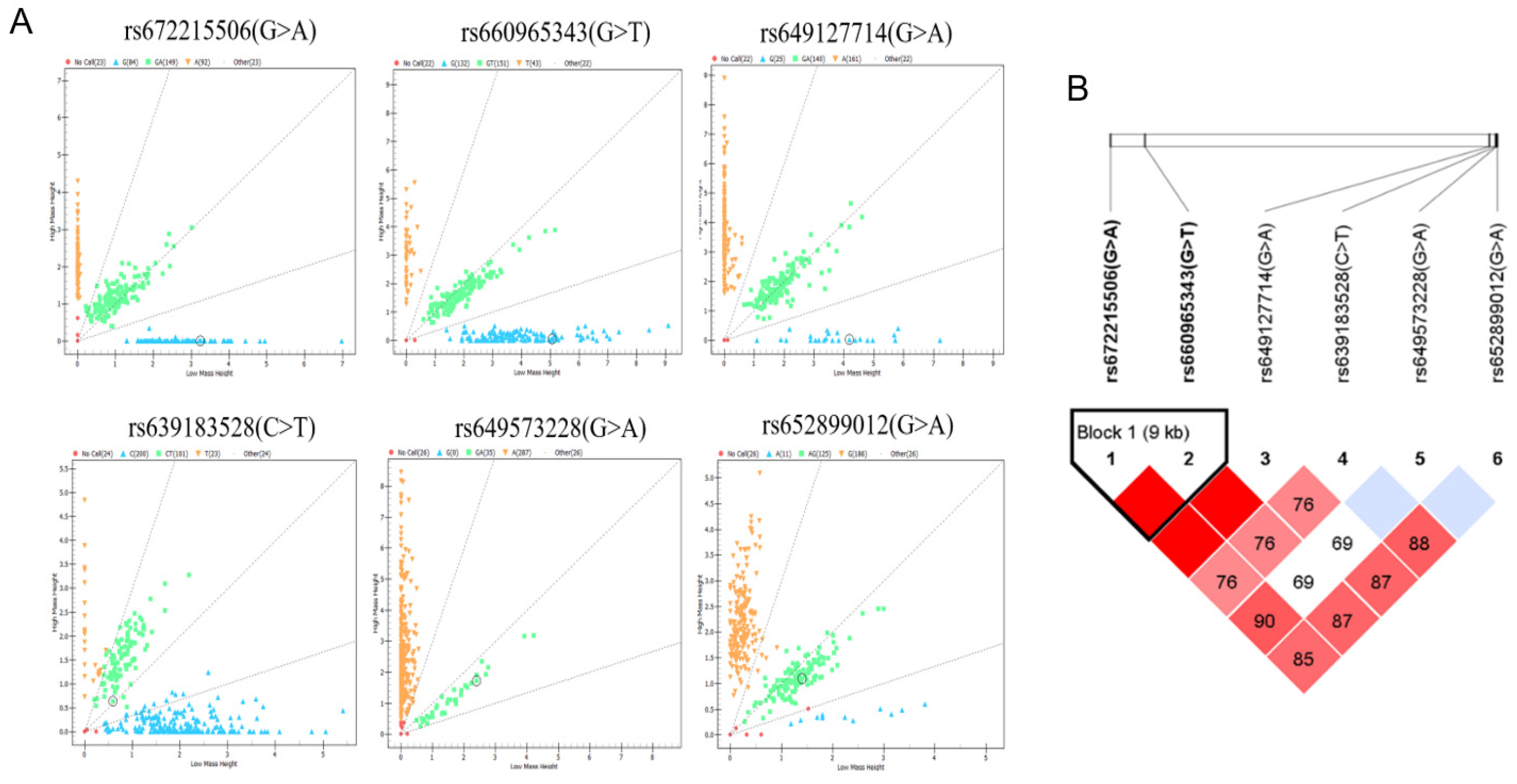

Table 1.
