
Submitted:
20 February 2024
Posted:
20 February 2024
You are already at the latest version
Abstract
This study aimed to evaluate the impact of Candida albicans on subgingival biofilm formation on dental implant surfaces. Scanning electron microscopy (SEM) and confocal laser scanning microscopy (CLSM) were used to compare biofilm structure and microbial biomass in the presence and absence of the fungus after periods of 24, 48, and 72 h. Quantitative polymerase chain reaction (qPCR) was used to quantify the number of viable and total micro-organisms for each of the biofilm-forming strains. A general linear model was applied to compare CLSM and qPCR results between control and test conditions. The biofilm developed with C. albicans at 72 h had a higher bacterial biomass, with a significantly higher cell viability (p < 0.05). After both 48 and 72 h of incubation, in the presence of C. albicans, there was a significant increase in counts of Fusobacterium nucleatum and Porphyromonas gingivalis and in the cell viability of Streptococcus oralis, Aggregatibacter actinomycetemcomitans, F. nucleatum, and P. gingivalis. Using a dynamic in vitro multispecies biofilm model, C. albicans exacerbated the development of the biofilm grown on dental implant surfaces, significantly increasing the number and cell viability of periodontal bacteria.
Keywords:
peri-implantitis
; oral biofilm
; Candida albicans
; scanning electron microscopy
; confocal laser scanning microscopy
; quantitative polymerase chain reaction
1. Introduction
The use of dental implants is currently the most widespread strategy for the rehabilitation of total or partial edentulism, resulting in long-term satisfactory success rates [1,2,3]. However, these implant-supported restorations are susceptible to complications during their function, both mechanical and biological. At the last World Workshop on the Classification of Periodontal and Peri-implant Diseases and Conditions (2017), peri-implant diseases were classified as peri-implant mucositis and peri-implantitis [4], with estimated prevalence ranges between 43% to 47% for peri-implant mucositis and 20% to 22% for peri-implantitis [5]. The primary etiological factor of peri-implantitis is the biofilms formed on dental implant and their restorative component surfaces, triggering a chronic inflammatory response, eventually leading to bone destruction and progressive loss of the implant osseointegration [4,6,7,8,9]. Although most of the studies have reported that oral bacteria are the main component of the submarginal biofilms associated with peri-implant diseases, viruses, protozoa, and, to a greater extent, fungi may also be part of the microbial communities within these biofilms [10,11,12,13].
The most frequent fungal pathogen in the oral cavity is Candida albicans, a dimorphic facultative anaerobic fungus, that is usually present as a yeast under favorable environments, although it usually presents as a filamentous fungus under unfavorable conditions [14]. These differential conditions are related to nutrient availability, environmental atmospheric composition or the presence of antifungal agents [15].
Presence of C. albicans in subgingival pockets has been reported in higher rates in subjects with periodontitis, compared with periodontally healthy individuals [16], although there is a high heterogeneity in the reported prevalence of C. albicans in the subgingival microbiota of periodontitis patients, ranging from 14.6% to 87.5% [17,18]. Similarly, in peri-implantitis, the presence of C. albicans is higher than around implants with healthy peri-implant tissues, or in peri-implant mucositis sites [19]. In different studies, C. albicans was detected in 27%, 15.8%, 77.6%, 76.2% of patients with peri-implantitis, versus 0%, 10%, 12.2%, 9.8% in patients with peri-implant health [7,20,21,22]. This high variability could be due to the different identification methods used in the different studies.
The pathogenicity of C. albicans is mediated by its adhesion to the implant surface in a process favored by salivary mucin and albumin [23]. Once adhered, its growth generates hyphae and secretes hydrolytic enzymes, basically proteases, lipases, and hemolysins, which may activate the inflammatory response of the host soft tissues [24,25,26]. The inflammatory response to C. albicans-infected epithelial cells is mediated by pro-inflammatory cytokines, which further contribute to the chronic inflammatory response characteristic of peri-implant diseases lesion and, eventually, to the tissue destruction and alveolar bone resorption, characteristic of peri-implantitis [27,28,29].
The pathogenic mechanisms of the potential interactions between C. albicans and the bacteria present in subgingival/submarginal biofilms have been mostly studied in vitro, using culturing or static biofilm models [30,31,32]. These studies have demonstrated that C. albicans influences biofilm architecture and favors the presence and virulence of certain periodontal pathogenic bacterial strains [33,34,35]. However, the overall effect of C. albicans on a multispecies biofilm or on biofilm formation on dental implant surfaces has not yet been studied. It was, therefore, the aim of this in-vitro study to evaluate the impact of C. albicans in the development, kinetics, structure, and viability of biofilm formation on dental implant surfaces, in a validated multispecies dynamic model.
2. Results
2.1. Scanning Electron Microscopy (SEM) analysis
Figure 1 depicts the SEM images of biofilms grown on the implant surfaces after 24, 48 and 72 h in the presence and absence of the fungus C. albicans, clearly showing a differential biofilm morphology. After 24 h of incubation, C. albicans presented as yeast (Figure 1G) and did not affect the structure of the biofilm, composed at this first stage by cocci (Streptococcus oralis and Veillonella parvula), and rods and spindle shaped bacteria, corresponding to Actinomyces naeslundii and Fusobacterium nucleatum (Figure 1A and Figure 1D).
At 48 h, C. albicans started to manifest as filaments, forming pseudohyphae to which F. nucleatum and cocci bacteria were anchored (Figure 1H). Compared to the 24 h biofilm, there was a higher density of spindle-shaped bacteria and cocci, corresponding to F. nucleatum and Aggregatibacter actinomycetemcomitans, respectively. In the test biofilms (with C. albicans) (Figure 2E), the presence of coccobacillary forms of Porphyromonas gingivalis was more frequent than in control biofilms (Figure 2B).
At 72 h, cellular aggregates formed by spindles, cocci, and coccobacilli (F. nucleatum, A. actinomycetemcomitans and P. gingivalis, respectively) were deposited on the pseudohyphae of C. albicans, presenting as a mixed mature biofilm with a higher biomass than in the previous time intervals (Figure 1I). The test biofilms demonstrated a higher bacterial density, as shown in Figure 1F and Figure 1C. In fact, control biofilms at 72 h presented a lower bacterial density than the test biofilms at 48 h.
2.2. Confocal Laser Scanning Microscopy (CLSM) analysis
C. albicans increased its size on the implant surface, as the biofilm was maturing after 24, 48 and 72 h of incubation (Figure 2 and Figure 3). Figure 2 A-C and Figure 2 D-F depict CLSM images representative of control and test biofilms, respectively, clearly showing the impact C. albicans on the biomass of the test biofilms. Figure 2 G-I show C. albicans in the test biofilms. The kinetics of the development of both biofilms are shown in Figure 3.
After the first 24 h, the bacterial biomass of biofilms grown in the absence of C. albicans (7.21 µm3/µm2 (standard deviation, SD = 4.50)) and in the presence of the fungus (6.63 µm3/µm2 (SD = 4.63)) showed no differences (Figure 2A and Figure 2D). The viability percentages, 60.20% (SD = 20.19%) and 63.37% (SD = 9.70%), respectively, were also similar (Figure 3). The biomass of C. albicans incorporated into the test biofilm was 5.95 µm3/µm2 (SD = 3.18) (Figure 2G).
Biofilms formed after 48 h also did not show differences between test and control biofilms. Control biofilms had a bacterial biomass of 10.02 µm3/µm2 (SD = 6.21) and test ones of 9.69 µm3/µm2 (SD = 2.14) (Figure 2B and Figure 2E). There were also no differences between cell viability of the two biofilms, 52.47% (SD = 5.00%) for control biofilms and 58.50% (SD = 18.07%) for test biofilms (Figure 3). C. albicans had a biomass of 8.86 µm3/µm2 (SD = 2.78) (Figure 2H).
In contrast, the mature biofilms developed in the presence of C. albicans after 72 h of incubation showed higher bacterial biomass than control biofilms (12.88 µm3/µm2 (SD = 8.77) and 8.31 µm3/µm2 (SD = 4.70, respectively)). Furthermore, as seen in Figure 2C, Figure 2F, and Figure 3, the bacterial viability of the biofilms developed without the fungus was 31.34% (SD = 9.81%), while in the mixed biofilm, it was 66.70% (SD = 10.05%), a difference that was statistically significant (p<0.05). Figure 2I shows the biomass corresponding to C. albicans, which was 18.91 µm3/µm2 (SD = 6.31).
