
Submitted:
20 February 2024
Posted:
21 February 2024
You are already at the latest version
Abstract
Abstract: Brassinosteroids (BR) are essential plant hormones that play a crucial role in regulating plant growth and development, as well as their resistance to abiotic and biotic stresses, including pathogen infections. In the context of plant im-munity, BR influence various processes such as the expression of genes related to pathogen defense, the production of antimicrobial compounds, and the re-inforcement of the plant cell structure. The aim of work is understand the role of BR in plant defense against abiotic and biotic stresses. BR are involved in the regulation of metabolic processes, water economy, and photosynthesis, contributing to increased plant resistance to abiotic stresses. The study discusses various mechanisms of BR action on plant immunity, including the regulation of gene expression related to defense, activation of the plant defense system, in-teraction with other plant hormones, regulation of water and salt balance, and influence on photosynthetic processes. In the context of plant immunity, BR influence various processes, including the expression of genes related to pathogen defense, the production of antimicrobial compounds, and the reinforcement of the plant cell structure. Additionally, BR are involved in the regulation of metabolic processes, water economy, and photosynthesis, con-tributing to increased plant resistance to abiotic and biotic stresses. The paper emphasizes the importance of further research on the role and mechanisms of action of BR in plant immunity, which may contribute to the development of more effective strategies for the use of BR in agriculture, horticulture, and plant protection.
Keywords:
plant hormones
; gene expression
; BR functions
; antimicrobial actions
; immunity mechanisms
; plant stress
1. Introduction
BR constitute a novel group of steroid hormones that share struc-tural similarities with hormones found in animals and insects. Initially identified in rapeseed pollen, their presence has been docu-mented across various plant species [1]. Despite their low concentra-tions in plants, BR profoundly influence numerous cellular and molec-ular func-tions [2]. They play a pivotal role in regulating physiological processes such as cell division, cell expansion, seed germination, de-velopment of conductive tissues, growth differentiation, reproduction, and responses to light (photomorphogenesis and scotomorphogenesis). Notably, in-appropriate levels of BR can result in phenotypic abnor-malities in plants. BR act as key growth regulators, offering plants the ability to counteract abiotic stresses such as drought, salinity, extreme tempera-tures, flooding, exposure to toxic heavy metals/metalloids, and other contaminants, all of which can lead to substantial agricultural crop losses.
Plants have evolved effective defense mechanisms, including the neutralization of Reactive Oxygen Species (ROS) generated under stress conditions. Over the last two decades, extensive research has enhanced our understanding of the complex of BR signaling pathway in response to various stresses. This knowledge has paved the way for the devel-opment of strategies to improve crop performance by manipu-lating BR signaling, biosynthesis, or perception [1,2,3].
BR, as phytohormones, play a crucial role in regulating plant growth and development, aiding plants in adapting to changing en-vi-ronmental conditions [3]. They are found in various plant organs and tissues, including roots, stems, leaves, pollen, anthers, and fruits [4,5]. The highest concentrations of BR, as reported by Li and He [6], are found in seeds, pollen, and fruits. Young plant tissues (1-100 ng/g fresh weight) generally contain higher BR concentrations than mature tissues (0.01-0.1 ng/g fresh weight) [7]. BR play a crucial role in various developmental and physiological processes, including cell expansion, cell division, maintenance of stem cells, vascular development, elongation of dif-fer-ent cell types [8,9]. They also influence hypocotyl growth [10], root and shoot development [11], stomatal regulation [12], seed germination [13], xylem shaping and differentiation [14], photomorphogenesis, plant re-production [15], and aging processes [16].
In the realm of plant immunity, BR regulate processes such as the expression of defense-associated genes, production of antimicrobial compounds, and reinforcement of plant cell structures. In response to pathogen infection, BR can enhance the production of lignin, phenolic compounds, and glycosides, fortifying cell walls against bacterial, viral, or fungal infections [2,17,18]. Regarding plant resilience to abiotic stresses, BR are involved in regulating metabolic processes, including the accumulation of osmotic substances and the adaptation of chlo-ro-plasts to light stress. Additionally, BR contribute to increasing the sta-bility of cell membranes, protecting them against oxidative stress [18,19,20]. Furthermore, BR stimulate root growth, aiding plants in water and mineral nutrient absorption from the soil, thereby enhancing re-sil-ience to abiotic stresses [17,20,21].
Manipulating the levels of BR in plants can impact their ability to adapt to both abiotic and biotic stress. For instance, increasing BR levels can enhance resistance to drought and cold but may simultaneously reduce resistance to pathogen infections [22,23,24,25,26]. The adjustment of BR signaling, biosynthesis pathways, or perception holds significant po-tential for improving crop yields [23,27]. Therefore, the objective of this study was to investigate and analyze the actual role of BR in sig-naling cascades, emphasizing their positive contribution to biotic and abiotic stress tolerance.
2. Structure and Functions of BR
2.1. Structure and Biosynthesis of BR
Structure and Biosynthesis: BR are steroid compounds charac-ter-ized by a complex structure, featuring a steroid ring with two side chains. Their structure is based on triterpenoids and includes several distinctive elements [14,23,28]. The general chemical structure of BR can be outlined as follows:
- −
- −
- −
- −
- −
Examples of BR include brassinolide (BL), castasterone (CS), and teasterone (TZ), which differ in chemical structure but exhibit similar functions as phytohormones influencing plant development and re-sponses to environmental stresses. BL and CS are considered the most important BR due to their significant biological activity and widespread occurrence in plants. However, due to commercial availa-bility, 24-epibrassinolide (EBR) is the most commonly used BR in studies on exogenous steroid phytohormones in plants [31]. Since the discovery of BL, approximately 70 different BR have been identified in 64 plant species, including 6 gymnosperms, 53 angiosperms, one horsetail spe-cies (Equisetum arvense), three algae species (Hydrodictyon reticulatum, Cystoseira myrica, and Chlorella vulgaris), one moss species (Marchantia polymorpha), one moss species (Physcomitrella patens), lycopods (Selagi-nella moellendorffii and S. uncinata), and thirteen fern species [6,32,33]. It has been observed that in Cupressus arizonica pollen, the concentration of 6-deoxotyphasterol (6-deoxoTY) was 6400 times higher than BL. The highest BR concentration, amounting to 6.4 mg 6-deoxoTY per kilogram (kg) of pollen, was found in C. arizonica [22]. To date, only fifty-two BR have been detailed in terms of their biological activity in plants [31]. CS, BL, TY, 6-deoxoCS, TE, and 28-norCS are the most common BR in plants [33]. The greatest diversity of BR (2 conjugated forms and 25 free forms) was found in immature common bean (Phaseolus vulgaris) seeds [34]. Various BR have also been identified in shoots, seeds, and pollen of other leguminous plants, with their content ranging from 0.007 to 628 μg/g fresh weight [35].
According to Chaudhuri et al. [27], BRs are a group of naturally occurring polyhydroxylated phytosterols that have at least one oxygen group at position C3, as well as additional groups at one or more carbon atoms, such as C2, C6, C22, and C23 [36]. Their chemical structure can be schematically represented (Figure 1), [28].
The transport and metabolism of brassinosteroids (BRs) in plants are complex processes with significant implications for growth, development, and stress responses. More than 70 BR derivatives can be produced through various metabolic transformations, including acylation, demethylation, epimerization, glucosylation, hydroxylation, side chain cleavage, sulfonation, and oxidation [22,28,30,31,37,38,39,40,41,42].
Exogenous BR transport in plants occurs in a directed manner, known as basipetal transport, from apices to the base. This process, independent of vessel movement, involves energy consumption and specific carrier proteins in the cell membrane. Calcium ion levels may influence this process. When applied to roots, BR transport becomes acropetal, moving toward leaves. Endogenous BR transport remains unclear [14,15,22].
BR perception occurs on the cell surface through receptors, leading to a phosphorylation cascade activating the transcription factor BZR1. BZR1 regulates BR-responsive gene transcription, influencing photosynthesis efficiency and plant biomass accumulation. Plant responses to BR vary, and genetic studies suggest a positive correlation between endogenous BR levels and abiotic stress tolerance [36,37,38,39,40,41,42,43].
Genetic studies show a positive correlation between the level of endogenous BR and tolerance to abiotic stress, although this assumption contradicts the behavior of some BR mutants under stressful conditions. It is worth noting that plant responses to BR vary significantly de-pending on the plant variety and species, developmental stages, and environmental conditions. Additionally, other hormones and signaling molecules participating in fine-tuning BR action also play a significant role in plant adaptation to stress [15,22,36,40]. An example is cucumber plants, where the application of exoge-nous 24-epibrassinolide (24-epiBL) to the roots causes acropetal transport of this compound in the plant. It has been shown that 24-epiBL applied to cucumber roots was metabolized only in the leaves, trans-forming into 2,24-diepiBL [35,41]. Fatty acids and/or sugars may also be attached to the hydroxyl groups of the A ring or the side chain of BR. Exogenous BRs seem to move very slowly or not at all within plants after application to leaves. Even when BR-deficient plant mutants are grafted onto wild-type plants, it does not lead to changes in the plant’s pheno-type, according to Bajguz & Hayat [36]. Removing the growth tip or mature leaves also does not lower the BR levels in the remaining parts of the plant. For example, in tomatoes, the presence of wild forms does not significantly affect the growth of BR-deficient forms when both types are present in the same leaf. Despite the fact that BRs do not undergo long-distance movements, they can still influence signaling over greater distances by modifying the transport of, for example, IAA produced in growth cones. Enzymes involved in BR biosynthesis are located inside cells, especially in the endoplasmic reticulum. BR reception occurs on the external surface of the cell, so BRs must move from inside the cell to the outside, where they are recognized by the same cell or neighboring cells [36,38,43,44].
Metabolic transformations of exogenous BR, such as in cucumber plants, result in acropetal transport, producing different derivatives like 2,24-diepiBL [37]. Despite limited long-distance movement, BRs influence signaling over greater distances by modifying the transport of other compounds. Enzymes involved in BR biosynthesis are located intracellularly, while reception occurs externally [14,45,46].
A feedback mechanism suggests cells can perceive and produce BRs, indicating complex transport and perception processes. The biosynthesis of BR involves sterol processing, hydroxylation, reduction, C-22 hydroxylation, cyclopropylation, C-23 hydroxylation, and the final reduction to form BR. BRs act as steroid hormones influencing plant growth, development, and stress responses [30,33].
Recent research by Aitken et al. [30] explores the synthesis and evaluation of brassinosteroid (BR) analogs with modified alkyl side chains, particularly incorporating phenyl or benzoate groups. Their study indicates that introducing fluorine atoms (F) enhances the bioactivity of these analogs. Building on this, a series of 23,24-bisnorcholenic analogs with benzoate groups substituted with F atoms in ortho or para positions was synthesized and assessed for their impact on plant growth stimulation. Results demonstrate that analogs with an F atom in the para position show the highest activity, often exceeding that of brassinolide. Docking studies reveal that these F-containing compounds adopt a similar orientation to brassinolide, with the F atom in the para position forming an additional hydrogen bond in the binding site. Overall, these findings suggest that BR analogs with fluorine substitutions exhibit promising biological activity, offering potential for enhancing plant growth under stressful conditions [47,48,49,50,51].
Exogenous BR application or manipulation of biosynthetic pathways can enhance yields. Signaling involving BR, IAA, and GA plays a crucial role in regulating various processes, including leaf angle development. A phosphorelay signaling system governs BR signal transduction, regulating plant growth. BR binding initiates phosphorylation events, activating downstream kinases and transcription factors, ultimately influencing gene expression [48,52,53]. Jo et al. [54] conducted a comprehensive study on cucumber metabolomics and gene expression to understand the metabolism of bioactive compounds. They found consistent expression patterns between carotenoid and chlorophyll-related genes and the metabolome analysis. The upregulation of carotenoid and chlorophyll metabolism in fruit peel and flesh, exhibiting the highest antioxidant activity, suggests their contribution to fruit quality. The identified bioactive compounds can serve as biomarkers for commercial fruit and vegetable quality.
The application of exogenous BR or manipulation of biosynthetic and signaling pathways can potentially increase yields. BR, IAA, and GA signaling plays a crucial role in regulating leaf blade bending in rice, and their manipulation could enhance crop yields [44,53,61]. A complex phosphorelay signaling system governs BR signal transduction, regulating plant growth and development through the activation of transcription factors like BZR1/2 (Figure 2).
BR binding phosphorylates BKI1, relieving BRI1 receptor inhibition. BRI1 activation involves mutual phosphorylation with BAK1 or other SERK family members. BRI1 then phosphorylates BSK1 and CDG1, activating BSU1 and PP2A. These dephosphorylate and deactivate BIN2, allowing BR response transcription factors like BZR1/2 to accumulate in the nucleus and bind to DNA, along with other transcription factors [48]. In summary, the transport, metabolism, and perception of BR in plants are intricate processes with significant implications for plant physiology, growth, and stress responses, necessitating further research in this area.
The biosynthetic pathway of BRs in rice involves enzymatic reactions:
- a)
- Squalene -> campesterol by SQE.
- b)
- Campesterol -> obtusifoliol by CYP90B2.
- c)
- Obtusifoliol -> epicastasterone by CYP90D1.
- d)
- Epicastasterone -> teasterone by CYP85A2.
- e)
Enzymes in the phytosterol biosynthetic pathway include squalene epoxidase (SQE), cycloartenol synthase, C24 methylation enzymes, cyclopropylsterol isomerase, CYP51, Fackel/Hydra2/Extra-Long-Lifespan 1 (Fk/Hyd2/Ell1), and Δ8-Δ7 sterol isomerase (HYDRA1/HYD1). They convert precursor molecules into phytosterols, serving as intermediates for BR biosynthesis [46,47,50,55,56].Typhasterone is the final product in the BR biosynthetic pathway, further modified into active forms like brassinolide (BL) and castasterone (CS). BR biosynthesis is a complex process occurring in different cellular compartments and regulates various physiological processes in plants, including growth, cell expansion, and their stress responses [4,47,54].
Receptor-like kinases (RLKs) are vital surface receptors in plants, crucial for signaling pathways. RLK structure includes three domains: extracellular for ligand binding, transmembrane for membrane anchoring, and cytoplasmic kinase for initiating intracellular signaling (Figure 3) [4,12,22,35,36].
RLKs play diverse roles in plant signaling, influencing defense responses, hormone perception, and development. They mediate extracellular signals, regulating growth, immunity, and environmental adaptation [51,57,58,59]. Recent research highlights their involvement in complex signaling networks, interacting with other proteins dynamically [57]. Understanding RLK-mediated pathways is crucial for enhancing crop performance amid changing conditions. Some RLKs share signaling cascades, like BRI1, EMS1, and NILR1, suggesting intricate signaling mechanisms [4,39,51,57]. Advanced techniques such as domain swapping and single-cell omics analysis can deepen our understanding of RLK pathways with precision.
New techniques are needed to identify RLK-binding ligands, complementing existing proteomic and genetic approaches. Functional redundancy among RLKs complicates their identification; for instance, BAK1 serves as a co-receptor for BRI1 and regulates immune responses through FLS2 and EFR pathways. Advanced methods like CRISPR and sophisticated phenotypic analysis can address this redundancy [4,57].
RLKs often participate in multiple pathways, posing challenges in understanding their specific functions. Technologies such as genetics, structural biology, and proteomics will be crucial for unraveling these complexities, ultimately improving crop yields and developing stress-tolerant varieties through bioengineering and breeding approaches [58,59,60].
BRI1 is a pivotal receptor for BR, crucial in plant physiology and development. Mutations in its gene can disrupt plant growth. Other BR signaling components include BAK1, BIN2, BSU1 phosphatase, and BZR transcription factors [40]. Despite progress, uncertainties remain in BRI1 kinase activation. Continuous discovery of new signaling components highlights the complexity of BR pathways and their integration with other signaling networks, essential for plant development and defense. Understanding these mechanisms may unlock new agricultural applications [35,60].
