
Submitted:
19 February 2024
Posted:
19 February 2024
You are already at the latest version
Abstract
The theory of water transport in trees, according to which the main driving force of water movement is the suction created by the evaporation of water by the meniscus (the curved surface of the capillary liquid column) on the evaporating elements, supported from below by root pressure, is flawed. The capillary effect in nature is around one meter. In the case of open-air gaps, the leaf cannot suck in the water against gravity because, in this case, the plant would not be sucking in water, but air through the open-air gap. If not, how do trees transport water? When evaporation occurs, there is no suction towards the canopy, but pressure is due to a reduction in the cross-section of the water pipe caused by heat loss through evaporation. At night, when evaporation stops, a thermal equilibration process is triggered, which restores the original cross-section of the pipe. This generates suction and draws water from the soil. As the hydrostatic pressure in the pipe is high for tall trees, the pipe is cut off. To prove my theory, I have given several examples. Water transport is not based on physical mechanical laws alone. There may be complex physiological, biochemical, and biophysical processes behind the operation of the pipe system.
Keywords:
plant water transport
; suction-pressure pipe
; pulsating system
; different time-phase cycle
; aquaporin
1. Introduction
Over the past seven years, I have published several articles on the electronic news portal Forest Press on the subject of water delivery to trees. In the present article, I am also looking at water and nutrient transport and the evidence for it. I also discuss two concepts that do not exist in trees: water potential and root pressure.
From plant anatomy, we know quite well that the dividing tissue of the cambium gives rise to the fascicles, which are dedicated to different tasks, including the cells specialized in water transport, which are transformed into water transport tubes by longitudinal interconnection. Towards the outer sheath of the tree, they form chain cells that transport nutrients. In the following, I want to deal with water transport when water moves in the so-called xylem elements. In vascular plants, the xylem, or woody part, is one of the two transport tissues of the phloem, or limpet part, is the other (Figure 1).
2. The Current Theory and Its Refutation
The theories of water transport that have been studied so far differ only slightly, with no substantial differences. The main driving force for the movement of xylem sap is the evaporation of water from the surface of the mesophyll (leaf litter cells) into the atmosphere. Vaporation causes the walls of mesophilic cells to form millions of tiny meniscus (the capillary fluid column curved surface of the capillary). Their surface tension exerts a negative pressure or pulling force on the xylem, which pulls water up from the roots and soil. Root pressure is commonly cited as the downstream driver of water transport. If the water potential of the roots is more negative than that of the soil (more precisely, the soil solution), water can usually enter the roots from the soil by osmosis due to the high concentration of solutes. This creates a positive pressure that moves the xylem sap towards the leaves. This phenomenon is known in the literature as root pressure. For a comprehensive overview of water transport, see the following article [1].
Figure 1.
Cross section of the tree trunk.

The question arises whether water transport in the tree can work as described. A system will work as intended if it has a functioning mechanism, if there is a matching force of sufficient magnitude, acting constantly, and if the structural elements can withstand it.
Let's look at each of these separately.
One of the forces mentioned is capillarity, or the capillary effect, the property of a fluid to move in a narrow, confined space against the force of gravity. If the diameter of the tube is small enough, the combined forces of surface tension and adhesion forces between the tubes will lift the liquid into the capillaries of the plant. However, the capillary rise in nature is around one meter, and water in capillaries cannot rise higher than this. The root pressure (pushing force) from osmosis can be small or large, it doesn't matter, it is not suitable for continuous water delivery because it needs to be worked to keep the water flowing. This requires a mechanism to ensure that the force can move constantly on the way.
The system mechanism is flawed. The only way to deliver water is to squeeze the water out of the system. So there is both suction and pressure. The delivery of water from trees can be compared to the operation of water pumps because similar systems work in a similar way. There are two types of these pumps in terms of the way they deliver water.
At the same time phase suction-pressure pumps are for example gear pumps (Figure 2).
The other is the differential time-phase pump, best known as a suction-pressure well, or the laterally driven piston pump (Figure 3).
The operating principles of the two types of pumps are described in more detail below. Water cannot be transported (this is also true for organic systems) by suction or pressure alone. Nor can water be transported by pushing at the bottom and sucking at the top.
In the case of evaporation, the pipes must be saturated with water, because evaporation can only take place in this case. Force balance condition for any height. The downward gravitational force must be in balance with the suction force in the opposite direction. For water to enter the leaves at a certain height, a force greater than the suction force creating the force balance is required, depending on the internal resistance. If I want to illustrate this with a practical example, for example, a beech twenty meters high, which can be considered medium height, this means that there must be a suction force in the leaves at twenty meters greater than two atmospheres. With two atmospheres of pressure, you can pump the tires of a mid-size car hard. How the leaf tissue can withstand the reverse, a suction of two atmospheres (two bars)? Not to mention the fact that you can't even create that much suction. The analysis shows that the theory of water transport as understood so far is flawed. It is faulty because there is not enough force in the system to match the mechanism. So there is no guarantee that the force will work and move along the path. The mechanism is faulty because water cannot be transported in this way. The biggest contradiction is that at high altitudes, the leaf tissue is unable to withstand the suction generated.
2.1. Water Potential, the Big Revealer
However, the concept of tolerability is not a category of proof. The untenability of traditional theories is highlighted by the water potential theory. In 1960, an Australian and an American researcher, Taylor, and Slatyer, proposed to use the chemical potential of water as a basis for evaluating the soil-plant-air system, which they argued had a water potential difference (Figure 4).
It was also suggested that the chemical potential of clean water should be considered as zero. The water potential is the chemical potential of a system, or of water in a part of a system, expressed in terms of pressure, of pure water chemical potential of a water system. If the water potential of pure water is the greatest, (0), then the water potential of solutions is less than this, i.e. a negative value [2]. Water diffuses from the higher water potential to the lower water potential, just as heat diffuses from the higher temperature to the lower temperature (Jannes Sutcliffe) [3].
In open-ended pipes (stomachs) there is no suction force and associated suction mechanism, so water cannot flow in the pipes against gravity. This phenomenon will be analyzed in more detail later in the description of pump types. If there were a suction force in the leaves and an associated mechanism, the plant would not suck up water but would suck in air through the air gap. Since the concept of water potential was created to evaluate the soil-plant-air system, it is quite clear that when evaporation occurs, suction continues through the air. So the air sucks water out through the air gap. The problem: we live in a physical atmosphere at approximately sea level. Evangelista Torricelli proved this in 1643, and Otto von Gueircke presented his famous experiment to the princes gathered in Regensburg. In this experiment, he used an air pump he had developed to pump air out of two empty hemispheres. By placing eight to eight horses on opposite sides of the sphere, he found that the sixteen horses were unable to pull apart the two hemispheres, which were forty-five centimeters in diameter. Otto von Guericke was a famous German physicist and the mayor of Magdeburg, and his experiment has gone down in the history of physics as the Magdeburg hemisphere. How much atmosphere did the physicist suck out of the hemispheres? Slightly less than he locked in. Nearly one atmosphere, and then he reached a state close to interstellar space (zero atmosphere). So the maximum suction value that exists in the air under terrestrial conditions is less than a physical atmosphere, but in this case, the air must be enclosed. The lowest sea level pressure of eight hundred sixty hectopascals was measured in the funnel of a tornado on 24 June 2003 in South Dakota, which is not a significant drop from the normal 1013 hectopascals.
Let's put into words what happened. The water potential has failed the conventional theory while failing itself. So there is no water potential. This also made it clear that water can only be released into the atmosphere by pressure when evaporating, as pumps do. The question arises as to how such high suction values could be detected in tall trees. The measurement was carried out with a Scholander pressure chamber (Figure 5).
"This method consists of placing a leaf or a leaf stem in a chamber that can withstand a gas pressure of 5000 kPa, so that the stem or the cut end of the stem protrudes from a locking device fixed to the mouth of the vessel. Then, a gas, such as nitrogen gas, is released into the chamber and the pressure of the gas is increased until the liquid appears on the cut surface of the plant protruding from the chamber. Then equilibrium between the leaf cells and the sap in the wood, and the gas pressure is exactly balances the water potential of the leaf cells. " (James Sutcliffe).
As you can see in the diagram, when the branch is cut, the plant's sap is retracted. The perceived suction value is read when the sap appears. The most fundamental flaw in the measurement method is that when the branch or leaf is cut off, both suction and pressure tubes are cut through according to the conventional theory. According to the theory, the xylem is a suction tube, but the nutrient-transporting phloem's rust tube is a pressure tube. This was found by cutting through the mouthparts of sucking insects under the tip bud of the leader drive and pressing out sugary sap. The conventional theory is that a large suction in the leaf should be converted into a small pressure. Such a mechanism to convert a large suction into a small pressure is not found in the leaf. The water transport researchers are very self-critical in pointing out that with this instrument, despite the fact that many people have even climbed trees with it, they have not been able to detect differences in water potential. The measurements ranged from 0.5 to – 0.8 MPa. Zimmerman modified the device, but the suction-pressure tube anomaly persisted [4]—no wonder the non-existent water potential could not be measured with an instrument based on false principles. The error was apparently due to the fact that the water delivery researchers forgot to tell Scholander that they had cut both suction and discharge pipes during the stem cutting. If not, how do trees transport water?
3. The Principle of Operation of the Water Conveyance System
The leaf surface has small openings (stomata). These air slits, which are mostly located on the back of the leaf, provide air exchange and evaporation. During evaporation the air slits are open. At higher energy levels, water molecules are released into the air. Consequently, the water remaining in the tube cools down. A heat dissipation phenomenon occurs. Since it is an organic system, the cells, which are located above each other and are connected to each other by their walls, respond by a reduction in cross-section. Of course, only the elastic annual ring formed in the last year can do this (this is certainly the case for the sessile oak). In this case, we are not talking about suction, but about pressure towards the atmosphere. The tubes can only shrink up to a certain limit. Once the contraction is complete, the breathing openings close. Evaporation and heat extraction cease. A thermal equilibration process is triggered, which slowly restores the original cross-section of the tubes. This increase in cross-section, in turn, generates suction forces towards the roots, which draws water from the soil. Similar to a medical eye-dropper, when a compressed cylindrical flexible rubber with a narrowed cross-section is released, the liquid is sucked into the tube. As a pressure gradient develops in the tube (the hydrostatic pressure gradually decreases from the bottom to the top), a variable suction or pressure force is required, and the tube is therefore cut off. Thus, the movement of water can be achieved with a force of nearly equal magnitude, but also with a smaller force depending on the number of cells that are cut off. This disconnection results in a peristaltic movement. Another important aspect of peristaltic motion is that this form of motion ensures that the system, actuated by a lateral force, can move along a constant path. Therefore, permanent work is also possible.
The mechanism for closing and opening valves (the operation of the system is illustrated in Figure 6).