2.3. Quantitative Polymerase Chain Reaction (qPCR) analysis
Table 1 shows counts of total and viable cells in biofilms developed at 24, 48 and 72 h, expressed as colony forming units (CFU)/mL for each bacterial species and C. albicans in test biofilms, together with cell viability, and the percentage of live cells to total counts.
At 24 h of incubation, the growth and viability of C. albicans was limited. There were no statistically significant differences in counts for any of the six biofilm-forming bacterial species. When comparing test and control biofilms.
At 48 h, the development of C. albicans increased, and in the test biofilms, counts and viability of F. nucleatum and P. gingivalis were significantly higher. The same pattern was observed for A. naeslundii. Cell viability of A. actinomycetemcomitans was also significantly higher in test biofilms.
At 72 h, a similar pattern occurred, with higher growth of C. albicans, when compared to previous intervals, and larger counts of F. nucleatum, P. gingivalis and A. naeslundii in test biofilms when compared to control biofilms. At this stage, also the number of viable cells of S. oralis, F. nucleatum, P. gingivalis and A. actinomycetemcomitans were significantly higher compared to the controls.
3. Discussion
In the present study, a validated in vitro multispecies dynamic biofilm model was used to assess the influence of C. albicans on biofilms developed on dental implant surfaces. Bacterial counts determined by qPCR analysis indicated that, after 48 and 72 h of growth, the number and cell viability of F. nucleatum and P. gingivalis were significantly higher in biofilms developed in the presence of the fungus. Similarly, the proportion of live cells of A. actinomycetemcomitans and S. oralis also increased significantly in the presence of C. albicans in mature biofilms (72 h.) (Table 1). Similar results were obtained by CLSM analyses, depicting a significantly higher overall size and cell viability in mature biofilms (72 h) in the presence of C. albicans (Figure 2 and Figure 3).
The progressive filamentation of C. albicans cells observed in the SEM analysis (Figure 1) may have been favored by the experimental conditions of the biofilm model used, since they simulate oral cavity conditions (pH 7, 37°C, anaerobic environment) and the presence of Gram-negative bacteria (V. parvula. F. nucleatum. P. gingivalis and A. actinomycetemcomitans) [15]. In the maturation of C. albicans-associated biofilms, the filamentation process led to the attachment to implant surfaces of hyphae and yeast-like (sessile) cells, associated with microcolonies of rods and spindle shaped bacteria, embedded in an extracellular matrix. This morphology coincides with other previous descriptions of C. albicans-associated biofilms [36]. The impact of C. albicans on biofilm formation shown in the present study, demonstrating significantly higher biofilm biomass and higher percentages/counts of total and viable bacterial strains, may be exacerbated by the demonstrated activation in the expression of hydrolytic enzymes by C. albicans in presence of periodontal bacteria, what may further compromise the host immune response and enhance the resistance of the resulting biofilm to antifungal agents [30,37].
One of the possible relevant findings of the present study is the specific impact of C. albicans on the percentage of viable cells of P. gingivalis (Table 1). This effect may be due to the enhanced anaerobic environment generated by the fungal hyphae, clearly depicted within the biofilm architecture shown by SEM. Also, Interlin InlJ has been involved in the expression of P. gingivalis genes responsible for the interaction with C. albicans hyphae [38]. It has also been reported that adhesins Als3 and the proteases Sap6 and Sap9 of C. albicans, together with the gingipains of P. gingivalis may favor the invasion of these microorganisms in epithelial cells and fibroblasts [33,39]. Along the same lines, citrullination, mediated by peptidyl arginine deiminase (PPAD) secreted by P. gingivalis, may favor the adhesion of this bacterial species to the cell wall of C. albicans, thus increasing its viability under aerobic conditions [40]. The competition for iron sources that may occur between P. gingivalis and C. albicans, under conditions such as those of the present study where this nutrient is limited, may also explain the increased viability of the bacteria in the mixed biofilm. Furthermore, this competition may also favor the resistance of P. gingivalis to antimicrobial substances by increasing the expression of virulence genes [41]. In contrast to the results from the present investigation, Cavalcanti et al. (2016) reported that P. gingivalis exerted an opposite influence on C. albicans by inhibiting its hyphal production. In the model used in the present investigation, the concomitant presence of Streptococcus and Actinomyces species may have reverted this inhibition [30,31]. In fact, other authors have argued that the effect of this interaction is dependent on the fungal strain, the composition of the medium, and the streptococcal population present [42].
C. albicans also significantly increased the vitality of F. nucleatum in the multispecies biofilm (Table 1), what could be due to the interaction between the bacterial adhesin radD with the fungal cell wall mannoprotein FLO9, thus facilitating a specific dual aggregation [43]. This increased growth may enhance the bridging role of F. nucleatum between primary colonizers and the late colonizers P. gingivalis and A. actinomycetemcomitans, and effect that has already been attributed to C. albicans [35]. Conversely, another in vitro study indicated that F. nucleatum could inhibit the filamentation process of C. albicans by limiting its ability to kill macrophages and, thus, attenuating its pathogenic potential [44]. Similarly, the presence of A. actinomycetemcomitans through its autoinducer Quorum Sensing-2 (AI-2) molecule inhibits fungal hyphal formation and C. albicans aggregation [32]. However, the quantitative results from the present study indicated that C. albicans increased the survival rate of A. actinomycetemcomitans in mature biofilms. This phenomenon may suggest that the protective anaerobic environment generated by the hyphae, and the consequent increased development of F. nucleatum spindles, would favor the survival of A. actinomycetemcomitans in mature biofilms. Further studies are needed to elucidate this specific dual interaction.
The increase of live cells of the initial colonizer S. oralis in the mature biofilm was also favored by C. albicans. This effect can be explained by the binding of the cocci to the gtfR glucan-binding domain, the main component of the cell wall of C. albicans [45]. S. oralis is also supposed to induce filamentation of C. albicans, which may enhance the invasiveness of fungal and bacterial cells into host epithelial cells [46,47].
Consistent with the results from the present investigation, in vivo studies have also reported that C. albicans may exert an important pathogenic effect in the later stages of peri-implantitis, when the biofilm is already established [30]. In fact, case-control studies have demonstrated a higher presence of C. albicans in the peri-implant sulcus of patients with peri-implantitis, compared with those with healthy peri-implant tissues [48]. Similarly, the presence of hyphae in connective tissue specimens of peri-implantitis has been demonstrated in association with P. gingivalis, A. actinomycetemcomitams and P. intermedia [49], as well as with V. parvula, Tannerella forsythia and Parvimonas micra [50]. A deeper understanding of the interactions of C. albicans with the virulence of the different individual bacterial species within the subgingival/submarginal biofilms may help to better understand its pathogenicity and its resistance to antimicrobial strategies. For example, the β,1-3 glucan in the cell wall of C. albicans has shown to modulate the tolerance of periodontal bacterial anaerobes to different antibiotics [51]. This knowledge may also help to design more effective preventive strategies, such those based on the use of pre- or probiotics [31]
The experimental procedures used for the development of the present study are not free of limitations, that should be acknowledged. First, although the biofilm model attempts to mimic the conditions of the oral cavity, there are specific individual variables that cannot be reproduced. In addition, natural subgingival/submarginal biofilms may be composed of hundreds of species, whereas the model used is composed of six bacterial species that are intended to be representative of different types of colonizers. Finally, the accuracy of the data obtained is limited due to the high experimental variability linked to in vitro work with live microorganisms.
Considering the acknowledged limitations, the statistical evaluation of the obtained results allows to conclude that C. albicans has a significant impact on the growth, dynamics, structure and viability of subgingival/submarginal biofilms formed on implant surfaces, favoring an increase in the development of P. gingivalis, F. nucleatum, A. actinomycetemcomitans and S. oralis.
4. Materials and Methods
4.1. Microbial strains and culture conditions
Bacterial strains Streptococcus oralis CECT 907T, Actinomyces naeslundii ATCC 19039, Veillonella parvula NCTC 11810, Fusobacterium nucleatum DMSZ 20482, Porphyromonas gingivalis ATCC 33277 and Aggregatibacter actinomycetemcomitans DSMZ 8324 were used. They were grown on blood agar plates (Blood Agar Oxoid No 2; Oxoid, Basingstoke, UK), supplemented with 5% (v/v) sterile horse blood (Oxoid), 5.0 mg/L haemin (Sigma, St. Louis, MO, USA) and 1.0 mg/L menadione (Merck, Darmstadt, Germany) at 37°C for 24-72 h under anaerobic conditions (10% H2, 10% CO2 and N2 balance). The fungal strain Candida albicans SC 5314 was grown on yeast-peptone-glucose (YPD) agar plates (2% glucose (Panreac, Barcelona, Spain), 2% peptone (Life Technologies, Detroit, MI, USA), 1% peptone yeast extract (Life Technologies, Detroit, MI, USA) and 2% agar (Becton, Dickinson and Company, Sparks, MD, USA)) at 37°C for 24 h under aerobic conditions.
