
Submitted:
17 February 2024
Posted:
20 February 2024
You are already at the latest version
Abstract
Plants can utilize the different forms of nitrogen (N), including inorganic and organic N nutrients, and show the great difference in the utilizing efficiency of N-form among species and genotypes within a species. In our previous study, we found that Tibetan wild barley (XZ16) was better in utilization of organic nitrogen in comparison with cultivated barley (Hua30). In this study, metabolite profiles of the two barley genotypes were comprehensively compared in their response to four N forms, including nitrate (NO3-), ammonium (NH4+), urea and glycine. Macro-and micro-nutrient concentrations in shoots were mostly found higher in nitrate and urea treatments than in ammonium and glycine in both the genotypes. XZ16 had higher concentrations of nutrient ions in glycine treatment, but Hua30 accumulated more nutrients in ammonium treatment. Among the total 163 differentially expressed metabolites, highest up-regulation and highest down-regulation was found in XZ16 in glycine and urea treatments respectively. Some important metabolites such as proline, glutamine, serine, asparagine, L-homoserine, aspartic acid, putrescine, ornithine, 4-aminobutyrate were up-regulated in glycine treatment in both the genotypes with higher fold change in XZ16 than in Hua30. Similarly, fructose-6-PO4, aconitic acid and isocitrate were only up-regulated in XZ16 in glycine treatment. Here, we concluded that the genotype XZ16 exhibited higher response to glycine treatment, while Hua30 showed higher response to NH4+ treatment which is attributed to their better utilization of glycine-N and NH4+-N, respectively. .
Keywords:
barley
; shoot tissue
; genotypes
; metabolomics
; N fertilizers forms
1. Introduction
Nitrogen (N) is a crucial component for crop growth, biomass accumulation and yield formation [1], because it is a constituent of amino acids, which are the elemental units of protein and enzymes. However, nitrogen fertilizer application increases crop yield, often at the rates far greater than the rates at which crops can consume, resulting in a leftover of N in soils [2]. Improper application of nitrogen fertilizers not only reduces crop nitrogen utilization efficiency, but also has the negative impact on the environment [3]. Therefore, precise management of crop nitrogen status are important to improve crop yield and reduce environmental pollution [4].
Nitrogen is mainly available to plants in the form of inorganic nitrogen such as nitrate (NO3−) and ammonium (NH4+), but organic sources such urea, free amino acids, and short peptides are also acquired by plants. These nitrogen sources acquisitions by plant depends mainly on pant habitat and time, soil heterogeneity, microbial community, agronomic practices and other environmental conditions [5]. In aerated soils, NO3− is the most dominant nitrogen source with concentrations range between 1 to 5 mM, while NH4+ concentration usually ranges between 20 to 200 μM [6]. On the contrary free amino acids and urea are usually present in small concentrations, ranging from 1 to 150 µM and <70 µM, respectively [6,7,8]. Urea is the major nitrogen (N) form supplied as fertilizer in agriculture, instead, it is also important N metabolite in plants [9]. The low manufacturing cost and the high nitrogen content of urea (46% N) make this organic compound a common N fertilizer used in arable soils [10]. Currently in terrestrial N cycle, organic N uptake by plants is considered to be a critical component [11]. Before, the uptake of organic N was considered as secondary source of N in low inorganic N soils, such as arctic or boreal ecosystems [12,13,14] but conversely, recent studies on agricultural [15,16,17] and sub-tropical plants [18] confirmed that organic nitrogen uptake accounts for a significant portion of the N budget in plants.
Different N forms can affect the physiological and metabolic processes of plants, such as nutrient uptake, enzyme activity, photosynthesis rate, respiration rate, water balance, and signaling pathways, thus eventually influencing plant growth and crop yield [19,20,21,22]. NH4+ as a sole N source provokes negative effects on NH4+−sensitive plants. The effects of NH4+ and NO3− nutrition on plant growth have been intensively studied; however, the results are not consistent and depend mainly on plant species. Wheat and maize [23], sugar beet [24], beans [25],, tobacco [26] and canola [27], grow preferentially on NO3− nutrition; whereas, rice [28], pine and larch [29], grow preferentially on NH4+ nutrition. Urea fertilizer containing 46% of increased N utilization as compared to other fertilizer sources as well as better than other reduced N sources and appeared to be the best source which gave the highest dry matter yield compared to other nitrogen sources [30,31]. Several studies on Arabidopsis [32], pakchoi [33] and tomato [34] revealed that utilization of glycine could result in high biomass production compared with plants grown under deficiency of N supply. Wang et al. [35] once found that glycine as a nitrogen source influences a series of physiological and biochemical reactions, including plant defense systems, redox homeostasis, amino acids metabolism and protein synthesis.
To understand the adaptive mechanisms of crops to nutrient deficiency, a large number of nutrient-responsive genes or proteins have been identified and characterized through high-throughput omics techniques, such as genomics, transcriptomics, and proteomics [36,37]. Due to the changes in gene transcripts, protein levels, and enzyme activities, metabolites, are regarded as the direct readouts of plant growth or developmental status [38]. Metabolome analysis has been performed to categorize nitrogen deficiency responsive metabolites and their respective pathways, in many crops, such as rice, maize, wheat, barley, soybean, tomato, and rapeseed [39,40,41,42,43,44,45,46]. Previously, a metabolomics study performed with Arabidopsis leaf tissues, found alteration in plant phenotype, and biomass, along with regulation of enzymes and major metabolites under moderate N starvation [47]. Wang et al. [35] found varying physiological responses of the two cultivars to glycine as a nitrogen source and relate it with the up-regulation or down-regulation of glycine specific responsive proteins.
Wild barley (Hordeum spontaneum) is the progenitor of cultivated barley (Hordeum vulgare) and provides a rich source of genetic variations for barley improvement [48]. In our previous findings, wild genotype XZ16 had shown much better growth at urea, and was less affected under glycine-nitrogen treatment as compared to Hua30 in comparison with the plants exposed to NO3− fertilizer. The objective of this study was to find out the shoot metabolites responding to four N-form nutritions, to reveal the specific metabolites differing among N-form nutritions and the two barley genotypes.
2. Materials and Methods
2.1. Germination and Growth Conditions
Seeds of two barley genotypes, wild (XZ16, low-N tolerant) and cultivated (Hua30, low-N sensitive) were surface sterilized with 2% H2O2 for 20 min and then thoroughly washed with distilled water five times. Seeds were soaked for five hours and put in sterilized sand for germination. After germination, light was supplied with fluorescent lamps at 250 μmol m−2s−1 until two-leaf stage. Uniform seedlings were then transferred to 1 L black pots containing modified Hoagland solutions with KNO3 as a sole nitrogen source, and aerated with air pumps. The solution was renewed every day and the pH of the solution was adjusted to 5.8±1. Seedlings were grown in a controlled growth room at temperature 22℃ of 14 h day/18℃ of 10 h night until three-leaf stage, and then treatments were performed.
2.2. Treatment and Sampling
Barley seedlings were treated with four nitrogen forms (NO3-, NH4+, urea and glycine) at N level of 2 mM based on Hoagland solution. NO3- and NH4+ were supplied as KNO3 and (NH4)2SO4, respectively, and K concentration in other three N treatments without addition of KNO3 was adjusted with KCl. Basic nutrient solution for barley was modified to make the nitrogen level same for all the treatments [49]. Shoots were sampled in five replicates after 7 days treatments. Samples were frozen in liquid nitrogen immediately, and stored in a refrigerator at −80℃ for use in metabolomics analysis..
2.3. Elemental Analysis
For element analysis, shoots were collected after 7 days of treatments. Samples were washed with dH2O and blotted on filter paper. To attain a constant weight, samples were dried in an oven at 80°C for 72 hours. Approximately 0.1 g dry samples were used for element analysis. Tissue digestion was carried out in 5 ml HNO3 at 120°C using a microwave digester (MCA 3000, Anton Paar, Beijing, China) and final volume was made 15 ml by adding dH2O to the tube. Finally inductively coupled plasma optical emission spectrometer (ICP-OES, Optima 8000DV, PerkinElmer, USA) was used for mineral concentration determination.
2.4. Metabolites Extraction
Frozen shoots, weighing 10±1 mg were placed into a 2 mL test tube. Then 500 μL of pre-cold extraction mixture (methanol/dH2O (v:v) = 3:1) along with 10 μL internal standard (adonitol, 0.5 mg/mL stock) were added. The samples were vortexed for 30 s and homogenized with ball mill for 4 min at 45 Hz, followed by ultra-sonication for 5 min in ice water. After centrifugation at 4 ℃ for 15 min at 12000 rpm, 200 μL supernatant was added to a fresh test tube. To prepare the QC (Quality control) samples, 60 μL of each sample was taken out and combined together. After evaporation in a vacuum concentrator, 80 μL of methoxyamination hydrochloride (20 mg/mL in pyridine) was added and incubated at 80 ℃ for 30 min, and then derivatized by 100 μL of BSTFA regent (1% TMCS, v/v) at 70 ℃ for 1.5 h. Samples were gradually cooled to room temperature and 5 μL of FAMEs (in chloroform) was added. All samples were then analyzed by a gas chromatograph system (Agilent, Santa Clara, CA) coupled with a time-of-flight mass spectrometer (GC-TOF-MS) (Leco, St. Joseph, MI).