In the first column of Figure 6, you can see that the plant is pushing water from segment number two into segment number one above it. (Fny = pushing force, Fsz = suction force, Fe = counterforce, which consists of several forces and is therefore not shown.) The valve opening condition Fny is greater than Fe. In the pushing process, the counterforce must increase in order to close the valve. The moment Fny is less than Fe, the valve can close. Pressure can then be applied from segment 3 to segment 2 above, which has been reduced by the previous pressure. The results are illustrated in the second column (B). The precondition for opening - and closing the valve is the same as before. This process is repeated over and over again, and the peristaltic motion shown in Figure 6 is obtained.
The mechanism of uprooting at night is illustrated by the two columns on the right in Figure 6. Here, the suction starts from the first segment of the third column (A), which sucks water from segment 2 below. The suction continues as long as Fsz is greater than, Fe. The moment Fe is greater than Fsz the valve closes. The rest of the suction process is similar to that described for the pressure process. Here too, a peristaltic motion is formed, the result of which is shown in the fourth column (B).
In the pressure process, all segments become smaller and smaller during the day, which assumes that there is a shut-off valve at the bottom at the beginning of the pulsation. At night, the valve at the bottom opens at the upstream suction and the whole system becomes thicker and thicker, sucking in water. Figure 7 illustrates the peristaltic movement of the xylem tube during the pressure and suction stages.
An explanatory note below the picture text now summarizes the movement events, complete with the visual image.
3.1. How the Water Pump Works?
A peristaltically pulsating suction-pressure system is important, the similarly operating water pumps principle of operation of the same function. Rajkovits Zsuzsanna, a physicist, describes the sinking and rising of a snail octopus, which J. Verne's imaginary submarine the Nautilus. (J. Verne's imaginary Nautilus was known as the scientific name of the snail octopus [5].) The basic law of the bionics principle, similar systems work in the same way, one assumes the functional similarity between pumps and water pipes. As it have already mentioned, two types of pumps are known [6,7]. One is the same time-phase suction-pressure pump. Such as the ordinary well pump or the gear pump (Figure 2). These pumps are characterized by the continuous flow of water through the inlet and outlet openings, and both ends of the pump are open. In such pumps, the water flow is ensured by a rotating mechanism between the inlet and outlet, which divides the system into two parts. Between the inlet and one-half of the rotating mechanism is the suction, and between the other half and the outlet is the discharge. The other type of pump is where the suction and discharge are not in the same time phase. Examples include the commonly known suction-pressure well, or the less well-known piston pump, which is operated by a lateral force similar to a water delivery pipe(Figure 3). These pumps are characterized by the fact that they draw water into a tank. In this case, the valve between the tank and the suction pipe opens, but the valve between the tank and the air space remains closed. The next step is to squeeze out the water sucked into the tank. The valve opens at the suction is then closed, but the valve between the air space and the tank is opened. The result of this process is that the end from which I suck is open, and the end to which I suck/tank/tank is closed. This is the suction phase with the suctioned water section. And in pressure, where I push, the end is open, where I push/tank/ from, the end is closed. The conventional theory is that during evaporation, the flow between the inlet (root) and the outlet (breather) is continuous and both ends of the pipe are open. This is almost reminiscent of the operating principle of a pump with the same time phase. But where is the rotating mechanism that produces a suction at the bottom and a pressure above? As far as I know, there is no such thing in the plant. Then the plant still sucks water into a tank by closing and opening valves in a similar way to the differential pumps.
Figure 7.
A pulsating system.
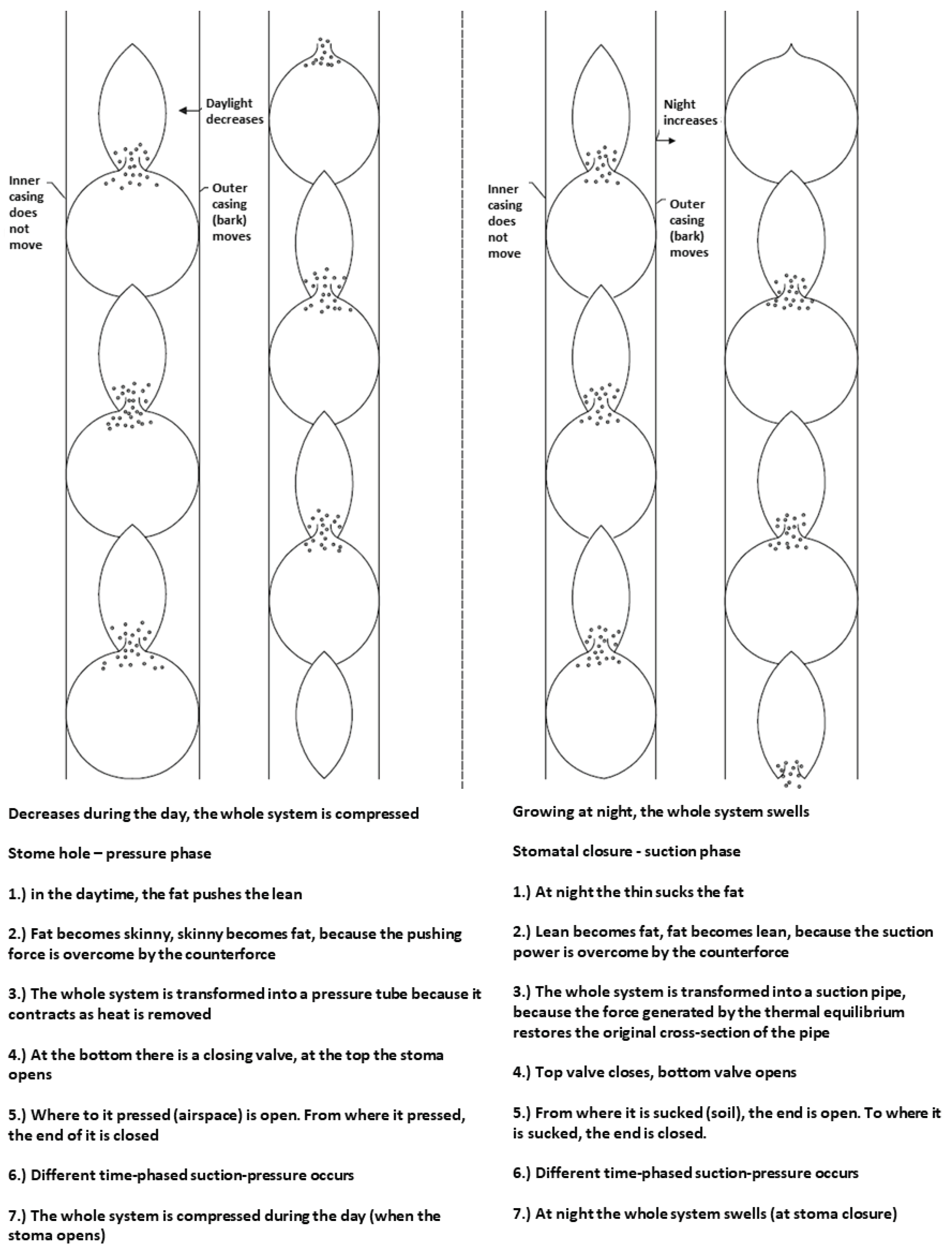
Figure 8 illustrates the movement of water during the pressure and suction stages. In the pressure stage, both the piston pump and the water delivery pipe have the upper valve open and the lower valve closed. In both cases, the water volume of the tank is reduced. In the suction stage, the situation is reversed. Here both the organic water pipe and the piston pump draw water. Therefore, the lower valves are open but the upper valves are closed. In both systems, the tank water volume increases. As shown in the diagram, the operation of the water hose and the piston pump requires power, a variable volume tank, valve, and mechanism. The latter ensures that the force can move on its way. Force in an organic system is provided by the presence or absence of evaporation. In the case of a piston pump, this is provided by the power source rotating the wheel, (manually or mechanically), to which an eccentrically mounted crankshaft is connected. The mechanism is provided by the pulsating motion of the organic water pipe and the piston motion of the piston pump.
Where are the tanks and where are the valves?
The question is raised for the organic system because it is obvious for the piston pump. One tank is the volume difference due to the expansion and contraction of the water transport system. I can prove this with an instrument I have developed and also visualize the movement of the water. The principle of construction and operation of the instrument is as follows: a tight-fitting rubber tube is inserted into a bicycle tube filled with water, and a capillary tube is inserted into a hole drilled in the rubber tube. This structure is clamped to the tree by means of four screw spindles and two Plexiglas plates so that the two flexible systems (the tree and the water-filled bicycle stick with the capillary tube) are placed between the two rigid Plexiglas plates. I then place the water nozzle in the middle of the capillary tube using the wing nuts on the screw spindles, taking care to keep the two plex sheets parallel. This allowed me to follow the bidirectional movement. The pitch of the screw spindle is one millimeter. The displacements in the capillary tube were linear during calibration. At 90 degrees of rotation of the wing nuts, the dragonfly pitch changed four centimeters, at 180 degrees eight centimeters, at 270 degrees twelve centimeters, and at a full 360 degrees of rotation 16 centimeters. Thus, for a one mm change in plant size, the vertical increase in the water level was 16 cm. The instrument converted the horizontal movement of the tree from an actual change in diameter to a vertical movement. In the case of thick bark trees, the changes in tree diameter presumably do not follow the changes in the diameter of the water pipe because their movement is absorbed by the elastic strap. My measurements showed that during the night suction phase, the tree diameter increased and the water intake crept up the capillary tube. During daytime evaporation, the displacement was in the opposite direction. It should be noted that the length of the day in plants is from the opening of the air gap to the air partition. Often in the early afternoon on hot summer days, the plant closes its respiratory aperture. It switches to night mode. I have also experienced this because I have had the water intake move in the opposite direction at noon, so the diameter of the tree has increased because it has been sucking water. The early afternoon air gap closure is well known among plant life scientists. I will discuss later that suction is a slower, energy-saving process. Hence, frequent daytime aeration is an important factor in plant water management. However, there is another reservoir in the tree, the existence and location of which have been detected by a research team organized by Csilla Béres in the Flatwoods project, using a portable computer tomography (CT) and a high-resolution magnetic resonance imaging (MRI) scanner (Figure 9)
The image shows a three-dimensional CT image of the trunk of a sessileoak. The figure is very informative because it tells us that the pulsation system is forming water reservoirs inside the trunk by means of channels. The image shows a very narrow circular ring that is permanently connected to the canals and the water reservoirs. The article by Csilla Béres also shows that the plant can use water from the water reservoir, the stored water, and return it to the active pulsating phase. It follows that there is water movement inside the tree even in the already dead part. The movement is presumably controlled by the pulsation system. The experiment also involved placing isotopes in the tree and using scintillation detectors at different heights to follow the isotope path. They found that the flow was not uniform. In addition to the fast flow, there was a much slower flow, and this persisted throughout the night. At night the air vents are closed. Csilla Béres also noted that the pull of evaporation cannot explain this phenomenon. It is also interesting to note that the rapid flow in the tracheae formed in the very narrow outer band of the tree in the last year of in the last year of the tree. They called this free water, and the water in the canals and water reservoirs bound water, either to a material or a structural element [8]. Given the fact that the deposition and extraction do not occur at the same height, we can conclude that water flow against gravity can also occur in the already dead wood. The mechanism of storage and evaporation is completely unknown in the world of physiology. At the end of the last millennium, water transport experiments on Sikfőkút were a milestone in physiological research, involving several famous institutions and universities were involved [9]. Without being exhaustive ATOMKI Budapest, Hungary, DOTE Debrecen, Hungary, University of Marburg, IATA Florence, University of Forestry and Woodworking Sopron, Hungary. The research in Síkfőkút has been reported in several articles by the institutions listed [8,10,11,12]. Another important question is where the valves are. The valves are located on the surface connecting the ridge walls of young one-year-old cells. Their existence has long been known (called transmembrane protein), only their function was unknown. Aquaporin, the membrane water channel, was discovered in 2003 by Peter Agre and Roderick MacKinnon. (They were awarded the Nobel Prize in 2003.) This transmembrane protein molecule allows valve closure, and valve opening [13]. It also has the last link in the chain, the valve, without which the pulsating system would not work (Figure 10).
