
Submitted:
21 February 2024
Posted:
22 February 2024
You are already at the latest version
Abstract
Iron is an essential nutrient and necessary for biological functions from DNA replication and repair to transcriptional regulation, mitochondrial respiration, electron transfer, oxygen transport, photosynthesis, enzymatic catalysis, and nitrogen fixation. However, due to iron’s propensity to generate toxic radicals which can cause damage to DNA, proteins, and lipids, multiple processes regulate uptake and distribution of iron in living systems. Understanding how intracellular iron metabolism is optimized and how iron is utilized to regulate other intracellular processes is important to our overall understanding of a multitude of biological processes. One of the tools that the cell utilizes to regulate a multitude of functions is ligation of the iron-sulfur (Fe-S) cluster cofactor. Fe-S clusters comprised of iron and inorganic sulfur are ancient components of living matter on earth that are integral for physiological function in all domains of life. Here we will explore the ways in which the cell utilizes Fe-S clusters to sense the intracellular environment and respond to maintain equilibrium.
Keywords:
Iron-sulfur clusters
; iron metabolism
; iron sensing
; IRE
; PAIR
; Aft1/2
; IRP1/2
; RirA
; Yap5
; ACO1
; IREB2
; HSCB
1. Introduction
Fe-S clusters are comprised of iron and inorganic sulfur and are one of the most ancient components of living matter on Earth. They are integral co-factors for physiological function in all domains of life [1,2,3,4]. Fe-S clusters act as cofactors for reduction-oxidation (redox) reactions that can change the structure and therefore function of proteins when they bind and/or change oxidation state[5,6]. In addition to their structural and catalytic roles, they may also function as sensors of iron and oxygen levels, interfacing between the intracellular environment and proteins involved in metabolic pathways [7,8,9,10,11]. Despite their ancient lineage and their key roles in many diverse biological functions, Fe-S clusters were not discovered and subsequently characterized until the late 1950s and early 1960s [2,3,12] when spectroscopic techniques and technologies were developed that enabled researchers to observe their intrinsic magnetic properties [1,13] (Figure 1). Since then, and especially in the last decade, numerous Fe-S containing proteins have been discovered in mammalian systems; however, due to the inherently labile nature and oxygen sensitivity of these cofactors, there are potentially many more proteins dependent upon Fe-S clusters than previously imagined [13] (Figure 1).
Recently, DNA replication and repair enzymes have been found to contain Fe-S clusters [14,15,16,17,18,19,20,21,22,23,24]. For example, the helicase-nuclease DNA2, glycosylases, helicases, base excision repair enzymes, and nuclear replicative DNA polymerases were found to ligate Fe-S cluster cofactors. Defects in Fe-S cluster biogenesis cause a concurrent increase in mitochondrial and nuclear DNA damage and ultimately genomic instability [17,25]. In addition to being largely unstable co-factors as mentioned above, one of the main threats to DNA integrity is oxidation of nucleic acid bases. As iron often contributes to damaging oxidation through Fenton chemistry [26,27,28,29], it was initially surprising when many DNA replication and repair enzymes were found to require FeS co-factors for function, though their intrinsic properties could potentially increase DNA damage and genomic instability. In some instances, the Fe-S clusters act as redox switches that provide a means to detect oxidative stress and modulate replication [6,22,30,31]. Also, some Fe-S clusters provide electrons for DNA Charge Transfer, which is then used by DNA repair enzymes to signal one another and search for DNA lesions [6,22]. Most recently, several Fe-S clusters have been discovered to be integral co-factors involved in viral replication of Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2), the pandemic-causing, causative agent of COVID-19 [32,33] (Figure 1). Three integral Fe-S cluster cofactors have been discovered in two SARS-CoV-2 proteins thus far. Two Fe-S clusters were found in SARS-CoV-2 RNA-dependent RNA polymerase (RdRp-nsp12), which is responsible for genomic replication and transcription, and one Fe-S cluster was discovered in the SARS-CoV-2 RNA helicase (nsp13) [33]. These clusters represent interesting novel broad spectrum anti-viral targets that are susceptible to oxidative damage and potentially unlikely to become refractory to targeted degradation [34].
Utilizing Fe-S clusters as cofactors in aerobic organisms is risky for the same reason it has been so difficult to discover new Fe-S cluster containing proteins. Fe-S clusters readily decompose upon exposure to oxygen [5,13], which leads to one of the largest unresolved questions in the field: Why did evolution favor continued dependence on Fe-S clusters after the increased oxygen levels in the atmosphere changed to largely disfavor their stability? When life began on earth, there was plenty of available iron and sulfur for easy and spontaneous incorporation into enzymes in the form of Fe-S cluster co-factors [5]. However, as oxygen levels rose, more iron was oxidized to less bioavailable forms while the lability and oxygen sensitivity of Fe-S clusters became a larger problem. This paradox suggests that utilization of Fe-S cluster co-factors has been under positive selection pressure, and Fe-S cofactors perform an essential role that other, less labile co-factors cannot replicate. Some of the most important Fe-S cluster functions include sensing the intracellular environment and regulating cellular processes in response [10,13,35].
2. FeS Cluster Structure, Geometries, and Biogenesis
Iron-sulfur clusters exhibit a variety of structures and geometries depending on the number of iron and sulfur atoms in the cluster and the coordinating residues present in the protein. The coordinating amino acids found in most Fe-S cluster proteins are largely comprised of cysteine and histidine, but occasionally aspartate, glutamate, lysine, serine, and threonine are also coordinating residues[36,37]. The most common iron-sulfur cluster geometries found in biological systems are [2Fe-2S], [4Fe-4S], and [3Fe-4S] [38]. Rhomboid [2Fe-2S] clusters consist of two iron atoms bridged by two amino acids residues, cubane [4Fe-4S] clusters consist of four iron atoms coordinated by four inorganic sulfur atoms and ligated by four amino acids forming a cubane-like structure with a central cavity, whereas pyramidal or triangular [3Fe-4S] clusters consist of three iron atoms coordinated by four inorganic sulfur atoms ligated by three amino acid ligands.
The transition metal iron is key to one of the main underlying functions of iron sulfur clusters, the ability to accommodate reversible binding of a single electron and to enhance movement of single, unpaired electrons through a relay system, such as that found in respiratory complex I [39]. Fe-S clusters’ unique capacity to accept and donate unpaired electrons in a non-energy intensive, disruptive manor has cemented their place in key biological functions across the cell. The most notable examples of electron relay systems are found in multiprotein complexes involved in mitochondrial respiration and photosynthesis [10,11,40,41].
The Fe-S cluster biogenesis machinery is highly conserved between eukaryotes and prokaryotes [11]. In mammalian cells, initial Fe-S cluster biogenesis occurs in parallel in both the mitochondrial matrix and cytosol [1,10,42,43,44,45,46,47,48]. The biogenesis of nascent Fe-S clusters is a highly energy intensive process that requires multiple proteins. Fe-S cluster proteins are de novo assembled on scaffold protein ISCU [46,49,50,51,52]. Inorganic sulfur atoms are supplied by the cysteine desulfurase NFS1, which requires ISD11 to stabilize the interaction and the cofactor pyridoxal phosphate [53,54,55]. Release of sulfur atoms from cysteine, and ligation to ISCU is promoted by transient binding of frataxin. Sulfur combines with 2 Fe2+ atoms and 2 reducing equivalents to form a [2Fe-2S] cluster on ISCU [56,57]. The iron necessary for cluster formation originates from an incompletely characterized pool of available iron sometimes referred to as the “chelatable or labile iron pool”[58,59]. Ligands for available iron may include glutathione [60] cytosolic iron chaperones PCPB1 and BOLA2 [61,62,63], and perhaps ATP, citrate, GAPDH, and other proteins that are not yet characterized. In the mitochondria ISCA1, ISCA2, and IBA57 proteins have been implicated in conversion of [2Fe-2S] clusters to [4Fe-4S] [64]. There is evidence that NFU plays an important role in maturation from [2Fe-2S] clusters to [4Fe-4S] clusters in both the mitochondrial matrix and the cytosol[50], but more details about the formation of cubane [4Fe-4S] clusters from the initial [2Fe-2S] building blocks formed on ISCU are needed. The newly formed Fe-S cluster can then be transferred to recipient proteins via a chaperone/cochaperone system. Briefly, holo-ISCU binds to the chaperone co-chaperone pair, HSC20 (aka HSCB) and HSPA9. HSPA9 is an ATPase that uses the energy from ATP to drive a conformational change in the Fe-S cluster transfer complex, enabling the complex to deliver Fe-S cofactors to specific recipients. An important advance in understanding how recipients are correctly identified emerged from studies of SDHB [39]. A three amino acid motif in recipient proteins that consists of an aliphatic amino acid in position one, followed by a large hydrophobic residue, either phenylalanine or tyrosine, and ending with a positively charged amino acid, either arginine or lysine, fits into a pocket in HSC20 which has been defined by mutagenesis [65]. Such motifs, known as LYR-like motifs, aid in the specificity of Fe-S cluster delivery to recipient proteins [65]. Specifically, recipient proteins are bound to the transfer complex when the nascent Fe-S cluster is released, leading to the safe, efficient, and specific transfer of the cluster from the biogenesis machinery to recipient proteins [45]. The fact that the transfer process is guided by direct interactions between biogenesis machinery and Fe-S insertion into recipients likely ensures that the vulnerable nascent Fe-S cluster will not be degraded by unprotected exposure to solvents and oxygen. HSC20 stimulates the ATPase activity of HSPA9 resulting in HSC20 dimerization via its J domain. Dimerized HSC20-ISCU may then bind to the cytosolic Fe-S platform protein, CIAO1 via CIOA1’s LYR motif. CIAO1 is a part of a large multimeric iron sulfur cluster delivery complex including MMS19, and FAM96B. Apo Fe-S cluster proteins likely interact with the large complex, likely due to the presence of a LYR-like motif, to receive their clusters. They then remain in the cytosol or are translocated to the nucleus to participate in DNA metabolism.
Much of our knowledge about FeS biogenesis arose from studies in bacteria, in which an operon devoted to FeS biogenesis was discovered [66,67]. Homology and processes were sufficiently conserved so that bacterial studies informed many studies performed in eukaryotes, including yeast, plants, and mammalian cells and other organisms.
3. Human Iron Metabolism
Iron is efficiently recycled and therefore moves throughout the body into different tissues and cell types in a tightly controlled cycle. Typically, only 1 to 1.5 mg of dietary iron needs to be absorbed per day to replace iron loss from epithelial sloughing and minor blood loss in order to maintain sufficient iron stores to support normal physiological processes within the human [68]. Non-heme iron is absorbed from the diet by polarized gut enterocytes through the apical Divalent Metal Transporter 1 (DMT1, also known as NRAMP2, DCT1, or SLC11A2) [69], and secreted into the blood on the basolateral side of the epithelium by Ferroportin (FPN1, also known as IREG1, MTP1, or SLC40A1) [70] where ferrous iron is oxidized by membrane-bound hephaestin [71] and bound by transferrin (Tf) that circulates in the bloodstream [72]. Diferric transferrin then circulates iron throughout the body, delivering most transferrin-bound iron to the bone marrow, where developing erythroid cells endocytose the transferrin after it binds to transferrin receptor (TFRC, aka TfR1) on the surface of the plasma membrane, strip it of iron and then pump the iron in the endosome through DMT1 into the cytosol and eventually to the mitochondria [73]. Iron enters the mitochondrial matrix through the inner membrane iron transporter, Mitoferrin (MFRN1, also known as SLC25A37) [74,75], where heme synthesis and iron-sulfur cluster biogenesis occurs [1,47,76,77]. Once the hemoglobinization process has started, erythroid cells begin the differentiation process [78,79] into mature red blood cells, which circulate throughout the body delivering oxygen from the lungs to other tissues and organs. When red blood cells become damaged, they are recycled by specialized macrophages primarily in the spleen, and also in the liver in cells known as Kupffer cells [80]. Macrophages phagocytose the damaged red blood cells, strip the iron from heme and secrete it into the blood stream via the iron exporter, FPN1 [81,82,83,84] where transferrin once again binds iron and the cycle continues [85,86] . If perturbations due to environmental or genetic causes arise at any point in this process, the tightly controlled system can break down, resulting in symptomatic disease. For instance, when macrophages lack inducible heme oxygenase, they are unable to defend themselves from heme toxicity, and macrophage death causes a severe disease in mice and human patients, which can be overcome when normal functional macrophages are introduced [87,88,89].