Genetic parameters and Hardy-Weinberg equilibrium test of 6 SNPs in the goat Population.
| Locus | Genotype | Genotype frequency | Allele frequency | Ho | He | Ne | P | PIC |
|---|---|---|---|---|---|---|---|---|
| rs672215506(G>A) | AA | 0.28 | 0.51(A) | 0.5 | 0.5 | 2 | 0.16 | 0.37 |
| GA | 0.46 | 0.49(G) | ||||||
| GG | 0.26 | |||||||
| rs660965343(G>T) | GG | 0.4 | 0.64(G) | 0.54 | 0.46 | 1.85 | 1 | 0.35 |
| GT | 0.46 | 0.36(T) | ||||||
| TT | 0.14 | |||||||
| rs649127714(G>A) | GG | 0.08 | 0.29(G) | 0.59 | 0.41 | 1.7 | 0.58 | 0.33 |
| GA | 0.43 | 0.71(A) | ||||||
| AA | 0.49 | |||||||
| rs639183528(C>T) | CC | 0.62 | 0.77(C) | 0.65 | 0.35 | 1.54 | 0.07 | 0.29 |
| CT | 0.31 | 0.23(T) | ||||||
| TT | 0.07 | |||||||
| rs649573228(G>A) | AA | 0.89 | 0.95(A) | 0.9 | 0.1 | 1.11 | 0.75 | 0.1 |
| GA | 0.11 | 0.05(G) | ||||||
| rs652899012(G>A) | GG | 0.58 | 0.65 | 0.35 | 1.54 | 0.09 | 0.29 | |
| AG | 0.39 | 0.77(G) | ||||||
| AA | 0.03 | 0.23(A) |
Note: Ho, He, Ne and PIC represent homozygosity, heterozygosity, effective number of alleles and polymorphism information content respectively; P > 0.05 indicates the locus was under Hardy–Weinberg equilibrium.
Table 2.
Haplotype and haplotype combination construction.
| Haplotype and Haplotype combination | Type | Genotype | Frequency |
|---|---|---|---|
| Haplotype | H1 | GG | 0.487 |
| H2 | AG | 0.141 | |
| H3 | AT | 0.372 | |
| H1H3 | GGAT | 0.379 | |
| Haplotype combination | H2H2 | GGGG | 0.241 |
| H3H3 | ATAT | 0.124 | |
| H2H3 | AGAT | 0.118 | |
| H1H2 | GGAG | 0.112 | |
| H1H1 | AGAG | 0.026 |
Table 3.
Association analysis of six SNPs with birth weight in Nanjiang Yellow goat Population.
| Locus | Genotype | Number | Birth weight |
|---|---|---|---|
| rs672215506(G>A) | AA | 92 | 2.29±0.28a |
| GA | 149 | 2.26±0.29ab | |
| GG | 84 | 2.20±0.31b | |
| rs660965343(G>T) | TT | 43 | 2.29±0.26a |
| GT | 151 | 2.27±0.33ab | |
| GG | 132 | 2.22±0.27b | |
| rs649127714(G>A) | AA | 161 | 2.27±0.34 |
| GA | 140 | 2.24±0.24 | |
| GG | 25 | 2.22±0.30 | |
| rs639183528(C>T) | CC | 200 | 2.26±0.32 |
| CT | 101 | 2.24±0.26 | |
| TT | 23 | 2.23±0.25 | |
| rs649573228(G>A) | AA | 287 | 2.25±0.31 |
| GA | 35 | 2.24±0.24 | |
| AA | 11 | 2.26±0.26 | |
| rs652899012(G>A) | AG | 125 | 2.22±0.27 |
| GG | 186 | 2.27±0.32 |
Note: Different small letters in the same group mean significant difference (p < 0.05).
Table 4.
Association analysis of 6 SNPs with different month body size trait in the Nanjiang Yellow goat population.
Table 4.
Association analysis of 6 SNPs with different month body size trait in the Nanjiang Yellow goat population.
