
Submitted:
23 February 2024
Posted:
24 February 2024
You are already at the latest version
Abstract
Aquaculture is rapidly becoming one of the pivotal sectors in the farm economy, driven by the increasing demand for high-quality animal protein at an affordable cost, especially with the escalating human population. However, the expansion of high-density fish populations also brings forth a challenge – the rapid transmission and spread of infectious disease agents among them. To combat this, vaccination is emerging as a reliable and standardized method for providing immunity against viral and bacterial outbreaks. The ideal vaccine is expected to be safe, effective, economical, and easily administered. The fish vaccination industry continually publishes new information on fish immunology and vaccinology, contributing to the improvement of vaccine formulation and efficacy. This review aims to offer insights into the current status of bacterial, viral, and parasitic diseases, discuss existing vaccinations, and address potential industry-threatening diseases like infectious edwardsiellosis, motile aeromonas septicemia, TiLV, salmon anemia, vibriosis, and white spot disease. Technological advancements have played a crucial role in enhancing our understanding of fish immunological mechanisms, leading to improved vaccine administration and the development of recombinant live attenuated, subunit, DNA, and RNA vaccines. However, challenges such as oral tolerance, vaccine degradation, and stressful environments persist, impacting vaccine efficacy. Addressing these challenges and gaining a deeper understanding of the fish immune system and host-pathogen interactions will be pivotal for future improvements, contributing to the sustainability of aquaculture and enhancing global food security.
Keywords:
Infectious disease agents in aquaculture
; Fish Vaccine
; Fish immunology
; Bacterial
; viral
; and parasitic diseases in aquaculture
; Recombinant live attenuated
; subunit
; DNA
; and RNA vaccines for fish
; Sustainability of aquaculture
; The innate and adaptive immune system of fish
; Fish immunological mechanisms
1. Introduction
Fish farming is a billion-dollar industry and stands out as one of the fastest-growing sectors in animal food production. Beyond providing a stable source of income to millions of individuals, it plays a crucial role in ensuring food security and driving the economic development of several [1]. Environmental factors in the culture environment, including low water quality, high stocking density, and low oxygen levels, can induce stress in fish, elevating their vulnerability to infectious diseases. While effective management practices and prophylactic treatments significantly diminish disease susceptibility, the aquaculture industry still faces a substantial challenge, with more than 10% of all cultured fish being lost annually due to infectious diseases. This amounts to over 10 billion USD globally, underscoring the persistent impact of fish diseases on the aquaculture sector [1,2]. Disease outbreaks in fish cultures are attributed to various agents, with bacterial pathogens accounting for 54.9%, viruses for 22.6%, parasites for 19.4%, and mycotic agents for 3.1% of the reported cases [3,4]. This poses a significant issue, not only leading to substantial losses in aquaculture production but also giving rise to economic concerns in the developing world, which accommodates 90% of the aquaculture industry [5]. In Chile, for instance, infectious salmon anemia alone resulted in the loss of 20,000 jobs, costing the country 2 billion dollars[6]. China, the producer of 70% of the world’s farmed fish, experiences a 15% loss in total fish production due to diseases [7].
Hence, there is a significant emphasis on developing methods to prevent and control the spread of diseases. In the early stages, antibiotics served as a primary tool to combat infections due to their accessibility, affordability, and effectiveness in treating bacterial diseases. However, the repeated use of antibiotics was observed to suppress the fish's immune system and induce antibiotic resistance, thereby posing a threat to consumer health and safety[8,9]. As an alternative approach, vaccinations were introduced to decrease reliance on antibiotics.
The initiation of fish vaccinations dates back to the 1940s, and since that time, vaccines have played a pivotal role in mitigating the impact of bacterial and viral diseases on fish [10,11]. In the 1980s, only two commercial fish vaccines were available [1]. Presently, the market boasts over 50 vaccines catering to more than 30 different fish species. A majority of these vaccines have received approval from the United States Department of Agriculture (USDA) and are produced using methods that involve culturing target pathogens [12,13]. Administration primarily occurs through intraperitoneal injections. Ultimately, vaccination has proven highly successful, annually safeguarding thousands of fish from severe diseases [1].
When discussing the design and application of fish vaccination, it is important to have a good understanding of the fish immune system. Typically, the vaccination process involves trial-and-error with regard to pathogen identification, cultivation, and vaccine formulation [14]. Once a fish is vaccinated with the trial vaccine, the duration and intensity of protective immunity developed is examined to help provide a basis of whether the antigen is immunogenic and elicits the right ‘type’ of response, and so that researchers can identify restrictions within the vaccine that need to be addressed [15]. The ideal vaccine combines the most efficient antigens and adjuvant systems to produce an effective response against a specific pathogen with minimal side effects [14].
Recent studies in fish immunology have discovered that using cytokines as adjuvants instead of oil adjuvants has the advantage of stimulating the expression of co-stimulatory molecules and polarizing antigen-presenting cells [14]. Studies have also reported that using interferon responsive genes (IRGs) provides a form of innate immunity when encountering viral infections [16,17].
Advanced information in the field of fish immunology and microbiology are also directing new approaches toward fish vaccinology. Modern vaccine technology includes developments in the realm of recombinant DNA, which targets specific components within the pathogen using various expression systems. mRNA vaccines are also currently being studied and have been found to provide greater levels of immunity. In terms of vaccine administration, oral vaccines are being explored as an alternative strategy to commercially vaccinate fish, with the added advantage that it causes no stress to fish and is easier for fish farmers to administer [10].
Live attenuated vaccines are more efficacious, as they mimic natural infection conditions and have the ability to stimulate both humoral and mucosal immunity to generate a strong antibody response [18,19,20]. Some live attenuated vaccines were developed in 1990s and have been successfully implemented in aquaculture to increase production and reduce the use of antibiotics [21,22]. Recombinant attenuated Edwardsiella piscicida vaccines (RAEVs) have been explored as a vaccine delivery platform for heterologous antigens in aquaculture [23]. In carp, an E. tarda live attenuated vaccine induced greater levels of cellular immunity and conferred higher levels of protection compared to a formalin-killed vaccine against the wild-type strain.
While overall progress in fish vaccination is promising, certain limitations persist within the field. One notable challenge lies in the efficiency of administration, particularly given the individualized nature of vaccine delivery. Additionally, the aqueous environment poses a potential hindrance to the optimal efficacy of the vaccines. Various journals have extensively detailed the techniques employed in vaccine administration, current vaccination models, and the challenges confronting the fish vaccinology industry today. Bearing these considerations in mind, we will delve into the trends in fish vaccination, explore different vaccine types and administration methods, assess their advantages and limitations, and discuss the prospects of effectively controlling infectious fish diseases through vaccination
2. Fish Immune System
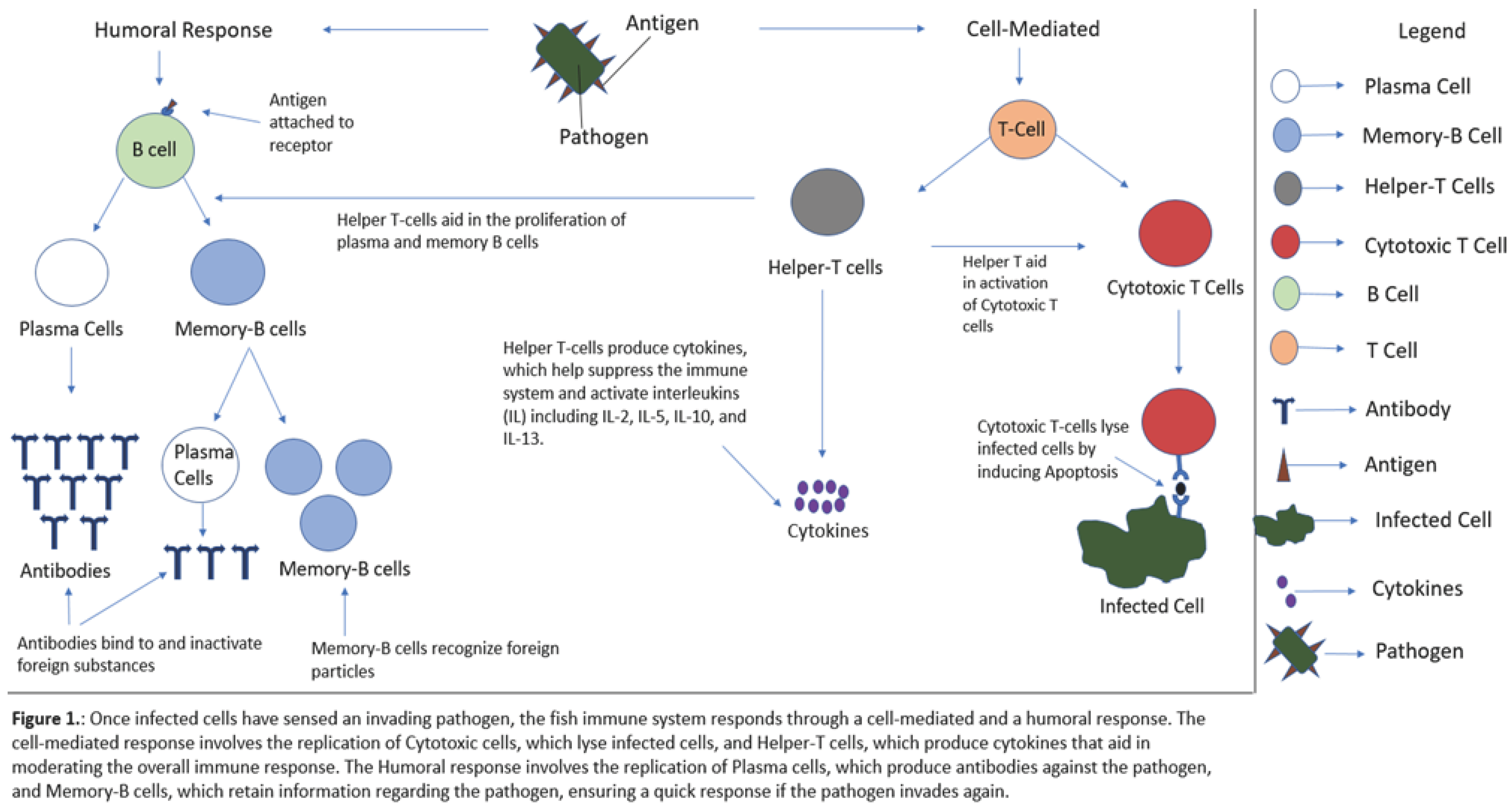
The fish immune system functions to identify and eliminate foreign material. It can be divided into two subsystems: the innate and adaptive immune system. While both subsystems aim to protect the body, adaptive immunity can be characterized as a system in which the organism, through genetic mutations and recombination, generates immunological memory against the pathogen. Namely, it provides immunity by identifying and eliciting the appropriate antigen response through the production of memory cells and specific receptors, including immunoglobins, B lymphocytes, and T-cells [24,25].
On the other hand, innate immunity is non-specific, wherein preexistent mechanisms, such as the epithelium and mucosa, form a barrier or chemically respond to anything that is foreign or non-self [26]. The innate immune system can be divided into physical, cellular and humoral components [27]. The physical barriers consist of mucosal routes, particularly in the gills, skin, gut, and epidermis [28]. The humoral components include cytokines, protease inhibitors, agglutinins, and antibacterial peptides. The cellular components consist of epidermal, neural, and phagocytic cells [27].
It is important to note that certain chemical components are a part of this system and play a vital role in the immune response. In particular, lysozymes, a component of the mucosa and the intestinal tract system, induce cell lysis to prevent infected viral cells from replicating. Cytokines, such as IL-1β and TNF-α, can trigger inflammation in response to gram-negative bacteria [18,29].
