
Submitted:
20 February 2024
Posted:
27 February 2024
You are already at the latest version
Abstract
A great deal of studies have assessed the expression of noncoding RNAs (ncRNAs) as well as their relevant molecular functions and biological mechanisms in the individuals of alcohol dependence. Alcohol dependence is one of the most prevalent neuropsychological disorders worldwide, and its pathogenesis is intricate and inadequately understood. There is considerable evidence demonstrating substantial links between multiple genetic factors and the development of alcohol dependence. Critical roles of ncRNAs have been emphasized in the pathophysiology of mental diseases including alcohol dependence. In the comprehension of ncRNAs action and their mechanisms of modification, they have emerged as therapeutic targets for a variety of psychiatric illness including alcohol dependence. Worth mentioning, dysregulated expression of ncRNAs has been regularly detected in individuals of alcohol dependence. An in-depth knowledge of roles of ncRNAs and m6A modification may be valuable for the development of a novel treatment against alcohol dependence. In general, a deeper comprehension of functional roles ncRNAs may make significant contributions to the precise diagnosis and/or actual treatment of alcohol dependence. Here in this review, we mostly focused on the up-to-date knowledge regarding alterations and/or modifications in the expression of ncRNAs in alcohol dependence, and then, presented their prospects for future research and therapeutic application with a novel concept of engram-system.
Keywords:
ncRNA
; lncRNA
; miRNA
; autophagy
; gut microbiota
; gut-brain axis
; alcohol dependence
; alcohol use disorder
1. Introduction
Alcohol dependence, which may include alcohol abuse and/or alcohol use disorder, is a kind of prolonged recurring psychiatric disease described by being unable to stop drinking without suffering withdrawal symptoms and/or continuing alcohol use in spite of destructive consequences. The representative symptoms may include habitual alcohol use, loss of regulation over alcohol consumption, and alcohol removal symptoms [1,2]. Probably, genes might be responsible for around half of the risk of alcohol dependence [1]. The consequences are connected with extensive disability as well as substantial medical and/or economic burdens [3], which becomes one of the most general mental diseases globally [4]. Hereafter, the term alcohol dependence practically equates with alcohol addiction, alcohol abuse, alcoholism, and/or alcohol use disorder.
Chronic alcohol exposure yields general neuroadaptations or alterations in gene expression in alcohol dependence [5]. It has been assumed that the controlling system in which non-coding RNAs (ncRNAs) may partake in affecting potential molecular targets of certain signaling pathway that control biological and cellular outcomes, eventually leading to the incidence and/or progress of alcohol dependence. ncRNAs are various bioactive molecules that are generally existing in organisms, mediating multiple biological processes including mRNA splicing, regulation of translation, and/or post-transcriptional modification for altered intracellular signal transduction [6]. A number of investigations grounded on recent biotechnologies have shown that ncRNAs are typically plentiful in central nervous system (CNS) and play a key role in brain homeostasis as well as the pathological progressions of a psychiatric disease via epigenetic mechanisms [7]. ncRNAs have been shown to regulate a variety of ion channels and/or intercellular linking proteins/molecules. Thousands of unique ncRNAs sequences subsist inside cells, which organize a part of the transcriptional background [8]. ncRNAs are kinds of well-designed machineries for defined gene expression with heterogeneous subsets, which could be further separated into microRNAs (miRNAs), long non coding RNAs (lncRNAs), circular RNAs (circRNAs), piwi interacting RNAs (piRNAs), small interfering RNAs (siRNAs), and so forth [9]. Understandings into the possible roles of these ncRNAs in the development of several diseases, predominantly in brain neurological diseases, have compelled ncRNAs potential tools for innovative therapeutic approaches. Dysregulation of these ncRNAs, particularly circRNAs, miRNAs, and lncRNAs has been detected in human individuals of alcohol dependence, which might be related with the beginning and/or advancement of the disorder [10]. (Figure 1)
Epigenetics is a field that investigates genetic alterations in gene expression that do not include changing the DNA sequence. Foremost epigenetic mechanisms contain the well-known regulation by above-mentioned ncRNAs, histone modifications, and DNA/RNA methylation, which are in an area of continuing investigation. In particular, a diverse of studies have suggested that ncRNAs may play an important role in epigenetic control [11]. In addition to genetic variation, various stressors including psychological stressors and environmental social factors can lead to alcohol dependence via epigenetic modifications at the transcriptional level of RNAs [12,13,14,15]. Here, we digest recent research progresses for the purpose of approaching to a superior understanding of ncRNAs/epigenetics and their mechanisms during the pathogenesis of alcohol dependence, which may contribute to gaining a comprehension on the underlying mechanisms for the superior development of tactics against alcohol dependence.
2. Alcohol Dependence and ncRNA
The profile of miRNAs expression has shown that some miRNAs are abnormally expressed in patients with alcohol dependence, which may be involved in the progression of alcohol dependence through several biological mechanisms. For example, more than 35 miRNAs including hsa-miR-553 and let-7f are considerably up-regulated in the individuals of alcohol dependence compared with healthy controls [16]. Plasma miRNAs profiling/analysis has also shown that the concentrations of miR-122-5p, miR-193b-3p, miR-3937 and miR-4507 in plasma are associated to the alcohol consumption, which may play a key role in the pathogenesis of alcohol dependence [17]. The miRNAs-dependent regulation of target genes might be also critical for the pathogenesis of alcohol dependence. Likewise, dysregulated lncRNAs may be found in the brain of individuals with alcohol dependence being related to their pathophysiology, one of which may contribute to the abnormal expression of brain-derived neurotrophic growth factor (BDNF) in patients with alcohol dependence [18]. The expression of several lncRNAs including SNORD3C, HSPA7, and RP11-543H23.2 has been aberrantly detected in different brain regions of patients with alcohol dependence. Some lncRNAs including NCRNA-00051 or 00176 and 00107 are also expressed higher in the prefrontal cortex of individuals of alcohol dependence compared with healthy controls. These lncRNAs may be related to the dysfunction of splicing factors by regulating some post-transcriptional processes [19]. There are more than a few lncRNAs, which are significantly decreased in individuals with alcohol dependence [20].
Furthermore, an accumulating body of research has also emphasized the stimulating views of circRNAs as diagnostic markers for alcohol dependence. Expression of serum hsa-circ-0002130 or 0004771 in patients with alcohol dependence are considerably higher than those of healthy control individuals. Therefore, the hsa-circ-0004771 could be a susceptible diagnostic biomarker. Mechanistically, differentially expressed circRNAs may interact with several alcohol dependence -related miRNAs, which could affect inflammatory pathways [21]. Interestingly, decreased expression of circRNA-406742 can be found in patients with alcohol dependence, which is negatively correlated with miR-1200 expression [21]. Besides, the circRNA-000480 and/or 104942 are highly enriched in patients with alcohol dependence and/or psychiatric disorders [22]. CircRNAs may function as a sponge of miRNAs to affect neural function through the regulation of target genes.
In these ways, a substantial body of research conducted by evolving biotechnologies has highlighted the crucial roles of several ncRNAs in the pathophysiology of mental diseases and alcohol dependence [7,23]. About 10 % of miRNAs are downregulated in alcoholism, including miR-126, miR-153, miR-432, and miR-567 [23]. Unlike the relatively stable genetic code, this combinatorial ncRNAs epigenetic code may be vigorously reprogrammed as a cause or consequence of psychiatric disorders and/or alcohol dependence [24]. From normal development and physiology to the regulation of diseases including alcoholism and/or several psychiatric disorders, some ncRNA molecules have been discovered to mediate diverse processes in CNS [5,25]. For example, ncRNAs may be developed as therapeutic agents to protect the blood-brain barrier for CNS damage patients [25].