| Locus | rs672215506(G>A) | rs660965343(G>T) | rs649127714(G>A) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Genotype | AA | GA | GG | TT | GT | GG | AA | GA | GG | |
| Number | 60 | 88 | 49 | 26 | 89 | 82 | 100 | 82 | 15 | |
| BW-6 | 25.99±3.87 | 25.83±4.45 | 26.28±4.58 | 25.42±3.67b | 25.99±4.16ab | 26.17±4.64a | 26.66±4.42A | 25.26±3.91B | 25.53±5.03B | |
| BL-6 | 58.85±5.21 | 59.06±5.42 | 59.73±5.53 | 58.00±4.43B | 59.16±5.23A | 59.54±5.79A | 59.79±5.59A | 58.37±4.98B | 59.33±5.77AB | |
| BH-6 | 55.80±4.44B | 56.11±4.96A | 56.11±4.96A | 55.23±3.92B | 56.21±4.79A | 56.43±4.80A | 56.71±4.99A | 55.50±4.24B | 56.27±4.67AB | |
| CC-6 | 65.19±4.02 | 65.06±4.56 | 65.71±4.83 | 64.37±3.65B | 65.17±4.23AB | 65.65±4.92A | 65.89±4.69Aa | 64.54±4.09Bb | 65.07±4.48ABb | |
| BW-12 | 34.08±4.54 | 34.22±5.04 | 34.49±5.31 | 33.37±4.18Bb | 34.37±4.80ABa | 34.39±5.33Aa | 35.02±5.23A | 33.38±4.33B | 33.83±5.55B | |
| BL-12 | 65.70±4.76Bb | 66.11±5.14ABb | 66.80±5.65Aa | 64.73±4.19Bc | 66.13±5.01Ab | 66.63±5.54Aa | 66.72±5.46A | 65.44±4.68B | 66.33±5.39AB | |
| BH-12 | 62.75±4.04b | 63.13±4.55b | 63.73±4.71a | 62.15±3.56Bb | 63.08±4.37ABa | 63.57±4.74Aa | 63.61±4.69a | 62.55±4.03b | 63.53±4.70a | |
| CC-12 | 74.86±4.93 | 75.26±5.28 | 75.87±5.20 | 74.12±4.73Bb | 75.23±4.92ABa | 75.73±5.49Aa | 75.76±5.51 | 74.76±4.74 | 75.10±4.73 | |
| BW-18 | 47.23±6.43 | 48.39±7.62 | 48.20±7.45 | 46.33±6.08B | 48.20±7.37A | 48.29±7.39A | 48.67±7.79a | 47.10±6.08b | 48.30±8.80ab | |
| BL-18 | 72.78±4.80 | 73.56±5.42 | 73.53±5.65 | 71.35±4.23B | 73.48±5.11A | 73.76±5.68A | 73.77±5.58a | 72.82±4.84b | 73.00±5.66b | |
| BH-18 | 69.17±3.95 | 69.30±4.31 | 69.41±4.38 | 68.38±3.42B | 69.25±4.22AB | 69.61±4.40A | 69.69±4.43A | 68.70±3.69AB | 69.80±5.10B | |
| CC-18 | 85.52±4.66 | 85.85±5.10 | 85.71±4.88 | 84.46±4.34B | 85.93±4.62A | 85.88±5.33A | 86.32±5.13a | 85.04±4.37b | 85.37±5.80b | |
| Locus | rs639183528(C>T) | rs649573228(G>A) | rs652899012(G>A) | |||||||
| Genotype | CC | CT | TT | AA | GA | AA | AG | GG | ||
| Number | 119 | 63 | 15 | 174 | 21 | 7 | 71 | 117 | ||
| BW-6 | 26.38±4.45 | 25.47±3.85 | 25.10±4.69 | 26.14±4.28 | 24.86±4.56 | 25.71±5.19 | 25.19±4.00 | 26.46±4.41 | ||
| BL-6 | 59.45±5.54a | 58.89±5.03ab | 58.00±5.57b | 59.30±5.41A | 58.05±5.31B | 60.14±6.15A | 58.49±5.09B | 59.56±5.52AB | ||
| BH-6 | 56.45±4.97 | 55.76±4.08 | 55.67±4.82 | 56.32±4.70A | 55.14±4.67B | 57.29±5.22A | 55.48±4.16B | 56.53±4.97AB | ||
| CC-6 | 65.55±4.72 | 64.92±4.07 | 64.40±3.96 | 65.44±4.47A | 64.00±4.42B | 65.71±4.03a | 64.65±4.23b | 65.62±4.64ab | ||
| BW-12 | 34.60±5.17 | 33.79±4.47 | 33.30±5.02 | 34.43±4.95A | 32.67±4.91B | 34.07±5.76 | 33.48±4.48 | 34.71±5.18 | ||
| BL-12 | 66.39±5.34 | 65.87±4.76 | 65.47±5.44 | 66.29±5.22A | 65.24±4.82B | 66.57±6.16a | 65.58±4.72b | 66.51±5.39ab | ||
| BH-12 | 63.34±4.66 | 62.86±3.95 | 63.00±4.77 | 63.29±4.46A | 62.24±4.41B | 63.86±5.24a | 62.65±4.01b | 63.43±4.67ab | ||
| CC-12 | 75.45±5.34 | 75.30±5.02 | 73.97±3.95 | 75.41±5.18 | 74.52±5.06 | 75.36±5.02 | 74.80±4.80 | 75.58±5.40 | ||
| BW-18 | 48.41±7.58 | 47.40±6.42 | 47.10±7.66 | 48.14±7.34 | 47.05±6.45 | 47.79±8.32 | 47.20±6.63 | 48.53±7.55 | ||
| BL-18 | 73.61±5.52 | 72.98±4.91 | 72.40±5.03 | 73.40±5.32a | 73.00±5.2b | 73.29±5.88 | 72.77±4.96 | 73.7±5.48 | ||
| BH-18 | 69.47±4.34 | 68.94±3.82 | 69.27±4.76 | 69.4±4.24A | 68.57±4.01B | 68.71±4.82 | 68.83±3.81 | 69.55±4.34 | ||
| CC-18 | 86.02±5.10 | 85.33±4.53 | 84.87±4.81 | 85.84±4.92A | 84.60±4.91B | 85.00±4.93 | 85.08±4.72 | 86.13±5.03 | ||
Note: Values are shown as means ± standard deviation; a , b means with different superscripts are significantly different (p < 0.05);A , B means with different superscripts are very significantly different (p < 0.01) ; body weight for six months (BW-6), body length for six months (BL-6), body height for six months (BH-6), chest circumference for six months (CC-6) and so on.