Pure cultures of each strain were grown for 24 h under anaerobic conditions in protein-enriched brain heart infusion (BHI) medium (Becton, Dickinson and Company, Franklin Lakes, NJ, USA), supplemented with 2.5 g/L mucin (Oxoid), 1.0 g/L yeast extract (Oxoid), 0.1 g/L cysteine (Sigma), 2.0 g/L sodium bicarbonate (Merck), 5.0 mg/L haemin (Sigma), 1.0 mg/L menadione (Merck) and 0.25 % (v/v) glutamic acid (Sigma). After incubation, microbial growth was measured spectrophotometrically to develop a microbial suspension containing 106 colony-forming units (CFU)/mL of each bacterium and, where appropriate, 104 CFU/mL of C. albicans.
4.2. In vitro dynamic multispecies biofilm model
An in vitro multispecies dynamic biofilm model was used [52,53], which has been validated on biofilms growing on implant surfaces [54,55]. Basically, the system consists of a sterile vessel, where the liquid culture medium, namely the previously described protein-enriched BHI medium, is pumped into the bioreactor by means of a peristaltic pump at constant pressure. The bioreactor (Lambda Minifor© bioreactor, LAMBDA Laboratory Instruments, Sihlbruggstrasse, Switzerland) maintains the culture medium under stable conditions at 37°C, pH 7.2, and an anaerobic atmosphere (10% H2, 10% CO2 and N2 balance) during the whole incubation process. These conditions are maintained by directly pumping an anaerobic gas mixture (10% H2, 10% CO2 and equilibrium N2) through a filter into the incubation vessel, keeping the pressure constant. The system is inoculated with 5 mL of the previously described microbial suspension and maintained for 12 h under the described conditions. Subsequently, once the mixed culture reached the exponential growth phase, the continuous culture is activated through a second peristaltic pump with a flow rate of 30 mL/h to transfer the culture to Robbins devices placed in series that carry the sterile dental implant units on which the biofilm was developed (Straumann® Tissue Level Standard, 8 mm in length and 3.3 mm in diameter, with the patented moderately rough sandblasted and acid-etched surface [Straumann Institute AG, Basel, Switzerland]). Inside the Robbins device, anaerobic conditions and constant temperature (37°C) are maintained at during each experimental interval to allow biofilm development.
4.3. Experimental groups
To evaluate the effect of C. albicans on the dynamics of subgingival biofilm formation on implant surfaces, three time-intervals were analyzed: 24, 48, and 72 h. For each time interval, the developed biofilms were incubated under two different conditions, the test biofilms included a mixed culture composed of the bacterial strains S. oralis, A. naeslundii, V. parvula, F. nucleatum, P. gingivalis, A. actinomycetemcomitans, and the fungus C. albicans, while the control biofilms included only from the six bacterial strains. At each time and in each condition, three implants were analyzed by confocal microscopy (CLSM) (n=3), three by scanning electron microscopy (SEM) (n=3), and nine by real-time polymerase chain reaction (qPCR) (n=9).
4.4. Scanning Electron Microscopy (SEM)
After removal of the implants from the Robbins device, the implants were sequentially washed three times with 2 ml of phosphate-buffered saline (PBS) (immersion time per rinse, 10 s) to remove unattached bacteria. The implants were then fixed in a solution of 4% paraformaldehyde (Panreac Química, Barcelona, Spain) and 2.5% glutaraldehyde (Panreac Química) for 4 h, at 4ºC. They were then washed in PBS and sterile water (immersion time per wash 10 min) and dehydrated through a series of graded ethanol solutions (30%, 50%, 70%, 80%, 90% and 100%; immersion time per series 10 min). Then the specimens were dried, coated with gold, and analyzed using a JSM 6400 electron microscope (JSM6400, JEOL, Tokyo, Japan), with a backscatter electron detector and an image resolution of 25 kV.
This analysis was carried out at the National Centre of Electron Microscopy (Instalación Científico-Técnico singular; ICTS) at the Moncloa Campus of the Complutense University of Madrid (Madrid, Spain).
4.5. Confocal Laser Scanning Microscopy (CSLM)
For the non-invasive confocal imaging of biofilms, an Olympus Ix83 inverted microscope coupled to an Olympus® FV1200 confocal system (Olympus; Shinjuku, Tokyo, Japan) was used. The CLSM software was configured to take a z-series of scans (XYZ) of 1 µm thickness (8 bits, 512x512 pixels).
Prior to the microscopic analysis, the Robbins device was taken from the bioreactor and carefully removed the implants, which were then washed three times with 2 mL PBS (immersion time per rinse, 10 s) to remove unattached bacteria.
For observing and quantifying the biofilm bacteria, the samples were stained with the LIVE/DEAD® BacLightTM bacterial viability kit solution (Molecular Probes, The Netherlands), which contains propidium iodide (PI) and SYTO9 nucleic acid dyes. With this method, dead cells or those with a compromised viability are stained in red (PI), while cells with an intact membrane are stained in green (SYTO9). Implants were then coated with fluorochromes in a 1:1 ratio and incubated for 9 ± 1 min to obtain the optimal fluorescence signal at the corresponding wavelengths (SYTO9: 515-530 nm; PI: >600 nm). To observe and quantify C. albicans implants were stained for 10 min with 3% Calcofluor White (CFW), thus obtaining an optimal signal using a wavelength of 405 nm.
Representative implant surface locations involving both the peak of a thread and the bottom of the valley were selected for the CLSM analyses.
The COMSTAT software (www.comstat.dk) was used to calculate the biomass in micrometres3/micrometres2 (µm3/µm2) of the CLSM images.
The analysis was performed at the Biological Research Centre Margarita Salas (Centro de Investigaciones Biológicas, Consejo Superior de Investigaciones Científicas - CIB-CSIC), located at the Moncloa Campus of the Complutense University of Madrid (Madrid, Spain).
4.6. Quantitative Polymerase Chain Reaction (qPCR)
Prior to DNA isolation, the test and control implants were rinsed sequentially in 2 ml sterile PBS three times (immersion time per rinse 10 s) to remove unattached bacteria. To disaggregate the biofilms, implants immersed in 1 mL sterile PBS were vortexed at maximum power at room temperature for 2 min. To exclude genetic material from non-viable cells, 100 µL of the obtained suspension was incubated with 100 µM PMA (propidium monoazide) prior to DNA extraction [53] and 100 µL of the same suspension was analyzed without the PMA treatment to calculate the viability percentages of each strain.
DNA was isolated using the commercial MolYsisComplete5 kit, Molzym (GmbH & CoKG, Bremen, Germany), according to the manufacturer's instructions. Primers and probes were supplied by Life Technologies Invitrogen (Carlsbad, CA, USA), Applied Biosystems (Carlsbad, CA, USA) and Roche (Roche Diagnostic GmbH, Mannheim, Germany).
The amplification reaction was performed in a total mix volume of 10 μL. Reaction mixtures contained 5 μL of Master Mix 2x (LC 480 Probes Master, Roche), optimal concentrations of primers and probe (900, 900 and 300 nM for S. oralis; 300, 300 and 300 nM for A. naeslundii and P. gingivalis; 750, 750 and 400 nM for V. parvula; 300, 300 and 200 nM for A. actinomycetemcomitans; and 600, 600 and 300 nM for F. nucleatum), and 2.5 μL of DNA extracted from the samples. The negative control was 2.5 μL of sterile water [non-template control (NTC)] (Roche). The primers and probes used were previously described [53].
