
Submitted:
22 February 2024
Posted:
26 February 2024
You are already at the latest version
Abstract
Low efficiency of in vivo transfection of a few fibers revealed a novel tissue network which temporally amplified growth stimulation in the entire regenerating rat soleus muscle. This acupuncture-like effect was demonstrated when the fibers began to grow after complete fiber degradation, synchronous inflammation, myoblast and myotube formation. The first detection of neonatal sarcoplasmic/endoplasmic reticulum ATPase (SERCA1b) has been made in this system. The neonatal, fast and slow SERCA isoforms displayed consequent change with innervation and differentiation, recapitulating events in muscle development. In vivo transfection of myotubes with plasmids expressing dominant negative Ras or a calcineurin inhibitor peptide (Cain/cabin) proved that the expression of the slow myosin heavy chain and the slow muscle type SERCA2a are dif-ferently regulated. In vivo transfection of a few nuclei of myotubes with dnRas or SER-CA1b shRNA stimulated fiber size growth in the whole regenerating muscle but only until the full size had been reached. Growth stimulation by Ras and SERCA1b antisense was abolished in co-transfection with Cain or in case of perimuscular injection with IL4 antibody. This revealed a novel signalling network resembling to scale-free networks which, starting from transfected fiber myonuclei as “hubs”, can amplify growth stimula-tion uniformly in the entire regenerating muscle.
Keywords:
skeletal muscle
; regeneration
; molecular acupuncture
; SERCA
; Ras
; in vivo transfection
1. Introduction
Skeletal muscle is a favourite object of differentiation studies. Not just because it is the largest tissue in mammals contributing to around 40% of body weight [1] but also for its importance in movement and healthy life [2]. To maintain integrity of such organ is crucial therefore the skeletal muscle can adapt with remarkable plasticity, be trained, respond to environmental constrains and to pathological conditions [3,4]. The most dramatic response is needed in regeneration when the muscle rebuilds itself after damage [5]. The main source of regeneration are the muscle stem cells singled out from the satellite cells located between the basal lamina and the sarcolemma [6,7,8]. However, satellite cells behave differently in regeneration than in hypertrophy [9]. In extensive muscle damage bone marrow cells [10,11,12] and non-muscle-resident fibroblasts [13,14] may also transform to myoblast and contribute to myotubes at the beginning and later during the process. Other stem cells called fibro-adipogenic progenitors can regulate the activity of muscle satellite cells [15,16]. A number of experimental models have been developed for muscle regeneration, but only a few based on complete damage of muscle fibres [17]. Regeneration is more even after complete fibre degradation [18] although the kinetics of changings does not seem to depend on the extent of damage [19]. If fibre degradation is partial the inflammation is sustained by type I macrophages that suppress myoblast formation [20,21]. Experimental regenerations can be induced by local anaesthetics or naturally occurring toxins like snake venoms [22,23]. Bupivacain is probably one of the most often used local anaesthetic for this purpose [24,25] and notexin from the Australian tiger snake [26] is a frequently applied venom. However, notexin induces a more complete muscle damage in mouse tibialis anterior (TA) than bupivacaine [27]. It was also found that rat soleus regenerates with a 1-2 days delay after intramuscular injection of notexin compared to that after bupivacaine [28,29]. This is because notexin results in a more extensive muscle fibre damage than bupivacaine [27].