2.5. GC-TOF-MS Analysis
GC-TOF-MS analysis was performed using an Agilent 7890 gas chromatograph coupled with a time-of-flight mass spectrometer. The system utilized a DB-5MS capillary column. Aliquot of 1 μL extraction was injected in a splitless mode. Helium was used as the carrier gas. Front inlet purge flow was 3 mL min−1, and the gas flow rate through the column was 1 mL min−1. The initial temperature was kept at 50 °C for 1 min, and then raised to 310 °C at a rate of 10 °C min−1, finally kept for 8 min at 310 °C. The injection, transfer line, and ion source temperatures were 280, 280 and 250 °C, respectively. The energy was -70 eV in electron impact mode. The mass spectrometry data were acquired in full-scan mode with the m/z range of 50-500 at a rate of 12.5 spectra per second after a solvent delay of 6.30 min.
2.6. Data Preprocessing and Annotation
Chroma TOF (V 4.3x, LECO) software was used for raw data analysis, including peak extraction, baseline adjustment, deconvolution, alignment and integration. LECO-Fiehn Rtx5 database was used for metabolite identification by matching the mass spectrum and retention index. Lastly, the peaks detected in less than half of QC samples or RSD > 30% in QC samples was removed [50].
2.7. Data Analysis
Differentially expressed metabolites were presented by Venn diagrams drawn by online jvenn software (http://genoweb.toulouse.inra.fr:8091/app/index.html). Enriched KEGG pathway mapping was performed by the online database (http://www.genome.jp/kegg/). PCA, PLSDA, and heatmap analysis were done by Metaboanalyst 6.0 (https://www.metaboanalyst.ca). The metabolites with VIP > 1 and p < 0.05 (student t test) were considered as significantly changed metabolites. For element analysis, statistical analysis was performed with statistical software package (Statistix 8.1v) using two-way analysis of variance (ANOVA). The significant difference between treatments was observed using Fisher’s least significant difference (LSD) test at 95 % probability.
3. Results
3.1. Elemental Concentrations
Effect of nitrogen forms is obvious on element concentrations in shoots of the two genotypes (Table 1). Glycine and ammonium treatments were significantly lower in the concentrations of macro and micro elements in shoots relative to the two other N treatments, but the two genotypes showed the different response to the N form treatments, with lower K concentration for XZ16 in ammonium treatment, and for Hua30 in glycine treatment. Similarly Mg was much lower in glycine treatment for Hua30, for XZ16 in ammonium treatments. For Ca, S, and P, XZ16 showed non-significant difference between ammonium and glycine treatments. For micro elements the same trend was found between two genotypes. Urea treatment showed non-significant difference with NO3- treatment for most of the elements in both the genotypes while XZ16 had much higher concentrations than Hua30.
3.2. Metabolomics Profiles of XZ16 and Hua30
Totally 163 metabolites (Table S1) were differentially expressed in shoots of XZ16 and Hua30 among which up-regulated and down-regulated metabolites in each treatment are presented in Figure 1. Highest up-regulation was found in glycine and ammonium treatments for XZ16 (47) and Hua30 (37), respectively (Figure A), while lowest up-regulation was observed in urea treatment for XZ16 (18). Furthermore up-regulated metabolites in Hua30 in urea and glycine treatments were 28 and 29, respectively, while up-regulated metabolites in XZ16 in ammonium treatment were 33. An opposite trend was observed in down-regulated metabolites where highest down-regulation was found in urea treatment in XZ16 (36) (Figure 1B). In detail, down-regulated metabolites in ammonium (24, 23) and glycine (28, 23) treatments were found in XZ16 and Hua30, respectively.
To differentiate between the genotypes and treatments, PCA analysis was carried out (Figure 2E). The PC1 differentiate nitrate (NO3−) and urea samples from ammonium and glycine treated samples, but the PC1 and PC2 did not clearly differentiate the genotypes. Thus Partial Least Squares - Discriminant Analysis (PLS-DA) was carried out. In NO3- treatment purine riboside, galactinol and 13 others differentiate the two genotypes (Figure 2A), while in ammonium treatment d-fructose 1,6-bisphosphate, 3,6-anhydro-d-galactose and 13 others differentiated the two genotypes (Figure 2 2B). Similarly the dominated metabolites in urea treatment were ornithine, linolenic acid, 8-Aminocaprylic acid, and 12 others (Figure 2C). In glycine treatment the dominated metabolites which contributed to the genotypes separation were glycine, allose, glucoheptonic acid, methyl phosphate, D-glyceric acid and 10 others (Figure 2D). Among the top 15 metabolites responsible for genotypes separations, 8, 9, 11 and 8 were found unique in nitrate, ammonium, urea and glycine treatments, respectively. Methyl phosphate was common in nitrate, ammonium and glycine treatments, while d-fructose 1, 6-bisphosphate and glucoheptonic acid were only commonly found in ammonium and glycine treatments.
3.3. Differentially Expressed Metabolites
From the heatmap analysis (Figure 3) it is clear that nitrate and urea had the similar metabolites expression which can also be seen from PCA analysis that PC1 grouped these two together (Figure 2E). Similarly ammonium and glycine treated samples were grouped together. However there was distinctive difference in each N treatment between the two genotypes. Hence 3-hydroxypropionic, isopropyl-ß-d-thiogalacto-pyranoside, glutamine, D-(glycerol 1-phosphate), allose and methyl phosphate were up-regulated in Hua30 and down-regulated in XZ16 in ammonium treatment, while 4-acetamidobutyric acid, alpha-ketoglutaric acid, mucic acid were up-regulated in XZ16 and down-regulated in Hua30. Mucic acid was also up-regulated in urea, ammonium and glycine treatments only in XZ16. Ornithine, 3-cyanoalanine, asparagine were up-regulated in ammonium and glycine treatments in both the genotypes, while these metabolites were down-regulated in nitrate and urea treatments except Hua30 which showed a little up-regulation in urea treatment. Similarly, glutamine was down-regulated in nitrate and urea treatments, while up-regulated in ammonium and glycine treatments, except XZ16 which showed down regulation of glutamine in ammonium treatment. Some metabolites were treatment specific such as putrescine, citric acid, quinic acid which were strongly down-regulated in ammonium treatment for both the genotypes, while up-regulated or unchanged in other treatments. Similarly, lyxose and sulfuric acid was greatly reduced only in glycine treatment for both the genotypes. On the other hand, some metabolites were genotypes specific such as methyl phosphate which was up-regulated in Hua30 in nitrate, urea, and ammonium treatments. Similarly mucic acid was up-regulated in XZ16 in ammonium, urea and glycine treatments (Figure 3).
In detail, we compared the whole metabolic profiles of the two genotypes in ammonium, urea, and glycine treatments with respect to nitrate (as control) by characterizing all the differentially expressed metabolites into major groups (Table S1). Also we mapped the important metabolites to the common pathways of carbohydrates and amino acid metabolisms (Figure 4). Mostly carbohydrates were up-regulated in ammonium and urea treatments but down-regulated under glycine treatment in both the genotypes, but there were distinct variations between the wild and cultivated genotypes (Table S1). In ammonium treatment more carbohydrates were up-regulated in Hua30, but turanose, allose, D-(glycerol 1-phosphate), and 3,6-anhydro-D-galactose were commonly up- regulated in both the genotypes with greater fold changes in Hua30 as compared to XZ16. In urea treatment sorbose, turanose, mannose, digitoxose were also commonly up-regulated in both the genotypes but with greater change in Hua30. Very few carbohydrates were up-regulated in glycine treatment, but a number of amino acid and organic acids were up-regulated in glycine treatment for both the genotypes with greater number in XZ16 than Hua30. Some important amino acids such as aspartic acid, serine, L-allothreonine, asparagine, glutamine, 3-cyanoalanine, L-homoserine, and ornithine were commonly up-regulated in both the genotypes with greater fold change in XZ16 (Table S1). Similarly malonic acid, 4-aminobutyric acid, pyrrole-2-carboxylic acid, toluenesulfonic acid, guanidinosuccinic acid, dehydroascorbic Acid, and 3-hydroxypropionic acid were up-regulated in both the genotypes but with significantly higher change in XZ16. Some fatty acids were significantly up-regulated only in XZ16 in glycine treatment. Nitrogen containing compound putrescine was also up-regulated in both wild and cultivated genotypes with greater change in wild genotype XZ16. Glycolysis and TCA cycle intermediates were strongly down-regulated in both the genotypes with Hua30 being higher down regulated as compared to XZ16. Also fructose-6-phosphate, isocitric acid, aconitic acid were only significantly up-regulated in XZ16 (Figure 4).
Except carbohydrates metabolism, all other metabolites specially amino acids and organic acids were greatly reduced in urea treatment, but there was a great difference between the wild and cultivated genotypes with XZ16 showing more down regulation as compared to Hua30.
3.4. Pathway Analysis
Pathway analysis was carried out by using online metaboanalyst 6.0. Total differentially expressed metabolites were analyzed on the basis of KEGG pathway enrichment and topology analysis. The most important pathways on the basis of p-value of pathway enrichment and impact scores from topology analysis, were alanine, aspartate and glutamate metabolism (0.597), glyoxylate and dicarboxylate metabolism (0.328), glycine, serine and threonine metabolism (0.437), beta-alanine metabolism (0.325), pentose phosphate pathway (0.386), and citrate cycle (TCA cycle) (0.279) (Table S2). In glycine treatment glyoxylate and dicarboxylate metabolism and pentose phosphate pathway important intermediate 2-oxoglytarate was less down-regulated in XZ16 as compared to Hua30 (Figure 4), while in ammonium treatment glycine, serine and threonine pathway were also up-regulated in XZ16. TCA cycle and amino acid metabolism was greatly affected in the four nitrogen treatments (Table S1 and Figure 4). Aspartate and asparagine belonging to alanine, aspartate and glutamate pathway were highly up-regulated in ammonium and glycine treatments in XZ16 as compared to Hua30 (Table S1 and Figure 4).