The target used for the detection and quantification of the six selected bacterial species was the 16S rRNA gene of each of them. For the detection and quantification of C. albicans, an optimization process of a qPCR targeting the ribosomal internal transcribed spacer (ITS) region was previously performed. The primers and probe designed by He et al. (Forward: 5′-GGT GTT GAG GAG CAA TAC GAC-3′; Reverse: 5′-AGA CCT AAG CCA TTG TCA-3′; Probe: 5′-FAM-ATC CCG CCT TAC CAC TAC CG-TAMRA-3′) were used [56]. Primer concentrations of 600 and 600 nM and probe concentration of 300 nM were set as optimal. generating a standard curve whose equation was y = -3.3598x + 42.944, R2 = 0.9996, and a detection limit set at 102 colony-forming units (CFU)/mL, with a 95% confidence interval. No cross-reaction with DNA from the bacterial strains used was observed.
The amplification program consisted of an initial cycle at 95°C for 10 min, followed by 40 cycles at 95°C for 15 s and 60°C for 1 min. It was performed on a LightCycler® 480 II thermal cycler (Roche Diagnostic GmbH, Mannheim, Germany). The microplates used for qPCR were LightCycler480 Multiwell-384 (Roche).
Each DNA sample was analyzed in duplicate. The quantification cycle (Cq) values were determined using the provided software (LC 480 Software 1.5, Roche). Quantification of cells was based on extrapolation with previously designed standard curves with the Cq values generated in qPCR vs log CFU/mL. Correlation between Cq and CFU/mL values was automatically generated by the software (LC 480 Software 1.5, Roche).
4.7. Statistical analysis
Quantitative data was expressed in colony-forming units per millilitre (CFU/mL) (qPCR) and bacterial biomass obtained by CLSM was expressed in µm3/µm2. Data were reported as means and standard deviations (SDs)) and the Shapiro-Wilk goodness-of-fit tests was used to assess data normality. When the two sets of data compared showed a normal distribution, a T-test with Welch's correction was applied. When at least one of the two groups did not show a normal distribution, a Mann Whitney test was applied. Statistically significant differences were considered for p-values <0.05. A GraphPad Prism software was used for all data analysis.
Author Contributions
E.B., D.H. and M.S. designed the study. E.B. have performed the experiments and analyzed and interpreted the collected data. E.B. and M.A. performed the statistical analysis. E.B. has drafted the manuscript. M.A., H.R-V., D.H. and M.S. have critically revised the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
Internal funds from the ETEP Research Group were used for this investigation.
Acknowledgments
The authors would like to acknowledge the technical support of A.M. Vicente, at the ICTS National Centre of Electron Microscopy (University Complutense, Madrid, Spain). M.G. Elvira and M.T. Seisdedos, at the Confocal and Fluorescence Microscopy Service of Margarita Salas Biological Research Center (Superior Center for Scientific Research) and R. Ayuso, at the Platform of Mechanic Workshops (University Complutense, Madrid, Spain). We also thank J. Arroyo and R. García, at the Faculty of Pharmacy (University Complutense, Madrid, Spain), for the loan of the SC 5314 strain of Candida albicans. M. Arce is a recipient of scholarship ANID 21221003 from the Chilean Government and Osteology Scholar 2023 for the Osteology Foundation.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Buser, D.; Janner, S.F.; Wittneben, J.G.; Bragger, U.; Ramseier, C.A.; Salvi, G.E. 10-year survival and success rates of 511 titanium implants with a sandblasted and acid-etched surface: a retrospective study in 303 partially edentulous patients. Clin Implant Dent Relat Res 2012, 14, 839–851. [Google Scholar] [CrossRef]
- Buser, D.; Sennerby, L.; De Bruyn, H. Modern implant dentistry based on osseointegration: 50 years of progress, current trends and open questions. Periodontol 2000 2017, 73, 7–21. [Google Scholar] [CrossRef]
- Gotfredsen, K. A 10-year prospective study of single tooth implants placed in the anterior maxilla. Clin Implant Dent Relat Res 2012, 14, 80–87. [Google Scholar] [CrossRef]
- Berglundh, T.; Armitage, G.; Araujo, M.G.; Avila-Ortiz, G.; Blanco, J.; Camargo, P.M.; Chen, S.; Cochran, D.; Derks, J.; Figuero, E.; et al. Peri-implant diseases and conditions: Consensus report of workgroup 4 of the 2017 World Workshop on the Classification of Periodontal and Peri-Implant Diseases and Conditions. J Periodontol 2018, 89 Suppl 1, S313–S318. [Google Scholar] [CrossRef]
- Herrera, D.; Berglundh, T.; Schwarz, F.; Chapple, I.; Jepsen, S.; Sculean, A.; Kebschull, M.; Papapanou, P.N.; Tonetti, M.S.; Sanz, M.; et al. Prevention and treatment of peri-implant diseases-The EFP S3 level clinical practice guideline. J Clin Periodontol 2023, 50 Suppl 26, 4–76. [Google Scholar] [CrossRef]
- Derks, J.; Tomasi, C. Peri-implant health and disease. A systematic review of current epidemiology. J Clin Periodontol 2015, 42 Suppl 16, S158–171. [Google Scholar] [CrossRef]
- Leonhardt, A.; Renvert, S.; Dahlen, G. Microbial findings at failing implants. Clin Oral Implants Res 1999, 10, 339–345. [Google Scholar] [CrossRef]
- Burgers, R.; Gerlach, T.; Hahnel, S.; Schwarz, F.; Handel, G.; Gosau, M. In vivo and in vitro biofilm formation on two different titanium implant surfaces. Clin Oral Implants Res 2010, 21, 156–164. [Google Scholar] [CrossRef] [PubMed]
- Ferreira Ribeiro, C.; Cogo-Muller, K.; Franco, G.C.; Silva-Concilio, L.R.; Sampaio Campos, M.; de Mello Rode, S.; Claro Neves, A.C. Initial oral biofilm formation on titanium implants with different surface treatments: An in vivo study. Arch Oral Biol 2016, 69, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Dewhirst, F.E.; Chen, T.; Izard, J.; Paster, B.J.; Tanner, A.C.; Yu, W.H.; Lakshmanan, A.; Wade, W.G. The human oral microbiome. J Bacteriol 2010, 192, 5002–5017. [Google Scholar] [CrossRef] [PubMed]
- Kato, A.; Imai, K.; Sato, H.; Ogata, Y. Prevalence of Epstein-Barr virus DNA and Porphyromonas gingivalis in Japanese peri-implantitis patients. BMC Oral Health 2017, 17, 148. [Google Scholar] [CrossRef]