3. Alcohol Dependence and m6A Modification of RNAs
Alterations of epitranscriptome could authenticate their investigation as an imperative modulator. In general, the epitranscriptome encompasses all post-transcriptional modifications that occur on RNAs. The most prevalent modification is methylation of N6 adenosine (m6A) that occurs on specific sequence contexts of RNAs [26], which can change the function and/or regulation of their RNA targets. Environmental factors including anxiety, stress and/or social pressure as bad as chronic alcohol usage could lead to alcohol dependence through epigenetic regulation and/or remodeling of chromatin. Environmental effects could also lead to epigenetic modifications at post-transcriptional levels. The m6A is the most well known modification of eukaryotic RNAs, which could regulate transcript splicing, stability, translation, and ncRNA binding. [27]. Chronic ethanol exposure may alter levels of some ncRNAs methylation as well as certain mRNA methylation and their expression levels, suggesting a potential mechanism of epitranscriptome by which chronic alcohol usage could remodel the expression of alcohol responsive genes in the brain, therefore enhancing the risk of development of alcohol dependence [28]. Strikingly, in human postmortem amygdala of early onset alcohol dependence, brain-derived neurotrophic factor (BDNF)-antisense lncRNA is hypomethylated leading to decreased expression levels of BNDF [29]. In the context of alcohol intoxication, the up-regulation of the lncRNA could ameliorate BDNF expression, in which BDNF-AS seems to be regulated by diminished levels of m6A [29]. BDNF belongs to the neurotrophin family with familiar roles in neural development and/or synaptic plasticity. Therefore, lncRNAs may play imperial roles in the control of BDNF expression [30].
The m6A is a pervasive mRNA modification in eukaryotic cells, which arises from the action of methyltransferases, methylation-binding proteins, and demethylases. m6A methylation of RNAs is related with various neurological disorders including depression, epilepsy, Parkinson's disease, Alzheimer's disease, brain injury, and brain gliomas. Therefore, m6A-related drugs have appealed great concerns in the therapeutic fields of neurological disorders [31]. A number of the signaling pathways in brain have been realized to be mediated through m6A modification, but only a few investigations have directly explored the effects of m6A on depression and/or depressive-like behaviors. Depression is a frequent psychiatric disorder described by continued low mood which may be associated with m6A methylation. Therefore, the m6A related molecules including METTL3, METTL14, ALKBH5 and WTAP are associated with major depression [32]. In addition, the gene expression level of m6A controllers in depressive-like behaviors [33]. Interestingly, it has been reported that regulation of m6A is compromised in patients of major depressive disorder following glucocorticoid receptor stimulation [34].
Related proteins of m6A modification could play key roles in the progress of several neuropsychiatric disorders including depression, Parkinson’s disease, and Alzheimer’s disease. The m6A modification regulation mechanism in CNS during the development of neuropsychiatric disorders may provide some insight into new research targets and treatment directions [35]. Similarly, the disturbance of m6A modification may be one of the most important causes for the atypical function of CNS leading to the occurrence of CNS diseases including depression [36]. Serious drinking may lead to neuronal atrophy associated with increased risk for anxiety, depression, cognitive deficits, and altered regulation over drinking behaviors [37]. In addition, chronic stress, anxiety, and depression may be key risk factors for developing alcohol dependence [38]. In fact, depression is often comorbid with alcohol dependence with severe stress components [39]. (Figure 1)
4. Individual Epigenetic Mechanisms for Alcohol Dependency
There is a huge body of evidence exhibiting that alcohol could alter gene expression through epigenetic processes [40]. Epigenetic mechanisms, such as acetylation of the N-terminal tails of histones that pack up DNAs to nucleosomal remodeling, could cause transcriptional change in addiction, which may recompense related genes in specific bain regions contributing to the helpful phenotype such as alcohol tolerance. Studies that exploit alcohol withdrawal to bring depressive-like behaviors have assumed different ways and intervals of alcohol exposure and withdrawal. However, a relationship might exist between individual sensitivity to the aversive properties of ethanol and risk for alcohol dependency. An important confusing factor to deliberate is that the molecular changes induced by alcohol consumption itself and withdrawal from habitual alcohol use may not be related to the depressive-like behavior. Therefore, it might be important to establish a causal role for specific epigenetic mechanisms and alterations of gene expression induced by alcohol explicitly in depressive-like condition [41]. Epigenetic mechanisms may also play an imperative role in depression [42]. Analytical methods of genome-wide DNA methylation and histones modification profile have delivered respected information to establish the functional role of histone-modification could indicate on specific genes [43]. Alcohol dependency may actively lead to relaxed chromatin because of the downregulation of DNA/histone methylation. Otherwise, chronic exposure might in part lead to close-fitting chromatin bundle. Consequently, alcohol drinking may affect epigenetic mechanisms responsible for adaptation alterations of several brain paths probably linked to stress management [44]. After withdrawal, chromatin may tend to the condensed state via the upregulation of DNA and/or histones [45]. For example, mRNA expression levels are significantly lower compared to controls, which correspond to alterations in DNA methylation in a rodent model [46]. Therefore, DNA methylation might be a target for pharmacological interventions for alcohol dependency [46]. In addition, DNA methylation could be a good biomarker of alcohol consumption [47]. Exposure to ethanol during adolescence might upregulate DNA methyltransferase activity, which can induce hypermethylation of various genes such as coding for neuropeptide Y (NPY) and BDNF [48]. Furthermore, prenatal exposure to alcohol may generally trigger epigenetic modifications depending on the development stage, which may contain increased histone acetylation and/or reduced DNA/histone methylation [49]. In this regards, alcohol withdrawal may lead to dysregulated histone acetylation via the increased expression of histone deacetylase (HDAC) in some brain areas [50]. Hence, the treatment with HDAC inhibitors can amend negative emotional conditions brought by alcohol withdrawal [50]. However, histone acetylation in the brain of depression-like behavior through withdrawal after alcohol exposure may require further intensive examination.
5. A Relationship between Gut Microbiota and Alcohol Dependency
Gut microbiota have various effects on host physiology, including host metabolism, the development of immune system, and even behaviors [51]. The intricate interaction among the gut, stress, and eating/drinking behavior may simplify new therapeutic targets for stress-related psychiatric disorders [51]. Remarkably, short-chain fatty acids (SCFAs), explicitly acetate, propionate, and butyrate, might be interaction-mediators of microbiota-gut-brain axis on the stress response and/or eating/drinking behavior. In fact, various metabolites from the gut microbiome including SCFAs have been proved to regulate the histone acetylation process [52]. The microbiota-gut-brain axis may be a bidirectional route of homeostatic communication via epigenetic mechanisms of diverse metabolites such as SCFAs. Thus, a modulation of gut microbiota via diet or lifestyle can regulate neuron/brain inflammation via certain epigenetic mechanisms [53], which might be effective for emotional status and/or depressive disorders [54]. As important constituents of epigenetics by gut microbial metabolites and/or fermentation products, several miRNAs with epigenetic mechanisms have vital roles in various physiological homeostasis mechanisms [55,56]. For example, microbial acetate and/or butyrate might alleviate obesity with the regulation of host miRNAs [57]. In addition, there were close intricate interactions among gut microbiota, inflammation and differential miRNAs suggesting that ncRNA may possess a potential role in the protection of host against life-related diseases such as atherosclerosis [58]. Interestingly, it has been shown that circRNAs and the gut microbiome can interact to influence the growth of cancer cells [59]. Similarly, expression of lncRNAs could be repressed by gut microbiota [60]. Increasing data suggest that regulatory ncRNAs including miRNAs, circRNAs, and lncRNAs may influence host-microbe interactions showing as potential biomarkers in microbiome-associated disorders including diabetes and cancers [61]. (Figure 2)
In addition, the gut microbiota has an effect on host m6A mRNA modifications, which is another demonstration of the interaction between gut commensal bacteria and their hosts [62]. As well, the host m6A modification can also influence the gut microbiome by provoking gut inflammatory responses [63]. Possibly, Lactobacillus plantarum and/or Akkermansia muciniphila can influence the specific m6A modifications, which might emphasize epitranscriptomic modifications as a communication between gut commensal bacteria and host [62,64]. The presence of certain gut microbiome may also account for the considerably elevated m6A levels in the intestine [65]. Thus, m6A methylation is indeed implicated in the host-gut microbiota crosstalk. Actually, substantial studies have suggested that the enteric microbiome is a key mediator of m6A modification. In general, a number of ncRNAs and/or m6A modification have been associated in the beginning and progression of drug addiction [66,67]. Therefore, the gut-brain axis may be the key to the homeostasis of CNS, which may regulate several neuro-behaviors [68]. The gut microbiome could also affect drug bioavailability, blood-brain barrier (BBB) permeability, and social behaviors [69]. Emerging microbiota-based interventions such as prebiotics, probiotics, FMT, or metabolites supplementation might advocate an exciting tactic to treating psychiatric disorders probably including alcohol dependency.