In the past few years a significant progress has been made in understanding the mechanism of notexin effect [30]. This A2 phospholipase (PLA2) analogue toxin binds to unidentified receptors on muscle fibres and neuromuscular junctions, it causes sarcolemma damages and starts perturbation that elevates sarcoplasmic Ca2+ level [31,32]. The myotoxic effect is fairly selective for type I and type IIA fibres and apparently spares early differentiated cells like satellite cells, myoblast precursors and myoblasts [18]. The different pathological consequences are probably not due to different mechanisms of cell toxicity but rather to the intrinsic physiological and anatomical properties of the targeted cells [31]. Accordingly, crotoxin, a myotoxin similar to notexin can be antagonised by cyclosporine A (CSA). This can be interpreted so that CSA inhibits opening of the transient permeability pores of the inner membrane of mitochondria [31]. Because oxidative type I and type IIA muscle fibres are more depending on mitochondria than type IIb and type IIx/d glycolytic fibres they are more vulnerable to PLA2 toxins. Notexin can induce muscle necrosis relatively distant from the site of administration but it does not appear to cross blood brain barrier explaining while it could induce muscle regeneration so effectively after subcutaneous injection [33]. Notexin, in contrast to viper venoms [34] do not create extensive haemorrhage since it leaves the blood vessels, the extracellular tissue and the nerves relatively intact, these all act in favour of an even regeneration [35].
Our aim was to establish an experimental system of complete regeneration in order to monitor developmental events. We intend to align the received information with the pattern of in vivo muscle differentiation. Myotube formation and innervation were considered as crucial landmarks. The synchrony of events was studied with morphological, biochemical and molecular biological methods. The whole extract of muscles from each stages of regeneration was studied with practically no contamination of non-regenerating or out-of-stage regenerating tissues. The complete necrosis and subsequent regeneration was achieved by intramuscular injection of ten times higher dose of the purified snake venom (Sigma) than it was administered subcutaneously in earlier reports [33] reviewed in [36]. Such treatment induced a full regeneration process of the soleus muscle of three months old male Wistar rats of 300-360 grams.
2. Muscle degradation, inflammation and increase of TNF-α level
After 6-24 hrs of the intramuscular injection of appropriate amount of snake venom, the soleus became swollen and lost its red colour. This was probably due to the inflammation and the degradation of myoglobin in mitochondria rich fibres that were the most sensitive to the Ca2+ entry through the sarcolemma [31,32,37]. The venom toxins bind to a receptor on the sarcolemma and tissue damage is achieved by amplification of the initial effect via signal networks [38]. As the rat soleus muscle contain about 70% type I and 28% type IIA and only 1-2% type IIx fibres [39] this meant practically total fibre degradation. The muscle pallor and weight loss, lasted until day 4-5 of regeneration, then the colour was restored by day 5 but the weight was regained only around day 28 of regeneration [40]. The microscopic changes were the most evident on transversal sections of the necrotised-regenerating muscle; the fibre outline faded already at 6 hours and practically diminished by day 3 after venom administration. A number of mononucleated cells populated at endomysial sites and throughout the cross section of the fibers [18]. In the beginning when the optimal dose of the snake venom was established transversal sections were taken along the spindle shaped soleus scanning from proximal to distal tendons in order to check completeness of muscle degradation. This was also confirmed by the dramatic decline or lack of detection of muscle fibre specific transcripts and proteins (i.e. MyHC-s and SERCA-s, skeletal actin) in the whole muscle homogenate [28,40,41]. However, blood vessels and the endomysia outlines were partially preserved until 3 days after toxin administration [28].