While iron is an essential nutrient that is required for many important functions such as oxygen transport, electron transfer, oxidation-reduction reactions and catalytic activity in metalloproteins, it can also be very toxic when it builds up in excess within the body and cells [90,91,92,93] . Iron is an efficient catalyst for biological reactions due to it redox capabilities, and it is therefore a cofactor in many enzymes. Unfortunately, the same properties that make it indispensable to life also lead to its toxicity. When too much free iron overwhelms a biological system, it can lead to production of reactive oxygen species (ROS) and toxicity caused by the formation of superoxide followed by the formation of a hydroxyl radical [58,94,95,96]. Hydroxyl radicals lead to DNA and protein damage as well as lipid peroxidation, which can cause genetic mutations, necrosis, and tissue damage[90,97,98,99]. Therefore, even though iron deficiency is a worldwide problem, iron uptake, utilization, and storage mechanisms within biological systems have evolved to be highly regulated phenomena. Iron uptake is regulated by a complex system of iron sensing proteins in concert with transcriptional and translational regulation because there is no efficient mechanism for the human body to release iron once it is acquired[90,100,101,102].
In addition to its potential to cause toxicity, iron is at the forefront of host pathogen interactions[103,104,105,106]. Iron is a required nutrient for both pathogens and their hosts, leading to a tug of war between pathogens, the microbiome and the host. This competition for iron causes each group to balance its efforts to obtain sufficient iron while simultaneously preventing bacterial competitors from successfully acquiring iron[103,104,105,106,107]. Often, microbes biosynthesize siderophores to scavenge for iron in their environment[105,108,109,110], while hosts sequester iron away from pathogens by binding it with calprotectin in blood cells such as neutrophils[111,112], storing it in the protein ferritin[113], or allowing it to circulate throughout the body tightly bound to the protein transferrin[107,114,115]. It is very rare for any iron to be unbound to some type of chelator within the body. This sequestration of iron away from potentially pathogenic microorganisms is known as nutritional immunity[104,105,107,116]. Excess transferrin helps to keep pathogens at bay by minimizing presence of non-transferrin bound iron (NTBI)[103,104,105,115,117,118,119].
Because of iron’s important role in biological systems, and the need for tight regulation of iron within the body, it makes sense that hundreds of diseases are associated with impaired iron metabolism. Disorders of iron metabolism are the most common genetic deficiencies world-wide, and because iron is such an integral metal ion for many biological processes, iron overload or deficiency can affect almost every system and process of the body[72,114,120,121,122]. Currently, there are more than 25 known hereditary diseases caused by deficiencies in iron regulation and homeostasis, and additional disorders can be caused by acquired loss of protein function due to chronic illness, inflammation, and other environmental factors[122,123,124] .
4. Intracellular Iron and Oxygen Sensing
There is significant cross-talk between oxygen-sensing and iron-sensing pathways[8,9,35,125]. Proteins involved in both pathways can influence each other's activity. The availability of iron can influence the assembly and stability of Fe-S clusters in oxygen sensing proteins. Additionally, when oxygen levels are high, Fe-S clusters in iron response and iron regulatory proteins can be degraded, resulting in changes in the Fe-S cofactor that normally result during a low iron state [35,125,126,127,128,129,130,131,132,133]. Cells therefore use Fe-S clusters as highly sensitive and efficient cofactors to sense and respond to intracellular iron and oxygen levels. Mammalian, bacterial, and yeast cells utilize Fe-S cluster containing iron regulatory proteins for intracellular iron sensing. In the broadest sense, iron regulatory proteins, such as IRP1 (aka ACO1) in humans[1,7,10,102,134,135,136,137], IscR/RirA in bacteria[8,27,138,139,140,141,142], and Aft1 in yeast[143,144,145,146] sense iron levels via the presence or absence of Fe-S clusters. In iron replete conditions, ligation of Fe-S cluster cofactors is favored. However, in cases of iron depletion or oxidative stress, Fe-S cluster incorporation is disfavored due to decreased Fe-S biogenesis, or oxidative degradation respectively. The ligation of iron-sulfur clusters causes functional and conformational changes of the protein’s activity and structure, allowing for modulation of expression of downstream iron metabolic proteins.
4.1. Mammalian Iron Sensing Iron-Sulfur Clusters
In mammalian cells, iron transport and storage proteins are transcriptionally, translationally, and post translationally regulated to maintain physiological intracellular labile (or chelatable) iron levels while avoiding functional iron overload and toxicity[101]. Specifically, levels of the iron transporter Divalent Metal Transporter 1 (DMT1), the iron sequestration and storage protein ferritin (heavy, FTH1 and light, FTL1 chains), the sole iron efflux protein ferroportin (FPN1), and the iron uptake mediator transferrin receptor 1 (TfR1) are translationally or post-transcriptionally regulated through short hairpin iron response elements (IREs) located at the 5’- and 3’-untranslated regions (UTR) of the corresponding mRNA transcripts that impair translation (5’UTR IRE) or prolong mRNA half-life (3’UTR IRE)[10]. Iron-sensing iron response proteins (IRP1 and IRP2) bind to these IREs to block translation (5’-IRE: FTH1, FTL1, FPN1, HIF2α, ACO2, ALAS2) or stabilize mRNA (3’-IRE: DMT1, TfR1, CDC14A, MRCKα) under iron starvation[7,132,133,147,148,149,150,151,152,153,154,155,156,157,158].
Upon iron stimulation and binding, the IRPs dissociate from the mRNAs, reversing their described effects. Transcriptional regulation is achieved through the transcriptional activator, hypoxia-inducible factor 2-alpha (Hif2α, aka EPAS1), which is degraded after oxygen and iron-mediated proline hydroxylation[101]. Interestingly, Hif2α activates transcription of a transcript of Fpn1 that evades IRE-mediated translational repression under iron deprivation conditions[131,149].
The mechanisms that IRP1/2 utilize to sense and react to intracellular iron levels are elegant (Figure 2). IRP1 exists in two mutually exclusive forms, either as a cytosolic aconitase or as an RNA binding protein[7,158,159]. The dual nature of IRP1 allows this unique protein to sense iron levels within the cytosol by conditionally ligating an Fe-S cluster in it’s active site cleft. In iron replete conditions, IRP1 acquires a [4Fe-4S] cluster. The holo-IRP1 enzyme acts as a cytosolic aconitase, isomerizing citrate and isocitrate[10,134]. In iron starvation conditions, the Fe-S cluster of IRP1 is absent, and IRP1 becomes an IRE binding protein[35,78,102,137]. Activation of IRE binding activity increases cytosolic iron levels by reducing iron sequestration in ferritin and by increasing transferrin receptor activity[7]. These two major responses are accomplished by repressing translation of ferritin and stabilizing mRNA levels of TFRC (TFR1), which allows increased synthesis of transferrin receptor. Whereas IRP1 utilizes an Fe-S cluster to switch its activity from aconitase to IRE binding, IRP2 (aka IREB2) does not contain an Fe-S cluster and does not exhibit aconitase activity[1,72,136,160,161]. This divergence in activity and cluster ligation is surprising due to the high sequence similarity (~60%) that IRP1 and IRP2 share[7,162,163]. IRP2 therefore only exhibits iron regulatory properties via its IRE-binding activity. Despite not containing an Fe-S cluster directly, IRP2 levels are still influenced by an oxygen sensing Fe-S cluster[160,164]. It is not surprising that two proteins would have the same regulatory function but differ in how they are regulated, as many genes in the mammalian genome likely arose during whole genome duplication of an evolutionary precursor[165] enabling organisms to develop systems that are robustly supported by multiple pathways[166], some of which have mechanistically diverged over time. For instance, degradation of IRP2 when iron levels are high is a more robust response than turning off IRE binding activity through insertion of an Fe-S cluster cofactor that occludes the IRE-binding site.
The protein FBXL5 recruits ubiquitin ligase to specifically bind to and target IRP2 for ubiquitination and proteosomal degradation[135,160,164,167,168,169,170]. FBXL5 does this in an iron and oxygen dependent manner. When iron levels are low, destabilization and degradation of FBXL5 renders it unable to bind to and target IRP2 for proteasomal degradation[160]. The [2Fe-2S] cluster in the C-terminal domain of FBXL5 is highly redox-sensitive[167]. FBXL5 can therefore utilize this redox sensitive [2Fe-2S] cluster to sense oxygen levels and regulate IRP2 protein levels[164]. When the [2Fe-2S] cluster is in the 2+ oxidized state, a conformational change occurs that allows for a rearrangement of the c-terminus and binding of FBXL5 to IRP2, allowing for the ubiquitination and proteasomal degradation pathway to move forward. Therefore, while IRP2 itself does not directly ligate an intracellular environment sensing Fe-S cluster, it is still regulated by one that occupies a different position in the overall regulatory scheme. Thus, activity of each IRP depends on the status of an Fe-S cofactor which determines how much IRE-binding activity each IRP contributes to regulation of its nine major target transcripts, either through regulating the rate of translation, or by stabilizing the transcript to allow increased protein synthesis. Interestingly, parallel IRE/IRP systems have been discovered in other eukaryotes including protozoans like Trichomonas vaginalis[171].
Recently, NCOA4, the master regulator of ferritinophagy, has gotten a lot of attention from the iron biology field. Ferritinophagy is the process via which ferritin is degraded in the autolysosome so that the iron it has stored can be recycled for use within the cell. Aberrant ferritinophagy has been linked to neurodegeneration and pancreatic cancer [28,172,173,174,175]. NCOA4 was known to bind iron and regulate ferritinophagy in response to iron levels, but until recently it was unknown how NCOA4 sensed iron[28,173]. NCOA4 has now been proposed as both an oxygen sensor[176] and an intracellular iron sensor[177] due to its ligation of an Fe-S cluster. These new discoveries highlight the inextricable link between Fe-S clusters and the regulation of intracellular regulation of iron metabolism.
4.2. Bacterial Iron Sensing Iron-Sulfur Clusters
Iron is a limiting reagent for microorganism growth both pathogenic and free living. As is the case with mammalian and multicellular organisms, iron is both necessary for survival but also highly toxic at high concentrations[178]. Bacteria therefore require the ability to sense intracellular and extracellular iron to maintain optimal iron concentrations for growth. Bacteria have a large arsenal of iron acquisition techniques which range from simple membrane bound iron transporters to small molecule siderophore scavengers[108,110,178,179,180]. Iron uptake needs to be tightly controlled to prevent iron overload and subsequent toxicity.
Bacteria use many environmental sensing Fe-S clusters in order to react to intracellular and extracellular conditions. These sensing Fe-S clusters have been covered extensively in the literature[9]. One recently discovered example of an iron-sensing Fe-S cluster protein in bacteria is the iron responsive Rhizobium protein rhizobial iron regulator (RirA)[27,181]. Mutations in RirA cause deregulation, and therefore constitutively high transcription rates of many operons involved in iron acquisition. Rhizobium is a genus of nitrogen fixing, gram-negative bacteria that associate with the root systems of plants in structures called root nodules[182]. The plant, often in the legume or flowering plant family, and the rhizobium bacteria have a symbiotic relationship wherein the rhizobium utilizes its nitrogenase to convert atmospheric nitrogen in the soil to bioavailable nitrogen for the plant to use to optimize growth conditions [182]. The plant supplies the bacterial colonies with nutrients through their association with the root system, which allows both organisms to thrive. Similar to IRP1, RirA exists both in an apo- and holo [4Fe-4S] cluster form[138]. Like the mammalian system, there is redundancy to modulate such an important physiological function, and rhizobium has an additional non Fe-S cluster binding co-regulator similar to IRP2, known as the iron response regulator (Irr), which aids in the modulation of intracellular iron levels[179]. When RirA is in its holo- [4Fe-4S] cluster form, it represses iron uptake by binding to the iron response operator (IRO) box, a DNA sequence present in promoters of multiple genes[8,27,37,138]. Binding to the IRO box by RirA suppresses transcription of iron uptake genes (Figure 2). When RirA loses its cluster due to low iron and/or aerobic conditions, it undergoes a conformational change and can no longer bind to the IRO box, which then allows unfettered transcription and the expression of numerous iron acquisition genes [8,27,138,139,140].
Numerous other bacterial regulatory proteins utilize Fe-S cofactors to sense levels of nitric oxide, oxidative stress, Fe-S cluster sufficiency, and oxygen levels to regulate transcription of genes needed to combat environmental challenges. Responses range from upregulation of DNA repair enzymes and siderophore synthesis, to dramatic global changes such as sporulation, increased motility, and biofilm production [183]. Since Fe-S cofactors are often lost or replaced by zinc during aerobic purification, it is reasonable to predict that more bacterial Fe-S cluster sensors will be discovered in the future[13,184].