Once the antigen enters the body, the immune system launches a humoral and cell-mediated response. B cells identify and bind the antigen to specific receptors. Exposure to the antigen and the activation of helper T cells facilitates proliferation and differentiation of B cells into memory B and plasma cells during primary activation [30,31]. Plasma cells construct high-affinity antibodies against the antigen. Memory B cells contain antigen-recognizing receptors that can continue to identify the pathogen even after the virus has been controlled [32].
A cell-mediated response will also be activated. This involves the activation of T cells, which can act as both helper T cells or cytotoxic T cells. Cytotoxic T cells can lyse virus-infected cells and protect the organism from re-infection. T-helper cells, defined as CD4+, CD25+ T cells, act as a regulatory mechanism, stimulating Cytotoxic T cell production, influencing cytotoxic T cells, and suppressing certain immune responses deleterious to the host if needed [33]. The cell-mediated response is an imperative component of the viral response. It can protect against intracellular bacterial infection by pathogens like E. tarda with CD8α+, CD4+ T cells and sIgM+ cells, which possess antibacterial properties [34].
Cell-mediated responses work to terminate infected cells while humoral responses provide a form of resistance against the virus through antibody production. Thus, an effective vaccine entails the successful identification of virulence factors, the goal being to stimulate the immune system such that the fish develops protective immunity against the pathogen [7]. In most fish vaccinations, the organism is exposed to either a live attenuated pathogen, a non-replicating pathogen, an inactive pathogen or its subunits. This elicits a response from the immune system and aids in the development of antibodies against the pathogen [8,9].
Vaccination methods can stimulate activity of certain components within the immune system. For instance, oral vaccinations act in the gut and in mucosal sites, they trigger a systemic and humoral response while also increasing genetic expression of cytokines and IgM production. Bath vaccinations induce an innate and adaptive response, resulting in the upregulation of toll-like receptors and proinflammatory genes [18,35,36]. Mucosal vaccinations stimulate a form of frontline humoral immunity, wherein local immunoglobin proteins are produced in the mucosa-associated lymphoid tissue (MALT) and act against the pathogens in the mucosal tract [37]. Overall, a clear understanding of the immune system and its influence on vaccine administration can provide insight on vaccination trends, but more importantly, provide context on how the method of administration influences the effects of vaccination.
3. Bacterial, Viral, and Parasitic Diseases in Fish
Fish are susceptible to a variety of diseases, both infectious and non-infectious. Infectious diseases are caused by a pathogenic organism, either encountered in the environment or spread through contact. Non-infectious diseases are caused by a combination of environmental and genetic anomalies and are not contagious. Infectious diseases are transmitted to fish either directly or indirectly and are expediated by external conditions such as poor water quality and contaminated culture. There are three types of infections that fish commonly suffer from: parasitic, bacterial and viral. The most common bacterial, viral and parasitic diseases and their current vaccination status are listed in Table 1.
Fish are commonly infected by pathogenic bacterial species belonging to the genera Edwardsiella, Vibrio, Aeromonas, Streptococcus, and Flavobacterium [38]. Bacterial pathogens are ubiquitous in the aquatic environment [39]. Induced stress and causative factors (e.g., organic pollution) can prompt a bacterial infection outbreak [40]. Symptoms of bacterial infection are often exhibited through exophthalmia, epithelial lesions and anorexia, all of which can be treated with antibiotics and vaccination. Intraperitoneal or bath vaccination are favored to treat bacterial infections, but in severe cases, antibiotics are administered [1]. There are existing vaccinations for several, but not all, bacterial infections.
Virus-host interactions can cause alterations at the cellular level and the viral infection can spread rapidly through manipulation of host DNA. Well-characterized DNA viruses include iridoviruses, herpesviruses, and adenoviruses [41]. However, some organisms are able to replicate inside the host organism through the insertion of RNA contents. RNA viruses are represented in the aquaculture world in the form of Retroviridae, Rhabdoviridae, Paramyxoviridae, and Orthomyxoviridae [41].
Viral symptoms are specific to the infection. For example, paramyxovirus evokes inflammatory gill disease, whereas infectious salmon anemia virus (ISAV) causes liver necrosis and anemia [42]. Given that viral symptoms include bacterial infections, it can be difficult to distinguish between viral and bacterial diseases. Thus, laboratory tests are the primary method of viral identification. Today, there are vaccines for both DNA and RNA viruses. DNA viruses are treated through the administration of pDNA via intramuscular injection, which yields ‘strong expression of transgenes at the injection site’. RNA vaccines utilize reverse genetics systems wherein RNA viral genomes are targeted and genetically manipulated by cDNA [43].
Parasitic organisms can enter through the gills and skin or be ingested with its intermediate host. They are grouped into two categories: ectoparasites and endoparasites. Ectoparasites live on the external surface of hosts, while endoparasites live within the host [44]. The spread of parasitic infections is facilitated via anthropogenic factors; low pH, low oxygen concentration in reservoirs and lakes, and thermal pollution can cause infections such as ichthyobodosis and myxosporeoses [40]. Parasitic infections are often diagnosed using a combination of observational and laboratory techniques. Salt-grain blisters and emerging white spots are often considered to signs of white spot disease, however, the presence of Ichthyophthirius mulifiliis (‘Ich’) can only be confirmed by analyzing the organism’s epithelial tissue under a compound microscope [45]. Apart from increased mucus secretion and gill discoloration, parasites evoke cell proliferation and immunomodulation, which are detrimental to the fish host [46]. There is a high emphasis on preventative measures, such as biochemical and bath techniques, to prevent and decrease the severity of the disease in culture environments. However, there are no vaccinations for parasitic infections currently on the market.
4. Current Licensed Vaccines for Bacterial Diseases
Bacterial diseases occur as a result of interactions between bacteria, host, and the surrounding environment [47]. Over the past few years, these interactions have been studied and have led to the production of various vaccines. Table 2 covers the list of licensed commercial vaccines available for bacterial diseases in fish.
4.1. Edwardsiellosis in Fish
Edwardsiellosis is one of the most important bacterial diseases in fish. It is caused by Edwardsiella piscicida, a gram-negative, facultative anaerobic bacterium of the family Enterobacteriaceae. It affects many economically important fish species, including catfish (Ictalurus furtatus), tilapia (Tilapia nilotica), European eel (Anguilla anguilla) and Indian major carp (Catla catla) [48,49,50]. Having endured the overuse of antibiotics, the pathogen boasts several antibiotic resistance genes, making it a complex and difficult pathogen to protect against. Vaccination would be an effective method to prevent and control Edwardsiella outbreaks. There are no commercial vaccines available to prevent disease.
Current research is focusing on developing live attenuated and subunit vaccines, with live attenuated vaccines targeting the PhoP-PhoQ complex, known to sense and respond to homeostatic changes, as well as Type III and Type IV secretion systems, often considered to be influential virulent determinants [51,52,53]. Several subunit vaccines impact cytokines, specifically interleukins, to provide immunity to the host (Tang et al 2020) (Guo et al 2018). A recombinant attenuated Edwardsiella piscicida vaccine (RAEV) vector system with a regulated-delayed attenuation phenotype has been used to deliver heterologous protective antigen. This novel vaccine system induces both systemic and mucosal IgM titer against E. piscicida and heterologous antigen in zebrafish [54].
4.2. Enteric Septicemia of Catfish
Enteric septicemia of catfish (ESC), also known as hole-in-the-head disease, is caused by Edwardsiella ictaluri. Given that E. ictaluri is intracellular pathogen, ESC cannot be treated using killed vaccines [47]. Instead, a vaccine that targets T cells by antigen presenting cells (APC) can induce innate immunity [55]. The only commercially available vaccine for ESC has not been widely accepted due to lack of efficacy as well as nominal economic returns [56]. Another live attenuated vaccine developed by MAFES and MSU, which contains increasing concentrations of rifamycin, is under consideration and is undergoing commercial-scale trials [57]. To be able to produce more vaccines and better understand how the pathogen operates, researchers are investigating the the immune response once the vaccine is administered [55,58,59].
4.3. Bacterial Kidney Disease:
Bacterial Kidney Disease (BKD) is a chronic infection which impacts salmonids at low temperatures. The causative agent, Renibacterium salmoninarum, contains the p57 protein, which agglutinates and suppresses the host’s defense mechanism [60]. The predominant effect of the disease is granulomatous inflammation, which spreads throughout the body and causes lesions in the kidney [61]. There is only one vaccine commercially available in the market, sold under the trade name Renogen. It is a live vaccine prepared through lyophilization and contains a live version of the bacteria [61]. In one trial, it was found that a combination of Renogen and MT-239 resulted in increased survival [61]. However, immune response varies and a single vaccine is not completely reliable. There are several recently published papers that focus on the characteristics of the bacterium, but there are few papers published on vaccination trials and formulation in this decade [62,63,64].
4.4. Flavobacteriosis/Columnaris
Flavobacteriosis is caused by bacteria in the Flavobacteriaceae family, and impacts both farmed and wild fish. The most well-characterized bacteria within this family are F. psychrophilum and F. columnare, both of which are responsible for devastating losses. Current vaccinations on the market include Aquavac-Col, Fryvacc 1 and Fryvacc 2. When conducting trials, Aquavac-Col was developed using genomovar I and was found to be effective in protecting catfish from columnaris, though it was found to be ineffective against genomovar II [65]. Rifampicin-resistant mutants from F. columnare genomovar II strains were found to be stable and effective against the more virulent genomovar II strain [66]. Fryvacc1 is specifically administered to salmonids and FryVacc 2 is the first bivalent immersion vaccine created [67]. Fryvacc II works against Columnaris and Yersiniosis [68]. Though both vaccines are legally licensed, there are few articles assessing their efficacy. Extensive research has provided insight on the genomic sequences and characterization of the variants of the disease, and has led to the development of mucosal vaccines which incorporate a rifampicin-resistant mutant strain [69,70]. Moreover, it has led to the formulation of the B.17-ILM vaccine, a live attenuated vaccine that protects against F. psychrophilum in salmonids and rainbow trout [71,72]. The vaccine is composed of a rifampicin resistant strain of F. psychrophilum, has considerable research supporting its protection potency, and is currently undergoing trials so that it can become licensed and commercialized in the US [72,73,74].
4.5. Furunculosis
Furunculosis is caused by the gram-negative, bacillus bacterium Aeromonas salmonicida. Symptoms of the disease include dermal ulcerations, skin darkening and bacterial infiltration in several organs, and are elicited by the type 3 secretion system, which serves as a conduit for the passage of effector proteins into host cells [75,76,77]. This is best described as an ‘immune invasion,’ as it inhibits phagocytosis and intracellular killing, making it a high-mortality disease [77,78]. Currently, there are several vaccines are available on the market to treat furunculosis. Many of the vaccines, including those manufactured by Alpha Ject Norvax Minova 6, Forte VI and Lipogen Forte, are polyvalent vaccines. It is difficult to distinguish whether this provides more protection against the bacterin or not, some studies indicate that this form of vaccine provides better immunity than monovalent vaccines and that it enhances the humoral immune response, and others found that both provide proper protection [79,80]. Various studies have concluded that mineral oil-adjuvant vaccines have provided the most protection when challenged by this disease, as it is known to induce a non-specific immunity [81,82,83]. However, this form of vaccination induces adverse reactions, including internal lesions and reduction in weight [81]. There is an emphasis on identifying what can be done to reduce the severity of these reactions, and on developing safer, yet effective vaccines [84,85,86]. Live attenuated vaccines containing A-layer and O-deficient strains of Aeromonas salmonicida have been shown to confer significant protection, and there are researchers looking into recombinant vaccines for other species impacted by the disease, including rainbow trout [67,87,88,89].