3.2. The Mechanism of Nutrient Transport
The physiological literature explains the mechanism of nutrient transport in many different ways. The best-known theory is that there is a potential difference between the place of loading and the place of use, based on the phenomenon of osmosis. This results in a turgor pressure gradient, which drives the sap by mass flow. However, calculations have not always confirmed the existence of a gradient. The probable reason is the high internal resistance. Other mechanisms were thought to be involved in the formation of the flow. I theorize that the pulsating motion of the xylem tube pushes the glucose out of the roasting tube through the tiny pores. The nutrient-transporting roasting tube has an inverse motion to the xylem tube. This is illustrated by the following phenomenon: I break off a leaf from a fig tree at the base of the petioles and then turn it vertically upwards towards the sky. Pressing the broken part, latex sap drips out, similar to that of herbaceous plants such as bleeding dovetails or dog milkweed. Then, when I started to cut the leaf down towards the leaf plate, I experienced the same thing. That is, the sap squeezed out by the xylem tube remained pressed inside the space between the segments formed by the two transport beams. The theory is supported by my experiment, which I carried out after dark around the time of the longest day. The flow of sap continued until midnight, after which it stopped. That's how long it took for the sap to incorporate.
3.3. Evidence for the Water Transport Theory
1.) The plane-root isotope experiments have clearly shown that the conventional theory is wrong. In his summary paper, János Attila Tóth mentions that the transport of water by trees has been shed in a completely new light. In particular, his findings were decisive, as I have already described. Upward flow was also observed at night, which is impossible for closed breathing holes. However, some writers explain the night flow by root pressure. This is not correct, because a poplar's stake shoots out even if it has not yet formed roots. Roots are formed only after the shoot has been established. I will explain the physiological reasons for this later.
2.) Figure 11 shows time-dependent velocity patterns in the trunks of different tree species on a day after watering. On the vertical axis, K= 0.206 U0.814, where U is the flow velocity/m/min [10,11,14]. In the figure, the sawtooth shape at the bottom may indicate when water in the still organic xylem tube of the annual ring formed in the last year flows through valves from one cell to the cell above. The pattern in the image reflects the flow of water from cell to cell. As it is a flow in an elastic system, there is no uniform motion. The opening and closing of the cell valves involves the storage and recovery of kinetic energy. In the early afternoon, the flow is decreasing. This could mean, as I have described earlier, that the plant has started to take up water and store it in the woody debris. This is when I noticed a change in the direction of the water intake of my instrument, i.e. the tubes were swollen. There was no condensation in the nylon bag on the branch. That the plant could have done this could be explained by the fact that the xylem tubes had already used up a lot of water and could move it in another direction. This phenomenon is particularly important in thin-corted tree species such as beech. If there is enough water in the soil, the tree does not shed its bark in the open, because it switches to night mode, even several times a day. Moisture circulation does not stop and cooling is achieved by suction from the roots. If there is not enough water in the soil, it will immediately drop its bark in the open. This is the phenomenon of pieling.
3.) The law of continuity in itself proves the theory correct, although Figure 11 shows the image of a function of a half-sine curve. I thought that the nocturnal part of it could be found somewhere. Csilla Béres showed it on her computer before her death in Szombathely in the early 2010s [15]. The other half of the sine curve was also plotted. This was important for me because it gave me the right to invoke the law of continuity. The average speed is higher during the day and lower at night. According to the law of continuity, a high speed has a small cross-section and a low speed has a large cross-section. (This can of course be distorted by the amount of inward and outward movement.) So the tree's casing is moving. Further mobile computer tomography (MCT) measurements by Csilla Béres and the research team mentioned above confirm beyond any doubt that the rate of tree sap flow shows a regular periodic diurnal oscillation and that a 24-hour periodic sinusoidal curve can be perfectly fitted to this rate variation [10] (Figure 12).
4.) One of the most mysterious secrets of tree life: how does life start in spring, how does a dormant shoot in a bud turn into a leafy shoot? Let's not forget that, at this stage, there is neither evaporation nor photosynthesis. In my opinion, in autumn the leaves fall off the tree so that the pressure remains in the flexible transport system. This is easy to see, as water is constantly flowing upwards at suction and pressure. At the end of the winter, some of the living elements are absorbed in the tubes, and thus the water is released from the pressure tube and combines with the sugary compounds next to it. This pressure and excess compound is a prerequisite for life to restart. The diameter of the fold is then reduced. This phenomenon, that one is suspected, was pointed out by my forest engineer colleague Gergely Levente Kökény. His article can be read in the March 2022 issue of Forestry Journal. The article is titled Tree Moisture Cycle, Twitter, and YouTube [16].
5.) The term root pressure is often mentioned in the literature as an important factor in water transport. In my opinion, there is no root pressure. As I described earlier, a one-and-a-half meter long stake can sprout without roots, but only if the last living annual ring is full of life. A prerequisite for the onset of life is the partial death of the last living growth ring, which results in a reduction in the cross-section of the plant. Of course, all xylem elements cannot die at the same time, because then the growth of the plant is not guaranteed. The floodplain foresters store the summer and the willow in a cool, humid place and plant them immediately after delivery. It is true, however, that the delivery system has a variable pressure rhythm. This pressure is highest in the morning and lowest in the evening. There is also an annual rhythm. It is highest in the winter, during the dormant season, when the tree is saturated with water. This is when the shrubs are submerged in water. The inner water reservoirs of the tree are almost saturated. I did an experiment on this in the birch tree in front of my house. In winter, before the buds had set, I cut off a ten-centimeter thick branch and smeared it with a liquid, gel-like wound resin. Water dripped down the cross-section of the bush when it budded, but there were also areas where the wound resin bulged out. An air bubble formed and later burst. This phenomenon also proves that the wood has a large water reserve in winter and that the wood body is under pressure. This daily and yearly pressure rhythm of the transport system therefore applies not only to the root but to the whole transport system. The rootless shoot emergence and the spring initiation of life in the stake can be traced back to the same analogous phenomenon. In some of the tubes, the living elements are absorbed, causing the water to escape from the pressurized tube and merge with the sugar solution next to it. The diameter of the stem is then reduced since the excess material is realized in the leaf shoot. When the branch is cut off, the excess compound flows out of the wood. This phenomenon is observed in walnuts and birch. The phenomenon of guttation is also due to the pressure of the transport system. This is when plant fluid is forced out through the hydatodes on the leaf plate. It is no coincidence that this phenomenon occurs late at night in closed breathing openings when the pressure of the transport system is high. It is particularly common in the tropics, where high humidity is common in crow flies. So it's not root pressure either.
In Figure 13, only the dead water transport cells of various shapes are visible. The negatives of the dead protein molecules across the membranes are clearly visible. There is no need for organic transport tissues inside the tree, but solidification is needed. It is particularly important to achieve high bending strength.
6.) A team of Hungarian, Finnish, and Austrian researchers has used an infrared laser scanner to investigate what happens to tree canopies when night falls. The scientists concluded that the trees were asleep. they used a laser scanner to create a highly accurate and detailed model of the trees they studied, which they repeated every hour during the night. It was confirmed that the branches and leaves of the trees were dropping by up to ten centimeters at night. According to András Zlinszky, a biologist at the Hungarian Academy of Sciences, they reached their lowest position just before sunrise and then returned to their previous position within a few hours at dawn. It is not yet known whether the trees were woken up by the sun or by their own internal rhythm independent of the sun (Eetu Puttonen) [17]. The research team measured the shift in old birch trees The scientific name of the birch is Betula pendula. The name is derived from the Latin word pendere, which means drooping. The branches of the old birch tree hang long. I think that nothing proves the theory more clearly than the sleep movement of trees. At night, as the branches are sucked up from the root, the drooping branches become heavier and heavier from the water flowing into the transport system and into the water tanks, so they keep dropping down [18]. During the day, when the pressure starts to build up, the process is reversed. The reason for the rapid rise is the difference between the daytime and night-time power play. Later, András Zlinszky and his research colleague repeated the experiment with exotic tree species of a few meters in height and a few centimeters in diameter. The measurements were carried out in a semi-enclosed greenhouse in Balatonfüred. They found that, unlike birch trees, these trees move their foliage up and down [19]. This movement may be related to the water saturation of the trees, as they correctly guessed. Csilla Béres' MRI experiment shows that the tree stores water in different places through channels during the night. This causes a continuous change in the bending moment in the cross-section of the tree, causing the branches to move up and down. This has been called the pulse of the tree. That's why it was deduced that not all trees are asleep, but they all have a pulse. I met András Zlinszky several times, so he knew my theory and referred to me as the author who described the flow of water in a tree with different time phases of suction and pressure.