4.3. Yeast Iron Sensing Iron-Sulfur Clusters
Like mammalian and bacterial intracellular iron sensing systems, yeast utilize iron sensing Fe-S clusters to regulate iron metabolism. Yeast react to iron deficiency quickly and efficiently via transcriptional regulation implemented by three main transcription factors, Aft1, Aft2, and Yap5. Aft1/2 are paralogs that share 26% sequence identity and regulate yeast iron metabolism via turning on and off the yeast iron regulon[143,145,146]. Specifically, similarly to IRP1 which regulates translation, when iron levels are low, apo- Aft1/2 monomers bind to DNA sites to increase transcription of members of the iron regulon[143,185,186]. When cytosolic iron levels rise, Aft1/2 ligate a [2Fe-2S] cluster causing a conformational change that does not allow for DNA binding, thus shutting off transcription of the iron regulon and thereby reducing iron uptake. Aft1/2 gain their cluster by forming a complex with Grx4/5 and Bol2[145] (Figure 2). Members of the regulon aid in the acquisition of iron via cellular membrane iron transporters (e.g. FET3/FTR1), siderophore binding/uptake (eg ARN1-4, FIT1-3), Fe-S biogenesis machinery (ISCU), and mRNA binding regulatory proteins (CTH1/2)[185,187]. The iron-transporting complex Fet3Ftr1 is required for growth on low-iron media and is one of the main ways in which yeast acquire iron [122,188,189]. ARN1-4 are responsible for transporting siderophore-iron chelates into the cell and these transporters are therefore important for iron scavenging in scarce iron conditions[122,190,191]. CTH1/2 target and degrade RNA transcripts of nonessential proteins that require copious amounts of iron for their function. A more extensive list of genes regulated by Aft1/2 has been compiled (Kaplan & Kaplan 2009)[185]. Opposing Aft1/2, Yap5 constitutively binds to the Yap Responsive Element within the genome[143,186,187,192,193,194,195]. When iron levels are low, Yap5 represses transcription of iron storage and sequestration proteins. When intracellular iron levels are high, YAP5 ligates a [2Fe-2S] cluster, which changes its conformation to promote the transcription of iron storage and sequestration proteins among other proteins necessary to adapt to high intracellular iron [193,194]. Specifically, Yap5 regulates a yeast glutaredoxin (Grx4), a wybutosine modified tRNA synthesis enzyme (Tyw1), a vacuolar iron-copper transporter Ccc1, and a copper chelator (metallothionein Cup1)[145]. The proteins up-regulated by Yap5 aid in protecting the cell from protein and DNA damage due to iron overload by sequestering iron in vacuoles (Ccc1), turning off the iron acquisition pathway via delivery of iron sulfur clusters to Aft1/2 (Grx4), binding copper to halt ferrireductases from aiding iron import (Cup1), and increasing fidelity of tRNAs to ensure proper translation of proteins when iron concentrations are high (Tyw1). Though yeast model systems are among the most studied model organisms for very good reason, more questions remain about how they regulate iron metabolism and interact with their environment.
4.4. Plant Iron Sensing Iron-Sulfur Clusters
Although iron is the most abundant element on earth, the vast majority of iron is in a biologically inaccessible, insoluble ferric hydroxide state[5]. Plants, like the other organisms above, deal with nutritional iron deficiency in areas where bioavailable iron is reduced. Conversely, plants need to be able to reduce iron uptake in iron rich soils. Unlike more motile forms of life, most plants cannot physically move to find the perfect balance of nutrients once they have established themselves. Lack of motility means that they need to have the ability to effectively sense and react to changes in their environment at the molecular level. Therefore, plants need to be able to sense the iron availability or lack thereof in their environment as well as intracellularly.
Currently, there are hints that Fe-S clusters may be involved in iron sensing pathways, but no definitive evidence has been found to date[196]. As mentioned above, IRP1 has dual function as an RNA binding, iron response protein, and alternatively, as a cytosolic aconitase. Interestingly, a recent study in rice by Senoura et al seems to promise the possibility of an iron-sensing Fe-S cluster in rice[197]. When one of three aconitase genes, OsACO1, is knocked down in rice, changes in iron homeostasis and iron-responsive genes are observed. OsACO1 is an aconitase that binds a [4Fe-4S] cluster and is expressed ubiquitously throughout the plant. Rice heterozygous for OsACO1, OsACO1(+/-), have reduced aconitase activity as well as reduced expression of genes involved in iron trafficking and regulation. Homozygous deletion of OsACO1 was embryonic lethal. A reduction in the nicotianamine (NA) synthase gene, OsNAS2, lowers plant NA synthesis, which reduces long-distance transport of iron due to decreased of NA [197,198]. NA is an iron chelator that plants utilize to enhance iron trafficking throughout the plant[197], analogous to the function of transferrin in iron trafficking and circulation throughout the body in humans. Additionally, lowered gene expression of OsYSL15, a protein involved in iron uptake in roots[199], and OsIRO2, a transcription factor involved in iron homeostasis[200], is observed in the roots of OsACO1(+/-) rice plants. There is also evidence that OsACO1 has RNA-binding activity. Similar to mammalian IREs to which IRP1/2 bind, the RNA probes bound by OsACO1 in a gel-shift assay contain an RNA stem-loop structure composed of a 7-nucleotide loop on a stem of five-paired nucleotides with an unpaired bulge on the 5′ strand of the stem. In IRP1/2-IRE binding, the loop, 5 base-paired stem, and unpaired bulge has been shown to be vital for IRP1/2 recognition of the IRE[197] (Figure 2). The authors called this new RNA stem-loop structure Plant ACO-Interacting RNA element or PAIR. Subsequent studies confirming that the [4Fe-4S] in rice OsACO1 is utilized as an iron-sensing Fe-S cluster are likely to be interesting and will help fill in the missing link of how plant cells physically sense iron to better adapt to their environment.
4.5. Archaea Iron Sensing Iron-Sulfur Clusters
Archaea are an ancient form of prokaryotic life that is estimated to have diverged over 3.9 billion years ago [201,202,203]. The archaeal iron-sulfur biogenesis pathway is not highly conserved between archaea species or between archaea, bacteria, and eukaryotes[204]. While archaea have more in common with eukaryotes than prokaryotes, those species that do have proteins associated with canonical Fe-S cluster biogenesis machinery most likely acquired them via lateral gene transfer from bacteria throughout their evolutionary journey [205,206]. Like the majority of life, most species of archaea rely heavily on Fe-S clusters in a variety of processes including electron transfer performed in large part by ferredoxins[207]. A subtype of archaea called Methanogens are thought to contain the highest percentage of Fe-S cluster containing proteins per genome of any other organism[208]. Currently, it is thought that the anaerobic environments that methanogens inhabit contribute to their ability to contain such a uniquely large percentage of Fe-S cluster proteins in their proteome[208]. It is therefore surprising that iron-sulfur clusters have not been more heavily implicated in intracellular iron sensing in archaea, especially considering their heavy reliance on Fe-S cluster cofactors. Though a paper published in 2002 suggested that the aconitase of Sulfolobus solfataricus could be acting as an iron-sensing Fe-S cluster similarly to IRP1[209] (Figure 2), no subsequent confirmational studies have been published. The domain archaea comprises a vast and diverse group of organisms that inhabit some of the most extreme environments on earth. Archaea therefore represent a treasure trove in which new discoveries and yet to be elucidated biological breakthroughs may be uncovered.
5. Conclusions, Future Perspectives and Remaining Questions
Tight control of iron metabolism and therefore intracellular iron sensing is critical to all forms of life. Iron sensing Fe-S Clusters represent a rapid and highly sensitive co-factor for detection of iron levels. When these ancient co-factors are incorporated (or not) into transcription factors and posttranscriptional modulators, they allow cells to sense and react to their intracellular environment through alteration of protein expression. While iron-sulfur clusters are ubiquitous and essential co-factors in all domains of life, many unanswered questions remain. Fe-S cofactors are hard to study and isolate due to the same properties that enable them to be good iron and oxygen sensors. Their labile nature and propensity to degrade in the presence of oxygen often cause them to disappear when proteins are purified on the bench in the presence of oxygen. Therefore, it is very possible that Fe-S cluster containing proteins are even more ubiquitous than they are currently thought to be. They are hiding in plain sight waiting for an anaerobic chamber and a determined researcher to reveal their existence. Recently, essential Fe-S clusters were found to be necessary for the causative agent of the pandemic causing COVID-19 (SARS-CoV-2) virus to replicate. It is very possible that even more viruses depend on Fe-S clusters to replicate their genetic material, or perhaps sense and modulate the metabolism of the cells they are infecting. How many more human disease relevant Fe-S cluster cofactors are yet undiscovered?
Interestingly there is very little data on potential iron sensing Fe-S clusters in plants and archaea. With archaea being some of the most ancient forms of life on earth and known to contain many Fe-S cluster proteins, it would not be surprising if at least some species utilize Fe-S clusters to sense iron and oxygen levels within their cytosol. An additional large and multipart remaining question regarding sensing iron sulfur clusters is: how do the ligating proteins switch between their apo and holo forms? In order to sense the intracellular environment, do the proteins lose their clusters, or are the Fe-S cofactors never incorporated due to lack of iron? If they lose their cluster due to low iron levels, are they able to regain a cluster under iron replete conditions? How does the stability of the cluster compare to its turnover due to damage and degradation? When does normal acquisition of the cluster occur? While the protein is folding or after the protein is folded?
The field of iron-sulfur cluster biology has come a long way over the past few decades with an explosion of newly discovered Fe-S cluster containing proteins. These discoveries open new chemical and biological vistas to study these fascinating, ancient, and essential metal co-factors. As always questions remain, and new discoveries await!
References
- Rouault, T.A. The indispensable role of mammalian iron sulfur proteins in function and regulation of multiple diverse metabolic pathways. Biometals 2019, 32, 343–353. [CrossRef]
- Beinert, H. Iron-sulfur proteins: ancient structures, still full of surprises. J Biol Inorg Chem 2000, 5, 2–15. [CrossRef]
- Beinert, H.; Holm, R.H.; Münck, E. sIron-sulfur clusters: Nature's modular, multipurpose structures. Science 1997, 277, 653–659. [CrossRef]
- Koonin, E.V.; Martin, W. On the origin of genomes and cells within inorganic compartments. Trends in Genetics 2005, 21, 647–654. [CrossRef]
- Imlay, J.A. Iron-sulphur clusters and the problem with oxygen. Mol Microbiol 2006, 59, 1073–1082. [CrossRef]
- Barton, J.K.; Silva, R.M.B.; O'Brien, E. Redox Chemistry in the Genome: Emergence of the [4Fe4S] Cofactor in Repair and Replication. Annual Review of Biochemistry 2019, 88, 163–190. [CrossRef]
- Maio, N.; Zhang, D.-L.; Ghosh, M.C.; Jain, A.; SantaMaria, A.M.; Rouault, T.A. Mechanisms of cellular iron sensing, regulation of erythropoiesis and mitochondrial iron utilization. Seminars in Hematology 2021, 58, 161–174. [CrossRef]
- Pellicer Martinez, M.T.; Crack, J.C.; Stewart, M.Y.; Bradley, J.M.; Svistunenko, D.A.; Johnston, A.W.; Cheesman, M.R.; Todd, J.D.; Le Brun, N.E. Mechanisms of iron- and O(2)-sensing by the [4Fe-4S] cluster of the global iron regulator RirA. Elife 2019, 8. [CrossRef]
- Mettert, E.L.; Kiley, P.J. Fe-S proteins that regulate gene expression. Biochimica et Biophysica Acta - Molecular Cell Research 2014, 1853, 1284–1293. [CrossRef]
- Rouault, T.A.; Maio, N. Biogenesis and functions of mammalian iron-sulfur proteins in the regulation of iron homeostasis and pivotal metabolic pathways. Journal of Biological Chemistry 2017, 292, 12744–12753. [CrossRef]
- Rouault, T.A. Biogenesis of iron-sulfur clusters in mammalian cells: New insights and relevance to human disease. DMM Disease Models and Mechanisms 2012, 5, 155–164. [CrossRef]
- Beinert, H.; Sands, R.H. Studies on succinic and DPNH dehydrogenase preparations by paramagnetic resonance (EPR) spectroscopy. 1960.
