
Submitted:
19 February 2024
Posted:
22 February 2024
You are already at the latest version
Abstract
Chagas disease is a Neglected Tropical Disease with limited and ineffective therapy. In a search for new antitrypanosomal compounds, we investigated the potential of metabolites from bacteria living in corals and sediments of the Southeastern Brazilian coast. Three corals Tubastraea coccinea, Mussismilia hispida, Madracis decactis and sediments yielded 11 bacterial strains, that were fully identified by MALDI-ToF/MS or gene sequencing, resulting in six genera - Vibrio, Shewanella, Mesoflavibacter, Halomonas, Bacillus and Alteromonas. To conduct this study, EtOAc extracts were prepared and tested against Trypanosoma cruzi. Crude extracts showed IC50 values ranging from 15 to 51 μg/mL against trypomastigotes. The bacterium Mesoflavibacter zeaxanthinifaciens was selected for fractionation, resulting in an active fraction (FII) with IC50 values of 17.7 μg/mL and 23.8 μg/mL against the trypomastigotes and amastigotes, respectively, with neither mammalian cytotoxicity nor hemolytic activity. Using NMR and ESI-HRMS analysis, FII revealed the presence of unsaturated iso-type fatty acids. Its lethal action was investigated, leading to a protein spectral profile of the parasite altered after treatment. FII also induced a rapid permeabilization of the plasma membrane of the parasite, leading to cell death. These findings demonstrate these unsaturated iso-type fatty acids as possible new hits against T. cruzi.
Keywords:
marine bacteria
; Trypanosoma cruzi
; iso-fatty acids
; metabolites
; antimicrobial
1. Introduction
Marine microorganisms have developed defense strategies that include the production of different metabolites and enzymes with important applications for humans [1]. These low molecular weight compounds are produced by living organisms to perform biological functions for their survival and as well as in defense [2]. Exploration of marine microbiota is recent, but already shows great promise in both medical and biotechnology fields [3]. Since the oceans cover most of the Earth’s surface, they host a substantial portion of the biodiversity, which lives under distinct and varied conditions and has evolved through a long period of metabolic adaptations. Exploration of the biodiversity and metabolic chemodiversity of the oceans has resulted in the discovery of thousands of structurally unique bioactive marine natural products [4]. Considering that oceans encompass typical microbial abundances of 106 microorganisms per mL in seawater and 109 per mL in ocean-bottom sediments [5], the exploration of marine microorganisms as sources of bioactive marine organisms has led to the discovery of promising new drug candidates [1,6]. A review by Newman and Craig (2020) [7] showed that about 56% of 1881 therapeutic products approved by the US FDA between 1981 and 2019 had natural prototypes, especially obtained from marine microorganisms.
Chagas disease is a parasitic infection considered neglected by the World Health Organization (WHO). This condition affects over eight million people worldwide, mainly in Latin America. It is caused by the protozoan parasite Trypanosoma cruzi and transmitted mostly by the insect Triatoma sp or by oral contamination of food. In non-endemic countries, it is present due to immigration and disseminated by contaminated food and organ transplantation [8,9]. The parasite can be identified in two clinically relevant forms: trypomastigotes, which are found in blood and extracellular compartments and are considered the infective form, and amastigotes, which are found in the affected tissues during latent and chronic disease and are the intracellular replicative forms. The current treatment employs only two highly toxic drugs, benznidazole or nifurtimox. Both drugs are associated with severe adverse effects, but only benznidazole is still approved in Brazil. A large multicentric study confirmed the low efficacy of benznidazole to eliminate the parasites in chronic patients [10], emphasizing the urgent need for safer and more effective treatments for Chagas disease.
The marine environment has been a source of more than 20,000 inspirational natural products discovered over the past 50 years. In view of the huge chemodiversity present in microorganisms living in the oceans, numerous biologically active compounds have been described [11]. For example, small molecules isolated from the marine bacterium Bacillus pumilus, present in the black coral Antipathes sp. of the Pacific coast of Panama have shown potent activity against Trypanosoma cruzi with IC50 values from 19 to 27 μM [12]. Penidigiamycin-A, an antiprotozoal compound isolated from the marine bacterium Paenibacillus sp. DE2SH, showed anti-Leishmania (IC50 7 µM), anti-Trypanosoma brucei (IC50 0.78 µM) and anti-Plasmodium falciparum (IC50 9.1 µM) activity [13]. The Blue Amazon is the Brazilian oceanic area of the Atlantic that comprises 3.6 million square kilometers and presents a huge biodiversity of potential pharmaceutical candidates to fight Neglected Tropical Diseases (NTDs). In this study, we examined the antiparasitic potential of bacterial metabolites isolated from three corals and sediments collected from the North coast of São Paulo, Brazil (Figure 1, orange arrow).
2. Results
2.1. Collection of Marine Invertebrates
In our study, we collected three different species of marine invertebrates and two sediment samples by scuba diving in the geographical area of the São Sebastião Channel and Buzios Island (Table 1). The depth of the collection ranged from 5 to 35 meters (Table 1) and the collected specimens are shown in Figure 2.
2.2. Isolation and Identification of Marine Bacteria.
The microbiota associated to the corals was isolated under sterile conditions and stored at -85°C. The microorganisms were identified with MALDI-ToF/MS (Bruker-Daltonics, Germany), according to the logarithmic system of intrinsic scoring of the program that attributes a degree of confidence (score) to the result (Table 2). After Gram staining, it was found that most of the isolated bacteria were Gram-negative bacilli (Table 2).
Considering the limited databank for marine bacteria, isolates that could not be identified by in the MALDI-TOF/MS were selected for partial sequence identification of the 16S rRNA gene. After sequencing, nucleotide sequences were compared to those deposited in the sequence banks EzTaxon (https://www.ezbiocloud.net/),(https://www.ezbiocloud.net/), SepsiTest (http://www.sepsitest-last.de/en/index) and Microbenet (https://microbenet.cdc.gov/) and BLAST 16S rRNA database. Table 3 shows the respective percentages of similarity based on the sequencing of the partial gene 16S rRNA, so that the identified organisms are compatible with the marine bacteria Alteromonas macleodii. Shewanella pneumatophori, Mesoflavibacter zeaxanthinifaciens and Halomonas aquamarina, respectively.
Thus, the study identified 11 strains belonging to 7 different species: Alteromonas macleodii, Vibrio harveyi, Vibrio alginolyticus, Shewanella pneumatophori, Mesoflavibacter zeaxanthinifaciens and Halomonas aquamarina.
2.3. Extraction of microbial metabolites and evaluation of the 50% inhibitory concentration (IC50) for T. cruzi trypomastigotes.
To evaluate the anti-Trypanosoma cruzi potential of microbial metabolites, crude organic extracts were obtained after cultivation in Marine Agar medium in Petri dishes of 140 x 15 mm. A total of seven plates were used and cultured for 120 h for each species. The organic extracts of the 11 strains isolated from marine and sediment were obtained, ranging from 2 to 11 mg (Table 4).
The 50% inhibitory concentration (IC50) of the organic extracts were quantified against trypomastigote forms of T. cruzi, and all 11 extracts had antiparasitic activity, with IC50 values ranging from 8 to 60 μg/mL. The least active extract was from Bacillus megaterium (60 µg/mL), isolated in the sediments of the São Sebastião Channel (Table 4). The most active extracts were from Vibrio spp. isolated from different corals and marine sediments, with IC50 values ranging from 8 to 51 μg/mL, as well as Shewanella pneumatophori (15 μg/mL), Mesoflavibacter zeaxanthinifaciens (18 μg/mL) and Halomonas aquamarina (15 μg/mL). Based on the dose-response sigmoidal curves (data not shown), six microbial extracts killed 100% of the parasites to the highest tested concentration, ranked as follows: TC 2.2 (V. alginolyticus), MH 3.3 (V. alginolyticus), SCSB 6.0.2.2 (Mesoflavibacter zeaxanthinifaciens), SCSB 6.2 (V. harveyi), SIBUZ 7 (V. harveyi), SIBUZ 7.2.2. (Halomonas aquamarina). In addition, based on the method of cellular viability of resazurin by mitochondrial oxidative activity, it was possible to verify that all the extracts showed a trypanocidal activity.
2.4. Fractionation of Mesoflavibacter zeaxanthinifaciens extract
Considering the potency of Mesoflavibacter zeaxanthinifaciens and the amount of the obtained EtOAc extract, it was selected for a chromatographic fractionation to produce four fractions (F0, FI, FII and FIII).
After evaluation of fractions against trypomastigotes, fraction II (FII) eliminated 100% of the parasite after 24 h at a tested dose of 100 µg/mL. Thus, to identify the main compounds in this bioactive fraction, it was analyzed by NMR and by ESI-HRMS. (Figure 3).
Initially, the NMR 1H spectrum indicated the predominance of fatty material due to the presence of an intense singlet at δ 1.2. However, the signal referring to the termination of the carbon chain, commonly observed as one triplet at δ 0.8, was detected as one doublet (J = 6.5 Hz), suggesting the presence of iso fatty acids. To identify these compounds, this fraction was analyzed by ESI-HRMS at negative mode (Figure 3).
According to the literature [14] the presence of iso-type fatty acids was previously described in the study species. Thus, the occurrence of such fatty acids with variation in the extension of the lateral chain was proposed based on the obtained results of ESI-HRMS data.
As indicated in the Figure 3, this spectrum show that four main [M – H]- ions at m/z 297.2790, 311.2954, 325.3104, and 339.260 were observed, which were compatible with molecular formulas C19H37O2- (calculated 297.2794), C20H39O2- (calculated 311.2950), C21H41O2-(calculated 325.3106) and C22H43O2- (calculated 339.3263), all with a degree of unsaturation, i.e. a carbonyl, containing, therefore, saturated side chains. This analysis allowed the identification of a homologous series of C19, C20, C21 and C22 iso-type fatty acids (compounds 1 – 4) as indicated in Figure 4.
2.5. Antitrypanosomal activity of Fraction II of Mesoflavibacter zeaxanthinifaciens EtOAc extract (FII)
The potency of fraction II of Mesoflavibacter zeaxanthinifaciens EtOAc extract was determined in trypomastigotes and intracellular amastigotes of T. cruzi. This fraction showed potent activity against the trypanomastigotes with an IC50 value of 17.7 μg/mL after 48 hours incubation. The intracellular amastigotes were also eliminated with an IC50 value of 23.8 μg/mL. The cytotoxicity of fraction II in NCTC clone 929 cells revealed no toxicity at the highest tested concentration of 200 μg/mL (Table 5).
2.6. Hemolytic Activity of fraction II of Mesoflavibacter zeaxanthinifaciens EtOAc extract
The hemolytic activity of fraction II was evaluated in murine erythrocytes after incubation for 2 h, using water as a positive control (Figure 5). No hemolysis could be detected up to 100 μg/mL and with; at 200 μg/mL, and less than 10% of the erythrocytes were hemolyzed when compared to untreated cell (negative control).
2.7. Protein profile of T. cruzi after treatment with fraction II of Mesoflavibacter zeaxanthinifaciens EtOAc extract (FII).