6. A Possible Tactic with Alteration of Gut Microbiota against Alcohol Dependency
Too much alcohol consumption may induce gut dysbiosis, an imbalance in gut microbiota, through numerous mechanisms. Consequently, chronic alcohol exposure can reduce the creation of mucus and several peptides, which may intrude the intestinal barrier [70]. Although pharmacological treatments are existent, their effectiveness depends on appropriate faithfulness to the prescribed regimen [71]. Therefore, most patients with alcohol dependency are left untreated, and there is a need for additional, more effective therapies. Identifying some biological markers predicting susceptibility to develop extreme alcohol consumption may lead to an enhancement of good clinical care. Interestingly, relationships between gut microbiota and behavioral characters that individualize alcohol dependency has been described [72]. Certain microbiota composition is linked to addiction actions in a realistic model of alcohol dependency [72]. Based on this findings, newfangled therapeutic regimens should embrace gut microbiome manipulation, which may lessen alcohol intake and/or drinking activities. Indeed, alcohol consumption produces both direct and/or indirect consequences on the gut microbiota through metabolism, neuronal response, and immune inflammatory cascades. In particular, chronic inflammatory condition may lead to alterations in several inflammatory mediators that can activate nuclear factor kappa B (NF-kB) signaling pathway leading to neuronal damage/apoptosis in glial and/or neuronal cells [73]. It is imperative to note that not all patients with alcohol dependency have dysbiosis and/or increased gut epithelial disrupton [74]. However, several effects of alcohol on the gut microbiome might contribute to the increased alcohol consumption. Therefore, use of probiotics, prebiotics, or FMT may deserve further investigation as therapeutic tactics for alcohol dependency [74]. At present, however, the application of FMT as a beneficial therapeutic approach is yet in the investigatory stages [74]. In addition, donor-to-recipient disease of stool transfer may be a great fear of the FMT. Moreover, it still requires to be determined what bad effect of FMT has on the gut microbiota and/or to the brain in the long run. [75]. (Figure 2)
Interestingly, an antidepressant arketamine (also termed R-ketamine) can renovate the altered composition of gut microbiota within patients of depression-like behaviors in rodents, contributing to valuable effects of the R-ketamine. Ketamine is a racemic mix composed of two enantiomers, R-ketamine and esketamine (S-ketamine). Importantly, both enantiomers have shown antidepressant effects, whose effects are attributed to distinct pharmacological activities including NMDA-channel and/or opioid receptor. It has been shown that antidepressant-like effects of both ketamines might be in part mediated by alteration of gut microbiota [76]. Ketamine could potentially activate several biochemical signaling pathways which may eventually lead to inhibitory phosphorylation of GSK3β molecule in microglia [77]. Remarkably, S-ketamine exerts the neuroprotective effects via enhancing autophagy lessening oxidative stress, whose mechanism comprises AMPK/mTOR-dependent autophagy and/or antioxidant system [78]. Amazingly, the S-ketamine considerably could change the abundance of gut microbiota including Adlercreutzia equolifaciens and Akkermansia muciniphila [79]. It has been revealed that the regulation of NFAT signaling by miR-149 might play a key role in tenacious prophylactic effects of R-ketamine in inflammation, and that gut microbiota can control the gene expression of miRNAs via the gut-brain axis [80]. Increased miR-149 expression may be related to the reduced glial cell numbers in patients diagnosed with familial bipolar disorder [81]. Because miR-149 has been revealed to inhibit glial proliferation, increased miR-149 expression is also consistent with the pathology of depressive disorder [82]. Still, there is rarely information presenting the role of other miRNAs or m6A modification in the prophylactic effects of ketamine and its enantiomers in brain neuroinflammation disorders.
7. Future Perspectives
Would you like to use ketamine for improving alcohol dependency? Come to think of it, there might be a dangerous choice. In other words, it would be difficult to answer, which is better and/or safer either ketamine abuse or alcohol abuse. Based on a hypothesis if the ketamine may improve alcohol dependency via the mechanism of improving autophagy in neuronal cells, dietary intervention would be possible for the treatment of alcohol dependency. Because, the modification of gut microbiome is safely imaginable via a diet, which could also contribute to the alteration of ncRNAs production and/or m6A modification in various cells [83,84]. In fact, some dietary supplements are dynamically performing through different mechanisms to reduce alcohol relapse [85]. As for prebiotics and/or probiotics, those interventions may be somewhat inadequate for a treatment in regard to the improvement by autophagy [86,87]. Some additional factors and/or signaling activation might be required for the superior dietary intervention even against alcohol dependency. In addition to modulating the gut microbiome, for example, metformin could exert its positive effect by affecting mitochondrial function and restoring of redox balance [88]. In addition, several elements involved in the tryptophan and kynurenine pathway may also be plausible, which has been presented to be linked to various immune-related diseases including major depressive and bipolar disorders [89]. Formed by recurring inflammatory conditions, an “engram” might devote to a gentle development of these diseases [89]. Engram memory system in brain might retain the knowledge of a certain inflammation in a body which would be involved in the pathogenesis of immune-related diseases, in which the immunity-linked routes might be also related with the neuronal responses to memory engrams [90]. Clearance of the bad memory “engrams” could be promising for the prevention and/or treatment against an immune-related disease even as well a cancer, a cardiac arrhythmia, and/or a neurodegenerative disease [83,91,92]. (Figure 3) If that is the case of alcohol dependence, a certain adjustment of “engram” with the alteration of gut microbiota might be helpful for a notable treatment tactic against alcohol dependence. Future work should precisely explain at the molecular level how this engram pathway could interfere to progress the alcohol dependency. (Figure 3)
8. Conclusion
Several ncRNAs and/or m6A modification might be involved in the instigation of alcohol dependency, in which the relationship between gut and brain can play an important role. Also, the correlation between brain and immunity might also influence the development of alcohol dependency. An in-depth knowledge of roles of ncRNAs and m6A in gut microbiome may be valuable for the development of a novel treatment against alcohol dependence.
Abbreviations
| BBB | blood-brain barrier |
| BDNF | brain-derived neurotrophic growth factor |
| CNS | central nervous system |
| circRNA | circular RNA |
| FMT | fecal microbiota transplantation |
| HDAC | histone deacetylase |
| lncRNAs | long non-coding RNAs |
| mRNA | messenger RNA |
| m6A | methylation of N6 adenosine |
| ncRNA | non-coding RNA |
| NF-kB | nuclear factor kappa B |
| NPY | neuropeptide Y |
| piRNAs | piwi interacting RNAs |
| siRNAs | small interfering RNAs |
| ROS | reactive oxygen species |
| SCFAs | short-chain fatty acids |
| siRNA | short interference RNA |
| R-ketamine | arketamine |
| S-ketamine | esketamine |
Author Contributions
Conceptualization, MN, NS, YI, and SM; original draft preparation and editing, MN, NS, SY, YI, and SM; visualization, MN, NS, YI and SM; supervision, SM. Each author (MN, NS, SY, YI, and SM) has participated satisfactorily in this work of drafting the article and/or revising the article for the important rational content. Then, all authors provided final approval of the version to be submitted. Lastly, all authors have read and approved to the published version of the manuscript.
Funding
This research presented here received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Conflicts of Interest
The authors declare that they have no competing financial interests.