2.2. Physiological Functions of BR in Plant Growth and Development
Research on BR-deficient mutants has identified them as a novel class of phytohormones, yet many of their functions remain undiscovered due to limitations in analyzing plant species. Specific inhibitors of BR biosynthesis can be valuable tools for studying their role in various plant developmental stages, such as germination, leaf development, pollen formation, and flowering. BR influences plant growth, development, and responses to abiotic and biotic stresses, including stomatal apparatus formation, sexual plant development, etiolation, elongation of the pistil, leaf morphology, responses to atmospheric pollution, and thermotolerance [27,28,61].
Manipulating BR levels, both through exogenous application and changes in biosynthesis and signaling pathways, holds the potential to increase crop yields. Studies on growth regulation in Arabidopsis thaliana have highlighted the significant role of factors like WRKY46, WRKY54, and WRKY70, activated by BR. The histone methyltransferase SDG8 plays a crucial role in regulating the expression of BR-related genes, affecting various aspects of plant development, yield, and quality. Inhibitors of BR biosynthesis, used to study BR-deficient mutants, can effectively elucidate the functions of these phytohormones [46,47,61,62,63].
Impact on Cell Growth: BR serves as a key regulator of cell growth, especially elongation, leading to increased shoot and root length. Approximately 70 different BRs have been identified, and they play essential roles in reactions such as stem growth, pollen tube growth, leaf bending, leaf unfolding, root growth inhibition, proton pump activation, ethylene production, and cell elongation [4,14,22,25,35,62,63,64].
Role of BR in Shaping Plant Morphology: BR influences various aspects of plant morphology, including leaf, bud, root, and flower shape. They are crucial for the development of cultivated plants and the production of secondary metabolites. BR also affects cell elongation, which is pivotal in the initial growth stages. Studies on BR-deficient mutants in cereal plants underscore the importance of these phytohormones in regulating plant morphology and architecture [15,40,65].
BR also influences cell elongation, crucial in the initial growth stages of plants. Research confirms the positive effect of BR on the formation of root nodules in pea plants (Pisum sativum) and the shaping of lateral roots, often regulated through synergistic interactions with auxins and other key plant hormones. Changes in signal transduction pathways and BR synthesis at the molecular level can lead to metabolic defects, influencing plant morphology and architecture, particularly in cereals. For example, dwarf cereal plants show resistance to lodging, enabling increased fertilizer doses and higher yields [9,10,66].
BR in Root Growth: Exogenous phytohormones like MRW or EBL promote marigold’s lateral root growth, suggesting that methane (CH4) induces lateral roots, possibly by increasing endogenous BR levels. CH4 affects cell wall components and enzyme activity during root formation, with BR potentially involved, highlighting BR’s role in CH4-induced lateral root growth and cell wall regulation [60].
BR in Xylem Development: BR influences xylem development in woody plants via the BR signaling pathway. Exogenous BR increases stem length and diameter, enhancing xylem area in stems and roots. Gene expression changes, including upregulation of EgrBRI1, EgrBZR1, and EgrBZR2, along with alterations in HDZIPIII and cellulose synthase genes, were observed. Six-month-old trees treated with BR showed increased xylem area, fiber cell length, and cell number, along with elevated expression of BR signaling and xylem-related transcription factor genes [69,70].
BR significantly impacts plant photomorphogenesis, influencing various morphological and physiological processes in response to light. Key aspects include:
- −
- −
- −
- −
- −
These mechanisms underscore BR’s role in plant adaptation to environmental conditions and the efficiency of growth and development.
3. Mechanisms of Defense Related to BR
3.1. Impact of BR on the Expression of Genes Related to Plant Defense
Regulation of defense-related gene expression: BR can influence the expression of genes related to plant defense, such as genes encoding antioxidant enzymes, anti-inflammatory proteins, or proteins involved in the synthesis of secondary compounds that play a role in plant defense against biotic and abiotic stresses [35,42,65].
BR can stimulate the production of secondary compounds, such as alkaloids, flavonoids, or phytoalexins, which play a role in plant resistance to pathogen attacks [65]. BR influences the activation of signaling pathways that regulate the plant’s defense response. They can activate kinases and transcription factors involved in the regulation of the expression of defense genes [14,76,77]. BR also influences the growth and development of plants, which may increase their resistance to mechanical stresses, such as damage caused by herbivores or wind. They can also regulate interactions with other plant hormones, such as jasmonates or salicylates, which are involved in plant defense responses. This can affect the complex regulation of plant defense [78,79].
3.2. Interactions between Electrical Signals and Plant Hormones
Interactions between electrical signals and plant hormones are vital for plant responses to environmental stresses. Plants employ a three-stage strategy involving cellular homeostasis, detoxification, and restoring growth processes [78,80,81]. Brassinosteroids (BR) become more active under stress, aiding in scavenging reactive oxygen species (ROS), maintaining redox balance, improving photosynthesis, and enhancing antioxidant systems [5,80,82,83]. Local stimuli trigger long-distance electrical signals that influence hormone levels in distant plant parts, mediating important integration points involving changes in pH, calcium ion levels, and ROS [5,83,84,85].
Research by Ladeynova et al. [47] sheds light on intricate interactions between electrical signals and plant hormones, elucidating systemic responses. Further studies are crucial to deepen our understanding of these interactions, particularly in managing environmental stress [47,80,86]. Action potentials (AP) and other electrical activity patterns influence physiological processes like photosynthesis and plant reactions to environmental stimuli [78].
Plant electrical activity is vital for understanding adaptive strategies with implications for agriculture and environmental conservation. Despite progress, unanswered questions persist, necessitating ongoing research [47,86,87].
Plants utilize a three-stage strategy to cope with environmental stresses: maintaining cellular homeostasis, detoxification, and restoring growth processes. Brassinosteroids (BR) play a significant role in stress alleviation by scavenging reactive oxygen species (ROS), enhancing photosynthesis, and strengthening the antioxidant system [88,89].
Local stimuli can trigger long-distance signals that affect hormone levels throughout the plant. Electrical signals, pH changes, and ROS are crucial in mediating these responses. Further research is needed to fully understand these interactions and their applications in agriculture, horticulture, forestry and plants protection [90,91,92,93,94].
Research highlights BR’s role in plant defense mechanisms, including the induction of pathogenesis-related genes (PR), increased expression of defense-related transcription factors, and regulation of the JA signaling pathway. BR also enhances plant tolerance to oxidative stress and influences plant-microbe interactions, improving overall plant health and productivity [2,17,80].
In summary, understanding the interplay between electrical signals, hormones, and defense mechanisms is crucial for developing effective stress management strategies in plants, with BR emerging as a key player in this process.
3.3. Regulation of Gene Expression Related to Immunity
Brassinosteroids (BRs) exert a significant influence on the expression of genes associated with plant defense against both biotic and abiotic stresses [3,6,70,94,95,96,97]. Here, we delve into the mechanisms underlying this regulation:
Gene Responses to Stresses: Genes modulated by BR respond to diverse stressors, encompassing those encoding antioxidant enzymes, anti-inflammatory proteins, signal receptors, and other factors implicated in plant defense [45,98,99,100].
Signaling Mechanisms Involved in Gene Expression Regulation via BR Signaling Pathways:
- −
- −
- −
- −
- Expression of Defense Genes: Transcription factors like BZR1 and BES1 wield influence over the expression of defense-related genes. They orchestrate the activation of genes involved in plant defense against a spectrum of stresses, both biotic and abiotic. These genes encompass those encoding enzymes for synthesizing chemical compounds, receptors for pathogen recognition, and factors pivotal for the growth and development of defense cells. Consequently, BR-mediated signaling pathways modulate the expression of defense genes, bolstering plants’ resilience against stressors and pathogens. Nonetheless, the specific targets and mechanisms underlying these processes may vary contingent upon the plant species and the nature of the stressors [14,15,58].
Pathogen Resistance: A plethora of studies underscores the pivotal role of BR in enhancing plant resistance to pathogen assaults, including those by fungi or bacteria. Alterations in the expression of defense-related genes significantly contribute to this heightened resistance [14,39,90]. Research conducted on model plants has unveiled the intricate interplay between signaling pathways involving salicylic acid (SA), jasmonic acid (JA), and ethylene (Et) in modulating plant defense specificity. Investigations into the biosynthesis and signaling of these compounds during infections have corroborated SA’s protective role against biotrophic pathogens, which thrive in living cells, while JA and Et synergize to combat necrotrophic organisms that feed on dead or dying cells [17,91,101].
3.4. Regulation of Water and Salt Economy
3.4.1. Drought Response
Brassinosteroids (BRs), a class of plant steroid hormones, play pivotal roles in modulating plant growth, development, and responses to abiotic stresses like drought. Recent research has shed light on several mechanisms through which BR influences plant responses to drought [92,93,94]:
- −
- −
- Regulation of Stomatal Dynamics: BRs are involved in the regulation of stomatal opening and closing on leaf surfaces. This regulation is vital for managing water loss through transpiration and optimizing gas exchange. By modulating stomatal behavior, BRs help plants conserve water during drought stress [12,94,95].
- −
- Protection Against Oxidative Stress: Acting as antioxidants, BRs play a role in safeguarding plant cells from oxidative stress induced by drought. Oxidative damage often arises from an imbalance between the production of reactive oxygen species and the cell’s antioxidant defense mechanisms. BRs contribute to maintaining cellular redox homeostasis, thereby protecting plants from oxidative injury [23,24,63,71].
- −
- Regulation of Drought-Responsive Gene Expression: BRs are implicated in the regulation of gene expression associated with drought response. Studies have identified genetic links between BR signaling and the activation of drought-responsive genes, highlighting the involvement of BRs in orchestrating molecular responses to water scarcity [56,99].
- −
- Enhanced Water Use Efficiency: Emerging evidence suggests that BR-treated plants may exhibit improved water use efficiency, allowing them to utilize available water resources more effectively. This enhanced efficiency can confer a competitive advantage to plants facing drought stress, enabling them to maintain essential physiological processes despite limited water availability [55,96,97].
These insights into the multifaceted roles of BRs in drought response underscore their significance as potential targets for enhancing crop resilience to water scarcity and advancing sustainable agricultural practices in the face of changing climatic conditions.
3.4.2. Salinity Protection
Soil salinity, affecting approximately 6% of the Earth’s land area, poses a significant challenge to plants and crop productivity worldwide. Saline soils are prevalent in dry and semi-arid climates, but anthropogenic activities such as excessive fertilization, pesticide use, and the application of de-icing salts contribute to salt accumulation even in humid regions. Soil salinity imposes both ionic and osmotic stresses on plants, disrupting metabolic processes, inducing nutrient deficiencies, and exacerbating oxidative stress. Plants respond to salinity through mechanisms aimed at minimizing salt entry and reducing salt concentrations within cells. These adaptive responses encompass a wide range of strategies, from morphological adaptations to alterations in gene expression, underscoring the complexity of plant responses to salinity stress. Understanding these mechanisms is crucial for developing strategies in plant breeding and genetic engineering to enhance salt tolerance in crops [95,96].
BRs play a significant role in plant defense against salinity stress through a variety of adaptive mechanisms, including:
- −
- −
- Stimulation of Root Development: BRs promote increased root development, enhancing the plant’s capacity to absorb water and nutrients from the soil, particularly crucial in saline environments where water availability is limited [23].
- −
- −
- −
- Osmotic Adjustment: BRs influence the accumulation of osmoprotectants such as proline and polyols, aiding in the maintenance of osmotic balance within cells under saline conditions, thus minimizing cellular damage [99].
- −
- Regulation of Gene Expression: BRs modulate the expression of stress-responsive genes, including those involved in salinity defense mechanisms, orchestrating molecular responses to mitigate the adverse effects of salinity stress [56].
- −
- −
- Ion Transport Regulation: BRs influence ion transport processes, including the uptake and translocation of ions such as sodium (Na+) and potassium (K+), crucial for maintaining ionic balance within plant cells and mitigating the harmful effects of excess salt accumulation [100].
In summary, BRs play a multifaceted role in enhancing plant tolerance to salinity stress, integrating various physiological and molecular processes to bolster plant resilience against this environmental challenge. Understanding the mechanisms underlying BR-mediated salinity protection holds promise for the development of strategies aimed at improving salt tolerance in crops, thereby contributing to sustainable agriculture in saline-affected regions.
3.5. Activation of Plant Defense System by BR
BRs play a pivotal role in activating the plant defense system against both abiotic and biotic stresses by regulating various metabolic and signaling processes. The mechanisms through which BR influences plant resistance to stressors are multifaceted and involve intricate interactions with various defense-related pathways and molecules [37,65,88,101].
Regulation of Phytohormone Production: BRs can stimulate the production of phytohormones such as jasmonic acid (JA) and salicylic acid (SA), which are essential for inducing plant defense responses. JA is primarily associated with defenses against pests, while SA is crucial for defense against pathogens [22,35,38,47]. By regulating the expression of defense-related genes, BRs activate signaling pathways that induce the synthesis of defense proteins like Pathogenesis-Related (PR) and Resistance (R) proteins, aiding the plant in combatting pathogens [78,91].
Enhancement of Surface Barriers: BRs may reinforce the plant’s surface barriers, making it more challenging for pests and pathogens to attack. This reinforcement could involve the production of physical deterrents such as trichomes or the secretion of chemicals that deter pathogens [91,92].
Stimulation of ROS Production: BRs stimulate the production of ROS, such as hydrogen peroxide (H2O2), which serve as signaling molecules to activate defense responses and exhibit toxic effects on pathogens [80]. Additionally, ROS influence the regulation of autophagy, a process in which plant cells eliminate damaged organelles and pathogens, aiding in the plant’s self-cleansing of infections [22].
Reinforcement of Cell and Tissue Structures: BRs may strengthen plant cell and tissue structures, limiting pathogen access to sensitive plant parts [43,69].
Control of Photosynthesis Process: BRs enhance the efficiency of the photosynthesis process, conserving energy and resources that can be allocated towards defense reactions [22,35,69,73,74].
Overall, BRs play a significant role in reinforcing plant resistance to biotic stresses, aiding in the fight against pathogens and pests, and stimulating defense responses [28,93]. However, the specific mechanisms and effects of BR-mediated defense activation may vary depending on factors such as the type of pathogen, plant variety, and species.
3.5.1. Mechanisms of Phytoalexin Induction by BR
Transcription Factor Activation: BR activates transcription factors like BZR1 and BES1, which regulate gene expression. These factors directly or indirectly influence genes involved in phytoalexin production by binding to their regulatory sites [88].
Interactions with Other Hormones: BR interacts with other plant hormones such as salicylic acid (SA) or jasmonic acid (JA), which play roles in regulating the plant’s defense response. By influencing interactions between different hormonal pathways, BR can activate genes encoding enzymes involved in phytoalexin biosynthesis [78,81].
MAPK Signaling Pathway Activation: BR may activate key MAPK enzymes associated with stress responses, thereby influencing the expression of genes involved in phytoalexin production [78,81].
Regulation of Phytoalexin Biosynthetic Enzymes: BR can affect the activity of enzymes responsible for phytoalexin biosynthesis, such as chalcone synthase and stilbene synthase [81].
3.5.2. BR Impact on Plant Defense Response
Pattern Recognition and PTI: BR influences the plant’s first line of defense, known as pattern-triggered immunity (PTI), by modulating gene expression related to defense proteins, cell wall reinforcement, and PTI-associated signaling pathways [65].
Induction of Phytoalexin Production: BR triggers the production of phytoalexins, antimicrobial compounds, through activated signaling pathways, bolstering the plant’s defense against pathogens [22,28,65,102].
Transcription Factors Involvement: BR activates transcription factors like BZR1 and BES1, which regulate gene expression involved in biotic stress responses, including the production of defense proteins [78,81].