- Montelongo-Jauregui, D.; Srinivasan, A.; Ramasubramanian, A.K.; Lopez-Ribot, J.L. An In Vitro Model for Candida albicans(-)Streptococcus gordonii Biofilms on Titanium Surfaces. J Fungi (Basel) 2018, 4. [Google Scholar] [CrossRef]
- Sahrmann, P.; Gilli, F.; Wiedemeier, D.B.; Attin, T.; Schmidlin, P.R.; Karygianni, L. The Microbiome of Peri-Implantitis: A Systematic Review and Meta-Analysis. Microorganisms 2020, 8. [Google Scholar] [CrossRef]
- Ghannoum, M.A.; Jurevic, R.J.; Mukherjee, P.K.; Cui, F.; Sikaroodi, M.; Naqvi, A.; Gillevet, P.M. Characterization of the oral fungal microbiome (mycobiome) in healthy individuals. PLoS Pathog 2010, 6, e1000713. [Google Scholar] [CrossRef]
- Chow, E.W.L.; Pang, L.M.; Wang, Y. From Jekyll to Hyde: The Yeast-Hyphal Transition of Candida albicans. Pathogens 2021, 10. [Google Scholar] [CrossRef] [PubMed]
- Canabarro, A.; Valle, C.; Farias, M.R.; Santos, F.B.; Lazera, M.; Wanke, B. Association of subgingival colonization of Candida albicans and other yeasts with severity of chronic periodontitis. J Periodontal Res 2013, 48, 428–432. [Google Scholar] [CrossRef] [PubMed]
- Urzua, B.; Hermosilla, G.; Gamonal, J.; Morales-Bozo, I.; Canals, M.; Barahona, S.; Coccola, C.; Cifuentes, V. Yeast diversity in the oral microbiota of subjects with periodontitis: Candida albicans and Candida dubliniensis colonize the periodontal pockets. Med Mycol 2008, 46, 783–793. [Google Scholar] [CrossRef] [PubMed]
- Arumugam, M.; Iyer, S.S. A Comparative Evaluation of Subgingival Occurrence of Candida Species in Chronic Periodontitis and Peri-implantitis: A Clinical and Microbiological Study. International Journal of Clinical Implant Dentistry With Dvd 2015, 1, 95–100. [Google Scholar] [CrossRef]
- Salvi, G.E.; Cosgarea, R.; Sculean, A. Prevalence and Mechanisms of Peri-implant Diseases. J Dent Res 2017, 96, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, F.; Becker, K.; Rahn, S.; Hegewald, A.; Pfeffer, K.; Henrich, B. Real-time PCR analysis of fungal organisms and bacterial species at peri-implantitis sites. Int J Implant Dent 2015, 1, 9. [Google Scholar] [CrossRef] [PubMed]
- Alrabiah, M.; Alshagroud, R.S.; Alsahhaf, A.; Almojaly, S.A.; Abduljabbar, T.; Javed, F. Presence of Candida species in the subgingival oral biofilm of patients with peri-implantitis. Clin Implant Dent Relat Res 2019, 21, 781–785. [Google Scholar] [CrossRef]
- Alsahhaf, A.; Al-Aali, K.A.; Alshagroud, R.S.; Alshiddi, I.F.; Alrahlah, A.; Abduljabbar, T.; Javed, F.; Vohra, F. Comparison of yeast species in the subgingival oral biofilm of individuals with type 2 diabetes and peri-implantitis and individuals with peri-implantitis without diabetes. J Periodontol 2019, 90, 1383–1389. [Google Scholar] [CrossRef]
- Samaranayake, L.P.; McCourtie, J.; MacFarlane, T.W. Factors affecting th in-vitro adherence of Candida albicans to acrylic surfaces. Arch Oral Biol 1980, 25, 611–615. [Google Scholar] [CrossRef] [PubMed]
- Berman, J.; Sudbery, P.E. Candida Albicans: a molecular revolution built on lessons from budding yeast. Nat Rev Genet 2002, 3, 918–930. [Google Scholar] [CrossRef] [PubMed]
- Mayer, F.L.; Wilson, D.; Hube, B. Candida albicans pathogenicity mechanisms. Virulence 2013, 4, 119–128. [Google Scholar] [CrossRef]
- De-La-Torre, J.; Quindos, G.; Marcos-Arias, C.; Marichalar-Mendia, X.; Gainza, M.L.; Eraso, E.; Acha-Sagredo, A.; Aguirre-Urizar, J.M. Oral Candida colonization in patients with chronic periodontitis. Is there any relationship? Rev Iberoam Micol 2018, 35, 134–139. [Google Scholar] [CrossRef]
- Lamster, I.B.; Grbic, J.T.; Mitchell-Lewis, D.A.; Begg, M.D.; Mitchell, A. New concepts regarding the pathogenesis of periodontal disease in HIV infection. Ann Periodontol 1998, 3, 62–75. [Google Scholar] [CrossRef] [PubMed]
- Martin, R.; Wachtler, B.; Schaller, M.; Wilson, D.; Hube, B. Host-pathogen interactions and virulence-associated genes during Candida albicans oral infections. Int J Med Microbiol 2011, 301, 417–422. [Google Scholar] [CrossRef]
- Dongari-Bagtzoglou, A.; Kashleva, H.; Villar, C.C. Bioactive interleukin-1alpha is cytolytically released from Candida albicans-infected oral epithelial cells. Med Mycol 2004, 42, 531–541. [Google Scholar] [CrossRef]
- Cavalcanti, Y.W.; Wilson, M.; Lewis, M.; Del-Bel-Cury, A.A.; da Silva, W.J.; Williams, D.W. Modulation of Candida albicans virulence by bacterial biofilms on titanium surfaces. Biofouling 2016, 32, 123–134. [Google Scholar] [CrossRef]
- Morse, D.J.; Wilson, M.J.; Wei, X.; Bradshaw, D.J.; Lewis, M.A.O.; Williams, D.W. Modulation of Candida albicans virulence in in vitro biofilms by oral bacteria. Lett Appl Microbiol 2019, 68, 337–343. [Google Scholar] [CrossRef] [PubMed]
- Bachtiar, E.W.; Bachtiar, B.M.; Jarosz, L.M.; Amir, L.R.; Sunarto, H.; Ganin, H.; Meijler, M.M.; Krom, B.P. AI-2 of Aggregatibacter actinomycetemcomitans inhibits Candida albicans biofilm formation. Front Cell Infect Microbiol 2014, 4, 94. [Google Scholar] [CrossRef] [PubMed]
- Bartnicka, D.; Karkowska-Kuleta, J.; Zawrotniak, M.; Satala, D.; Michalik, K.; Zielinska, G.; Bochenska, O.; Kozik, A.; Ciaston, I.; Koziel, J.; et al. Adhesive protein-mediated cross-talk between Candida albicans and Porphyromonas gingivalis in dual species biofilm protects the anaerobic bacterium in unfavorable oxic environment. Sci Rep 2019, 9, 4376. [Google Scholar] [CrossRef]
- Guo, Y.; Wei, C.; Liu, C.; Li, D.; Sun, J.; Huang, H.; Zhou, H. Inhibitory effects of oral Actinomyces on the proliferation, virulence and biofilm formation of Candida albicans. Arch Oral Biol 2015, 60, 1368–1374. [Google Scholar] [CrossRef]
- Thurnheer, T.; Karygianni, L.; Flury, M.; Belibasakis, G.N. Fusobacterium Species and Subspecies Differentially Affect the Composition and Architecture of Supra- and Subgingival Biofilms Models. Front Microbiol 2019, 10, 1716. [Google Scholar] [CrossRef]
- Finkel, J.S.; Mitchell, A.P. Genetic control of Candida albicans biofilm development. Nat Rev Microbiol 2011, 9, 109–118. [Google Scholar] [CrossRef]