4. Discussion
Leaf nutrient concentration and resorption play an important role in determining plant nutrient-use strategies [51]. High leaf nutrient concentration and low leaf nutrient resorption generally reflect a “resource spending” nutrient use strategy for plants to quickly grow and develop [52]. However, low leaf nutrient concentration and high leaf nutrient resorption usually reflect a “conservative consumption” nutrient use strategy for plant survival and reproduction [53]. Leaf nutrient concentration and stoichiometry also reflect plant nutrient status and the strength of nutrient limitation in plants [54,55]. In our study, we found dramatic reduction in shoot concentrations of macro and micro nutrients in glycine and ammonium treatments relative to other two N-form treatments. Still there was significant difference in the two genotypes in nutrient concentrations in each N treatment. XZ16 had higher concentrations of nutrient ions in glycine treatment as compared to Hua30, but Hua30 accumulated more nutrients in ammonium treatment (Table 1). These results are consistent with our previous results [49] where some parameters like leaf surface area and tiller per plant increased in ammonium treatment for the cultivated barley as compared to wild genotypes. Similarly higher biomass of wild genotypes was found in urea treatment in our previous study and here also we found higher nutrient concentrations in both the genotypes in urea treatment in comparison with other two treatments, with XZ16 being higher than Hua30. Dubey et al. [56] found that ammonium nitrogen not only induced disorder in nutrient balance but also reduced calcium (Ca) uptake. In an another study more ammonium uptake resulted in reduced plants shoot Ca and Mg concentrations and increased Mn and Al uptake [57,58].
Appropriate nitrogen fertilizer input can significantly increase the activity of enzymes related to nitrogen metabolism in leaves [59], and promote photosynthesis and biosynthesis of organic acids, amino acids, proteins and other nitrogen-containing secondary metabolites in plants [60]. In the present study, we found different pattern of carbohydrates, amino acids and organic acids among four nitrogen forms. In root metabolomics we had found down regulation of most carbohydrates, amino acids and organic acids in glycine treatment, but in shoot metabolomics, we found the opposite trend. Carbohydrate metabolism in shoots was greatly increased in ammonium and urea treatments in both the genotypes, with urea treatment being higher than ammonium treatment. But we can observe the difference between two genotypes (Table S1). In ammonium treatment some commonly regulated sugar metabolites such as turanose, allose, D-(glycerol 1-phosphate), 3,6-anhydro-d-galactose had greater fold change in Hua30 (11.13, 3.23, 1.84, 8.12) as compared to XZ16 (2.57, 1.68, 1.94, 1.62), respectively (Table S1). As osmolytes and signaling molecules, soluble sugars participate in the response and adaptation of plants to environmental stresses [61]. The beta-mannosylglycerate is a potential protein thermostabilizer [62] and significantly up-regulation of this metabolite in Hua30 may increase leaves protein stability in ammonium treatment [63]. More accumulation of soluble sugar may confer higher resistance to abiotic stresses in ammonium treatment to Hua30 as compared to XZ16. From figure 4, it is clear that glycolysis intermediate (fructose-6-P) was more up-regulated and pyruvate was less down-regulated in XZ16 as compared to Hua30, thus adding the property of more glycine utilization in XZ16. Similarly, TCA cycle intermediates (aconitic acid and isocitrate) were more up-regulated and citrate, alpha ketoglutarate, and fumarate were less down-regulated in XZ16 as compared to Hua30, which may also be the contributive metabolites in glycine for more uptakes (Figure 4). Overall, glycine reduced TCA intermediate in both the genotypes and these results are consistent with former studies conducted by Wang et al. [64] and Liu et al. [65], which partly explains the lower N use efficiency in response to glycine [64].
The amino acid pool of plants is characterized by high flexibility and varies among species, cell types, and physiological situations [39,66,67,68]. In the environments of nutritional, low temperature and drought stresses, it might stimulate the synthesis of some amino acids [60,69]. Amino acids up-regulation may be also due to degradation of proteins to single amino acids, which can provide carbon skeleton for TCA cycle for enhancement of energy under stress [70]. In the current study, we found more accumulation of amino acids in ammonium and glycine treatments in both the genotypes (Table S1) with Hua30 being more affected than XZ16 under glycine treatment. Perchlik and Tegeder [71] also found the increased amino acid delivery to aap2 leaves, resulting in an enhanced sink C supply for higher NUE under N limitation. Some studies support that amino acid transport processes in the shoot and/or associated changes in the shoot N status exert regulatory control over N uptake in roots [72,73,74,75]. Stress responsive amino acids such as proline, asparagine, homoserine were greatly induced in ammonium and glycine treatments but less responsive in urea treatment. Accumulation of asparagine is regarded as a self-regulation mechanism for plant adaptation to organic N sources [32,76], this effect was also observed in our study (Figure 4). Intermediates of arginine and proline metabolism (L-ornithine and putrescine) and 4-aminobutanoic acid are associated with plant stress responses [77,78,79]. Glycine treatment led to higher concentrations of these components (Figure 4), suggesting that supply of glycine as an N source may increase stress resistance in barley as reported in lettuce earlier [80]. Considering these changes in plant metabolism, it is expected that glycine fertilizers could enhance the levels of antioxidants which might improve tolerance to abiotic stresses [81].
Certain metabolites; including carbon compounds from photosynthesis, starch and sucrose metabolism plus oxidative pentose phosphate pathway, tricarboxylic acid (TCA) cycle and glycolysis metabolites and N-containing metabolites such as glutamine, have been seen to be at low levels in plants undergoing high growth. This suggests that these metabolites are providing the major building blocks for bio-macromolecules such as proteins and that growth drives metabolism [82]. In urea treatment, amino acid and organic acids were greatly down-regulated in XZ16 as compared to Hua30. Urea is a good N fertilizer form to barley only after nitrate [83]. Thus for the barley seedlings exposed to urea treatment, it is not necessary for them to accumulate more amino acids or organic acids as stress responsive. Also the amino acids produced may be directly used for protein synthesis that may be the reason for less accumulation of amino acids. Furthermore more down regulation of these metabolites in XZ16 may be related to the more biomass production in urea treatment as reported earlier [83].
Glyoxylate and dicarboxylate metabolism is an important abiotic stress-related pathway, providing a metabolic balance to improve tolerance [84]. In glycine treatment this was greatly affected (Table S6). Oxalic acid, isocitrate, and glutamin were significantly up-regulated in XZ16 as compared to Hua30 in glycine of this pathway which may be the reason for more utilization of glycine in XZ16. Similarly arginine and proline pathway is related to nitrogen metabolism in plants, essential for production of nucleic acids and proteins. Arginine is a precursor of polyamines and has a role in proline biosynthesis when glutamate is not available. In ammonium treatment this pathway is greatly affected (Table S4). The influence of ammonium treatment is highly expected in this pathway. Neto et al. [85] also found these pathways as stress responsive.
5. Conclusions
Among the 163 differentially expressed metabolites identified in the two barley genotypes exposed to four forms of N, highest up-regulation was observed in glycine treatment, while highest down-regulation was observed in urea treatment in wild genotype XZ16. Except carbohydrates, all other metabolites, especially amino acids were greatly down-regulated in urea treatment, which may be directly utilized for protein synthesis to produces higher biomass as reported earlier in our previous study [83]. On the other hand, up-regulation of amino acids and stress responsive metabolites in glycine treatment, as glycine did not provide the sufficient N for normal metabolisms of the two genotypes. Furthermore, higher concentrations of macro and micro nutrients were found in urea and nitrate as compared to ammonium and glycine treatment. Relatively XZ16 had higher nutrient concentrations than Hua30 in glycine treatment, while it was just opposite when the two genotypes were subjected to ammonium treatment. More up-regulation of soluble sugar in Hua30 in ammonium treatment may contribute to reduce ammonium toxicity. Similarly glycolysis and TCA intermediates up-regulated only in XZ16 in glycine treatment along with stress responsive metabolites such as proline, asparagine, putrescine more up-regulation in XZ16 as compared to Hua30 may enable the wild genotypes for better utilization of glycine. In short, we suggested that nitrate and urea are best nitrogen sources for barley, and wild barley can utilize N from glycine.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1: title; Table S1: title.
Author Contributions
Conceptualization, S.N. and G.Z.; sample preparation, S.N., F.K. and A.M.; formal analysis, S.N. and S.U.K.; investigation, S.N.; writing—original draft preparation, S.N and S.U.K.; writing, review and editing, S.U.K and G.Z.; project administration, G.Z.; funding acquisition, G.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Key Research Foundation of Science and Technology Department of Zhejiang Province of China (2021C02064-3) and Jiangsu Collaborative Innovation Center for Modern Crop Production (JCIC-MCP).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data sets supporting the conclusions of this article are included within the article and its additional files.