References
- Ducci, F.; Goldman, D. Genetic approaches to addiction: genes and alcohol. Addiction. 2008, 103, 1414–1428. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, AF.; Heilig, M.; Perez, A.; Probst, C.; Rehm, J. Alcohol use disorders. The Lancet. 2019, 394, 781–792. [Google Scholar] [CrossRef] [PubMed]
- Rehm, J.; Shield, KD. Global burden of disease and the impact of mental and addictive disorders. Current psychiatry reports. 2019, 21, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Hasin, DS.; Wall, M.; Witkiewitz, K.; Kranzler, HR.; Falk, D.; Litten, R.; et al. Change in non-abstinent WHO drinking risk levels and alcohol dependence: a 3 year follow-up study in the US general population. The Lancet Psychiatry. 2017, 4, 469–476. [Google Scholar] [CrossRef] [PubMed]
- Mayfield, RD. Emerging roles for ncRNAs in alcohol use disorders. Alcohol. 2017, 60, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Slack, FJ.; Chinnaiyan, AM. The role of non-coding RNAs in oncology. Cell. 2019, 179, 1033–1055. [Google Scholar] [CrossRef] [PubMed]
- Wu, YY.; Kuo, HC. Functional roles and networks of non-coding RNAs in the pathogenesis of neurodegenerative diseases. Journal of Biomedical Science. 2020, 27, 1–23. [Google Scholar] [CrossRef]
- Anastasiadou, E.; Jacob, LS.; Slack, FJ. Non-coding RNA networks in cancer. Nature Reviews Cancer. 2018, 18, 5–18. [Google Scholar] [CrossRef]
- Zhang, P.; Wu, W.; Chen, Q.; Chen, M. Non-coding RNAs and their Integrated Networks. Journal of integrative bioinformatics. 2019, 16, 20190027. [Google Scholar] [CrossRef]
- Wang, JQ.; Liu, YR.; Xia, QR.; Liang, J.; Wang, JL.; Li, J. Functional roles, regulatory mechanisms and theranostics applications of ncRNAs in alcohol use disorder. Int J Biol Sci. 2023, 19, 1316–1335. [Google Scholar] [CrossRef]
- Wei, JW.; Huang, K.; Yang, C.; Kang, CS. Non-coding RNAs as regulators in epigenetics (Review). Oncol Rep. 2017, 37, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Berkel, TD.; Pandey, SC. Emerging Role of Epigenetic Mechanisms in Alcohol Addiction. Alcohol Clin. Exp. Res. 2017, 41, 666–680. [Google Scholar] [CrossRef] [PubMed]
- Dai, Y.; Hu, R.; Pei, G.; Zhang, H.; Zhao, Z.; Jia, P. Diverse types of genomic evidence converge on alcohol use disorder risk genes. J. Med. Genet. 2020, 57, 733–743. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Wang, F.; Rosato, AJ.; Farrer, LA.; Henderson, DC.; Zhang, H. Prefrontal cortex eQTLs/mQTLs enriched in genetic variants associated with alcohol use disorder and other diseases. Epigenomics. 2020, 12, 789–800. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Zhang, Y.; Zhang, Y.; Wang, Q.; Miao, Q.; Xu, Y.; et al. Correlation between the epigenetic modification of histone H3K9 acetylation of NR2B gene promoter in rat hippocampus and ethanol withdrawal syndrome. Mol. Biol. Rep. 2019, 46, 2867–2875. [Google Scholar] [CrossRef] [PubMed]
- Santos-Bezerra, DP.; Cavaleiro, AM.; Santos, AS.; Suemoto, CK.; Pasqualucci, CA.; Jacob-Filho, W.; et al. Alcohol Use Disorder is Associated with Upregulation of MicroRNA-34a and MicroRNA-34c in Hippocampal Postmortem Tissue. Alcoholism: Clinical and Experimental Research. 2021; 45, 64–68. [Google Scholar]
- Karabegović, I.; Abozaid, Y.; Maas, SCE.; Labrecque, J.; Bos, D.; De, Knegt, RJ.; et al. Plasma microRNA signature of alcohol consumption: the Rotterdam Study. J Nutr. 2023, 152, 2677–2688.
- Pandey, SC. A critical role of brain-derived neurotrophic factor in alcohol consumption. Biological psychiatry. 2016, 79, 427–429. [Google Scholar] [CrossRef] [PubMed]
- Van, Booven, D.; Mengying, Li.; Sunil, Rao, J.; Blokhin, IO.; Dayne, Mayfield, R.; Barbier, E.; et al. Alcohol use disorder causes global changes in splicing in the human brain. Translational psychiatry. 2021, 11, 2. [CrossRef]
- Farris, SP.; Mayfield, RD. RNA-Seq reveals novel transcriptional reorganization in human alcoholic brain. International review of neurobiology. 2014, 116, 275–300. [Google Scholar]
- Liu, Y.; Li, J.; Bu, H.; Wang, H.; Zhang, Y.; Shen, Q.; et al. Circular RNA expression alteration identifies a novel circulating biomarker in serum exosomal for detection of alcohol dependence. Addiction Biology. 2021, 26, e13031. [Google Scholar] [CrossRef]
- Vornholt, E.; Drake, J.; Mamdani, M.; McMichael, G.; Taylor, ZN.; Bacanu, SA.; et al. Identifying a novel biological mechanism for alcohol addiction associated with circRNA networks acting as potential miRNA sponges. Addiction biology. 2021, 26, e13071. [Google Scholar] [CrossRef] [PubMed]
- Mohebbati, R.; Sadeghnia, HR. The Role of microRNAs in Alcoholism: A Meta-analytic Review. Curr Pharm Des. 2022, 28, 1926–1931. [Google Scholar] [CrossRef] [PubMed]
- Jung, Y.; Goldman, D. Role of RNA modifications in brain and behavior. Genes Brain Behav. 2018, 17, e12444. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Bai, W.; Sun, L.; Lin, Y.; Tian, M. Targeting Non-Coding RNA for CNS Injuries: Regulation of Blood-Brain Barrier Functions. Neurochem Res. 2023, 48, 1997–2016. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Yue, Y.; Han, D.; Wang, X.; Fu, Y.; Zhang, L.; et al. A METTL3-METTL14 complex mediates mammalian nuclear RNA N6-adenosine methylation. Nat. Chem. Biol. 2014, 10, 93–95. [Google Scholar] [CrossRef]
- Enge,l M; Eggert, C.; Kaplick, PM.; Eder, M.; Röh, S.; Tietze, L.; et al. The role of m(6)A/m-RNA methylation in stress Response regulation. Neuron 2018, 99, 389–403. e9. [CrossRef] [PubMed]
- Liu, Y.; Koo, JS.; Zhang, H. Chronic intermittent ethanol exposure-induced m6A modifications around mRNA stop codons of opioid receptor genes. Epigenetics. 2024, 19, 2294515. [Google Scholar] [CrossRef]
- Bohnsack, JP.; Teppen, T.; Kyzar, EJ.; Dzitoyeva, S.; Pandey, SC. The lncRNA BDNF-AS is an epigenetic regulator in the human amygdala in early onset alcohol use disorders. Transl Psychiatry. 2019, 9, 34. [Google Scholar] [CrossRef]
- Khani-Habibabadi, F.; Zare, L.; Sahraian, MA.; Javan, M.; Behmanesh, M. Hotair and Malat1 Long Noncoding RNAs Regulate Bdnf Expression and Oligodendrocyte Precursor Cell Differentiation. Mol Neurobiol. 2022, 59, 4209–4222. [Google Scholar] [CrossRef]
- Lv, J.; Xing, L.; Zhong, X.; Li, K.; Liu, M.; Du, K. Role of N6-methyladenosine modification in central nervous system diseases and related therapeutic agents. Biomed Pharmacother. 2023, 162, 114583. [Google Scholar] [CrossRef]
- Du, T.; Rao, S.; Wu, L.; Ye, N.; Liu, Z.; Hu, H.; et al. An association study of the m6A genes with major depressive disorder in Chinese Han population. J. Affect Disord. 2015, 183, 279–286. [Google Scholar] [CrossRef] [PubMed]
- Joshi, K.; Wang, DO.; Gururajan, A. The m6A-methylome in major depression: A bioinformatic analysis of publicly available datasets. Psychiatry Res. Commun. 2022, 2, 100089. [Google Scholar] [CrossRef]
