
Submitted:
20 February 2024
Posted:
20 February 2024
You are already at the latest version
Abstract
Seeds from several Ribes taxa were surveyed for phenolic compounds and in vitro antiproliferative activity against HT-29 colorectal cancer cells. Total phenolic compounds were analysed through the Folin-Ciocalteu procedure, while phenolic profiles were analysed by LC coupled to a single mass spectrometer Orbitrap using an electrospray interface (ESI). Antitumor effects were established using the MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide) assay. Total phenolics ranged from 11.4 in R. alpinum to 94.8 mg of caffeic acid equivalents (CAE)/g in R. nigrum 'Koksa'. Concerning phenolic compounds, four were hydroxylated benzoic acids, four cinnamic acid derivatives, eight flavonoids, and nine flavonoid glycosides. The growth inhibition against HT-29 cancer cells was exercised much better by R. nigrum 'Koksa' and Ribes 'Erkeeni' (GI50 37 and 42 µg/ml). All Ribes extracts, except R. nigrum 'Hara katarlik', showed higher activity than that of R. rubrum (GI50 at 72 h: 99 µg/ml). Interestingly, the highly bioactive extract from Ribes 'Erkeeni' contains all detected phenolics, while R. nigrum 'Koksa' lacks only populnin. Therefore, the high bioactivity found for such extracts could be due to a synergy due to all detected compounds. This work constitutes a comprehensive action for expanding knowledge on the phenolic profiles and antitumor activity of GLA-rich Ribes seeds.
Keywords:
Ribes
; blackcurrant cultivars
; phenolic compounds
; LC-MS
; HT-29
; MTT assay
1. Introduction
The genus Ribes (fam. Grossulariaceae) includes more than 150 diploid species, distributed in the temperate regions of the Northern Hemisphere and South America (Brennan, 1996). At present, 10-12 species of Ribes are propagated for fruit production, the majority of which are black (Ribes nigrum L.), red, and white currant (R. rubrum L., synonyms R. vulgare Jancz. and R. sativum Syme) and gooseberry (e.g., European gooseberry: R. uva-crispa L., synonym R. grossularia L., and American hairystem gooseberry: Ribes hirtellum Michx.) (Brennan, 2009). Black currants are considered as promising crops with high economic value, especially since they are ranked second in consumer preferences after strawberries (Brennan, 2009; Jiménez-Aspee et al., 2015).
Fresh edible Ribes fruits are promising crops with high economic value, not only due to their desired taste and nutritional value but also for their well-known health-promoting properties (Mikulic-Petkovsek et al., 2015). Furthermore, Ribes seed oil has immunomodulation and anti-inflammatory effects, i.e., the use of blackcurrant seed oil in preventing illness like hypertension, psoriasis, and atopic dermatitis has been reported (Jurgoński et al., 2015; Michalak & Kiełtyka-Dadasiewicz, 2018).
Moreover, the fruit industry generates year by year increased amounts of fruits by-products, in which occurs tonnage amounts of seeds, while the seeds of some species/cultivars constitute a valuable source of γ-linolenic acid (GLA, 18:3n-6), which can account for more than 20% of total fatty acids (FA) (e.g., Lyashenko et al., 2019; Golovenko et al., 2021). Furthermore, in addition to its high content in GLA, blackcurrant seed oil has also been reported to be a good source of phenolic compounds (Bakowska-Barczak et al., 2009), which constitute a large fraction of the unsaponifiable material of most vegetable oils (Fabrikov et al., 2019).
The chemical profiling of European currants has been extensively described, mainly for R. nigrum (black currants) and R. rubrum (red currants), while data on the composition of phenolic compounds and biological activity of the remaining species of Ribes genus and cultivars is extremely limited. Taking into account the benefits of the unsaponifiable material contained in the seed oil and the lack of information about several Ribes taxa, the aim of present study was to determine the phenolic profiles and the in vitro antitumor activity against colorectal cancer cells of the phenolic-containing seeds extracts from selected Ribes taxa. All studied species/cultivars have been previously typified as potential GLA producers (Lyashenko et al., 2019).
2. Results and Discussion
2.1. Total Phenolics and Oil Content
Table 1 shows the amounts of oil content, the total phenolic content (TPC) in mg of caffeic acid equivalents (CAE)/g seeds and mg of CAE/g oil, as well as data on GLA content in seeds (% of total FA) previously reported by our Research Team (Lyashenko et al., 2019). Notably, seed samples vary in their total oil content, with values ranging from 12.7 (R. alpinum 1B) to 25.6 g/100 g of seeds (R. hudsonianum). A significant variability was observed in TPC amounts among different samples and sections. In sect. Berisia, R. alpinum 1A have a lower oil content compared to 1B, but show higher TPC, while R. pulchellum stands out with the highest content of oil and TPC. As for sect. Coreosma, R. hudsonianum has the highest oil content and TPC in oil, and R. nigrum ‘Koksa’ has the highest TPC in seeds. Concerning sect. Ribes, R. rubrum has the lowest oil and TPC content in seeds. Regarding GLA values, Ribes ‘Myuryucheene’ shows the highest percentages of total FA, but unfortunately neither its oil content nor TPC are notable. Conversely, R. nigrum ‘Koksa’ has 17.0% GLA of total FA, and given its high TPC (15.5 mg CAE/g oil), this cultivar constitutes a promising source of GLA-rich oil containing good amounts of phenolics, thus, hosting healthy properties related to such compounds.
In short, a proper selection of Ribes varieties can significantly influence the content of bioactive compounds in seeds, i.e., GLA and TPC, whose values in the species and cultivars focused here are among the highest reported for Ribes taxa. Previous analysis effected to the seeds of other R. nigrum cultivars revealed that ‘Ben Tirran’ and ‘Ben Sarek’ ones are good sources of GLA (15.2-16.7% of total FA), although these have very low TPC amounts quantified by the Folin Ciocalteu (F-C) method: 1.99 and 2.31 mg of Gallic Acid Equivalents (GAE)/g seed residue, respectively (Bakowska-Barczak et al., 2009. However, Van Hoed et al. (2011) indicated figures obtained by the F-C methodology in the range showed here for several Ribes cultivars (5.6-11.3 mg CAE/g oil). Other works focused on residues from the extraction of Ribes fruits cannot be compared with the results obtained here, since such works were focused on the residual cake from the extraction of the fruit, which in addition to the seeds contains several other tissues of the fruit (e.g., Kapasakalidis et al., 2006; Granato et al., 2022).
2.2. Phenolic Compounds Profiles
The phenolic compounds profiles obtained by the LC-MS system of the seeds of Ribes species/cultivars focused here are reported in Table 2. The identification was achieved by means of retention time (Rt) and m/z of both, the adducts and fragment ions. All compounds were properly identified, and the bases for the identification of each compound are described in Table 2.
Among detected phenolics, four consisted of hydroxylated derivatives of benzoic acids (3,4-dihydroxybenzoic, salicylic, vanillic, and 4-hydroxybenzoic acid) (Figure 1A); four were cinnamic acid derivatives (caffeic, p-coumaric, chlorogenic, and ferulic acids) (Figure 1B); two flavone derivatives (luteolin and apigetrin) (Figure 1C); three flavanone derivatives (juncein, eriodictyol, and naringenin) (Figure 1D). The largest group was the flavonol derivatives, with eleven compounds (populnin, quercetin, isoquercitrin, rutin, kaempferol, quercitrin, myricetin, fisetin, astragalin, nicotiflorin, and galangin) (Figure 1E). Finally, a dihydrochalcone glucoside was also detected (phloridzin) (Figure 1F). Anthocyanins, usually found in blackcurrant and redcurrant pomaces, such as delphinidin-3-glucoside and cyanidin-3-glucoside (Mikulic-Petkovsek e al., 2015), were not found, which was due to the fact that the analyzed material only included seeds and no other fruit tissues where such compounds occur. However, some authors reported low quantities of anthocyanins (3−6 mg/100 g), such as delphinidin and cyaniding glycosides, in black currant seeds (e.g., Bakowska-Barczak et al., 2009), which can be due to an incomplete removal of pulp tissues from seeds.