In contrast to the decline of the muscle specific proteins, inflammation markers dramatically increased with the muscle necrosis. The lymphocyte/macrophage specific protein MAC 387 increased after 6 hours showing a maximum on day 3 after toxin injection on immunoblot of whole muscle extracts [42]. This marker then gradually declined until 28 days after venom injection but still remained above the normal level [42]. The persistence of MAC 387 indicated that lymphocytes/macrophages apparently accompanied muscle regeneration long after necrosis and probably influenced the autocrine-paracrine environment when regeneration looked macro- and microscopically complete [43]. Tumor necrosis factor α (TNF-α), a pleiotropic pro-inflamatory cytokine increased with about two magnitude at the mRNA and protein levels. The comparison of the position of desmin positive muscle cells and the in situ hybridization of TNF-α mRNA on cross sections showed that the cytokine is not expressed in myoblasts and myotubes at day four of regeneration when its level was still around the maximum. The lack of TNF-α expression in muscle myoblasts and myotubes was supported by the absence of its transcript from differentiating in vitro muscle cell cultures like C2C12 and BC3H1 cells however the mRNAs of TNF receptors were expressed in these cell lines. In the in vivo muscle regeneration the change of TNF-α was paralleled by the increase of TNFR-60 but not TNFR-80 mRNA levels suggesting that of the smaller receptor has primary role in the TNF-α response. The role of this cytokine is complex in muscle regeneration. It has been shown that TNF-α acts on muscle cells, just like in other tissues, through more than one pathway [44,45]. One of these pathways at high level of TNF-α, inhibits myogenesis activating NF-χB which translocates to the nucleus and induces iNOS, IL-1 and again TNF-α [46]. The other pathway prevents differentiation through destabilization of myoD [47]. This leads to pro-proliferative and differentiation-suppressing changes [48,49]. The other pathway is through the p38 kinase which phosphorylates essential myogenic transcription factors like MyoD, myogenin and MEF-2 and therefore promotes differentiation to myotubes [50,51]. The later happens at low TNF concentration. TNF-α is also a chemoattractant of satellite cells and mesangioblasts which are able to contribute to regeneration in dystrophic muscle [51,52]. The balance between inflammation and regeneration might be essential for in vivo muscle regeneration [53]. In a preliminary study, we have administered Remicade (cV1q), a drug containing chimeric antibody to TNF-α, subcutaneously and perimuscularly immediately after the venom injection [54]. The Remicade treatment increased fibre size and the intensity of desmin specific immunostaining at 4 days compared to the control regenerating muscle. This supported that TNF-α inhibits early events of muscle regeneration including the formation and growth of myotubes/primitive fibres. This was in agreement with the ameliorating effect of Remicade on skeletal muscle fibrosis in mdx mice, where, however, it also had a negative impact on cardiac function [55]. Myostatin, the first discovered myokine also appeared in this system, according to its growth inhibitory role, suppressed satellite cell activation [56] and later increasingly maintained levels with differentiation [57].