Table 5.
Association analysis between haplotype combination and growth traits of Nanjiang Yellow goat.
Table 5.
Association analysis between haplotype combination and growth traits of Nanjiang Yellow goat.
| Combined Haplotypes | H2H3 | H3H3 | H1H2 | H1H3 | H2H2 |
|---|---|---|---|---|---|
| Number | 27 | 26 | 26 | 75 | 49 |
| BW-6 | 26.30±3.63a | 25.42±3.67b | 25.77±4.67ab | 25.66±4.42ab | 26.28±4.58ab |
| BL-6 | 59.52±5.4Aa | 58±4.43Bb | 59.19±6.06Aa | 58.8±5.29ABab | 59.73±5.53Aa |
| BH-6 | 56.11±4.46Aa | 55.23±3.92Bb | 55.77±5.04ABa | 56.07±5.02ABa | 56.73±4.51Aa |
| CC-6 | 65.70±3.71Aa | 64.37±3.65Bc | 65.35±4.91Aab | 64.74±4.40ABbc | 65.71±4.83Aab |
| BW-12 | 34.33±4.46a | 33.37±4.18b | 33.83±5.30ab | 34.14±4.95ab | 34.49±5.31a |
| BL-12 | 66.07±4.82Aab | 64.73±4.19Bc | 66.00±5.29Aab | 66.02±5.09Ab | 66.80±5.65Aa |
| BH-12 | 62.85±4.03ABa | 62.15±3.56Bb | 63.00±4.67ABa | 63.01±4.47ABb | 63.73±4.71Aa |
| CC-12 | 75.35±4.90Aa | 74.12±4.73Bb | 75.46±6.05Aa | 74.88±4.94Bb | 75.87±5.20Aa |
| BW-18 | 47.76±6.53Aa | 46.33±6.08Bb | 48.38±7.45Aa | 47.93±7.45Ba | 48.20±7.45Aa |
| BL-18 | 73.56±4.55Aab | 71.35±4.23Bc | 73.81±5.64Aa | 73.2±5.22Ab | 73.53±5.65Aab |
| BH-18 | 69.52±4.05Aab | 68.38±3.42Bc | 69.69±4.34Aa | 69.03±4.16Ab | 69.41±4.38Aab |
| CC-18 | 86.33±4.07Aa | 84.46±4.34Bb | 86.08±5.73Aa | 85.61±4.88ABab | 85.71±4.88ABa |
Note: Values are shown as means ± standard deviation; a , b means with different superscripts are significantly different (p < 0.05);A , B means with different superscripts are very significantly different (p < 0.01) ; body weight for six months (BW-6), body length for six months (BL-6), body height for six months (BH-6), chest circumference for six months (CC-6) and so on.
Table 6.
Association analysis between haplotype combination and lambing number of Nanjiang Yellow goat.
Table 6.
Association analysis between haplotype combination and lambing number of Nanjiang Yellow goat.
| Combined Haplotypes | Number | Primiparity | Multiparity |
|---|---|---|---|
| H2H3 | 21 | 1.52±0.51 | 1.82±0.26Cc |
| H3H3 | 23 | 1.74±0.45 | 1.95±0.15BCb |
| H1H2 | 18 | 1.67±0.49 | 1.94±0.24BCb |
| H1H3 | 47 | 1.57±0.54 | 1.98±0.21Bb |
| H2H2 | 34 | 1.65±0.54 | 2.13±0.17Aa |
Note: Values are shown as means ± standard deviation; a , b means with different superscripts are significantly different (p < 0.05);A , B means with different superscripts are very significantly different (p < 0.01).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