As for the distribution of compounds among the various taxa, the highest diversity of compounds was detected in Ribes ‘Algo’ Yakutskaya and R. nigrum ‘Koksa’, while the compounds that were identified in all Ribes samples were ferulic and 4-hydroxybenzoic acids. Some compounds were restricted to few taxa; for instance, eriodictyol and phloridzin occurred only in R. nigrum ‘Koksa’, Ribes ‘Algo’ Yakutskaya, and Ribes ‘Myuryucheene’. It has not been possible to establish a correlation between the presence of phenolic compounds and taxonomic category. This is interpreted as meaning that, within Ribes genus, environmental factors (temperature, soil, light, fertilizer, etc.) are more decisive in terms of the occurrence of phenolic compounds than any genetic proximity. This has been investigated in berries, and it has been reported that TPC was higher in fruits cultured in the north than in the south, and also that high radiation and temperature were associated with low contents of the major phenolic compounds in all the cultivars studied (Yang et al., 2013). However, the influence of the environment on the phenolic content of seeds remains unstudied.
Some authors reported flavonols as the main phenolic group in black currant seeds. Among flavonols, quercetin-3-glucoside, myricetin-3-glucoside, and kaempferol-3-glucoside were the major compounds detected (Bakowska-Barczak et al., 2009). In this study, kaempferol-7-glucoside was detected instead the 3-glusoside, in addition to the aglycone of these compounds, but not the glycosides of myricetin. Any case, most compounds found here were previously reported from Ribes species. For instance, Wójciak et al. (2022), reported for black currant seeds glucosides and rutinosides derivatives of quercetin and kaempferol, besides the aglycones of most compounds reported here. However, to the best of our knowledge, this work constitutes the first report for some compounds detectedt in Ribes taxa: e.g., fisetin, luteolin, eriodictyol, phloretin, galangin, and naringenin, as well as some of their glycosides.
The great variety of flavonoids found in the focused Ribes seeds has deep significance for health. Such compounds exhibit high activity against several diseases including cancer, without showing significant toxicity towards normal cells. Flavonoids can enhance drug sensitivity and suppress proliferation, metastasis, and angiogenesis of cancer cells by modulating several oncogenic or oncosuppressor microRNAs (miRNAs, miRs) (Tuli et al., 2023). For instance, quercetin is active against lung, breast, and prostate cancer cells, and luteolin is active against glioblastoma and colon cancers (Tuli et al., 2023). The same is true for flavonoid glycosides; e.g., populnin (kaempferol-7-O-glucoside) has potent anti-Herpes simplex virus activity, and significant anti-HIV-1 reverse transcriptase activity, which leads to consider it as an anti-HIV potential drug for the early treatment of HIV infection (Behbahani et al. 2014).
Previous reports indicated that berries cultivated in cold climates accumulate significantly higher levels of phenolic compounds than those grown in milder climates. In this way, flavonoids are accumulated in response to abiotic stresses such as low temperature, which increased the abundance of enzymes involved in flavonoid biosynthesis and the expression of flavonoid biosynthesis genes in various plant species (Schulz al., 2016). Consistently, the great diversity of phenolic compounds found in this work could be interpreted considering that plants were cultivated in very difficult climatic conditions of Siberia (Yakutia and Krasnoyarsk krai). These are the coldest regions of Russia, characterized by a protracted cold season and exceptionally low winter temperatures (-40 °C). Over time, the evolutionary processes in these challenging conditions have shaped a unique gene pool in plants, endowing them with complex resistance, such as frost resistance, high levels of biologically active compounds and key nutritional components. In fact, Ribes cultivars from Yakutia are characterized by higher levels of polyunsaturated FA (PUFA) than other Ribes species, particularly GLA (Lyashenko et al., 2019).
2.3. Antiproliferative Activity of the Water:Methanol Seed Extracts on HT-29 Cancer Cells
To determine the in vitro anticancer activity, we selected blackcurrant cultivars from Yakutia, since their seed oils showed high content of GLA (Lyashenko et al., 2019). A sample of red currant (R. rubrum) was also checked with comparative purposes since it is a widely cultured and commercialized berry. Previously, the antitumor activity of phenolic extracts from fruits and leaves of Ribes species has been studied. For instance, the apoptotic effects and mechanisms of blackcurrant extracts in MKN-45 (human gastric adenocarcinoma) and TE-1 (human esophageal cancer) cells were assessed. It was demonstrated that such extracts induced caspase-dependent apoptosis through downregulation of Bcl-2, a mitochondrial pathway involving activation of p38 (mitogen-activated protein kinases) and JNK (c-Jun N-terminal kinase), and inactivation of Akt (a central kinase that controls diverse processes including cell survival and apoptosis). Thus, such extract has been proposed as a potential candidate for cancer therapy (Liu and Li, 2016). However, the antitumor activity of phenolic-rich seed extracts of any Ribes species remains unchecked.
Figure 2 shows the results of the MTT assay. The concentration-response plots for HT-29 cells after exposure to seed extracts after 48 and 72 h of treatment are drawn in Figures 2A and 2B, respectively, while GI50 for the previous assays and for ferulic acid and doxorubicin (positive control) is depicted in Figure 3. The Selectivity Index (SI) for 72-h exposed cells to seeds extracts is shown in parentheses over columns. After 48 and 72 h of treatment, the MTT assay revealed concentration- and time-dependent inhibitory effects on HT-29 cells for all assayed extracts. Cell viability at 48 h at the maximum concentration tested (300 μg/mL) and for the different species, was ~20% lower than that obtained at 72 h. After 72 h culture, cells growth inhibition was exercised much better by R. nigrum ‘Koksa’ and Ribes ‘Erkeeni’, which shows a GI50 value of 37 and 42 µg/ml. All assayed Ribes extracts, except R. nigrum ‘Hara katarlik’, showed activity higher than R. rubrum extract (GI50 at 72 h of cell exposure to extract of 99 µg/ml). The extracts of R. nigrum ‘Hara katarlik’ showed an undetermined GI50 value higher than 300 μg/mL. Interestingly, the highly bioactive extract from Ribes ‘Erkeeni’ contains all detected phenolics, while R. nigrum L. ‘Koksa’ lacks only populnin. Therefore, the high bioactivity found for such extracts could be due to a synergy between all the compounds detected. However, need to be considered that the polysaccharide-rich fraction from R. nigrum has been characterized as highly bioactive (Yang et al., 2020), and that the water:methanol extracts obtained from the seeds assayed in this work can include some amounts of polysaccharides, thus, a synergy between phenolic compounds and seed polysaccharides could be also responsible for the noted antitumor effects against HT-29 cells, especially considering that these cells are not too sensitive to phenolic compounds (Lyashenko et al., 2021; Chelh et al., 2022). Finally, the selectivity index (SI) of HT- 29 versus normal cells CCD-18 was calculated (see Material and Methods section, 3.7). An extract with SI value greater than 2 is considered as high selectivity against cancer cells, whereas one with SI value less than 2 demonstrates general toxicity to normal cells (Vichitsakul et al., 2023). Such value for any research on herbal drug and/or isolated compound is critical for determining whether further research can be continued (Peña-Morán et al., 2016). SI at 72 h ranged from 17 (Ribes ‘Myuryucheene’) to 32 (Ribes ‘Erkeeni’), from which it can be concluded that the seed extracts of Ribes have a high selectivity against HT-29 human colorectal cancer cells. This means that the extracts evaluated here constitute promising candidates for obtaining active compounds against colorectal cancer.