5. Current Licensed Vaccines for Viral Diseases
In comparison to bacterial infections, viral disease outbreaks are more difficult to control due to lack of anti-viral therapeutics and information regarding the mechanism of viruses, specifically, how they impact the organism. Several research trials have been conducted on the latter and have provided crucial information that has allowed companies and academic organizations alike to develop effective vaccines. Table 3 displays a list of available licensed viral vaccines for fish.
Koi Herpes Virus (KHV)
The koi herpesvirus causes significant mortality in common carp types. It is a double-stranded DNA virus that belongs to the family Alloherpresviridae. The virus can replicate within a host organism and release infectious agents without showing physical or clinical signs, resulting in its association with viral outbreaks in the aquatic world.
Not much is understood about its pathogenesis and how the virus is transmitted, it can go undetected for weeks. KV-3, produced by KoVax, is the only commercially available vaccine for KHV. Autogenous, inactivated vaccines have been developed and administered to protect carp in countries such as Germany and Indonesia [90]. DNA vaccines are being tested and developed, and Arthrospira platensis is being explored as an alternative to prevent an outbreak of KHV and reduce its transmission rate [90,91,92,93].
Infectious hematopoietic necrosis virus (IHNV)
Infectious hematopoietic necrosis virus is a single-stranded RNA virus belonging to the family Rhabdoviridae and the genus Novirhabdovirus. The virus can be isolated into 3 genogroups (U, M, and L), with the M genogroup containing a higher genetic diversity and a higher mortality (CFSPH) [94]. There are several patented vaccines against IHNV, including live-attenuated vaccines, inactivated and reverse genetic vaccines [95,96,97]. However, safety and environmental concerns have prevented commercial availability. APEX-IHN was commercially marketed due to its protective efficacy and reduction in transmission [98]. Currently, DNA vaccine trials are being conducted, the majority of which are developed based on the G protein unique to the IHNV U and M genotypes. Multivalent vaccines are being researched to combat both IHNV and VHSV [99].
Red Sea Bream Iridovirus (RSIV)
Red Sea Bream Iridovirus (RSIV) is a single-stranded DNA virus belonging to the Iridoviridae family and the Megalocytivirus genus. It utilizes a horizontal mode of transmission and has been found to infect juveniles more than adults. There have been some trials attempting to develop a genetic vaccine and a recombinant yeast cell oral vaccine [100]. However, more trials must be conducted in order to certify and license them as commercially available vaccines. A formalin-based vaccine was developed by Nakajima et al 1999 [101]. While this vaccine is currently available for fish species of the Seriola genus, it is not effective on rock bream [102]. Currently, there are trials being conducted to determine the efficacy of DNA vaccines and vaccines effective on rock bream [102,103]. When conducting cross-examination against other Megalocytiviruses, AQUAVAC® IridoV showed partial protection against infectious spleen and kidney necrosis virus (ISKNV) and no cross-protection against scale drop disease virus (SDDV) [104]. As a result, a bivalent or polyvalent vaccine is being considered. One example is the PISCIVAC™ Irido Si vaccine, which specifically protects red sea bream against both RSIV and Streptococcus iniae, and is licensed in Japan [105].
Salmonid Alphavirus (SAV)
The salmonid alphavirus itself is a positive-sense RNA virus belonging to the family Toogaviridae and the genus Alphavirus. The diseases caused by the virus are pancreatic disease (PD) and sleeping disease (SD), and while both are classified as separate diseases, a histopathological study conducted found that both diseases are caused by a similar or identical agent. Additionally, both have similarities at the nucleotide and amino acid level [106]. There are 6 different strains or subtypes of SAV. That being said, it was found that a vaccine containing a single subtype or strain can protect against PD caused by different strains of SAV, regardless of their subtype grouping [107].
Currently, only inactivated intraperitoneal vaccines are on the market. Though they are licensed for commercial use, it was found that Norvax Compact PD reduced the mortality rate by 50% [108]. Few studies have been conducted to evaluate the efficacy of all 4 vaccines. In a recent study, researchers compared the efficacy of 2 DNA vaccines, one based on a plasmid expressing the whole SAV structural polyprotein C-E3-E2-6K-E1 (pCSP) and one based on a plasmid encoding the SAV3 surface protein E2 alone (pE2), and Norvax Compact PD containing the inactivated SAV subtype 1 [109]. They found that pCSP and the Norvax vaccine reduced mortality rates, but when comparing all three vaccines, it was found that pCSP provided far more protection against the virus than Norvax and the p53 plasmid [110]. Thus, there is more research being done on developing a DNA vaccine against the virus. Recently, a DNA vaccine under the name of Clynav with SAV subtype 3 was approved by the European Medicine Agency and is currently undergoing commercial trials in salmon farms [111].
Infectious Pancreatic Necrosis Virus (IPNV)
Infectious pancreatic necrosis virus is a double stranded RNA (dsRNA) virus belonging to the Birnavirus family. The virus affects salmonids, namely the Rainbow trout and Atlantic salmon populations in North America and Europe. IPNV has a double-stranded genome, consisting of fragments A and B. Out of the 5 proteins that make up the virus, Fragment A is responsible for encoding VP2 and VP3, 2 major structural proteins of the virus. Specifically, VP2 proteins contain the most antigenic determinants [112]. Therefore, many vaccines against the IPNV contain the immunogenic protein to neutralize antibodies and stimulate an immune response.
There are many commercially available vaccines currently on the market to combat IPNV (Table 2). Some of the first viral vaccines developed to combat IPNV and many other viruses were inactivated vaccines, and they continue to be a reliable method, which may be why there are many inactivated vaccines on the market for IPNV. Inactivated viral vaccines also induce strong responses, as they retain the inactivated genomic component [113]. Along with inactivated vaccines, the Microtek Trivalent vaccine has also been highly successful and provides the user the convenience of providing protection against 3 pathogens [113]. There are DNA vaccines are being developed and tested currently, including a DNA vaccine wherein the VP2 gene was incorporated into a DNA vector and added into alginate microspheres [114]. This alginate-bound DNA vaccine was distributed into food pellets and was found to provide high protection in juvenile Rainbow Trout [114]. Another DNA vaccine encodes the VP2 gene with P217, T221, A247 (PTA) motif, which was found to elicit a strong immune response [115].
Infectious Salmon Anemia (ISA)
Infectious salmon anemia is caused by a single-stranded RNA virus (ssRNA) that belongs to the Orthomyxoviridae family. It is one of the deadliest viruses that affect Atlantic salmon. The virus’s biochemical and chemical structures are similar to that of the influenza virus [116]. After examining more than 160 ISAV isolates, it was found that there are 2 hemagglutinin subtypes of the vaccine, one predominant in America, and the other dominant in Europe [117]. Therefore, many vaccines contain an inactivated whole virus as an antigen [116].
As seen in Table 3, there are many inactivated polyvalent vaccines and subunit vaccines available. While subunit vaccines can be difficult to formulate, given that the proteins can quickly degenerate, they yield effective results. A highly successful example is the Blueguard ISA Oral vaccine [3]. Recently, an adjuvant vaccine, wherein an SAV-based replicon, was found to provide immunity against the ISA when administered via intramuscular injection [10,118]. There are also studies investigating interferon-based vaccines [119].
Tilapia Lake Virus
Tilapia lake virus (TiLV) is a pathogen that has recently emerged and is responsible for significant mortality and economic losses in global tilapia aquaculture. Tilapia Lake Virus (TiLV) is a negative-sense single-stranded RNA virus belonging to the family Amnoonvirdae and the genus Tilapinevirus [120,121]. First documented in 2013, TiLV made headlines as it spread across continents in record time with a mortality rate above 80% [122]. The virus consists of 10 segments that encode 10 proteins. Segment 1 exhibits slight similarity to the polymerase proteins of influenza and is believed to encode for PB1, a protein that initiates RNA synthesis and plays a vital role in integrating viral genetic material into the host cell [122,123]. Currently, research has focused on understanding the etiology of the virus and its method of transmission. There are currently no vaccines commercially available on the market. Several investigations on the effect of the VP20 protein on segment 8, which serves as a vaccine antigen, have made creating DNA-based vaccines against the virus more favorable [124,125].
5.1. Current Licensed Vaccines for Parasitic Diseases
Parasitic diseases can have devastating effects on the aquaculture industry, and these effects are not limited to monetary and economic loss. If not handled properly and eliminated, the consumption of raw diseased fish can cause infections in other organisms, and even humans.
Although parasitic infections are known to be as impactful as viruses and bacteria, there is only one commercially licensed vaccine to treat a parasitic infection (a vaccine against Sea lice manufactured by Aquatec) [126,127]. This is because very little is understood about the immune response to parasites. Parasitic diseases invade host cells and feed off of the host to obtain nutrients for growth, reproduction and transmission. Additionally, parasites have mechanisms in place to prevent and cope with the piscine immune response. These include parasitic migration, wherein the parasite migrates to host sites where the immune response has not reached or is weak, anti-immune mechanisms, which allow them to resist innate humoral factors, and immunodepression, where parasites induce apoptosis of the host’s leukocytes [128,129]. However, modern science has made advances in understanding parasitic infections. Neutrophils have been found to play a crucial role in defending against parasites, MC degranulation has been observed in several parasitic infections and eosinophilic granule cells have been found to release various tryptases, lysosomes and antimicrobial peptides at the site of parasitic invasion [129,130,131]. Current literature review focuses on investigating ectoparasites and their proteomic expression to determine vaccination mechanisms and methods. For instance, in Tetracapsuloides bryosalmonae, known to cause polycystic kidney disease, the activation of a micro-exon gene Tb-MEG1 has been found to avert immune subversion [132]. An anti-Tb-MEG1-based DNA vaccine has been shown to activate IgM responses against the infection [126]. Additionally, the injectable, commercially available vaccine produced by Aquatec utilizes a synthetic subunit antigen along with the keyhole limpet hemocyanin (KLH) to elicit protection against sea lice [133,134]. Studies investigating I. multifiliis have found success expression of a DNA vaccine using Iag52B in rainbow trout and channel catfish, the recurrent theme in vaccine trials against this parasite seems to be the use of 48kDa immobilization antigens [135,136,137].
Overall, parasitic vaccines rely on DNA and subunit-based vaccines through intraperitoneal injection [126]. It is important to note the discrepancy in the number of vaccine trials done on ectoparasitic diseases in comparison to endoparasitic diseases. Little is known about the genetic composition and protein expression of myxozoans, which play a role in pathogenicity of the fish host. Analysis of their microbiology and DNA composition will facilitate the isolation of proteins directly involved in pathogenesis and aid in developing vaccines.
5.2. Challenges and Limitations in Developing Vaccines for Fish
The ideal vaccine provides long-term immunity, is easily distributed and is economical. Oral vaccination is the preferred method of administration due to its versatility and ease of use, it doesn’t cause stress to fish and can be distributed in large numbers at once. However, as much as oral vaccination is considered a future prospect, it comes with its limitations. Due to its route of entry, this form of vaccination can cause oral tolerance. Defined as the suppression of humoral and cellular immune responses, oral tolerance is one of the main issues that hinder the development of effective oral vaccines [28]. Though there is some information regarding the mechanism responsible for this phenomenon in mammals and humans, there is little information on how it works in fish [138,139].
Similar to oral, mucosal vaccines are widely distributed, easy to administer, and can provide optimal administration efficacy as fish have large mucosal surfaces [1]. The issue in this case involves efficacy. There are no immunostimulants in non-replicating mucosal vaccines that can impact the vaccine’s performance and there is also the risk of prolonged exposure to vaccination [140]. Because most mucosal vaccinations utilize bath administration, it can be difficult to calculate how much of the antigen was administered in relation to the functional testing of mucosal T cells, which can prevent proper evaluation of vaccination efficacy [141].