- Schaller, M.; Borelli, C.; Korting, H.C.; Hube, B. Hydrolytic enzymes as virulence factors of Candida albicans. Mycoses 2005, 48, 365–377. [Google Scholar] [CrossRef] [PubMed]
- Sztukowska, M.N.; Dutton, L.C.; Delaney, C.; Ramsdale, M.; Ramage, G.; Jenkinson, H.F.; Nobbs, A.H.; Lamont, R.J. Community Development between Porphyromonas gingivalis and Candida albicans Mediated by InlJ and Als3. mBio 2018, 9. [Google Scholar] [CrossRef]
- Tamai, R.; Sugamata, M.; Kiyoura, Y. Candida albicans enhances invasion of human gingival epithelial cells and gingival fibroblasts by Porphyromonas gingivalis. Microb Pathog 2011, 51, 250–254. [Google Scholar] [CrossRef]
- Karkowska-Kuleta, J.; Bartnicka, D.; Zawrotniak, M.; Zielinska, G.; Kieronska, A.; Bochenska, O.; Ciaston, I.; Koziel, J.; Potempa, J.; Baster, Z.; et al. The activity of bacterial peptidylarginine deiminase is important during formation of dual-species biofilm by periodontal pathogen Porphyromonas gingivalis and opportunistic fungus Candida albicans. Pathog Dis 2018, 76. [Google Scholar] [CrossRef]
- Guo, Y.; Wang, Y.; Wang, Y.; Jin, Y.; Wang, C. Heme Competition Triggers an Increase in the Pathogenic Potential of Porphyromonas gingivalis in Porphyromonas gingivalis-Candida albicans Mixed Biofilm. Front Microbiol 2020, 11, 596459. [Google Scholar] [CrossRef] [PubMed]
- Arzmi, M.H.; Dashper, S.; Catmull, D.; Cirillo, N.; Reynolds, E.C.; McCullough, M. Coaggregation of Candida albicans, Actinomyces naeslundii and Streptococcus mutans is Candida albicans strain dependent. FEMS Yeast Res 2015, 15, fov038. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Cen, L.; Kaplan, C.; Zhou, X.; Lux, R.; Shi, W.; He, X. Cellular Components Mediating Coadherence of Candida albicans and Fusobacterium nucleatum. J Dent Res 2015, 94, 1432–1438. [Google Scholar] [CrossRef] [PubMed]
- Bor, B.; Cen, L.; Agnello, M.; Shi, W.; He, X. Morphological and physiological changes induced by contact-dependent interaction between Candida albicans and Fusobacterium nucleatum. Sci Rep 2016, 6, 27956. [Google Scholar] [CrossRef] [PubMed]
- Souza, J.G.S.; Bertolini, M.; Thompson, A.; Mansfield, J.M.; Grassmann, A.A.; Maas, K.; Caimano, M.J.; Barao, V.A.R.; Vickerman, M.M.; Dongari-Bagtzoglou, A. Role of glucosyltransferase R in biofilm interactions between Streptococcus oralis and Candida albicans. ISME J 2020, 14, 1207–1222. [Google Scholar] [CrossRef]
- Xu, H.; Sobue, T.; Bertolini, M.; Thompson, A.; Dongari-Bagtzoglou, A. Streptococcus oralis and Candida albicans Synergistically Activate mu-Calpain to Degrade E-cadherin From Oral Epithelial Junctions. J Infect Dis 2016, 214, 925–934. [Google Scholar] [CrossRef]
- Bertolini, M.M.; Xu, H.; Sobue, T.; Nobile, C.J.; Del Bel Cury, A.A.; Dongari-Bagtzoglou, A. Candida-streptococcal mucosal biofilms display distinct structural and virulence characteristics depending on growth conditions and hyphal morphotypes. Mol Oral Microbiol 2015, 30, 307–322. [Google Scholar] [CrossRef]
- Rosa, E.A.R.; Rached, R.N.; Ignacio, S.A.; Rosa, R.T.; Jose da Silva, W.; Yau, J.Y.Y.; Samaranayake, L.P. Phenotypic evaluation of the effect of anaerobiosis on some virulence attributes of Candida albicans. J Med Microbiol 2008, 57, 1277–1281. [Google Scholar] [CrossRef]
- Reynaud, A.H.; Nygaard-Østby, B.; Bøygard, G.K.; Eribe, E.R.; Olsen, I.; Gjermo, P. Yeasts in periodontal pockets. J Clin Periodontol 2001, 28, 860–864. [Google Scholar] [CrossRef]
- Schwarz, F.; Derks, J.; Monje, A.; Wang, H.L. Peri-implantitis. J Periodontol 2018, 89 Suppl 1, S267–s290. [Google Scholar] [CrossRef]
- Kong, E.F.; Tsui, C.; Kucharikova, S.; Andes, D.; Van Dijck, P.; Jabra-Rizk, M.A. Commensal Protection of Staphylococcus aureus against Antimicrobials by Candida albicans Biofilm Matrix. mBio 2016, 7. [Google Scholar] [CrossRef]
- Blanc, V.; Isabal, S.; Sanchez, M.C.; Llama-Palacios, A.; Herrera, D.; Sanz, M.; Leon, R. Characterization and application of a flow system for in vitro multispecies oral biofilm formation. J Periodontal Res 2014, 49, 323–332. [Google Scholar] [CrossRef]
- Sanchez, M.C.; Llama-Palacios, A.; Fernandez, E.; Figuero, E.; Marin, M.J.; Leon, R.; Blanc, V.; Herrera, D.; Sanz, M. An in vitro biofilm model associated to dental implants: structural and quantitative analysis of in vitro biofilm formation on different dental implant surfaces. Dent Mater 2014, 30, 1161–1171. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, M.C.; Alonso-Espanol, A.; Ribeiro-Vidal, H.; Alonso, B.; Herrera, D.; Sanz, M. Relevance of Biofilm Models in Periodontal Research: From Static to Dynamic Systems. Microorganisms 2021, 9. [Google Scholar] [CrossRef] [PubMed]
- Alonso-Espanol, A.; Bravo, E.; Ribeiro-Vidal, H.; Virto, L.; Herrera, D.; Alonso, B.; Sanz, M. The Antimicrobial Activity of Curcumin and Xanthohumol on Bacterial Biofilms Developed over Dental Implant Surfaces. Int J Mol Sci 2023, 24. [Google Scholar] [CrossRef] [PubMed]
- He, Z.X.; Zhao, H.H.; Wang, F.K. PCR-detectable Candida DNA exists a short period in the blood of systemic candidiasis murine model. Open Life Sci 2020, 15, 677–682. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements. opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas. methods. instructions or products referred to in the content. |
Figure 1.
Images obtained by scanning electron microscope (SEM) with 2500x magnification of control biofilms developed at 24, 48, and 72 h (A, B, and C, respectively) and of test biofilms developed at the same intervals (D, E, and F, respectively). Images G, H and I show test biofilms with 5000x magnification after 24, 48, and 72 h of incubation, respectively (n=6).
Figure 1.
Images obtained by scanning electron microscope (SEM) with 2500x magnification of control biofilms developed at 24, 48, and 72 h (A, B, and C, respectively) and of test biofilms developed at the same intervals (D, E, and F, respectively). Images G, H and I show test biofilms with 5000x magnification after 24, 48, and 72 h of incubation, respectively (n=6).
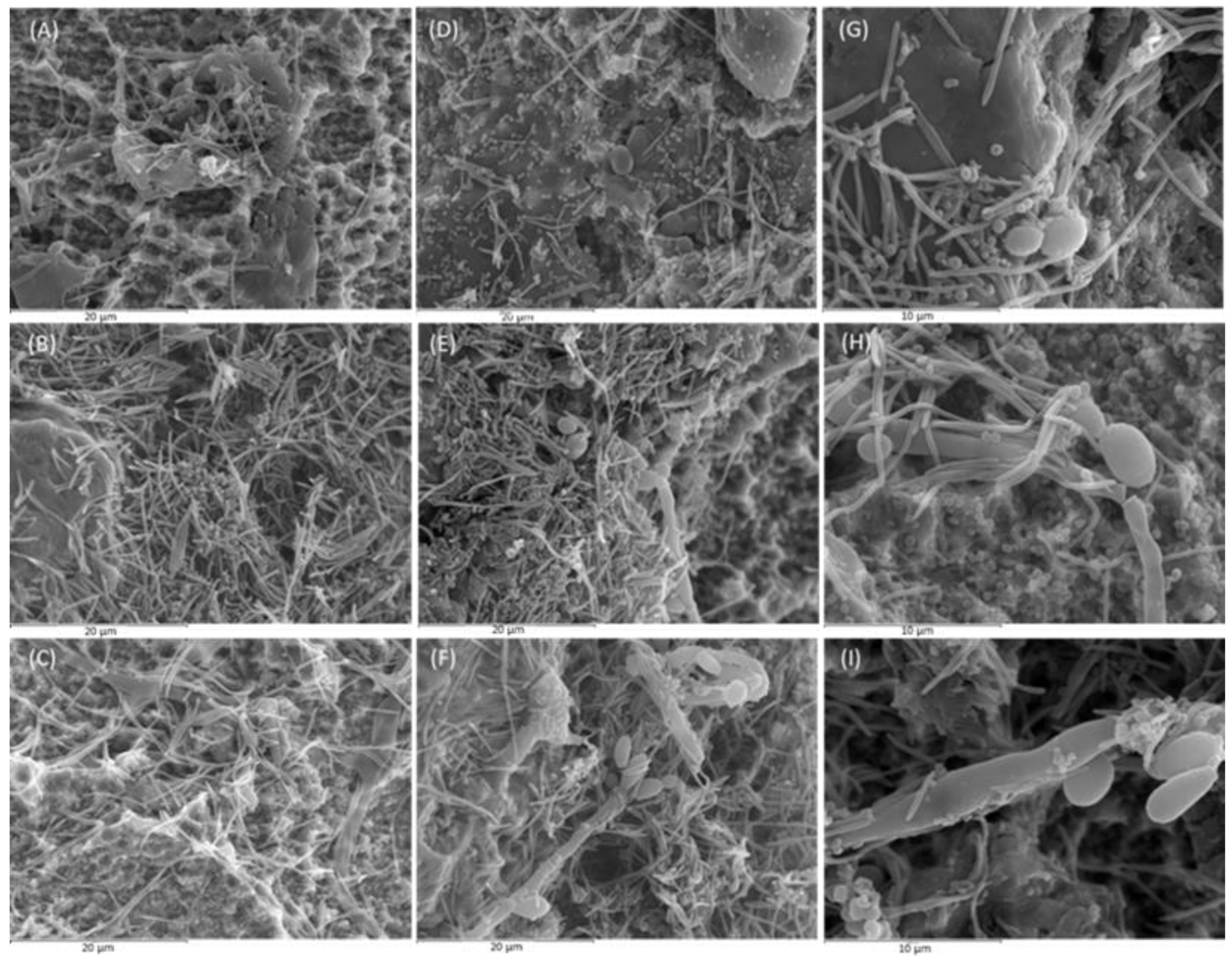
Figure 2.