Conclusions
As shown, the seeds of some unexplored taxa belonging to Ribes genus constitute potentially raw sources of healthy phenolic compounds-rich seed oils, in addition to their already known GLA-rich FA profiles. Among the different taxa analyzed here highlights R. nigrum cultivars, due to the diversity of compounds they show, especially Ribes ‘Algo’ Yakutskaya and R. nigrum ‘Koksa’. The compounds that were identified in all Ribes samples were ferulic and 4-hydroxybenzoic acids, while some compounds were restricted to few taxa. It was not possible to establish a correlation between the presence of phenolic compounds and taxonomic rank and, probably, environmental factors were more decisive for compound occurrence than any genetic proximity. It highlights the antiproliferative activity of some seed extracts against HT-29 cells. Cell growth inhibition was strongly effected by R. nigrum ‘Koksa’ and Ribes ‘Erkeeni’, which shows very low GI50 values and contains most identified phenolics. Future research on this subject should be carried out to elucidate the composition of other components of the unsaponifiable of these oils, such as sterols and tocols, while it is recommended to obtain Ribes oils exclusively by cold pressing, so that they can be enriched in the healthy components contained in the unsaponifiable fraction. Other actions should be carried out to deepen the knowledge of the quantification of the detected phenolics, and their individual action against various cancer cell lines.
3. Materials and Methods
3.1. Reagents and Chemicals
Unless otherwise indicated, all chemicals and solvents were purchased from Merck (Madrid, Spain). Aluminum chloride and sodium carbonate were obtained from Sigma-Aldrich Co. (St Louis, MO, USA). Water was purified using a Milli-Q system (Millipore, Burlington, MA, USA). All the chemicals used, including the solvents, were of analytical grade.
3.2. Plant Material
Data on analyzed Ribes seeds are detailed in Table 3. Seeds were donated by several botanical gardens. R. alpinum (1B) seeds were collected three well-differentiated subpopulations from their natural habitats in Sierra de Baza (Granada, Spain). Upon receipt, after cleaning, adequate amounts of each sample (2 g) were used for moisture analysis. This was carried out in a forced air oven at 103 ºC for 8 h, and all results in tables and figures are expressed on dry weight (dw) basis. Moisture content ranged from 7.1 (R. rubrum) to 8.3 g/100 g (R. Erkeeni). The remaining seeds were labeled and placed in plastic containers at -18 ºC until lab analysis. Immediately before starting each experiment, seeds were dried and grounded into fine powder using mortar and pestle.
3.3. Seed oil Extraction
Wild-collected seeds were separated from the pulp, exposed to air to dry at room temperature, and ground into powder. The powders from all seeds were analyzed without delay after crushing and the oil content was determined by the Weibull and Stoldt method (AOAC, 2000).
3.4. Extraction of Phenolics from Ribes Seeds
Extraction and analysis of phenolic compounds from Ribes seeds were carried out according to Lyashenko et al. (2021) with some modifications. Powdered seeds (~0.2 g) were extracted three times with 3 mL of methanol:water, (60:40, v/v). After centrifuging at 1,000×g for 10 min, the supernatants were collected, combined and the solvent was evaporated under vacuum at 60 ºC to dryness. The residue was dissolved in 1 mL of methanol:water (60:40, v/v), and filtered through a 0.22 μm membrane filter prior to the chromatographic analysis.
3.5. Determination of Total Phenol Content
Total phenolics content (TPC) was measured using the F-C assay as developed by Singleton et al. (1999) with minor modifications. Briefly, 10 μL of phenolic seed extracts, prepared as above described, 0.79 mL of MiliQ water and 50 μL of the F-C reagent were mixed, vortex and allowed to stand for 5 min at room temperature. Next, 150 μL of a 20% sodium carbonate solution were added and vortex. A control sample was also prepared. After incubation at room temperature for 2 h in darkness, the absorbance of the mixture was read at 765 nm on a UV-VIS spectrophotometer using either water as blanks. The results were expressed as mg of Caffeic Acid Equivalents (CAE) per 100 g of sample using a standard curve of caffeic acid (ranging from 50 to 900 μg/mL). Determinations were done in triplicate.
3.6. Characterization of Phenolics by Liquid Chromatography-Mass Spectrometry
The chromatographic separation was performed on a Thermo Fisher Scientific Transcend 600 LC (Thermo Scientific TranscendTM, Thermo Fisher Scientific, San Jose, CA, USA) by using a Hypersil Gold column (250 x 4.6mm, 5 µm). A flow rate of 0.65 mL/min was set. The compounds were separated with gradient elution using aqueous acetic acid (acetic acid: H2O, 1:99, v/v) (A) and methanol (B) as eluents at ambient temperature. The step gradient was as follows: 0–20 min 80% of A; then, it was linearly decreased to 25% in 10 min and remained constant during 10 min. Later, it was increased to 80% in 10 min and remained constant during 5 min. The total running time was 55 min. The column temperature was 25 °C, and the injection was 10 μL.
The LC system is coupled to a single mass spectrometer Orbitrap Thermo Fisher Scientific (ExactiveTM, Thermo Fisher Scientific, Bremen, Germany) using an electrospray interface (ESI) (HESI-II, Thermo Fisher Scientific, San Jose, CA, USA) in positive and negative ion mode. ESI parameters were as follows: spray voltage, 4 kV; sheath gas (N2>95%), 35 (adimensional); auxiliary gas (N2>95%), 10 (adimensional); skimmer voltage, 18 V; capillary voltage, 35 V; tube lens voltage, 95 V; heater temperature, 305 °C; capillary temperature, 300 °C. The mass spectra were acquired employing two alternating acquisition functions: (1) full MS, ESI+, without fragmentation (higher collisional dissociation (HCD) collision cell was switched off), mass resolving power = 25,000 FWHM; scan time = 0.25 s; (2) all-ions fragmentation (AIF), ESI+, with fragmentation (HCD on, collision energy 30 eV), mass resolving power = 10,000 FWHM; scan time = 0.10 s, (3) full MS, ESI- using the aforementioned settings and (4) AIF, ESI- using the settings explained for (2). Mass range in the full scan experiments was set at m/z 50–1000. LC chromatograms were acquired using the external calibration mode and they were processed using XcaliburTM version 3.0, with Qualbrowser and Trace Finder 4.0 (Thermo Fisher Scientific, Les Ulis, France). Unknown analysis was carried out with Compound DiscovererTM version 2.1.
3.7. Cell assays on Cancer and Normal Cell Lines
The anticancer activity was determined for seed extracts from Ribes cultivars, and R. rubrum extract, a widely used commercial Ribes species, was used for comparison. The HT-29 colon cancer cells line and the CCD-18 colonic human myofibroblasts cells line was used to check antiproliferative activities. Cultures were supplied by the Technical Instrumentation Service of University of Granada (Granada, Spain). First, it was checked for the absence of Mycoplasma and bacteria. Then, cells were grown at 37 °C and 5% CO2 humidified atmosphere in medium RPMI-1640 supplemented with 5% fetal bovine serum, 2 mM L-Glutamine, 1 mM sodium pyruvate, 0.125 mg/mL amphotericin and 100 mg/mL penicillin-streptomycin.
All cultures were plated in 25 cm2 plastic tissue culture flasks (Sarstedt, USA). All culture media and reagents were purchased from Sigma-Aldrich (St. Louis, MO, USA). Cell culture and cell assay, that is, the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) test were accomplished as previous described (Ramos-Bueno et al., 2016).