Acknowledgments
This work was supported by the Key Research Foundation of Science and Technology Department of Zhejiang Province of China (2021C02064-3) and Jiangsu Collaborative Innovation Center for Modern Crop Production (JCIC-MCP).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Lemaire, G.; Sinclair, T.; Sadras, V.; Bélanger, G. Allometric Approach to Crop Nutrition and Implications for Crop Diagnosis and Phenotyping: A Review. Agron. Sustain. Dev. 2019, 39. [Google Scholar] [CrossRef]
- Dong, N.Q.; Lin, H.X. Higher Yield with Less Nitrogen Fertilizer. Nat. Plants 2020, 6, 1078–1079. [Google Scholar] [CrossRef]
- Zhao, B.; Ata-Ul-Karim, S.T.; Duan, A.; Liu, Z.; Wang, X.; Xiao, J.; Liu, Z.; Qin, A.; Ning, D.; Zhang, W.; Lian, Y. Determination of Critical Nitrogen Concentration and Dilution Curve Based on Leaf Area Index for Summer Maize. Field Crops Res. 2018, 228, 195–203. [Google Scholar] [CrossRef]
- Ge, J.; Tang, H.; Yang, N.; Hu, Y. Rapid Identification of Damaged Buildings Using Incremental Learning with Transferred Data from Historical Natural Disaster Cases. ISPRS J. Photogramm. Remote Sens. 2023, 195, 105–128. [Google Scholar] [CrossRef]
- Bloom, A.J. The Increasing Importance of Distinguishing among Plant Nitrogen Sources. Curr. Opin. Plant Biol. 2015, 25, 10–16. [Google Scholar] [CrossRef]
- Owen, A.G.; Jones, D.L. Competition for Amino Acids between Wheat Roots and Rhizosphere Microorganisms and the Role of Amino Acids in Plant N Acquisition. Soil biol. biochem. 2001, 33, 651–657. [Google Scholar] [CrossRef]
- Becker-Ritt, A.B.; Martinelli, A.H.S.; Mitidieri, S.; Feder, V.; Wassermann, G.E.; Santi, L.; Vainstein, M.H.; Oliveira, J.T.A.; Fiuza, L.M.; Pasquali, G. Antifungal Activity of Plant and Bacterial Ureases. Toxicon 2007, 50, 971–983. [Google Scholar] [CrossRef] [PubMed]
- Perchlik, M.; Foster, J.; Tegeder, M. Different and Overlapping Functions of Arabidopsis LHT6 and AAP1 Transporters in Root Amino Acid Uptake. J. Exp. Bot. 2014, 65, 5193–5204. [Google Scholar] [CrossRef]
- Mérigout, P.; Lelandais, M.; Bitton, F.; Renou, J.-P.; Briand, X.; Meyer, C.; Daniel-Vedele, F. Physiological and Transcriptomic Aspects of Urea Uptake and Assimilation in Arabidopsis Plants. Plant Physiol. 2008, 147, 1225–1238. [Google Scholar] [CrossRef] [PubMed]
- Pinton, R.; Tomasi, N.; Zanin, L. Molecular and Physiological Interactions of Urea and Nitrate Uptake in Plants. Plant signal. behav. 2016, 11, e1076603. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Yu, M.; Xu, F.; Ouyang, S.; Xu, X.; Gao, Q.; Li, X. Uptake of Amino Acids and Inorganic Nitrogen by Two Dominant Temperate Grasses. Rhizosphere 2020, 14, 100199. [Google Scholar] [CrossRef]
- Näsholm, T.; Ekblad, A.; Nordin, A.; Giesler, R.; Högberg, M.; Högberg, P. Boreal Forest Plants Take up Organic Nitrogen. Nature 1998, 392, 914–916. [Google Scholar] [CrossRef]
- Henry, H.A.L.; Jefferies, R.L. Interactions in the Uptake of Amino Acids, Ammonium and Nitrate Ions in the Arctic Salt-marsh Grass, Puccinellia Phryganodes. Plant Cell Environ. 2003, 26, 419–428. [Google Scholar] [CrossRef]
- Hu, B.; Zhou, M.; Bilela, S.; Simon, J.; Dannenmann, M.; Liu, X.; Alfarraj, S.; Hou, L.; Chen, H.; Zhang, S.; et al. Nitrogen Nutrition of Native and Introduced Forest Tree Species in N-Limited Ecosystems of the Qinling Mountains, China. Trees 2017, 31, 1189–1202. [Google Scholar] [CrossRef]
- Ge, T.; Song, S.; Roberts, P.; Jones, D.L.; Huang, D.; Iwasaki, K. Amino Acids as a Nitrogen Source for Tomato Seedlings: The Use of Dual-Labeled (13C, 15N) Glycine to Test for Direct Uptake by Tomato Seedlings. Environ. Exp. Bot. 2009, 66, 357–361. [Google Scholar] [CrossRef]
- Gioseffi, E.; de Neergaard, A.; Schjørring, J.K. Interactions between Uptake of Amino Acids and Inorganic Nitrogen in Wheat Plants. Biogeosciences 2012, 9, 1509–1518. [Google Scholar] [CrossRef]
- Brackin, R.; Näsholm, T.; Robinson, N.; Guillou, S.; Vinall, K.; Lakshmanan, P.; Schmidt, S.; Inselsbacher, E. Nitrogen Fluxes at the Root-Soil Interface Show a Mismatch of Nitrogen Fertilizer Supply and Sugarcane Root Uptake Capacity. Sci. Rep. 2015, 5, 15727. [Google Scholar] [CrossRef]
- Wei, L.; Chen, C.; Yu, S. Uptake of Organic Nitrogen and Preference for Inorganic Nitrogen by Two Australian Native Araucariaceae Species. Plant Ecol. Divers. 2015, 8, 259–264. [Google Scholar] [CrossRef]
- Guo, S.; Zhou, Y.; Shen, Q.; Zhang, F. Effect of Ammonium and Nitrate Nutrition On Some Physiological Processes in Higher Plants - Growth, Photosynthesis, Photorespiration, and Water Relations. Plant Biol. 2007, 9, 21–29. [Google Scholar] [CrossRef]
- Szczerba, M.W.; Britto, D.T.; Ali, S.A.; Balkos, K.D.; Kronzucker, H.J. NH4+-Stimulated and -Inhibited Components of K+ Transport in Rice (Oryza Sativa L.). J. Exp. Bot. 2008, 59, 3415–3423. [Google Scholar] [CrossRef]
- Yang, X.; Li, Y.; Ren, B.; Ding, L.; Gao, C.; Shen, Q.; Guo, S. Drought-Induced Root Aerenchyma Formation Restricts Water Uptake in Rice Seedlings Supplied with Nitrate. Plant Cell Physiol. 2012, 53, 495–504. [Google Scholar] [CrossRef]
- Ding, L.; Gao, C.; Li, Y.; Li, Y.; Zhu, Y.; Xu, G.; Shen, Q.; Kaldenhoff, R.; Kai, L.; Guo, S. The Enhanced Drought Tolerance of Rice Plants under Ammonium Is Related to Aquaporin (AQP). Plant Sci. 2015, 234, 14–21. [Google Scholar] [CrossRef]
- Cramer, M.D.; Lewis, O.A.M. The Influence of Nitrate and Ammonium Nutrition on the Growth of Wheat (Triticum Aestivum) and Maize (Zea Mays) Plants. Ann. Bot. 1993, 72, 359–365. [Google Scholar] [CrossRef]
- Raab, T.K.; Terry, N. Nitrogen Source Regulation of Growth and Photosynthesis in Beta Vulgaris L. Plant Physiol. 1994, 105, 1159–1166. [Google Scholar] [CrossRef]
- Guo, S.; Kaldenhoff, R.; Uehlein, N.; Sattelmacher, B.; Brueck, H. Relationship between Water and Nitrogen Uptake in Nitrate- and Ammonium-Supplied Phaseolus Vulgaris L. Plants. J. Plant Nutr. Soil Sci. 2007, 170, 73–80. [Google Scholar] [CrossRef]
- Lu, Y.X.; Li, C.J.; Zhang, F.S. Transpiration, Potassium Uptake and Flow in Tobacco as Affected by Nitrogen Forms and Nutrient Levels. Ann. Bot. 2005, 95, 991–998. [Google Scholar] [CrossRef]
- Gao, L.; Liu, M.; Wang, M.; Shen, Q.; Guo, S. Enhanced Salt Tolerance under Nitrate Nutrition Is Associated with Apoplast Na+ Content in Canola (Brassica. Napus L.) and Rice (Oryza Sativa L.) Plants. Plant Cell Physiol. 2016, 57, 2323–2333. [Google Scholar] [CrossRef]
- Li, Y.; Gao, Y.; Ding, L.; Shen, Q.; Guo, S. Ammonium Enhances the Tolerance of Rice Seedlings (Oryza Sativa L.) to Drought Condition. Agric. Water Manag. 2009, 96, 1746–1750. [Google Scholar] [CrossRef]