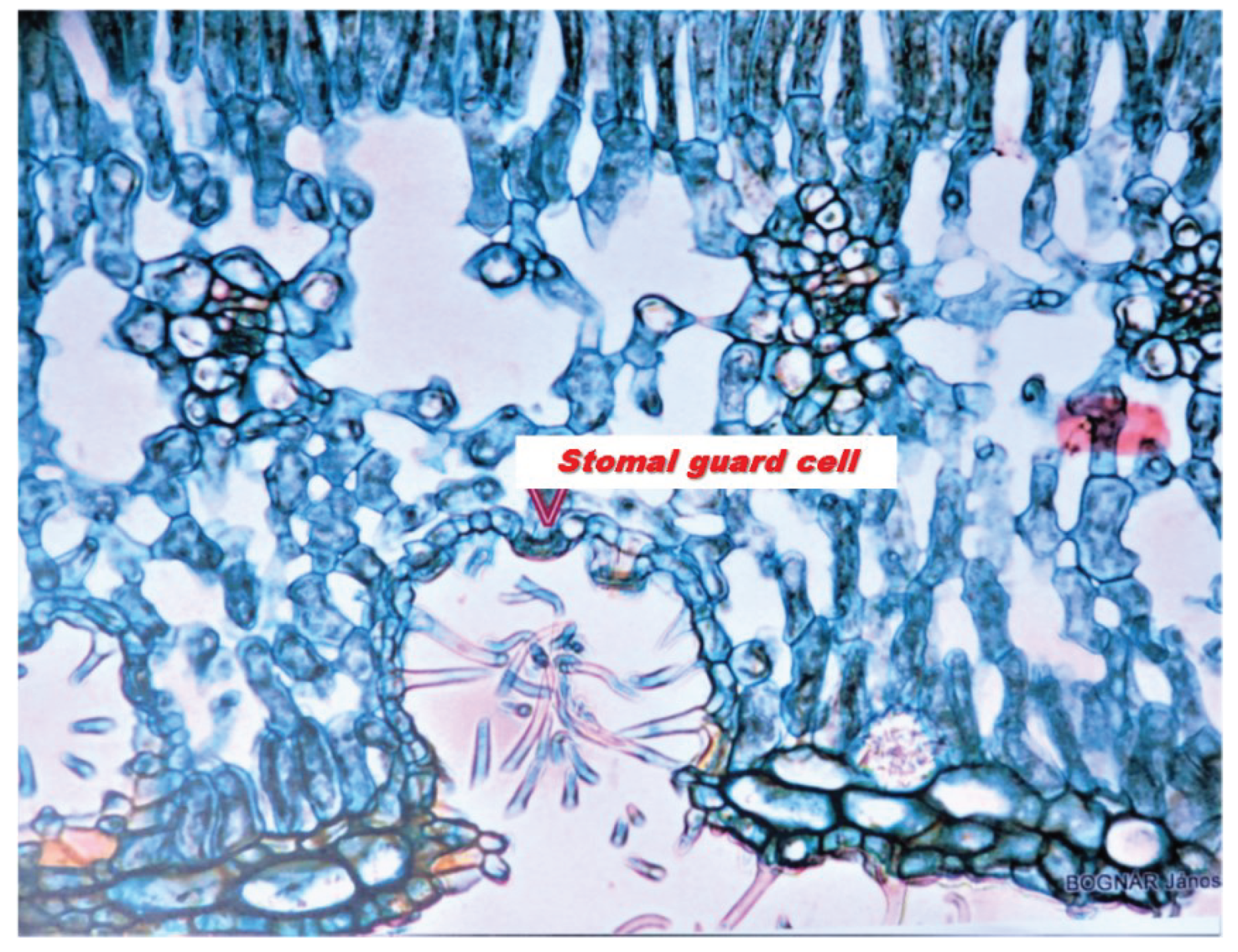
7.) Why do the trees make music? Reading books on plant biology and notes, I have come to the conclusion that there is no plausible explanation for the air exchange in trees. I found an interesting engraving of a succulent plant leaf in János Bognár's plantarium [20].
In the section, the valves were distinctly outlined on the stomatal crypt vault, connected to the spongy parenchyma cells, the double vessel ray, and the columnar parenchyma (Figure 14). The double vessel ray pulsates. As the leaf vessels become thinner, the amplitude of the oscillation decreases, but the frequency increases. The vibration is transmitted through the spongy parenchyma to the vault of the crypt, which it vibrates. This causes air to flow in and out of the valves. . This vibration, caused by the movement of the valves, is converted into sounds that the human ear can hear by an instrument called "music of the plants". This music is played at two bus stops in Buda. You can find it on the Internet by typing the following into your computer's search engine: “the music of the plants at two bus stops in Buda” [21]. I feel that solving the mystery of the trees making music is one of the proofs of the theory.
8.) Fictional evidence. How does a Scots pine absorb water from frozen ground? Forests usually plant two-year-old trained Scots pine seedlings with roots and stems about twenty centimeters long. At this depth, this layer of soil freezes in colder winters. The roots would then have to take up water from the frozen soil, as the pine also evaporates in winter. The skater slides on the water because the pressure and sliding caused by the movement have melted the ice. In my opinion, the pulsating motion is true of the entire transportation system. It may also be true that as you get closer to the thinner roots, the amplitude decreases, but the frequency increases, which may be greatest in the capillaries. The resulting vibration may then be able to thaw frozen soil particles and absorb water from the plant.
4. Epilogue
To conclude, I think it is important to give some thoughts on bionics, as I have compared living and non-living systems in my paper. Bionics is an interdisciplinary discipline that aims to translate solutions developed in living nature into technical practice, based on the consideration that natural selection in nature represents the optimal solution to a problem [22]. The main principle of bionics is that similar systems work in similar ways. The reason is thought to lie in the similarity of their mechanisms. A mechanism is a self-contained symmetrical form of motion that is kept in a symmetrical state by opposing effects. In the case of a water tube formed in the last year of a tree's organic life, several opposing effects can be found: suction-pressure, cross-sectional increase-cross-sectional decrease, warming-cooling, valve opening-valve closing. If one of the listed pairs of opposites is missing, or if the pair of opposites is missing, even in one case, the mechanism of the organic system becomes inoperative. For water pumps delivering water, we cannot speak of so many pairs of opposites. The water pump was not prepared by man for what the plant's water transport system had to be prepared for. They are adapting to a changing climate with changing available water supplies. Therefore the mechanisms of organic systems are much more complex. I don't want to fall into the trap of that the operation of the water pipe is based on physical and mechanical laws alone. . Behind the contraction of water as it cools, complex physiological and biochemical processes may be at work. Likewise, physiological phenomena may also control the storage movements. In this case, the last living ring of living cells stores water in and removes water from lifeless, dead water tubes. The water transport of trees is thus further complicated by the interconnection of the two reservoirs. It was no accident that Ferenc Móra wrote: "the human seed had not yet been sown in the mud of the seas when the trees were already working with pillars and traverses." [23] Finally, just one thought. How do trees transport water? By pulsating peristaltically with different time phases of suction and pressure. How do we humans transport? The same way. By peristaltic pulsations of different time phases of suction and pressure. Perhaps not by accident.
5. Conclusions
The theory of water transport in trees, according to which the main driving force of water movement is the suction created by the evaporation of water by the meniscus (the curved surface of the capillary liquid column) on the evaporating elements, supported from below by root pressure, is flawed. The capillary effect in nature is around one meter. In the case of open-air gaps, the leaf cannot suck in the water against gravity because, in this case, the plant would not be sucking in water, but air through the open-air gap. If not, how do trees transport water [24]?
When evaporation occurs, there is no suction towards the canopy, but pressure is due to a reduction in the cross-section of the water pipe caused by heat loss through evaporation. At night, when evaporation stops, a thermal equilibration process is triggered, which restores the original cross-section of the pipe. This generates suction and draws water from the soil. As the hydrostatic pressure in the pipe is high for tall trees, the pipe is cut off. To prove my theory, I have given several examples. Water transport is not based on physical mechanical laws alone. There may be complex physiological, biochemical, and biophysical processes behind the operation of the pipe system [25].
References
- Bentrup, F.-W. Water ascent in trees and lianas: the cohesion-tension theory revisited in the wake of Otto Renner. Protoplasma 2017, 254, 627–633. [Google Scholar] [CrossRef] [PubMed]
- Taiz, L.; Zeiger, E.; Møller, I.M.; Murphy, A. Plant physiology and development; Sinauer Associates Incorporated: 2015.
- Sutcliffe, J. Plants and water, 2nd. Edward Arnold, London. 122p 1979.
- Balling, A.; Zimmermann, U. Comparative measurements of the xylem pressure of Nicotiana plants by means of the pressure bomb and pressure probe. Planta 1990, 182, 325–338. [Google Scholar] [CrossRef] [PubMed]
- Zsuzsanna, R. Gyűjtemény a fizika interdiszciplináris oktatásához; GlobeEdit: 2015.
- Horváth, B.; Czupy, I. Erdészeti gépek; Szaktudás Kiadó Ház: 2003.
- Csizmadia, F. Géptan; Tankönyvmester: 2005.
- Béres, C.; Fenyvesi, A.; Raschi, A.; Ridder, H. Field experiment on water transport of oak trees measured by computer tomograph and magnetic resonance imaging. Chemosphere 1998, 36, 925–930. [Google Scholar] [CrossRef]
- Tóth, J.A. 40 éves a Síkfőkút Project. BOTANIKAI KÖZLEMÉNYEK 2013, 100, 21–45. [Google Scholar]
- Tognetti, R.; Raschi, A.; Béres, C.; Fenyvesi, A.; Ridder, H.W. Comparison of sap flow, cavitation and water status of Quercus petraea and Quercus cerris trees with special reference to computer tomography. Plant Cell Environ. 1996, 19, 928–938. [Google Scholar] [CrossRef]
- Raschi, A.; Tognetti, R.; Ridder, H.W.; Béres, C. Water in the stems of sessile oak (Quercus petraea) assessed by computer tomography with concurrent measurements of sap velocity and ultrasound emission. Plant Cell Environ. 1995, 18, 545–554. [Google Scholar] [CrossRef]
- Béres, C.; Fenyvesi, A.; Jakucs, P.; Mahunka, I.; Kovács, Z.; Molnár, T.; Szabó, L.; Ditroi, F. Application of an MGC-20 cyclotron and methods of radioecology in solution of problems of forestry and the wood industry. Nucl. Instrum. Methods Phys. Res. Sect. B: Beam Interact. Mater. At. 1989, 43, 101–103. [Google Scholar] [CrossRef]
- Agre, P. Aquaporin water channels (Nobel lecture). Angewandte Chemie-International Edition 2004, 43, 4278–4290. [Google Scholar] [CrossRef] [PubMed]
- Németh László, L.; Béres, C. Terepi mérések a környezeti oktatásban. NYUGAT-MAGYARORSZÁGI EGYETEM SAVARIA EGYETEMI KÖZPONT TUDOMÁNYOS KÖZLEMÉNYEI 2016, 21, 103–114. [Google Scholar]
- László, N.; Béres, C. Korszerű mérési módszerek a környezettan oktatásában. 2015.
- Kökény, G.; Levente, A. Fák nedvkeringée, a Twitter és a Youtube. Erdészeti Lapok 2022, CLVII, 97–100. [Google Scholar]
- Puttonen, E.; Briese, C.; Mandlburger, G.; Wieser, M.; Pfennigbauer, M.; Zlinszky, A.; Pfeifer, N. Quantification of overnight movement of birch (Betula pendula) branches and foliage with short interval terrestrial laser scanning. Frontiers in Plant Science 2016, 7, 222. [Google Scholar] [CrossRef] [PubMed]
- Zlinszky, A.; Molnár, B.; Barfod, A.S. Not all trees sleep the same—high temporal resolution terrestrial laser scanning shows differences in nocturnal plant movement. Frontiers in Plant Science 2017, 8, 1814. [Google Scholar] [CrossRef] [PubMed]
- Zlinszky, A.; Barfod, A. Short interval overnight laser scanning suggest sub-circadian periodicity of tree turgor. Plant Signaling & Behavior 2018, 13, e1439655. [Google Scholar]
- Bognár, J. stoma complex. Available online: http://www.plantarium.hu/2012/07/24/szovettan-gazcserenyilassztomakomplex/ (accessed on 2023. 10. 25).
- Musical chairs – plants inspire sounds at two Budapest tram stops. Available online: https://welovebudapest.com/en/article/2021/12/1/budapest-musical-chairs-plants-inspire-sounds-at-two-budapest-tram-stops (accessed on 2023. 10. 25.).