In the MTT assay, cells were divided into 96-well microtiter plates, adjusted of 1 × 104 cell/well and cultivated in medium at 37°C, 5% CO2 prior to adding the different extracts dissolved in medium. The phenolics-containing extracts were supplied to cells dissolved in a mixture of methanol:water, (60:40, v/v), and then in the culture medium at designed concentrations (0–300 μg/mL). After 48 and 72 h of cell exposure, 5 mg/mL of an MTT solution was added to the culture medium to determine the viability of cells. The absorbance was recorded at 570 nm on an enzyme-linked immunosorbent assay (ELISA) plate reader (Thermo Electron Corporation, Sant Cugat del Valles, Barcelona, Spain). The formazan crystals produced were solubilized using 100 μL dimethylsulfoxide (DMSO). Cells without phenolic extracts were considered as negative controls.
Cell survival in exposed cultures relative to unexposed cultures (negative controls) was calculated, and the number of viable cells was calculated using the following equation: Percentage of viable cell (%) = (Absorbance of treated cells/Absorbance of untreated cells) × 100%.
The concentrations causing 50% cell growth inhibition (GI50) were calculated from the growth curves. Doxorubicin (98.0–102%, D1515), from Sigma-Aldrich (Madrid, Spain) was used as a positive control, while DMSO and methanol were used as the negative (vehicle) controls. Phenolic extracts and controls were evaluated in three independent assays. Values presented are mean ± standard error of the mean. The selectivity index (SI) of each compound was calculated as GI50 of the extract against the CCD-18 normal cell line/GI50 of the same extract against the HT-29 cancer cell line (Vichitsakul et al., 2023).
3.8. Statistical Analysis
Data on seeds from botanical gardens correspond to the analyses effected to seeds received in a single shipment, which were analyzed three times in triplicate each. Seeds from the wild were collected from three different species populations and each of which was analyzed in triplicate. All data in tables were analyzed using one-way ANOVA (Statgraphics Centurion XVI.I, Warrenton, VA, USA) and expressed as the average ± SD. Differences among mean values were tested by Duncan’s test at P<0.05.
Funding
The authors acknowledge the financial support of Junta de Andalucía (Project P20_00806), University of Almería (P_LANZ_2023/003), Campus de Excelencia Internacional Agroalimentario (ceiA3), and Centro de Investigación en Agrosistemas Intensivos Mediterráneos y biotecnología Agroalimentaria (CIAMBITAL).
Data Availability Statement
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author/s.
Acknowledgments
The authors acknowledged the botanical gardens listed in Table 1 for providing plant material.
Conflicts of Interest
The sponsors had no role in the design, execution, interpretation, or writing of the study.
References
- AOAC (2000). Fat (crude) in nuts and nut products. Gravimetric method (AOAC Official Method 948.22) W. Horwitz (Ed.), Officials methods of analysis of AOAC International (17th ed.), AOAC, Gaithersburg, MD.
- Bakowska-Barczak, A. M., Schieber, A., & Kolodziejczyk, P. (2009). Characterization of Canadian black currant (Ribes nigrum L.) seed oils and residues. Journal of agricultural and food chemistry, 57, 11528-11536. [CrossRef]
- Behbahani, M., Sayedipour, S., Pourazar, A., & Shanehsazzadeh, M. (2014). In vitro anti-HIV-1 activities of kaempferol and kaempferol-7-O-glucoside isolated from Securigera securidaca. Research in pharmaceutical sciences, 9, 463.
- Brennan, R. M. (1996). Currants and Gooseberries. Chapter 3 pp. 191-295 in: J. Janick and J. N. Moore (eds.) Fruit Breeding, Vol. II Vine and Small Fruit Crops. John Wiley & Sons. Inc. N.Y.
- Brennan, J., & Graham, J. (2009). Improving Fruit Quality in Rubus and Ribes Through Breeding. Functional Plant Science and Biotechnology, 3 (1), 22-29.
- Chelh, T. C., Lyashenko, S., Lahlou, A., Belarbi, E. H., Rincón-Cervera, M. Á., Rodríguez-García, I., ... & Guil-Guerrero, J. L. (2022). Buglossoides spp. seeds, a land source of health-promoting n-3 PUFA and phenolic compounds. Food Research International, 157, 111421. [CrossRef]
- https://doi.org/10.1016/j.foodres.2022.111421. [CrossRef]
- Chernonosov, A. A., Karpova, E. A., & Lyakh, E. M. (2017). Identification of phenolic compounds in Myricaria bracteata leaves by high-performance liquid chromatography with a diode array detector and liquid chromatography with tandem mass spectrometry. Revista Brasileira de Farmacognosia, 27, 576-579. [CrossRef]
- Dantas, C. A. G., Abreu, L. S., da Cunha, H. N., Veloso, C. A. G., Souto, A. L., de Fatima Agra, M., ... & Tavares, J. F. (2021). Dereplication of phenolic derivatives of three Erythroxylum species using liquid chromatography coupled with ESI-MSn and HRESIMS. Phytochemical Analysis, 32, 1011-1026. [CrossRef]
- Dos Santos, C., Galaverna, R. S., Angolini, C. F., Nunes, V. V., De Almeida, L. F., Ruiz, A. L., ... & Eberlin, M. N. (2018). Antioxidative, antiproliferative and antimicrobial activities of phenolic compounds from three Myrcia species. Molecules, 23, 986. [CrossRef]
- https://doi.org/10.3390/molecules23050986. [CrossRef]
- Fabre, N., Rustan, I., de Hoffmann, E., & Quetin-Leclercq, J. (2001). Determination of flavone, flavonol, and flavanone aglycones by negative ion liquid chromatography electrospray ion trap mass spectrometry. Journal of the American society for mass spectrometry, 12, 707-715. [CrossRef]
- Fabrikov, D., Guil-Guerrero, J. L., González-Fernández, M. J., Rodríguez-García, I., Gómez-Mercado, F., Urrestarazu, M., ... & Lyashenko, S. (2019). Borage oil: Tocopherols, sterols and squalene in farmed and endemic-wild Borago species. Journal of Food Composition and Analysis, 83, 103299. [CrossRef]
- https://doi.org/10.1016/j.jfca.2019.103299. [CrossRef]
- Golovenko, E., Lyashenko, S., Akimova, S., Mitina, L., Mulenkova, E., Belarbi, E. H., & Guil-Guerrero, J. L. (2021). Gamma-linolenic acid from fifty-seven Ribes species and cultivars. Plant Foods for Human Nutrition, 76, 385-393. [CrossRef]
- Granato, D., Fidelis, M., Haapakoski, M., dos Santos Lima, A., Viil, J., Hellström, J., ... & Pap, N. (2022). Enzyme-assisted extraction of anthocyanins and other phenolic compounds from blackcurrant (Ribes nigrum L.) press cake: From processing to bioactivities. Food Chemistry, 391, 133240. [CrossRef]
- https://doi.org/10.1016/j.foodchem.2022.133240. [CrossRef]
- Jiménez-Aspee, F., Thomas-Valdés, S., Schulz, A., Ladio, A., Theoduloz, C., & Schmeda-Hirschmann, G. (2015). Antioxidant activity and phenolic profiles of the wild currant Ribes magellanicum from Chilean and Argentinean Patagonia. Food Science & Nutrition, 4(4), 595–610. [CrossRef]
- Jurgoński, A., Fotschki, B., & Juśkiewicz, J. (2015). Disparate metabolic effects of blackcurrant seed oil in rats fed a basal and obesogenic diet. European Journal of Nutrition, 54, 991–999. [CrossRef]
- Kapasakalidis, P. G., Rastall, R. A., & Gordon, M. H. (2006). Extraction of polyphenols from processed black currant (Ribes nigrum L.) residues. Journal of agricultural and food chemistry, 54, 4016-4021. [CrossRef]
- Liu, B., & Li, Z. (2016). Black currant (Ribes nigrum L.) extract induces apoptosis of MKN-45 and TE-1 cells through MAPK-and PI3K/Akt-mediated mitochondrial pathways. Journal of medicinal food, 19, 365-373. [CrossRef]
- Lyashenko, S., González-Fernández, M. J., Gomez-Mercado, F., Yunusova, S., Denisenko, O., & Guil-Guerrero, J. L. (2019). Ribes taxa: A promising source of γ-linolenic acid-rich functional oils. Food chemistry, 301, 125309. [CrossRef]
- Lyashenko, S., Fabrikov, D., González-Fernández, M. J., Gómez-Mercado, F., Ruiz, R. L., Fedorov, A., ... & Guil-Guerrero, J. L. (2021). Phenolic composition and in vitro antiproliferative activity of Borago spp. seed extracts on HT-29 cancer cells. Food Bioscience, 42, 101043. [CrossRef]
- Ma, Y. L., Li, Q. M., Van den Heuvel, H., & Claeys, M. (1997). Characterization of flavone and flavonol aglycones by collision-induced dissociation tandem mass spectrometry. Rapid communications in mass spectrometry, 11, 1357-1364. [CrossRef]
- https://doi.org/10.1002/(SICI)1097-0231(199708)11:12<1357::AID-RCM983>3.0.CO;2-9. [CrossRef]
- March, R. E., & Miao, X. S. (2004). A fragmentation study of kaempferol using electrospray quadrupole time-of-flight mass spectrometry at high mass resolution. International Journal of Mass Spectrometry, 231, 157-167. [CrossRef]
- Marcum, C. L. (2016). Fundamental studies of collision-activated dissociation (CAD) of deprotonated model compounds relevant to lignin degradation products (Doctoral dissertation, Purdue University).