- Rouault, T.A. Iron-sulfur proteins hiding in plain sight. Nature Chemical Biology 2015, 11, 442–445. [CrossRef]
- Oliveira, M.T.; Garesse, R.; Kaguni, L.S. Animal models of mitochondrial DNA transactions in disease and ageing. Experimental Gerontology 2010, 10.1016/j.exger.2010.01.019. [CrossRef]
- Prakash, A.; Doublié, S. Base Excision Repair in the Mitochondria. Journal of Cellular Biochemistry 2015, 116, 1490–1499. [CrossRef]
- Fuss, J.O.; Tsai, C.L.; Ishida, J.P.; Tainer, J.A. Emerging critical roles of Fe-S clusters in DNA replication and repair. In Biochimica et Biophysica Acta - Molecular Cell Research, Elsevier: 2015; Vol. 1853, pp 1253-1271.
- Puig, S.; Ramos-Alonso, L.; Romero, A.M.; Martinez-Pastor, M.T. The elemental role of iron in DNA synthesis and repair. Metallomics 2017, 9, 1483–1500. [CrossRef]
- Baranovskiy, A.G.; Siebler, H.M.; Pavlov, Y.I.; Tahirov, T.H. Iron–Sulfur Clusters in DNA Polymerases and Primases of Eukaryotes, 1 ed.; Methods in Enzymology. Elsevier Inc.: 2018; Vol. 599, pp. 1–20.
- Holt, M.E.; Salay, L.E.; Chazin, W.J. A Polymerase With Potential: The Fe–S Cluster in Human DNA Primase, 1 ed.; Methods in Enzymology. Elsevier Inc.: 2017; Vol. 595, pp. 361–390.
- Netz, D.J.A.; Stith, C.M.; Stümpfig, M.; Köpf, G.; Vogel, D.; Genau, H.M.; Stodola, J.L.; Lill, R.; Burgers, P.M.J.; Pierik, A.J. Eukaryotic DNA polymerases require an iron-sulfur cluster for the formation of active complexes. Nature Chemical Biology 2012, 8, 125–132. [CrossRef]
- Troadec, M.B.; Loréal, O.; Brissot, P. The interaction of iron and the genome: For better and for worse. Mutation Research - Reviews in Mutation Research 2017, 774, 25–32. [CrossRef]
- O'Brien, E.; Barton, J.K.; Holt, M.E.; Thompson, M.K.; Salay, L.E.; Ehlinger, A.C.; Chazin, W.J. The [4Fe4S] cluster of human DNA primase functions as a redox switch using DNA charge transport. Science 2017, 355. [CrossRef]
- Lukianova, O.A.; David, S.S. A role for iron-sulfur clusters in DNA repair. Current Opinion in Chemical Biology 2005, 9, 145–151. [CrossRef]
- ter Beek, J.; Parkash, V.; Bylund, G.O.; Osterman, P.; Sauer-Eriksson, A.E.; Johansson, E. Structural evidence for an essential Fe–S cluster in the catalytic core domain of DNA polymerase ϵ. Nucleic Acids Research 2019, 47, 5712–5722. [CrossRef]
- Veatch, J.R.; McMurray, M.A.; Nelson, Z.W.; Gottschling, D.E. Mitochondrial Dysfunction Leads to Nuclear Genome Instability via an Iron-Sulfur Cluster Defect. Cell 2009, 137, 1247–1258. [CrossRef]
- Yien, Y.Y.; Shi, J.; Chen, C.; Cheung, J.T.M.; Grillo, A.S.; Shrestha, R.; Li, L.; Zhang, X.; Kafina, M.D.; Kingsley, P.D.; et al. FAM210B is an erythropoietin target and regulates erythroid heme synthesis by controlling mitochondrial iron import and ferrochelatase activity. Journal of Biological Chemistry 2018, 293, 19797–19811. [CrossRef]
- Todd, J.D.; Wexler, M.; Sawers, G.; Yeoman, K.H.; Poole, P.S.; Johnston, A.W.B. RirA, an iron-responsive regulator in the symbiotic bacterium Rhizobium leguminosarum. Microbiology (Reading) 2002, 148 (Pt 12), 4059–4071. [CrossRef]
- Mancias, J.D.; Vaites, L.P.; Nissim, S.; Biancur, D.E.; Kim, A.J.; Wang, X.; Liu, Y.; Goessling, W.; Kimmelman, A.C.; Harper, J.W. Ferritinophagy via NCOA4 is required for erythropoiesis and is regulated by iron dependent HERC2-mediated proteolysis. eLife 2015, 4, 1–19. [CrossRef]
- Zhang, D.L.; Ghosh, M.C.; Rouault, T.A. The physiological functions of iron regulatory proteins in iron homeostasis - an update. Front Pharmacol 2014, 5, 124. [CrossRef]
- Couturier, J.; Przybyla-Toscano, J.; Roret, T.; Didierjean, C.; Rouhier, N. The roles of glutaredoxins ligating Fe-S clusters: Sensing, transfer or repair functions? Biochimica et Biophysica Acta - Molecular Cell Research 2015, 1853, 1513–1527. [CrossRef]
- Jain, R.; Vanamee, E.S.; Dzikovski, B.G.; Buku, A.; Johnson, R.E.; Prakash, L.; Prakash, S.; Aggarwal, A.K. An iron-sulfur cluster in the polymerase domain of yeast DNA polymerase epsilon. J Mol Biol 2014, 426, 301–308. [CrossRef]
- Maio, N.; Lafont, B.A.P.; Sil, D.; Li, Y.; Bollinger, J.M., Jr.; Krebs, C.; Pierson, T.C.; Linehan, W.M.; Rouault, T.A. Fe-S cofactors in the SARS-CoV-2 RNA-dependent RNA polymerase are potential antiviral targets. Science 2021, 373, 236–241. [CrossRef]
- Maio, N.; Raza, M.K.; Li, Y.; Zhang, D.L.; Bollinger, J.M., Jr.; Krebs, C.; Rouault, T.A. An iron-sulfur cluster in the zinc-binding domain of the SARS-CoV-2 helicase modulates its RNA-binding and -unwinding activities. Proc Natl Acad Sci U S A 2023, 120, e2303860120. [CrossRef]
- Maio, N.; Cherry, S.; Schultz, D.C.; Hurst, B.L.; Linehan, W.M.; Rouault, T.A. TEMPOL inhibits SARS-CoV-2 replication and development of lung disease in the Syrian hamster model. iScience 2022, 25, 105074. [CrossRef]
- Rouault, T.A.; Klausner, R.D. Iron-sulfur clusters as biosensors of oxidants and iron. Trends Biochem Sci 1996, 21, 174–177. [CrossRef]
- Bak, D.W.; Elliott, S.J. Alternative FeS cluster ligands: tuning redox potentials and chemistry. Curr Opin Chem Biol 2014, 19, 50–58. [CrossRef]
- Gray, E.; Stewart, M.Y.Y.; Hanwell, L.; Crack, J.C.; Devine, R.; Stevenson, C.E.M.; Volbeda, A.; Johnston, A.W.B.; Fontecilla-Camps, J.C.; Hutchings, M.I.; et al. Stabilisation of the RirA [4Fe-4S] cluster results in loss of iron-sensing function. Chem Sci 2023, 14, 9744–9758. [CrossRef]
- Rouault, T.A. Mammalian iron-sulphur proteins: Novel insights into biogenesis and function. Nature Reviews Molecular Cell Biology 2015, 16, 45–55. [CrossRef]
- Maio, N.; Kim, K.S.; Singh, A.; Rouault, T.A. A Single Adaptable Cochaperone-Scaffold Complex Delivers Nascent Iron-Sulfur Clusters to Mammalian Respiratory Chain Complexes I–III. Cell Metabolism 2017, 25, 945-953.e946. [CrossRef]
- Ibrahim, I.M.; Wu, H.; Ezhov, R.; Kayanja, G.E.; Zakharov, S.D.; Du, Y.; Tao, W.A.; Pushkar, Y.; Cramer, W.A.; Puthiyaveetil, S. An evolutionarily conserved iron-sulfur cluster underlies redox sensory function of the Chloroplast Sensor Kinase. Commun Biol 2020, 3, 13. [CrossRef]
- Lu, Y. Assembly and Transfer of Iron-Sulfur Clusters in the Plastid. Front Plant Sci 2018, 9, 336. [CrossRef]
- Kim, K.S.; Maio, N.; Singh, A.; Rouault, T.A. Cytosolic HSC20 integrates de novo iron-sulfur cluster biogenesis with the CIAO1-mediated transfer to recipients. Human Molecular Genetics 2018, 27, 837–852. [CrossRef]
- Sharma, A.K.; Pallesen, L.J.; Spang, R.J.; Walden, W.E. Cytosolic iron-sulfur cluster assembly (CIA) system: Factors, mechanism, and relevance to cellular iron regulation. Journal of Biological Chemistry 2010, 285, 26745–26751. [CrossRef]
- Shi, R.; Hou, W.; Wang, Z.Q.; Xu, X. Biogenesis of Iron-Sulfur Clusters and Their Role in DNA Metabolism. Front Cell Dev Biol 2021, 9, 735678. [CrossRef]
- Maio, N.; Rouault, T.A. Iron-sulfur cluster biogenesis in mammalian cells: New insights into the molecular mechanisms of cluster delivery. Biochimica et Biophysica Acta - Molecular Cell Research 2015, 1853, 1493–1512. [CrossRef]
- Maio, N.; Rouault, T.A. Outlining the Complex Pathway of Mammalian Fe-S Cluster Biogenesis. Trends Biochem Sci 2020, 45, 411–426. [CrossRef]
- Rouault, T.A.; Tong, W.H. Iron-sulfur cluster biogenesis and human disease. Trends in Genetics 2008, 24, 398–407. [CrossRef]
- Uhrigshardt, H.; Singh, A.; Kovtunovych, G.; Ghosh, M.; Rouault, T.A. Characterization of the human HSC20, an unusual DnaJ type III protein, involved in iron-sulfur cluster biogenesis. Hum Mol Genet 2010, 19, 3816–3834. [CrossRef]
- Esquilin-Lebron, K.; Dubrac, S.; Barras, F.; Boyd, J.M. Bacterial Approaches for Assembling Iron-Sulfur Proteins. mBio 2021, 12, e0242521. [CrossRef]
- Jain, A.; Singh, A.; Maio, N.; Rouault, T.A. Assembly of the [4Fe-4S] cluster of NFU1 requires the coordinated donation of two [2Fe-2S] clusters from the scaffold proteins, ISCU2 and ISCA1. Hum Mol Genet 2020, 29, 3165–3182. [CrossRef]
- Legati, A.; Reyes, A.; Ceccatelli Berti, C.; Stehling, O.; Marchet, S.; Lamperti, C.; Ferrari, A.; Robinson, A.J.; Muhlenhoff, U.; Lill, R.; et al. A novel de novo dominant mutation in ISCU associated with mitochondrial myopathy. J Med Genet 2017, 54, 815–824. [CrossRef]
- Srour, B.; Gervason, S.; Hoock, M.H.; Monfort, B.; Want, K.; Larkem, D.; Trabelsi, N.; Landrot, G.; Zitolo, A.; Fonda, E.; et al. Iron Insertion at the Assembly Site of the ISCU Scaffold Protein Is a Conserved Process Initiating Fe-S Cluster Biosynthesis. J Am Chem Soc 2022, 144, 17496–17515. [CrossRef]
- Herrera, M.G.; Noguera, M.E.; Sewell, K.E.; Agudelo Suarez, W.A.; Capece, L.; Klinke, S.; Santos, J. Structure of the Human ACP-ISD11 Heterodimer. Biochemistry 2019, 58, 4596–4609. [CrossRef]
- Friemel, M.; Marelja, Z.; Li, K.; Leimkuhler, S. The N-Terminus of Iron-Sulfur Cluster Assembly Factor ISD11 Is Crucial for Subcellular Targeting and Interaction with l-Cysteine Desulfurase NFS1. Biochemistry 2017, 56, 1797–1808. [CrossRef]
- Hershkovitz, T.; Kurolap, A.; Tal, G.; Paperna, T.; Mory, A.; Staples, J.; Brigatti, K.W.; Regeneron Genetics, C.; Gonzaga-Jauregui, C.; Dumin, E.; et al. A recurring NFS1 pathogenic variant causes a mitochondrial disorder with variable intra-familial patient outcomes. Mol Genet Metab Rep 2021, 26, 100699. [CrossRef]
- Patra, S.; Barondeau, D.P. Mechanism of activation of the human cysteine desulfurase complex by frataxin. Proc Natl Acad Sci U S A 2019, 116, 19421–19430. [CrossRef]
- Maio, N.; Rouault, T.A. Mammalian iron sulfur cluster biogenesis: From assembly to delivery to recipient proteins with a focus on novel targets of the chaperone and co-chaperone proteins. IUBMB Life 2022, 74, 684–704. [CrossRef]
- Kakhlon, O.; Cabantchik, Z.I. The labile iron pool: Characterization, measurement, and participation in cellular processes. Free Radical Biology and Medicine 2002, 33, 1037–1046. [CrossRef]
- Philpott, C.C.; Protchenko, O.; Wang, Y.; Novoa-Aponte, L.; Leon-Torres, A.; Grounds, S.; Tietgens, A.J. Iron-tracking strategies: Chaperones capture iron in the cytosolic labile iron pool. Front Mol Biosci 2023, 10, 1127690. [CrossRef]
- Hider, R.C.; Kong, X.L. Glutathione: a key component of the cytoplasmic labile iron pool. BioMetals 2011, 24. [CrossRef]
- Patel, S.J.; Frey, A.G.; Palenchar, D.J.; Achar, S.; Bullough, K.Z.; Vashisht, A.; Wohlschlegel, J.A.; Philpott, C.C. A PCBP1-BolA2 chaperone complex delivers iron for cytosolic [2Fe-2S] cluster assembly. Nat Chem Biol 2019, 15, 872–881. [CrossRef]
- Li, H.; Mapolelo, D.T.; Randeniya, S.; Johnson, M.K.; Outten, C.E. Human glutaredoxin 3 forms [2Fe-2S]-bridged complexes with human BolA2. Biochemistry 2012, 51, 1687–1696. [CrossRef]
- Li, H.; Outten, C.E. Monothiol CGFS glutaredoxins and BolA-like proteins: [2Fe-2S] binding partners in iron homeostasis. Biochemistry 2012, 51, 4377–4389. [CrossRef]
- Sheftel, A.D.; Wilbrecht, C.; Stehling, O.; Niggemeyer, B.; Elsasser, H.P.; Muhlenhoff, U.; Lill, R. The human mitochondrial ISCA1, ISCA2, and IBA57 proteins are required for [4Fe-4S] protein maturation. Mol Biol Cell 2012, 23, 1157–1166. [CrossRef]
- Maio, N.; Singh, A.; Uhrigshardt, H.; Saxena, N.; Tong, W.H.; Rouault, T.A. Cochaperone binding to LYR motifs confers specificity of iron sulfur cluster delivery. Cell Metabolism 2014, 19, 445–457. [CrossRef]
- Zheng, L.; Cash, V.L.; Flint, D.H.; Dean, D.R. Assembly of Iron-Sulfur Clusters. Journal of Biological Chemistry 1998, 273, 13264–13272. [CrossRef]
- Frazzon, J.; Dean, D.R. Formation of iron-sulfur clusters in bacteria: an emerging field in bioinorganic chemistry. Curr Opin Chem Biol 2003, 7, 166–173. [CrossRef]
- Saito, H.; Sargent, T.; Parker, H.G.; Lawrence, J.H. Whole-body Iron Loss In Normal Man Measured with a Gamma Spectrometer'. Journal of Nuclear Medicine 1964, 5, 571–580.