The protein profile of trypomastigotes was evaluated by MALDI-TOF/MS (Bruker-Daltonics) after treatment with fraction II of Mesoflavibacter zeaxanthinifaciens EtOAc extract (FII) at two concentrations (20 and 50 μg/mL). The parasites were also treated with benznidazole (BZN-40 μM) as comparison. Based on the spectral data of untreated parasites, it was possible to observe significant changes in the protein profile of fraction II treated parasites (Figure 6).
Significant spectral modifications were observed at peaks m/z 4,417, with decreased intensities in the group treated with FII at 50 µg/mL, when compared to untreated parasites and also benznidazole. Similarly, the areas of the peaks at m/z 2,690, 2,780 and 3,0975 were significantly decreased at FII-treated parasites (50 μg/mL), as well as those above m/z 10,000. But an increased peak intensity was found at treated group FII 50 µg/mL at m/z 8,075 and, a similar effect also detected at the benznidazole-treated group. The effect of FII treatment in T. cruzi was clearly dose-dependent, and the alterations of the protein spectral were similar to those found for benznidazole, the standard drug.
2.8. Plasma Membrane Permeability of Trypomastigotes treated with fraction II of Mesoflavibacter zeaxanthinifaciens EtOAc extract
The plasma membrane permeability of trypomastigotes was analyzed in the presence of fraction II of Mesoflavibacter zeaxanthinifaciens EtOAc extract (FII), using the fluorescent probe SYTOX Green. After 10 min incubation, FII induced a rapid permeabilization of the membrane, with significant increase of the fluorescence levels when compared to untreated control (Figure 7). Triton X-100 was used as a positive control and showed the highest levels. After 60 min incubation, the fluorescence levels of the fraction II-treated group were like those that had been treated with Triton X-100.
Using fluorescence digital microscopy, it was also possible to corroborate the spectrofluorimetric data, by taking images of the parasites after incubation with FII (Figure 8). In the FII-treated group, the parasites presented an altered morphology, with a round shape and an intense green fluorescence in the cytoplasm, caused by penetration of the vital SYTOX Green dye. The untreated group showed no fluorescence, and the parasites demonstrated a normal elongated morphology (Figure 8). The positive control with Triton X-100 demonstrated an intense alteration of the morphology with an elevated green fluorescence.
3. Discussion
Chagas disease is a serious public health problem in 21 developing countries. In Brazil, only one drug is approved for the treatment, with a limited efficacy and high toxicity. In 2015, Morillo and co-workers published the most complete multicentric and randomized clinical study with benznidazole, involving 2,854 patients with Chagas cardiomyopathy [10]. The results showed that the therapy significantly reduced parasitemia, however, showed no efficacy to reduce the cardiac damage over the 5-year follow-up of the study [10]. In the literature there is a consensus about the urgent need for new therapeutic candidates for Chagas disease.
Considering the huge chemodiversity found in marine microorganisms, we investigated for the first time the anti-Trypanosoma cruzi potential of metabolites produced by bacteria that had been previously isolated from invertebrates and sediments from Sao Paulo north coast in Brazil. Among the most active extracts, it was observed that metabolites of the bacterium Vibrio harveyi were amongst the most potent.
This strain was isolated from four sources, one from the coral Mussismilia hispida and two from the sediments from the São Sebastião Channel and the Buzios Island at 35- and 13-meters m depth. Members of the Vibrionaceae family are freely in tropical marine waters or in the microbiota of marine animals [15]. The genus Vibrio was the most common one found in this study, covering 7 of the 11 identified strains and it has been recognized as part of the microbiome of several invertebrates. It is a holobiont that participates in nitrogen fixation, food resource, chitin decomposition, and the production of antimicrobial metabolites ([16,17]).
The Vibrionaceae family has yielded 93 metabolites with different biological activities, including antifungal, antibacterial and anticancer. Most have been isolated from three species, namely V. parahaemolyticus, V. anguillarum, and V. vulnificus. However, only one compound, prodigiosin, presented antiprotozoal activity against Plasmodium spp. [18]
To our knowledge, our study demonstrated for the first time, the anti-trypanosomal activity of metabolites from Vibrio spp. It was noteworthy that the same species, but collected in different locations, resulted in different potencies against the parasites. This was verified with V. harveyi, which resulted in metabolites with IC50 values against T. cruzi ranging from 8 to 51 µg/mL. These differences in the metabolic profile of genetically similar strains have been reported for Streptomyces griseus, the most prolific genus in the production of bioactive compounds [19]. Species closely associated by 16S rRNA maintain a common set of chemical compounds. However, despite the identical genetic sequences of this gene, a noticeable difference was found in the accessory set of accumulation of metabolites, which are unique for each bacterium [19]. Sottorff and colleagues suggested that, based on phylogenetic proximity and the similarity of metabolites, both strains of Streptomyces spp. had a common origin that underwent posterior specialization as a function of their habitat [19]. In our studies, the Vibrio spp. strains were isolated from completely different environments, which included deep sediments (close to the coast), sediments from shallow waters of an island (approximately 20 miles from the coast) and other three coral species. It is possible that the different microbiomes may have contributed to the process of specialization of their metabolites.
Another potent extract with anti-T. cruzi activity was produced from Shewanella pneumatophori, isolated from marine sediments of the Buzios island. This bacterium belongs to a genus generally found in extreme temperatures and has shown antimicrobial [20] and antifungal activities [21,22,23]. Our study demonstrated, for the first time in the literature, the anti-T. cruzi activity of metabolites of this genus.
The genus Bacillus has already shown promising activities in the literature, ranging from antimicrobial activities e.g. B. subtilis [24], to antiparasitic activities e.g. B. pulmilus against Trypanosoma cruzi [12]. In our study, Bacillus megaterium, isolated from deep sediments of the São Sebastião Canal at 35 meters depth, showed anti-T. cruzi activity, but presented the lowest potency among all tested extracts. Generally found in terrestrial environments, this species has the capacity to solubilize natural phosphates in soil and are of great interest for the industry of bioproducts [25,26]. The organisms have also shown antimicrobial and antifungal activities, with potential use against dermatophytosis and in aquaculture for biocontrol of Aspergillus flavus [27,28,29].
Our study is also the first to describe the anti-T. cruzi potential of Halomonas aquamarina metabolites. The extract of H. aquamarina showed high potency activity against T. cruzi, amongst all the tested bacteria. The Halomonas genus has a wide geographical distribution and promising antimicrobial and antitumor biological activities [30,31]. Halomonas is a common genus of halophilic bacteria, which are inhabitants of hypersaline environments. These extremophiles have special abilities to produce extremozymes and other bioactive molecules as potential antibiotics [32].
Another potent microbial extract against T. cruzi was obtained from Mesoflavibacter zeaxanthinifaciens, a bacterium isolated from marine sediments of Buzios island. This is an aerobic Gram-negative, rod-shaped, halo- and mesophilic bacterium, first isolated from a seawater sample collected from the Pacific coastline of Japan [14]. Considering the antiparasitic potential of these metabolites, we conducted an OSMAC (One-Strain Many Compounds) study, a promising approach to activate cryptic genes and enhance the production of bioactive compounds. Using solid and liquid mediums for cultivation, our results showed that the cultivation in liquid medium with rotation, resulted in a higher mass of microbial metabolites. However, the potency of these anti-T. cruzi metabolites decreased 8-fold when compared to those produced by bacteria in solid medium (Marine Agar, Difco). English and co-workers used a similar OSMAC approach to test the ability of Streptomyces sp. to produce antibiotics against S. aureus. No antimicrobial activity was detected in metabolites isolated from bacteria cultured in liquid medium, but only in agar medium [33]. Similarly, Guo and co-workers also observed that the marine fungi Penicillium sp. F23-2 showed different chemical compounds when cultivated in solid medium, leading to the isolation of five new analogues of ambuic acid (penicyclones A-E), which antibacterial activity [34].
Using solid-phase extraction, we conducted a fractionation of the crude EtOAc extract from Mesoflavibacter zeaxanthinifaciens, resulting in FII, a fraction with trypanocidal effect, as confirmed by the lack of mitochondrial activity, detected by the resazurin assay. The studies with the intracellular amastigotes revealed a potent activity of FII against the clinically relevant form of the parasite. Despite the importance of the trypomastigotes during the acute phase of the disease, the amastigotes are the major parasitic form living inside the cells, contributing to the prevalence of the disease for decades [35]. It was noteworthy that FII eliminated both forms of the parasite without affecting the mammalian cells (up to a tested concentration of 200 μg/mL), as demonstrated by the lack of cytotoxicity against fibroblasts. Additionally, FII was also evaluated in erythrocytes for cytotoxicity, but resulted in a low hemolysis at the highest tested concentration. These studies confirmed that FII presented selective compounds against T. cruzi, suggesting a promising biological activity for drug discovery exploration.
Using 1H NMR and ESI-HRMS analysis, four iso fatty acids (1: C19H37O2, 2: C20H39O2, 3: C21H41O2, and 4: C22H43O2) were identified as main compounds in FII. Our study corroborated the known predominance of branched saturated fatty acids, in addition to branched monounsaturated and branched hydroxyacids in specimens of the family Flavobacteriaceae [14,36]. In the literature, studies about the antiparasitic activity of saturated fatty acids are limited. Londero and co-workers showed acetylenic fatty acids can change the membrane potential and kill T. cruzi [37]. Other fatty acids, with antimalarial, antimycobacterial and anfungal properties have been reported. They might be able to cause pores in the bacterial membrane and alter cell permeability, activating signaling pathways that lead to the microorganism to death [38].
Considering the promising results of fraction II of Mesoflavibacter zeaxanthinifaciens EtOAc extract against T. cruzi, we investigated the mechanism of action in the parasite. Using MALDI-ToF/MS, our studies demonstrated that FII induced significant dose-dependent alterations of the protein profile of trypomastigotes after treatment, reducing the intensities of protein signals when compared to untreated parasites, similarly to the standard drug benznidazole. This effect has also been shown for 6-brome-2’-de-N-methylaplysinopsin, a marine alkaloid isolated from the coral Tubastrea tagusensis with potent activity against T. cruzi [39].
The T. cruzi plasma membrane acts as a physical barrier between the external environment and the inner cell organelles, allowing the change of molecules, maintenance of the cell potential, and plays a vital role in the shape of the cell [40]. In our study, the plasma membrane of trypomastigotes was rapidly permeabilized in the presence of fraction II and this was accompanied by the loss of the elongated shape of trypomastigotes to an amastigote-like form. It is known that permeabilization of plasma membrane can contribute to the leakage of intracellular components leading to the death of the cell. For example, amphotericin B, an antifungal approved drug used for the treatment of Leishmaniasis, alters the permeability of the parasite, causing pores in the membrane of the parasite [41]. It is possible that a similar mechanism occurs in T. cruzi treated with FII.
4. Materials and Methods
4.1. Mice
BALB/c mice were obtained from the animal breeding facility at the Instituto Adolfo Lutz, Brazil. The animals were maintained in sterilized cages under a controlled environment and received water and food ad libitum. All procedures were approved by the Animal Care and Use Committee from Instituto Adolfo Lutz – Secretary of Health of Sao Paulo State (Project number CEUA 05/2018) in agreement with the Guide for the Care and Use of Laboratory Animals from the National Academy of Sciences.