- Mikulic-Petkovsek, M., Rescic, J., Schmitzer, V., Stampar, F., Slatnar, A., Koron, D., & Veberic, R. (2015). Changes in fruit quality parameters of four Ribes species during ripening. Food Chemistry, 173, 363-374. [CrossRef]
- Peña-Morán, O. A., Villarreal, M. L., Álvarez-Berber, L., Meneses-Acosta, A., & Rodríguez-López, V. (2016). Cytotoxicity, post-treatment recovery, and selectivity analysis of naturally occurring podophyllotoxins from Bursera fagaroides var. fagaroides on breast cancer cell lines. Molecules, 21, 1013. [CrossRef]
- Ramos-Bueno, R. P., González-Fernández, M. J., & Guil-Guerrero, J. L. (2016). Various acylglycerols from common oils exert different antitumor activities on colorectal cancer cells. Nutrition and Cancer, 68, 518-529. [CrossRef]
- Schulz, E., Tohge, T., Zuther, E., Fernie, A. R., & Hincha, D. K. (2016). Flavonoids are determinants of freezing tolerance and cold acclimation in Arabidopsis thaliana. Scientific reports, 6, 34027. [CrossRef]
- https://doi.org/10.1038/srep34027. [CrossRef]
- Singleton, V. L., Orthofer, R., & Lamuela-Raventós, R. M. (1999). Analysis of total phenols and other oxidation substrates and antioxidants by means of folin-ciocalteu reagent. In Methods in enzymology (Vol. 299, pp. 152-178). Academic press. [CrossRef]
- Śliwka-Kaszyńska, M., Anusiewicz, I., & Skurski, P. (2022). The Mechanism of a Retro-Diels–Alder Fragmentation of Luteolin: Theoretical Studies Supported by Electrospray Ionization Tandem Mass Spectrometry Results. Molecules, 27, 1032. [CrossRef]
- Tuli, H. S., Garg, V. K., Bhushan, S., Uttam, V., Sharma, U., Jain, A., ... & Sethi, G. (2023). Natural flavonoids exhibit potent anticancer activity by targeting microRNAs in cancer: A signature step hinting towards clinical perfection. Translational Oncology, 27, 101596. [CrossRef]
- https://doi.org/10.1016/j.tranon.2022.101596. [CrossRef]
- Van Hoed, V., Barbouche, I., De Clercq, N., Dewettinck, K., Slah, M., Leber, E., & Verhé, R. (2011). Influence of filtering of cold pressed berry seed oils on their antioxidant profile and quality characteristics. Food Chemistry, 127, 1848-1855. [CrossRef]
- Vichitsakul K, Laowichuwakonnukul K, Soontornworajit B, Poomipark N, Itharat A, & Rotkrua P. (2023). Anti-proliferation and induction of mitochondria-mediated apoptosis by Garcinia hanburyi resin in colorectal cancer cells. Heliyon 22, e16411. [CrossRef]
- Wójciak, M., Mazurek, B., Tyśkiewicz, K., Kondracka, M., Wójcicka, G., Blicharski, T., & Sowa, I. (2022). Blackcurrant (Ribes nigrum L.) seeds—a valuable byproduct for further processing. Molecules, 2, 8679. [CrossRef]
- Yang, B., Zheng, J., Laaksonen, O., Tahvonen, R., & Kallio, H. (2013). Effects of latitude and weather conditions on phenolic compounds in currant (Ribes spp.) cultivars. Journal of Agricultural and Food Chemistry, 61, 3517-3532. [CrossRef]
- Yang, Y., Lei, Z., Zhao, M., Wu, C., Wang, L., & Xu, Y. (2020). Microwave-assisted extraction of an acidic polysaccharide from Ribes nigrum L.: Structural characteristics and biological activities. Industrial Crops and Products, 147, 112249. [CrossRef]
Figure 1.
Structure of phenolic compounds detected in Ribes seeds.