- Malagoli, M.; Dal Canal, A.; Quaggiotti, S.; Pegoraro, P.; Bottacin, A. Differences in Nitrate and Ammonium Uptake between Scots Pine and European Larch. Plant Soil 2000, 221, 1–3. [Google Scholar] [CrossRef]
- Nasir, C.; Malik, M.; Qadir, G.; Rafique, M.; Nawaz, N. Influence of Temperature and Osmotic Stress on Germination Induction of Different Castor Bean Cultivars. Pak. J. Bot. 2010, 42, 4035–4041. [Google Scholar]
- Khalilzadeh, R.; Tajbakhsh, M.; Jalilian, J. Growth Characteristics of Mung Bean (Vigna Radiata L.) Affected by Foliar Application of Urea and Bio-Organic Fertilizers. Int. J. Agric. Crop Sci. 2012, 4, 637–642. [Google Scholar]
- Forsum, O.; Svennerstam, H.; Ganeteg, U.; Näsholm, T. Capacities and Constraints of Amino Acid Utilization in Arabidopsis. New Phytol. 2008, 179, 1058–1069. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.L.; Yang, D.N.; Huang, D.F. Effects of different applied nitrogen forms on pakchoi (Brassica chinensis) growth and its carbon and nitrogen accumulation. Ying yong sheng tai xue bao = J. appl. ecol. 2012, 23, 1042–1048. [Google Scholar]
- Ti-Da, G.E.; Shi-Wei, S.; Wu, J.; Dong-Mei, T.; Dan-Feng, H. Influence of Glycine-N Concentration on the Growth and Nitrogen Metabolism of Tomato Seedlings under Sterile Hydroponics Cultivation. Shengtai Xuebao/ Acta Ecologica Sinica 2009, 29, 1994–2002. [Google Scholar]
- Wang, X.; Tang, D.; Huang, D. Proteomic Analysis of Pakchoi Leaves and Roots under Glycine–Nitrogen Conditions. Plant Physiol. Biochem. 2014, 75, 96–104. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.; Tian, J.; Liao, H. Proteomics Dissection of Plant Responses to Mineral Nutrient Deficiency. Proteomics 2013, 13, 624–636. [Google Scholar] [CrossRef]
- Zhou, M.; Zhu, S.; Mo, X.; Guo, Q.; Li, Y.; Tian, J.; Liang, C. Proteomic Analysis Dissects Molecular Mechanisms Underlying Plant Responses to Phosphorus Deficiency. Cells 2022, 11, 651. [Google Scholar] [CrossRef]
- Luo, J. Metabolite-Based Genome-Wide Association Studies in Plants. Curr. Opin. Plant Biol. 2015, 24, 31–38. [Google Scholar] [CrossRef]
- Schlüter, U.; Mascher, M.; Colmsee, C.; Scholz, U.; Bräutigam, A.; Fahnenstich, H.; Sonnewald, U. Maize Source Leaf Adaptation to Nitrogen Deficiency Affects Not Only Nitrogen and Carbon Metabolism but Also Control of Phosphate Homeostasis. Plant Physiol. 2012, 160, 1384–1406. [Google Scholar] [CrossRef]
- Sung, J.; Lee, S.; Lee, Y.; Ha, S.; Song, B.; Kim, T.; Waters, B.M.; Krishnan, H.B. Metabolomic Profiling from Leaves and Roots of Tomato (Solanum Lycopersicum L.) Plants Grown under Nitrogen, Phosphorus or Potassium-Deficient Condition. Plant Sci. 2015, 241, 55–64. [Google Scholar] [CrossRef]
- Quan, X.; Qian, Q.; Ye, Z.; Zeng, J.; Han, Z.; Zhang, G. Metabolic Analysis of Two Contrasting Wild Barley Genotypes Grown Hydroponically Reveals Adaptive Strategies in Response to Low Nitrogen Stress. J. plant physiol. 2016, 206, 59–67. [Google Scholar] [CrossRef]
- Tawaraya, K.; Horie, R.; Wagatsuma, T.; Saito, K.; Oikawa, A. Metabolite Profiling of Shoot Extract, Root Extract, and Root Exudate of Rice under Nitrogen and Phosphorus Deficiency. Soil Sci. Plant Nutr. 2018, 64, 312–322. [Google Scholar] [CrossRef]
- Xin, W.; Zhang, L.; Zhang, W.; Gao, J.; Yi, J.; Zhen, X.; Li, Z.; Zhao, Y.; Peng, C.; Zhao, C. An Integrated Analysis of the Rice Transcriptome and Metabolome Reveals Differential Regulation of Carbon and Nitrogen Metabolism in Response to Nitrogen Availability. Int. J. Mol. Sci. 2019, 20, 2349. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Xue, Y.; Xie, B.; Zhu, S.; Lu, X.; Liang, C.; Tian, J. Complex Gene Regulation between Young and Old Soybean Leaves in Responses to Manganese Toxicity. Plant Physiol. Biochem. 2020, 155, 231–242. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, D.; Tao, Z.; Yang, Y.; Gao, Z.; Zhao, G.; Chang, X. Impacts of Nitrogen Deficiency on Wheat (Triticum Aestivum L.) Grain During the Medium Filling Stage: Transcriptomic and Metabolomic Comparisons. Front. Plant Sci. 2021, 12, 674433. [Google Scholar] [CrossRef]
- Shen, X.; Yang, L.; Han, P.; Gu, C.; Li, Y.; Liao, X.; Qin, L. Metabolic Profiles Reveal Changes in the Leaves and Roots of Rapeseed (Brassica Napus L.) Seedlings under Nitrogen Deficiency. Int. J. Mol. Sci. 2022, 23, 5784. [Google Scholar] [CrossRef]
- Tschoep, H.; Gibon, Y.; Carillo, P.; Armengaud, P.; Szecowka, M.; Nunes-Nesi, A.; Fernie, A.R.; Koehl, K.; Stitt, M. Adjustment of Growth and Central Metabolism to a Mild but Sustained Nitrogen-Limitation in Arabidopsis. Plant Cell Environ. 2009, 32, 300–318. [Google Scholar] [CrossRef]
- Schlüter, U.; Colmsee, C.; Scholz, U.; Bräutigam, A.; Weber, A.P.M.; Zellerhoff, N.; Bucher, M.; Fahnenstich, H.; Sonnewald, U. Adaptation of Maize Source Leaf Metabolism to Stress Related Disturbances in Carbon, Nitrogen and Phosphorus Balance. BMC genomics 2013, 14, 1–25. [Google Scholar] [CrossRef]
- Naz, S.; Shen, Q.; Wa Lwalaba, J.L.; Zhang, G. Genotypic Difference in the Responses to Nitrogen Fertilizer Form in Tibetan Wild and Cultivated Barley. Plants 2021, 10, 595. [Google Scholar] [CrossRef]
- Tsugawa, H.; Tsujimoto, Y.; Arita, M.; Bamba, T.; Fukusaki, E. GC/MS Based Metabolomics: Development of a Data Mining System for Metabolite Identification by Using Soft Independent Modeling of Class Analogy (SIMCA). BMC Bioinformatics 2011, 12, 131. [Google Scholar] [CrossRef]
- See, C.R.; Yanai, R.D.; Fisk, M.C.; Vadeboncoeur, M.A.; Quintero, B.A.; Fahey, T.J. Soil Nitrogen Affects Phosphorus Recycling: Foliar Resorption and Plant–Soil Feedbacks in a Northern Hardwood Forest. Ecology 2015, 96, 2488–2498. [Google Scholar] [CrossRef] [PubMed]
- Kobe, R.K.; Lepczyk, C.A.; Iyer, M. Resorption Efficiency Decreases With Increasing Green Leaf Nutrients in A Global Data Set. Ecology 2005, 86, 2780–2792. [Google Scholar] [CrossRef]
- Lü, X.T.; Reed, S.C.; Yu, Q.; Han, X.G. Nutrient Resorption Helps Drive Intra-Specific Coupling of Foliar Nitrogen and Phosphorus under Nutrient-Enriched Conditions. Plant Soil 2016, 398, 111–120. [Google Scholar] [CrossRef]
- Grime, J.P. Benefits of Plant Diversity to Ecosystems: Immediate, Filter and Founder Effects. J. Ecol. 1998, 86, 902–910. [Google Scholar] [CrossRef]
- Güsewell, S. Nutrient Resorption of Wetland Graminoids Is Related to the Type of Nutrient Limitation. Funct. Ecol. 2005, 19, 344–354. [Google Scholar] [CrossRef]
- Dubey, R.S.; Srivastava, R.K.; Pessarakli, M. Physiological mechanisms of nitrogen absorption and assimilation in plants under stressful conditions. In Handbook of plant and crop physiology; CRC Press, 2021; pp. 579–616.