Bath immersion, also known as immersion vaccination, involves the administration of vaccine antigens via immersive techniques and is mostly used to provide booster vaccinations [71]. Immersion vaccines facilitate mass vaccination and can provide vaccination to smaller fish that are not able to endure intraperitoneal vaccination. Though new research has found that live attenuated vaccines, recombinant vaccines and chitosan-based vaccines increase immersion vaccine efficacy, it is difficult to produce a bath administration vaccine in the lab due to the denaturation of antigens when crossing digestive and mucosal barriers, variability in results, and difficulty in determining improvement measures from experimental trials [1,142,143,144].
Although there such vaccine models have the potential to be successful, there needs to be more research conducted on immunological mechanisms to understand how to formulate a vaccine that will be able to successfully produce the desired result without severe side-effects, such as chronic stress. Researchers should focus on producing vaccines that induce long-term immunity. This way, fish do not have to repeatedly undergo handling and anesthetic procedures, which are necessary for injection-based vaccines administration [145]. This would also reduce susceptibility to oral tolerance. When discussing administration methods, bath and immersion vaccines are also noted. Bath vaccination is noted as inducing long-term immunity.
Although such limitations are directly related to vaccine formulation and administration, certain factors such as cost efficiency are indirectly related. As much as cost-efficiency is one of the strengths of vaccination, a lot of money is needed to produce a strong vaccine. From antigen isolation, to producing the most favorable adjuvant, to evaluating the mortality of the vaccine in a commercial setting, there are several steps required for the production of a strong vaccine, all of which can be costly.
There should also be more information on the efficacy of commercial vaccines. Although there are certain regulations and standards that a certified vaccination must meet to become commercially available, there are not many scientific papers comparing the efficacy of different vaccines that claim to protect against the same pathogen. This would aid farmers in determining which vaccine is best for them to use. Though vaccines have the ability to prevent viral outbreaks, they cannot be used on fish weighing less than 20g [145]. Many of these fish are in the larval or fry stage. This is a challenge, as fish in these stages are the most susceptible to major disease outbreaks [146].
6. Summary and Conclusions
Pharmaq, a leading aquaculture vaccine-producing company, has reported the distribution of over 300 million doses of salmon-helping vaccines to Norway. In Chile, the world’s second-largest producer of farmed fish, the majority, if not all, of farmed salmonids are vaccinated [147]. Globally, millions of fish vaccines are distributed with the aim of reducing mortality rates in aquaculture farms and preventing disease outbreaks. The vaccination process is complex, requiring a deep understanding of the host’s genomic components, cellular defenses, and their interactions. While several licensed vaccines are available, there is currently no commercially available vaccine to protect against certain parasitic, bacterial, and viral diseases, such as white spot disease, Edwardsiellosis, and Tilapia Lake Virus (TiLV). Nevertheless, publications have provided insights into potential prospects and outlined preventive measures, such as adhering to strict aquaculture guidelines to reduce stress and maintain sanitation. It is crucial to acknowledge advancements in fish immunology and vaccinology, which have introduced various administration methods to vaccinate against diseases like Vibriosis and IPNV. These advancements have also facilitated the formulation of cost-effective polyvalent vaccines, allowing the simultaneous vaccination against multiple diseases with just one injection.
Acknowledgments
This work was supported by the U.S. Department of Agriculture (USDA) - National Institute of Food and Agriculture – USDA-NIFA Grant No. 2022-70007-38287.
References
- A. Adams, Progress, challenges and opportunities in fish vaccine development, Fish & shellfish immunology 90 (2019) 210-214. [CrossRef]
- D.W. Wanja, P.G. Mbuthia, R.M. Waruiru, J.M. Mwadime, L.C. Bebora, P.N. Nyaga, H.A. Ngowi, Fish Husbandry Practices and Water Quality in Central Kenya: Potential Risk Factors for Fish Mortality and Infectious Diseases, Veterinary medicine international 2020 (2020) 6839354. [CrossRef]
- A.K. Dhar, S.K. Manna, F.C. Thomas Allnutt, Viral vaccines for farmed finfish, Virusdisease 25(1) (2014) 1-17. [CrossRef]
- W. Surachetpong, S.R.K. Roy, P. Nicholson, Tilapia lake virus: The story so far, Journal of Fish Diseases 43(10) (2020) 1115-1132. [CrossRef]
- F.A.O. Fisheries, Aquaculture Department, The state of world fisheries and aquaculture (2012) 1-153.
- A. Assefa, F. Abunna, Maintenance of Fish Health in Aquaculture: Review of Epidemiological Approaches for Prevention and Control of Infectious Disease of Fish, Veterinary Medicine International 2018 (2018) 5432497. [CrossRef]
- E.I. Broughton, D.G. Walker, Policies and practices for aquaculture food safety in China, Food Policy 35(5) (2010) 471-478. [CrossRef]
- G.T. Rijkers, A.G. Teunissen, R. Van Oosterom, W.B. Van Muiswinkel, The immune system of cyprinid fish. The immunosuppressive effect of the antibiotic oxytetracycline in carp (Cyprinus carpio L.), Aquaculture 19(2) (1980) 177-189. [CrossRef]
- M.A. Salam, M.Y. Al-Amin, M.T. Salam, J.S. Pawar, N. Akhter, A.A. Rabaan, M.A.A. Alqumber, Antimicrobial Resistance: A Growing Serious Threat for Global Public Health, Healthcare (Basel, Switzerland) 11(13) (2023). [CrossRef]
- J. Ma, T.J. Bruce, E.M. Jones, K.D. Cain, A Review of Fish Vaccine Development Strategies: Conventional Methods and Modern Biotechnological Approaches, Microorganisms 7(11) (2019). [CrossRef]
- M. Horzinek, V. Schijns, M. Denis, P. Desmettre, L. Babiuk, General description of vaccines, Veterinary Vaccinology. Amsterdam: Elsevier (1997) 131-52.
- V.f. Aquaculture, USDA, AMS, LPS Agricultural Analytics Division for the USDA National Organic Program, USDA (2014).
- Y. Du, X. Hu, L. Miao, J. Chen, Current status and development prospects of aquatic vaccines, Frontiers in immunology 13 (2022). [CrossRef]
- M. Guo, C. Li, An overview of cytokine used as adjuvants in fish: current state and future trends, Reviews in Aquaculture 13(2) (2021) 996-1014. [CrossRef]
- C. Secombes, Will advances in fish immunology change vaccination strategies?, Fish & shellfish immunology 25(4) (2008) 409-16. [CrossRef]
- C. Langevin, E. Aleksejeva, G. Passoni, N. Palha, J.-P. Levraud, P. Boudinot, The Antiviral Innate Immune Response in Fish: Evolution and Conservation of the IFN System, Journal of Molecular Biology 425(24) (2013) 4904-4920. [CrossRef]
- E.R. Verrier, C. Langevin, C. Tohry, A. Houel, V. Ducrocq, A. Benmansour, E. Quillet, P. Boudinot, Genetic resistance to rhabdovirus infection in teleost fish is paralleled to the derived cell resistance status, PLoS One 7(4) (2012) e33935-e33935. [CrossRef]
- B. Swain, C.T. Powell, R. Curtiss, Virulence, immunogenicity and live vaccine potential of aroA and phoP mutants of Edwardsiella piscicida in zebrafish, Microbial Pathogenesis 162 (2022) 105355. [CrossRef]
- B. Swain, C.T. Powell, R. Curtiss, Pathogenicity and immunogenicity of Edwardsiella piscicida ferric uptake regulator (fur) mutations in zebrafish, Fish & Shellfish Immunology 107 (2020) 497-510. [CrossRef]
- B. Swain, V.A. Campodonico, R. Curtiss, Recombinant Attenuated Edwardsiella piscicida Vaccine Displaying Regulated Lysis to Confer Biological Containment and Protect Catfish against Edwardsiellosis, Vaccines 11(9) (2023) 1470.
- C.A. Shoemaker, P.H. Klesius, J.J. Evans, C.R. Arias, Use of Modified Live Vaccines in Aquaculture, Journal of the World Aquaculture Society 40(5) (2009) 573-585. [CrossRef]
- R. Gudding, W.B. Van Muiswinkel, A history of fish vaccination: Science-based disease prevention in aquaculture, Fish & Shellfish Immunology 35(6) (2013) 1683-1688. [CrossRef]
- B. Swain, C.T. Powell, R. Curtiss, Construction and Evaluation of Recombinant Attenuated Edwardsiella piscicida Vaccine (RAEV) Vector System Encoding Ichthyophthirius multifiliis (Ich) Antigen IAG52B, Frontiers in Immunology 12 (2022). [CrossRef]
- P.R. Rauta, B. Nayak, S. Das, Immune system and immune responses in fish and their role in comparative immunity study: a model for higher organisms, Immunology letters 148(1) (2012) 23-33. [CrossRef]
- N.C. Smith, M.L. Rise, S.L. Christian, A Comparison of the Innate and Adaptive Immune Systems in Cartilaginous Fish, Ray-Finned Fish, and Lobe-Finned Fish, Frontiers in immunology 10 (2019) 2292. [CrossRef]
- G.Á. Aristizábal B, Innate immune system." In Autoimmunity: From Bench to Bedside, El Rosario University Press Chapter 2 (2013).
- B. Magnadóttir, Innate immunity of fish (overview), Fish & shellfish immunology 20(2) (2006) 137-51. [CrossRef]
- S. Mutoloki, H.M. Munang'andu, Ø. Evensen, Oral Vaccination of Fish - Antigen Preparations, Uptake, and Immune Induction, Front Immunol 6 (2015) 519-519. [CrossRef]
- B. Swain, M. Samanta, M. Basu, P. Panda, B.R. Sahoo, N.K. Maiti, B.K. Mishra, A.E. Eknath, Molecular characterization, inductive expression and mechanism of interleukin-10 gene induction in the Indian major carp, catla (Catla catla), Aquaculture Research 43(6) (2012) 897-907. [CrossRef]
- J. Playfair, Living with Germs: In sickness and in health, 2004.
- C.A. Janeway, Paul Travers, Mark Walport, and Donald J. Capra., Immunobiology, UK: Garland Science: Taylor & Francis Group 5th edition (2001).
- J. Gregory, Taylor, Dennis., Fosbery, Richard., Jones, Mary, Cambridge International AS and A Level Biology Coursebook with CD-ROM. United Kingdom, Cambridge University Press (2014).
- T. Nakanishi, Y. Shibasaki, Y. Matsuura, T Cells in Fish, Biology (Basel) 4(4) (2015) 640-663. [CrossRef]
- M. Yamasaki, K. Araki, T. Nakanishi, C. Nakayasu, Y. Yoshiura, T. Iida, A. Yamamoto, Adaptive immune response to Edwardsiella tarda infection in ginbuna crucian carp, Carassius auratus langsdorfii, Veterinary immunology and immunopathology 153(1-2) (2013) 83-90. [CrossRef]
- Q. Wang, W. Ji, Z. Xu, Current use and development of fish vaccines in China, Fish & shellfish immunology 96 (2020) 223-234. [CrossRef]
- B. Swain, C.T. Powell, R. Curtiss, 3rd, Pathogenicity and immunogenicity of Edwardsiella piscicida ferric uptake regulator (fur) mutations in zebrafish, Fish Shellfish Immunol 107(Pt B) (2020) 497-510. [CrossRef]
- Y. Yu, Q. Wang, Z. Huang, L. Ding, Z. Xu, Immunoglobulins, Mucosal Immunity and Vaccination in Teleost Fish, Front Immunol 11 (2020) 567941. [CrossRef]
- S.P. Wamala, K.K. Mugimba, S. Mutoloki, Ø. Evensen, R. Mdegela, D.K. Byarugaba, H. Sørum, Occurrence and antibiotic susceptibility of fish bacteria isolated from Oreochromis niloticus (Nile tilapia) and Clarias gariepinus (African catfish) in Uganda, Fisheries and Aquatic Sciences 21(1) (2018) 6. [CrossRef]
- M.K. Viršek, M.N. Lovšin, Š. Koren, A. Kržan, M. Peterlin, Microplastics as a vector for the transport of the bacterial fish pathogen species Aeromonas salmonicida, Marine Pollution Bulletin 125(1) (2017) 301-309. [CrossRef]
- S. Kumari, Freshwater and marine water fish diseases: A review, Int J Fish Aquat Stud 8(4) (2020) 4.