- Andrews, N.C. The iron transporter DMT1. The international journal of biochemistry & cell biology 1999, 31, 991–994. [CrossRef]
- Zoller, H.; Theurl, I.; Koch, R.; Kaser, A.; Weiss, G. Mechanisms of iron mediated regulation of the duodenal iron transporters divalent metal transporter 1 and ferroportin 1. Blood cells, molecules & diseases 2002, 29, 488–497. [CrossRef]
- Ma, Y.; Yeh, M.; Yeh, K.-y.; Glass, J. Iron Imports. V. Transport of iron through the intestinal epithelium. American Journal of Physiology-Gastrointestinal and Liver Physiology 2006, 290, G417-G422. [CrossRef]
- Andrews, N.C. Iron homeostasis: Insights from genetics and animal models. Nature Reviews Genetics 2000, 1, 208–217. [CrossRef]
- Canonne-Hergaux, F.; Zhang, A.S.; Ponka, P.; Gros, P. Characterization of the iron transporter DMT1 (NRAMP2/DCT1) in red blood cells of normal and anemic mk/mk mice. Blood 2001, 98, 3823–3830. [CrossRef]
- Shaw, G.C.; Cope, J.J.; Li, L.; Corson, K.; Hersey, C.; Ackermann, G.E.; Gwynn, B.; Lambert, A.J.; Wingert, R.A.; Traver, D.; et al. Mitoferrin is essential for erythroid iron assimilation. Nature 2006, 440, 96–100. [CrossRef]
- Chung, J.; Anderson, S.A.; Gwynn, B.; Deck, K.M.; Chen, M.J.; Langer, N.B.; Shaw, G.C.; Huston, N.C.; Boyer, L.F.; Datta, S.; et al. Iron regulatory protein-1 protects against mitoferrin-1-deficient porphyria. Journal of Biological Chemistry 2014, 289, 7835–7843. [CrossRef]
- Friend, C.; Scher, W.; Holland, J.G.; Sato, T. Hemoglobin synthesis in murine virus-induced leukemic cells in vitro: stimulation of erythroid differentiation by dimethyl sulfoxide. Proceedings of the National Academy of Sciences of the United States of America 1971, 68, 378–382. [CrossRef]
- Levi, S.; Rovida, E. The role of iron in mitochondrial function. Biochimica et Biophysica Acta - General Subjects 2009, 1790, 629–636. [CrossRef]
- Hentze, M.W.; Muckenthaler, M.U.; Andrews, N.C. Balancing Acts: Molecular Control of Mammalian Iron Metabolism. Cell 2004, 117, 285–297. [CrossRef]
- Andrews, N.C.; Andrews, N.C. Forging a field : the golden age of iron biology ASH 50th anniversary review Forging a field : the golden age of iron biology. Blood 2009, 112, 219–230. [CrossRef]
- Knutson, M.D.; Oukka, M.; Koss, L.M.; Aydemir, F.; Wessling-Resnick, M. Iron release from macrophages after erythrophagocytosis is up-regulated by ferroportin 1 overexpression and down-regulated by hepcidin. Proceedings of the National Academy of Sciences 2005, 102, 1324–1328. [CrossRef]
- Donovan, A.; Brownlie, A.; Zhou, Y.; Shepard, J.; Pratt, S.J.; Moynihan, J.; Paw, B.H.; Drejer, A.; Barut, B.; Zapata, A.; et al. Positional cloning of zebrafish ferroportin1 identifies a conserved vertebrate iron exporter. Nature 2000, 403, 776–781. [CrossRef]
- Sabelli, M.; Montosi, G.; Garuti, C.; Caleffi, A.; Oliveto, S.; Biffo, S.; Pietrangelo, A. Human macrophage ferroportin biology and the basis for the ferroportin disease. Hepatology 2017, 65, 1512–1525. [CrossRef]
- Garrick, M.D.; Garrick, L.M. Cellular iron transport. Biochimica et Biophysica Acta - General Subjects 2009, 1790, 309–325. [CrossRef]
- Korolnek, T.; Hamza, I. Macrophages and iron trafficking at the birth and death of red cells. Blood 2015, 125, 2893–2897. [CrossRef]
- Garrick, M.D. Human iron transporters. Genes and Nutrition 2011, 6, 45–54. [CrossRef]
- Lane, D.J.R.; Merlot, A.M.; Huang, M.L.H.; Bae, D.H.; Jansson, P.J.; Sahni, S.; Kalinowski, D.S.; Richardson, D.R. Cellular iron uptake, trafficking and metabolism: Key molecules and mechanisms and their roles in disease. Biochimica et Biophysica Acta - Molecular Cell Research 2015, 1853, 1130–1144. [CrossRef]
- Kim, K.S.; Zhang, D.L.; Kovtunovych, G.; Ghosh, M.C.; Ollivierre, H.; Eckhaus, M.A.; Rouault, T.A. Infused wild-type macrophages reside and self-renew in the liver to rescue the hemolysis and anemia of Hmox1-deficient mice. Blood Adv 2018, 2, 2732–2743. [CrossRef]
- Kovtunovych, G.; Eckhaus, M.A.; Ghosh, M.C.; Ollivierre-Wilson, H.; Rouault, T.A. Dysfunction of the heme recycling system in heme oxygenase 1-deficient mice: effects on macrophage viability and tissue iron distribution. Blood 2010, 116, 6054–6062. [CrossRef]
- Kovtunovych, G.; Ghosh, M.C.; Ollivierre, W.; Weitzel, R.P.; Eckhaus, M.A.; Tisdale, J.F.; Yachie, A.; Rouault, T.A. Wild-type macrophages reverse disease in heme oxygenase 1-deficient mice. Blood 2014, 124, 1522–1530. [CrossRef]
- Papanikolaou, G.; Pantopoulos, K. Iron metabolism and toxicity. Toxicology and Applied Pharmacology 2005, 202, 199–211. [CrossRef]
- Cherayil, B.J. Iron and immunity: Immunological consequences of iron deficiency and overload. Archivum Immunologiae et Therapiae Experimentalis 2010, 58, 407–415. [CrossRef]
- Gattermann, N.; Muckenthaler, M.U.; Kulozik, A.E.; Metzgeroth, G.; Hastka, J. The Evaluation of Iron Deficiency and Iron Overload. Dtsch Arztebl Int 2021, 118, 847–856. [CrossRef]
- Pasricha, S.R.; Tye-Din, J.; Muckenthaler, M.U.; Swinkels, D.W. Iron deficiency. Lancet 2021, 397, 233–248. [CrossRef]
- Bresgen, N.; Eckl, P.M. Oxidative stress and the homeodynamics of iron metabolism. Biomolecules 2015, 5, 808–847. [CrossRef]
- Steinbicker, A.U.; Muckenthaler, M.U. Out of balance-systemic iron homeostasis in iron-related disorders. Nutrients 2013, 5, 3034–3061. [CrossRef]
- Mackenzie, E.L.; Iwasaki, K.; Tsuji, Y. Intracellular Iron Transport and Storage: From Molecular Mechanisms to Health Implications. Antioxidants & Redox Signaling 2008, 10, 997–1030. [CrossRef]
- Aisen, P.; Enns, C.; Wessling-Resnick, M. Chemistry and biology of eukaryotic iron metabolism. International Journal of Biochemistry and Cell Biology 2001, 33, 940–959. [CrossRef]
- Pietrangelo, A. Hereditary Hemochromatosis — A New Look at an Old Disease. New England Journal of Medicine 2004, 350, 2383–2397. [CrossRef]
- Haley, H.M.S.S.; Hill, A.G.; Greenwood, A.I.; Woerly, E.M.; Rienstra, C.M.; Burke, M.D. Peridinin Is an Exceptionally Potent and Membrane-Embedded Inhibitor of Bilayer Lipid Peroxidation. Journal of the American Chemical Society 2018, 140, 15227–15240. [CrossRef]
- Muckenthaler, M.U.; Galy, B.; Hentze, M.W. Systemic Iron Homeostasis and the Iron-Responsive Element/Iron-Regulatory Protein (IRE/IRP) Regulatory Network. Annual Review of Nutrition 2008, 28, 197–213. [CrossRef]
- Hentze, M.W.; Muckenthaler, M.U.; Galy, B.; Camaschella, C. Two to Tango: Regulation of Mammalian Iron Metabolism. Cell 2010, 142, 24–38. [CrossRef]
- Rouault, T.A. The role of iron regulatory proteins in mammalian iron homeostasis and disease. Nature Chemical Biology 2006, 2, 406–414. [CrossRef]
- Nairz, M.; Haschka, D.; Demetz, E.; Weiss, G. Iron at the interface of immunity and infection. Frontiers in Pharmacology 2014, 5 JUL, 1-11. [CrossRef]
- Nairz, M.; Weiss, G. Iron in infection and immunity. Mol Aspects Med 2020, 75, 100864. [CrossRef]
- Marchetti, M.; De Bei, O.; Bettati, S.; Campanini, B.; Kovachka, S.; Gianquinto, E.; Spyrakis, F.; Ronda, L. Iron Metabolism at the Interface between Host and Pathogen: From Nutritional Immunity to Antibacterial Development. Int J Mol Sci 2020, 21. [CrossRef]
- Gerner, R.R.; Nuccio, S.P.; Raffatellu, M. Iron at the host-microbe interface. Mol Aspects Med 2020, 75, 100895. [CrossRef]
- Iatsenko, I.; Marra, A.; Boquete, J.P.; Pena, J.; Lemaitre, B. Iron sequestration by transferrin 1 mediates nutritional immunity in Drosophila melanogaster. Proc Natl Acad Sci USA 2020, 117, 7317–7325. [CrossRef]
- Caza, M.; Kronstad, J.W. Shared and distinct mechanisms of iron acquisition by bacterial and fungal pathogens of humans. Frontiers in Cellular and Infection Microbiology 2013, 3, 1–23. [CrossRef]