4.2. Parasites and mammalian cell maintenance
Macrophages were collected from the peritoneal cavity of BALB/c mice by washing the cavities with RPMI-1640 medium (Sigma-Aldrich, USA) supplemented with 10% (v/v) fetal calf serum (Sigma-Aldrich, USA) and were maintained at 37ºC in a 5% (v/v) CO2 humidified incubator. Trypomastigotes of T. cruzi (Y strain) were maintained in Rhesus monkey kidney cells (LLC-MK2 - ATCC CCL 7), cultivated in RPMI-1640 medium supplemented with 2% (v/v) fetal calf serum at 37ºC in a 5% (v/v) CO2 humidified incubator. The murine conjunctival cells (NCTC clone 929, ATCC) were maintained in RPMI-1640 supplemented with 10% (v/v) FBS at 37ºC in a 5% (v/v) CO2 humidified incubator.
4.3. Analytical methods
1H NMR spectra (500 MHz) were recorded on a Varian INOVA 500 spectrometer using CD3OD (Sigma-Aldrich) as the solvent and internal standard. ESI-HRMS spectra were obtained with electrospray ionization in the positive ion mode on a Bruker Daltonics MicroTOF QII spectrometer. The MALDI-TOF/MS data was analyzed on a Bruker MicroFlex spectrometer at a 20 kV accelerating voltage, in the positive mode (500 laser shots). The signals were collected in a range between m/z 2,.000 -20,.000 with the AutoXecute tool (Bruker-Daltonics).
4.4. Collection of marine corals and sediments
The collections were made by scuba diving in the São Sebastião region at CEBIMar-USP at depths ranging from 5 to 35 meters. The marine invertebrates were identified by Dr. Álvaro E. Migotto (ICMBio MMA 10186-2). The sea sediments were collected with the help of a dredge in the São Sebastião Channel (at 35 meters depth) (S 23°49’40.7" W045°24’44.7"), as well as on Buzios Island, by scuba diving (13 meters deep) (23°48’05.2"; W 45°08’;32.2").
4.5. Isolation and cultivation of microorganisms
After collection, the samples were immediately processed at the Centro de Biologia Marinha (CEBIMar), Universidade de São Paulo, under sterile conditions and the contents of the corals and sediment samples were inoculated on to 90x15 mm Petri dishes in a BOD incubator (SolidSteel, Brazil) at 25ºC, using Marine in an agar medium rich in nutrients and specific for heterotrophic marine bacteria (Marine Agar (Difco, USA). From this material, several isolates were obtained. The isolates were stored in a liquid medium specific for heterotrophic marine bacteria (Marine Broth, Difco, USA) containing 15% (v/v) DMSO (Merck, USA) at -85°C. Gram staining of bacterial isolates was done and the bacteria morphology was observed [42].
4.6. Bacteria Identification by Mass Spectrometry (MALDI-ToF/MS)
MALDI-TOF/MS (Matrix Associated Laser Desorption-Ionization - Time of Flight) analysis was used to identify the microorganisms. The samples were extracted in 1.5 mL tubes or directly on the plate. For tube extraction, up to five colonies were suspended in 300 µL of ultrapure water (Milli-Q) and mixed by vortex. Next, 900 µL of EtOH was added, vortex mixed, and centrifuged (Eppendorf, USA) at maximum speed for 1 minute. After elimination of EtOH, 50 µL of 70% (v/v) formic acid and 50 µL of acetonitrile (100%) were added to the pellet. The tube was centrifuged for 2 min and the supernatant was dispensed in duplicates (1 µL) into a 96-well plate for MALDI-TOF/MS and air-dried. The matrix, α-cyano-4-hydroxy-cinnamic acid (HCCA) (Bruker-Daltonics), was prepared at a concentration of 50 mg/mL in 50% (v/v) acetonitrile and 50% (v/v) water with 2.5% (v/v) trifluoroacetic acid and 1 µL of the matrix was added to the dried sample. In the direct plate extraction, a few colonies were transferred to the 96-well plate and covered with the matrix as described above [43]. A DH5-α protein extract from Escherichia coli (Bruker-Daltonics) was added to the plate as an external control. The analyses were performed on the MALDI-TOF/MS Microflex mass spectrometer (Bruker-Daltonics) with a nitrogen laser (337 nm) operating in linear mode with delayed extraction (260 ns) at 20 kV accelerating voltage. Each spectrum was automatically collected in positive-ion mode as an average of 500 laser shots (50 laser shots at 10 different point positions). A mass range between 2,000 and 20,000 m/z (mass-to-charge ratio) was selected to collect the signals with the Auto Xecute tool of the flexcontrol acquisition software (Version 2.4; Bruker-Daltonics). Only peaks with a signal-to-noise ratio were considered. The software utilizes a logarithmically transformed similarity score that is computed by evaluating several factors. Log(score) values above 2 indicate species-level identification of bacteria. Scores between 1.7 and 2.0 suggest identification of microorganisms at the genus level. Scores below 1.7 signify that the spectrum cannot be effectively identified using the MALDI Biotyper method ([43]
4.7. Bacterial identification by sequencing of the partial 16S rRNA gene
Some strains were not identified by MALDI- ToF/MS method and were selected for genetic sequencing. Genomic DNA from the bacterial culture was isolated using the Wizard® Genomic DNA Purification Kit, with some modifications. In this modified protocol we increased the incubation period with Proteinase K (2 hours) and used a higher volume of RNase (10 ul). The partial amplification of the 16S rRNA gene was performed with the GoTaq Master Mix kit (Promega) and universal primers for eubacteria [44]. The PCR products were purified using the ExoSAP-IT (Thermo Fisher Scientific) and sequencing reactions were prepared using the BigDye® Terminator v3.1 Cycle Sequencing Kit (Thermo Fisher Scientific). A precipitation reaction with ethanol-sodium acetate was performed and the products were sequenced using the ABI 3730 DNA Analyzer, a 48-capillary analysis system (Applied Biosystems). Sequences were analyzed using BioNumerics v.8.0 software (Applied Maths, BioMerieux) in comparison with the sequences present in the Ezbiocloud and Microbenet databases for species database and also in NCBI (http://www.ncbi.nlm.nih.gov/) using the Basic Local Alignment Search Tool (BLAST) and the 16S ribosomal RNA sequences database.
4.8. Extraction of metabolites from marine bacteria grown in agar and broth medium
4.8.1. Agar
The bacterial strains were seeded in Petri dishes (140 x 15 mm) and grown for 120 h at 25°C. Colonies were scraped with a cell scraper (Corning) and diluted in ultrapure water (Milli-Q) in a glass vial. After vortex mixing for 2 min and sonication in an ultrasonic bath for 10 min, 200 mL of EtOAc (JT Baker) were added and the solution was transferred to the ultrasonic bath for 40 min. After partitioning, the organic phase was filtered, and the solvent was removed under reduced pressure at 40°C to afford the crude EtOAc extract.
4.8.2. Broth
Colonies grown in Marine Agar (Difco) were transferred to 20 mL of liquid medium Marine Broth and cultured for 24h in a BOD incubator (SolidSteel, Brazil) at 25ºC. The optical density of this culture was measured and adjusted to 0.5 on the McFarland scale, i.e. 1.5x108 CFU/mL. The culture was kept at 25°C in an incubator (SolidSteel, Brazil) with an orbital shaker or static for a total of 120h. After this period, the cultures were transferred to 50 mL plastic conical tubes and centrifuged at 4000g for 20 min. The cell precipitate was separated from the supernatant and concentrated in 100 mL ultrapure water (Milli-Q). After vortex mixing for 2 min and sonication in an ultrasonic bath for 10 min, 100 mL EtOAc was added to this aqueous phase. The extraction procedure was then followed.
4.9. Bio-Guided Fractionation of Mesoflavibacter zeaxanthinifaciens organic extract
Fractionation of EtOAc extract of Mesoflavibacter zeaxanthinifaciens (500 mg) was performed using a solid phase extraction cartridge (SPE Bond Elute C18 - Agilent). The cartridge was activated with MeOH, conditioned with MeOH:H2O (9:1) and after application of the crude EtOAc extract (500 mg), four fractions (F0 – 158,2 mg, FI– 11,9 mg, FII 10,2 and FIII – 161,6 mg) were collected using different mixtures of MeOH:H2O (7:3, 4:6, 9:13:2 and pure MeOH, respectively) as eluents. Fractions were dried over reduced pressure and subjected to evaluation of anti-T. cruzi activity.
4.10. Evaluation of the anti-Trypanosoma cruzi activity
LLC-MK2-derived trypomastigotes were seeded (1 x 106 cells/well) in 96-well plates and incubated with the extracts/compounds serially diluted 2-fold (200 to 1.5 µg/mL), during 24h in RPMI 1640 medium at 37ºC in a 5% (v/v) CO2 humidified incubator. Subsequently, resazurin (0.011% (w/v) in PBS) was added to check for parasite viability for 24h. The optical density was determined in the FilterMax F5 (Molecular Devices) at λ570 nm. Benznidazole was used as the standard drug [45].
For anti-amastigote assay, mouse peritoneal macrophages (1 x 105 cells/well, were seeded in 16-well chamber slide (NUNC, Thermo Fisher Scientific) and infected with trypomastigotes (10:1 parasite:macrophage ratio). After 2h, the extracts / compounds were diluted in different concentrations (200 to 1.5 µg/mL) and incubated with infected macrophages for 48h at 37ºC in a 5% (v/v) CO2 humidified incubator. Finally, slides were fixed with methanol (100%), stained with Giemsa and observed under a light microscope (EVOS M5000, Termo, USA) with digital image acquirement. The 50% inhibitory concentration (IC50) values were determined by the infection index. Benznidazole was used as standard drug [39,45]
4.11. Cytotoxicity against mammalian cells
Fibroblast NCTC cells (clone 929) (6 x 104 cells/well) were seeded in 96-well plates and incubated with the extracts / compounds (1.5 to 200 µg/mL) for 48h at 37ºC in a 5% (v/v) CO2 humidified incubator. The 50% cytotoxic concentration (CC50) was determined by the MTT colorimetric assay, as described previously [39].
4.12. Hemolytic activity
Whole blood was collected from male BALB/c (25 g) mice after euthanasia. Erythrocytes were collected by centrifugation and used to prepare a 3% (v/v) solution in PBS. In a round-bottomed titration microplate, the fraction obtained from active extract from Mesoflavibacter zeaxanthinifaciens (FII), was dispensed in serial dilutions of concentrations from 200 to 1.5 µg/mL. Then the suspension of erythrocytes (100 µL/well) was incubated for 2 hours at 24°C and incubated for 2h at 25ºC in a BOD incubator (Solidsteel, Brazil). The supernatant was collected, and the optical density was determined at λ570 nm (FilterMax F5 Multi-Mode Microplate Reader, Molecular Devices). The maximum hemolysis was obtained using hemocytes suspended in ultrapure distilled water pure distilled water (Milli-Q) (positive control) and untreated hemocytes as the negative control [46].