Regulation of MAPK Signaling: BR influences MAPK kinase activity, essential for relaying stress signals, thereby regulating the expression of genes related to phytoalexin production and other defense mechanisms [58,81].
Phytoalexin Biosynthetic Enzyme Regulation: BR modulates the activity of enzymes like chalcone synthase and stilbene synthase, boosting phytoalexin production upon pathogen detection [53,55].
Cell Wall Reinforcement: BR affects cell wall structure, impeding pathogen penetration and reinforcing the plant’s defense mechanisms [48,88].
These findings underscore BR’s intricate role in plant defense responses against pathogens. However, further research is needed to fully elucidate the mechanisms underlying BR-mediated defense regulation [4,37,65,88], as highlighted in recent studies such as those by Sirangelo et al. [103], which delve into the molecular intricacies of plant immunity against diverse pathogens.
3.5.3. BR Modulation of Pathogen-Responsive Gene Expression
BR influences gene expression associated with pathogen response, including the induction of phytoalexin production, through interactions with other hormones and signaling pathways [88]. Recent research by Sirangelo et al. [103] sheds light on the molecular mechanisms underlying potential pathogen resistance in Cannabis sativa.
The initial defense response in C. sativa involves recognition of pathogen-associated molecular patterns (PAMP) by plant pattern recognition receptors (PRR), activating PAMP-triggered immunity (PTI) against nonspecific pathogens. Two key molecules, Harpin and Flg22, induce genes like FRK1 and PR1, with Harpin additionally activating ERF1, a transcription factor crucial for ethylene-mediated defense responses. Ethylene response factor 1 (ERF1) plays a pivotal role in regulating gene expression in response to ethylene signals, essential for plant defense mechanisms. Furthermore, receptor-like kinases (RLK) participate in defense against Fusarium, while reactive oxygen species (ROS) and mitogen-activated protein kinase (MAPK) activation contribute to PTI [103].
These findings underscore BR’s comprehensive role in enhancing plant defense against pathogens and biotic stresses, emphasizing the intricate interactions within the signaling network that warrant further investigation for a complete understanding of BR’s mechanisms of action.
BR and Abiotic Stresses
Plants respond differently to temperature changes, with mild to moderate increases leading to thermomorphogenesis, influencing plant development. However, heat stress negatively affects growth and can cause cellular damage. Plants have sophisticated mechanisms to cope with heat stress, including thermosensors, heat shock proteins, and systemic responses, crucial for agricultural adaptation [104,105].
BR Responses to Abiotic Stress: BR aids plants in adapting to various stresses like drought, extreme temperatures, salinity, UV radiation, and heavy metal pollution. It regulates processes such as stomatal aperture control and osmotic compound accumulation, influencing the plant’s defense system. BR’s role in activating the plant defense system is vital, as these hormones play a key role in plant responses to diverse threats and their ability to survive in changing environmental conditions (Figure 4) [22,35].
4.1. Role of BR in Abiotic Stress Responses
Plants rely on receptor-like kinases (RLKs) to perceive stress signals and initiate intracellular responses. While RLKs are well-known for their role in defense responses to biotic stresses, their involvement in abiotic stress responses is less documented. RLKs consist of extracellular, transmembrane, and intracellular kinase domains, enabling them to respond to various stimuli and regulate growth, reproduction, hormone perception, and defense responses [45]. The functions of RLKs in various abiotic stress responses are discussed in Table 1S.
Polyhydroxylated steroidal phytohormones, such as BR, are essential for regulating plant growth, reproduction, and stress responses throughout the plant’s life cycle. BRs impact various physiological processes, including stomatal regulation, photosynthesis, biomass increase, and gene expression stimulation, thereby enhancing stress tolerance [27,56,79,93].
BRs play a crucial role in helping plants combat abiotic stresses induced by factors like drought, temperature extremes, salinity, heavy metals, and air pollution. They influence stomatal regulation, photosynthetic efficiency, antioxidant defense mechanisms, and gene expression, enhancing stress tolerance [28,40,55,56,106,107]. Moreover, BRs interact with other hormones to modulate stress responses, such as increasing ABA content to protect against drought [108].
Temperature fluctuations profoundly affect plant growth and development, with heat stress causing cellular damage and inhibiting photosynthesis. BRs improve thermal tolerance by enhancing antioxidant enzyme levels, mitigating lipid peroxidation, and preventing leaf necrosis and chlorosis under high-temperature conditions [71,109,110,111]. Additionally, BR application in heat-sensitive plants increases photosynthetic pigments, assimilation rates, and water-use efficiency [43].
Cold stress negatively impacts plant growth, but BRs alleviate oxidative damage, maintain photosynthetic efficiency, and activate antioxidant enzymes to enhance cold tolerance. BR-induced cold tolerance involves signaling cascades that regulate gene expression, antioxidant capacity, and cellular redox homeostasis [89].
BRs play a multifaceted role in plant responses to abiotic stresses, influencing various physiological and molecular processes to enhance stress tolerance and improve plant resilience in changing environmental conditions (Figure 5).
Water stress in plants, caused by soil water deficiency, results in osmotic stress, disrupting plant homeostasis and leading to developmental changes like inhibited shoot growth and intensified root growth [66,82,109]. Plant hormones play a crucial role in long-distance communication within plants, with recent research highlighting the significance of hormone movement in the vascular system [9,38,78,103].
Heavy metals, such as chromium, cadmium, nickel, and lead, induce oxidative stress in plants, decreasing photosynthetic efficiency and limiting growth. Excessive accumulation of these metals in plant tissues elevates reactive oxygen species levels, exacerbating oxidative stress [92,112,113,114,115].
Brassinosteroids (BR) alleviate heavy metal stress in plants by promoting phytochelatin synthesis, facilitating metal ion complexation, and enhancing antioxidant enzyme activity to mitigate reactive oxygen species accumulation [116]. Similar benefits are observed in plants exposed to cobalt salts, where BR application enhances antioxidant systems and promotes growth [24]. Additionally, BR application to tomato seedlings growing in cadmium-contaminated medium improves fruiting and fruit antioxidant capacity [117].
Salt stress affects plant growth through ionic and osmotic stress components, leading to inhibited cell growth and stomatal closure. Excessive Na+ ions cause further defense responses, reducing sodium transport and storing ions in roots [31,76,82].
Irfan et al. [68] found that tomato seeds germinated under salt stress and treated with exogenous BR (0.25 and 0.5 ppm via roots and foliar spray) showed improved growth compared to salt-stressed controls. Plants treated with BR exhibited increased plant height, root length, leaf and fruit number, and biomass, mitigating typical salt stress symptoms.
BR, as internal regulators, play a crucial role in plant responses to environmental stresses and developmental processes. They regulate physiological and metabolic processes, acting in a paracrine or autocrine manner. BR biosynthesis genes are widely distributed, and their long-term effects depend on interactions with other hormones [2,20,36,38,118].
BR contribute to plant protection by enhancing physiological processes, such as growth, organogenesis, and adaptation to environmental changes. They interact with other hormones like auxins and gibberellins and are transported intracellularly from synthesis sites to cell membranes and endosomal compartments [38,118]. However, BR transport occurs from their synthesis sites in the endoplasmic reticulum to the cell membrane and early endosomal compartments, where they are received through passive or active intracellular transport. Here are several ways in which BR can contribute to ensuring plant protection against environmental stresses:
- −
- Growth stimulation: BR promotes plant growth and aids in faster recovery from stress-induced damage by increasing cell length and numbers [38].
- −
- −
- −
- −
- −
4.1.1. Leaf Rust Kinase (LRK 10-like)
The LRK10 protein, found in wheat (TaLRK10), and its homolog AtLRK10L1 in Arabidopsis are associated with resistance to leaf rust caused by Puccinia recondita. AtLRK10L1.2, a transcript of AtLRK10L1, plays a crucial role in regulating flowering time, pathogen defense, and ABA signaling during seed germination, especially in drought responses by aiding stomatal closure [25,100,119,120]. Ongoing research investigates LRK10’s domains in response to various stresses [25,100,120,121]. BR are pivotal in enhancing plant resilience to environmental stresses by regulating growth, adaptive mechanisms, and damage prevention [65,79,100,112,120], influencing developmental processes and gene expression [23,28,43]. See Figure 6 for a schematic diagram illustrating BR’s key roles in plant life.
BR plays a crucial role in plant responses to abiotic stresses by:
- −
- −
- −
- −
- Reducing oxidative stress by influencing antioxidant enzyme activity to neutralize reactive oxygen species [38].
- −
- −
Overall, BR plays a pivotal role in regulating plant responses to diverse abiotic stresses, aiding plants in survival and adaptation to challenging environmental conditions.
4.2. BR and Plant Immune Activation through Antimicrobial Production
BR influences the production of antimicrobial substances and activates the plant immune system through various mechanisms:
- −
- Stimulation of Phytohormone Production: BR stimulates the production of phytohormones like JA and SA, crucial for activating the plant immune system against pests and pathogens [124].
- −
- Regulation of Defensive Gene Expression: BR regulates the expression of genes encoding defensive proteins, including Pathogenesis-Related (PR) proteins and Resistance (R) proteins, aiding in combating pathogens [90].
- −
- −
- Regulation of Plant-Pathogen Interactions: BR influences interactions between plants and pathogens, affecting pathogen recognition mechanisms and intercellular communication [125].
- −
- −
BR’s influence on antimicrobial substance production and plant immune activation varies based on factors like pathogen type, plant species, and ecological context. Despite ongoing research, the precise molecular mechanisms underlying plant tolerance to abiotic stresses remain incompletely understood. Understanding these mechanisms is crucial for addressing global food production challenges caused by various stressors like drought, temperature fluctuations, salinity, and more, impacting plant growth, physiology, and photosynthesis [63,113,124].
4.3. Role of BR in Biotic Stresses
BR plays a crucial role in regulating defense processes against pathogens, including the production of antimicrobial compounds like phytoalexins and the activation of the plant’s immune response. Manipulating BR levels can impact a plant’s ability to combat pathogens, with excessive production potentially reducing resistance to diseases and deficiencies increasing susceptibility to attacks [21,33,42,115,127].
BR significantly contributes to plant defense against various pathogens and pests. It influences plant responses to biotic stresses through several mechanisms:
- Enhanced Pathogen Resistance: BR stimulates the production of phytoalexins, strengthening the plant’s defense system against infections [127].
- Reinforcement of Immunity: BR stimulates the production of chemical substances with anti-parasitic and antibacterial properties, enhancing overall plant immunity.
- Regulation of Defensive Cell Development: BR influences the development of defense cells like trichomes, aiding in defense against pests and pathogens [36].
Overall, BR’s role in plant defense against pathogens and pests is crucial, although its effects can vary depending on factors such as the type of pathogen, plant species, and ecological context.
Interactions Between Plant Hormones in Immunity
Plant responses to stress involve intricate interactions between hormones and signaling pathways, particularly those involving jasmonic acid (JA) and salicylic acid (SA) production. These pathways regulate various stages, including transcription, protein-protein interaction, and targeted protein degradation [36]. Additionally, numerous phytohormones influence plant growth and development, with brassinosteroids (BR) notably enhancing resistance to biotic stresses. BR’s impact involves regulating protein and nucleic acid production, influencing metabolic processes like photosynthesis, and modulating enzymatic reactions crucial for stress resilience [30,33,42,92]. Through complex biochemical reactions, BR acts as immunomodulators, strengthening plant ability to combat pathogens by regulating enzymatic reactions, protein production, and gene expression [33,42]. Examples include their cross-talk with JA and SA, modulation of enzymatic reactions, induction of defense-related protein production, and modification of gene expression. In summary, BR orchestrates interactions between plant hormones, ensuring a coordinated response to environmental stresses and bolstering plant immunity against pathogens [102,124].
5.1. Abscisic Acid and Salicylic Acid
Interactions between ABA and SA are complex and context-dependent. In some cases, ABA may suppress defense responses triggered by SA, while in others, it may act synergistically or independently. JA is an important hormone in defense against pests and plant pathogens, such as feeding insects and fungi. SA is often involved in defense against pathogens, especially in infections by viruses, fungi, and bacteria. Interactions between JA and SA are complex and depend on the type of pathogen and plant. Antagonism often occurs between these hormones, meaning that an increase in one may inhibit responses induced by the other hormone [124,127].
5.2. Interaction of ABA and JA
Interactions between BR and other plant hormones, ABA, and JA, play a significant role in regulating various growth and development processes in response to plant stresses. Key interactions of BR-ABA include:
- −
- −
- −
- Gene Expression Regulation: Both BR and ABA influence gene expression in response to various stresses. In drought conditions, BR and ABA may regulate different sets of genes related to their absence. During drought, ABA levels increase, leading to the activation of signaling pathways controlling the expression of genes responsible for adapting to water deficiency. These genes may be associated with stomatal closure, activation of genes involved in osmotic compound synthesis, or maintaining water homeostasis. On the other hand, BR may be involved in regulating genes related to long-term stress adaptation, such as controlling the expression of genes associated with processes like stomatal closure, root growth, or activation of signaling pathways related to plant defense against stress, while ABA focuses on faster adaptive responses [69,81,84].
In summary, the interplay between BR and ABA involves complex synergistic and antagonistic interactions, influencing various aspects of plant growth and stress responses. These interactions are context-dependent, and their understanding is crucial for unraveling the intricacies of plant hormone signaling.
5.3. BR-JA Interactions
BR and JA often act antagonistically in regulating various processes. For instance, BR may stimulate shoot and flower growth, while JA is known for its role in defense against pests and immune responses against pathogens. In some cases, acting antagonistically, BR may suppress JA activity, and vice versa, helping plants adapt to changing conditions. Hayat et al. demonstrated the influence of rootstocks on scion vigor and hormonal mechanisms related to dwarfing, such as IAA, CK, ABA, GA, and BR. Both BR and ABA play a crucial role in regulating gene expression in response to drought, adapting plants to challenging environmental conditions. Together and individually, they influence various aspects of responses to water deficiency, adjusting plant growth, development, and defense reactions [69,81,102].
The interaction of BR-JA may be crucial in plant defense against pathogens and pests. JA is often involved in defensive responses against these threats, while BR may assist the plant in maintaining growth and development in the face of attacks, simultaneously stimulating defense by regulating the expression of defense genes [69,84,102,124].
Both BR and JA can influence the production of phytoalexins, chemical substances with antibacterial and antiparasitic properties, crucial components of plant defense against pathogens [69,84,102].
Interactions between BR, ABA, and JA are context-dependent and may vary depending on the plant variety and species, the type of stress, and the plant’s developmental stage. These plant hormones collaborate to provide a response to changing environmental conditions, balancing growth, development, and defense as needed. BR can interact with both ABA and JA, influencing plant growth and development processes as well as the regulation of defense responses [36,81,84].
Interactions between BR and other plant hormones are complex and context-dependent. They can act synergistically, antagonistically, or independently concerning other hormones, depending on the type of stress and the plant. For instance, in the case of CK and GA, interactions between CK and GA are essential for regulating plant growth and development processes, impacting the plant’s ability to counteract biotic and abiotic stresses through changes in plant architecture. Interactions between plant hormones are dynamic and depend on many factors [69,78,81,93,102]. Plants adjust their defense responses based on the type of threat and hormonal signals they receive. Therefore, researchers continue to delve into these complex mechanisms to better understand how plants regulate their immunity and growth in changing environmental conditions.
6. Practical Applications of BR
BR enhances plant resistance to both biotic and abiotic stresses, such as drought, extreme temperatures, salinity, oxidative stress, toxic heavy metals, UV radiation, and attacks by pathogens. Their ability to defend against stress and regulate development is associated with the regulation of specific proteins and nucleic acids, as well as their influence on metabolic processes related to photosynthesis [21,30,33,42,92].