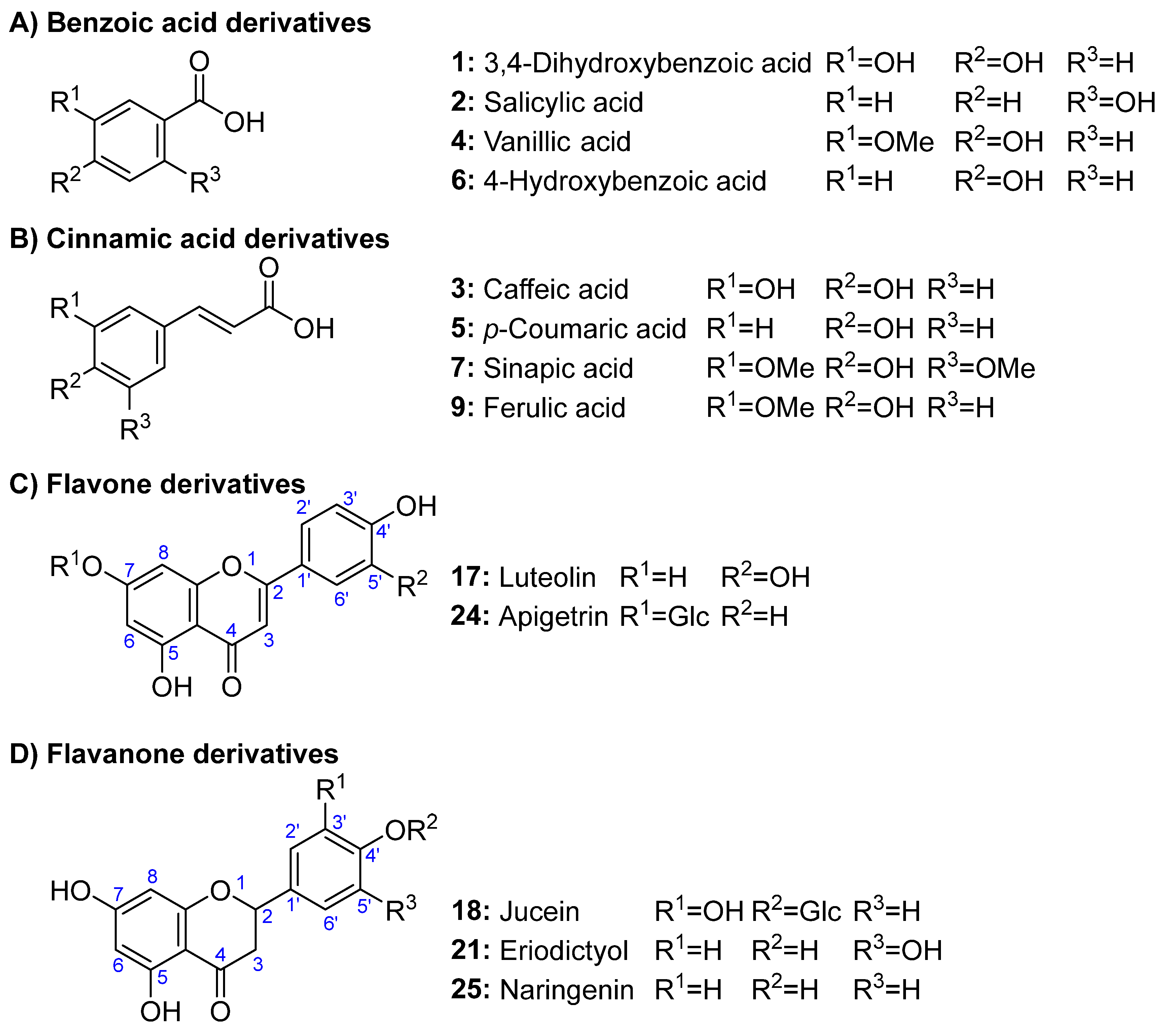
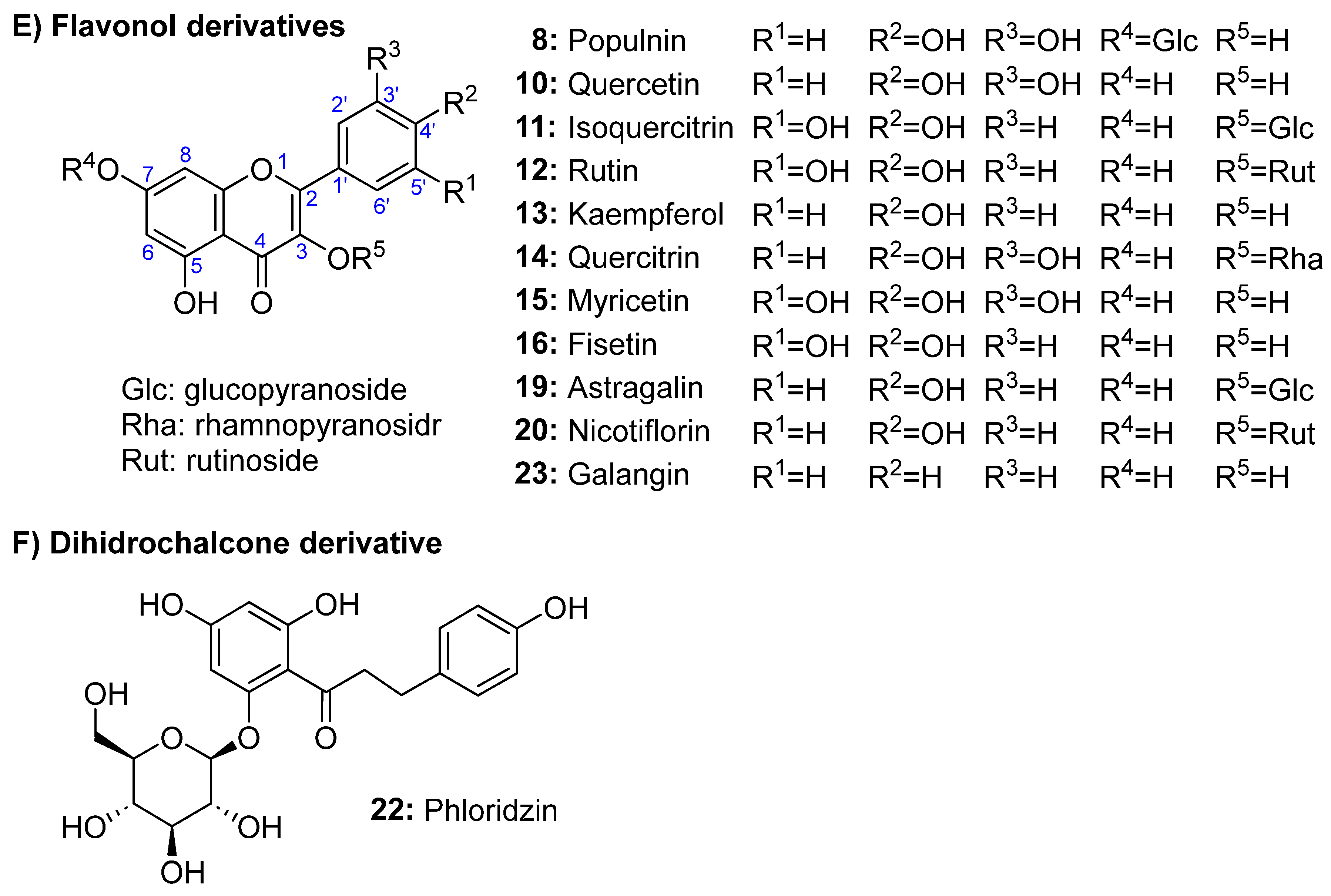
Figure 2.
MTT assay. A: Concentration-response plot for HT-29 cells after exposure to seed extracts for 48 (A) and 72 h (B). Data represent the mean of three complete independent experiments ± SD (error bars).
Figure 2.
MTT assay. A: Concentration-response plot for HT-29 cells after exposure to seed extracts for 48 (A) and 72 h (B). Data represent the mean of three complete independent experiments ± SD (error bars).
Figure 3.
MTT assay. GI50 after HT-29 cells exposure for 48 and 72 h to seed extracts, as well as to ferulic acid and doxorucicin (positive control). The GI50 value is detailed over columns, and the Selectivity Index (SI) for 72-h exposed cells to seeds extracts is shown in parentheses. Data represent the mean of three complete independent experiments ± SD (error bars). In a bar, means followed by different lower case letters (for 48 h treatment) and capital letters (for 72 h) are significantly different at p<0.05.
Figure 3.
MTT assay. GI50 after HT-29 cells exposure for 48 and 72 h to seed extracts, as well as to ferulic acid and doxorucicin (positive control). The GI50 value is detailed over columns, and the Selectivity Index (SI) for 72-h exposed cells to seeds extracts is shown in parentheses. Data represent the mean of three complete independent experiments ± SD (error bars). In a bar, means followed by different lower case letters (for 48 h treatment) and capital letters (for 72 h) are significantly different at p<0.05.