4.13. Evaluation of T. cruzi trypomastigotes protein profile
Trypomastigotes (1x107 /well) were treated with FII fraction from Mesoflavibacter zeaxanthinifaciens (25 µg/mL and 50 µg/mL) or benznidazole (40 µM) for 24 hours in RPMI medium. After this period, the sample suspensions from the expanded cultures were centrifuged, the supernatant removed, and the precipitate was washed twice in Milli-Q water. The precipitate was resuspended in 300 µL of Milli-Q water before adding 900 µL of 70% EtOH. After further centrifugation, 20 µL of 70% formic acid and 20 µL of acetonitrile were added to the precipitate and the solution was vortexed and centrifuged. Each centrifugation step was performed at 10,000 g for 10 min at room temperature. Untreated parasites were used as control.
The supernatant was dispensed (3 µL) in duplicates into a 96-well steel plate for MALDI-ToF/MS (Bruker-Daltonics) and dried at room temperature. The matrix, α-cyano-4- hydroxy-cinnamic acid (HCCA) (Bruker-Daltonics), was prepared at a concentration of 50 mg/mL in 50% acetonitrile and 50% H2O with 2.5% TFA, and was added (1 µL) on the plate. A DH5-alpha protein extract from Escherichia coli (Bruker-Daltonics) was added to the plate for external control. The analyses were performed on a Bruker-Daltonics Microflex MALDI-ToF/MS mass spectrometer with a nitrogen laser (337 nm) operating in linear mode with delayed extraction (260 ns) at 20 kV accelerating voltage. Each spectrum was automatically collected in positive ion mode as an average of 500 laser shots (50 laser shots at 10 different point positions). A mass range between 3,000 and 20,000 m/z (mass-to-charge ratio) was selected to collect the signals with the Auto Xecute tool of the FlexControl acquisition software (Version 2.4; Bruker-Daltonics). Only peaks with signal-to-noise ratio were considered [47,48].
4.14. Evaluation of Plasma Membrane Permeability
Sytox® Green, a fluorescent nucleic acid marker, impermeable to viable cells, was used to evaluate possible changes in the permeability of the plasma membrane. T. cruzi
trypomastigotes were added to 96-well black microtiter plates (2x106 parasites/well) and incubated with Sytox® Green (1 µM) in HBSS medium supplemented with NaHCO3 (4.2 mM) and D-glucose (10 mM). Subsequently, a basal reading of the plate was done, and the FII fraction from Mesoflavibacter zeaxanthinifaciens was added (t=0) at IC50 value and the plate was incubated at 24ºC for 15 min [48]. The fluorescence signal was monitored every 20 min for 180 min. Measurements were made in a spectrofluorometer (FilterMax F5 Multi-Mode Microplate Reader) with excitation filters of λ485 nm and emission filters of λ535 nm. The maximum permeabilization was obtained in the presence of 0.5% (v/v) Triton X-100, and untreated parasites were used as negative control (100% viability, integral membrane) [48]. Digital Images of the trypomastigotes after treatment with FII fraction from Mesoflavibacter zeaxanthinifaciens were obtained in a digital fluorescence microscope (EVOS M 5000, Thermo, USA) using SYTOX Green [49].
4.15. Statistical Analysis
IC50 and CC50 values were calculated from sigmoidal dose-response curves. Unless otherwise stated, the data reported were the mean ± standard error of at least two independent experiments performed with duplicate samples. For hemolytic activity, One-way ANOVA with Tukey’s Multiple Comparison test was applied for significance (p value< 0.05) using GraphPad Prism 6.0 software. The samples were tested in duplicate/triplicate and the experiments were repeated at least twice.
5. Conclusions
These findings demonstrate for the first time the anti-trypanosomal potential of microbial metabolites from six genera of marine bacteria collected from corals and sediments from the Blue Amazon. Saturated iso-type fatty acids, identified as main compounds from bioactive fraction II of Mesoflavibacter zeaxanthinifaciens EtOAc extract (FII) showed a promising potential against trypomastigotes and the intracellular amastigotes of the parasite, causing a trypanocidal effect by altering the permeability of the membrane, without affecting mammalian cells. These marine bacteria can be considered promising sources for the selection of new hit compounds against Chagas disease.
Author Contributions
Author contributions were as follows: Conceptualization, diving, funding acquisition, methodology, data curation, supervision and coordination AGT.; Collection of marine material: AGT.; Identification of marine corals: A.E.M; Investigation, biological assays, data analysis, original manuscript writing and editing: D.A.S.F; methodology, anti-trypanossomal assays development: EVLC; 16S rRNA sequencing: CHC and AYY; methodology and structural identification of compounds: JHGL; ESI-HRMS studies: LMSC and ARC; data evaluation and discussion, manuscript review: MC. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the São Paulo Research State Foundation (FAPESP, Projects 2021/04464-8, 2017/50333-7, 2023/12447-1) and Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq 405691/2021-1). The authors AGT and JHGL are thankful to CNPq for the scientific research awards.
Institutional Review Board Statement
Animal Care and Use Committee from Instituto Adolfo Lutz – Secretary of Health of Sao Paulo State (Project number CEUA 05/2018) in agreement with the Guide for the Care and Use of Laboratory Animals from the National Academy of Sciences.
Data Availability Statement
The 16S rRNA data presented in this study are openly available in GeneBank (NCBI), reference number OP163900, OP163959, OR479885 and OP163958. The raw data supporting the conclusions of this article will be made available by the authors on request.
Acknowledgments
We also acknowledge CAPES for DASF scholarship. This work was developed with members of the Research Network Natural Products against Neglected Diseases (ResNetNPND): http://www.uni-muenster.de/ResNetNPND/.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Santos, J.D.; Vitorino, I.; Reyes, F.; Vicente, F.; Lage, O.M. From Ocean to Medicine: Pharmaceutical Applications of Metabolites from Marine Bacteria. Antibiotics 2020, 9, 1–30. [Google Scholar] [CrossRef] [PubMed]
- Atanasov, A.G.; Zotchev, S.B.; Dirsch, V.M.; Orhan, I.E.; Banach, M.; Rollinger, J.M.; Barreca, D.; Weckwerth, W.; Bauer, R.; Bayer, E.A.; et al. Natural Products in Drug Discovery: Advances and Opportunities. Nature Reviews Drug Discovery 2021 20:3 2021, 20, 200–216. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.; Buddie, A.G.; Goss, R.J.M.; Overmann, J.; Lepleux, C.; Brönstrup, M.; Kloareg, B.; Meiners, T.; Brennecke, P.; Ianora, A.; et al. Discovery Pipelines for Marine Resources: An Ocean of Opportunity for Biotechnology? World J Microbiol Biotechnol 2019, 35. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; She, J.; Fu, J.; Wang, J.; Ye, Y.; Yang, B.; Liu, Y.; Zhou, X.; Tao, H. Advances in Natural Products from the Marine-Sponge-Associated Microorganisms with Antimicrobial Activity in the Last Decade. Mar Drugs 2023, 21. [Google Scholar] [CrossRef] [PubMed]
- Fenical, W.; Jensen, P.R. Developing a New Resource for Drug Discovery: Marine Actinomycete Bacteria. Nat Chem Biol 2006, 2, 666–673. [Google Scholar] [CrossRef] [PubMed]
- Romano, S.; Jackson, S.A.; Patry, S.; Dobson, A.D.W. Extending the “One Strain Many Compounds” (OSMAC) Principle to Marine Microorganisms. Mar Drugs 2018, 16, 1–29. [Google Scholar] [CrossRef] [PubMed]
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs over the Nearly Four Decades from 01/1981 to 09/2019. J Nat Prod 2020, 83, 770–803. [Google Scholar] [CrossRef]
- Schmunis, G.A.; Yadon, Z.E. Chagas Disease: A Latin American Health Problem Becoming a World Health Problem. Acta Trop 2010, 115, 14–21. [Google Scholar] [CrossRef]
- Moreno, A.M.H. Mecanismos de Transmissão Da Doença de Chagas. Available online: http://chagas.fiocruz.br/doenca/transmissao/#controle (accessed on 1 May 2022).
- Morillo, C.A.; Marin-Neto, J.A.; Avezum, A.; Sosa-Estani, S.; Rassi, A.; Rosas, F.; Villena, E.; Quiroz, R.; Bonilla, R.; Britto, C.; et al. Randomized Trial of Benznidazole for Chronic Chagas’ Cardiomyopathy. New England Journal of Medicine 2015, 373, 1295–1306. [Google Scholar] [CrossRef]
- Gerwick, W.H.; Fenner, A.M. Drug Discovery from Marine Microbes. Microb Ecol 2013, 65, 800–806. [Google Scholar] [CrossRef]
- Martínez-Luis, S.; Gómez, J.F.; Spadafora, C.; Guzmán, H.M.; Gutiérrez, M. Antitrypanosomal Alkaloids from the Marine Bacterium Bacillus Pumilus. Molecules 2012, Vol. 17, Pages 11146-11155 2012, 17, 11146–11155. [Google Scholar] [CrossRef] [PubMed]
- Osei, E.; Kwain, S.; Mawuli, G.T.; Anang, A.K.; Owusu, K.B.A.; Camas, M.; Camas, A.S.; Ohashi, M.; Alexandru-Crivac, C.N.; Deng, H.; et al. Paenidigyamycin A, Potent Antiparasitic Imidazole Alkaloid from the Ghanaian Paenibacillus Sp. DE2SH. Marine Drugs 2019, Vol. 17, Page 9 2018, 17, 9. [Google Scholar] [CrossRef]
- Asker, D.; Beppu, T.; Ueda, K. Mesoflavibacter Zeaxanthinifaciens Gen. Nov., Sp. Nov., a Novel Zeaxanthin-Producing Marine Bacterium of the Family Flavobacteriaceae. Syst Appl Microbiol 2007, 30, 291–296. [Google Scholar] [CrossRef] [PubMed]
- Austin, B.; Zhang, X.H. Vibrio Harveyi: A Significant Pathogen of Marine Vertebrates and Invertebrates. Lett Appl Microbiol 2006, 43, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Chimetto, L.A.; Brocchi, M.; Thompson, C.C.; Martins, R.C.R.; Ramos, H.R.; Thompson, F.L. Vibrios Dominate as Culturable Nitrogen-Fixing Bacteria of the Brazilian Coral Mussismilia Hispida. Syst Appl Microbiol 2008, 31, 312–319. [Google Scholar] [CrossRef] [PubMed]
- Mustapha, S.; Mustapha, E.; Nozha, C. Vibrio Alginolyticus: An Emerging Pathogen of Foodborne Diseases. Int. J. Sci. Technol 2013, 2, 302–309. [Google Scholar]
- Mansson, M.; Gram, L.; Larsen, T.O. Production of Bioactive Secondary Metabolites by Marine Vibrionaceae. Marine Drugs 2011, Vol. 9, Pages 1440-1468 2011, 9, 1440–1468. [Google Scholar] [CrossRef] [PubMed]
- Sottorff, I.; Wiese, J.; Lipfert, M.; Preußke, N.; Sönnichsen, F.D.; Imhoff, J.F. Different Secondary Metabolite Profiles of Phylogenetically Almost Identical Streptomyces Griseus Strains Originating from Geographically Remote Locations. Microorganisms 2019, 7. [Google Scholar] [CrossRef]
- Hirota, K.; Nodasaka, Y.; Orikasa, Y.; Okuyama, H.; Yumoto, I. Shewanella Pneumatophori Sp. Nov., an Eicosapentaenoic Acid-Producing Marine Bacterium Isolated from the Intestines of Pacific Mackerel (Pneumatophorus Japonicus). Int J Syst Evol Microbiol 2005, 55, 2355–2359. [Google Scholar] [CrossRef]
- Portela, R.W.S. Isolamento e Caracterização de Cepas Shewanella Sp. Do Cultivo Heterotrófico de Litopenaeus Vannamei (Boone, 1931) Isolamento e Caracterização de Cepas Shewanella sp . Do Cultivo Heterotrófico de Litopenaeus Vannamei (BOONE, 1931), 2014.