6.1. Potential Applications of BR in Agriculture and Horticulture
BR has several practical applications in agriculture and plant protection, although its use is relatively new and still under investigation [130,131,132]. The potential applications of BR in these fields are illustrated in Figure 7.
Enhancing Crop Yield and Quality: BR stimulates plant growth and development, potentially increasing crop yield and improving characteristics like size, shape, texture, taste, and color of fruits or flowers [68,128,132,133].
Protection Against Abiotic Stresses: BR aids plants in coping with extreme environmental conditions like drought, low temperatures, and soil salinity by enhancing plant tolerance to these stresses. They regulate stomatal opening and closing, controlling transpiration and water loss [44,93,97].
Protection Against Pathogens and Pests: BR stimulates the plant’s immune system, aiding defense against fungal, bacterial, and viral infections. They influence the production of antimicrobial substances and activate defense reactions against pests, affecting plant development and reducing pest and disease damage [69,130].
Root System Stimulation: BR impacts root development, increasing the plant’s ability to absorb water and nutrients, especially under abiotic stress conditions [53].
Acceleration of Maturation and Fruiting Processes: BR can accelerate fruit maturation, benefiting agricultural and horticultural production and fruit availability in the market [60,86].
Mitigation of Oxidative Stress Effects: BR helps plants protect against oxidative stress by neutralizing reactive oxygen species and preventing cell damage [24,118].
Applications in Plant Breeding and Biotechnology: BR can be used in breeding programs to obtain varieties more resistant to stresses and in genetic engineering to modify plants for improved tolerance to environmental conditions. Studies on Arabidopsis mutants affecting the BR biosynthesis pathway have contributed to understanding BR’s role in regulating plant growth and the role of BR receptors in this signaling pathway [20,21,134].
While BR shows potential for improving plant productivity and resilience, further research optimizing doses, application timing, and considering ecological and environmental safety is necessary for effective application in agriculture, horticulture, and plant protection.
6.2. Significance of BR in Cell Division and Elongation
BRs participate in the regulation of plant growth by influencing two cell processes: cell expansion and division [9,28]. They impact the elongation of hypocotyls, epicotyls, and stems in dicot plants, as well as coleoptiles and mesocotyls in monocot plants [26,135]. These hormones play a role in cell enlargement, involving gene expression and enzymatic activity [78]. In Arabidopsis, leaves emerge on the sides of the shoot apical meristem through cell division, followed by a gradual maturation characterized by the completion of the cell cycle and cell expansion [59]. Genetic, hormonal, and environmental factors determine leaf growth and development [96]. Das et al. [131] demonstrated the bene-ficial impact of BR on cell elongation and division in studies on the second internode of beans. Stimulating effects of BR, auxin, and cyto-kinin on cell division in bean parenchyma cells were also shown by Bariş and Sağlam-Çağ [25], Li et al. [136], as well as in cabbage protoplasts by Cimino et al. [137] and petunia by Cui et al. [138].
6.2.1. BR in Seed Development and Germination
Seed germination in plants like Arabidopsis thaliana and Nicotiana tabacum relies on a delicate balance between the growth potential of the embryo and the mechanical constraints imposed by the micropylar endosperm [129,139]. BR application has been shown to promote seed germination and counteract the dormancy-inducing effects of abscisic acid (ABA), particularly in stressful conditions [64].
Chakma et al. [64] studied cotton seed germination under various hormone treatments and found that BR facilitated germination even under stress conditions like heat and salinity. It also promoted cotyledon opening and lateral root development. Similarly, research on rice seeds by Xiong et al. [13] demonstrated that both GA and BR enhanced germination and post-germination growth, with GA compensating for BR deficiency or insensitivity. Baset et al. [63] observed increased germination parameters in rice with BR treatment, despite the presence of toxic aluminum.
Thongsri et al. [140] investigated the effects of BR and GA3 on soybean seed germination at low temperatures, where BR and GA3 application helped overcome delayed germination due to cold stress. Additionally, Hayat and Ahmad [141] showed positive effects of 28-homobrassinolide (HBR) treatment on wheat seed germination and enzyme activity.
In another study, Kumar et al. [133] examined mung bean response to zinc stress and BR treatment, noting significant improvements in germination percentage and seedling growth with BR application, albeit with reduced antioxidant enzyme activity.
Finally, Kiliç et al. [98] found that 24-epibrassinolide (EBR) positively impacted barley seed germination and root elongation under saline stress conditions.
In summary, BR application shows promise in enhancing seed germination and seedling growth, particularly under adverse environmental conditions. However, further research is needed to optimize its application for maximum benefit while considering potential trade-offs in antioxidant activity.
6.2.2. BR in Root and Shoot Growth
The impact of BR on root and shoot growth is influenced by factors such as concentration, treatment duration, plant stage, and application method. Lower concentrations of BR typically stimulate root growth, while higher concentrations can inhibit it [7].
In a study on Eucalyptus grandis Hill, Chinese researchers investigated the effects of arbuscular mycorrhizal fungi symbiosis and BR application on root growth. They found that increasing concentrations of exogenous 24-epibrassinolide initially increased the mass and length of fresh taproots but eventually led to a decrease in growth [142].
Liu et al. [57] studied the effects of homobrassinolide (HBR) on barley root growth and observed enhanced mitotic activity and significant root tip enlargement compared to the control group.
Meanwhile, research by Nováková et al. [143] focused on the application of BR and its effects on Scots pine seedlings. They noted that BR treatment slowed down both the height and radial growth of the seedlings. Specifically, the height increment of BR-treated seedlings was 11% lower, and the diameter increment at the root collar was 13% lower compared to the control. Additionally, BR-treated pine trees exhibited higher mortality rates compared to the control group.
Overall, while BR can have varying effects on root and shoot growth depending on concentration and application method, it is crucial to consider its impact on overall plant health and survival when applying it in agricultural or horticultural settings.
6.2.3. BR Effects on Plant Growth, Photosynthesis, and Aging
The influence of BR on root and shoot growth is concentration-dependent, with lower concentrations stimulating root growth and higher concentrations inhibiting it [7]. Studies on Eucalyptus grandis Hill roots showed increased mass and length with higher exogenous BR concentration [142]. Homobrassinolide (HBR) positively affected barley root growth, demonstrating higher mitotic activity compared to the control [57]. However, BR application in Scots pine seedlings slowed down height and radial growth, with increased mortality compared to the control [143].
BR plays a crucial role in maintaining photosystem II (PSII) performance, especially under stress conditions. Exogenous BR application in rice under heat stress helped maintain net photosynthetic rates and improved water use efficiency [128]. Similar positive effects on rice photosynthesis were observed with the application of HBL under cold stress [72].
BL, including 24-epibrassinolide (EBR), proved effective in alleviating growth inhibition in melon under high-temperature conditions [43]. Cucumber seedlings treated with 28-homobrassinolide showed higher antioxidant enzyme activity and proline content under chilling stress [72].
The impact of BR analogs on leaf aging varied among plants. In papaya, BR maintained the greenness of older leaves, while in soybean, aging was accelerated after epibrassinolide (EBL) application [25,61,144,145,146]. Sunflower leaf aging, influenced by EBL, showed differences between upper and lower leaves, possibly linked to auxin distribution [147].
In summary, BR plays a multifaceted role in regulating plant growth, photosynthesis, and aging under various environmental conditions.
6.3. BR under Salinity and Heavy Metal Concentrations
Soil salinity poses a significant challenge to plant growth, often leading to toxic metal ion accumulation and nutrient imbalances. Studies have shown that the application of brassinosteroids (BRs) can mitigate the negative effects of salinity stress on plants.
In research conducted by Pakistani scientists, Lycopersicon esculentum (tomato) plants treated with NaCl experienced decreased growth. However, the application of BR enhanced various plant parameters, both in stressed and non-stressed soil conditions [68].
Optimizing mineral nutrient levels and the application of plant growth regulators are crucial strategies for enhancing crop yield under stress conditions. Abbas et al. [17] suggested potassium and jasmonic acid supplementation for crops irrigated with water containing heavy metals, providing a potential solution to mitigate the adverse effects of heavy metal stress.
Rice plants subjected to salt stress (100 and 200 mM NaCl) showed improved germination, growth, and photosynthesis when treated with BR. Additionally, BR treatment enhanced the activities of antioxidant enzymes, helping to counteract the oxidative stress induced by salinity [100].
Pea seeds soaked in 24-epibrassinolide (EBL) along with sodium chloride (NaCl) exhibited increased germination rates and growth parameters, as well as enhanced photosynthesis and antioxidant activities, indicating the protective role of BR against salt stress in pea plants [122].
In rapeseed plants subjected to salt stress, exogenous application of 24-EBL mitigated the negative effects of salinity, resulting in improved growth, photosynthesis, and nutrient content in both roots and shoots [31]. Additionally, research by Efimova et al. [148] confirmed the protective action of EBL on rapeseed, highlighting its role in restoring growth and preventing lipid peroxidation. Furthermore, investigations by Houimli et al. [149] explored the role of 24-EBL in enhancing salt tolerance in pepper plants. Their findings indicated improved growth and maintenance of cell membrane integrity in pepper plants treated with BR under salt stress conditions [149].
A mechanism depicting the involvement of 24-epibrassinolide (EBR) application in conferring salt stress tolerance in perennial ryegrass can be outlined as follows [150] (Figure 8):
- −
- Activation of Stress Signaling Pathways: Upon exposure to salt stress, perennial ryegrass plants perceive the stress signals, leading to the activation of stress signaling pathways.
- −
- Recognition and Uptake of EBR: Exogenous application of EBR to the plants allows for its recognition and uptake, either through the roots or foliar application.
- −
- Brassinosteroid Signaling Cascade Activation: EBR binds to its receptor, initiating a signaling cascade that involves various downstream components, such as BRASSINAZOLE-RESISTANT (BZR) transcription factors and BRI1-EMS-SUPPRESSOR 1 (BES1), leading to the activation of stress-responsive genes.
- −
- Enhanced Ion Homeostasis: EBR application modulates ion homeostasis by regulating the uptake and compartmentalization of ions, particularly sodium (Na+) and chloride (Cl-), thus preventing their toxic accumulation in plant tissues.
- −
- Osmotic Adjustment: EBR promotes osmotic adjustment by enhancing the accumulation of compatible solutes such as proline and sugars, which help maintain cellular turgor pressure and osmotic balance under salt stress conditions.
- −
- Antioxidant Defense Activation: EBR treatment boosts the activity of antioxidant enzymes such as superoxide dismutase (SOD), catalase (CAT), and peroxidase (POD), scavenging reactive oxygen species (ROS) and minimizing oxidative damage to cellular components.
- −
- Maintenance of Photosynthetic Machinery: EBR application preserves the integrity and functionality of the photosynthetic apparatus by mitigating the adverse effects of salt stress on chlorophyll content, photosystem efficiency, and carbon assimilation rates.
- −
- Stimulation of Growth and Development: Despite salt stress, EBR promotes plant growth and development by modulating hormone signaling pathways, particularly auxins, cytokinin’s, and gibberellins, which contribute to enhanced root and shoot growth and biomass accumulation.
- −
- Regulation of Stress-Responsive Gene Expression: EBR-mediated regulation of stress-responsive genes involved in various physiological and biochemical processes helps orchestrate adaptive responses to salt stress, thereby improving overall stress tolerance in perennial ryegrass.
- −
- Overall Improvement in Salt Stress Tolerance: Through these mechanisms, EBR application confers enhanced salt stress tolerance in perennial ryegrass, enabling the plant to withstand adverse environmental conditions and maintain optimal growth and productivity [150].
- −
- As a result, exogenous 24-epibrassinolide (EBR) may: improve the integrity and stability of the cell membrane (lower MDA, EL, higher proline), enhance photosynthetic function (higher Pn, gs, and chlorophyll), enhance antioxidant defense (resulting in higher activity of SOD, CAT, and APX), increase hormonal metabolism (higher levels of IAA, ZR, iPA, JA, SA, and ABA), and reduce ion toxicity (lower Na+ and Na+/K+, higher K+, Mg2+, and Ca2+), leading to increased tolerance to salt stress and visual quality of plants [151] (Figure 8).
- −
- These studies collectively demonstrate the potential of BRs to alleviate the adverse effects of salinity and heavy metal stress on plants, highlighting their significance in enhancing plant resilience to environmental challenges.
6.4. Enhancing Crop Yield and Resilience with BR: Potential Strategies and Benefits
BR manipulation holds promise for enhancing crop yield and resilience against various stresses in agriculture. Key strategies and potential benefits include:
- −
- Growth Promotion: Elevating BR levels stimulates the growth of stems, leaves, and roots, potentially leading to increased yield mass [51].
- −
- −
- −
- −
- −
It’s important to acknowledge that manipulating BR levels is nuanced and context-dependent, requiring precise control of doses and application timing. Environmental and safety considerations are paramount when using these substances in agriculture. Ongoing research aims to fully grasp the potential benefits and complexities of BR manipulation for agriculture and plant protection [149,150,151].
7. Towards the Future
In the face of climate change challenges, leveraging BR offers promising strategies to improve plant tolerance to abiotic stresses. Key approaches include:
- −
- Advanced Research Tools: Precision translatomics and fluorescence-activated cell sorting (FACS) enable a deeper understanding of how plants respond to BR under stress, facilitating the mapping of BR interactions with other hormones.
- −
- Exogenous BR Application: Utilizing BR application can boost plant resilience, particularly in cereal crops. Genetic manipulation of BR-related genes can pave the way for developing transgenic varieties with heightened stress resistance.
- −
- Exploring Hormone Interactions: Investigating how exogenous BR impacts the synthesis and signaling of other hormones under stress conditions is crucial. Combining different plant hormones may offer synergistic strategies to enhance stress tolerance.
- −
- Genetic Engineering: Manipulating BR-related genes through genetic engineering, coupled with advanced research tools, holds promise for increasing crop yields by bolstering tolerance to abiotic stresses.
While these strategies require further research, their potential to address global food demand by bolstering plant resilience underscores the importance of BR in agriculture.
8. Conclusions
Understanding the complex interactions between BR and other plant hormones is crucial for harnessing their potential in agriculture.
Manipulating BR levels shows promising results in increasing yields, enhancing resistance to abiotic stresses, and defending against pathogens and pests. Advanced research tools, such as precision translatomics and genetic engineering, offer opportunities for deeper understanding of BR mechanisms and their applications in agriculture.
Further research and experiments are necessary to optimize strategies for manipulating BR levels and ensure their effectiveness across different plant species and environmental conditions.
Despite challenges, the potential of utilizing BR in agriculture is significant and offers promising solutions that may contribute to addressing global food security issues in the face of climate change.
BR plays a crucial role as plant hormones influencing various aspects of plant development and adaptation to changing environmental conditions. Conscious manipulation of BR levels can significantly impact crop productivity and their resilience to stresses and pathogens.
Understanding these mechanisms can contribute to the development of more effective strategies to improve the resilience of cultivated plants to various environmental challenges.
List abberavions
| ABA – Abscisic Acid BL – Brassinolide BR – Brassinosteroid BZR – Brasinozol CAT – Catalase CH4 - methane CK – Cytokinins CS – Castasterone EBL – benzylaminopurine EBL – Epibrassinolide EBR – 24-epibrassinolide ERF – Ethylene Response Factor FRK1 - Fusarium-Induced Receptor Kinase GA – Gibberelins HBR – Homobrassinolide IAA – indolyl-3-acetic acid IAN – indolylacetonitrile IBA – Indolyl-3-butyric acid IBA – indolyl-3-butyric acid IPA – indolyl-3-propionic acid JA – jasmonate MAPK – Mitogen-Activated Protein Kinase MRW - indole-3-butyric acid NiRA – Nitrate Reductase PAA – phenylacetic acid, PAMPs – Pathogen-associated molecular proteins POD – Peroxidase PR - Pathogenesis-Related Genes PRRs – Pattern Recognition Receptors, PSII – photosystem II PTI – Pattern Triggered Immunity RLK – Receptor-Like Kinases ROS – Production of reactive oxygen species ROS – Reactive Oxygen Species SA – salicylic acid SERK – Somatic Embryogenesis Receptor-Like Kinase SOD – Superoxide Dismutase SQE – Squalene Monooxygenase Enzyme TZ – Tezasterone WRKY46, WRKY54, WRKY70 – regulators of proteins from the WRKY family activated by BR |
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1: title: Functions of RLKs in different abiotic stress responses.