- Bowman, W.D.; Cleveland, C.C.; Halada, Ĺ.; Hreško, J.; Baron, J.S. Negative Impact of Nitrogen Deposition on Soil Buffering Capacity. Nature Geoscience 2008, 1, 767–770. [Google Scholar] [CrossRef]
- Tian, D.; Niu, S. A Global Analysis of Soil Acidification Caused by Nitrogen Addition. Environ. Res. Lett. 2015, 10, 24019. [Google Scholar] [CrossRef]
- Wang, J.; Fu, Z.; Chen, G.; Zou, G.; Song, X.; Liu, F. Runoff Nitrogen (N) Losses and Related Metabolism Enzyme Activities in Paddy Field under Different Nitrogen Fertilizer Levels. Environ. Sci. Pollut. Res. Int. 2018, 25, 27583–27593. [Google Scholar] [CrossRef]
- Cao, Y.; Qu, R.; Tang, X.; Sun, L.; Chen, Q.; Miao, Y. UPLC-Triple TOF-MS/MS Based Metabolomics Approach to Reveal the Influence of Nitrogen Levels on Isatis Indigotica Seedling Leaf. Sci. Hortic. 2020, 266, 109280. [Google Scholar] [CrossRef]
- Wang, H.; Gong, M.; Xin, H.; Tang, L.; Dai, D.; Gao, Y.; Liu, C. Effects of Chilling Stress on the Accumulation of Soluble Sugars and Their Key Enzymes in Jatropha Curcas Seedlings. Physiol. Mol. Biol. Plants 2018, 24, 857–865. [Google Scholar] [CrossRef]
- Ramos, A.; Raven, N.; Sharp, R.J.; Bartolucci, S.; Rossi, M.; Cannio, R.; Lebbink, J.; J, V.D.O.; M, D.V.W.; Santos, H. Stabilization of Enzymes against Thermal Stress and Freeze-Drying by Mannosylglycerate. Appl. Environ. Microbiol. 1997, 63, 4020–4025. [Google Scholar] [CrossRef]
- Lin, Z.H.; Chen, C.S.; Zhong, Q.S.; Ruan, Q.C.; Chen, Z.H.; You, X.M.; Shan, R.Y.; Li, X.L. The GC-TOF/MS-Based Metabolomic Analysis Reveals Altered Metabolic Profiles in Nitrogen-Deficient Leaves and Roots of Tea Plants (Camellia Sinensis). BMC Plant Biol. 2021, 21, 506. [Google Scholar] [CrossRef]
- Wang, X.; Yu, W.; Zhou, Q.; Han, R.; Huang, D. Metabolic Response of Pakchoi Leaves to Amino Acid Nitrogen. J. Integr. Agric. 2014, 13, 778–788. [Google Scholar] [CrossRef]
- Liu, X.; Yang, X.; Wang, L.; Duan, Q.; Huang, D. Comparative Analysis of Metabolites Profile in Spinach (Spinacia Oleracea L.) Affected by Different Concentrations of Gly and Nitrate. Sci. Hortic. 2016, 204, 8–15. [Google Scholar] [CrossRef]
- Millward, D.J.; Layman, D.K.; Tomé, D.; Schaafsma, G. Protein Quality Assessment: Impact of Expanding Understanding of Protein and Amino Acid Needs for Optimal Health1. Am. J. Clin. Nutr. 2008, 87, 1576S–1581S. [Google Scholar] [CrossRef]
- Tessari, P.; Lante, A.; Mosca, G. Essential Amino Acids: Master Regulators of Nutrition and Environmental Footprint? Sci. Rep. 2016, 6, 26074. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Sharma, A.; Kaur, R.; Thukral, A.K.; Bhardwaj, R.; Ahmad, P. Differential Distribution of Amino Acids in Plants. Amino Acids 2017, 49, 821–869. [Google Scholar] [CrossRef]
- Deng, Y.; Lu, S. Biosynthesis and Regulation of Phenylpropanoids in Plants. Crit. Rev. Plant Sci. 2017, 36, 257–290. [Google Scholar] [CrossRef]
- Anzano, A.; Bonanomi, G.; Mazzoleni, S.; Lanzotti, V. Plant Metabolomics in Biotic and Abiotic Stress: A Critical Overview. Phytochem. Rev. 2022, 21, 503–524. [Google Scholar] [CrossRef]
- Perchlik, M.; Tegeder, M. Leaf Amino Acid Supply Affects Photosynthetic and Plant Nitrogen Use Efficiency under Nitrogen Stress. Plant physiol. 2018, 178, 174–188. [Google Scholar] [CrossRef]
- Tan, Q.; Zhang, L.; Grant, J.; Cooper, P.; Tegeder, M. Increased Phloem Transport of S-Methylmethionine Positively Affects Sulfur and Nitrogen Metabolism and Seed Development in Pea Plants. Plant Physiol. 2010, 154, 1886–1896. [Google Scholar] [CrossRef]
- Zhang, L.; Garneau, M.G.; Majumdar, R.; Grant, J.; Tegeder, M. Improvement of Pea Biomass and Seed Productivity by Simultaneous Increase of Phloem and Embryo Loading with Amino Acids. The Plant J. 2015, 81, 134–146. [Google Scholar] [CrossRef]
- Santiago, J.P.; Tegeder, M. Connecting Source with Sink: The Role of Arabidopsis AAP8 in Phloem Loading of Amino Acids. Plant Physiol. 2016, 171, 508–521. [Google Scholar] [CrossRef]
- Perchlik, M.; Tegeder, M. Improving Plant Nitrogen Use Efficiency through Alteration of Amino Acid Transport Processes. Plant Physiol. 2017, 175, 235–247. [Google Scholar] [CrossRef]
- Oliveira, I.C.; Brenner, E.; Chiu, J.; Hsieh, M.H.; Kouranov, A.; Lam, H.M.; Shin, M.J.; Coruzzi, G. Metabolite and Light Regulation of Metabolism in Plants: Lessons from the Study of a Single Biochemical Pathway. Braz. J. Med. Biol. Res. 2001, 34, 567–575. [Google Scholar] [CrossRef]
- Kinnersley, A.M.; Turano, F.J. Gamma Aminobutyric Acid (GABA) and Plant Responses to Stress. Crit. Rev. Plant Sci. 2000, 19, 479–509. [Google Scholar] [CrossRef]
- Bown, A.W.; Shelp, B.J. Plant GABA: Not Just a Metabolite. Trends Plant Sci. 2016, 21, 811–813. [Google Scholar] [CrossRef] [PubMed]
- Alcázar, R.; Planas, J.; Saxena, T.; Zarza, X.; Bortolotti, C.; Cuevas, J.; Bitrián, M.; Tiburcio, A.F.; Altabella, T. Putrescine Accumulation Confers Drought Tolerance in Transgenic Arabidopsis Plants Over-Expressing the Homologous Arginine Decarboxylase 2 Gene. Plant Physiol. Biochem. 2010, 48, 547–552. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Feng, L.; Zhao, L.; Liu, X.; Hassani, D.; Huang, D. Effect of Glycine Nitrogen on Lettuce Growth under Soilless Culture: A Metabolomics Approach to Identify the Main Changes Occurred in Plant Primary and Secondary Metabolism. J. Sci. Food Agric. 2018, 98, 467–477. [Google Scholar] [CrossRef] [PubMed]
- Luna, M.C.; Tudela, J.A.; Tomás-Barberán, F.A.; Gil, M.I. Modified Atmosphere (MA) Prevents Browning of Fresh-Cut Romaine Lettuce through Multi-Target Effects Related to Phenolic Metabolism. Postharvest Biol. Technol. 2016, 119, 84–93. [Google Scholar] [CrossRef]
- Beatty, P.H.; Klein, M.S.; Fischer, J.J.; Lewis, I.A.; Muench, D.G.; Good, A.G. Understanding Plant Nitrogen Metabolism through Metabolomics and Computational Approaches. Plants 2016, 5, 39. [Google Scholar] [CrossRef] [PubMed]
- Naz, S.; Shen, Q.; Lwalaba, J.L.; Zhang, G. Genotypic Difference in the Responses to Nitrogen Fertilizer Form in Tibetan Wild and Cultivated Barley. Plants 2021, 10, 595. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Zeng, X.; Wu, J.; Zhang, F.; Li, C.; Jiang, J.; Wang, Y.; Sun, W. ITRAQ-Based Quantitative Proteome Revealed Metabolic Changes in Winter Turnip Rape (Brassica Rapa L.) under Cold Stress. Int. J. Mol. Sci. 2018, 19, 3346. [Google Scholar] [CrossRef] [PubMed]
- Neto, J.C.R.; Vieira, L.R.; de Aquino Ribeiro, J.A.; de Sousa, C.A.F.; Júnior, M.T.S.; Abdelnur, P.V. Metabolic Effect of Drought Stress on the Leaves of Young Oil Palm (Elaeis Guineensis) Plants Using UHPLC–MS and Multivariate Analysis. Sci. Rep. 2021, 11, 18271. [Google Scholar] [CrossRef]
Figure 1.
Venn diagram of differentially expressed metabolites in shoot tissues of two barley genotypes (XZ16 and Hua30). (A) Upregulated metabolites and (B) downregulated metabolites.
Figure 1.
Venn diagram of differentially expressed metabolites in shoot tissues of two barley genotypes (XZ16 and Hua30). (A) Upregulated metabolites and (B) downregulated metabolites.