- Fitriani, A.; Ihsan, F.; Hamdiyati, Y. Antibacteria Activity of Shewanella and Pseudomonas as Endophytic Bacteria from the Root of Ageratum Conyzoides L. Asian Journal of Applied Sciences 2015, 03, 2321–2893. [Google Scholar]
- Gong, A.D.; Li, H.P.; Shen, L.; Zhang, J.B.; Wu, A.B.; He, W.J.; Yuan, Q.S.; He, J. De; Liao, Y.C. The Shewanella Algae Strain YM8 Produces Volatiles with Strong Inhibition Activity against Aspergillus Pathogens and Aflatoxins. Front Microbiol 2015, 6, 1091. [Google Scholar] [CrossRef] [PubMed]
- Kaspar, F.; Neubauer, P.; Gimpel, M. Bioactive Secondary Metabolites from Bacillus Subtilis: A Comprehensive Review. J Nat Prod 2019, 82, 2038–2053. [Google Scholar] [CrossRef]
- Vary, P.S.; Biedendieck, R.; Fuerch, T.; Meinhardt, F.; Rohde, M.; Deckwer, W.-D.; Jahn, D. Bacillus Megaterium—from Simple Soil Bacterium to Industrial Protein Production Host. Appl Microbiol Biotechnol 2007, 76, 957–967. [Google Scholar] [CrossRef]
- Wyciszkiewicz, M.; Saeid, A.; Chojnacka, K.; Górecki, H. Production of Phosphate Biofertilizers from Bones by Phosphate-Solubilizing Bacteria Bacillus Megaterium. Open Chem 2015, 13, 1063–1070. [Google Scholar] [CrossRef]
- A. Malanicheva, D.G.K. et al Antimicrobial Activity of Bacillus Megaterium Strains. Mikrobiologiya 2012, 81, 196–204.
- Viet Cuong, P.; Thi Kim Cuc, N.; Thi Quyen, V.; Thanh Binh, P.; Van Kiem, P.; Hoai Nam, N.; Tien Dat, N. Antimicrobial Constituents from the Bacillus Megaterium LC Isolated from Marine Sponge Haliclona Oculata. Natural Product Sciences 2014, 20, 202–205. [Google Scholar]
- Kong, Q.; Shan, S.; Liu, Q.; Wang, X.; Yu, F. Biocontrol of Aspergillus Flavus on Peanut Kernels by Use of a Strain of Marine Bacillus Megaterium. Int J Food Microbiol 2010, 139, 31–35. [Google Scholar] [CrossRef]
- Cheffi, M.; Maalej, A.; Mahmoudi, A.; Hentati, D.; Marques, A.M.; Sayadi, S.; Chamkha, M. Lipopeptides Production by a Newly Halomonas Venusta Strain: Characterization and Biotechnological Properties. Bioorg Chem 2021, 109, 104724. [Google Scholar] [CrossRef]
- Wang, L.; Große, T.; Stevens, H.; Brinkhoff, T.; Simon, M.; Liang, L.; Bitzer, J.; Bach, G.; Zeeck, A.; Tokuda, H.; et al. Bioactive Hydroxyphenylpyrrole-Dicarboxylic Acids from a New Marine Halomonas Sp.: Production and Structure Elucidation. Appl Microbiol Biotechnol 2006, 72, 816–822. [Google Scholar] [CrossRef]
- Biswas, J.; Jana, S.K.; Mandal, S. Biotechnological Impacts of Halomonas: A Promising Cell Factory for Industrially Relevant Biomolecules. [CrossRef]
- English, A.L.; Boufridi, A.; Quinn, R.J.; Kurtböke, D.I. Evaluation of Fermentation Conditions Triggering Increased Antibacterial Activity from a Near-Shore Marine Intertidal Environment-Associated Streptomyces Species. Synth Syst Biotechnol 2017, 2, 28–38. [Google Scholar] [CrossRef]
- Guo, W.; Zhang, Z.; Zhu, T.; Gu, Q.; Li, D. Penicyclones A-E, Antibacterial Polyketides from the Deep-Sea-Derived Fungus Penicillium Sp. F23-2. J Nat Prod 2015, 78, 2699–2703. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Molina, J.A.; Molina, I. Chagas Disease. The Lancet 2018, 391, 82–94. [Google Scholar] [CrossRef] [PubMed]
- Nichols, C.M.; Bowman, J.P.; Guezennec, J. Olleya Marilimosa Gen. Nov., Sp. Nov., an Exopolysaccharide-Producing Marine Bacterium from the Family Flavobacteriaceae, Isolated from the Southern Ocean. Int J Syst Evol Microbiol 2005, 55, 1557–1561. [Google Scholar] [CrossRef]
- Londero, V.S.; da Costa-Silva, T.A.; Gomes, K.S.; Ferreira, D.D.; Mesquita, J.T.; Tempone, A.G.; Young, M.C.M.; Jerz, G.; Lago, J.H.G. Acetylenic Fatty Acids from Porcelia Macrocarpa (Annonaceae) against Trypomastigotes of Trypanosoma Cruzi: Effect of Octadec-9-Ynoic Acid in Plasma Membrane Electric Potential. Bioorg Chem 2018, 78, 307–311. [Google Scholar] [CrossRef] [PubMed]
- Carballeira, N.M. New Advances in Fatty Acids as Antimalarial, Antimycobacterial and Antifungal Agents. Prog Lipid Res 2008, 47, 50–61. [Google Scholar] [CrossRef] [PubMed]
- Romanelli, M.M.; Amaral, M.; Thevenard, F.; Santa Cruz, L.M.; Regasini, L.O.; Migotto, A.E.; Lago, J.H.G.; Tempone, A.G. Mitochondrial Imbalance of Trypanosoma Cruzi Induced by the Marine Alkaloid 6-Bromo-2′-de- N-Methylaplysinopsin. ACS Omega 2022, 7, 28561–28570. [Google Scholar] [CrossRef]
- Galhardo, T.S.; Ueno, A.K.; Costa-Silva, T.A.; Tempone, A.G.; Carvalho, W.A.; Fischmeister, C.; Bruneau, C.; Mandelli, D.; Lago, J.H.G. New Derivatives from Dehydrodieugenol B and Its Methyl Ether Displayed High Anti-Trypanosoma Cruzi Activity and Cause Depolarization of the Plasma Membrane and Collapse the Mitochondrial Membrane Potential. Chem Biol Interact 2022, 366, 110129. [Google Scholar] [CrossRef]
- Efimova, S.S.; Malykhina, A.I.; Ostroumova, O.S. Triggering the Amphotericin B Pore-Forming Activity by Phytochemicals. Membranes (Basel) 2023, 13. [Google Scholar] [CrossRef]
- Coico, R. Gram Staining. Curr Protoc Immunol 2001, Appendix 3. [CrossRef]
- Emami, K.; Askari, V.; Ullrich, M.; Mohinudeen, K.; Anil, A.C. Characterization of Bacteria in Ballast Water Using MALDI-TOF Mass Spectrometry. PLoS One 2012, 7, 38515. [Google Scholar] [CrossRef]
- Weisburg, W.G.; Barns, S.M.; Pelletier, D.A.; Lane, D.J. 16S Ribosomal DNA Amplification for Phylogenetic Study. J Bacteriol 1991, 173, 697. [Google Scholar] [CrossRef]
- Lima, M.L.; Romanelli, M.M.; Borborema, S.E.T.; Johns, D.M.; Migotto, A.E.; Lago, J.H.G. Tempone Antitrypanosomal Activity of Isololiolide Isolated from the Marine Hydroid Macrorhynchia Philippina (Cnidaria, Hydrozoa). Bioorg Chem 2019, 89, 103002. [Google Scholar] [CrossRef] [PubMed]
- Amaral, M.; de Sousa, F.S.; Silva, T.A.C.; Junior, A.J.G.; Taniwaki, N.N.; Johns, D.M.; Lago, J.H.G.; Anderson, E.A.; Tempone, A.G. A Semi-Synthetic Neolignan Derivative from Dihydrodieugenol B Selectively Affects the Bioenergetic System of Leishmania Infantum and Inhibits Cell Division. Scientific Reports 2019 9:1 2019, 9, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Varela, M.T.; Amaral, M.; Romanelli, M.M.; de Castro Levatti, E. V.; Tempone, A.G.; Fernandes, J.P.S. Optimization of Physicochemical Properties Is a Strategy to Improve Drug-Likeness Associated with Activity: Novel Active and Selective Compounds against Trypanosoma Cruzi. European Journal of Pharmaceutical Sciences 2022, 171. [Google Scholar] [CrossRef]
- de Castro Levatti, E. V.; Costa-Silva, T.A.; Morais, T.R.; Fernandes, J.P.S.; Lago, J.H.G.; Tempone, A.G. Lethal Action of Licarin A Derivatives in Leishmania (L.) Infantum: Imbalance of Calcium and Bioenergetic Metabolism. Biochimie 2023, 208, 141–150. [Google Scholar] [CrossRef]
- Londero, V.S.; Rosa, M.E.; Baitello, J.B.; Costa-Silva, T.A.; Monteiro Cruz, L.S.; Tempone, A.G.; Caseli, L.; Henrique Lago, J.G. Barbellatanic Acid, a New Antitrypanosomal Pseudo-Disesquiterpenoid Isolated from Nectandra Barbellata, Displayed Interaction with Protozoan Cell Membrane. BBA-Biomembranes 2023, 1865, 184184. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Map comprising the Blue Amazon area of Brazil. Orange arrow indicates the approximate location where samples were collected by scuba diving.
Figure 1.
Map comprising the Blue Amazon area of Brazil. Orange arrow indicates the approximate location where samples were collected by scuba diving.

Figure 2.
Marine invertebrates collected in São Sebastião Channel and Buzios Island, São Paulo, Brazil. TC: Tubastraea coccinea; MH:Mussismilia hispida; MD: Madracis decactis; Photos from Álvaro E. Migotto and Marcelo Visentini Kitahara. Available at The Cifonauta Image Bank (University of São Paulo, http://cifonauta.cebimar.usp.br/).
Figure 2.