- L. Gui, V.G. Chinchar, Q. Zhang, Molecular basis of pathogenesis of emerging viruses infecting aquatic animals, Aquaculture and Fisheries 3(1) (2018) 1-5. [CrossRef]
- J.C. Leong, Fish Viruses, Encyclopedia of Virology (2008) 227-234. [CrossRef]
- L.B. Hølvold, A.I. Myhr, R.A. Dalmo, Strategies and hurdles using DNA vaccines to fish, Veterinary research 45(1) (2014) 21-21. [CrossRef]
- F. Athanasopoulou, I. Pappas, K. Bitchava, An overview of the treatments of parasitic disease in Mediterranean aquaculture, Options Méditerranéennes 86 (2009).
- R.Y. Ruth Francis-Floyd, and Deborah Pouder, Ichthyophthirius multifiliis (White Spot) Infections in Fish, Florida Cooperative Extension Service, Institute of Food and Agricultural Sciences, University of Florida (2018).
- Bridging America and Russia with shared perspectives on aquatic animal health: Proceedings of the Third Bilateral Conference Between the United States and Russia, Aquatic Animal Health 2009, Khaled bin Sultan Living Oceans Foundation, Landover, MD, 2011.
- A.E. Toranzo, J.L. Romalde, B. Magariños, J.L. Barja, Present and future of aquaculture vaccines against fish bacterial diseases, in: B. Basurco, C. Rogers (Eds.), The use of veterinary drugs and vaccines in Mediterranean aquaculture, Zaragoza : CIHEAM2009, pp. 155–176.
- N. Bujan, A. Toranzo, B. Magarinos, Edwardsiella piscicida: a significant bacterial pathogen of cultured fish, Diseases of Aquatic Organisms 131 (2018). [CrossRef]
- K.Y. Leung, Q. Wang, Z. Yang, B.A. Siame, Edwardsiella piscicida: A versatile emerging pathogen of fish, Virulence 10(1) (2019) 555-567. [CrossRef]
- S.B. Park, T. Aoki, T.S. Jung, Pathogenesis of and strategies for preventing Edwardsiella tarda infection in fish, Veterinary Research 43(1) (2012) 67. [CrossRef]
- B. Swain, C.T. Powell, R. Curtiss, Virulence, immunogenicity and live vaccine potential of aroA and phoP mutants of Edwardsiella piscicida in zebrafish, Microbial Pathogenesis (2021) 105355. [CrossRef]
- M. Sayed, O. Ozdemir, M. Essa, A. Olivier, A. Karsi, M.L. Lawrence, H. Abdelhamed, Virulence and live vaccine potential of Edwardsiella piscicida phoP and phoQ mutants in catfish against edwardsiellosis, J Fish Dis 44(9) (2021) 1463-1474. [CrossRef]
- W. Yang, L. Wang, L. Zhang, J. Qu, Q. Wang, Y. Zhang, An invasive and low virulent Edwardsiella tarda esrB mutant promising as live attenuated vaccine in aquaculture, Appl Microbiol Biotechnol 99(4) (2015) 1765-77. [CrossRef]
- B. Swain, C.T. Powell, R. Curtiss, 3rd, Construction and Evaluation of Recombinant Attenuated Edwardsiella piscicida Vaccine (RAEV) Vector System Encoding Ichthyophthirius multifiliis (Ich) Antigen IAG52B, Frontiers in immunology 12 (2021) 802760. [CrossRef]
- A.O. Kordon, H. Abdelhamed, H. Ahmed, W. Baumgartner, A. Karsi, L.M. Pinchuk, Assessment of the Live Attenuated and Wild-Type Edwardsiella ictaluri-Induced Immune Gene Expression and Langerhans-Like Cell Profiles in the Immune-Related Organs of Catfish, Front Immunol 10 (2019) 392. [CrossRef]
- H. Abdelhamed, M.L. Lawrence, A. Karsi, Development and Characterization of a Novel Live Attenuated Vaccine Against Enteric Septicemia of Catfish, Front Microbiol 9 (2018) 1819-1819. [CrossRef]
- D.J. Wise, T.E. Greenway, T.S. Byars, M.J. Griffin, L.H. Khoo, Oral Vaccination of Channel Catfish against Enteric Septicemia of Catfish Using a Live Attenuated Edwardsiella ictaluri Isolate, Journal of aquatic animal health 27(2) (2015) 135-43. [CrossRef]
- N. Dahal, H. Abdelhamed, J. Lu, A. Karsi, M.L. Lawrence, Tricarboxylic Acid Cycle and One-Carbon Metabolism Pathways Are Important in Edwardsiella ictaluri Virulence, PLoS One 8(6) (2013) e65973. [CrossRef]
- S.W. Nho, H. Abdelhamed, A. Karsi, M.L. Lawrence, Improving safety of a live attenuated Edwardsiella ictaluri vaccine against enteric septicemia of catfish and evaluation of efficacy, Veterinary Microbiology 210 (2017) 83-90. [CrossRef]
- M.R. Delghandi, M. El-Matbouli, S. Menanteau-Ledouble, Renibacterium salmoninarum-The Causative Agent of Bacterial Kidney Disease in Salmonid Fish, Pathogens 9(10) (2020) 845. [CrossRef]
- D.G. Elliott, G.D. Wiens, K.L. Hammell, L.D. Rhodes, Vaccination against Bacterial Kidney Disease, Fish Vaccination2014, pp. 255–272. [CrossRef]
- O. Brynildsrud, E.J. Feil, J. Bohlin, S. Castillo-Ramirez, D. Colquhoun, U. McCarthy, I.M. Matejusova, L.D. Rhodes, G.D. Wiens, D.W. Verner-Jeffreys, Microevolution of Renibacterium salmoninarum: evidence for intercontinental dissemination associated with fish movements, Isme j 8(4) (2014) 746-56. [CrossRef]
- I. Matejusova, N. Bain, D.J. Colquhoun, E.J. Feil, U. McCarthy, D. McLennan, M. Snow, D. Verner-Jeffreys, I.S. Wallace, S.J. Weir, M. Hall, Multilocus variable-number tandem-repeat genotyping of Renibacterium salmoninarum, a bacterium causing bacterial kidney disease in salmonid fish, BMC Microbiology 13(1) (2013) 285. [CrossRef]
- J. Bethke, M. Poblete-Morales, R. Irgang, A. Yáñez, R. Avendaño-Herrera, Iron acquisition and siderophore production in the fish pathogen Renibacterium salmoninarum, J Fish Dis 39(11) (2016) 1275-1283. [CrossRef]
- T. Zhou, Z. Yuan, S. Tan, Y. Jin, Y. Yang, H. Shi, W. Wang, D. Niu, L. Gao, W. Jiang, D. Gao, Z. Liu, A Review of Molecular Responses of Catfish to Bacterial Diseases and Abiotic Stresses, Front Physiol 9 (2018) 1113-1113. [CrossRef]
- H. Mohammed, O. Olivares-Fuster, S. LaFrentz, C.R. Arias, New attenuated vaccine against columnaris disease in fish: choosing the right parental strain is critical for vaccine efficacy, Vaccine 31(45) (2013) 5276-80. [CrossRef]
- G. Rathore, Bacterial Vaccines for Fishes: Current Status, PROPHYLAXIS IN AQUACULTURE (2017) 2.
- S. Bravo, Environmental impacts and management of veterinary medicines in aquaculture: the case of salmon aquaculture in Chile, Improving biosecurity through prudent and responsible use of veterinary medicines in aquatic food production (2012) 11.
- A. Gavriilidou, J. Gutleben, D. Versluis, F. Forgiarini, M.W.J. van Passel, C.J. Ingham, H. Smidt, D. Sipkema, Comparative genomic analysis of Flavobacteriaceae: insights into carbohydrate metabolism, gliding motility and secondary metabolite biosynthesis, BMC Genomics 21(1) (2020) 569. [CrossRef]
- F. Dumetz, S.E. Lapatra, E. Duchaud, S. Claverol, M. Le Hénaff, The Flavobacterium psychrophilum OmpA, an outer membrane glycoprotein, induces a humoral response in rainbow trout, J Appl Microbiol 103(5) (2007) 1461-70. [CrossRef]
- J. Bøgwald, R.A. Dalmo, Review on Immersion Vaccines for Fish: An Update 2019, Microorganisms 7(12) (2019) 627. [CrossRef]
- P.S. Sudheesh, K.D. Cain, Prospects and challenges of developing and commercializing immersion vaccines for aquaculture, International Biology Review 1(1) (2017).
- J. Ma, T.J. Bruce, P.S. Sudheesh, C. Knupp, T.P. Loch, M. Faisal, K.D. Cain, Assessment of cross-protection to heterologous strains of Flavobacterium psychrophilum following vaccination with a live-attenuated coldwater disease immersion vaccine, J Fish Dis 42(1) (2019) 75-84. [CrossRef]
- T.J. Bruce, J. Ma, C. Knupp, T.P. Loch, M. Faisal, K.D. Cain, Cross-protection of a live-attenuated Flavobacterium psychrophilum immersion vaccine against novel Flavobacterium spp. and Chryseobacterium spp. strains, J Fish Dis 43(8) (2020) 915-928. [CrossRef]
- M.D. Fast, B. Tse, J.M. Boyd, S.C. Johnson, Mutations in the Aeromonas salmonicida subsp. salmonicida type III secretion system affect Atlantic salmon leucocyte activation and downstream immune responses, Fish Shellfish Immunol 27(6) (2009) 721-8. [CrossRef]
- B.K. Gudmundsdottir, Bjornsdottir, B., Aeromonas salmonicida and A. hydrophila., 2017. [CrossRef]
- L.M. Braden, S.K. Whyte, A.B.J. Brown, C.V. Iderstine, C. Letendre, D. Groman, J. Lewis, S.L. Purcell, T. Hori, M.D. Fast, Vaccine-Induced Protection Against Furunculosis Involves Pre-emptive Priming of Humoral Immunity in Arctic Charr, Frontiers in Immunology 10(120) (2019). [CrossRef]
- P. Vanden Bergh, M. Heller, S. Braga-Lagache, J. Frey, The Aeromonas salmonicida subsp. salmonicida exoproteome: determination of the complete repertoire of Type-Three Secretion System effectors and identification of other virulence factors, Proteome Sci 11(1) (2013) 42. [CrossRef]
- H. Mikkelsen, M.B. Schrøder, V. Lund, Vibriosis and atypical furunculosis vaccines; efficacy, specificity and side effects in Atlantic cod, Gadus morhua L, Aquaculture 242(1-4) (2004) 81-91. [CrossRef]
- G. Rørstad, P.M. Aasjord, B. Robertsen, Adjuvant effect of a yeast glucan in vaccines against furunculosis in Atlantic salmon (Salmo salar L.), Fish & Shellfish Immunology 3(3) (1993) 179-190. [CrossRef]
- A. Toranzo, J. Romalde, B. Magariños, J. Barja, Present and future of aquaculture vaccines against fish bacterial diseases, Options Mediterraneennes 86 (2009) 155-176.