Images obtained by confocal laser scanning microscopy (CLSM) on control biofilms developed at 24, 48, and 72 h (A, B, and C, respectively) and on test biofilms developed at the same intervals (D, E, and F, respectively). Images G, H, and I show Candida albicans at 24, 48, and 72 h, respectively (scale bar = 200 µm). LIVE/DEAD® BackLight Kit was used to stain live bacteria (green), dead bacteria (red), and implant surfaces (white). Calcofluor White (CFW) was used to stain C. albicans (blue) (n=6).
Figure 2.
Images obtained by confocal laser scanning microscopy (CLSM) on control biofilms developed at 24, 48, and 72 h (A, B, and C, respectively) and on test biofilms developed at the same intervals (D, E, and F, respectively). Images G, H, and I show Candida albicans at 24, 48, and 72 h, respectively (scale bar = 200 µm). LIVE/DEAD® BackLight Kit was used to stain live bacteria (green), dead bacteria (red), and implant surfaces (white). Calcofluor White (CFW) was used to stain C. albicans (blue) (n=6).

Figure 3.
Kinetics of control and test biofilms and Candida albicans [expressed as microbial biomass of biofilm (µm3/µm2)] obtained by quantification of images of confocal laser scanning microscopy (CLSM). Percentages show the proportion of viable cells at each Interval of incubation. * p < 0.05, statistically significant differences when comparing between test and control biofilms, at each time interval.
Figure 3.
Kinetics of control and test biofilms and Candida albicans [expressed as microbial biomass of biofilm (µm3/µm2)] obtained by quantification of images of confocal laser scanning microscopy (CLSM). Percentages show the proportion of viable cells at each Interval of incubation. * p < 0.05, statistically significant differences when comparing between test and control biofilms, at each time interval.
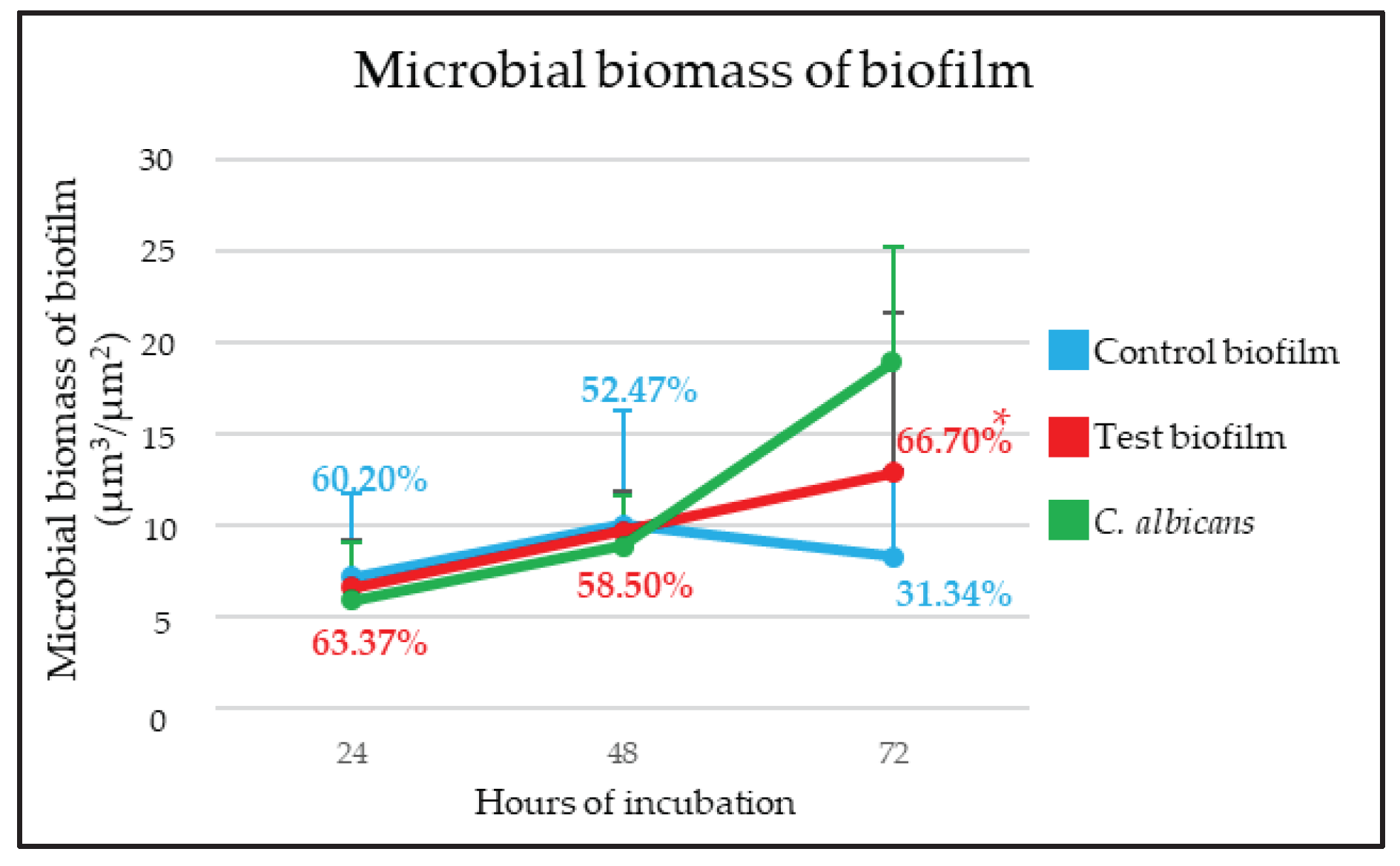

Table 1.
Counts (expressed as mean and standard deviation (SD) and 95% confidence intervals (CI) for means) of microbial species (colony-forming units (CFUs)/mL) determined by quantitative Polymerase chain reaction (qPCR) in 24, 48, and 72 h biofilms on dental implants in the presence (test) and absence (control) of Candida albicans (n=9), using specific primers and probes directed to the 16S rRNA gene. * p < 0.05, statistically significant differences when comparing CFU/mL between test and control biofilms at each time interval. Comparisons between groups were performed considering viable cells and total cells.
Table 1.
Counts (expressed as mean and standard deviation (SD) and 95% confidence intervals (CI) for means) of microbial species (colony-forming units (CFUs)/mL) determined by quantitative Polymerase chain reaction (qPCR) in 24, 48, and 72 h biofilms on dental implants in the presence (test) and absence (control) of Candida albicans (n=9), using specific primers and probes directed to the 16S rRNA gene. * p < 0.05, statistically significant differences when comparing CFU/mL between test and control biofilms at each time interval. Comparisons between groups were performed considering viable cells and total cells.