Marine invertebrates collected in São Sebastião Channel and Buzios Island, São Paulo, Brazil. TC: Tubastraea coccinea; MH:Mussismilia hispida; MD: Madracis decactis; Photos from Álvaro E. Migotto and Marcelo Visentini Kitahara. Available at The Cifonauta Image Bank (University of São Paulo, http://cifonauta.cebimar.usp.br/).

Figure 3.
High resolution mass spectra of the fraction II of Mesoflavibacter zeaxanthinifaciens EtOAc extract (FII) obtained in spectrometer (Bruker-Daltonics MicroTOF QII), using an electrospray ionization source, operating in negative mode via direct injection.
Figure 3.
High resolution mass spectra of the fraction II of Mesoflavibacter zeaxanthinifaciens EtOAc extract (FII) obtained in spectrometer (Bruker-Daltonics MicroTOF QII), using an electrospray ionization source, operating in negative mode via direct injection.
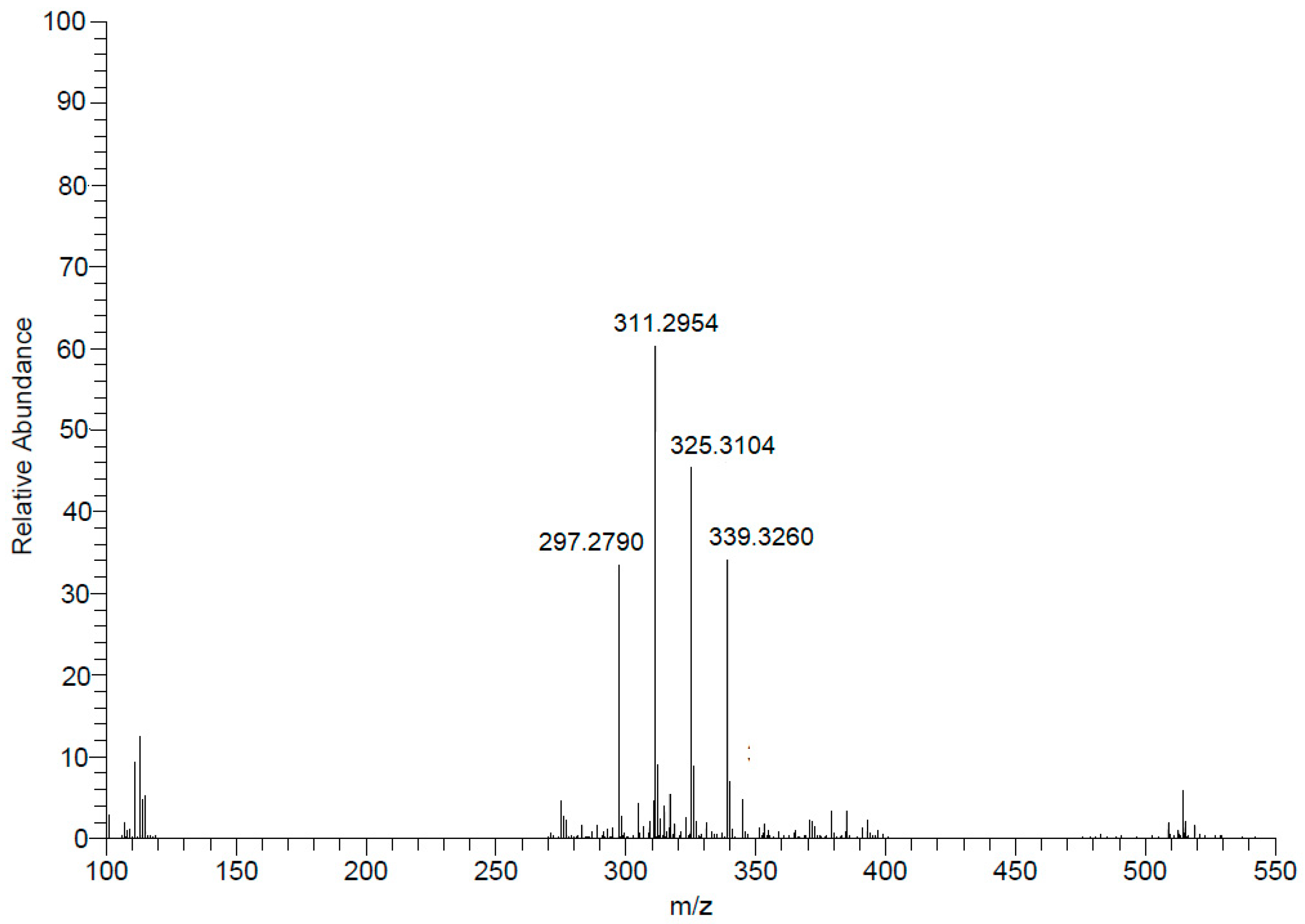
Figure 4.
Molecular structures of the iso chain fatty acids 1 – 4 identified in the bioactive fraction II from Mesoflavibacter zeaxanthinifaciens EtOAc extract.
Figure 4.
Molecular structures of the iso chain fatty acids 1 – 4 identified in the bioactive fraction II from Mesoflavibacter zeaxanthinifaciens EtOAc extract.
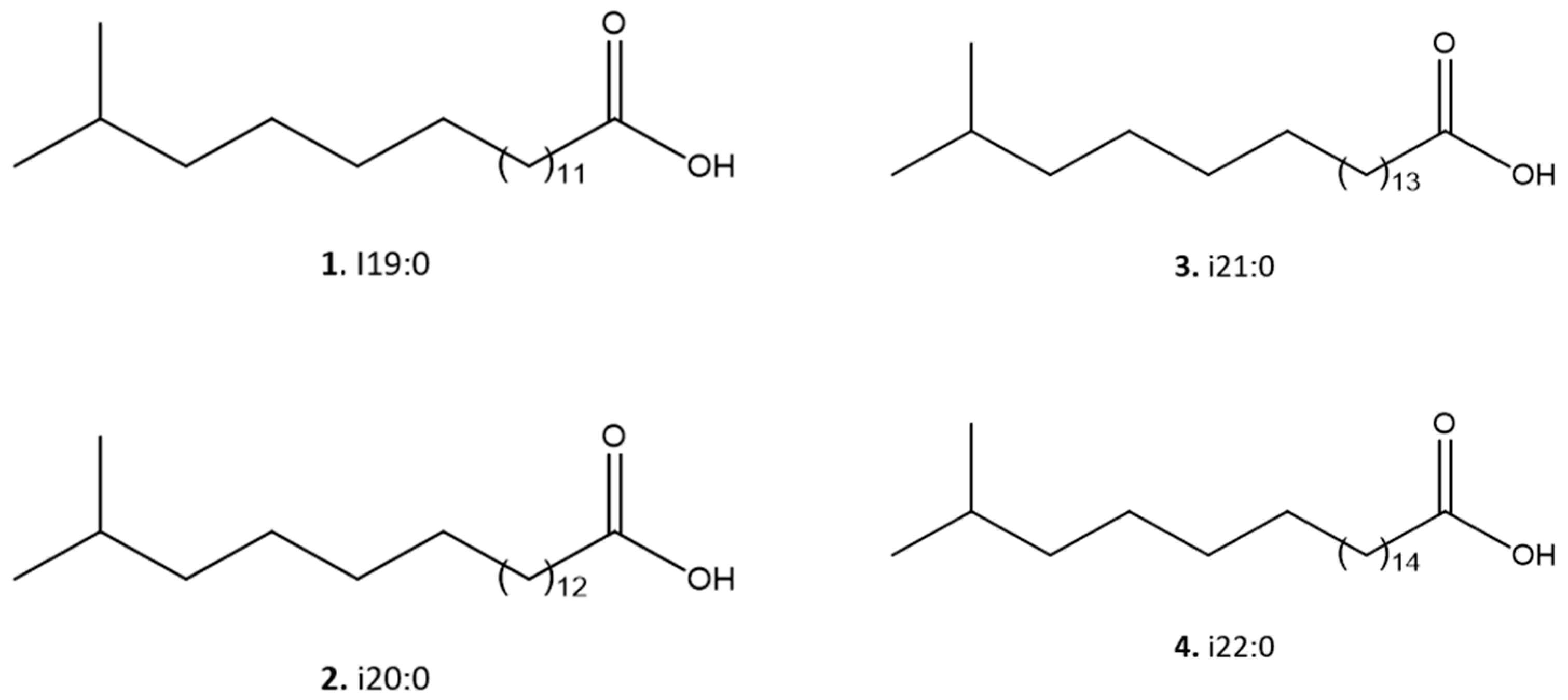
Figure 5.
Hemolytic activity of fraction II of Mesoflavibacter zeaxanthinifaciens EtOAc extract (FII) in murine erythrocytes after 2 h incubation. CT- (negative control); CT+ (positive control). The absorbance was measured in a plate spectrophotometer (λ=570 nm) (FilterMax F5, Molecular Devices).***p <0.005.
Figure 5.
Hemolytic activity of fraction II of Mesoflavibacter zeaxanthinifaciens EtOAc extract (FII) in murine erythrocytes after 2 h incubation. CT- (negative control); CT+ (positive control). The absorbance was measured in a plate spectrophotometer (λ=570 nm) (FilterMax F5, Molecular Devices).***p <0.005.