- T. Erkinharju, M.R. Lundberg, E. Isdal, I. Hordvik, R.A. Dalmo, T. Seternes, Studies on the antibody response and side effects after intramuscular and intraperitoneal injection of Atlantic lumpfish (Cyclopterus lumpus L.) with different oil-based vaccines, J Fish Dis 40(12) (2017) 1805-1813. [CrossRef]
- M.H. Marana, D. Sepúlveda, D. Chen, A. Al-Jubury, R.M. Jaafar, P.W. Kania, N.H. Henriksen, B. Krossøy, I. Dalsgaard, N. Lorenzen, K. Buchmann, A pentavalent vaccine for rainbow trout in Danish aquaculture, Fish Shellfish Immunol 88 (2019) 344-351. [CrossRef]
- K.R. Villumsen, P.W. Kania, D. Christensen, E.O. Koppang, A.M. Bojesen, Injection Vaccines Formulated with Nucleotide, Liposomal or Mineral Oil Adjuvants Induce Distinct Differences in Immunogenicity in Rainbow Trout, Vaccines (Basel) 8(1) (2020). [CrossRef]
- R. Hoare, S.J. Jung, T.P.H. Ngo, K. Bartie, J. Bailey, K.D. Thompson, A. Adams, Efficacy and safety of a non-mineral oil adjuvanted injectable vaccine for the protection of Atlantic salmon (Salmo salar L.) against Flavobacterium psychrophilum, Fish Shellfish Immunol 85 (2019) 44-51. [CrossRef]
- H. Tziouvas, a.P. Varvarigos, Intensity scale of side effects in European sea bass (Dicentrarchus labrax) post intraperitoneal injection with commercial oil-adjuvanted vaccines Bulletin of the European Association of Fish Pathologist 41(3) (2021) 8. [CrossRef]
- J.C. Thornton, R.A. Garduq o, W.W. Kay, The development of live vaccines for furunculosis lacking the A-layer and O-antigen of Aeromonas salmonicida, Journal of Fish Diseases 17 (1994) 195-204. [CrossRef]
- S. Menanteau-Ledouble, M. El-Matbouli, Antigens of Aeromonas salmonicida subsp. salmonicida specifically induced in vivo in Oncorhynchus mykiss, Journal of fish diseases 39(8) (2016) 1015-1019. [CrossRef]
- X.-D. Ling, W. Dong, Y. Zhang, J. Hu, J.-x. Liu, X. Zhao, A recombinant adenovirus targeting typical Aeromonas salmonicida induces an antibody-mediated adaptive immune response after immunization of rainbow trout, Microbial pathogenesis 133 (2019) 103559. [CrossRef]
- S.M. Bergmann, Y. Jin, K. Franzke, B. Grunow, Q. Wang, S. Klafack, Koi herpesvirus (KHV) and KHV disease (KHVD) – a recently updated overview, Journal of Applied Microbiology 129(1) (2020) 98-103. [CrossRef]
- L. Schröder, S. Klafack, S.M. Bergmann, D. Fichtner, Y. Jin, P.Y. Lee, D. Höper, T.C. Mettenleiter, W. Fuchs, Generation of a potential koi herpesvirus live vaccine by simultaneous deletion of the viral thymidine kinase and dUTPase genes, J Gen Virol 100(4) (2019) 642-655. [CrossRef]
- L.-C. Cui, X.-T. Guan, Z.-M. Liu, C.-Y. Tian, Y.-G. Xu, Recombinant lactobacillus expressing G protein of spring viremia of carp virus (SVCV) combined with ORF81 protein of koi herpesvirus (KHV): A promising way to induce protective immunity against SVCV and KHV infection in cyprinid fish via oral vaccination, Vaccine 33(27) (2015) 3092-3099. [CrossRef]
- F. Hu, Y. Li, Q. Wang, G. Wang, B. Zhu, Y. Wang, W. Zeng, J. Yin, C. Liu, S.M. Bergmann, C. Shi, Carbon nanotube-based DNA vaccine against koi herpesvirus given by intramuscular injection, Fish & Shellfish Immunology 98 (2020) 810-818. [CrossRef]
- M.M.D. Peñaranda, S.E. LaPatra, G. Kurath, Specificity of DNA vaccines against the U and M genogroups of infectious hematopoietic necrosis virus (IHNV) in rainbow trout (Oncorhynchus mykiss), Fish & Shellfish Immunology 31(1) (2011) 43-51. [CrossRef]
- E.T. Larragoite, L. Tacchi, S.E. LaPatra, I. Salinas, An attenuated virus vaccine appears safe to the central nervous system of rainbow trout (Oncorhynchus mykiss) after intranasal delivery, Fish Shellfish Immunol 49 (2016) 351-4. [CrossRef]
- L. Tang, H. Kang, K. Duan, M. Guo, G. Lian, Y. Wu, Y. Li, S. Gao, Y. Jiang, J. Yin, M. Liu, Effects of Three Types of Inactivation Agents on the Antibody Response and Immune Protection of Inactivated IHNV Vaccine in Rainbow Trout, Viral Immunol 29(7) (2016) 430-5. [CrossRef]
- R.N. Rouxel, C. Tafalla, E. Mérour, E. Leal, S. Biacchesi, M. Brémont, D.S. Lyles, Attenuated Infectious Hematopoietic Necrosis Virus with Rearranged Gene Order as Potential Vaccine, Journal of Virology 90(23) (2016) 10857-10866. [CrossRef]
- A. Long, J. Richard, L. Hawley, S.E. LaPatra, K.A. Garver, Transmission potential of infectious hematopoietic necrosis virus in APEX-IHN®-vaccinated Atlantic salmon, Dis Aquat Organ 122(3) (2017) 213-221. [CrossRef]
- C.Y. Yong, H.K. Ong, H.C. Tang, S.K. Yeap, A.R. Omar, K.L. Ho, W.S. Tan, Infectious hematopoietic necrosis virus: advances in diagnosis and vaccine development, PeerJ 7 (2019) e7151. [CrossRef]
- C.M. Caipang, T. Takano, I. Hirono, T. Aoki, Genetic vaccines protect red seabream, Pagrus major, upon challenge with red seabream iridovirus (RSIV), Fish Shellfish Immunol 21(2) (2006) 130-8. [CrossRef]
- K. Nakajima, Y. Maeno, A. Honda, K. Yokoyama, T. Tooriyama, S. Manabe, Effectiveness of a vaccine against red sea bream iridoviral disease in a field trial test, Dis Aquat Organ 36(1) (1999) 73-5. [CrossRef]
- M.H. Jung, C. Nikapitiya, S.J. Jung, DNA vaccine encoding myristoylated membrane protein (MMP) of rock bream iridovirus (RBIV) induces protective immunity in rock bream (Oplegnathus fasciatus), Vaccine 36(6) (2018) 802-810. [CrossRef]
- S.Y. Oh, M.J. Oh, T. Nishizawa, Potential for a live red seabream iridovirus (RSIV) vaccine in rock bream Oplegnathus fasciatus at a low rearing temperature, Vaccine 32(3) (2014) 363-8. [CrossRef]
- S. Senapin, H.T. Dong, W. Meemetta, W. Gangnonngiw, P. Sangsuriya, R. Vanichviriyakit, M. Sonthi, B. Nuangsaeng, Mortality from scale drop disease in farmed Lates calcarifer in Southeast Asia, J Fish Dis 42(1) (2019) 119-127. [CrossRef]
- K. Thanasaksiri, K. Fukuda, S. Tsubone, H. Miyadai, T. Murakami, A. Murakami, R. Takano, Efficacy of a bivalent inactivated vaccine against red seabream iridovirus and Streptococcus iniae in red seabream, Pagrus major, Aquaculture 492 (2018) 132-136. [CrossRef]
- I. Deperasińska, P. Schulz, A.K. Siwicki, Salmonid Alphavirus (SAV), J Vet Res 62(1) (2018) 1-6. [CrossRef]
- D.A. Graham, H.R. Rowley, P. Frost, Cross-neutralization studies with salmonid alphavirus subtype 1-6 strains: results with sera from experimental studies and natural infections, J Fish Dis 37(8) (2014) 683-91. [CrossRef]
- B. Bang Jensen, A.B. Kristoffersen, C. Myr, E. Brun, Cohort study of effect of vaccination on pancreas disease in Norwegian salmon aquaculture, Dis Aquat Organ 102(1) (2012) 23-31. [CrossRef]
- C.J. Chang, J. Gu, B. Robertsen, Protective effect and antibody response of DNA vaccine against salmonid alphavirus 3 (SAV3) in Atlantic salmon, J Fish Dis 40(12) (2017) 1775-1781. [CrossRef]
- C.J. Chang, J. Gu, B. Robertsen, Protective effect and antibody response of DNA vaccine against salmonid alphavirus 3 (SAV3) in Atlantic salmon, Journal of Fish Diseases 40(12) (2017) 1775-1781. [CrossRef]
- C. Collins, K. Lester, J. Del-Pozo, B. Collet, Non-Lethal Sequential Individual Monitoring of Viremia in Relation to DNA Vaccination in Fish-Example Using a Salmon Alphavirus DNA Vaccine in Atlantic Salmon Salmo salar, Vaccines (Basel) 9(2) (2021). [CrossRef]
- J. Pajdak-Czaus, P. Schulz, E. Terech-Majewska, W. Szweda, A.K. Siwicki, A. Platt-Samoraj, Influence of Infectious Pancreatic Necrosis Virus and Yersinia ruckeri Co-Infection on a Non-Specific Immune System in Rainbow Trout (Oncorhynchus mykiss), Animals (Basel) 11(7) (2021) 1974. [CrossRef]
- S. Kanrar, A.K. Dhar, Complete Genome Sequence of a Novel Mutant Strain of Vibrio parahaemolyticus from Pacific White Shrimp (Penaeus vannamei), Genome Announcements 6(24) (2018) e00497-18. [CrossRef]
- N.A. Ballesteros, S. Rodriguez Saint-Jean, S.I. Perez-Prieto, Food pellets as an effective delivery method for a DNA vaccine against infectious pancreatic necrosis virus in rainbow trout (Oncorhynchus mykiss, Walbaum), Fish & Shellfish Immunology 37(2) (2014) 220-228. [CrossRef]
- S. Ahmadivand, M. Soltani, M. Behdani, Ø. Evensen, E. Alirahimi, E. Soltani, R. Hassanzadeh, J. Ashrafi-Helan, VP2 (PTA motif) encoding DNA vaccine confers protection against lethal challenge with infectious pancreatic necrosis virus (IPNV) in trout, Molecular Immunology 94 (2018) 61-67. [CrossRef]
- F.S. Kibenge, K. Munir, M.J. Kibenge, T. Joseph, E. Moneke, Infectious salmon anemia virus: causative agent, pathogenesis and immunity, Anim Health Res Rev 5(1) (2004) 65-78. [CrossRef]
- F.S. Kibenge, Kibenge, M. J., Joseph, T., & McDougall, J. , The development of infectious salmon anemia virus vaccines in Canada, In Miller, Otis; Cipriano, Rocco C., tech. coords. International response to infectious salmon anemia: prevention, control, and eradication: proceedings of a symposium, US Department of Agriculture, Animal and Plant Health Inspection Service; US Department of the Interior, US Geological Survey; US Department of Commerce, National Marine Fisheries Service, New Orleans, LA. Tech. Bull. 1902. Washington, DC, 2003, pp. 39–49.