Author Contributions
For research articles with several authors, a short paragraph specifying their individual contributions must be provided. The following statements should be used “Conceptualization: B.S., B.B., and P.B.; methodology: B.B., B.S., P.P.; software: D.S., P.P.; validation: D.S.; formal analysis: P.B., B. K-M., investigation: P.P.; resources, B.B., P.B., B. K-M.; data curation: D.S., P.P.; B.K-M.; writing—original draft preparation: B.B., B.S., P.B., D.S.; writing—review and editing, B.S., P.P., visualization: D.S., supervision, B.S.; B.K-M.; project administration: P.B.; funding acquisition: B.S. All authors have read and agreed to the published version of the manuscript.”.
Funding
“This research received no external funding”.
Institutional Review Board Statement
“Not applicable”.
Informed Consent Statement
“Not applicable.”.
Data Availability Statement
Data sharing not applicable. No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Acknowledgments
We would like to thank the University of Life Sciences in Lublin for administrative and technical support.
Conflicts of Interest
“The authors declare no conflict of interest.”.
Data Availability Statement
no new test results were generated.
References
- Liu, X.; Igarashi, D.; Hillmer, R.A.; Stoddard, H.; Lu, Y.; Tsuda, K.; Myers, C.L.; Katagiri, F. Dynamic decomposition of transcriptome responses during plant effector-triggered immunity revealed conserved responses in two distinct cell populations. Preprint, 2022, Chrome.extension://efaidnbmnnnibpcajpcglclefindmkaj/https://www.biorxiv.org/content/10.1101/2022.12.30.522333v1.full.pdf.
- Hewedy, O.A.; Elsheery, N.I.; Karkour, A.l.M.; Elhamouly, N.; Arafa, R.A.; Mahmoud, G.A.-E.; et al. Jasmonic acid regulates plant development and orchestrates stress response during tough times. Environmental and Experimental Botany 2023, 208, 105260. [Google Scholar] [CrossRef]
- Nolan, T.M.; Vukašinović, N.; Liu, D.; Russinova, E.; Yin, Y. Brassinosteroids: multidimensional regulators of plant growth, development, and stress responses. The Plant Cell 2020, 32, 295–318. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Su, P.; Meng, X.; Liu, P. Phylogeny of the plant receptor-like kinase (RLK) gene family and expression analysis of wheat RLK genes in response to biotic and abiotic stresses. BMC Genomics 2023, 24, 224. [Google Scholar] [CrossRef] [PubMed]
- Yan, M.Y.; Xie, D.L.; Cao, J.J.; Xia, X.J.; Shi, K.; Zhou, Y.H.; Zhou, J.; Foyer, C.H.; Yu, J.Q. Brassinosteroid-mediated reactive oxygen species are essential for tapetum degradation and pollen fertility in tomato. The Plant Journal 2020, 102, 931–947. [Google Scholar] [CrossRef]
- Li, Z.; He, Y. Roles of Brassinosteroids in Plant Reproduction. Int. J. Mol. Sci. 2020, 21, 872. [Google Scholar] [CrossRef]
- Kanwar, M.K.; Bajguz, A.; Zhou, J.; Bhardwaj, R. Analysis of brassinosteroids in plants. Journal of Plant Growth Regulation 2017, 36, 1002–1030. [Google Scholar] [CrossRef]
- Siddiqui, H.; Hayat, S.; Bajguz, A. Regulation of photosynthesis by brassinosteroids in plants. Acta Physiologiae Plantarum 2018, 40, 1–15. [Google Scholar] [CrossRef]
- Oh, M.H.; Honey, S.H.; Tax, F.E. The control of cell expansion, cell division, and vascular development by brassinosteroids: a historical perspective. International Journal of Molecular Sciences 2020, 21, 1743. [Google Scholar] [CrossRef]
- Oh, E.; Zhu, J.Y.; Bai, M.Y.; Arenhart, R.A.; Sun, Y.; Wang, Z.Y. Cell elongation is regulated through a central circuit of interacting transcription factors in the Arabidopsis hypocotyl. Life 2014, 3, e03031. [Google Scholar] [CrossRef]
- Wei, Z.; Li, J. Brassinosteroids regulate root growth, development, and symbiosis. Mol. Plant 2016, 9, 86–100. [Google Scholar] [CrossRef]
- Li, J.G.; Fan, M.; Hua, W.; Tian, Y.; Chen, L.G.; Sun, Y.; Bai, M.Y. Brassinosteroid and hydrogen peroxide interdependently induce stomatal opening by promoting guard cell starch degradation. The Plant Cell 2020, 32, 984–999. [Google Scholar] [CrossRef]
- Xiong, M.; Feng, G.N.; Gao, Q.; Zhang, C.Q.; Li, Q.F.; Liu, Q.Q. Brassinosteroid regulation in rice seed biology. Seed Biology 2022, 1, 1–9. [Google Scholar] [CrossRef]
- Yamamoto, R.; Fujioka, S.; Iwamoto, K.; Demura, T.; Takatsuto, S.; Yoshida, S.; Fukuda, H. Co-regulation of brassinosteroid biosynthesis-related genes during xylem cell differentiation. Plant and Cell Physiology 2007, 48, 74–83. [Google Scholar] [CrossRef]
- Wang, Z.Y.; Bai, M.Y.; Oh, E.; Zhu, J.Y. Brassinosteroid signaling network and regulation of photomorphogenesis. Annual Review of Genetics 2012, 46, 701–724. [Google Scholar] [CrossRef]
- Kovtun, I.S.; Kukharenko, N.E.; Kusnetsov, V.V.; Khripach, V.A.; Efimova, M.V. Effect of lactone-and ketone-containing brassinosteroids on photosynthetic activity of barley leaves during aging. Russian Journal of Plant Physiology 2021, 68, 440–450. [Google Scholar] [CrossRef]
- Abbas, T.; Fan, R.; Hussain, S.; Sattar, A.; Khalid, S.; Butt, M.; Shahzad, U.; Atif, H.M.; Batool, M.; Ullah, S.; Li, Y.; Al-Hashimi, A.; Elshikh, M.S.; Al-Yahyai, R. Protective effect of jasmonic acid and potassium against cadmium stress in peas (Pisum sativum L.). Saudi Journal of Biological Sciences 2022, 29, 2626–2633. [Google Scholar] [CrossRef]
- Abo-Ogiala, A., Carsjens, C., Diekmann, H., Fayyaz, P., Herrfurth, C., Feussner, I., Polle, A. Temperatureinduced lipocalin (TIL) is translocated under salt stress and protects chloroplasts from ion toxicity, Journal of Plant Physiology 2014, 171, 250–259, ISSN 0176-1617. [CrossRef]
- Aghaee, P.; Rahmani, F. Seed priming with 24-epibrassinolide alters growth and phenylpropanoid pathway in flax in response to water deficyt. J. Agric. Sci. Technol. 2020, 22, 1039–1052. [Google Scholar]
- Ahammed, G.J.; Yu, J.Q. Role of Hormones in Plant Adaptation to Heat Stress. In: Plant Hormones under Challenging Environmental Factors, Springer: Dordrecht, The Netherlands, 2016; 1–21.
- Ahammed, G.J.; Li, X.; Liu, A.; Chen, S. Brassinosteroids in plant tolerance to abiotic stress. Journal of Plant Growth Regulation 2020, 39, 1451–1464. [Google Scholar] [CrossRef]
- Bajguz, A. Brassinosteroids–occurence and chemical structures in plants. In Brassinosteroids: A Class of Plant Hormone; Springer: Berlin/Heidelberg, Germany, 2011; pp. 1–27. [Google Scholar]
- Alzahrani, S.M.; Alaraidh, I.A.; Migdadi, H.; Alghamdi, S.; Khan, M.A.; Ahmad, P. Physiological, biochemical, and antioxidant properties of two genotypes of Vicia faba grown under salinity stress. Pakistan Journal of Botany 2019, 51, 786–798. [Google Scholar] [CrossRef] [PubMed]
- Arora, P.; Bhardwaj, R.; Kanwar, M.K. Effect of 24-epibrassinolide on growth, protein content and antioxidative defense system of Brassica juncea L. subjected to cobalt ion toxicity. Acta Physiol. Plant. 2012, 34, 2007–2017. [Google Scholar] [CrossRef]
- Bariş, Ç.Ç.; Sağlam-Çağ, S. The effects of brassinosteroids on sequential leaf senescence occurring in Glycine max L. Int J Biotechnol. Res. 2016, 6, 7–16. [Google Scholar]
- Bhanu, A.N. Bhanu, A.N. Brassinosteroids: Relevance in biological activities of plants and agriculture. J. Plant Sci. Res 2019, 35, 1–15. [Google Scholar] [CrossRef]
- Chaudhuri, A.; Halder, K.; Abdin, M.Z.; Majee, M.; Datta, A. Abiotic Stress Tolerance in Plants: Brassinosteroids Navigate Competently, Int. J. Mol. Sci. 2022, 23, 14577. [Google Scholar] [CrossRef] [PubMed]
- Kour, J.; Kohli, S.K.; Khannika, K.; Bakshi, P.; Sharma, P.; Singh, A.D.; Ibrahim, M.; Devi, K.; Sharma, N.; Ohri, P.; Skalicky, M.; Brestic, M.; Bhardwaj, R.; Landi, M.; Sharma, A. Brassinosteroid Signaling, Crosstalk and, Physiological Functions in Plants Under Heavy Metal Stress, Frontiers in Plant Science 2021, 12. [CrossRef]
- Tang, J.; Han, Z.; Chai, J. Q&A: what are brassinosteroids and how do they act in plants? BMC Biol 2016, 14, 113. [Google Scholar] [CrossRef]
- Aitken, V.; Diaz, K.; Soto, M.; Olea, A.F.; Cuellar, M.A.; Nuñez, M.; Espinoza-Catalán, L. New Brassinosteroid Analogs with 23,24-Dinorcholan Side Chain, and Benzoate Function at C-22: Synthesis, Assessment of Bioactivity on Plant Growth, and Molecular Docking Study. International Journal of Molecular Sciences 2024, 25, 419. [Google Scholar] [CrossRef]
- Mehmood, S.; Siddiqi, E.H.; Nawaz, I.; Nasir, N. 24-Epibrassinolide modulates biomass production, gas exchange characteristics and inorganic nutrients in canola (Brassica napus L.) under salt stress. Pak. J. Bot 2022, 54, 1199–1209. [Google Scholar] [CrossRef]
- Yokota, T.; Ohnishi, T.; Shibata, K.; Asahina, M.; Nomura, T.; Fujita, T.; Ishizaki, K.; Kohchi, T. Occurrence of brassinosteroids in non-flowering land plants, liverwort, moss, lycophyte and fern. Phytochemistry 2017, 136, 46–55. [Google Scholar] [CrossRef]
- Hussain, M.A.; Fahad, S.; Sharif, R.; Jan, M.F.; Mujtaba, M.; Ali, Q.; Ahmad, A.; Ahmad, H.; Amin, N.; Ajayo, B.S.; sun, C.; Gu, L.; Ahmad, I.; Jiang, Z.; Hou, J. Multifunctional role of brassinosteroid and its analogues in plants. Plant Growth Regulation 2020, 92, 141–156. [Google Scholar] [CrossRef]
- Nafie, E. Physiological, Biochemical, Molecular and Hormonal Studies to Confirm Growth and Development Regulating Actions of Brassinosteroids in Phaseolus vulgaris L. cv. Bronco Seedlings. Life Science Journal 2014, 11, 974–991. [Google Scholar]
- Bajguz, A. The effect of brassinosteroids on cultures of the alga Chlorella vulgaris exposed to selected phytohormones and stress factors. PhD Thesis, University in Białystok, Publishing House of the University of Białystok, 2007. [Google Scholar]
- Bajguz, A.; Hayat, S. Effects of brassinosteroids on the plant responses to environmental stresses. Plant Physiology and Biochemistry 2009, 47, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Yuan, Y.; Du, J.; Sun, J.; Guo, S. Effects of 24-Epibrassinolide on Nitrogen Metabolism in Cucumber Seedlings under Ca[NO3]2 Stress. Plant Physiol. Biochem. 2012, 61, 29–35. [Google Scholar] [CrossRef]
- Lacombe, B.; Achard, B. Long-distance transport of phytohormones through the plant vascular system. Curr. Opin. Plant Biol. 2016, 34, 1–8. [Google Scholar]
- Gawande, N.D.; Sankaranarayanan, S. Genome wide characterization and expression analysis of CrRLK1L gene family in wheat unravels their roles in development and stress-specific responses. BioRxiv 2023. [Google Scholar] [CrossRef]
- Ibañez, C.; Delker, C.; Martinez, C.; Bürstenbinder, K.; Janitza, P.; Lippmann, R.; Ludwig, W.; Sun, H.; James, G.V.; Klecker, M. Brassinosteroids dominate hormonal regulation of plant thermo morphogenesis via BZR1. Curr. Biol. 2018, 28, 303–310. [Google Scholar] [CrossRef]
- Symons, G.M.; Ross, J.J.; Jager, C.E.; Reid, J.B. Brassinosteroid transport. J Exp Bot. 2008; 59(1):17-24. 59(1). [CrossRef] [PubMed]
- Manghwar, H.; Hussain, A.; Ali, Q.; Liu, F. Brassinosteroids (BRs) role in plant development and coping with different stresses. International Journal of Molecular Sciences 2022, 23, 1012. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.P.; Zhu, X.H.; Ding, H.D.; Yang, S.J.; Chen, Y.Y. Foliar Application of 24-Epibrassinolide Alleviates High-Temperature-Induced Inhibition of Photosynthesis in Seedlings of Two Melon Cultivars. Photosynthetica 2013, 51, 341–349. [Google Scholar] [CrossRef]
- Marková, H.; Tarkowská, D.; Čečetka, P.; Kočová, M.; Rothová Holá, D. Contents of endogenous brassinosteroids and the response to drought and/or exogenously applied 24-epibrassinolide in two different maize leaves. Frontiers in Plant Science 2023, 14, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Gandhi, A.; Oelmüller, R. Emerging Roles of Receptor-like Protein Kinases in Plant Response to Abiotic Stresses. International Journal of Molecular Sciences 2023, 24, 14762. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Zheng, J.; Huang, R.; Huang, Y.; Wang, H.; Jiang, L.; Fang, X. Phytohormones signaling and crosstalk regulating leaf angle in rice. Plant Cell Reports 2016, 2016. 35, 2423–2433. [Google Scholar] [CrossRef]
- Ladeynova, M.; Mudrilov, M.; Berezina, E.; Kior, D.; Grinberg, M.; Brilkina, A.; Sukhov, V.; Vodeneev, V. Spatial and temporal dynamics of electrical and photosynthetic activity and the content of phytohormones induced by local stimulation of pea plants. Plants. 2020, 9, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Fan, X.Y.; Cao, D.M.; Tang, W.; He, K.; Zhu, J.Y.; He, J.X.; Bai, M.Y.; Zhu, S.; Oh, E.; et al. Integration of brassinosteroid signal transduction into the transcriptional network to regulate plant growth in Arabidopsis. Development Cell. 2010, 19, 765–77. [Google Scholar] [CrossRef]
- Lee, J.; Han, S.; Lee, H.; Jeong, B.; Heo, T.; Hyun, T.K.; Kim, K.; Je, B.I.; Lee, H.; Shim, D.; et al. Brassinosteroids facilitate 1034 xylem differentiation and wood formation in tomato. Planta 2019, 249, 1391–1403. [Google Scholar] [CrossRef]