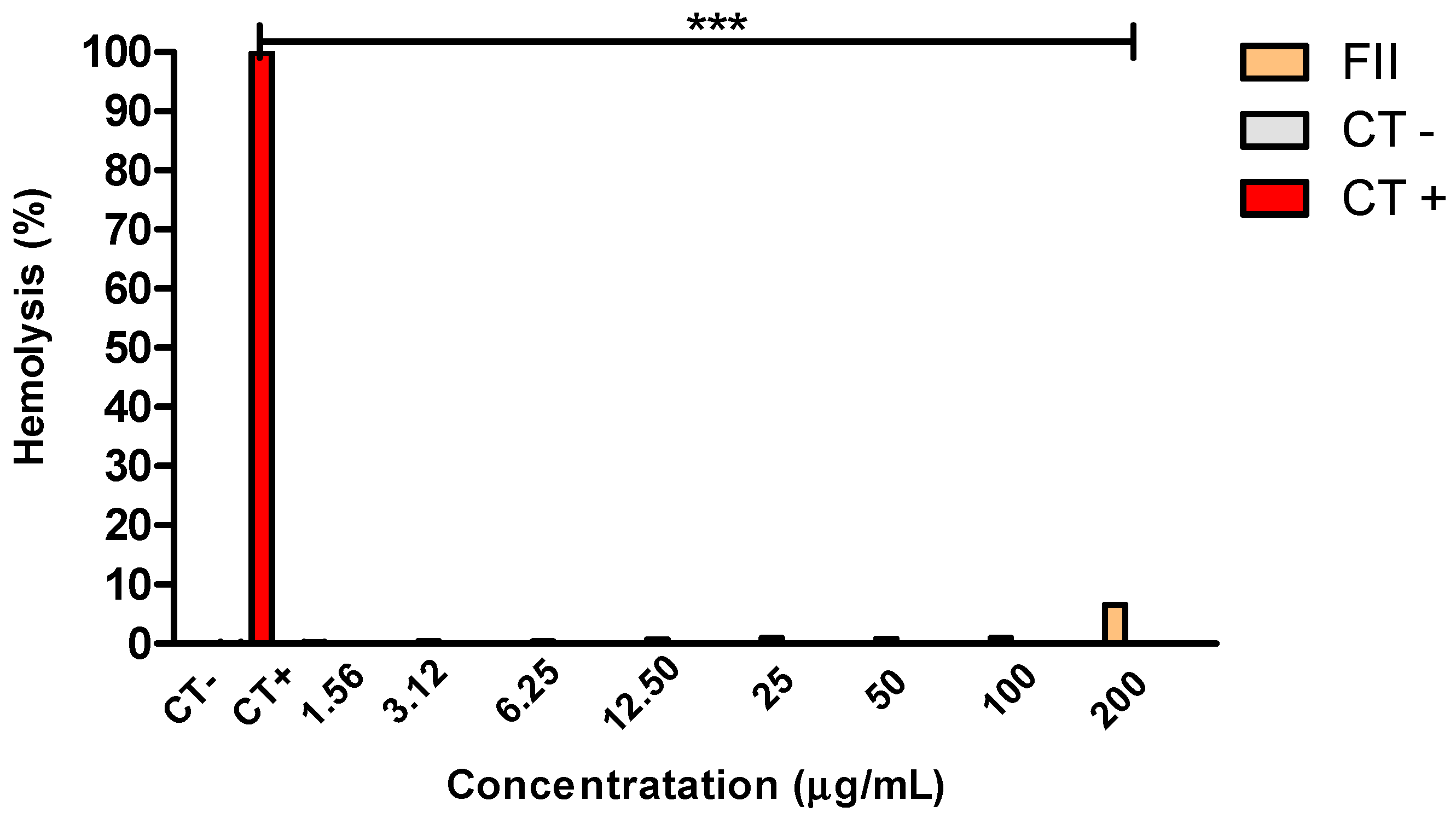
Figure 6.
Evaluation of the protein profile of trypanomastigotes of T. cruzi in the presence of fraction II of Mesoflavibacter zeaxanthinifaciens EtOAc extract (FII) by MALDI-TOF/MS Microflex. Spectrum of mass of trypanomastigotes treated FII (20 and 50 μg/mL), treated with benznidazole (BZN-40 μM) and untreated control (CT). The parasites were treated for 24 h and the protein profile was compared to untreated parasites.
Figure 6.
Evaluation of the protein profile of trypanomastigotes of T. cruzi in the presence of fraction II of Mesoflavibacter zeaxanthinifaciens EtOAc extract (FII) by MALDI-TOF/MS Microflex. Spectrum of mass of trypanomastigotes treated FII (20 and 50 μg/mL), treated with benznidazole (BZN-40 μM) and untreated control (CT). The parasites were treated for 24 h and the protein profile was compared to untreated parasites.
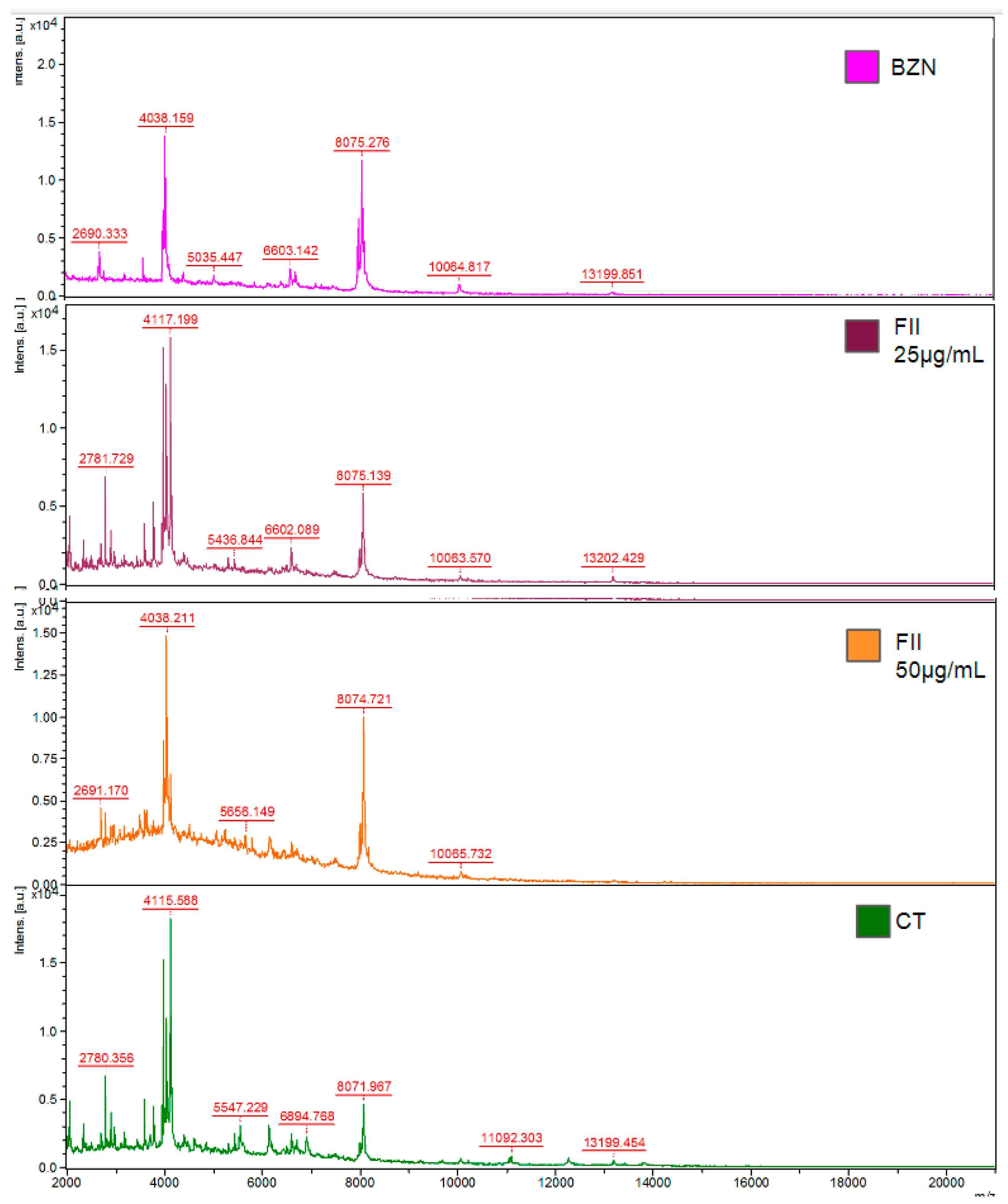
Figure 7.
Evaluation of membrane permeability of trypomastigotes after treatment with fraction II from Mesoflavibacter zeaxanthinifaciens EtOAc extract (FII), using the probe SYTOX Green dye and spectrofluorimeter. Untreated parasites (control group) and treated with 0.5 % Triton X-100 were eavaluated as minimal and maximal permeabilization, respectively. ***P<0.0001; ** p< 0,01.
Figure 7.
Evaluation of membrane permeability of trypomastigotes after treatment with fraction II from Mesoflavibacter zeaxanthinifaciens EtOAc extract (FII), using the probe SYTOX Green dye and spectrofluorimeter. Untreated parasites (control group) and treated with 0.5 % Triton X-100 were eavaluated as minimal and maximal permeabilization, respectively. ***P<0.0001; ** p< 0,01.
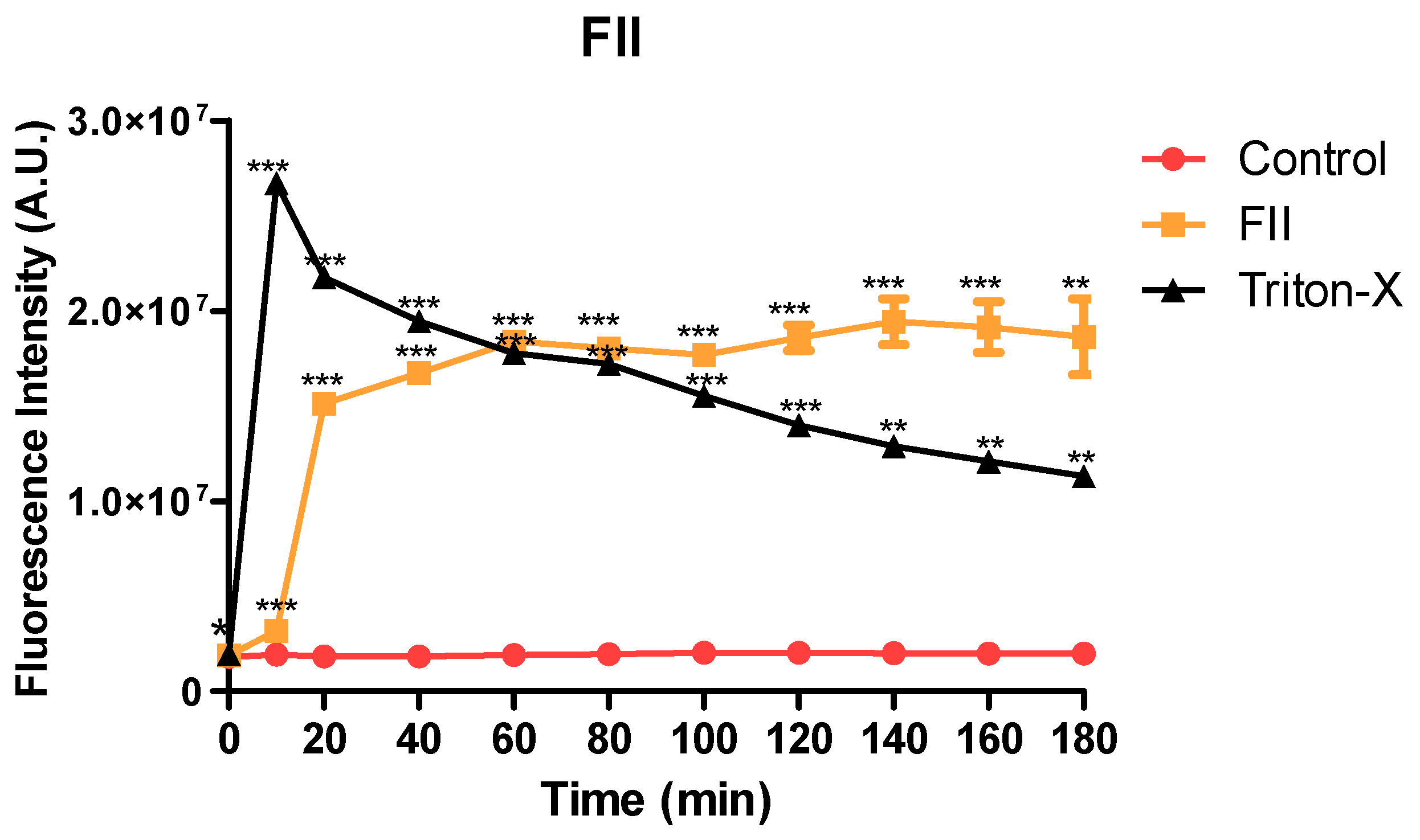
Figure 8.