| Species | Incubation time (hours) | Analysis | Condition | Microbial counts (CFU/ml) | % Viability | ||||
| Mean (SD) | 95% CI | ||||||||
| Lower limit | Upper limit | Mean | (SD) | ||||||
| Streptococcus oralis | 24 | Total | Control | 1.13x107 | 6.71x106 | 4.04x105 | 1.87x107 | ||
| Test | 1.16x107 | 4.69x106 | 6.52x106 | 1.80x107 | |||||
| Viable | Control | 5.85x106 | 3.76x106 | 1.31x106 | 1.10x107 | 51.78 | 14.69 | ||
| Test | 5.76x106 | 3.45x106 | 1.23x106 | 1.08x107 | 46.45 | 14.67 | |||
| 48 | Total | Control | 1.22x107 | 4.87x106 | 5.41x106 | 1.82x107 | |||
| Test | 1.65x107 | 6.86x106 | 7.20x106 | 2.59x107 | |||||
| Viable | Control | 6.51x106 | 2.71x106 | 3.51x106 | 1.08x107 | 56.54 | 19.16 | ||
| Test | 8.69x106 | 2.63x106 | 5.02x106 | 1.15x107 | 58.79 | 23.24 | |||
| 72 | Total | Control | 9.43x106 | 3.43x106 | 4.17x106 | 1.35x107 | |||
| Test | 1.24x107 | 7.25x106 | 7.06x106 | 2.98x107 | |||||
| Viable | Control | 3.95x106 | 1.54x106 | 2.21x106 | 6.26x106 | 45.24 | 18.15 | ||
| Test | 8.30x106* | 3.80x106 | 3.08x106 | 1.48x107 | 70.92* | 25.05 | |||
| Species | Incubation time (hours) | Analysis | Condition | Microbial counts (CFU/ml) | % Viability | ||||
| Mean (SD) | 95% Confidence interval for mean | ||||||||
| Lower limit | Upper limit | Mean | (SD) | ||||||
|
Actinomyces naeslundii |
24 | Total | Control | 7.58x105 | 4.95x105 | 1.14x105 | 1.31x106 | ||
| Test | 1.01x106 | 8.02x105 | 9.75x104 | 2.18x106 | |||||
| Viable | Control | 4.20x105 | 3.48x105 | 2.58x104 | 1.03x106 | 66.91 | 30.38 | ||
| Test | 5.65x105 | 3.90x105 | 1.01x105 | 1.03x106 | 68.79 | 20.82 | |||
| 48 | Total | Control | 5.51x105 | 3.48x105 | 1.10x105 | 1.09x106 | |||
| Test | 9.00x105 | 7.45x105 | 9.02x104 | 2.38x106 | |||||
| Viable | Control | 3.33x105 | 2.13x105 | 1.10x105 | 6.61x105 | 74.22 | 37.58 | ||
| Test | 5.72x105 | 3.96x105 | 9.98x104 | 1.31x106 | 83.24 | 38.63 | |||
| 72 | Total | Control | 4.43x105 | 2.93x105 | 7.39x104 | 8.09x105 | |||
| Test | 1.21x106 | 9.92x105 | 4.18x104 | 2.65x106 | |||||
| Viable | Control | 1.98x105 | 1.21x105 | 7.39x104 | 4.15x105 | 57.1 | 26.76 | ||
| Test | 4.11x105 | 3.05x105 | 5.14x104 | 9.60x105 | 58.47 | 39.11 | |||
|
Veillonella parvula |
24 | Total | Control | 5.18x107 | 4.45x107 | 6.70x106 | 1.49x108 | ||
| Test | 3.48x107 | 2.11x107 | 6.62x106 | 5.61x107 | |||||
| Viable | Control | 3.31x107 | 2.14x107 | 6.99x106 | 6.49x107 | 79.09 | 28.19 | ||
| Test | 2.31x107 | 1.34x107 | 4.97x106 | 4.38x107 | 76.76 | 30.06 | |||
| 48 | Total | Control | 4.58x107 | 3.08x107 | 6.79x106 | 9.32x107 | |||
| Test | 5.77x107 | 4.51x107 | 8.90x106 | 1.31x108 | |||||
| Viable | Control | 2.45x107 | 1.35x107 | 6.68x106 | 4.25x107 | 67.78 | 26.03 | ||
| Test | 3.57x107 | 2.79x107 | 5.63x106 | 8.43x107 | 67.2 | 17.34 | |||
| 72 | Total | Control | 3.04x107 | 2.26x107 | 5.15x106 | 6.81x107 | |||
| Test | 2.31x107 | 1.30x107 | 5.45x106 | 4.44x107 | |||||
| Viable | Control | 1.23x107 | 1.09x107 | 3.49x106 | 3.73x107 | 50.44 | 25.03 | ||
| Test | 1.05x107 | 5.73x106 | 3.41x106 | 1.78x107 | 51.21 | 21.46 | |||
|
Fusobacterium nucleatum |
24 | Total | Control | 1.95x106 | 1.04x106 | 3.64x105 | 3.27x106 | ||
| Test | 2.05x106 | 8.77x105 | 1.32x106 | 4.17x106 | |||||
| Viable | Control | 5.96x105 | 4.57x105 | 4.30x104 | 2.45x105 | 29.12 | 13.21 | ||
| Test | 9.39x105 | 6.15x105 | 1.16x106 | 1.77x106 | 46.1 | 18.89 | |||
| 48 | Total | Control | 2.85x106 | 2.08x106 | 1.70x105 | 5.09x106 | |||
| Test | 1.10x107* | 5.45x106 | 4.19x106 | 2.02x107 | |||||
| Viable | Control | 1.52x106 | 1.24x106 | 7.17x104 | 3.64x106 | 47.42 | 14.9 | ||
| Test | 5.15x106* | 3.04x106 | 1.20x106 | 8.35x106 | 48.25 | 12.38 | |||
| 72 | Total | Control | 4.15x106 | 3.45x106 | 6.15x104 | 9.60x106 | |||
| Test | 1.35x107* | 5.86x106 | 3.64x106 | 2.12x107 | |||||
| Viable | Control | 1.31x106 | 1.07x106 | 8.92x103 | 2.63x106 | 27.42 | 9.711 | ||
| Test | 7.53x106* | 5.74x106 | 4.54x105 | 1.86x107 | 54.36* | 26.21 | |||
| Species | Incubation time (hours) | Analysis | Condition | Microbial counts (CFU/ml) | % Viability | ||||
| Mean (SD) | 95% Confidence interval for mean | ||||||||
| Lower limit | Upper limit | Mean | (SD) | ||||||
|
Porphyromonas gingivalis |
24 | Total | Control | 3.31x104 | 1.50x104 | 5.20x103 | 4.79x104 | ||
| Test | 3.83x104 | 2.18x104 | 1.13x104 | 6.20x104 | |||||
| Viable | Control | 1.43x104 | 8.24x103 | 6.25x103 | 2.56x104 | 40.27 | 14.91 | ||
| Test | 2.21x104 | 1.27x104 | 6.41x103 | 3.87x104 | 57.3 | 19.82 | |||
| 48 | Total | Control | 2.02x105 | 1.03x105 | 7.29x104 | 3.44x105 | |||
| Test | 4.14x105* | 1.86x105 | 1.57x105 | 6.11x105 | |||||
| Viable | Control | 6.51x104 | 3.21x104 | 2.84x104 | 1.15x105 | 37.67 | 17.43 | ||
| Test | 2.25x105* | 1.13x105 | 7.34x104 | 4.07x105 | 54.38* | 9.468 | |||
| 72 | Total | Control | 3.65x105 | 1.97x105 | 1.10x105 | 5.81x105 | |||
| Test | 1.13x106* | 5.51x105 | 6.98x105 | 2.44x106 | |||||
| Viable | Control | 1.24x105 | 5.57x104 | 4.93x104 | 2.05x105 | 37.91 | 9.136 | ||
| Test | 6.78x105* | 1.84x105 | 4.75x105 | 9.28x105 | 65.29* | 18.16 | |||
| Aggregatibacter actinomycetemcomitans | 24 | Total | Control | 2.82x106 | 1.03x106 | 1.35x106 | 3.75x106 | ||
| Test | 2.72x106 | 8.14x105 | 1.68x106 | 4.07x106 | |||||
| Viable | Control | 1.10x106 | 4.24x105 | 1.27x105 | 1.52x106 | 44.34 | 23.92 | ||
| Test | 1.49x106 | 3.53x105 | 9.54x105 | 1.99x106 | 56.01 | 7.191 | |||
| 48 | Total | Control | 4.04x106 | 1.44x106 | 2.15x106 | 6.34x106 | |||
| Test | 3.79x106 | 1.24x106 | 1.45x106 | 5.41x106 | |||||
| Viable | Control | 1.38x106 | 2.43x105 | 9.64x105 | 1.76x106 | 38.36 | 15.88 | ||
| Test | 2.11x106* | 7.45x105 | 5.47x105 | 3.06x106 | 55.28* | 10.77 | |||
| 72 | Total | Control | 5.01x106 | 1.37x106 | 3.47x106 | 7.29x106 | |||
| Test | 4.90x106 | 1.65x106 | 2.52x106 | 6.60x106 | |||||
| Viable | Control | 1.56x106 | 3.28x105 | 1.21x106 | 2.16x106 | 32.17 | 6.379 | ||
| Test | 3.15x106* | 3.63x105 | 2.53x106 | 3.70x106 | 71.99* | 29.11 | |||
| Candida albicans | 24 | Total | Control | ||||||
| Test | 2.37x104 | 7.86x103 | 1.74x104 | 4.05x104 | |||||
| Viable | Control | ||||||||
| Test | 3.21x103 | 7.51x102 | 2.30x103 | 4.72x103 | 14.02 | 2.734 | |||
| 48 | Total | Control | |||||||
| Test | 7.65x104 | 2.72x104 | 3.87x104 | 1.13x105 | |||||
| Viable | Control | ||||||||
| Test | 3.02x104 | 1.04x104 | 1.62x104 | 4.36x104 | 45.37 | 23.73 | |||
| 72 | Total | Control | |||||||
| Test | 2.35x105 | 7.75x104 | 1.26x105 | 3.17x105 | |||||
| Viable | Control | ||||||||
| Test | 9.12x104 | 1.90x104 | 6.54x104 | 1.14x105 | 43.84 | 20.06 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