- Engel, M.; Eggert, C.; Kaplick, PM.; Eder, M.; Röh, S.; Tietze, L.; et al. The Role of m6A/m-RNA Methylation in Stress Response Regulation. Neuron. 2018; 99, 389–403.e9. [Google Scholar]
- Lei, C.; Wang, Q. The Progression of N6-methyladenosine Study and Its Role in Neuropsychiatric Disorders. Int J Mol Sci. 2022, 23, 5922. [Google Scholar] [CrossRef] [PubMed]
- Han, M.; Liu, Z.; Xu, Y.; Liu, X.; Wang, D.; Li, F.; et al. Abnormality of m6A mRNA methylation is involved in Alzheimer’s disease. Front. Neurosci. 2020, 14, 98. [Google Scholar] [CrossRef] [PubMed]
- Daviet, R.; Aydogan, G.; Jagannathan, K.; Spilka, N.; Koellinger, PD.; Koellinger, PD.; et al. Associations between alcohol consumption and gray and white matter volumes in the UK biobank. Nat. Commun. 2022, 13, 1175. [Google Scholar] [CrossRef] [PubMed]
- McCaul, ME.; Hutton, HE.; Stephens, MA.; Xu, X.; Wand, GS. Anxiety, anxiety sensitivity, and perceived stress as predictors of recent drinking, alcohol craving, and social stress response in heavy drinkers. Alcohol. Clin. Exp. Res. 2017, 41, 836–845. [Google Scholar] [CrossRef] [PubMed]
- Boden, JM.; Fergusson, DM. Alcohol and depression. Addiction. 2011, 106, 906–914. [Google Scholar] [CrossRef] [PubMed]
- Ajoolabady, A.; Aslkhodapasandhokmabad, H.; Zhou, Y.; Ren, J. Epigenetic modification in alcohol-related liver diseases. Med Res Rev. 2022, 42, 1463–1491. [Google Scholar] [CrossRef]
- Martins de Carvalho, L.; Chen, WY.; Lasek, AW. Epigenetic mechanisms underlying stress-induced depression. Int Rev Neurobiol. 2021, 156, 87–126. [Google Scholar]
- Misztak, P.; Panczyszyn-Trzewik, P.; Sowa-Kucma, M. Histone deacetylases (HDACs) as therapeutic target for depressive disorders. Pharmacol Rep, 2018; 70, 398–408. [Google Scholar]
- Boers, R.; Boers, J.; de Hoon, B.; Kockx, C.; Ozgur, Z.; Molijn, A.; et al. Genome-wide DNA methylation profiling using the methylation-dependent restriction enzyme LpnPI. Genome Res. 2018, 28, 88–99. [Google Scholar] [CrossRef]
- Palmisano, M.; Pandey, SC. Epigenetic mechanisms of alcoholism and stress-related disorders. Alcohol. 2017, 60, 7–18. [Google Scholar] [CrossRef] [PubMed]
- Ciafrè, S.; Carito, V.; Ferraguti, G.; Greco, A.; Chaldakov, GN.; Fiore, M. et al. How alcohol drinking affects our genes: An epigenetic point of view. Biochem. Cell Biol. 2019; 97, 345–356. [Google Scholar]
- Maier, HB.; Neyazi, M.; Neyazi, A.; Hillemacher, T.; Pathak, H.; Rhein, M.; et al. Alcohol consumption alters Gdnf promoter methylation and expression in rats. J Psychiatr Res. 2020, 121, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Mahna, D.; Puri, S.; Sharma, S. DNA methylation signatures: Biomarkers of drug and alcohol abuse. Mutat. Res. 2018, 777, 19–28. [Google Scholar] [CrossRef]
- Berkel, TDM; Zhang, H.; Teppen, T.; Sakharkar, AJ.; Pandey, SC. Essential role of histone methyltransferase G9a in rapid tolerance to the anxiolytic effects of ethanol. Int. J. Neuropsychopharmacol. 2019, 22, 292–302. [CrossRef] [PubMed]
- Jarmasz, JS.; Stirton, H.; Basalah, D.; Davie, JR.; Clarren, SK.; Astley, SJ.; et al. Global DNA methylation and histone posttranslational modifications in human and nonhuman primate brain in association with prenatal alcohol exposure. Alcohol. Clin. Exp. Res. 2019, 43, 1145–1162. [Google Scholar] [CrossRef] [PubMed]
- Chen, WY.; Zhang, H.; Gatta, E.; Glover, EJ.; Pandey, SC.; Lasek, AW. The histone deacetylase inhibitor suberoylanilide hydroxamic acid (SAHA) alleviates depression-like behavior and normalizes epigenetic changes in the hippocampus during ethanol withdrawal. Alcohol. 2019, 78, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Quagebeur, R.; Dalile, B.; Raes, J.; Van Oudenhove, L.; Verbeke, K.; Vrieze, E. The role of short-chain fatty acids (SCFAs) in regulating stress responses, eating behavior, and nutritional state in anorexia nervosa: protocol for a randomized controlled trial. J Eat Disord. 2023, 11, 191. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, A.; Chachra, P.; Kennedy, P.; Pena, CJ.; Desouza, LA.; Nestler, EJ.; et al. Hippocampal HDAC4 contributes to postnatal fluoxetine-evoked depression-like behavior. Neuropsychopharmacology, 2014; 39, 2221–2232. [Google Scholar]
- Ardizzone, A.; Capra, AP.; Repici, A.; Lanza, M.; Bova, V.; Palermo, N.; et al. Rebalancing NOX2/Nrf2 to limit inflammation and oxidative stress across gut-brain axis in migraine. Free Radic Biol Med. 2024, 213, 65–78. [Google Scholar] [CrossRef] [PubMed]
- Zalar, B.; Haslberger, A.; Peterlin, B. The Role of Microbiota in Depression - a brief review. Psychiatr Danub. 2018, 30, 136–141. [Google Scholar] [CrossRef]
- Woo, V.; Alenghat, T. Host–microbiota interactions: epigenomic regulation. Curr. Opin. Immunol. 2017, 44, 52–60. [Google Scholar] [CrossRef]
- Arner, P.; Kulyté, A. MicroRNA regulatory networks in human adipose tissue and obesity. Nat Rev Endocrinol. 2015, 11, 276. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Zhang, P.; Luo, J.; Shen, L.; Zhang, S.; Gu, H.; et al. Dietary betaine prevents obesity through gut microbiota-drived microRNA-378a family. Gut Microbes. 2021, 13, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Yang, L.; Liu, Y.; Yan, R.; Wang, R.; Zhang, P.; et al. Butyrate suppresses atherosclerotic inflammation by regulating macrophages and polarization via GPR43/HDAC-miRNAs axis in ApoE-/- mice. PLoS One. 2023, 18, e0282685. [Google Scholar] [CrossRef] [PubMed]
- Abdullah, ST.; Abdullah, SR.; Hussen, BM.; Younis, YM.; Rasul, MF.; Taheri, M. Role of circular RNAs and gut microbiome in gastrointestinal cancers and therapeutic targets. Noncoding RNA Res. 2023, 9, 236–252. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, M.; Chen, J.; Li, Y.; Kuang, Z.; Dende, C.; et al. The gut microbiota reprograms intestinal lipid metabolism through long noncoding RNA Snhg9. Science. 2023, 381, 851–857. [Google Scholar] [CrossRef] [PubMed]
- Fardi, F.; Khasraghi, LB.; Shahbakhti, N.; Salami Naseriyan, A.; Najafi, S.; Sanaaee, S.; et al. An interplay between non-coding RNAs and gut microbiota in human health. Diabetes Res Clin Pract. 2023, 201, 110739. [Google Scholar] [CrossRef] [PubMed]
- Jabs, S.; Biton, A.; Bécavin, C.; Nahori, MA.; Ghozlane, A.; Pagliuso, A.; et al. Impact of the gut microbiota on the m6A epitranscriptome of mouse cecum and liver. Nat Commun. 2020, 11, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.; Zhang, Z.; Xue, M.; Zhao, BS.; Harder, O.; Li, A.; et al. N6-methyladenosine modification enables viral RNA to escape recognition by RNA sensor RIG-I. Nat. Microbiol. 2020, 5, 584–598. [Google Scholar] [CrossRef] [PubMed]
- Kim, HJ.; Jeon, HJ.; Kim, JY.; Shim, JJ.; Lee, JH. Lactiplantibacillus plantarum HY7718 Improves Intestinal Integrity in a DSS-Induced Ulcerative Colitis Mouse Model by Suppressing Inflammation through Modulation of the Gut Microbiota. Int J Mol Sci. 2024, 25, 575. [Google Scholar] [CrossRef]