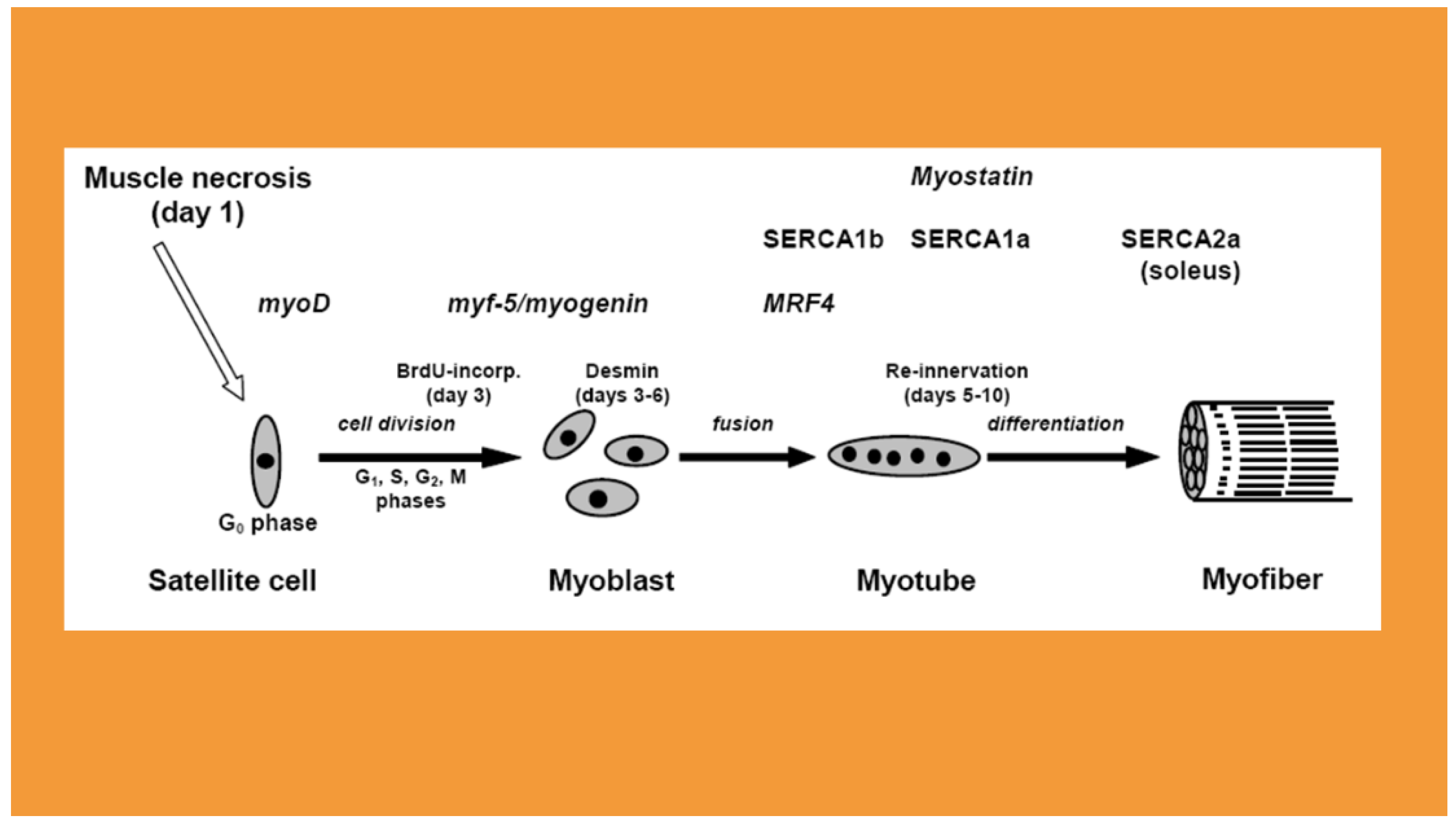
Figure 1.
Schematic summary of the studied gene expressions in rat soleus regeneration. The names MRF, SERCA, myostatin, TNF-α indicate the maximum levels of mRNAs/proteins during the characteristic morphological changes of regeneration. Note the consecutive neonatal (SERCA1b), fast (SERCA1a) and slow type (SERCA2a) calcium pump expressions and that SERCA2a remains the dominant form in the slow type soleus until the end of regeneration. The transfections were done to myotubes formed after for days of regeneration.
Figure 1.
Schematic summary of the studied gene expressions in rat soleus regeneration. The names MRF, SERCA, myostatin, TNF-α indicate the maximum levels of mRNAs/proteins during the characteristic morphological changes of regeneration. Note the consecutive neonatal (SERCA1b), fast (SERCA1a) and slow type (SERCA2a) calcium pump expressions and that SERCA2a remains the dominant form in the slow type soleus until the end of regeneration. The transfections were done to myotubes formed after for days of regeneration.
3. Myogenic regulatory factors
Skeletal muscle regeneration characteristically includes the selection of cells with mesodermal origin and transforming them into myoblasts [58]. It was a breakthrough in understanding of this process when the myogenic regulatory factors (MRFs), the myoD family had been discovered. The four main members of this family, myoD (myf-3 in human), myf-5, myogenin (myf-4 in human) and MRF-4 (herculin or myf-6) were found to transform fibroblasts or other non-muscle-like cells into myoblasts [59,60,61,62]. The MRFs belong to the group of “basic-helix-loop-helix” (bHLH) proteins of transcription factors that bind the E-box (CANNTAG) with their basic domain when form homodimers or heterodimers with each other or (in most cells) with non-myogenic E proteins [63,64,65]. Indeed, the E-box is present in the regulatory region of a number of muscle specific genes like the alpha subunit of nicotinic acetylcholine receptor, the myosin light chain, alpha-actin, desmin, troponin I, and creatine kinase and needed for muscle specific expression [66,67,68]. The different role of MRFs can be assumed according to their spatial and temporal expression and the diverse phenotype of their knock-out mutants [61,69]. The specificity of MRFs can be find in pursuit of interactions of their transactivation domain with other transcription factors. MyoD and myf-5 are playing important role in transforming muscle precursor cells of the somite into myoblasts during early embryogenesis of mammals by activating chromatin loci of muscle specific genes [67,70,71,72].