- Lesuisse, E.; Blaiseau, P.L.; Dancis, A.; Camadro, J.M. Siderophore uptake and use by the yeast Saccharomyces cerevisiae. Microbiology (Reading) 2001, 147 (Pt 12), 289–298. [CrossRef]
- Wilson, B.R.; Bogdan, A.R.; Miyazawa, M.; Hashimoto, K.; Tsuji, Y. Siderophores in Iron Metabolism: From Mechanism to Therapy Potential. Trends Mol Med 2016, 22, 1077–1090. [CrossRef]
- Martinelli, M.; Strisciuglio, C.; Alessandrella, A.; Rossi, F.; Auricchio, R.; Campostrini, N.; Girelli, D.; Nobili, B.; Staiano, A.; Perrotta, S.; et al. Serum hepcidin and iron absorption in paediatric inflammatory bowel disease. Journal of Crohn's and Colitis 2016, 10, 566–574. [CrossRef]
- Nakashige, T.G.; Zhang, B.; Krebs, C.; Nolan, E.M. Human calprotectin is an iron-sequestering host-defense protein. Nature Chemical Biology 2015, 11, 765–771. [CrossRef]
- Arosio, P.; Elia, L.; Poli, M. Ferritin, cellular iron storage and regulation. IUBMB Life 2017, 69, 414–422. [CrossRef]
- Andrews, N. Disorders of Iron Metabolism. The New England journal of medicine 1999, 341, 1986–1995. [CrossRef]
- Jabara, H.H.; Boyden, S.E.; Chou, J.; Ramesh, N.; Massaad, M.J.; Benson, H.; Bainter, W.; Fraulino, D.; Rahimov, F.; Sieff, C.; et al. A missense mutation in TFRC, encoding transferrin receptor 1, causes combined immunodeficiency. Nat Genet 2016, 48, 74–78. [CrossRef]
- Wessling-Resnick, M. Nramp1 and other transporters involved in metal withholding during infection. Journal of Biological Chemistry 2015, 290, 18984–18990. [CrossRef]
- Brittenham, G.M.; Andersson, M.; Egli, I.; Foman, J.T.; Zeder, C.; Westerman, M.E.; Hurrell, R.F. Circulating non-transferrin-bound iron after oral administration of supplemental and fortification doses of iron to healthy women: a randomized study. Am J Clin Nutr 2014, 100, 813–820. [CrossRef]
- Weinberg, E.D. Nutritional immunity. Host's attempt to withold iron from microbial invaders. JAMA 1975, 231, 39–41. [CrossRef]
- Murdoch, C.C.; Skaar, E.P. Nutritional immunity: the battle for nutrient metals at the host-pathogen interface. Nat Rev Microbiol 2022, 20, 657–670. [CrossRef]
- Anderson, G.J. Ironing Out Disease: Inherited Disorders of Iron Homeostasis. IUBMB Life (International Union of Biochemistry and Molecular Biology: Life) 2001, 51, 11–17. [CrossRef]
- Brissot, P.; Bardou-Jacquet, E.; Jouanolle, A.M.; Loréal, O. Iron disorders of genetic origin: A changing world. Trends in Molecular Medicine 2011, 17, 707–713. [CrossRef]
- Grillo, A.S.; SantaMaria, A.M.; Kafina, M.D.; Cioffi, A.G.; Huston, N.C.; Han, M.; Seo, Y.A.; Yien, Y.Y.; Nardone, C.; Menon, A.V.; et al. Restored iron transport by a small molecule promotes absorption and hemoglobinization in animals. Science 2017, 356, 608–616. [CrossRef]
- Gangat, N.; Wolanskyj, A.P. Anemia of chronic disease. Seminars in Hematology 2013, 50, 232–238. [CrossRef]
- Rivera, S.; Ganz, T. Animal Models of Anemia of Inflammation. Seminars in Hematology 2009, 46, 351–357. [CrossRef]
- Anderson, Sheila A.; Nizzi, Christopher P.; Chang, Y.-I.; Deck, Kathryn M.; Schmidt, Paul J.; Galy, B.; Damnernsawad, A.; Broman, Aimee T.; Kendziorski, C.; Hentze, Matthias W.; et al. The IRP1-HIF-2α Axis Coordinates Iron and Oxygen Sensing with Erythropoiesis and Iron Absorption. Cell Metabolism 2013, 17. [CrossRef]
- Crack, J.C.; Green, J.; Thomson, A.J.; Le Brun, N.E. Iron-sulfur clusters as biological sensors: the chemistry of reactions with molecular oxygen and nitric oxide. Acc Chem Res 2014, 47, 3196–3205. [CrossRef]
- Ghosh, M.C.; Zhang, D.; Ollivierre, H.; Noguchi, A.; Springer, D.; Linehan, W.M.; Rouault, T.A. Therapeutic inhibition of HIF2α reverses polycythemia and pulmonary hypertension in murine models of two human diseases. Blood 2020, 10.1182/blood.2020009138. [CrossRef]
- Wilkinson, N.; Pantopoulos, K. IRP1 regulates erythropoiesis and systemic iron homeostasis by controlling HIF2α mRNA translation. Blood 2013, 122. [CrossRef]
- Nandal, A.; Ruiz, J.C.; Subramanian, P.; Ghimire-Rijal, S.; Sinnamon, R.A.; Stemmler, T.L.; Bruick, R.K.; Philpott, C.C. Activation of the HIF prolyl hydroxylase by the iron chaperones PCBP1 and PCBP2. Cell Metab 2011, 14, 647–657. [CrossRef]
- Wang, D.; Wang, L.H.; Zhao, Y.; Lu, Y.P.; Zhu, L. Hypoxia regulates the ferrous iron uptake and reactive oxygen species level via divalent metal transporter 1 (DMT1) exon1B by hypoxia-inducible factor-1. IUBMB Life 2010, 62, 629–636. [CrossRef]
- Mastrogiannaki, M.; Matak, P.; Keith, B.; Simon, M.C.; Vaulont, S.; Peyssonnaux, C. HIF-2α, but not HIF-1α, promotes iron absorption in mice. Journal of Clinical Investigation 2009, 119. [CrossRef]
- Sanchez, M.; Galy, B.; Muckenthaler, M.U.; Hentze, M.W. Iron-regulatory proteins limit hypoxia-inducible factor-2α expression in iron deficiency. Nature Structural and Molecular Biology 2007, 14, 420–426. [CrossRef]
- Ghosh, M.C.; Zhang, D.-L.; Ollivierre, H.; Eckhaus, M.A.; Rouault, T.A. Translational repression of HIF2α expression in mice with Chuvash polycythemia reverses polycythemia. Journal of Clinical Investigation 2018, 128. [CrossRef]
- Holmes-Hampton, G.P.; Ghosh, M.C.; Rouault, T.A. Methods for Studying Iron Regulatory Protein 1: An Important Protein in Human Iron Metabolism. Methods Enzymol 2018, 599, 139–155. [CrossRef]
- Johnson, N.B.; Deck, K.M.; Nizzi, C.P.; Eisenstein, R.S. A synergistic role of IRP1 and FBXL5 proteins in coordinating iron metabolism during cell proliferation. Journal of Biological Chemistry 2017, 292. [CrossRef]
- Sanchez, M.; Galy, B.; Schwanhaeusser, B.; Blake, J.; Bähr-Ivacevic, T.; Benes, V.; Selbach, M.; Muckenthaler, M.U.; Hentze, M.W. Iron regulatory protein-1 and -2: transcriptome-wide definition of binding mRNAs and shaping of the cellular proteome by iron regulatory proteins. Blood 2011, 118. [CrossRef]
- Pantopoulos, K. Iron Metabolism and the IRE/IRP Regulatory System: An Update. Annals of the New York Academy of Sciences 2004, 1012, 1–13. [CrossRef]
- Pellicer Martinez, M.T.; Martinez, A.B.; Crack, J.C.; Holmes, J.D.; Svistunenko, D.A.; Johnston, A.W.B.; Cheesman, M.R.; Todd, J.D.; Le Brun, N.E. Sensing iron availability via the fragile [4Fe-4S] cluster of the bacterial transcriptional repressor RirA. Chem Sci 2017, 8, 8451–8463. [CrossRef]
- Hibbing, M.E.; Fuqua, C. Antiparallel and interlinked control of cellular iron levels by the Irr and RirA regulators of Agrobacterium tumefaciens. J Bacteriol 2011, 193, 3461–3472. [CrossRef]
- Rudolph, G.; Hennecke, H.; Fischer, H.M. Beyond the Fur paradigm: iron-controlled gene expression in rhizobia. FEMS Microbiol Rev 2006, 30, 631–648. [CrossRef]
- Vergnes, A.; Viala, J.P.; Ouadah-Tsabet, R.; Pocachard, B.; Loiseau, L.; Meresse, S.; Barras, F.; Aussel, L. The iron-sulfur cluster sensor IscR is a negative regulator of Spi1 type III secretion system in Salmonella enterica. Cell Microbiol 2017, 19. [CrossRef]
- Saninjuk, K.; Romsang, A.; Duang-Nkern, J.; Vattanaviboon, P.; Mongkolsuk, S. Transcriptional regulation of the Pseudomonas aeruginosa iron-sulfur cluster assembly pathway by binding of IscR to multiple sites. PLoS One 2019, 14, e0218385. [CrossRef]
- Outten, C.E.; Albetel, A.N. Iron sensing and regulation in Saccharomyces cerevisiae: Ironing out the mechanistic details. Curr Opin Microbiol 2013, 16, 662–668. [CrossRef]
- Yamaguchi-Iwai, Y.; Dancis, A.; Klausner, R.D. AFT1: a mediator of iron regulated transcriptional control in Saccharomyces cerevisiae. EMBO J 1995, 14, 1231–1239. [CrossRef]
- Gupta, M.; Outten, C.E. Iron-sulfur cluster signaling: The common thread in fungal iron regulation. Curr Opin Chem Biol 2020, 55, 189–201. [CrossRef]