- Roska, T.; Bálya, D.; Lázár, A.; Karacs, K.; Wagner, R.; Szuhaj, M. System aspects of a bionic eyeglass. In Proceedings of the 2006 IEEE International Symposium on Circuits and Systems, 2006; pp. 4 pp.-164.
- Móra, F. Foreword to the book The Wonderful Life of Plants by Pál Greguss. Available online: https://www.ponticulus.hu/rovatok/hidverok/mora-greguss.html#gsc.tab=0 (accessed on 2023. 10. 25).
- Török, A. Vízpotenciál, a növényélettan fehér akáca (Water potential, the white acacia of plant physiology). Erdészeti Lapok 2019, 154, 186–188. [Google Scholar]
- Török, A. A fák vízszállításának új elmélete (A new theory of water transport in trees). Available online: https://forestpress.hu/hu2/images/pdf/A%20f%C3%A1k%20nedv%C3%A1raml%C3%A1s%C3%A1nak%20%C3%BAj%20elm%C3%A9lete.pdf (accessed on 2023. 10. 25).
Figure 2.
Gear pump.
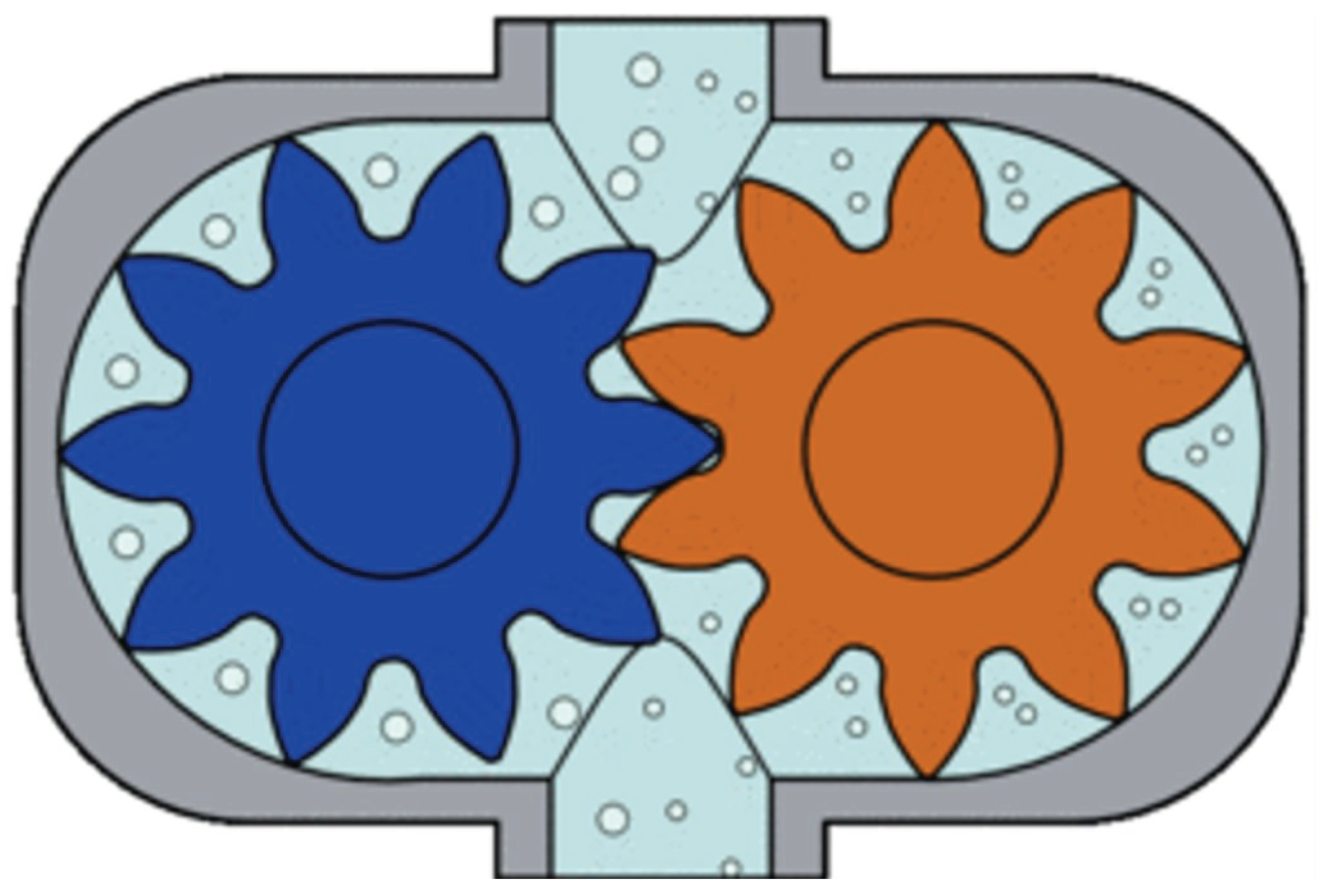
Figure 3.
Piston pump.
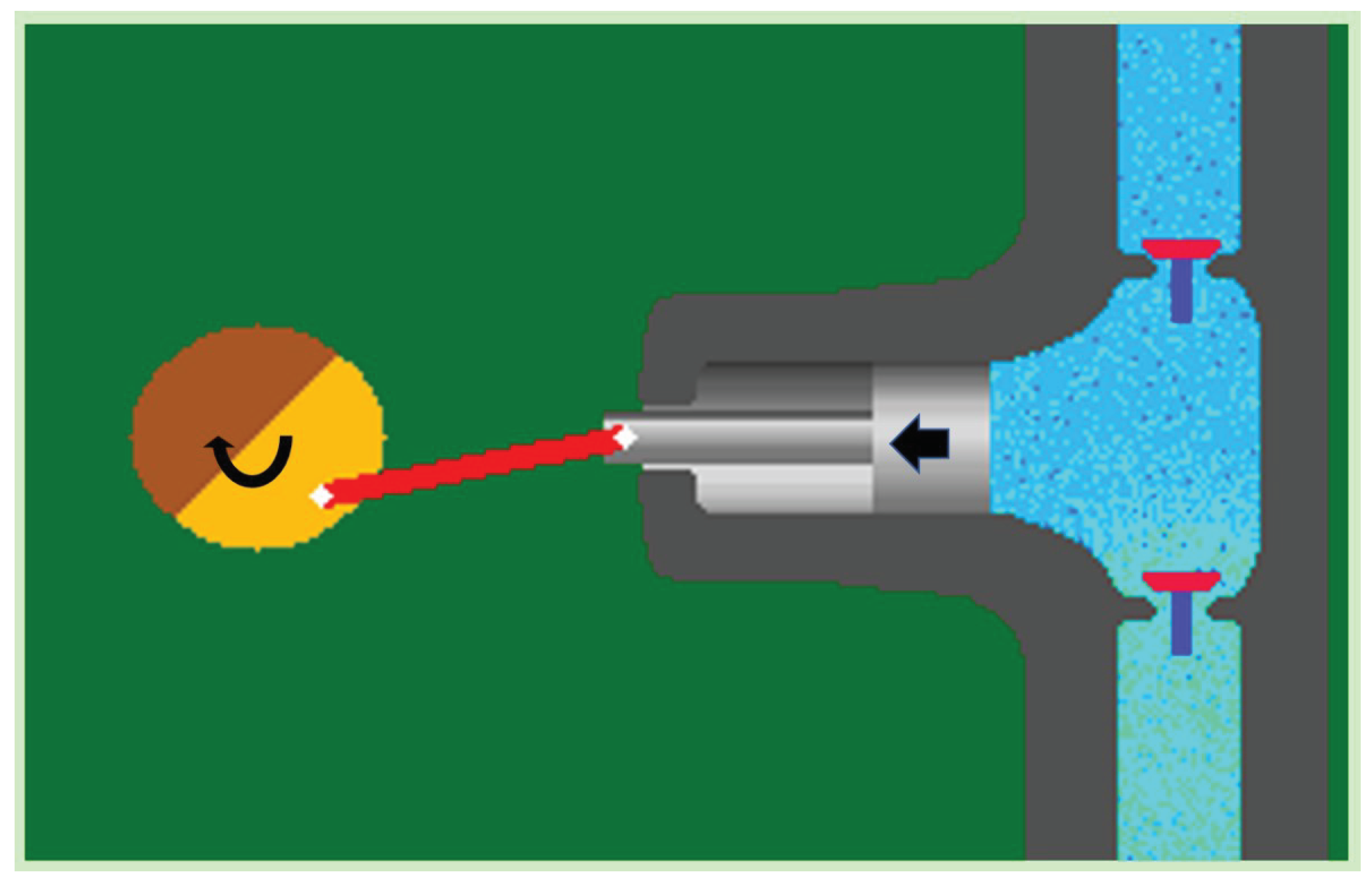
Figure 4.
Water potential difference in soil-plant-air system (https://courses.lumenlearning.com/wm-biology2/chapter/transpiration/).
Figure 4.
Water potential difference in soil-plant-air system (https://courses.lumenlearning.com/wm-biology2/chapter/transpiration/).
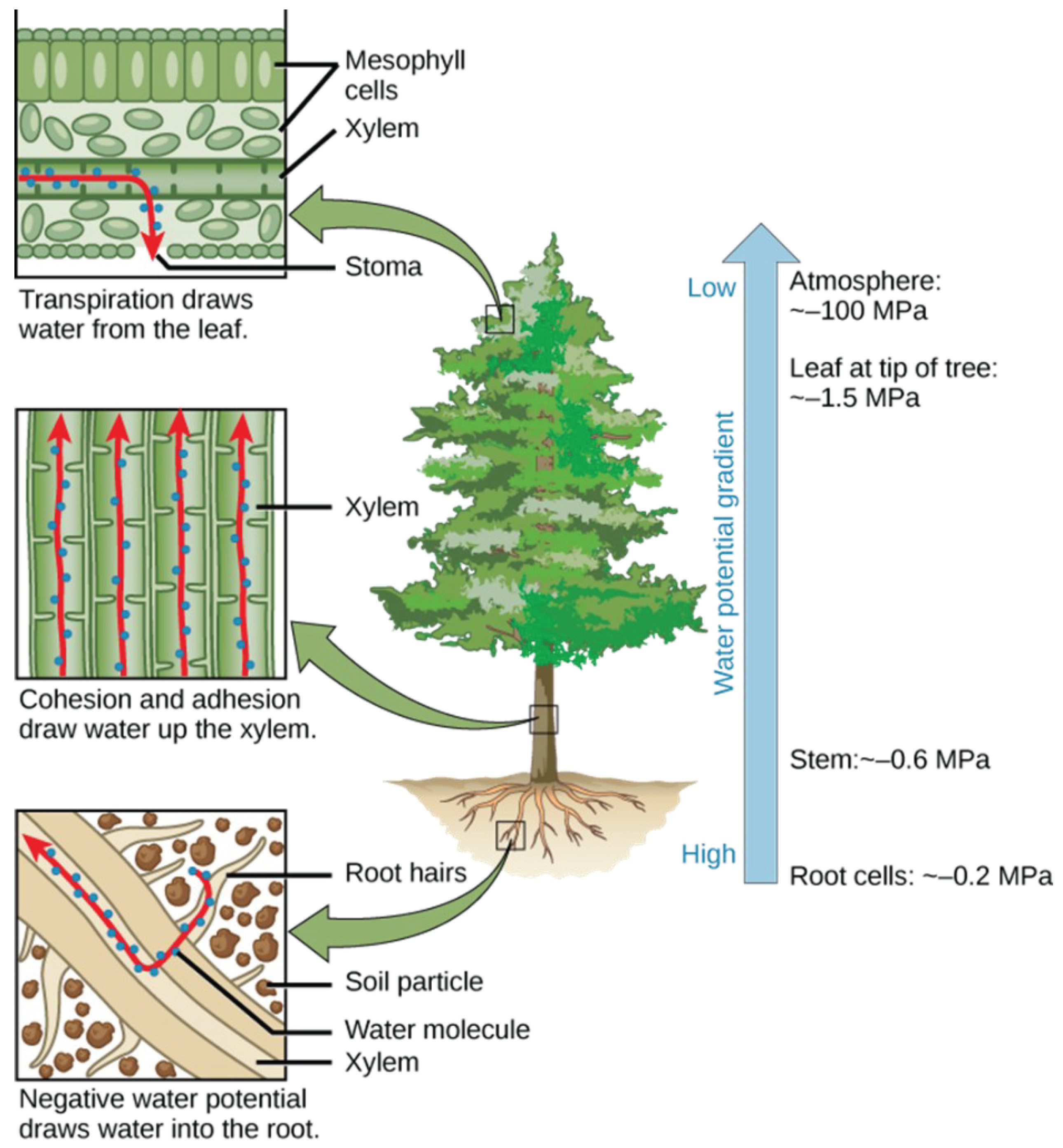
Figure 5.