- Wang, X.; Li, Y.; Chen, W.; Shi, H.; Eren, AM.; Morozov, A.; et al. Transcriptome-wide reprogramming of N(6)-methyladenosine modification by the mouse microbiome. Cell Res. 2019, 29, 167–170. [Google Scholar] [CrossRef]
- Zhao, Y.; Qin, F.; Han, S.; Li, S.; Zhao, Y.; Wang, H.; et al. MicroRNAs in drug addiction: Current status and future perspectives. Pharmacology & Therapeutics. 2022; 236, 108215. [Google Scholar]
- Liu, Y.; Zhang, H. RNA m6A Modification Changes in Postmortem Nucleus Accumbens of Subjects with Alcohol Use Disorder: A Pilot Study. Genes (Basel). 2022, 13, 958. [Google Scholar] [CrossRef] [PubMed]
- Huang, P.; Liu, M.; Zhang, J.; Zhong, X.; Zhong, C. YTHDF1 Attenuates TBI-Induced Brain-Gut Axis Dysfunction in Mice. Int J Mol Sci. 2023, 24, 4240. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, CL.; Doifode, T.; Rezende, VL.; Costa, MA.; Rhoads, JM.; Soutullo, CA. The many faces of microbiota-gut-brain axis in autism spectrum disorder. Life Sci. 2024, 337, 122357. [Google Scholar] [CrossRef] [PubMed]
- Fuenzalida, C.; Dufeu, MS.; Poniachik, J.; Roblero, JP.; Valenzuela-Pérez, L.; Beltrán, CJ. Probiotics-based treatment as an integral approach for alcohol use disorder in alcoholic liver disease. Front Pharmacol. 2021, 12, 729950. [Google Scholar] [CrossRef] [PubMed]
- Walker, JR.; Korte, JE.; McRae-Clark, AL.; Hartwell, KJ. Adherence across FDA-approved medications for alcohol use disorder in a Veterans Administration population. J Stud Alcohol Drugs. 2019, 80, 572–577. [Google Scholar] [CrossRef] [PubMed]
- Jadhav, KS.; Peterson, VL.; Halfon, O.; Ahern, G.; Fouhy, F.; Stanton, C.; et al. Gut microbiome correlates with altered striatal dopamine receptor expression in a model of compulsive alcohol seeking. Neuropharmacology. 2018, 141, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Flores-Bastías, O.; Karahanian, E. Neuroinflammation produced by heavy alcohol intake is due to loops of interactions between Toll-like 4 and TNF receptors, peroxisome proliferator-activated receptors and the central melanocortin system: A novel hypothesis and new therapeutic avenues. Neuropharmacology. 2018, 128, 401–407. [Google Scholar] [CrossRef]
- Wolstenholme, JT.; Duong, NK.; Brocato, ER.; Bajaj, JS. Gut-Liver-Brain Axis and Alcohol Use Disorder: Treatment Potential of Fecal Microbiota Transplantation. Alcohol Res. 2024, 44, 01. [Google Scholar] [CrossRef]
- Meng, Y.; Sun, J.; Zhang, G. Pick fecal microbiota transplantation to enhance therapy for major depressive disorder. Prog Neuropsychopharmacol Biol Psychiatry. 2024, 128, 110860. [Google Scholar] [CrossRef]
- Hashimoto, K. Neuroinflammation through the vagus nerve-dependent gut-microbiota-brain axis in treatment-resistant depression. Prog Brain Res. 2023, 278, 61–77. [Google Scholar]
- Kalkman, HO. Activation of σ1-Receptors by R-Ketamine May Enhance the Antidepressant Effect of S-Ketamine. Biomedicines. 2023, 11, 2664. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Liu, Y.; Zhou, H.; Lu, H.; Zhang, Y.; Hua, J.; et al. Esketamine is neuroprotective against traumatic brain injury through its modulation of autophagy and oxidative stress via AMPK/mTOR-dependent TFEB nuclear translocation. Exp Neurol. 2023, 366, 114436. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Ma, W.; Lin, H.; Gu, X.; Xie, H. The effects of esketamine on the intestinal microenvironment and intestinal microbiota in mice. Hum Exp Toxicol. 2023, 42, 9603271231211894. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Wang, L.; Chang, L.; Shan, J.; Qu, Y.; Wang, X.; et al. A role of microRNA-149 in the prefrontal cortex for prophylactic actions of (R)-ketamine in inflammation model. Neuropharmacology. 2022, 219, 109250. [Google Scholar] [CrossRef] [PubMed]
- Delalle, I. MicroRNAs as Candidates for Bipolar Disorder Biomarkers. Psychiatr Danub. 2021, 33, 451–455. [Google Scholar] [PubMed]
- Choi, JL.; Kao, PF.; Itriago, E.; Zhan, Y.; Kozubek, JA.; Hoss, AG.; et al. miR-149 and miR-29c as candidates for bipolar disorder biomarkers. Am J Med Genet B Neuropsychiatr Genet. 2017, 174, 315–323. [Google Scholar] [CrossRef] [PubMed]
- Suga, N.; Ikeda, Y.; Yoshikawa, S.; Taniguchi, K.; Sawamura, H.; Matsuda, S. Non-Coding RNAs and Gut Microbiota in the Pathogenesis of Cardiac Arrhythmias: The Latest Update. Genes. 2023, 14, 1736. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, M.; Suga, N.; Ikeda, Y.; Yoshikawa, S.; Matsuda, S. Circular RNAs, Noncoding RNAs, and N6-methyladenosine Involved in the Development of MAFLD. Non-Coding RNA. 2024, 10, 11. [Google Scholar] [CrossRef] [PubMed]
- Ezquer, F.; Quintanilla, ME.; Morales, P.; Santapau, D.; Munita, JM.; Moya-Flores, F.; et al. A dual treatment blocks alcohol binge-drinking relapse: Microbiota as a new player. Drug Alcohol Depend. 2022, 236, 109466. [Google Scholar] [CrossRef]
- Cooper, TE.; Khalid, R.; Chan, S.; Craig, JC.; Hawley, CM.; Howell, M.; et al. Synbiotics, prebiotics and probiotics for people with chronic kidney disease. Cochrane Database Syst. Rev. 2023, 10, CD013631. [Google Scholar]
- Yacoub, R.; Nadkarni, GN.; McSkimming, DI.; Chaves, LD.; Abyad, S.; Bryniarski, MA.; et al. Fecal microbiota analysis of polycystic kidney disease patients according to renal function: A pilot study. Exp. Biol. Med. 2019, 244, 505–513. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Zhu, YJ.; Zhou, YX.; Ding, J.; Liu, JY. Metformin in therapeutic applications in human diseases: Its mechanism of action and clinical study. Mol. Biomed. 2022, 3, 41. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, A.; Ikeda, Y.; Yoshikawa, S.; Taniguchi, K.; Sawamura, H.; Morikawa, S.; Nakashima, M.; Asai, T.; Matsuda, S. The Tryptophan and Kynurenine Pathway Involved in the Development of Immune-Related Diseases. Int. J. Mol. Sci. 2023, 24, 5742. [Google Scholar] [CrossRef] [PubMed]
- Håvik, B.; Røkke, H.; Dagyte, G.; Stavrum, A.K.; Bramham, C.R.; Steen, V.M. Synaptic activity-induced global gene expression patterns in the dentate gyrus of adult behaving rats: Induction of immunity-linked genes. Neuroscience. 2007, 148, 925–936. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, Y.; Taniguchi, K.; Yoshikawa, S.; Sawamura, H.; Tsuji, A.; Matsuda, S. A budding concept with certain microbiota, anti-proliferative family proteins, and engram theory for the innovative treatment of colon cancer. Explor Med. 2022, 3, 468–478. [Google Scholar] [CrossRef]
- Suga, N.; Ikeda, Y.; Yoshikawa, S.; Taniguchi, K.; Sawamura, H.; Matsuda, S. In Search of a Function for the N6-Methyladenosine in Epitranscriptome, Autophagy and Neurodegenerative Diseases. Neurol. Int. 2023, 15, 967–979. [Google Scholar] [CrossRef]
Figure 1.