- Kinoshita, T.; Cano-Delgado, A.; Seto, H.; Hiranuma, S.; Fujioka, S.; Yoshida, S.; Chory, J. Binding of brassinosteroids to 1036 the extracellular domain of plant receptor kinase BRI1. Nature 2005, 433, 164–167. [Google Scholar] [CrossRef]
- Ma, W.; Liu, X.; Chen, K.; Yu, X.; Ji, D. Genome-wide re-identification and analysis of crRLK1Ls in tomato. Int. J. Mol. Sci. 2023, 24, 3142. [Google Scholar] [CrossRef]
- Lozano-Elena, F.; Caño-Delgado, A.I. Emerging roles of vascular brassinosteroid receptors of the BRI1-like family. Curr. Opin. Plant Biol. 2019, 51, 105–113. [Google Scholar] [CrossRef]
- Sousa, V.Q.; Messias, W.F.S.; Pereira, Y.C.; da Silva, B.R.S.; Lobato, E.M.S.G.; Alyemeni, M.N.; Ahmad, P.; Lobato, A.K.D.S. Pretreatment with 24- Epibrassinolide Synergistically Protects Root Structures and Chloroplastic Pigments and Upregulates Antioxidant Enzymes and Biomass in Na+-Stressed Tomato Plants. Journal of Plant Growth Regulation 2021, 1–17. [Google Scholar] [CrossRef]
- Jo, H.E.; Son, S.Y.; Lee, C.H. Comparison of Metabolome and Functional Properties of Three Korean Cucumber Cultivars. Front. Plant Sci. 2022, 13, 1–14. [Google Scholar] [CrossRef]
- Lima, J.; Lobato, A. Brassinosteroids improve photosystem II efficiency, gas exchange, antioxidant enzymes and growth of cowpea plants exposed to water deficit. Physiol. Mol. Biol. Plants 2017, 23, 59–72. [Google Scholar] [CrossRef]
- Hakim, M.; Amjad, H.; Qurban, A.; Fen, L. Brassinosteroids [BRs] Role in Plant Development and Coping with Different Stresses. Int J Mol Sci. 2022, 23, 1012. [Google Scholar] [CrossRef]
- Liu, J.; Li, W.; Wu, G.; Ali, K. An update on evolutionary, structural, and functional studies of receptor-like kinases in plants. Front. Plant Sci., Sec. Plant Pathogen Interactions 2024, 15. [CrossRef]
- Bai, Q.; Wang, L.; Huang, S.; Ali, K.; Li, G.; Ren, H.; et al. The receptor-like kinase EMS1 and BRI1 coordinately regulate stamen elongation via the defense factors BES1/BZR1 in Arabidopsis. Plant Sci. 2023, 331, 111673. [Google Scholar] [CrossRef]
- Díaz, K.; Espinoza, L.; Carvajal, R.; Silva-Moreno, E.; Olea, A.F.; Rubio, J. Exogenous Application of Brassinosteroid 24-Norcholane 22(S)-23-Dihydroxy Type Analogs to Enhance Water Deficit Stress Tolerance in Arabidopsis thaliana. Int J Mol Sci. 2021, 22, 1158. [Google Scholar] [CrossRef]
- Li, B.; Zhang, C.; Cao, B.; Qin, G.; Wang, W.; Tian, S. Brassinolide enhances cold stress tolerance of fruit by regulating plasma membrane proteins and lipids. Amino Acids 2012, 43, 2469–2480. [Google Scholar] [CrossRef]
- de Assis-Gomes, M.D.M.; Pinheiro, D.T.; Bressan-Smith, R.; Campostrini, E. Exogenous brassinosteroid application delays senescence and promotes hyponasty in Carica papaya L. leaves. Theoretical and Experimental Plant Physiology 2018, 30, 193–201. [Google Scholar] [CrossRef]
- Ohnishi, T. Recent advances in brassinosteroid biosynthetic pathway: Insight into novel brassinosteroid shortcut pathway. J. Pestic. Sci. 2018, 43, 159–167. [CrossRef]
- Basit, F.; Liu, J.; An, J.; Chen, M.; He, C.; Zhu, X.; Li, Z.; Hu, J.; Guan, Y. Seed priming with brassinosteroids alleviates aluminum toxicity in rice via improving antioxidant defense system and suppressing aluminum uptake. Environmental Science and Pollution Research 2022, 1–15. [Google Scholar] [CrossRef]
- Chakma, S.P.; Chileshe, S.M.; Thomas, R.; Krishna, P. Cotton Seed Priming with Brassinosteroid Promotes Germination and Seedling Growth. Agronomy 2021, 11, 566. [Google Scholar] [CrossRef]
- Kumar, A.S.; Korra, T.; Thakur, R.; Arutsel van, R.; Kashyap, A.S.; Nehela, Y.; Chaplygin, V.; Minkina, T.; Lacombe, C.K.; Achard, B.P. Role of plant secondary metabolites in defense and transcriptional regulation in response to biotic stress. Plant Stress 2023, 8, 100154. [Google Scholar]
- Sarker, U.; Oba, S. The response of salinity stress-induced A. tricolor to growth, anatomy, physiology, non-enzymatic and enzymatic antioxidants. Front Plant Sci. 2020, 1354. [CrossRef]
- Dalio, R.J.D.; Pinheiro, H.P.; Sodek, L.; Haddad, C.R.B. The effect of 24-epibrassinolide and clotrimazole on the adaptation of Cajanus cajan (L.) Millsp. to salinity. Acta Physiol. Plant 2011, 33, 1887–1896. [Google Scholar] [CrossRef]
- Irfan, M.; Alam, J.; Ahmad, I.; Ali, I.; Gul, H. Effects of exogenous and foliar applications of Brassinosteroid (BRs) and salt stress on the growth, yield and physiological parameters of Lycopersicon esculentum (Mill.). Plant Science Today 2017, 4, 88–10. [CrossRef]
- Zhou, F.; Hu, B.; Li, J.; Yan, H.; Liu, Q.; Zeng, B.; Fan, C. Exogenous applications of brassinosteroids promote secondary xylem differentiation in Eucalyptus grandis. Peerj-16250 2024, 14. [CrossRef]
- Yoshizawa, S.; Azuma, T.; Kojima, K.; Inomura, K.; Hasegawa, M.; Nishimura, Y.; et al. Light-driven Proton Pumps as a Potential Regulator for Carbon Fixation in Marine Diatoms. Microbes Environ. 2023, 38. https://www.jstage.jst.go.jp/browse/jsme2. /. [CrossRef]
- Ogweno, J.O.; Song, X.S.; Shi, K.; Hu, W.H.; Mao, W.H.; Zhou, Y.H.; Yu, J.Q.; Nogués, S. Brassinosteroids Alleviate Heat-Induced Inhibition of Photosynthesis by Increasing Carboxylation Efficiency and Enhancing Antioxidant Systems in Lycopersicon esculentum. J. Plant Growth Regul. 2008, 27, 49–57. [Google Scholar] [CrossRef]
- Fariduddin, Q.; Yusuf, M.; Chalkoo, S.; Hayat, S.; Ahmad, A. 28-homobrassinolide improves growth and photosynthesis in Cucumis sativus L. through an enhanced antioxidant system in the presence of chilling stress. Photosynthetica 2011, 49, 55–64. [Google Scholar] [CrossRef]
- Gururani, M.A.; Venkatesh, J.; Tran, L.S.P. Regulation of photosynthesis during abiotic stress-induced photoinhibition. Molecular Plant 2015, 8, 1304–1320. [Google Scholar] [CrossRef]
- Derks, A.; Schaven, K.; Bruce, D. Diverse mechanisms for photoprotection in photosynthesis. Dynamic regulation of photosystem II excitation in response to rapid environmental change. Biochimica et Biophysica Acta (BBA)-Bioenergetics 2015, 184, 468–485. [Google Scholar] [CrossRef]
- Sonjaroon, W.; Jutamanee, K.; Khamsuk, O.; Thussagunpanit, J.; Kaveeta, L.; Suksamrarn, A. Impact of brassinosteroid mimic on photosynthesis, carbohydrate content and rice seed set at reproductive stage under heat stress. Agriculture and Natural Resources 2018, 52, 234–240. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, J.; Sarwar, R.; Zhang, W.; Geng, R.; Zhu, K.-M.; Tan, X.-L. Research progress on the physiological response and molecular mechanism of cold response in plants. Front. Plant Sci. 2024, Sec. Plant Abiotic Stress, 15 - 2024. [CrossRef]
- Zhu, X.; Liang, W.; Cui, X.; Chen, M.; Yin, C.; Luo, Z.; Zhu, J.; Lucas, W.J.; Wang, Z.; Zhang, D. Brassinosteroids promote development of rice pollen grains and seeds by triggering expression of Carbon Starved Anther, a MYB domain protein. Plant J. 2015, 82, 570–581. [Google Scholar] [CrossRef]
- Peres, A.L.G.; Soares, J.S.; Tavares, R.G.; Righetto, G.; Zullo, M.A.; Mandava, N.B.; Menossi, M. Brassinosteroids, the sixth class of phytohormones: a molecular view from the discovery to hormonal interactions in plant development and stress adaptation. International Journal of Molecular Sciences 2019, 20, 331. [Google Scholar] [CrossRef]
- Planas-Riverola, A.; Gupta, A.; Betegón-Putze, I.; Bosch, N.; Ibañes, M.; Caño-Delgado, A.I. Brassinosteroid signaling in plant development and adaptation to stress. Development 2019, 146: dev151894. [CrossRef]
- Choi, W.-G.; Miller, G.; Wallace, I.; et al. Orchestrating rapid long-distance signaling in plants with Ca2+, ROS and electrical signals. Plant J. 2017, 90, 698–707. [Google Scholar] [CrossRef] [PubMed]
- Hayat, F.; Li, J.; Iqbal, S.; Khan, U.; Ali, N.A.; Peng, Y.; Hong, L.; Asghar, S.; Javed, H.U.; Li, C.; et al. Hormonal Interactions Underlying Rootstock-Induced Vigor Control in Horticultural Crops. Applied Sci 2023, 13, 1237. [Google Scholar] [CrossRef]
- Młodzińska-Michta, E. Regulacja wzrostu i rozwoju systemu korzeniowego przez wybrane czynniki zewnętrzne i wewnętrzne. Kosmos. Problemy Nauk Biologicznych 2018, 4, 801–811. (in Polish). [Google Scholar] [CrossRef]
- Moustakas, M. Plant photochemistry, reactive oxygen species, and photoprotection. Photochem 2022, 2, 5–8. [CrossRef]
- Hafeez, M.B.; Zahra, N.; Zahra, K.; Raza, A.; Batool, A.; Shaukat, K.; Khan, S. Brassinosteroids: Molecular and physiological responses in plant growth and abiotic stresses, Plant Stress 2021, 2, 100029, ISSN 2667-064X. [CrossRef]
- Golam, J.A.; Xiao-Jian, X.; Kai, S.; Jing-Quan, Y.; Yan-Hong, Z. Role of Brassinosteroid in Plant Adaptation to Abiotic Stresses and its Interplay with their Hormones. Current Protein & Peptide Science 2015, 16. [CrossRef]
- Reissig, G.N.; de Carvalho Oliveira, T.F.; Parise, A.G.; Costa, Á.V.L.; Posso, D.A.; Rombaldi, C.V.; Souza, G.M. Approximate entropy: a promising tool to understand the hidden electrical activity of fruit. Commun Integr Biol. 2023, 16, 2195236. [Google Scholar] [CrossRef]
- Sheperd, V.A. From semi-conductors to the rhythms of sensitive plants: the research of J. C. Bose. Cell Mol Biol 2005, 51, 607–619. [Google Scholar]
- Sukhova, E.; Sukhov, V. Electrical signals, plant tolerance to actions of stressors, and programmed cell death: is interaction possible? Plants 2021, 10, 1704. [Google Scholar] [CrossRef]
- Xia, X.J.; Fang, P.P.; Guo, X.; Qian, X.J.; Zhou, J.; Shi, K.; Zhou, Y.H.; Yu, J.Q. Brassinosteroid-Mediated Apoplastic H2O2-Glutaredoxin 12/14 Cascade Regulates Antioxidant Capacity in Response to Chilling in Tomato. Plant Cell Environ. 2018, 41, 1052–1064. [Google Scholar] [CrossRef]
- Fuentes, D.E.; Acuña, L.G.; Calderón, I.L. Stress response and virulence factors in bacterial pathogens relevant for Chilean aquaculture: current status and outlook of our knowledge. Biol Res 2022, 55, 21. [Google Scholar] [CrossRef]
- Wilson, S.K.; Pretorius, T.; Naidoo, S. Mechanisms of systemic resistance to pathogen infection in plants and their potential application in forestry. BMC Plant Biology 2023, 23, 404. [Google Scholar] [CrossRef]
- Saini, S.; Kaur, N.; Pati, P.K. Phytohormones: Key players in the modulation of heavy metal stress tolerance in plants. Ecotoxicology and Environmental Safety 2021, 223, 112578. [Google Scholar] [CrossRef] [PubMed]
- Hafeez, M.B.; Raza, A.; Zahra, N.; Shaukat, K.; Akram, M.Z.; Iqbal, S.; Basra, S.M.A.; Irfana, L.; Syeda, F. Amjad, Nida Mansoora et al. Interactive effects of brassinosteroids and timber waste biochar enhances the drought tolerance capacity of wheat plant, 2022, PREPRINT (Version 2) available at Research Square. [CrossRef]
- Fricke, W. Fricke, W. Rapid and tissue-specific accumulation of solutes in the growth zone of barley leaves in response to salinity. Planta 2004, 219, 515–525. [Google Scholar] [CrossRef]
- Vikram et al. Role of Brassinosteroids in plants responses to salinity stress: A review. Journal of Applied and Natural Science 2022, 14, 582 -599. [CrossRef]
- Clauw, P.; Coppens, F.; Korte, A.; Herman, D.; Slabbinck, B.; Dhondt, S.; Van Daele, T.; De Milde, L.; Vermeersch, M.; Maleux, K.; Maere, S.; Gonzalez, N.; Inzé, D. Leaf growth response to mild drought: natural variation in Arabidopsis sheds light on trait architecture. The Plant Cell 2016, 28, 2417–2434. [Google Scholar] [CrossRef] [PubMed]
- Irfana, L.; Syeda, F. Amjad, Mansoora N., et al. Interactive effects of brassinosteroids and timber waste biochar enhances the drought tolerance capacity of wheat plant, 2022, PREPRINT (Version 2) available at Research Square. [CrossRef]
- Kiliç, S.; Çavuşoğlu, K.; Kabar, K. Effects of 24-epibrassinolide on salinity stress induced inhibition of seed germination, seedling growth and leaf anatomy of barley. Süleyman Demirel University Faculty of Arts and Science. Journal of Science 2007, 2, 41–52. [Google Scholar]
- Safdar, H.; Amin, A.; Shafiq, Y.; Ali, A.; Yasin, R.; Shoukat, A.; Ul Hussan, M.; Sarwar, M.I. A review: Impact of salinity on plant growth. Nat. Sci 2019, 17, 34–40. [Google Scholar]
- Das, T.; Shukla, Y.M.; Poonia, T.C.; Meena, M.; Meena, M.D. Effects of brassinolide on physiological characteristics of rice (Oryza sativa L.) with different salinity levels. Ann. Biol 2013, 29, 228–231. [Google Scholar]
- Ranf, S. Pattern recognition receptors—versatile genetic tools for engineering broad-spectrum disease resistance in crops. Agronomy 2018, 8, 134. [Google Scholar] [CrossRef]
- Wang, Y.H.; Irving, H.R. Developing a model of plant hormone interactions. Plant Signaling & Behavior 2011, 6, 494–500. [Google Scholar]
- Sirangelo, T.M.; Ludlow, R.A.; Spadafora, N.D. Molecular Mechanisms Underlying Potential Pathogen Resistance in Cannabis sativa. Plants. 2023, 12, 2764. [Google Scholar] [CrossRef]
- Anonimous 2023. Create Professional Science Figures in Minutes. https://www.biorender.com.