- A. Wolf, K. Hodneland, P. Frost, S. Braaen, E. Rimstad, A hemagglutinin-esterase-expressing salmonid alphavirus replicon protects Atlantic salmon (Salmo salar) against infectious salmon anemia (ISA), Vaccine 31(4) (2013) 661-9. [CrossRef]
- C.J. Chang, B. Sun, B. Robertsen, Adjuvant activity of fish type I interferon shown in a virus DNA vaccination model, Vaccine 33(21) (2015) 2442-8. [CrossRef]
- M.S. Ahasan, W. Keleher, C. Giray, B. Perry, W. Surachetpong, P. Nicholson, L. Al-Hussinee, K. Subramaniam, T.B. Waltzek, Genomic Characterization of Tilapia Lake Virus Isolates Recovered from Moribund Nile Tilapia (Oreochromis niloticus) on a Farm in the United States, Microbiol Resour Announc 9(4) (2020) e01368-19. [CrossRef]
- M. Abbadi, A. Basso, L. Biasini, R. Quartesan, A. Buratin, N. Davidovich, A. Toffan, Tilapia lake virus: A structured phylogenetic approach, Frontiers in genetics 14 (2023) 1069300. [CrossRef]
- M.D. Jansen, H.T. Dong, C.V. Mohan, Tilapia lake virus: a threat to the global tilapia industry?, Reviews in Aquaculture 11(3) (2019) 725-739. [CrossRef]
- Y. Thawornwattana, H.T. Dong, K. Phiwsaiya, P. Sangsuriya, S. Senapin, P. Aiewsakun, Tilapia lake virus (TiLV): Genomic epidemiology and its early origin, Transboundary and Emerging Diseases 68(2) (2021) 435-444. [CrossRef]
- W. Zeng, Y. Wang, H. Hu, Q. Wang, S.M. Bergmann, Y. Wang, B. Li, Y. Lv, H. Li, J. Yin, Y. Li, Cell Culture-Derived Tilapia Lake Virus-Inactivated Vaccine Containing Montanide Adjuvant Provides High Protection against Viral Challenge for Tilapia, Vaccines (Basel) 9(2) (2021). [CrossRef]
- T.T. Mai, P. Kayansamruaj, C. Soontara, P. Kerddee, D.-H. Nguyen, S. Senapin, J.Z. Costa, J. del-Pozo, K.D. Thompson, C. Rodkhum, H.T. Dong, Immunization of Nile Tilapia (Oreochromis niloticus) Broodstock with Tilapia Lake Virus (TiLV) Inactivated Vaccines Elicits Protective Antibody and Passive Maternal Antibody Transfer, Vaccines 10(2) (2022) 167. [CrossRef]
- S. Shivam, M. El-Matbouli, G. Kumar, Development of Fish Parasite Vaccines in the OMICs Era: Progress and Opportunities, Vaccines 9(2) (2021) 179. [CrossRef]
- L.T. Barrett, F. Oppedal, N. Robinson, T. Dempster, Prevention not cure: a review of methods to avoid sea lice infestations in salmon aquaculture, Reviews in Aquaculture 12(4) (2020) 2527-2543. [CrossRef]
- A. Sitjà-Bobadilla, Living off a fish: a trade-off between parasites and the immune system, Fish & shellfish immunology 25 4 (2008) 358-72. [CrossRef]
- R.A. Khan, Host-Parasite Interactions in Some Fish Species, Journal of Parasitology Research 2012 (2012) 237280. [CrossRef]
- A. Sfacteria, M. Brines, U. Blank, The mast cell plays a central role in the immune system of teleost fish, Mol Immunol 63(1) (2015) 3-8. [CrossRef]
- B.S. Dezfuli, T. Bo, M. Lorenzoni, A.P. Shinn, L. Giari, Fine structure and cellular responses at the host-parasite interface in a range of fish-helminth systems, Vet Parasitol 208(3-4) (2015) 272-9. [CrossRef]
- M.N. Faber, J.W. Holland, C.J. Secombes, Vaccination strategies and IgM responses against PKD in rainbow trout, Fish & Shellfish Immunology 91 (2019) 423. [CrossRef]
- Y. Carpio, L. Basabe, J. Acosta, A. Rodríguez, A. Mendoza, A. Lisperger, E. Zamorano, M. González, M. Rivas, S. Contreras, D. Haussmann, J. Figueroa, V.N. Osorio, G. Asencio, J. Mancilla, G. Ritchie, C. Borroto, M.P. Estrada, Novel gene isolated from Caligus rogercresseyi: A promising target for vaccine development against sea lice, Vaccine 29(15) (2011) 2810-2820. [CrossRef]
- P. Guragain, M. Tkachov, A.S. Båtnes, Y. Olsen, P. Winge, A.M. Bones, Principles and Methods of Counteracting Harmful Salmon–Arthropod Interactions in Salmon Farming: Addressing Possibilities, Limitations, and Future Options, Frontiers in Marine Science 8(965) (2021). [CrossRef]
- J.Y. Yao, X.M. Yuan, Y. Xu, W.L. Yin, L.Y. Lin, X.Y. Pan, G.L. Yang, C.F. Wang, J.Y. Shen, Live recombinant Lactococcus lactis vaccine expressing immobilization antigen (i-Ag) for protection against Ichthyophthirius multifiliis in goldfish, Fish Shellfish Immunol 58 (2016) 302-308. [CrossRef]
- L.v.G. Jørgensen, The fish parasite Ichthyophthirius multifiliis – Host immunology, vaccines and novel treatments, Fish & Shellfish Immunology 67 (2017) 586-595. [CrossRef]
- L. von Gersdorff Jørgensen, P.W. Kania, K.J. Rasmussen, A.H. Mattsson, J. Schmidt, A. Al-Jubury, A. Sander, A. Salanti, K. Buchmann, Rainbow trout (Oncorhynchus mykiss) immune response towards a recombinant vaccine targeting the parasitic ciliate Ichthyophthirius multifiliis, J Fish Dis 40(12) (2017) 1815-1821. [CrossRef]
- M. Kang, F. Feng, Y. Wang, L. Guo, L. Chen, K. Chen, Advances in Research into Oral Vaccines for Fish, 2018.
- C.W.E. Embregts, M. Forlenza, Oral vaccination of fish: Lessons from humans and veterinary species, Developmental & Comparative Immunology 64 (2016) 118-137. [CrossRef]
- H.M. Munang'andu, S. Mutoloki, Ø. Evensen, An Overview of Challenges Limiting the Design of Protective Mucosal Vaccines for Finfish, Front Immunol 6 (2015) 542-542. [CrossRef]
- M.R. Neutra, P.A. Kozlowski, Mucosal vaccines: the promise and the challenge, Nature Reviews Immunology 6(2) (2006) 148-158. [CrossRef]
- Z. Cao, S. Liu, H. Nan, K. Zhao, X. Xu, G. Wang, H. Ji, H. Chen, Immersion immunization with recombinant baculoviruses displaying cyprinid herpesvirus 2 membrane proteins induced protective immunity in gibel carp, Fish & shellfish immunology 93 (2019) 879-887. [CrossRef]
- C. Shoemaker, P. Klesius, J. Evans, C. Arias, Use of Modified Live Vaccines in Aquaculture, J World Aquac Soc 40 (2009). [CrossRef]
- S. Li, H. Xie, Z. Yan, B. Li, P. Wu, X. Qian, X. Zhang, J. Wu, J. Liu, X. Zhao, Development of a live vector vaccine against infectious hematopoietic necrosis virus in rainbow trout, Fish & shellfish immunology 89 (2019) 516-524. [CrossRef]
- J.L. Clarke, M.T. Waheed, A.G. Lössl, I. Martinussen, H. Daniell, How can plant genetic engineering contribute to cost-effective fish vaccine development for promoting sustainable aquaculture?, Plant molecular biology 83(1-2) (2013) 33-40. [CrossRef]
- M. Yimer, S. Tesfaye, B. Birhanu, Present Status and Future Prospects of Fish Vaccination: A Review, Journal of Veterinary Science & Technology 07 (2016). [CrossRef]
- C. Flores-Kossack, R. Montero, B. Köllner, K. Maisey, Chilean aquaculture and the new challenges: Pathogens, immune response, vaccination and fish diversification, Fish Shellfish Immunol 98 (2020) 52-67. [CrossRef]

Table 1.
List of the most common bacterial, viral and parasitic diseases in aquaculture.
| Name of the Diseases | Causative Agent | Fish it Affects |
|---|---|---|
| Bacterial | ||
| Atypical furunculosis | Aeromonas salmonicida | Salmonids |
| Motile aeromonid septicemia | Aeromonas hydrophila, A. caviae, A. sobria | Catfish, salmonids, cyprinids |
| Vibriosis | Vibrio anguillarum, V. ordalii | Marine Fish including: Salmonids, Yellowtail, Halibut, Amberjack |
| Enteric septicemia | Edwardsiella ictaluri | Catfish |
| Edwardsiellosis | Edwardsiella piscicida | Catfish, Striped Bass, Tilapia, Sea bream |
| Enteric septicemia | Edwardsiella tarda | Catfish, eel, hirame |
| Tuberculosis | Mycobacterium marinum, M. fortuitum, M. chelonae | Sea bass, tropical aquarium fish |
| Streptococcosis | Streptococcus parauberis | Turbot |
| Streptococcosis | Streptococcus phocae | Atlantic salmon |
| Flavobacteriosis | Flavobacterium psychrophilum | Salmonids, freshwater fish |
| Rainbow trout fry syndrome | Flavobacterium psychrophilum | Salmonids, freshwater Fish |
| Viral | ||
| Tilapia Lake Virus | Tilapia Tilapinevirus | Tilapia and hybrid tilapia fish |
| Infectious Hemorrhagic necrosis | Rhabdovirus | Snakehead, carp, barbs |
| Infectious salmon anemia | Orthomyxovirus | Atlantic salmon |
| Infectious pancreatic necrosis | Birnavirus | Salmonids, Sea brass, Sea bream, Pacific Cod |
| Koi Herpes Virus | Herpesvirus | Cyprinid Fish |
| Viral nervous necrosis | Betanovirus | Marine fish species including Sea brass, groupers, halibut |
| Salmonid Alphavirus | Alphavirus | Altantic Salmon, Rainbow Trout |
| Iridoviral disease | Iridovirus | Amberjack, Yellowtail, Red Sea Bream |
| Costiasis | Ichthyobodo necotor | Several Freshwater and Saltwater fish |
| Salmon Poisoning disease | Nanophyetus salmincola | Salmon, several Freshwater Fish |
| White Spot | Ichthyophthirius mulifiliis | Freshwater Fish |
| Sea Lice | Lepeophtheirus solmonis | Marine Salmonids |
| Whirling Disease | Myxobolus cerebralis | Trout, Salmon, Whitefish |
| Myxosporeans | Myxobolus genera | Freshwater and Marine Fish |
| Microsporean | Pleistophora genera | Freshwater and Marine Fish |
Table 2.