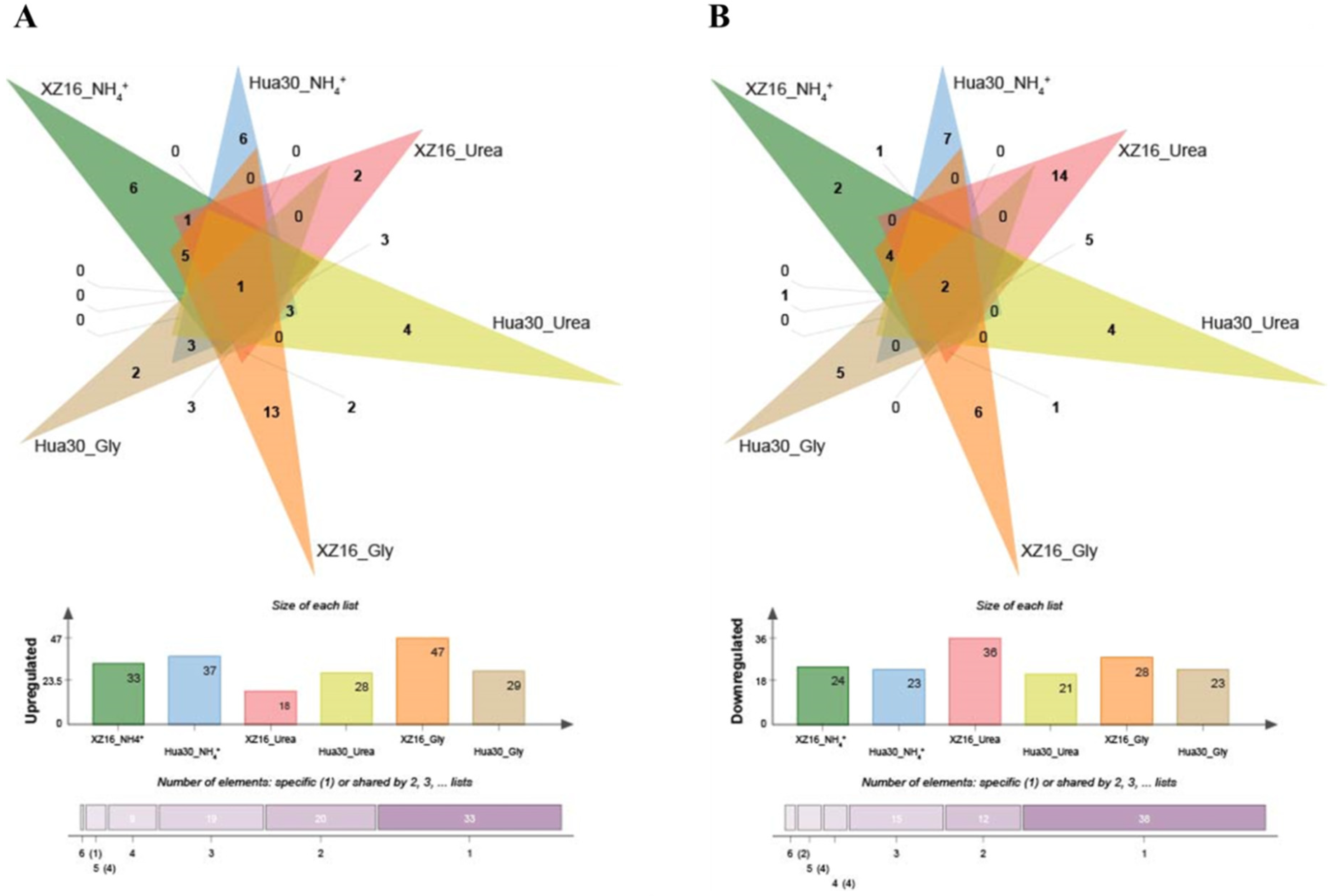
Figure 2.
Partial least square discriminate analysis (PLSDA) of metabolites in shoot tissues under control (A), Ammonium (B), Urea (C), and Glycine (D) along with VIP score contributing to the classification of genotypes. (E) Principal component analysis (PCA) of metabolites in XZ16 and Hua30 under four different nitrogen fertilizers treatments. PC1: first principal component; PC2: second principal component.
Figure 2.
Partial least square discriminate analysis (PLSDA) of metabolites in shoot tissues under control (A), Ammonium (B), Urea (C), and Glycine (D) along with VIP score contributing to the classification of genotypes. (E) Principal component analysis (PCA) of metabolites in XZ16 and Hua30 under four different nitrogen fertilizers treatments. PC1: first principal component; PC2: second principal component.
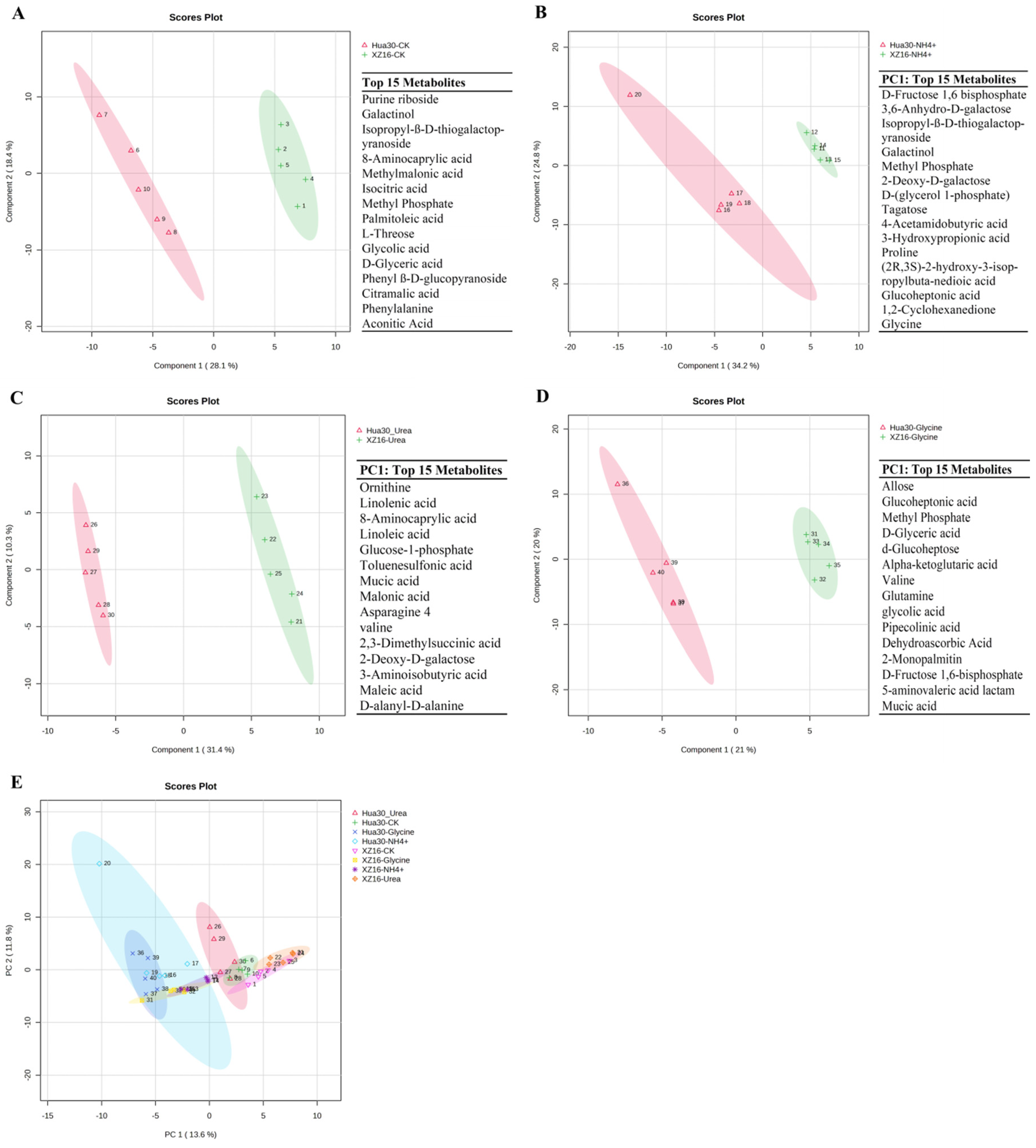
Figure 3.
Hierarchical clustering analysis for significant differentially expressed metabolites (DEMs) and the metabolomics view map of the significant metabolic pathways of the shoot tissues of XZ16 and Hua30 under different forms of nitrogen fertilizers. The relative metabolite level is shown according to color scale. Red indicates up-regulation, and blue indicates down-regulation.
Figure 3.
Hierarchical clustering analysis for significant differentially expressed metabolites (DEMs) and the metabolomics view map of the significant metabolic pathways of the shoot tissues of XZ16 and Hua30 under different forms of nitrogen fertilizers. The relative metabolite level is shown according to color scale. Red indicates up-regulation, and blue indicates down-regulation.