It was of interest to describe the mRNA levels of myogenic regulatory factors in muscle regeneration and as far as we know our work was the first to do this [18]. Originally we aimed to find homology between muscle regeneration and embryonic myogenesis and reveal differences in slow (soleus) and fast (EDL) muscle regeneration. In soleus regeneration, as it were expected, the myoD increased first on day one and reached a maximum on day 3 then gradually declined to near to normal level by days 21. The myf-5 and myogenin mRNA levels also declined on day one then increased to maximum on day 3 and decreased gradually. On day one the decline of MRF4 mRNA was the most dramatic compared to the other factors and to the normal muscle level and became restored the latest by day 5 of regeneration. This showed that MRF4 is more bound to the myotubes/fibers than to myoblasts. Interestingly the EDL muscle regeneration did not show much differences in changes of myoD, myf-5 and myogenin mRNA levels compared to the soleus. However the MRF4 was not declining significantly as in the soleus most likely because the EDL regeneration was less synchronous and had many not degraded/partially damaged large fibres (probably IIb and IIx types) [18]. Besides this the timing of expression of myogenic factors at the resolution of our work was largely similar to the order (myoD/myf-5, myogenin, MRF4) reported in myogenesis [68,73]. A remarkable difference was in the level myf-5 mRNA which increased in myogenesis at about the same time as the myoD message did [63] but it declined in our regeneration system in contrast to myoD. It has been reported that the myoD knock-out mice showed a slightly delayed but otherwise quite normal muscle development. This could happen because the increased myf-5 expression with the same cell specificity complemented for myoD function [69]. Such an altered regulation of myf-5 and myoD might happen in muscle regeneration as well. The other explanation for the decreased myf-5 message level might be that the satellite cells expressing myf-5 [74,75] were transformed into muscle progenitor cells and myoblasts and from that time expressed predominantly myoD.
The myoD protein followed its message in the regenerating soleus and showed a 6x increase compared to the normal level, a maximum on the 3rd day after venom injection. This was paralleled by the maximum level of BrdU incorporation in to the nucleus implicating that the proliferating cells, probably myoblasts expressed the transcription factor. We interfered with the myoD expression by injecting antisense oligonucleotides into the muscle on the 3rd day of regeneration [76]. The antisense decreased the mRNA and protein level of myoD dramatically at one hour after the treatment; the myoD protein was about 3-fold less in the isolated myonuclei compared to the control regenerating soleus injected with scrambled oligonucleotide. The progress of regeneration was also delayed by this treatment as it was judged two days later by desmin expression, myotube formation and the number of primitive endplates [76,77]. This was somewhat a surprise since myoD was reported to have a relatively quick turnover in cell culture [78,79]. Apparently our antisense oligo started to degrade after four hours and the myoD mRNA level was also restored by that time [76]. However the antisense still worked probably because it hybridized to the translation start of myoD [80] and contained a TCC sequence that enhanced the antisense effect by stimulating RNase H activity [81]. The antisense effect resulted in other interesting changes; the mRNA levels of myoD myf-5 and myogenin were elevated again after 24 hours probably to bring in the backlog of regeneration. However the mRNA levels of MRF4 and the myogenic factor inhibitory proteins Id1 and Id3 did not change. In the subsequent period no apparent difference was observed in the myoD antisense treated muscles except that after 28 days more small and split fibres were reflecting the delayed regeneration [76]. The above results clearly showed that myoD has a crucial role of in muscle regeneration and indicated that this system can be a tool