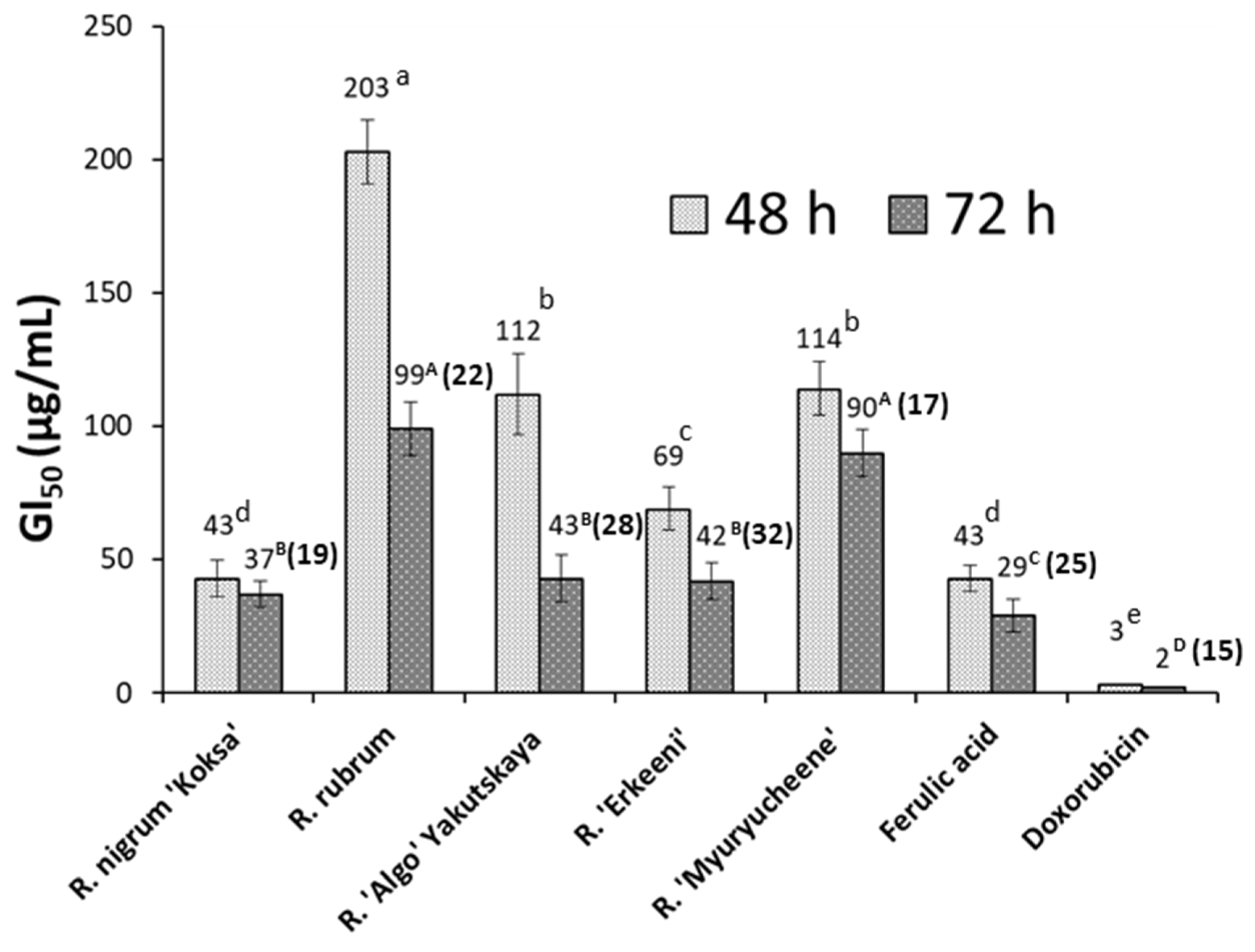

Table 1.
Data on collection, oil, and total phenolics content of Ribes samples.
| Code | Samples | Sample location | Total oil content g/100 g seeds |
TPC (mg CAE/g seeds)abc |
TPC (mg CAE/g oil)abc |
| Subgenus Ribes (Currants) | |||||
| Sect. Berisia Spach (Alpine currants) | |||||
| 1A | R. alpinum | Sukachev Institute of Forest of the Siberian Branch of the RAS, Krasnoyarsk, Russia | 19.9±0.5b | 36.9±1.8d | 7.3±0.3e |
| 1B | R. alpinum | Sierra de Baza, Granada, Spain | 12.7±0.4f | 33.4±0.9de | 4.2±0.1hi |
| 2 | R. pulchellum | Sukachev Institute of Forest of the Siberian Branch of the RAS, Krasnoyarsk, Russia | 23.0±1.0a | 34.2±1.2de | 7.9±0.2de |
| Sect. Coreosma (Spach) Jancz. (Black Currants) | |||||
| 3 | R. dikuscha | Botanical Garden of North-Eastern Federal University, Yakutsk, Russia | 17.8±0.2c | 30.5±2.4e | 5.4±0.0g |
| 4 | R. hudsonianum | Botanical Garden of North-Eastern Federal University, Yakutsk, Russia | 25.6±0.8a | 46.1±3.2c | 11.8±0.1b |
| 5A | R. nigrum ‘Hara katarlik’ | Botanical Garden of North-Eastern Federal University, Yakutsk, Russia | 18.4±0.1b | 53.4±2.5b | 9.8±0.2c |
| 5B | R. nigrum ‘Koksa’ | Botanical Garden of North-Eastern Federal University, Yakutsk, Russia | 16.3±0.0de | 94.8±3.4a | 15.5±0.1a |
| 6 | R. ‘Algo’ Yakutskaya | Botanical Garden of North-Eastern Federal University, Yakutsk, Russia | 17.0±0.3cd | 48.9±2.8bc | 8.3±0.2d |
| 7 | Ribes ‘Erkeeni’ | Botanical Garden of North-Eastern Federal University, Yakutsk, Russia | 18.3±0.2bc | 49.0±2.6b | 9.0±0.2cd |
| 8 | Ribes ‘Myuryucheene’ | Botanical Garden of North-Eastern Federal University, Yakutsk, Russia | 17.7±0.6c | 34.4±1.9de | 6.1±0.4f |
| Sect. Ribes (Red Currants) | |||||
| 9 | R. glabellum | Botanical Garden of North-Eastern Federal University, Yakutsk, Russia | 14.9±0.4e | 30.8±2.0e | 4.6±0.2h |
| 10 | R. triste | Dendropark “Alexandria” NAS of Ukraine, Belaja Tserkov, Ukraine | 18.5±0.5bc | 31.2±2.9e | 5.8±0.3fg |
| 11 | R. rubrum | Botanical Garden of North-Eastern Federal University, Yakutsk, Russia | 15.0±0.2e | 25.8±3.1f | 3.9±0.1i |
| aData represent means ± standard deviation of samples analyzed in triplicate; bDifferences in TPC amounts were tested according to one-way ANOVA followed by Duncan’s test; c Within a column, means followed by different letter are significantly different at P<0.05. | |||||
Table 2.