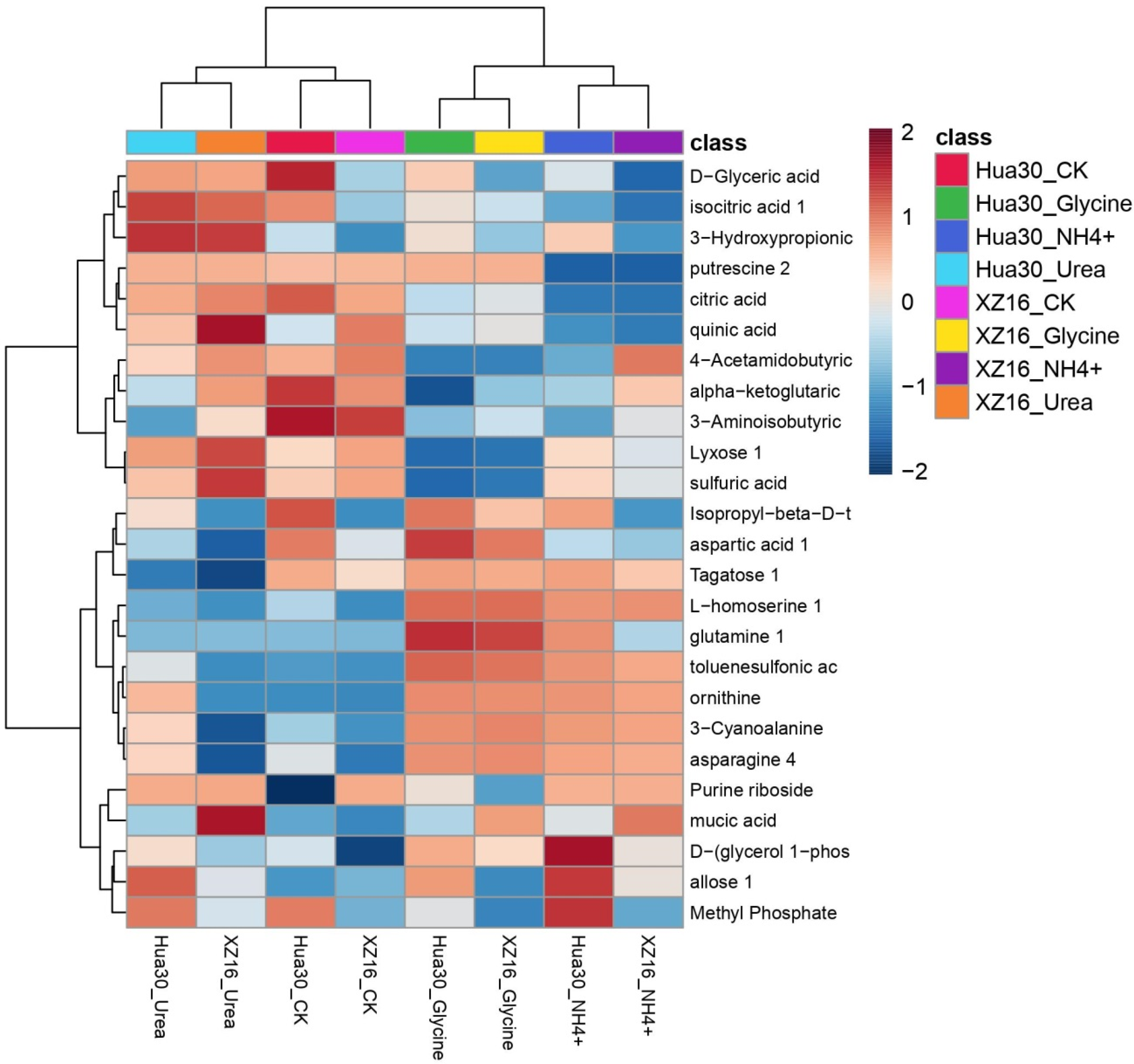
Figure 4.
Changes in the metabolites mapped in the metabolic pathways in shoot tissues of XZ16 and Hua30. Metabolites in green were the identified metabolites in this study. Fold changes of metabolites in red color indicated significant up-regulation (P ≤ 0.05 and fold change > 1) and in blue color indicated significant down-regulation (P ≤ 0.05 and fold change < 1), in comparison with normalized metabolite concentrations under different forms of nitrogen treatments and control conditions; and in white color indicated non-significant changes. Metabolic pathways were constructed on basis of KEGG database (http://www.genome.jp/kegg/).
Figure 4.
Changes in the metabolites mapped in the metabolic pathways in shoot tissues of XZ16 and Hua30. Metabolites in green were the identified metabolites in this study. Fold changes of metabolites in red color indicated significant up-regulation (P ≤ 0.05 and fold change > 1) and in blue color indicated significant down-regulation (P ≤ 0.05 and fold change < 1), in comparison with normalized metabolite concentrations under different forms of nitrogen treatments and control conditions; and in white color indicated non-significant changes. Metabolic pathways were constructed on basis of KEGG database (http://www.genome.jp/kegg/).
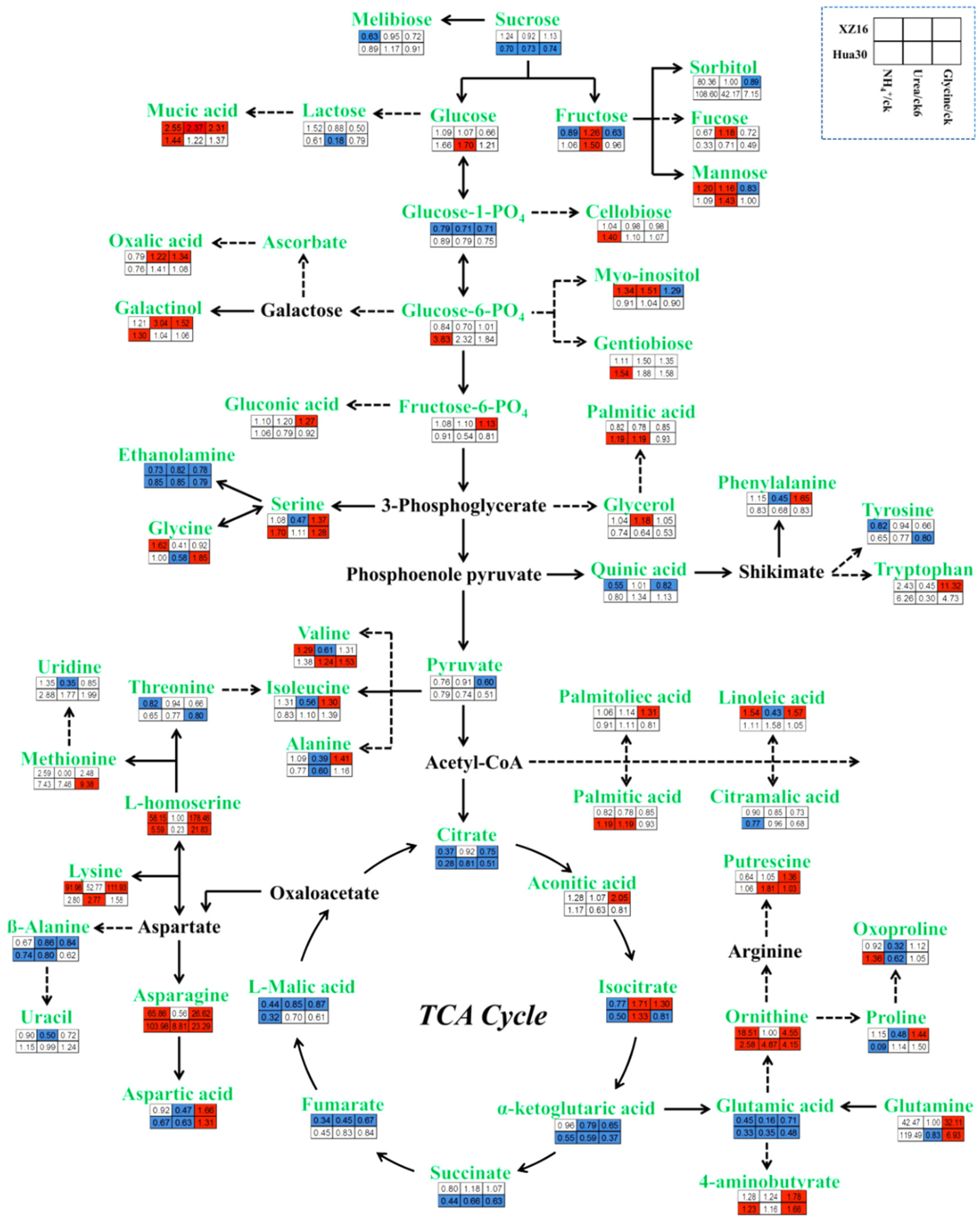

Table 1.
Analysis of element contents in the shoot tissues of wild and cultivated barley under different N fertilizers forms.
Table 1.
Analysis of element contents in the shoot tissues of wild and cultivated barley under different N fertilizers forms.
| Genotype | Treatments | K | Ca | Mg | P | S | Zn | Mn | Fe | Cu |
|---|---|---|---|---|---|---|---|---|---|---|
| mg g−1 DW | mg kg−1 DW | |||||||||
| XZ16 | NO3− | 55.75 b | 7.98 ab | 13.46 a | 13.10 a | 7.35 a | 2.30 a | 0.63 b | 2.84 a | 0.110 a |
| NH4+ | 46.82 d | 6.19 d | 10.18 de | 9.94 c | 6.09 cd | 1.79 bc | 0.42 de | 2.03 de | 0.087 b | |
| Urea | 58.48 a | 8.27 a | 12.51 b | 13.20 a | 7.11 ab | 2.03 ab | 0.73 a | 2.51 b | 0.093 ab | |
| Gly | 50.27 c | 6.42 cd | 11.03 c | 9.54 c | 6.55 bc | 2.16 ab | 0.53 c | 2.32 bc | 0.087 b | |
| Hua30 | NO3− | 46.27 de | 7.15 bc | 10.62 cd | 10.88 b | 6.51bcd | 1.33 cd | 0.43 de | 2.20 cd | 0.087 b |
| NH4+ | 40.68 f | 6.08 d | 8.44 f | 7.33 d | 5.87 d | 1.21 d | 0.34 ef | 1.83 ef | 0.073 bc | |
| Urea | 44.03 e | 6.58 cd | 9.47 e | 11.08 b | 6.77 ab | 1.46 cd | 0.46 cd | 1.84 ef | 0.083 bc | |
| Gly | 34.28 g | 4.55 e | 7.39 g | 5.48 e | 5.03 e | 1.17 d | 0.25 f | 1.57 f | 0.063 c | |
| Interaction (N×G) | *** | * | * | ** | * | ns | * | * | ns | |
| LSD(0.05) values | 2.6590 | 0.8377 | 0.7825 | 0.8229 | 0.6683 | 0.4676 | 0.095 | 0.2822 | 0.0223 | |
Different letters within a column show significant difference (P< 0.05). * (0.05), ** (0.01) and *** (0.001), ns; non-significant.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