- Kan, Y.; Mu, X.R.; Gao, J.; Lin, H.-X.; Lin, Y. The molecular basis of heat stress responses in plants. Molecular Plant 2023, 16, 1612–1634. [Google Scholar] [CrossRef] [PubMed]
- Eremina, M.; Unterholzner, S.J.; Rathnayake, A.I.; Castellanos, M.; Khan, M.; Kugler, K.G.; May, S.T.; Mayer, K.F.; Rozhon, W.; Poppenberger, B. Brassinosteroids participate in the control of basal and acquired freezing tolerance of plants. Proc. Natl. Acad. Sci. USA 2016, 113, E5982–E5991. [Google Scholar] [CrossRef]
- Zhu, T.; Deng, X.; Zhou, X.; Zhu, L.; Zou, L.; Li, P.; Zhang, D.; Lin, H. Ethylene and hydrogen peroxide are involved in brassinosteroid-induced salt tolerance in tomato. Sci. Rep. 2016, 6, 35392. [Google Scholar] [CrossRef]
- Rajewska, I.; Talarek, M.; Bajguz, A. Brassinosteroids and response of plants to heavy metals action. Front. Plant Sci. 2016, 7, 629. [Google Scholar] [CrossRef] [PubMed]
- Xiong, M.; Chu, L.; Li, Q.; Yu, J.; Yang, Y.; Zhou, P.; Zhou, Y.; Zhang, C.; Fan, X.; Zhao, D.; Yan, C.; Liu, Q. Brassinosteroid and gibberellin coordinate rice seed germination and embryo growth by regulating glutelin mobilization. The Crop Journal 2021, 9, 1039–1048. [Google Scholar] [CrossRef]
- Wang, Y.T.; Chen, Z.Y.; Jiang, Y.; Duan, B.B.; Xi, Z.M. Involvement of ABA and Antioxidant System in Brassinosteroid-Induced Water Stress Tolerance of Grapevine [Vitis vinifera L.]. Sci. Hortic 2019, 256, 108596. [Google Scholar] [CrossRef]
- Mazorraa, L.M.; Holtonb, N.; Bishopb, G.J.; Ineza, M. Heat shock response in tomato brassinosteroid mutants indicates that thermotolerance is independent of brassinosteroid homeostasis. Plant Physiol. Biochem. 2011, 49, 1420–1428. [Google Scholar] [CrossRef]
- Hussain, M.; Khan, T.A.; Yusuf, M.; Fariduddin, Q. Silicon-Mediated Role of 24-Epibrassinolide in Wheat under High-Temperature Stress. Environ. Sci. Pollut. Res. 2019, 26, 17163–17172. [Google Scholar] [CrossRef]
- Xi, Z.; Wang, Z.; Fang, Y.; Hu, Z.; Hu, Y.; Deng, M.; Zhang, Z. Effects of 24-Epibrassinolide on Antioxidation Defense and Osmoregulation Systems of Young Grapevines (V. vinifera L.) under Chilling Stress. Plant Growth Regul 2013, 71, 57–65. [Google Scholar] [CrossRef]
- Sawicka, B.; Kalembasa, D. Annual Variability of Some Toxic Element Contents (Cd, Cr, Co, Ni, and Pb) and Response of Two Jerusalem Artichoke Varieties to Increasing Nitrogen Fertilizer at Constant PK Levels. Pol. J. Environ. Stud. 2013, 22, 861–871. [Google Scholar]
- Biziewska, I.; Talarek, M.; Bajguz, A. Rola brassinosteroidów w odpowiedzi roślin na stres metali ciężkich. [w:] A. Bajguz, I. Ciereszko [red.] Różnorodność biologiczna – od komórki do ekosystemu Funkcjonowanie roślin i grzybów. Środowisko – eksperyment – edukacja, Polskie Towarzystwo Botaniczne 2015, 253–263. (in Polish). [Google Scholar]
- Sharma, I.; Pati, P.K.; Bhardwaj, R. Effect of 28-homobrassinolide on antioxidant defense system in Raphanus sativus L. under chromium toxicity. Ecotoxicology 2011, 20, 862–874. [Google Scholar] [CrossRef]
- Hayat, S.; Maheshwari, P.; Wani, A.S.; Irfan, M.; Alyemeni, M.N.; Ahmad, A. Comparative effect of 28 homobrassinolide and salicylic acid in the amelioration of NaCl stress in Brassica juncea L. Plant Physiol. Biochem. 2012, 53, 61–68. [Google Scholar] [CrossRef]
- Rampelotto, P.H.; Giannakos, N.R.O.; Mena Canata, D.A.; Pereira, F.D.; Hackenhaar, F.S.; Pereira, M.J.R.; Benfato, M.S. Oxidative Stress and Antioxidant Defense in the Brain of Bat Species with Different Feeding Habits. Int. J. Mol. Sci. 2023, 24, 12162. [Google Scholar] [CrossRef]
- Feuillet, C.; Schachermayr, G.; Keller, B. Molecular cloning of a new receptor-like kinase gene encoded at the Lr10 disease resistance locus of wheat. The Plant Journal 1997, 11, 45–52. [Google Scholar] [CrossRef]
- Lim, C.W.; Lee, S.C. Arabidopsis abscisic acid receptors play an important role in disease resistance. Plant Molecular Biology 2015, 88, 313–324. [Google Scholar] [CrossRef] [PubMed]
- Veley, K.M.; Michaels, S.D. Functional redundancy and new roles for genes of the autonomous floral-promotion pathway. Plant Physiology 2008, 147, 682–695. [Google Scholar] [CrossRef] [PubMed]
- Shahid, M.A.; Pervez, M.A.; Balal, R.M.; Mattson, N.S.; Rashid, A.; Ahmad, R.; Ayyub, C.M.; Abbas, T. Brassinosteroid (24-epibrassinolide) enhances growth and alleviates the deleterious effects induced by salt stress in pea (Pisum sativum L.). Australian Journal of Crop Science 2011, 5, 500–510. [Google Scholar]
- Soliman, M.; Elkelish, A.; Souad, T.; Alhaithloul, H.; Farooq, M. Brassinosteroid seed priming with nitrogen supplementation improves salt tolerance in soybean. Physiology and Molecular Biology of Plants 2020, 26, 501–511. [Google Scholar] [CrossRef] [PubMed]
- Sawicka, B.; Barbaś, P.; Skiba, D.; Pszczółkowski, P.; Yeganehpoor, F. Oxylipins in Plant Protection/Disease Management. [in:] Phyto-Oxylipins. 1st Edition, First Published 2023, Imprint: CRC Press, pp. 19; eBook ISBN: 9781003316558.
- Tang, K.W.K.; Millar, B.C.; Moore, J.E. Antimicrobial Resistance (AMR). Br J Biomed Sci. 2023, 80, 11387. [Google Scholar] [CrossRef] [PubMed]
- Joshi, T.; Sharma, P.; Chandra, S.; et al. A spotlight on the recent advances in bacterial plant diseases and their footprint on crop production [in:] Recent Advancements in Microbial Diversity, Editor(s): Surajit De Mandal, Pankaj Bhatt, Academic Press, 2020, 37-56, ISBN: 9780128212653. [CrossRef]
- Manghwar, H.; Zaman, W. Plant Biotic and Abiotic Stresses — Volume II. Editors H. Manghwar & W. Zaman. Editorial Office: MDPI, Basel, Switzerland, 2023, ISBN: 978-3-0365-8538-3(Hbk); ISBN: 978-3-0365-8539-0(PDF). [CrossRef]
- Thussagunpanit, J.; Jutamanee, K.; Sonjaroon, W.; Kaveeta, L.; Chai-Arree, W.; Pankean, P.; Suksamrarn, A. Effects of brassinosteroid and brassinosteroid mimic on photosynthetic efficiency and rice yield under heat stress. Photosynthetica 2015, 53, 312–320. [Google Scholar] [CrossRef]
- Hu, S.; Wang, C.; Sanchez, D.L.; Lipka, A.E.; Liu, P.; Yin, Y.; Blanco, M.; Lübberstedt, T. Gibberellins Promote Brassinosteroids Action and Both Increase Heterosis for Plant Height in Maize (Zea mays L.). Front. Plant Sci. 2017, Sec. Plant Breeding, 8 - 2017. [CrossRef]
- Mmbando, G.S. The recent relationship between ultraviolet-B radiation and biotic resistance in plants: a novel non-chemical strategy for managing biotic stresses. Plant Signaling & Behavior 2023. [CrossRef]
- Das, S.R.; Rout, GR. Role of Brassinosteroids on Plant Growth and Development. [In book:] Jasmonates and Brassinosteroids in Plants: Metabolisms, signaling and Biotechnological applications. Edited by Akul Ramakrishna, Geetika Sirhindi. Publisher: CRC Press, pp. 246. 2023.
- Divi, U.K.; Krishna, P. Brassinosteroid: A biotechnological target for enhancing crop yield and stress tolerance. New Biotechnol. 2009, 26, 131–136. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.; Sharma, V.; Kaur, G.; Lata, C.; Dasila, H.; Perveen, K.; Khan, F.; Gupta, V.K.; Khanam, M.N. Brassinosteroids as promoters of seedling growth and antioxidant activity under heavy metal zinc stress in mung bean (Vigna radiata L.). Front. Microbiol. 2023, 14, 1259103. [CrossRef]
- Bajguz, A.; Chmur, M.; Gruszka, D. Comprehensive Overview of the Brassinosteroid Biosynthesis Pathways: Substrates, Products, Inhibitors, and Connections. Frontiers in Plant Science 2020, 11, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Nelissen, H.; Gonzalez, N.; Inze, D. Leaf growth in dicots and monocots: so different yet so alike. Current Opinion in Plant Biology 2016, 33, 72–76. [Google Scholar] [CrossRef]
- Li, Y.; Hua, J.; Hou, X.; et al. Brassinosteroids is involved in methane-induced adventitious root formation via inducing cell wall relaxation in marigold. BMC Plant Biol 2023, 23. [Google Scholar] [CrossRef]
- Cimino, F.P.; Núñez, M.G.; Rosado-Abón, A.; Amesty, Á.; Estévez-Braun, A.; Díaz, K.; Espinoza, L.C.; Iglesias-Arteaga, M.A. Methyl esters of 23,24-Dinor-5α-cholan-22-oic acids as brassinosteroid Analogues. Synthesis, evaluation of plant growth promoting activity and Molecular docking. Steroids 2023, 196, 109248. [Google Scholar] [CrossRef]
- Cui, Z.; Zhao, Z.; Yao, L.; Hu, Y. Determination and analysis of solubility of brassinolide in different solvent systems at different temperatures (T = 278.15–323.15 K). Journal of Molecular Liquids 2021, 340, 117316. [Google Scholar] [CrossRef]
- Benson, C.L.; Kepka, M.; Wunschel, C.; Rajagopalan, N.; Nelson, K.M.; Christmann, A.; Abrams, S.R.; Grill, E.; Loewen, M.C. Abscisic acid analogs as chemical probes for dissection of abscisic acid responses in Arabidopsis thaliana. Phytochemistry 2015, 113, 96–107. [Google Scholar] [CrossRef]
- Thongsri, K.; Teingtham, K.; Duangpatra, J.; Romkaew, J. Effects of brassinosteroids and gibberellin on water uptake and performance of soya bean seeds under different temperatures. Seed Science and Technology 2021, 49, 141–157. [Google Scholar] [CrossRef]
- Hayat, S.; Ahmad, A. 28-Homobrassinolide induced changes favored germinability of wheat grains. Bulg. J., Plant Physiol 2003, 29, 55–62. [Google Scholar]
- Ren, Y.; Che, X.; Liang, J.; Wang, S.; Han, L.; Liu, Z.; Chen, H.; Tang, M. Brassinosteroids benefit plants performance by augmenting arbuscular mycorrhizal symbiosis. Microbiology Spectrum 2021, 9, e01645–21. [Google Scholar] [CrossRef]
- Nováková, O.; Kuneš, I.; Gallo, J.; Baláš, M. Effects of brassinosteroids on prosperity of Scots pine seedlings. Journal of Forest Science 2014, 60, 388–393. [Google Scholar] [CrossRef]
- Tikkanen, M.; Mekala, N.R.; Aro, E.M. Photosystem II photoinhibition-repair cycle protects Photosystem I from irreversible damage. Biochimica et Biophysica Acta (BBA) - Bioenergetics 2014, 837, 210–215. [Google Scholar] [CrossRef]
- Nishiyama, Y.; Murata, N. Revised scheme for the mechanism of photoinhibition and its application to enhance the abiotic stress tolerance of the photosynthetic machinery. Applied Microbiology and Biotechnology 2014, 98, 8777–8796. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.; Rozhon, W.; Poppenberger, B. The role of hormones in the aging of plants-a mini-review. Gerontology 2013, 60, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Kaplan-Dalyan, E.; Sağlam-Çağ, S. The effect of epibrassinolide on senescence in horizontal sunflower (Helianthus annuus L.) seedlings. European Journal of Biology 2013, 72, 33–44. [Google Scholar]
- Efimova, M.V.; Savchuk, A.L.; Hasan, J.A.K.; Litvinovskaya, R.P.; Khripach, V.A.; Kholodova, V.P.; Kuznetsov, V.V. Physiological mechanisms of enhancing salt tolerance of oilseed rape plants with brassinosteroids. Russian Journal of Plant Physiology 2014, 61, 733–743. [Google Scholar] [CrossRef]
- Houimli, S.I.M.; Denden, M.; Mouhandes, B.D. Effects of 24-epibrassinolide on growth, chlorophyll, electrolyte leakage and proline by pepper plants under NaCl-stress. EurAsian Journal of BioSciences 2010, 4, 96–104. [Google Scholar] [CrossRef]
- Khan, I.; Awan, S.A.; Ikram, R.; Rizwan, M.; Akhtar, N.; Yasmin, H.; Sayyed, R.Z.; Ali, S.; Ilyas, N. Effects of 24-epibrassinolide on plant growth, antioxidants defense system, and endogenous hormones in two wheat varieties under drought stress. Physiol Plant. 2021, 172, 696–706. [Google Scholar] [CrossRef] [PubMed]
- Jaiswal, S.; Båga, M.; Chibbar, R.N. Brassinosteroid receptor mutation influences starch granule size distribution in barley grains. Plant Physiol. Biochem. 2020, 154, 369–378. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The brassinosteroid biosynthesis pathway in rice and the enzymes that catalyze them. Abbreviations for each compound are given in parentheses below the full name. Source: own.
Figure 1.
The brassinosteroid biosynthesis pathway in rice and the enzymes that catalyze them. Abbreviations for each compound are given in parentheses below the full name. Source: own.