Scholander pressure chamber.
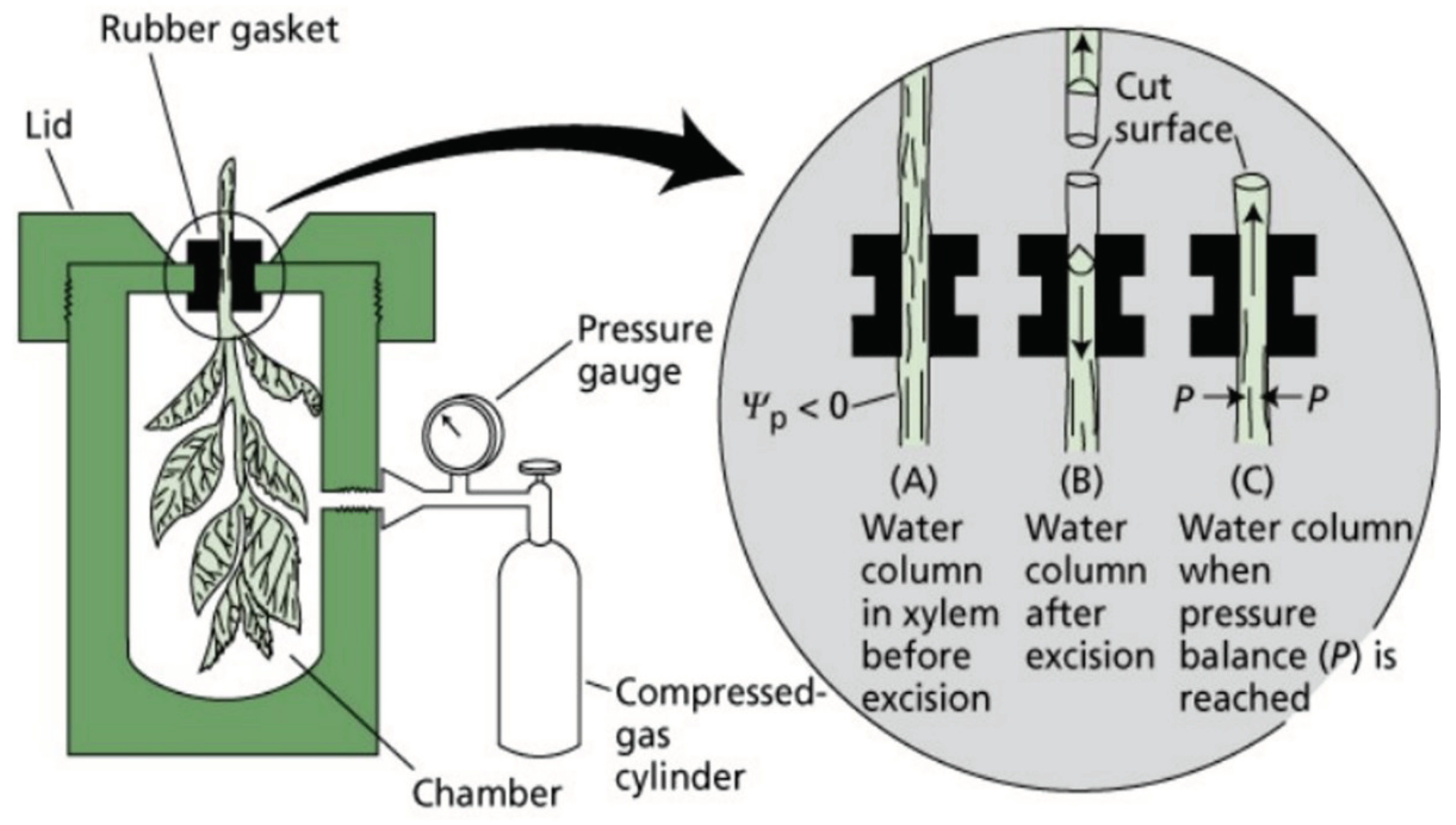
Figure 6.
The pulsating system in the pressure-suction phase.
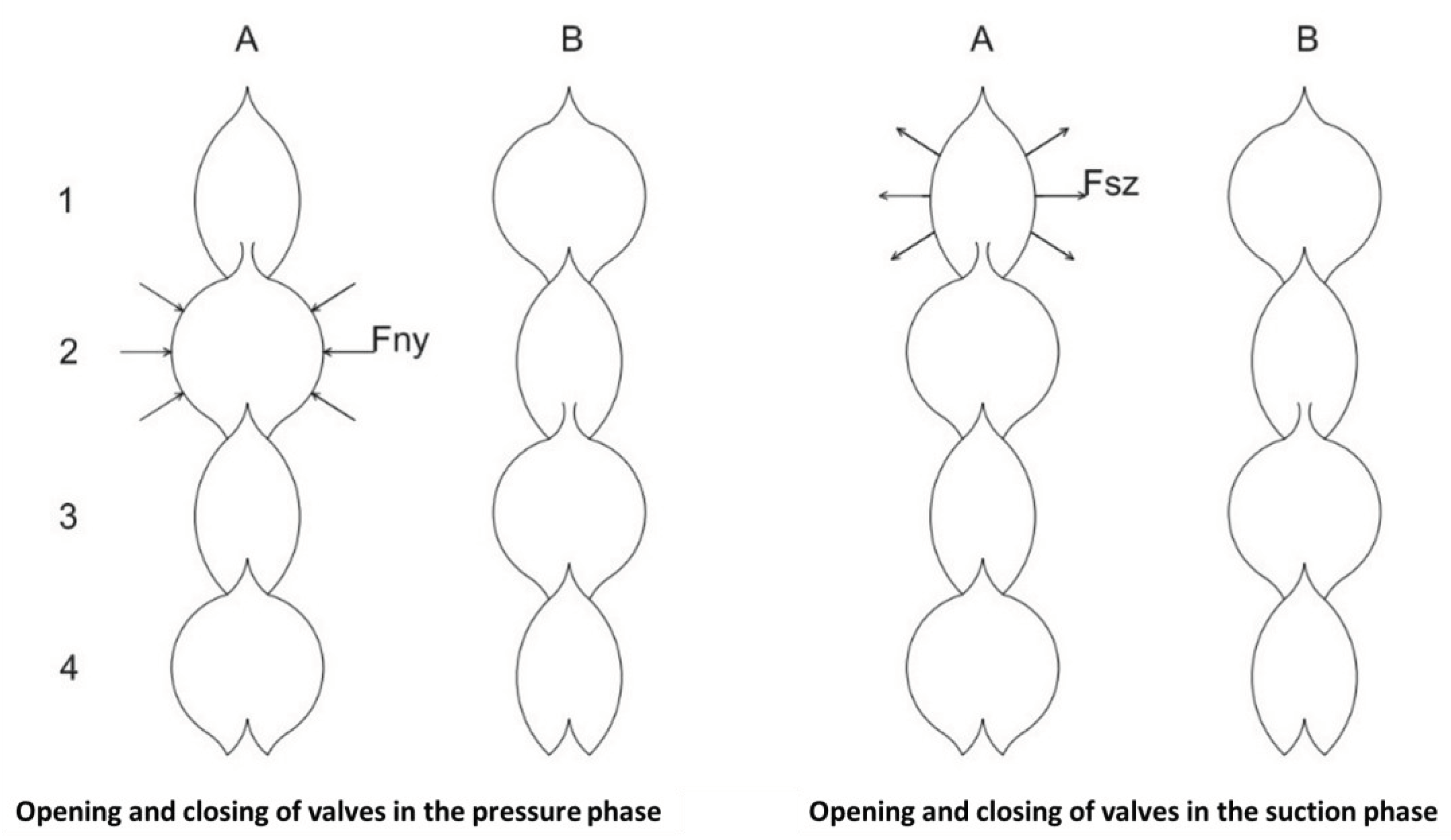
Figure 8.
Water movement in the piston pump and the water delivery pipe, in the pressure-suction phase.
Figure 8.
Water movement in the piston pump and the water delivery pipe, in the pressure-suction phase.