Representation for the association of non-coding RNAs (ncRNAs) and m6A modification (m6A+) of ncRNAs to psychiatric disorders and/or alcohol dependence. Firstly, the m6A modification may be controlled by methyltransferases “writers” and demethylases “erasers” by the stimulation of inflammation and/or oxidative stress with reactive oxygen species (ROS). The ncRNAs and m6A-ncRNAs with binding “readers” molecules may contribute to several RNAs activities including sponging, stability, processing, and/or translation of mRNAs, which could be consequently an important process several psychiatric disorders including alcohol dependence.
Figure 1.
Representation for the association of non-coding RNAs (ncRNAs) and m6A modification (m6A+) of ncRNAs to psychiatric disorders and/or alcohol dependence. Firstly, the m6A modification may be controlled by methyltransferases “writers” and demethylases “erasers” by the stimulation of inflammation and/or oxidative stress with reactive oxygen species (ROS). The ncRNAs and m6A-ncRNAs with binding “readers” molecules may contribute to several RNAs activities including sponging, stability, processing, and/or translation of mRNAs, which could be consequently an important process several psychiatric disorders including alcohol dependence.
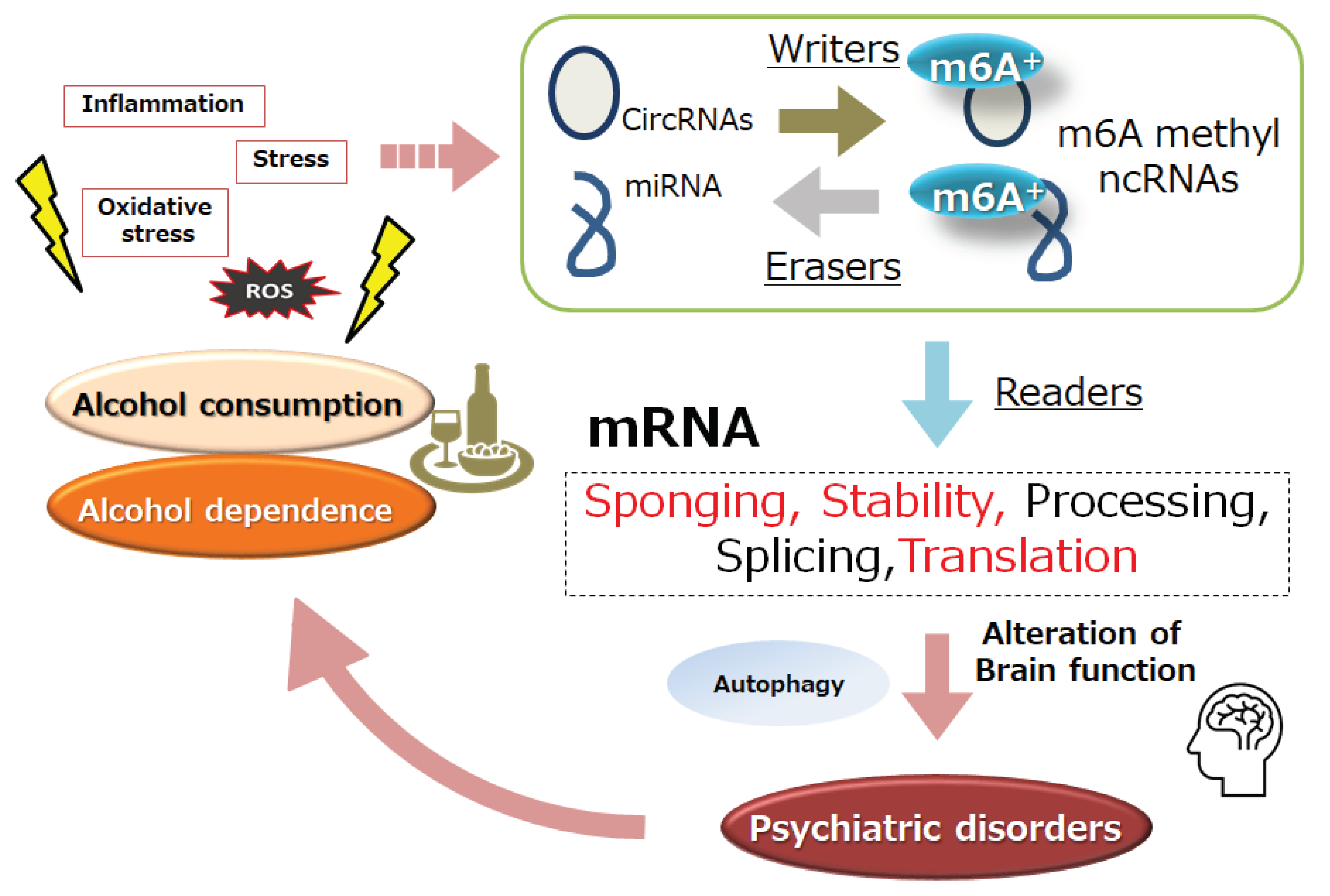
Figure 2.
Schematic demonstration of the potential strategies against the pathology of alcohol dependence. Various kinds of probiotics and/or fecal microbiota transplantation (FMT) may support the alteration of gut microbiota for the modification of autophagy, which might be advantageous for the inhibition of several engram formation, which may consequently improve the pathology of psychiatric disorders including alcohol dependence. Note that some of significant events such as autophagy initiation, ROS production, and inflammatory reactions have been misplaced for clarity.
Figure 2.
Schematic demonstration of the potential strategies against the pathology of alcohol dependence. Various kinds of probiotics and/or fecal microbiota transplantation (FMT) may support the alteration of gut microbiota for the modification of autophagy, which might be advantageous for the inhibition of several engram formation, which may consequently improve the pathology of psychiatric disorders including alcohol dependence. Note that some of significant events such as autophagy initiation, ROS production, and inflammatory reactions have been misplaced for clarity.