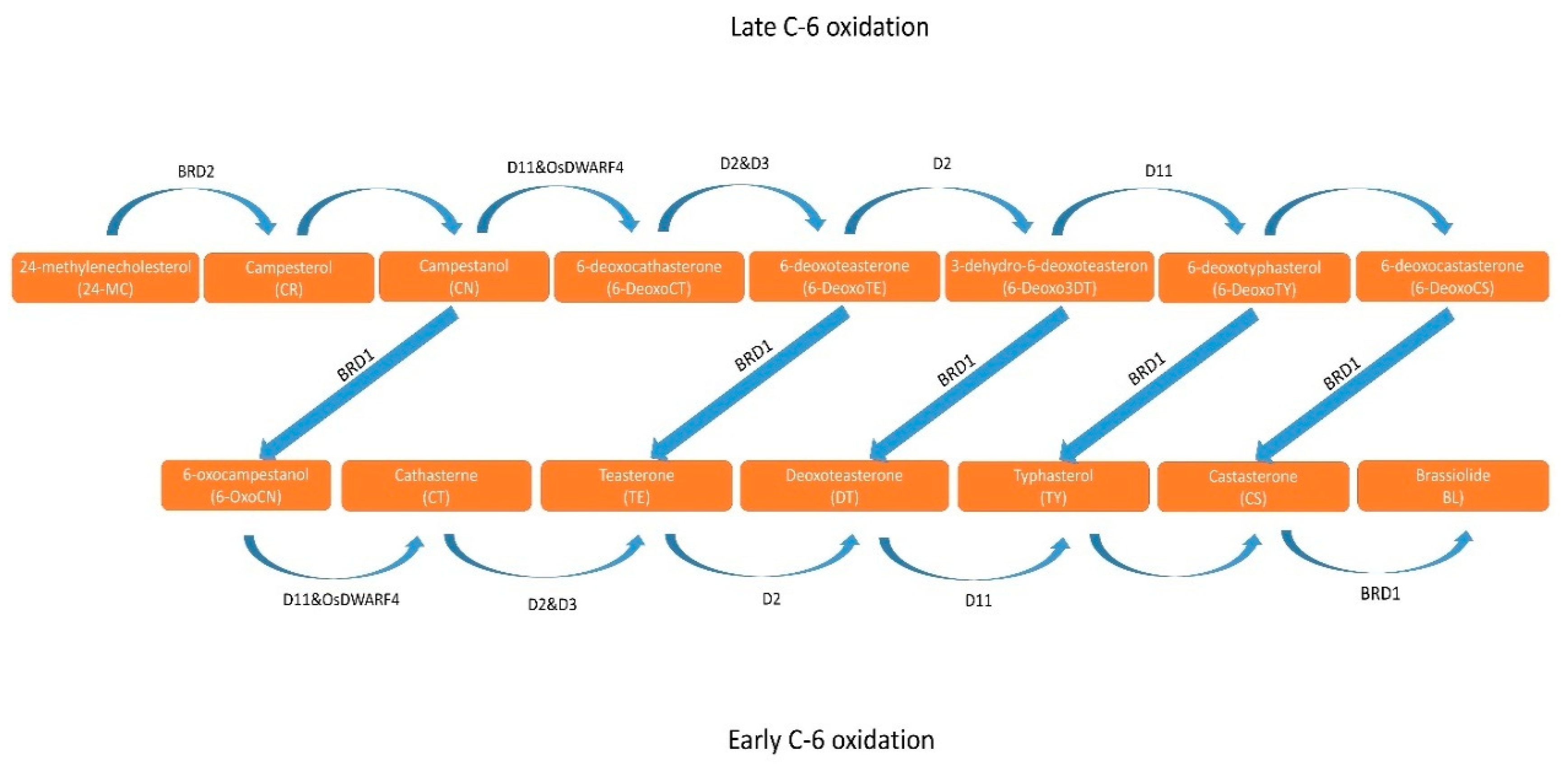
Figure 2.
Chemical structure of BL with steroid rings (A, B, C and D). Parts within the dashed lines can be replaced with different groups. Source: own.
Figure 2.
Chemical structure of BL with steroid rings (A, B, C and D). Parts within the dashed lines can be replaced with different groups. Source: own.
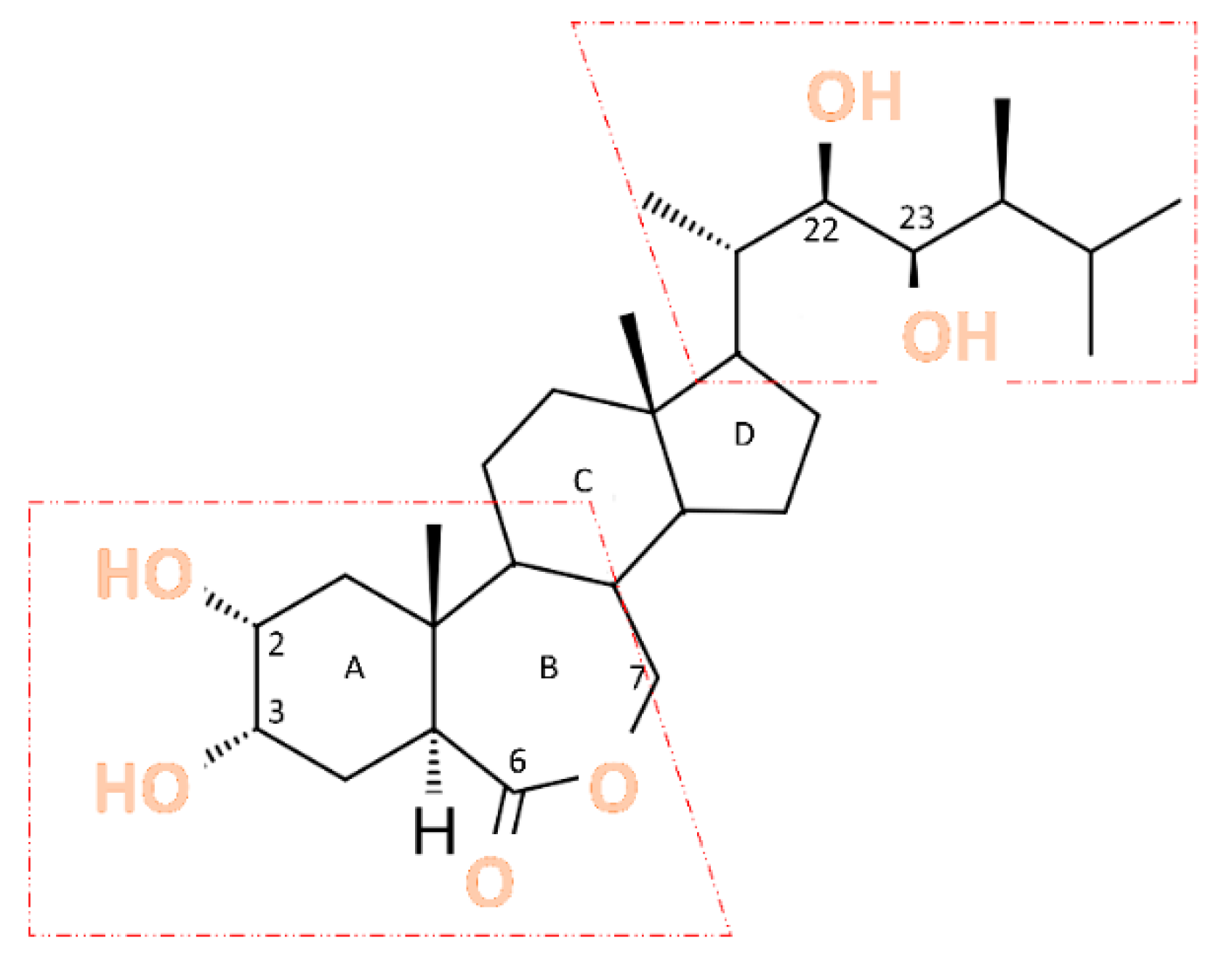
Figure 3.
RKL responses to abiotic stress. Source: own.
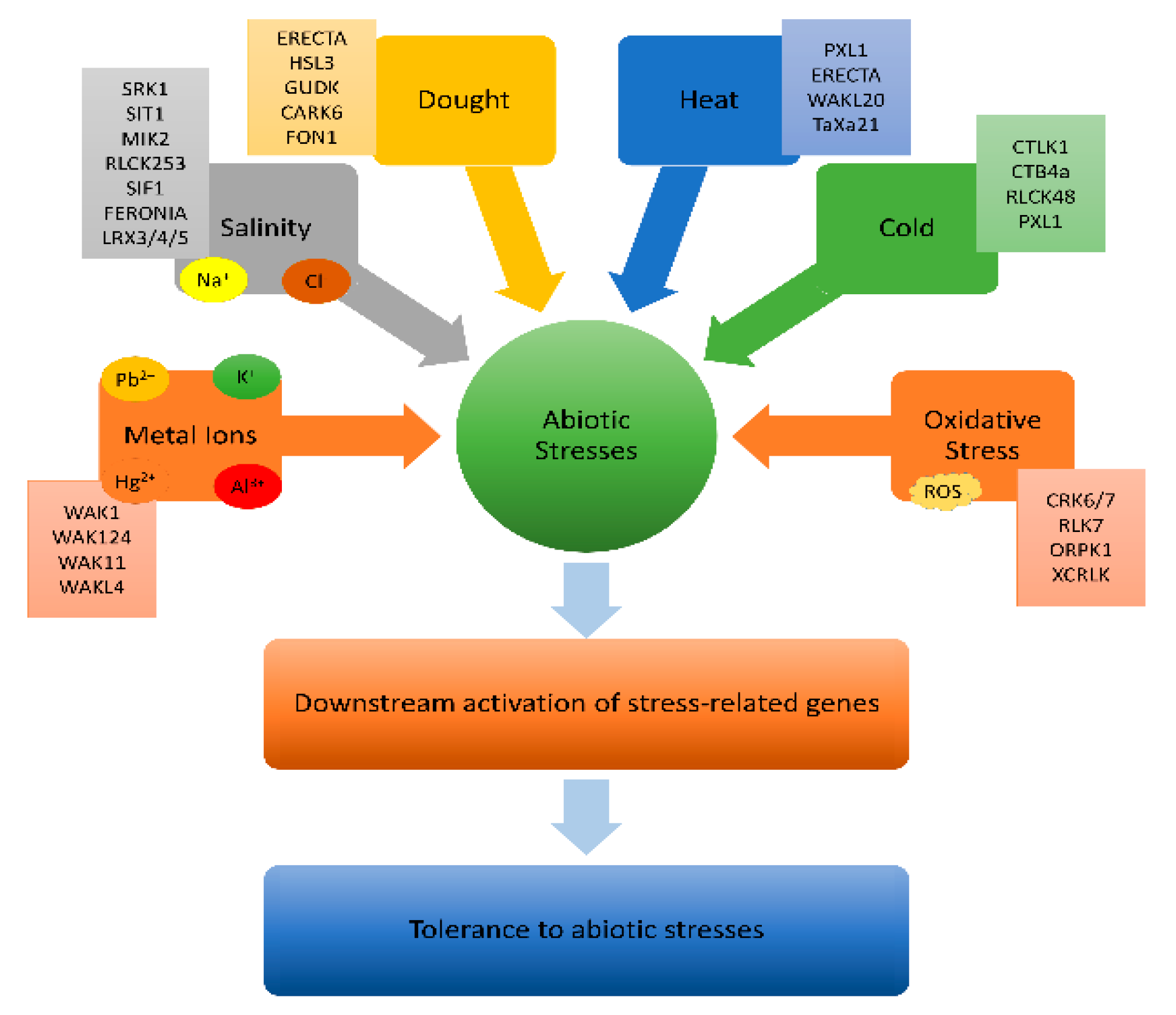
Figure 4.
Regulation of plant development processes by BR. Source: own adapted to Chaudhuri et al. [27].
Figure 4.
Regulation of plant development processes by BR. Source: own adapted to Chaudhuri et al. [27].
Figure 5.
Multidirectional effects of BR. Source: own.
Figure 6.
Selected key roles of BR in plants. Source: own.
Figure 7.
The importance of BR in agriculture, horticulture and plant protection. Source: own.
Figure 8.
This mechanism illustrates how exogenous application of 24-epibrassinolide (EBR) facilitates the adaptation of perennial ryegrass to salt stress by modulating multiple physiological and molecular processes involved in stress tolerance. Source: own.
Figure 8.
This mechanism illustrates how exogenous application of 24-epibrassinolide (EBR) facilitates the adaptation of perennial ryegrass to salt stress by modulating multiple physiological and molecular processes involved in stress tolerance. Source: own.
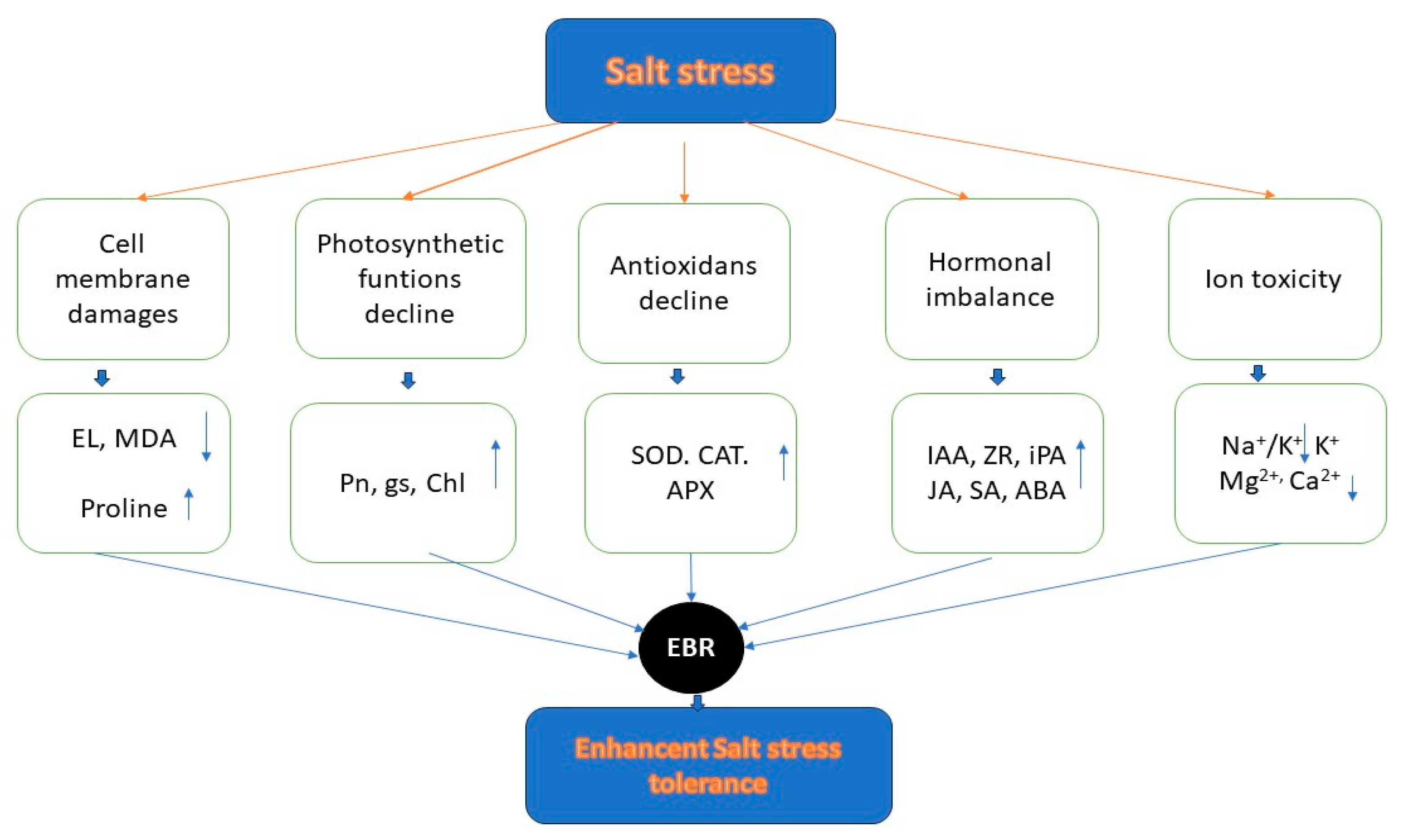
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