Identification of phenolic compounds in the seeds of selected Ribes taxa using LC-MS.
| N | Rt min | Massa m/z | Adduct | Fragmentb | Formula | Identification | Identification basis | Occurrence in samplesc |
| 1 | 3.88 | 153.01868 | [M-H]- | 109.02970 | C7H6O4 | 3,4-Dihydroxybenzoic (protocatechuic) acid | Molecular ion [M–H]− m/z 153, and at m/z 109, produced after the neutral loss of CO2 (44 Da) | 1B,5B,6,7,8,9,10 |
| 2 | 5.12 | 139.03909 | [M-H]- | 93.03460 | C7H6O3 | Salicylic acid | Molecular ion [M–H]− m/z 137, which further yielded a fragment ion at m/z 93, due to the loss of a CO2 group | 5A,5B,6,7,10,11 |
| 3 | 8.72 | 179.03498 | [M-H]- | 135.04810 | C9H8O4 | Caffeic acid | Molecular ion [M–H]− m/z 179 and its characteristic product ion 135, due to the loss of the CO2 group | 1A,1B,2,5A,5B,6,7,10 |
| 4 | 13.92 | 167.03498 | [M-H]- | 152.00996 | C8H8O4 | Vanillic acid | Molecular ion [M–H]− m/z 167 and its characteristic product ion 152, due to the loss of CH4 | 1A,1B,2,5B,6,7,8,10 |
| 5 | 16.68 | 163.04007 | [M-H]- | 119.04881 | C9H8O3 | p-coumaric acid | Molecular ion [M–H]− m/z 163 and its characteristic product ion 119, due to the loss of the CO2 group | 1A,1B,2,5A,5B,6,7,8,9,11 |
| 6 | 24.56 | 137.02442 | [M-H]- | 93.03325 | C7H6O3 | 4-hydroxybenzoic acid | Molecular ion [M–H]− m/z 137 and its characteristic product ion 93, generated by the loss of the CO2 group | 1A,1B,2,3,4,5A,5B,6,7,8,9,10,11 |
| 7 | 26.41 | 223.06120 | [M-H]- | 121.02821 | C11H12O5 | Sinapic acid | Molecular ion [M–H]− m/z 223, and the loss of 2CH3 – CO2 – CO (m/z 121) (Marcum 2016) | 1A,1B,5B,6,7,8,9,11 |
| 8 | 28.01 | 447.09328 | [M-H]- | 257.04496 | C21H20O11 | Populnin (kaempferol-7-O-glucoside) | Molecular ion [M–H]− m/z 447 and m/z 257, corresponding to the fragment [M–H–CO]− . The ejection of CO is notably followed by B-ring rotation and bonding with the A-ring to form the fused ring structure of m/z 257 (March and Miao, 2004). | 1A,1B,6,7,8 |
| 9 | 28.2 | 193.05063 | [M-H]- | 134.03690 | C10H10O4 | Ferulic acid | Molecular ion [M–H]− m/z 193, and m/z 134 corresponding to the loss of CO2 and CH3 | 1A,1B,2,4,5A,5B,6,7,8, 9,10,11 |
| 10 | 28.62 | 303.04993 | [M+H]+ | 178.99749 | C15H10O7 | Quercetin | Molecular ion [M–H]− m/z 303 and m/z 179, originated after cleavage of the B ring by a Retro Diels-Alder (RDA) mechanism (Dos Santos et al., 2018) | 1A,1B,5B,6,7,8,9,11 |
| 11 | 28.81 | 463.08820 | [M-H]- | 302.03696 | C21H20O12 | Isoquercitrin (quercetin-3-O-glucoside) | Molecular ion [M–H]− m/z 463, and m/z 302, corresponding to the aglycone of quercetin following the loss of a hexose ([M – H − 162]− | 1B,2,4,5A,5B,6,7,8 |
| 12 | 28.83 | 609.14611 | [M-H]- | 301.03474 | C27H30O16 | Rutin (quercetin 3-O rutinoside) | Molecular ion [M–H]− m/z 609, and fragment m/z 301 due to the loss of 308 Da (rutinose) | 1A,1B,2,3,4,5A,5B,6,7,8,11 |
| 13 | 29.57 | 287.05501 | [M+H]+ | 153.01760 | C15H10O6 | Kaempferol | Molecular ion [M–H]− m/z 287, and m/z 153 formed by RDA fragmentation wherein bonds 1 and 3 undergo scission leading to the formation of the A+ ion (m/z 153) (Ma et al., 1997) | 1A,1B,5B,6,7,8,9,11 |
| 14 | 29.75 | 447.09328 | [M-H]- | 230.98517 | C21H20O11 | Quercitrin (quercetin 3-O-rhamnoside) | Molecular ion [M–H]− m/z 447, and fragment m/z 231, corresponding to [quercetin-H-CO2-CO]- | 5B,6,7,8,9,10,11 |
| 15 | 29.77 | 317.03029 | [M-H]- | 151.00262 | C15H10O8 | Myricetin | Molecular ion [M–H]− m/z 317, and typical MS/MS fragment at m/z 151, that corresponded to retrocyclization on the A–C ring (1,2A−) and the consecutive loss of CO (1,2A−-CO) (Chernosov et al., 2017) | 1A,1B,2,3,4,5A,5B,6,7,8, 10 |
| 16 | 29.80 | 285.04046 | [M-H]- | 121.02799 | C15H10O6 | Fisetin | Molecular ion [M–H]− m/z 285, and m/z 121, that correspond to fragmentation of B ring (1,2B-), as described by Fabre et al. (2001) | 5B,6,7,9 |
| 17 | 29.80 | 285.04046 | [M-H]- | 175.03898 | C15H10O6 | Luteolin | Molecular ion [M–H]− m/z 285, and m/z 175, corresponding to the loss of C3O2 – C2H2O (Śliwka-Kaszyńska et al., 2022) | 1A,1B,5B,6,7,8,9 |
| 18 | 29.90 | 447.09328 | [M-H]- | 285.03995 | C21H20O11 | Juncein (luteolin-4′-O-glucoside) | Molecular ion [M–H]− m/z 447, and m/z 285 corresponding to luteolin aglycone, indicating the loss of a hexose | 1A,1B,5B,6,7,8,11 |
| 19 | 29.92 | 447.09328 | [M-H]- | 255.02924 | C15H10O6 | Astragalin (kaempferol-3-O-glucoside) | Molecular ion [M–H]− m/z 447, and m/z 255, corresponding to the loss of the CH2O from the aglycone (30 Da) (Dantas et al., 2021). | 1A,1B,5B,6,7,10,11 |
| 20 | 29.95 | 593.15119 | [M-H]- | 285.03973 | C27H30O15 | Nicotiflorin (kaempferol-3-O-rutinoside) | Molecular ion [M–H]− m/z 593, and m/z 285 corresponding to a deprotonated kaempferol aglycone, and further loss of the rutinoside moiety | 1A,1B,5B,6,7,8,10,11 |
| 21 | 30.06 | 287.05611 | [M-H]- | 135.04382 | C15H12O6 | Eriodictyol | Molecular ion [M–H]− m/z 287, and m/z 135 corresponding to fragmentation of the B ring (1,3B-), as described by Fabre et al. (2001) | 5B,6,7 |
| 22 | 30.53 | 435.12967 | [M-H]- | 273.07598 | C21H24O10 | Phloridzin (phloretin-2′-O-glucoside) | Molecular ion [M–H]− m/z 435, and m/z 273 corresponding to phloretin (dihydronaringenin), after the losses of a hexosyl (glucose, 162 Da) | 5B,6,7 |
| 23 | 30.78 | 269.04555 | [M-H]- | 213.0545 | C15H10O5 | Galangin | Molecular ion [M–H]− m/z 269, and m/z 213 corresponding to the loss of 2CO (56 Da) | 1A,3,4,5A,5B,6,7,8,9,10,11 |
| 24 | 30.87 | 433.11292 | [M+H]+ | 271.05908 | C21H20O10 | Apigetrin (apigenin-7-O-glucoside) | Molecular ion [M+H]− m/z 433, and m/z 271 corresponding to the aglycon apiginin, by the loss of a glucose (162 Da) | 5B,6,11 |
| 25 | 31.14 | 271.06120 | [M-H]- | 119.04879 | C15H12O5 | Naringenin | Molecular ion [M–H]− m/z 271, and m/z 119 that correspond to fragmentation of the B ring (1,3B-), as described by Fabre et al. (2001) | 1A,1B,2,4,5A,5B,6,7,10 |
| a mass error lower than 5 ppm; b mass error lower than 10 ppm; c Sample codes as in Table 1 | ||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