- Poor, C.B.; Wegner, S.V.; Li, H.; Dlouhy, A.C.; Schuermann, J.P.; Sanishvili, R.; Hinshaw, J.R.; Riggs-Gelasco, P.J.; Outten, C.E.; He, C. Molecular mechanism and structure of the Saccharomyces cerevisiae iron regulator Aft2. Proc Natl Acad Sci U S A 2014, 111, 4043–4048. [CrossRef]
- Shen, M.; Goforth, J.B.; Eisenstein, R.S. Iron-dependent post transcriptional control of mitochondrial aconitase expression. Metallomics 2023, 15. [CrossRef]
- Zimmer, M.; Ebert, B.L.; Neil, C.; Brenner, K.; Papaioannou, I.; Melas, A.; Tolliday, N.; Lamb, J.; Pantopoulos, K.; Golub, T.; et al. Small-molecule inhibitors of HIF-2a translation link its 5'UTR iron-responsive element to oxygen sensing. Mol Cell 2008, 32, 838–848. [CrossRef]
- Zhang, D.L.; Hughes, R.M.; Ollivierre-Wilson, H.; Ghosh, M.C.; Rouault, T.A. A Ferroportin Transcript that Lacks an Iron-Responsive Element Enables Duodenal and Erythroid Precursor Cells to Evade Translational Repression. Cell Metabolism 2009, 9, 461–473. [CrossRef]
- Garza, K.R.; Clarke, S.L.; Ho, Y.H.; Bruss, M.D.; Vasanthakumar, A.; Anderson, S.A.; Eisenstein, R.S. Differential translational control of 5' IRE-containing mRNA in response to dietary iron deficiency and acute iron overload. Metallomics 2020, 12, 2186–2198. [CrossRef]
- Cmejla, R.; Petrak, J.; Cmejlova, J. A novel iron responsive element in the 3′UTR of human MRCKα. Biochemical and Biophysical Research Communications 2006, 341. [CrossRef]
- Sanchez, M.; Galy, B.; Dandekar, T.; Bengert, P.; Vainshtein, Y.; Stolte, J.; Muckenthaler, M.U.; Hentze, M.W. Iron Regulation and the Cell Cycle. Journal of Biological Chemistry 2006, 281. [CrossRef]
- Gunshin, H.; Mackenzie, B.; Berger, U.V.; Gunshin, Y.; Romero, M.F.; Boron, W.F.; Nussberger, S.; Gollan, J.L.; Hediger, M.A. Cloning and characterization of a mammalian proton-coupled metal-ion transporter. Nature 1997, 388. [CrossRef]
- Tybl, E.; Gunshin, H.; Gupta, S.; Barrientos, T.; Bonadonna, M.; Celma Nos, F.; Palais, G.; Karim, Z.; Sanchez, M.; Andrews, N.C.; et al. Control of Systemic Iron Homeostasis by the 3' Iron-Responsive Element of Divalent Metal Transporter 1 in Mice. Hemasphere 2020, 4, e459. [CrossRef]
- Qatato, M.; Bonadonna, M.; Palais, G.; Ertl, A.; Schmidt, G.; Polycarpou-Schwarz, M.; Karim, Z.; Galy, B. IRE-dependent Regulation of Intestinal Dmt1 Prevails During Chronic Dietary Iron Deficiency but is Dispensable in Conditions of Acute Erythropoietic Stress. Hemasphere 2022, 6, e693. [CrossRef]
- Garrick, M.D.; Dolan, K.G.; Horbinski, C.; Ghio, A.J.; Higgins, D.; Porubcin, M.; Moore, E.G.; Hainsworth, L.N.; Umbreit, J.N.; Conrad, M.E.; et al. DMT1: A mammalian transporter for multiple metals. BioMetals 2003, 16, 41–54. [CrossRef]
- Ducamp, S.; Luscieti, S.; Ferrer-Cortès, X.; Nicolas, G.; Manceau, H.; Peoc’h, K.; Yien, Y.Y.; Kannengiesser, C.; Gouya, L.; Puy, H.; et al. A mutation in the iron-responsive element of ALAS2 is a modifier of disease severity in a patient suffering from CLPX associated erythropoietic protoporphyria. Haematologica 2021, 10.3324/haematol.2020.272450. [CrossRef]
- Wilkinson, N.; Pantopoulos, K. The IRP/IRE system in vivo: insights from mouse models. Front Pharmacol 2014, 5, 176. [CrossRef]
- Kaptain, S.; Downey, W.E.; Tang, C.; Philpott, C.; Haile, D.; Orloff, D.G.; Harford, J.B.; Rouault, T.A.; Klausner, R.D. A regulated RNA binding protein also possesses aconitase activity. Proc Natl Acad Sci U S A 1991, 88, 10109–10113. [CrossRef]
- Rouault, T.A.; Maio, N. How Oxidation of a Unique Iron-Sulfur Cluster in FBXL5 Regulates IRP2 Levels and Promotes Regulation of Iron Metabolism Proteins. Mol Cell 2020, 78, 1–3. [CrossRef]
- Zumbrennen-Bullough, K.B.; Becker, L.; Garrett, L.; Hölter, S.M.; Calzada-Wack, J.; Mossbrugger, I.; Quintanilla-Fend, L.; Racz, I.; Rathkolb, B.; Klopstock, T.; et al. Abnormal Brain Iron Metabolism in Irp2 Deficient Mice Is Associated with Mild Neurological and Behavioral Impairments. PLoS ONE 2014, 9. [CrossRef]
- Henderson, B.R. Iron regulatory proteins 1 and 2. Bioessays 1996, 18, 739–746. [CrossRef]
- Meyron-Holtz, E.G.; Ghosh, M.C.; Iwai, K.; LaVaute, T.; Brazzolotto, X.; Berger, U.V.; Land, W.; Ollivierre-Wilson, H.; Grinberg, A.; Love, P.; et al. Genetic ablations of iron regulatory proteins 1 and 2 reveal why iron regulatory protein 2 dominates iron homeostasis. The EMBO Journal 2004, 23. [CrossRef]
- Cluster, O.-r.; Wang, H.; Shi, H.; Rajan, M.; Stoll, S.; Leibold, E.A. Article FBXL5 Regulates IRP2 Stability in Iron Homeostasis. Molecular Cell 2020, 10.1016/j.molcel.2020.02.011, 1-11. [CrossRef]
- Holland, L.Z.; Ocampo Daza, D. A new look at an old question: when did the second whole genome duplication occur in vertebrate evolution? Genome Biol 2018, 19, 209. [CrossRef]
- Hsiao, T.L.; Vitkup, D. Role of duplicate genes in robustness against deleterious human mutations. PLoS Genetics 2008, 4. [CrossRef]
- Wang, H.; Shi, H.; Rajan, M.; Canarie, E.R.; Hong, S.; Simoneschi, D.; Pagano, M.; Bush, M.F.; Stoll, S.; Leibold, E.A.; et al. FBXL5 Regulates IRP2 Stability in Iron Homeostasis via an Oxygen-Responsive [2Fe2S] Cluster. Mol Cell 2020, 78, 31–41 e35. [CrossRef]
- Jiao, Q.; Du, X.; Wei, J.; Li, Y.; Jiang, H. Oxidative Stress Regulated Iron Regulatory Protein IRP2 Through FBXL5-Mediated Ubiquitination-Proteasome Way in SH-SY5Y Cells. Front Neurosci 2019, 13, 20. [CrossRef]
- Chollangi, S.; Thompson, J.W.; Ruiz, J.C.; Gardner, K.H.; Bruick, R.K. Hemerythrin-like Domain within F-box and Leucine-rich Repeat Protein 5 (FBXL5) Communicates Cellular Iron and Oxygen Availability by Distinct Mechanisms. Journal of Biological Chemistry 2012, 287. [CrossRef]
- Moroishi, T.; Nishiyama, M.; Takeda, Y.; Iwai, K.; Nakayama, Keiichi I. The FBXL5-IRP2 Axis Is Integral to Control of Iron Metabolism In Vivo. Cell Metabolism 2011, 14. [CrossRef]
- Leon-Sicairos, C.R.; Figueroa-Angulo, E.E.; Calla-Choque, J.S.; Arroyo, R. The Non-Canonical Iron-Responsive Element of IRE-tvcp12 Hairpin Structure at the 3'-UTR of Trichomonas vaginalis TvCP12 mRNA That Binds TvHSP70 and TvACTN-3 Can Regulate mRNA Stability and Amount of Protein. Pathogens 2023, 12. [CrossRef]
- Del Rey, M.Q.; Mancias, J.D. NCOA4-mediated ferritinophagy: A potential link to neurodegeneration. In Frontiers in Neuroscience, Frontiers Media S.A.: 2019; Vol. 13.
- Mancias, J.D.; Wang, X.; Gygi, S.P.; Harper, J.W.; Kimmelman, A.C. Quantitative proteomics identifies NCOA4 as the cargo receptor mediating ferritinophagy. Nature 2014, 508, 105–109. [CrossRef]
- Santana-Codina, N.; Mancias, J.D. The role of NCOA4-mediated ferritinophagy in health and disease; 2018; Vol. 11.
- Santana-Codina, N.; Del Rey, M.Q.; Kapner, K.S.; Zhang, H.; Gikandi, A.; Malcolm, C.; Poupault, C.; Kuljanin, M.; John, K.M.; Biancur, D.E.; et al. NCOA4-Mediated Ferritinophagy Is a Pancreatic Cancer Dependency via Maintenance of Iron Bioavailability for Iron-Sulfur Cluster Proteins. Cancer Discov 2022, 12, 2180–2197. [CrossRef]
- Kuno, S.; Iwai, K. Oxygen modulates iron homeostasis by switching iron sensing of NCOA4. J Biol Chem 2023, 299, 104701. [CrossRef]
- Zhao, H.; Lu, Y.; Zhang, J.; Sun, Z.; Cheng, C.; Liu, Y.; Wu, L.; Zhang, M.; He, W.; Hao, S.; et al. NCOA4 requires a [3Fe-4S] to sense and maintain the iron homeostasis. J Biol Chem 2023, 300, 105612. [CrossRef]
- Andrews, S.C.; Robinson, A.K.; Rodriguez-Quinones, F. Bacterial iron homeostasis. FEMS Microbiol Rev 2003, 27, 215–237. [CrossRef]
- Bradley, J.M.; Svistunenko, D.A.; Wilson, M.T.; Hemmings, A.M.; Moore, G.R.; Le Brun, N.E. Bacterial iron detoxification at the molecular level. J Biol Chem 2020, 295, 17602–17623. [CrossRef]