List of licensed bacterial vaccines.
| Disease | Pathogen | Host | Type of Vaccine | Route of delivery | Trade Name | Country |
|---|---|---|---|---|---|---|
| Enteric septicaemia of catfish (ESC) | Edwardsiella ictaluri | catfish | Live attenuated | Immersion | Aquavac-ESC | US |
| Bacterial Kidney Disease (BKD) | Renibacterium salmoninarum | salmonids | Live, Attenuated | IP | Renogen | US Canada Chile |
| Flavobacteriosis/ Columnaris |
Flavobacterium columnare Flavobacterium maritimus |
cyprinids, salmonids, catfish, carp | Live, Attenuated | Immersion | Aquavac-Col | US Canada Chile |
| Inactivated | IP | Alpha Ject® IPNVFlevo 0.025 | Chile | |||
| Killed bacterin | Immersion | FryVacc 1 | US Canada |
|||
| FryVacc 2 | Chile | |||||
| Furunculosis | Aeromonas salmonicida | Atlantic Salmon & Rainbow Trout |
Inactivated, oil-based | IP | AlphaJect 3000 | Denmark Finland Iceland Ireland Norway Sweden |
| Alpha Ject® 2.2 | UK | |||||
| Alpha Ject® 4-1, Alpha Ject® 5-1 | Chile | |||||
| Alpha Ject® 6-2 | Norway The Faroe Islands |
|||||
| Alpha Ject® micro 7 ILA | Norway The Faroe Islands |
|||||
| Subunit Vaccine | IP | Norvax® Minova 6 | Norway | |||
| Inactivated Bacterin | IP | AquaVac-FNM | UK Ireland Spain France |
|||
| Killed Bacterin | IP | Lipogen Forte, Furogen Dip, Forte VI | US Canada |
|||
| Streptococcosis | Streptococcus iniae | tilapia and seabass | Inactivated | IP or Bath | Norvax Strep Si, Aquavac Strep Sa | Vietnam Honduras Indonesia |
| tilapia | Killed | IP | Aquavac-Garvetil | Honduras Venezuela Ecuador The Philippines Indonesia |
||
|
Streptococcus agalactiae |
tilapia | Inactivated | IP | AlphaJect® micro1 TiLa | Brazil Colombia Honduras Indonesia Panama |
|
|
Streptococcus parauberis |
Turbot | Inactivated | IP | Icthiovac-STR | Spain | |
| Vibriosis |
V. anguillarum V. ordalii |
Atlantic Salmon | Inactivated, Oil-based | IP | Alpha Ject® micro 7 ILA, Alpha Ject® 6-2 | Norway The Faroe Islands |
| Inactivated, oil-based | IP | Alpha Ject® 5-1, Alpha Ject® 4-1, | Chile | |||
| Inactivated, oil-based | IP | Alpha Ject® Micro-4 | Canada | |||
| Subunit Vaccine | IP | Norvax® Minova 6 | Norway | |||
| Inactivated, Oil-based | IP | Alpha Ject® micro 6 | Ireland UK The Faroe Islands Norway |
|||
| Sea bass | Inactivated, Oil-based | IP | Alpha Ject® micro 2000 | Croatia Spain Greece France |
||
| Atlantic salmon | Inactivated, oil-based | IP | Alpha Ject® Micro-3 | Chile | ||
| Atlantic aalmon & Rainbow trout |
Inactivated, oil-based | IP | Alpha Ject® 5-3 | Iceland Norway |
||
| Atlantic salmon & Rainbow trout |
Inactivated, oil-based | IP | AlphaJect 3000 | Denmark Finland Iceland Ireland Norway Sweden |
||
| Sea bass | Inactivated, oil-based | Dip | ALPHA DIP® Vib | Croatia Cyprus Greece Italy Portugal Spain |
||
| Sea bass | Inactivated, oil-based | Bath/ Immersion |
ALPHA DIP® Vibrio | Turkey | ||
| Atlantic salmon | Inactivated, oil-based | IP | Alpha Ject® 2-2 | UK | ||
| Salmonids | Killed bacterin | IP | Furogen Dip, Forte VI, Lipogen Forte |
US Canada |
||
| Salmonids | Killed bacterin | Bath/ Immersion |
Vibrogen-2 | US Canada |
||
| European sea bass | Inactivated bacterin | IP | AquaVac Vibrio Pasteurella | Greece Middle East |
||
| Rainbow trout | Inactivated bacterin | Oral/ Immersion |
AquaVac Vibrio, AquaVac Vibrio Oral Boost | Finland UK Ireland Spain Greece |
Table 3.
List of licensed viral vaccines.
| Virus | Type of Virus (RNA/DNA) | Fish Host | Trade Name (if applicable) | Type of Vaccine | Delivery Method | Licensed for use in the following countries: | Description |
|---|---|---|---|---|---|---|---|
| SAV | RNA | Atlantic Salmon | Norvax Compact PD | Inactivated | Intraperitoneal Injection | Norway Chile UK |
A monovalent vaccine which contains an inactivated strain of SAV subtype 1. |
| SAV | RNA | Atlantic Salmon | Aquavac PD7 | Inactivated | Intraperitoneal Injection | Norway | A polyvalent vaccine which contains seven strains to protect against pancreatic disease, infectious pancreatic necrosis, furunculosis, cold-water vibriosis, vibriosis and winter ulcers.Specifically, to protect against SAV, it contains an inactivated strain of SAV subtype 1. |
| SAV | RNA | Atlantic Salmon | Aquavac PD3 | Inactivated | Intraperitoneal Injection | UK | A polyvalent vaccine which contains an inactivated strain of SAV subtype 1, as well as infectious pancreatic necrosis and furunculosis. |
| SAV | RNA | Atlantic Salmon | Alphaject Micro 1 PD | Inactivated | Intraperitoneal Injection | UK Norway |
A monovalent vaccine which contains the inactivated SAV subtype 3, the SAV strain most dominant in Norway. |
| IPNV | RNA | Atlantic Salmon, Rainbow Trout | AlphaJect 1000 | Inactivated | Intraperitoneal Injection | Chile Norway UK | A monovalent vaccine containing an inactivated form of the virus. |
| IPNV | RNA | Atlantic Salmon | Birnagen Forte | Inactivated | Intraperitoneal Injection | Canada UK |
A monovalent vaccine containing inactivated bacterins and virulins |
| IPNV | RNA | Atlantic Salmon | Aquavac IPN Oral | Recombinant | Oral | US Canada Chile Middle East |
A monovalent vaccine containing capsid proteins VP2 and VP3 |
| IPNV | RNA | Atlantic Salmon, Pacific Salmon, Chinook Salmon, Rainbow Trou | Blueguard IPNV Oral | Inactivated | Oral | Chile | A monovalent vaccine containing 2 inactivated strains of IPNV |
| IPNV | RNA | Rainbow Trout, Atlantic Salmon, Pacific Salmon, Chinook Salmon |
Blueguard IPN Inyectable | Inactivated | Intraperitoneal Injection | Chile | A monovalent vaccine containing 2 strains of inactivated IPNV |
| IPNV | RNA | Atlantic Salmon | AlphaJect IPNV-Flavo 0.025 | Inactivated | Intraperitoneal Injection | Chile | A bivalent vaccine protecting against IPNV and Flavobacteriosis. |
| IPNV | RNA | Atlantic Salmon, Pacific Salmon, Rainbow Trout | AlphaJect Micro 2 | Inactivated | Intraperitoneal Injection | Chile | A bivalent vaccine protecting against IPNV and SRS |
| IPNV | RNA | Atlantic Salmon | AlphaJect 2-2 | Inactivated | Intraperitoneal Injection | UK | A bivalent vaccine protecting against IPNV and Furunculosis. |
| IPNV | RNA | Atlantic Salmon | AlphaJect Micro 3 | Inactivated | Intraperitoneal Injection | Chile | A trivalent vaccine protecting against IPNV, SRS, and Vibriosis. |
| IPNV | RNA | Atlantic Salmon Rainbow Trout |
Blueguard SRS+IPN+Vibrio | Inactivated | Intraperitoneal Injection | Chile | A trivalent vaccine which includes 2 strains of inactivated IPNV and inactivated bacterins to protect against SRS and Vibrio |
| IPNV | RNA | Atlantic Salmon | AlphaJect 4-1 | Inactivated | Intraperitoneal Injection | Chile | A polyvalent vaccine protecting against Furunculosis, SRS, Vibriosis, and IPNV |
| IPNV | RNA | Atlantic Salmon | Pentium Forte Plus | Inactivated | Intraperitoneal Injection | Norway | Contains inactivated whole virus of IPNV, and also protects against Furunculosis, Classical Vibriosis, coldwater vibriosis, and Winter Ulcer. |
| IPNV | RNA | Atlantic Salmon | Norvax Minova 6 | Subunit, inactivated | Intraperitoneal Injection | UK Norway |
A multivalent vaccine which protects against Furunculosis, classical vibriosis, coldwater vibriosis, wound disease and IPNV. It contains a subunit VP2 capsid protein. |
| IPNV | RNA | Atlantic Salmon | AlphaJect Micro 6 | Inactivated | Intraperitoneal Injection | Norway United Kingdom The Faroe Islands Ireland |
A multivalent vaccine protecting against Furunculosis, Vibriosis, Coldwater vibriosis, Winter sore, and IPNV |
| IPNV | RNA | Atlantic Salmon | AlphaJect 6-2 | Inactivated | Intraperitoneal Injection | Norway The Faroe Islands |
A polyvalent vaccine protecting against Furunculosis, Vibriosis. Coldwater vibriosis, Winter sore, and IPNV. |
| IPNV and ISA | RNA | Atlantic Salmon | AlphaJect Micro 4-2 | Inactivated | Intraperitoneal Injection | Chile | A multivalent vaccine protecting against IPNV, Infectious Salmon Anemia (ISA), Vibriosis, and Furunculosis. |
| IPNV and ISA | RNA | Atlantic Salmon | AlphaJect 5-1 | Inactivated | Intraperitoneal Injection | Chile | A polyvalent vaccine protecting against Furunculosis, SRS, Vibriosis, ISA, and IPNV |
| IPNV and ISA | RNA | Atlantic Salmon | AlphaJect Micro 7 | Inactivated | Intraperitoneal Injection | Norway The Faroe Islands |
A multivalent vaccine protecting against, Furunculosis, Vibriosis, Coldwater vibriosis, Winter sore, IPNV, and (ISA). |
| IPNV and ISA | RNA | Atlantic Salmon | Blueguard SRS+IPN+VO+ISA | Subunit and Inactivated | Intraperitoneal Injection | Chile | A polyvalent vaccine containing subunit ISA, inactivated IPNV strain, and bacterins. It protects against ISA, IPNV, SRS, and Vibriosis. |
| IPNV and ISA | RNA | Altantic Salmon | Blueguard IPN+SRS+AS+VO+ISA inyectable | Subunit and Inactivated | Intraperitoneal Injection | Chile | A polyvalent vaccine containing subunit ISA, inactivated IPNV strain, and bacterins. It protects against ISA, IPN, SRS, vibriosis, and furunculosis. |
| ISA | RNA | Atlantic Salmon | AlphaJect Micro 1 ISA | Inactivated | Intraperitoneal Injection | Chile | A monovalent vaccine that includes an inactivated strain of ISA. |
| ISA | RNA | Salmonids | Forte VII | Inactivated | Intraperitoneal Injection | Canada | A polyvalent vaccine which contains inactivated ISA and bacterin. It protects against ISA, Furunculosis, and Vibriosis. |
| Koi Herpes Virus (KHV) | DNA | Cyprinid Fish | KV-3 | Attenuated | Immersion/Injection | Isreal, USA | A live, attenuated monovalent vaccine that protects against KHV. |
| RSIV | DNA | Red Sea Bream, along with cultured marine fish such as yellowtail and sea brass | n.a | Formalin | Intraperitoneal | Japan | A monovalent formalin-based vaccine that fights against RSIV. This was the first vaccine made against the virus. |
| RSIV | DNA | Red Sea Bream, along with cultured marine fish such as yellowtail and sea brass | AQUAVAC® IridoV | Formalin, oil-adjuvant | Intraperitoneal | Singapore | A monovalent vaccine with an inactivated strain of RSIV which targets tilapia and Asian Sea Bass. |
| IHNV | RNA | Salmonids including rainbow trout, steelhead trout and Atlantic Salmon | Apex-IHN | DNA | Intramuscular Injection | Canada, USA | A DNA plasmid vaccine targeting IHNV in salmonids |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