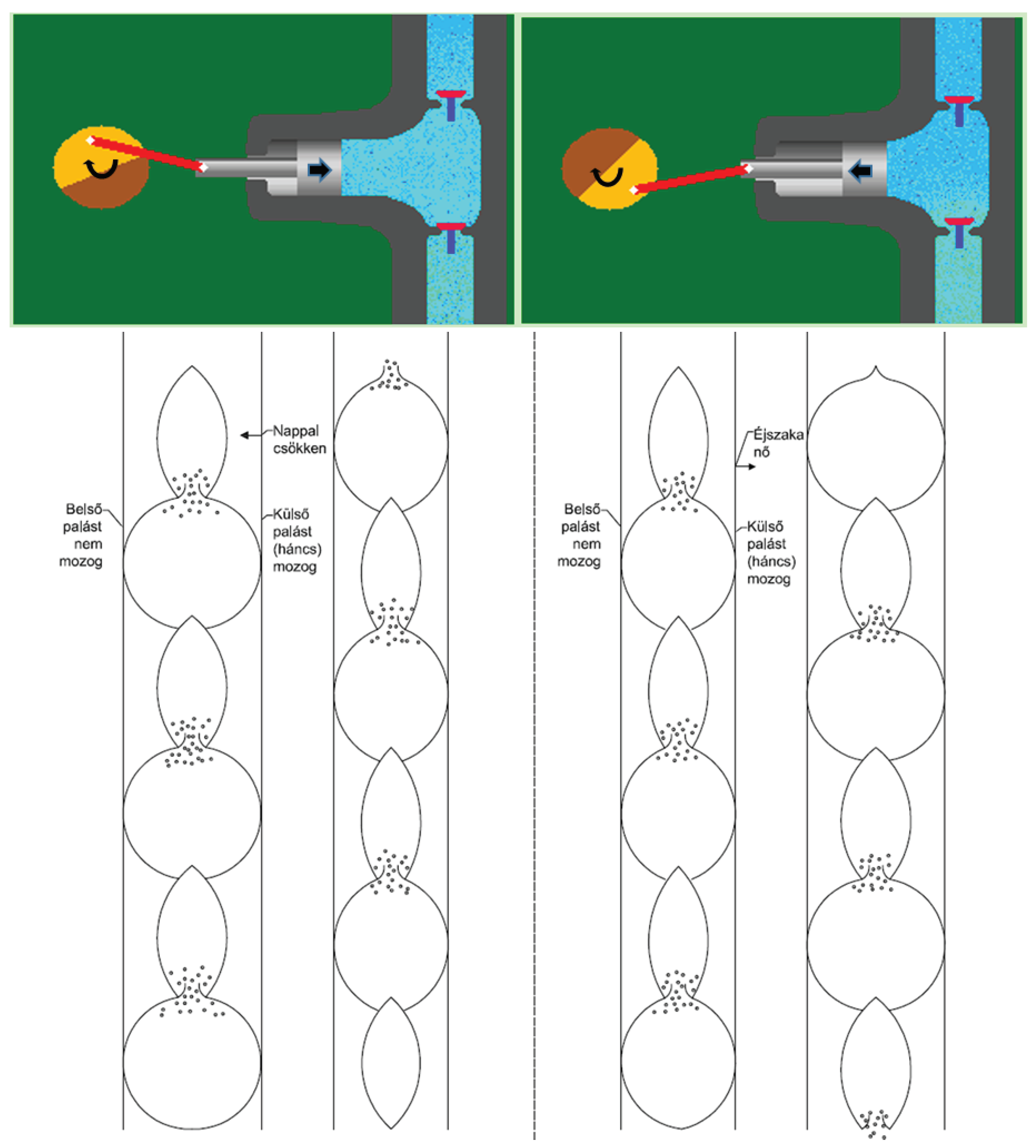
Figure 9.
Three-dimensional image of the trunk of a sessileoak.
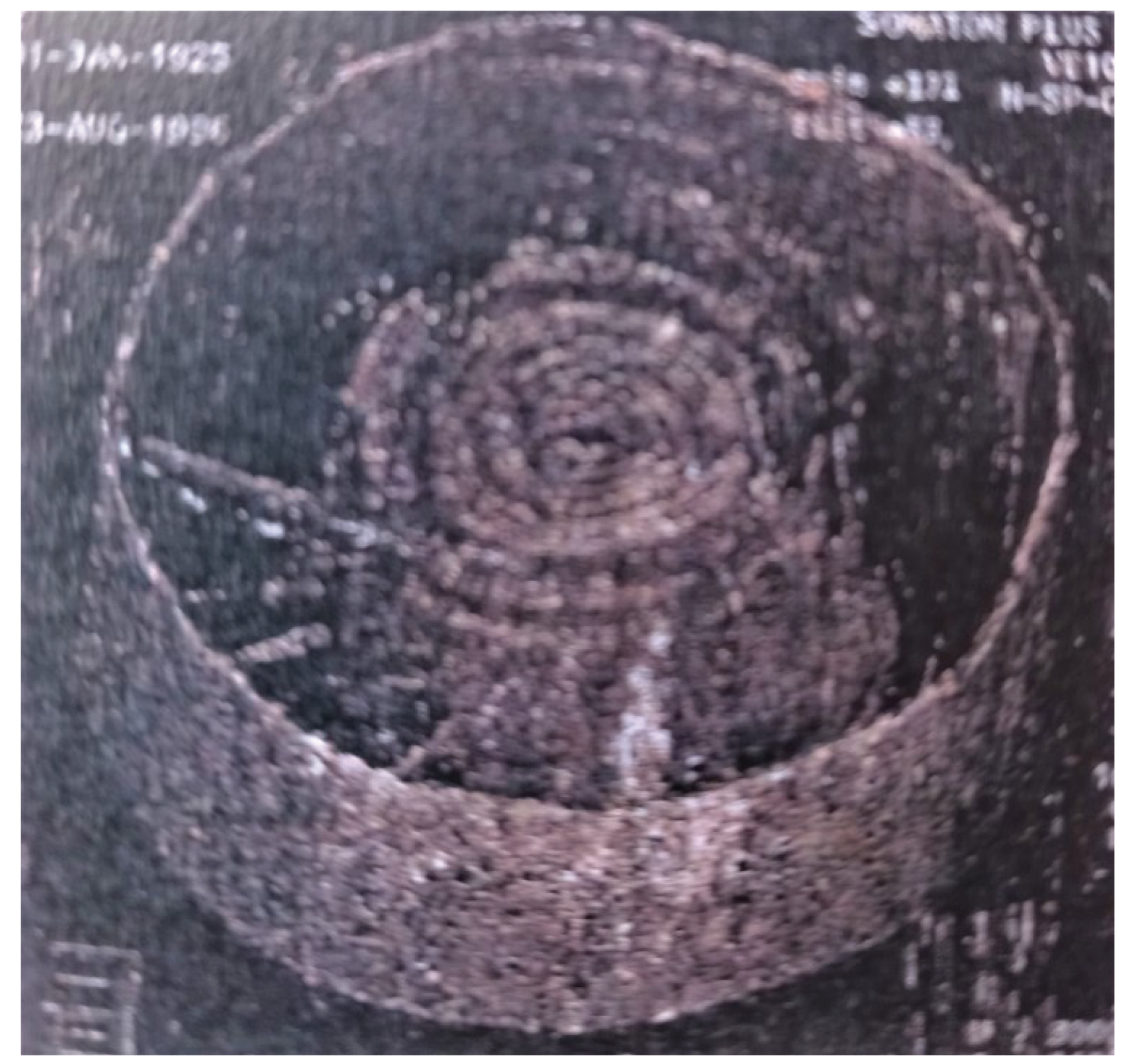
Figure 10.
Biological membrane.
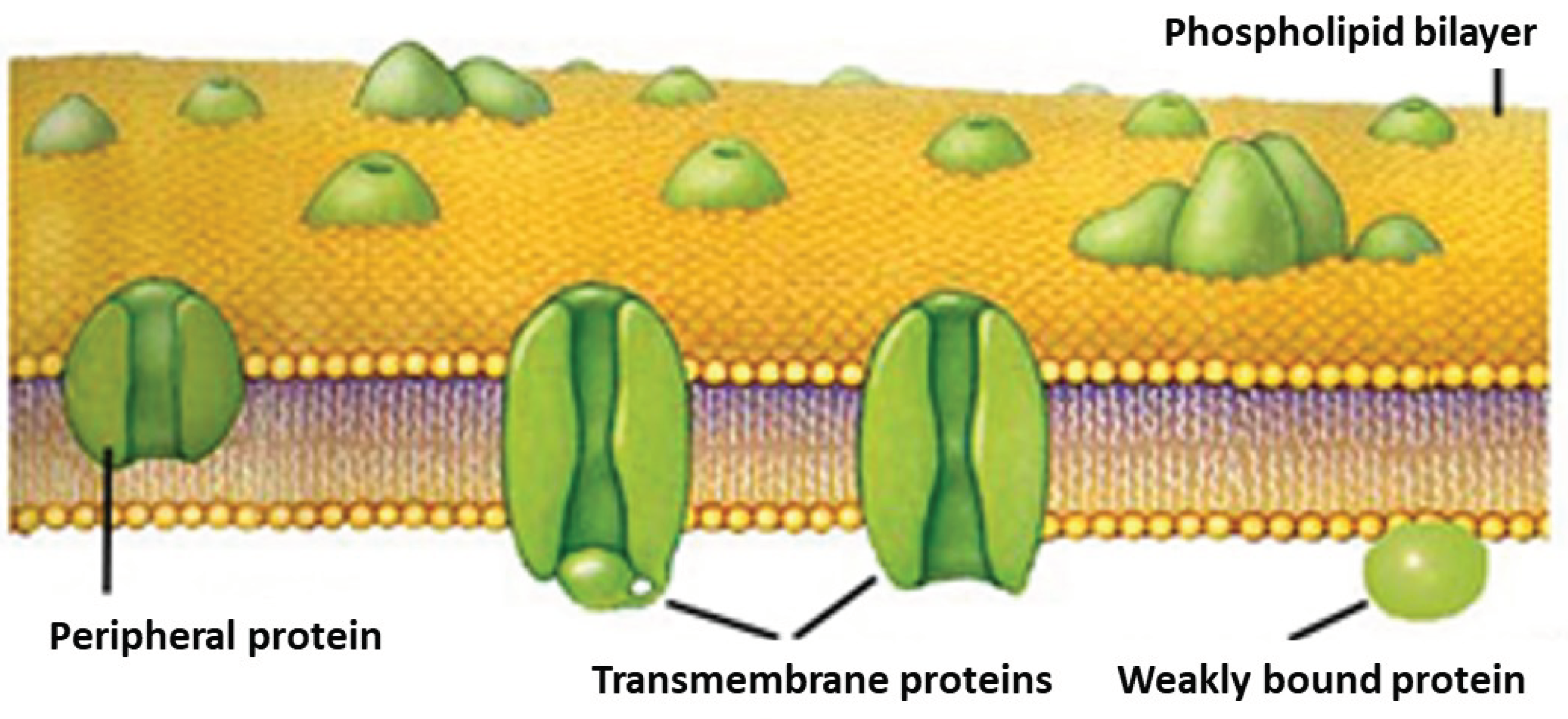
Figure 11.
Water flow measured in the trunks of cherry, sessileoak, and hornbeam the day after irrigation [15].
Figure 11.
Water flow measured in the trunks of cherry, sessileoak, and hornbeam the day after irrigation [15].