- Small, S.K.; Puri, S.; Sangwan, I.; O'Brian, M.R. Positive control of ferric siderophore receptor gene expression by the Irr protein in Bradyrhizobium japonicum. J Bacteriol 2009, 191, 1361–1368. [CrossRef]
- Yeoman, K.H.; Curson, A.R.; Todd, J.D.; Sawers, G.; Johnston, A.W. Evidence that the Rhizobium regulatory protein RirA binds to cis-acting iron-responsive operators (IROs) at promoters of some Fe-regulated genes. Microbiology (Reading) 2004, 150 (Pt 12), 4065–4074. [CrossRef]
- Pankievicz, V.C.S.; Irving, T.B.; Maia, L.G.S.; Ane, J.M. Are we there yet? The long walk towards the development of efficient symbiotic associations between nitrogen-fixing bacteria and non-leguminous crops. BMC Biol 2019, 17, 99. [CrossRef]
- Sevilla, E.; Bes, M.T.; Gonzalez, A.; Peleato, M.L.; Fillat, M.F. Redox-Based Transcriptional Regulation in Prokaryotes: Revisiting Model Mechanisms. Antioxid Redox Signal 2019, 30, 1651–1696. [CrossRef]
- Shimberg, G.D.; Pritts, J.D.; Michel, S.L.J. Iron-Sulfur Clusters in Zinc Finger Proteins. Methods Enzymol 2018, 599, 101–137. [CrossRef]
- Kaplan, C.D.; Kaplan, J. Iron acquisition and transcriptional regulation. Chem Rev 2009, 109, 4536–4552. [CrossRef]
- Pijuan, J.; Moreno, D.F.; Yahya, G.; Moisa, M.; Ul Haq, I.; Krukiewicz, K.; Mosbah, R.; Metwally, K.; Cavalu, S. Regulatory and pathogenic mechanisms in response to iron deficiency and excess in fungi. Microb Biotechnol 2023, 16, 2053–2071. [CrossRef]
- Ramos-Alonso, L.; Romero, A.M.; Martinez-Pastor, M.T.; Puig, S. Iron Regulatory Mechanisms in Saccharomyces cerevisiae. Front Microbiol 2020, 11, 582830. [CrossRef]
- Askwith, C.; Eide, D.; Van Ho, A.; Bernard, P.S.; Li, L.; Davis-Kaplan, S.; Sipe, D.M.; Kaplan, J. The FET3 gene of S. cerevisiae encodes a multicopper oxidase required for ferrous iron uptake. Cell 1994, 76, 403–410. [CrossRef]
- Stearman, R.; Yuan, D.S.; Yamaguchi-Iwai, Y.; Klausner, R.D.; Dancis, A. A permease-oxidase complex involved in high-affinity iron uptake in yeast. Science 1996, 271, 1552–1557. [CrossRef]
- Heymann, P.; Ernst, J.F.; Winkelmann, G. Identification and substrate specificity of a ferrichrome-type siderophore transporter (Arn1p) in Saccharomyces cerevisiae. FEMS Microbiol Lett 2000, 186, 221–227. [CrossRef]
- Yun, C.W.; Tiedeman, J.S.; Moore, R.E.; Philpott, C.C. Siderophore-iron uptake in saccharomyces cerevisiae. Identification of ferrichrome and fusarinine transporters. J Biol Chem 2000, 275, 16354–16359. [CrossRef]
- Martinez-Pastor, M.T.; Perea-Garcia, A.; Puig, S. Mechanisms of iron sensing and regulation in the yeast Saccharomyces cerevisiae. World J Microbiol Biotechnol 2017, 33, 75. [CrossRef]
- Rodrigues-Pousada, C.; Devaux, F.; Caetano, S.M.; Pimentel, C.; da Silva, S.; Cordeiro, A.C.; Amaral, C. Yeast AP-1 like transcription factors (Yap) and stress response: a current overview. Microb Cell 2019, 6, 267–285. [CrossRef]
- Rietzschel, N.; Pierik, A.J.; Bill, E.; Lill, R.; Muhlenhoff, U. The basic leucine zipper stress response regulator Yap5 senses high-iron conditions by coordination of [2Fe-2S] clusters. Mol Cell Biol 2015, 35, 370–378. [CrossRef]
- Sorribes-Dauden, R.; Peris, D.; Martinez-Pastor, M.T.; Puig, S. Structure and function of the vacuolar Ccc1/VIT1 family of iron transporters and its regulation in fungi. Comput Struct Biotechnol J 2020, 18, 3712–3722. [CrossRef]
- Velez-Bermudez, I.C.; Schmidt, W. Iron sensing in plants. Front Plant Sci 2023, 14, 1145510. [CrossRef]
- Senoura, T.; Kobayashi, T.; An, G.; Nakanishi, H.; Nishizawa, N.K. Defects in the rice aconitase-encoding OsACO1 gene alter iron homeostasis. Plant Mol Biol 2020, 104, 629–645. [CrossRef]
- Inoue, H.; Higuchi, K.; Takahashi, M.; Nakanishi, H.; Mori, S.; Nishizawa, N.K. Three rice nicotianamine synthase genes, OsNAS1, OsNAS2, and OsNAS3 are expressed in cells involved in long-distance transport of iron and differentially regulated by iron. Plant J 2003, 36, 366–381. [CrossRef]
- Inoue, H.; Kobayashi, T.; Nozoye, T.; Takahashi, M.; Kakei, Y.; Suzuki, K.; Nakazono, M.; Nakanishi, H.; Mori, S.; Nishizawa, N.K. Rice OsYSL15 is an iron-regulated iron(III)-deoxymugineic acid transporter expressed in the roots and is essential for iron uptake in early growth of the seedlings. J Biol Chem 2009, 284, 3470–3479. [CrossRef]
- Ogo, Y.; Itai, R.N.; Nakanishi, H.; Kobayashi, T.; Takahashi, M.; Mori, S.; Nishizawa, N.K. The rice bHLH protein OsIRO2 is an essential regulator of the genes involved in Fe uptake under Fe-deficient conditions. Plant J 2007, 51, 366–377. [CrossRef]
- Battistuzzi, F.U.; Feijao, A.; Hedges, S.B. A genomic timescale of prokaryote evolution: insights into the origin of methanogenesis, phototrophy, and the colonization of land. BMC Evol Biol 2004, 4, 44. [CrossRef]
- Brochier-Armanet, C.; Forterre, P.; Gribaldo, S. Phylogeny and evolution of the Archaea: one hundred genomes later. Curr Opin Microbiol 2011, 14, 274–281. [CrossRef]
- Hoshino, Y.; Villanueva, L. Four billion years of microbial terpenome evolution. FEMS Microbiol Rev 2023, 47. [CrossRef]
- Johnson, C.; England, A.; Munro-Ehrlich, M.; Colman, D.R.; DuBois, J.L.; Boyd, E.S. Pathways of Iron and Sulfur Acquisition, Cofactor Assembly, Destination, and Storage in Diverse Archaeal Methanogens and Alkanotrophs. J Bacteriol 2021, 203, e0011721. [CrossRef]
- Roche, B.; Aussel, L.; Ezraty, B.; Mandin, P.; Py, B.; Barras, F. Iron/sulfur proteins biogenesis in prokaryotes: formation, regulation and diversity. Biochim Biophys Acta 2013, 1827, 455–469. [CrossRef]
- Baker, B.J.; De Anda, V.; Seitz, K.W.; Dombrowski, N.; Santoro, A.E.; Lloyd, K.G. Diversity, ecology and evolution of Archaea. Nat Microbiol 2020, 5, 887–900. [CrossRef]
- Iwasaki, T. Iron-sulfur world in aerobic and hyperthermoacidophilic archaea Sulfolobus. Archaea 2010, 2010. [CrossRef]
- Saini, J.; Deere, T.M.; Lessner, D.J. The minimal SUF system is not required for Fe-S cluster biogenesis in the methanogenic archaeon Methanosarcina acetivorans. Sci Rep 2023, 13, 15120. [CrossRef]
- Uhrigshardt, H.; Zoske, S.; Anemuller, S. Sulfolobus aconitase, a regulator of iron metabolism? Biochem Soc Trans 2002, 30, 685–687. [CrossRef]
Figure 1.
A timeline illustrating select discoveries in the Fe-S cluster field ranging from the discovery of EPR and Mossbauer effects to recent discoveries of environmental sensing Fe-S clusters. Since their discovery in 1960, Fe-S clusters have been found to perform a variety of essential functions in all domains of life. With new scientific techniques and greater awareness of their important role within biology the pace of discovery has increased exponentially.
Figure 1.
A timeline illustrating select discoveries in the Fe-S cluster field ranging from the discovery of EPR and Mossbauer effects to recent discoveries of environmental sensing Fe-S clusters. Since their discovery in 1960, Fe-S clusters have been found to perform a variety of essential functions in all domains of life. With new scientific techniques and greater awareness of their important role within biology the pace of discovery has increased exponentially.
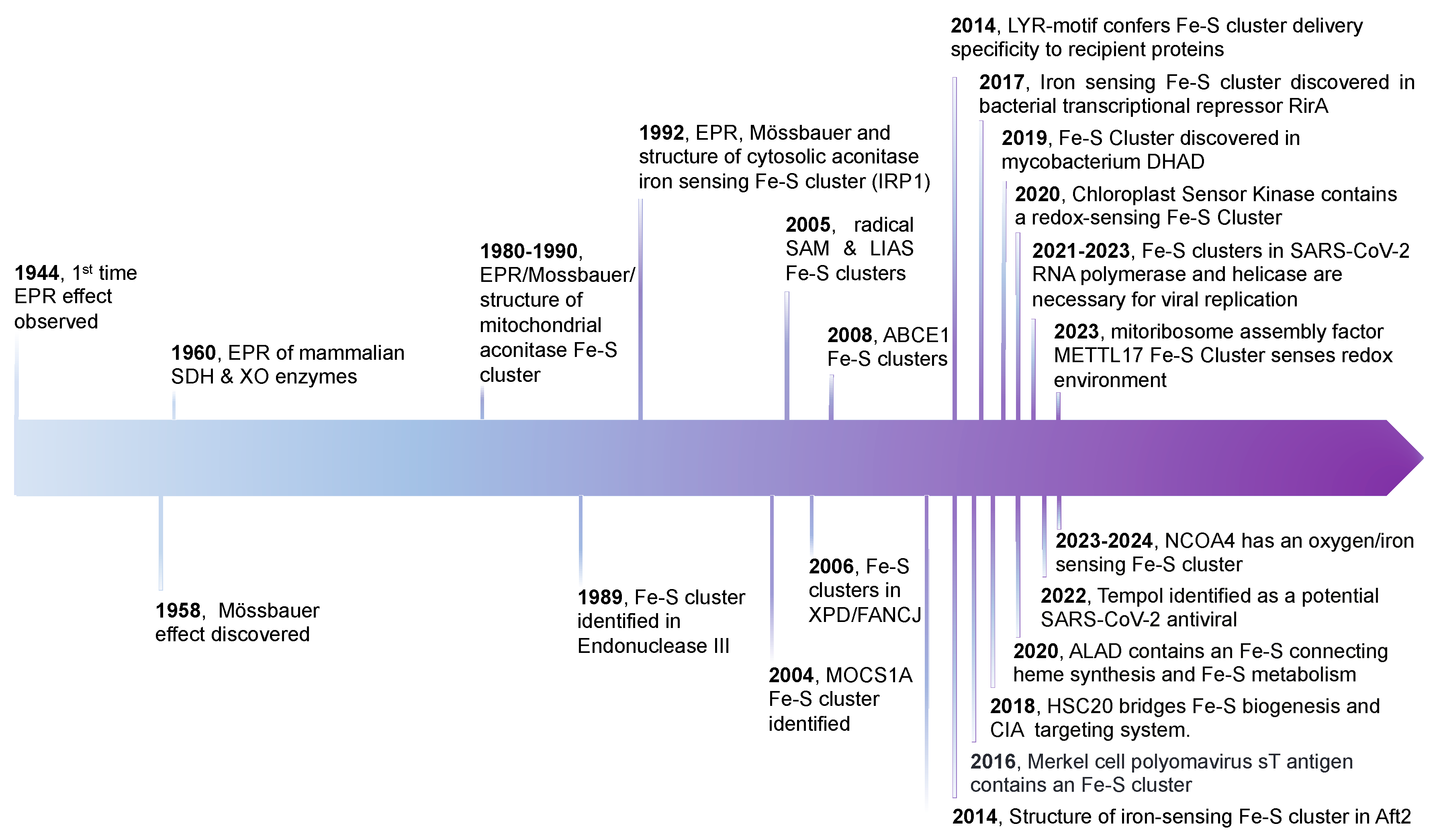
Figure 2.
Different iron sensing Fe-S cluster strategies across many forms of life. The labile nature of Fe-S clusters is taken advantage of by many organisms to sense and react to intracellular iron levels.
Figure 2.
Different iron sensing Fe-S cluster strategies across many forms of life. The labile nature of Fe-S clusters is taken advantage of by many organisms to sense and react to intracellular iron levels.
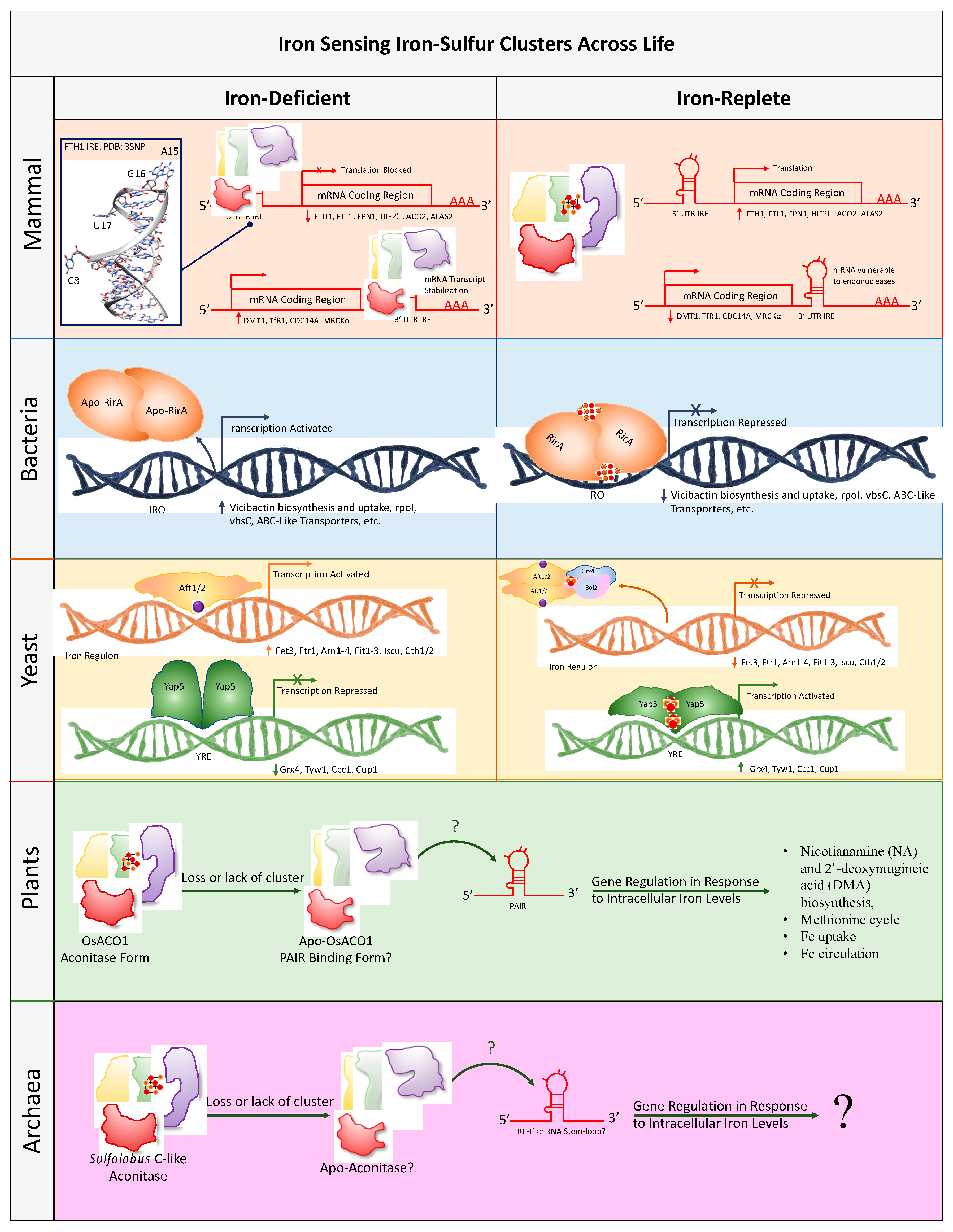
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