Digital fluorescence images (100× magnification) obtained after treatment of trypomastigotes with fraction II of Mesoflavibacter zeaxanthinifaciens EtOAc extract (FII) (D, E, F). Black arrows represent elongated forms of T. cruzi (trypomastigotes) and white arrows represent rounded-forms (amastigote-like forms) after treatment. SYTOX Green dye was used to evaluate the permeabilization effect. Untreated parasites were used as a negative control (A,B,C) and parasites treated with 0.5 % Triton X-100 were used as a positive control for maximal permeabilization (G, H, I). A representative experiment is shown.
Figure 8.
Digital fluorescence images (100× magnification) obtained after treatment of trypomastigotes with fraction II of Mesoflavibacter zeaxanthinifaciens EtOAc extract (FII) (D, E, F). Black arrows represent elongated forms of T. cruzi (trypomastigotes) and white arrows represent rounded-forms (amastigote-like forms) after treatment. SYTOX Green dye was used to evaluate the permeabilization effect. Untreated parasites were used as a negative control (A,B,C) and parasites treated with 0.5 % Triton X-100 were used as a positive control for maximal permeabilization (G, H, I). A representative experiment is shown.
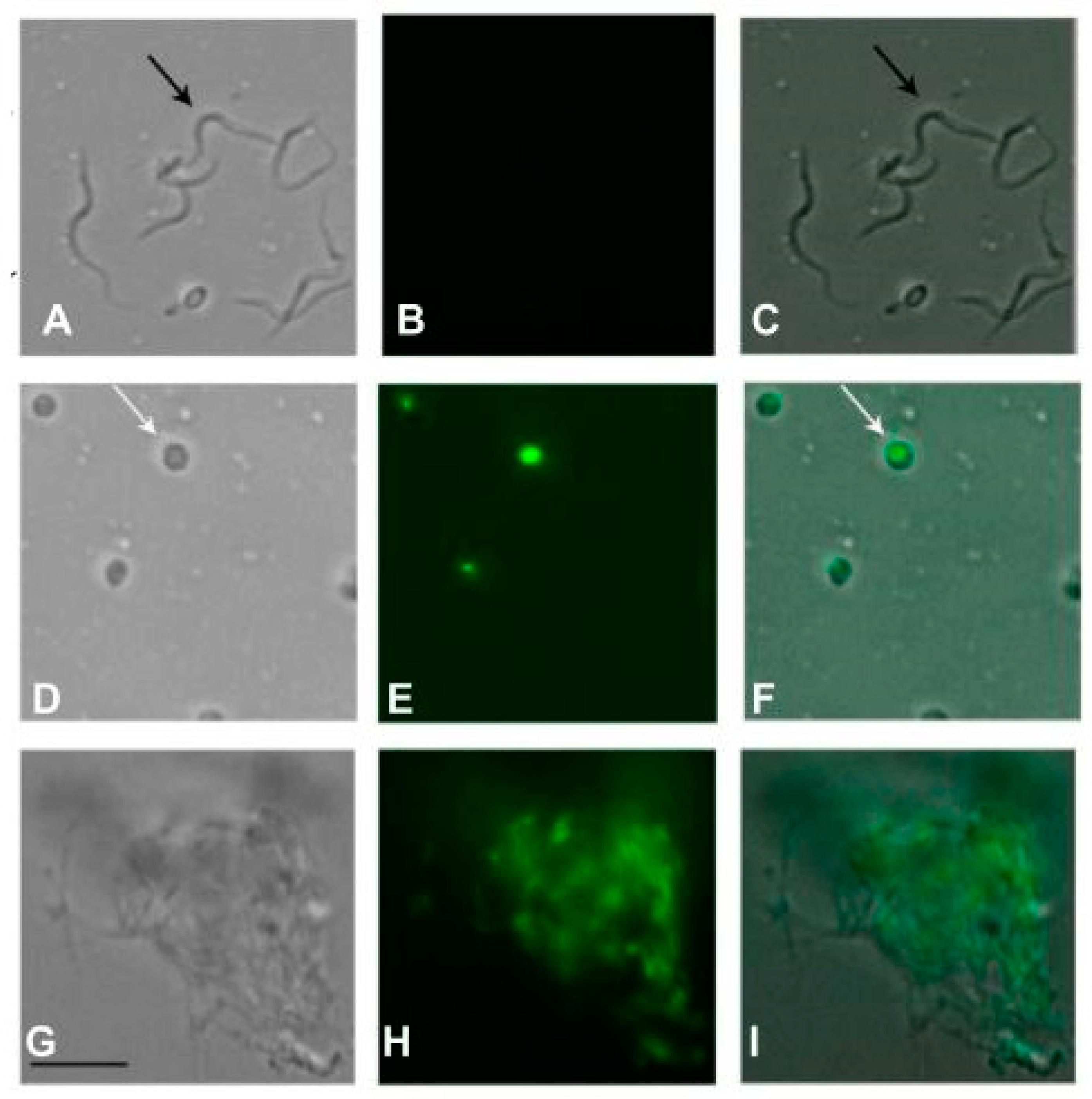

Table 1.
Origin of the microorganisms associated with corals or sediments.
| Acronym | Origin | Depth (meters) |
| TC | Tubastraea coccinea | 5 |
| MH | Mussismilia hispida | 10 |
| MD | Madracis decactis | 10 |
| SCSB | Sediment of the São Sabastião Channel | 35 |
| SIBUZ | Sediment of Buzios Island | 13 |
Table 2.
Identification and morphology of bacteria isolated from corals and marine sediments according to MALDI-TOF/MS score or sequencing.
Table 2.
Identification and morphology of bacteria isolated from corals and marine sediments according to MALDI-TOF/MS score or sequencing.
| Acronym | Morphology and Gram | Identification | Method |
|---|---|---|---|
| TC 2.0.2 | Bacillus (-) | Alteromonas macleodi | S |
| TC 2.2 | Bacillus (-) | Vibrio alginolyticus | MS |
| MH 3.0 | Bacillus (-) | Vibrio harveyi | MS |
| MH 3.3 | Bacillus (-) | Vibrio alginolyticus | MS |
| MD 5.0 | Coccobacillus (-) | Vibrio harveyi | MS |
| SCSB 6.0.2.1 | Bacillus (+) | Shewanella pneumatophori | S |
| SCSB 6.0.2.2 | Bacillus (+) | Mesoflavibacter zeaxanthinifaciens | S |
| SCSB 6.1 | Bacillus (-) | Bacillus megaterium | MS |
| SCSB 6.2 | Bacillus (-) | Vibrio harveyi | MS |
| SIBUZ 7 | Coccobacillus (-) | Vibrio harveyi | MS |
| SIBUZ 7.2.2 | Coccobacillus (-) | Halomonas aquamarina | S |
MS- identification by score obtained in MALDI-TOF/MS from two independent studies; S - identification by genetic sequencing; (-): Gram-negative bacteria. (+): Gram-positive bacteria; TC (Tubastraea coccinea), MH (Mussismilia hispida), MD (Madracis decactis), SCSB (Sediments of the São Sebastião Channel) and SIBUZ (Sediments of Buzios Island).
Table 3.
Identification of marine strains by partial sequencing of the 16S rRNA gene, according to the BLAST 16S rRNA sequences database.
Table 3.
Identification of marine strains by partial sequencing of the 16S rRNA gene, according to the BLAST 16S rRNA sequences database.
| Acronym | Microorganism | Identity (%) | Query Cover (%) | Acc number |
|---|---|---|---|---|
| TC 2.0.2 | Alteromonas macleodii | 99.78 | 99 | OP163900 |
| SCSB 6.0.2.1 | Shewanella pneumatophori | 99.72 | 99 | OP163959 |
| SCSB 6.0.2.2 | Mesoflavibacter zeaxanthinifaciens | 100 | 96 | OR479885 |
| SIBUZ 7.2.2 | Halomonas aquamarina | 99.72 | 100 | OP163958 |
Acc number: Access number in GenBank (NCBI).
Table 4.
Quantification of the 50% inhibitory concentration (IC50) of microbial metabolites against T. cruzi trypomastigotes, with the respective yields of extracted metabolites.
Table 4.
Quantification of the 50% inhibitory concentration (IC50) of microbial metabolites against T. cruzi trypomastigotes, with the respective yields of extracted metabolites.
| Acronym | Strain | Mass of metabolites (mg) | IC50 ± SD (μg/mL) |
|---|---|---|---|
| TC 2.0.2 | Alteromonas macleodi | 2.6 | 31.2 ± 2.6 |
| TC 2.2 | Vibrio alginolyticus | 2.1 | 13.9 ± 3.0 |
| MH 3.0 | Vibrio harveyi | 11.4 | 51.3 ± 1.1 |
| MH 3.3 | Vibrio alginolyticus | 8.4 | 25.4 ± 1.2 |
| MD 5.0 | Vibrio harveyi | 3.4 | 18.3 ± 2.6 |
| SCSB 6.0.2.1 | Shewanella pneumatophori | 5.5 | 15.1 ± 3.3 |
| SCSB 6.0.2.2 | Mesoflavibacter zeaxanthinifaciens | 4.8 | 17.9 ± 0.7 |
| SCSB 6.1 | Bacillus megaterium | 2.7 | 59.9 ± 0.1 |
| SCSB 6.2 | Vibrio harveyi | 3.9 | 32.8 ± 3.9 |
| SIBUZ 7 | Vibrio harveyi | 8.9 | 8.0 ± 0.7 |
| SIBUZ 7.2.2 | Halomonas aquamarina | 3.2 | 15.4 ± 0.5 |
TC (Tubastraea coccinea), MH (Mussismilia hispida), MD (Madracis decactis), SCSB (Sediments of the São Sebastião Channel), SIBUZ (Sediments of Buzios Island). SD (Standard Deviation). Benznidazole was used as standard drug (positive control).
Table 5.
Evaluation of the 50% Inhibitory Concentration (IC50) against trypomastigotes and amastigotes of T. cruzi and the 50% Cytotoxic Concentration (CC50) of fraction II from Mesoflavibacter zeaxanthinifaciens EtOAc extract and the positive control benznidazole.
Table 5.
Evaluation of the 50% Inhibitory Concentration (IC50) against trypomastigotes and amastigotes of T. cruzi and the 50% Cytotoxic Concentration (CC50) of fraction II from Mesoflavibacter zeaxanthinifaciens EtOAc extract and the positive control benznidazole.
| Compound | Trypomastigotes(IC50±SD) | Amastigotes (IC50±SD) | Cytotoxicity (CC50±SD) |
|---|---|---|---|
| Fraction II | 17.7 ± 3.5 μg/mL | 23.8 ± 2.7 μg/mL | >200 μg/mL |
| Benznidazole | 3.6 ± 0.9 μg/mL | 1.4 ± 0.5 μg/mL | 49.4 μg/mL |
SD - standard deviation; IC50: 50% Inhibitory Concentration; CC50: 50% Cytotoxic Concentration.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