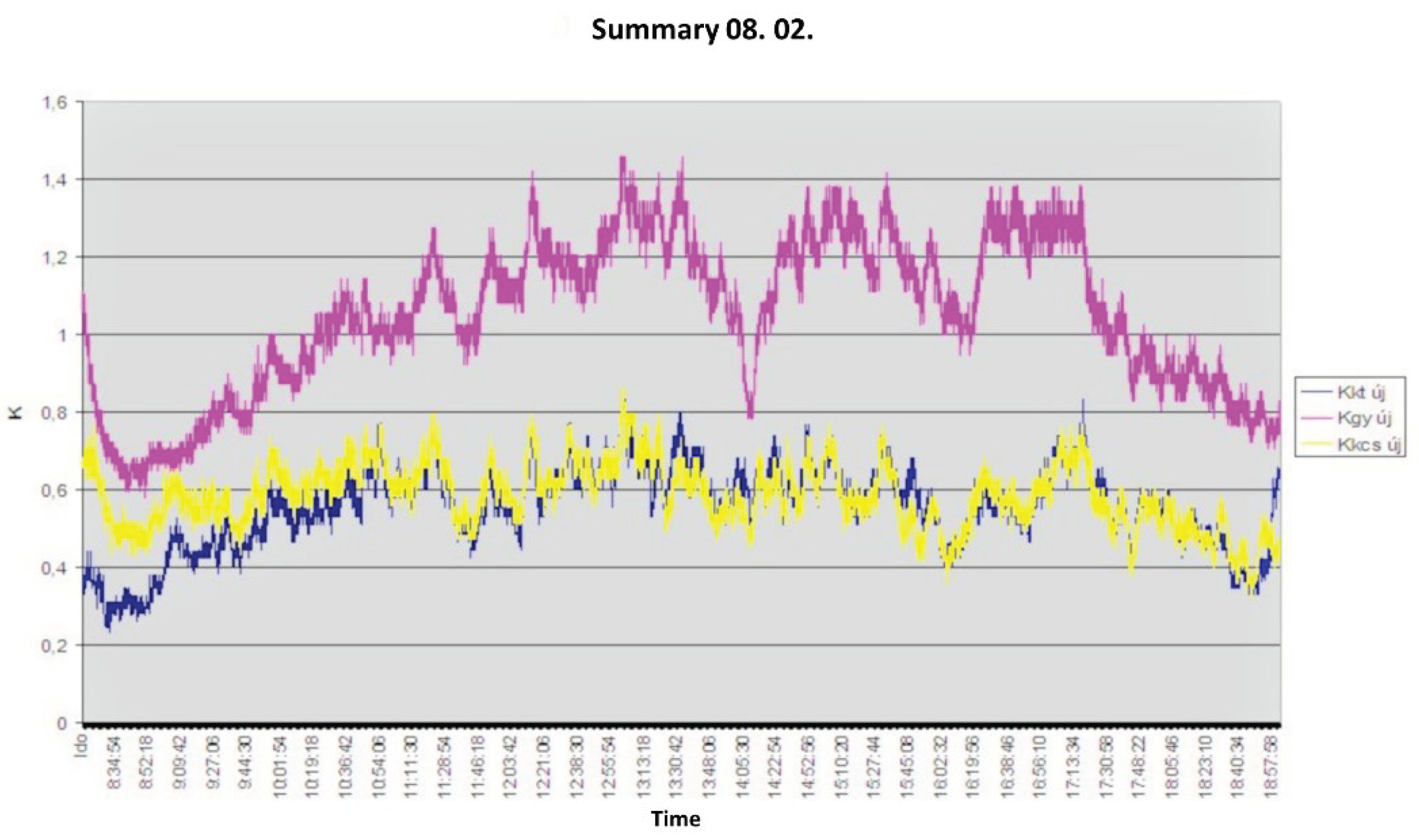
Figure 12.
Daily change of Sap flux of Q. cerris and Q. pertraea measured on 22-25 June 1996 by Tognetti et al [10].
Figure 12.
Daily change of Sap flux of Q. cerris and Q. pertraea measured on 22-25 June 1996 by Tognetti et al [10].
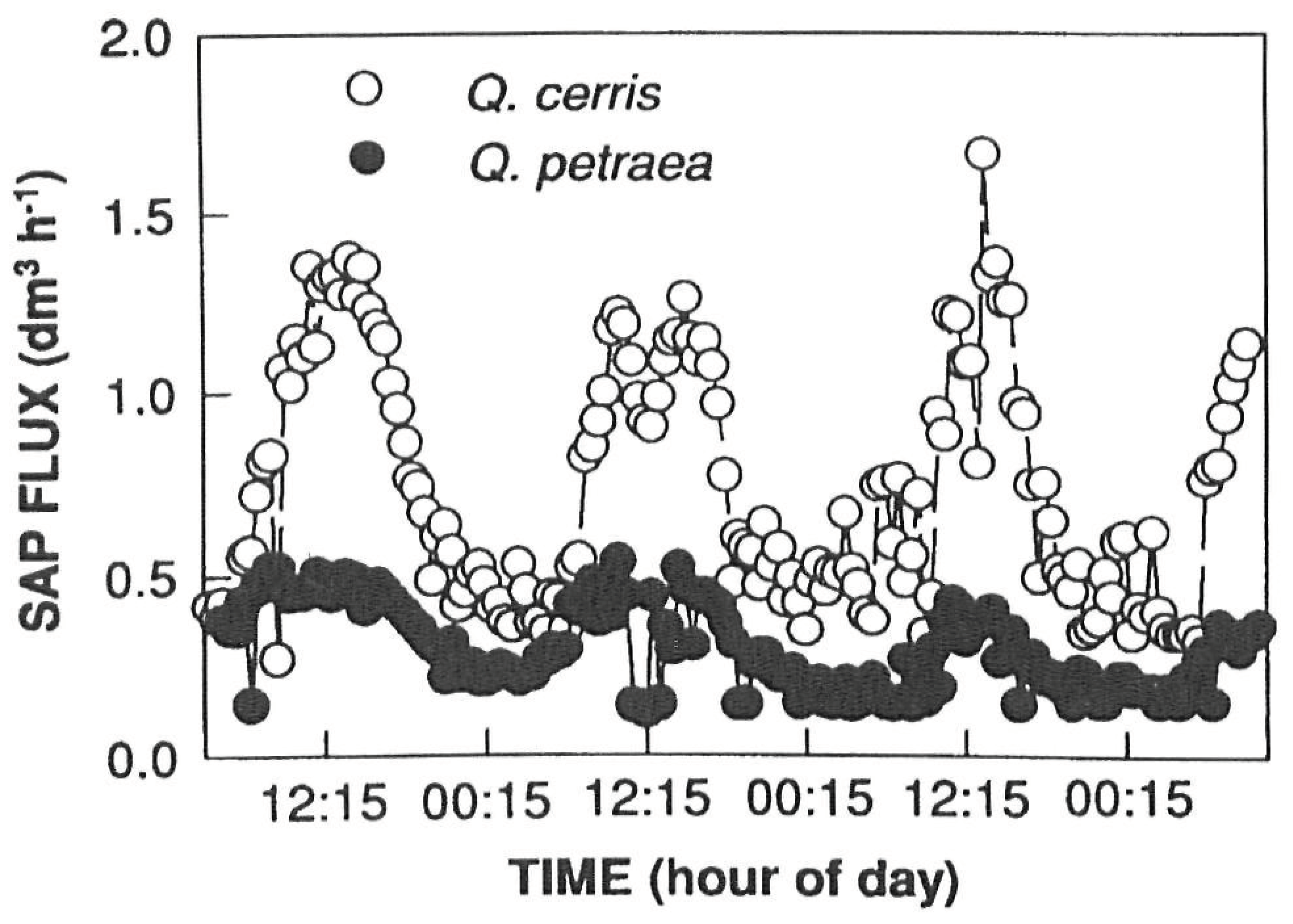
Figure 13.
Xylem elements.
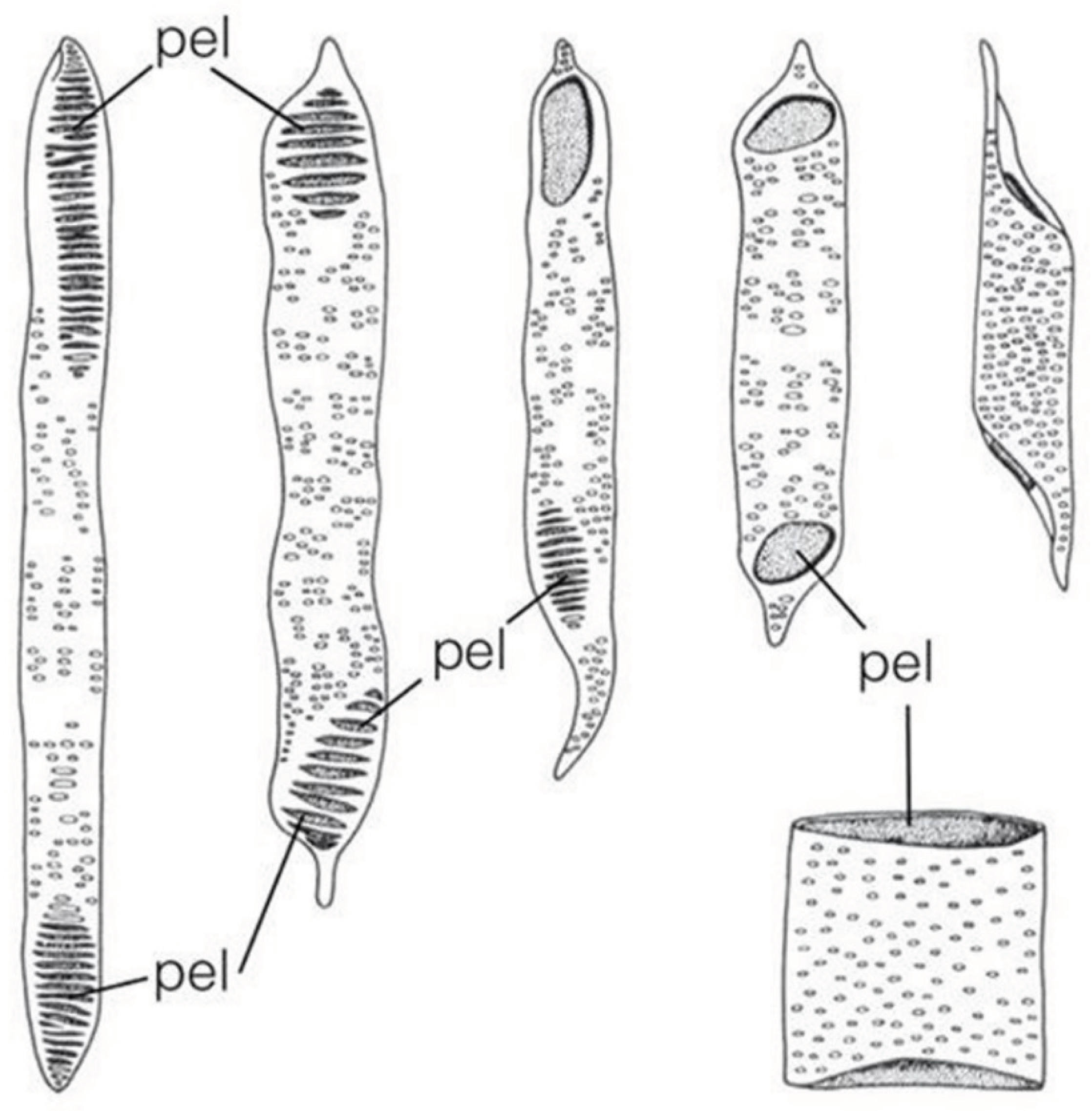
Figure 14.
Stomacript.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