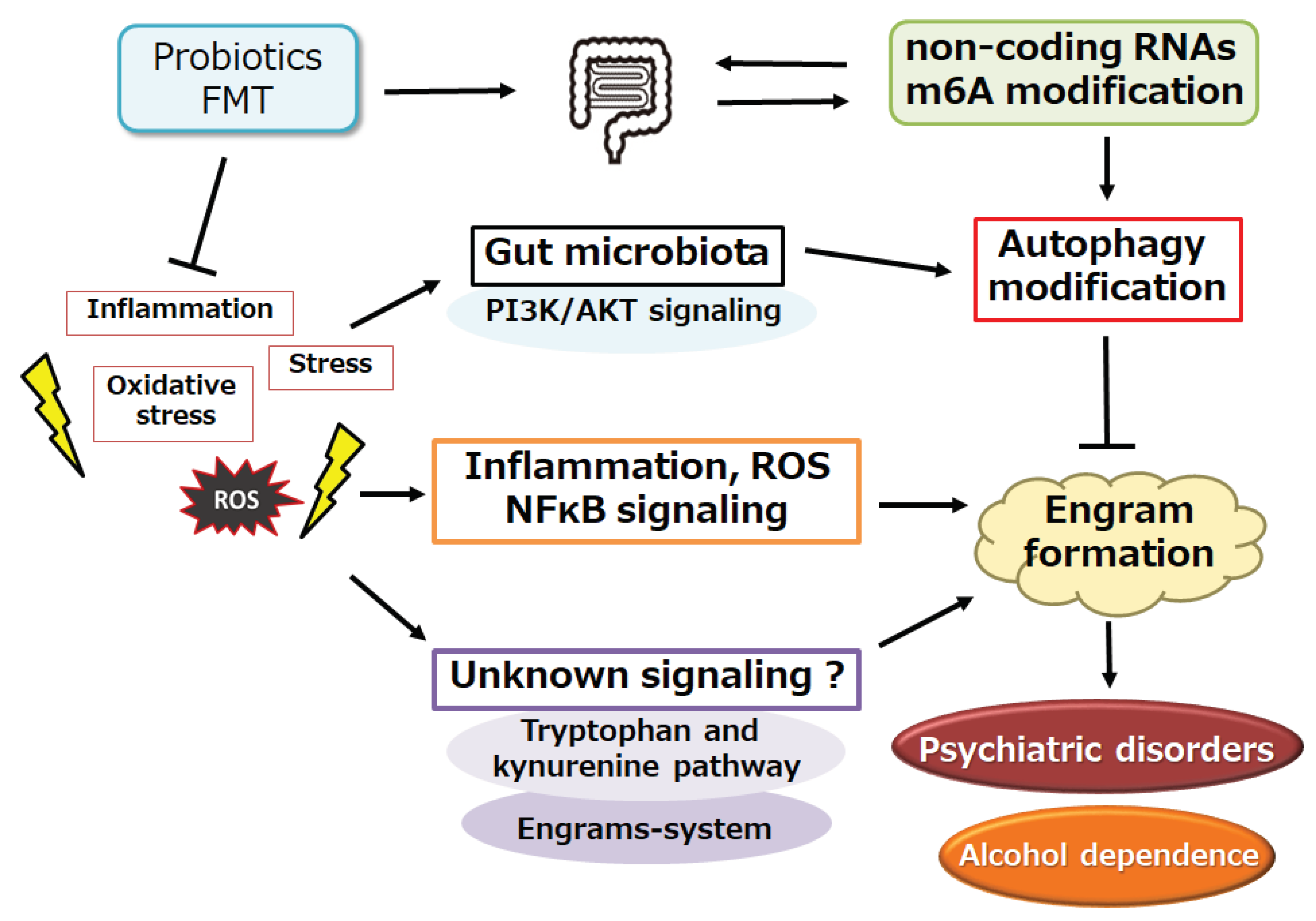
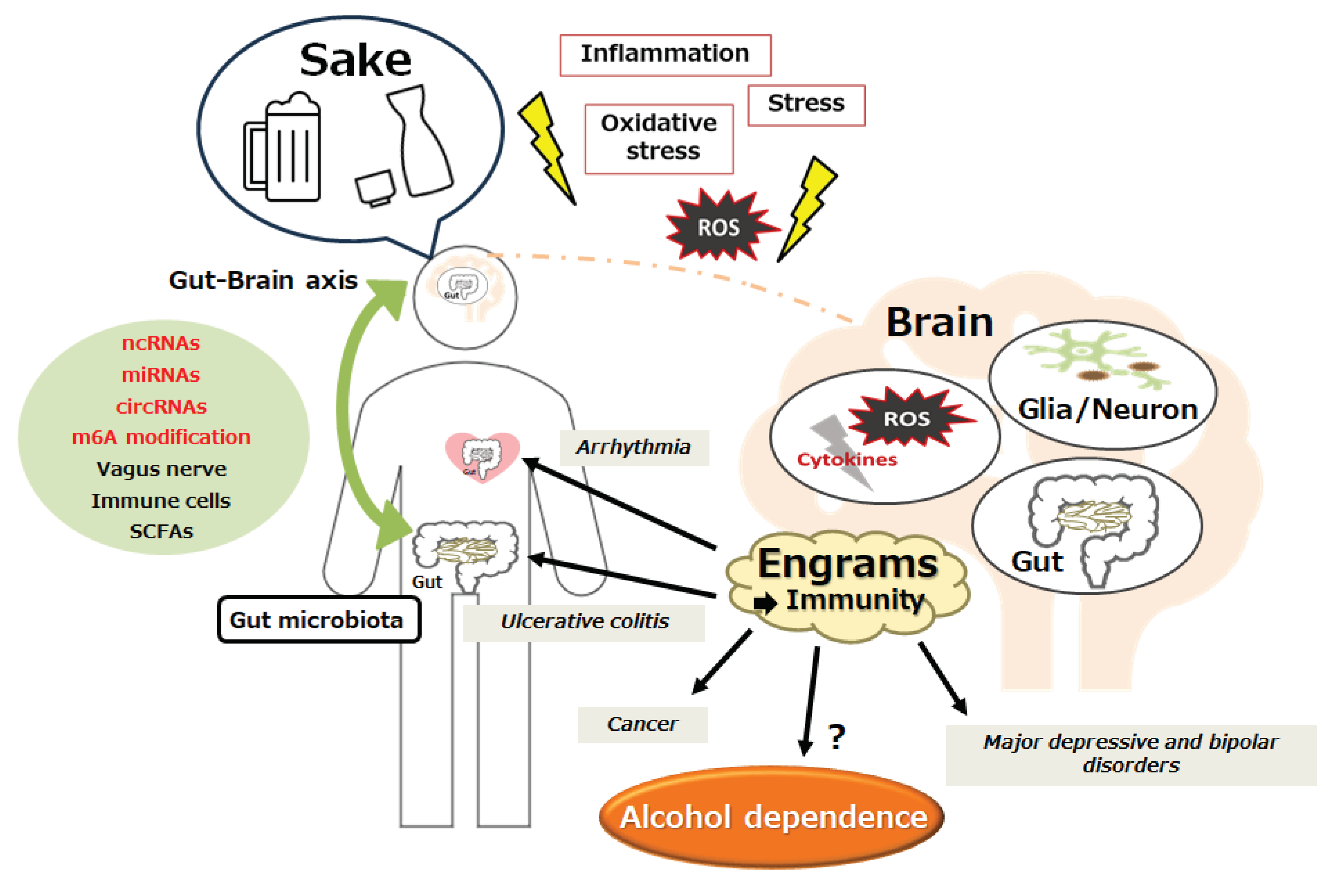
Figure 3.
Schematic impression for the pathogenesis of immune-related diseases such as cardiovascular diseases, acute kidney injury, chronic kidney disease, inflammatory bowel disease, major depressive disorder, bipolar disorder, and alcohol dependence. The gut–brain axis with the utilization of ncRNAs, m6A modification, and/or short chain fatty acids (SCFAs) may contribute to the pathogenesis of immune-related diseases through the formation of several “Engrams” in brain. Inflammation with reactive oxygen species (ROS) may be also involved in the pathway for the modification of immune cells. Note that several important events such as cytokine induction or anti-inflammatory reactions have been omitted for clarity. “?” means for author speculation.
Figure 3.
Schematic impression for the pathogenesis of immune-related diseases such as cardiovascular diseases, acute kidney injury, chronic kidney disease, inflammatory bowel disease, major depressive disorder, bipolar disorder, and alcohol dependence. The gut–brain axis with the utilization of ncRNAs, m6A modification, and/or short chain fatty acids (SCFAs) may contribute to the pathogenesis of immune-related diseases through the formation of several “Engrams” in brain. Inflammation with reactive oxygen species (ROS) may be also involved in the pathway for the modification of immune cells. Note that several important events such as cytokine induction or anti-inflammatory reactions have been omitted for clarity. “?” means for author speculation.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