for investigating similar functions of the other myogenic factors [82]. The expression of connexins have also been screened in notexin induced in vivo muscle regeneration by others. They found that only connexion 43 gap junction protein (Cx43) is present and it is upregulated transiently before myoblast fusion [83]. Dye injection of in vitro primary muscle cells revealed more intense gap junction coupling between sparsely located pre-fusion myoblasts than between the close fusion aligned myoblasts, suggesting that this tight junction protein takes part in synchronisation of cell cycle control and preparation for fusion to syncytial cell [84]. Transfection of skeletal muscle primary cultures with dominant negative Cx43 confirmed that activity of this protein is needed for myoblast proliferation and syncytial fusion into myotubes [85]. Later it has been found that in regenerating muscle the cluster of differentiated satellite cells which represent an advanced stage of fusion in the inner part of the basal membrane do not express Cx43, suggesting that this protein does not participate actively in myoblast fusion although it is required for preparation to that process [86].
4. The expression of sarco/endoplasmic reticulum Ca2+ ATPases (SERCAs) in regeneration
The SERCAs are a group of transmembrane proteins that pump the Ca2+ from the cyto/sarcoplasm to the endo/sarcoplasmic reticulum. In striated muscle this creates a situation in which the troponin complex can occupy the actin binding place of myosin head and prevent actomyosin formation [87]. This has an impact in aging and various pathological conditions [88]. There are three strongly homologous SERCA genes (SERCA1-3 or ATP2A1-3 according to the human nomenclature) but only two of them, SERCA1 and SERCA2 have splice variants expressed in striated muscle. SERCA1a is responsible for relaxation in adult fast skeletal muscle, SERCA1b is expressed in neonatal muscle and SERCA2a is found in slow skeletal muscle and cardio myocyte in heart ventricle [89,90,91]. The mRNA of SERCA1b was detected mostly in neonatal skeletal muscle and our work was the first to demonstrate that the protein is highly specific to myotubes and developing fibers but not transforming or adapting muscles [92]. We also showed that SERCA1b is a major isoform in regenerating (fast or slow) muscle but not expressed in a healthy adult rat or adult human muscle where otherwise its splicing could be several-fold increased i.e. by denervation or stretch [92,93,94,95,96,97]. We also found in accordance with others [98,99] that the SERCA1b protein was expressed in a pathological condition of adult human muscle called myotonic dystrophy type II [96]. Nevertheless, although the regenerative potential was up-built in ischemic lower limb muscle of atherosclerosis obliterans no SERCA1b only SERCA1a was upregulated [97].
In accordance the neonatal 1b is the first SERCA isoform that appears shortly after the myogenic regulatory factors in regenerating rat soleus and EDL muscles [28,41,100]. This predominant expression of SERCA1b indicates myotube formation and it is the forerunner of the orchestrated pattern of SERCA isoforms found characteristically at both message and protein levels in muscle regeneration. This motif is strongly dependent on the progress of innervation, the type of innervation (fast or slow) and it is expressed in developing myofibres before they become fast or slow. This means that the start of innervation will switch the splicing of SERCA1 transcript from neonatal to adult fast type mRNA in both fast and slow muscle regeneration [28,100]. The switch between SERCA1a and SERCA2a occurs in slow fibre regeneration when innervation is more established [77,101] and the fast SERCA1a declines while the slow SERCA2a elevates becoming exclusive (or at least predominant) [28,41]. Interestingly the fast twitch muscle follows an alternate motif which starts with a moderate elevation then decline of SERCA2a expression, paralleled with those of SERCA1b before the increase and predominance of SERCA1a while the innervation progresses [100].
The change of SERCA levels appears to be similar to the innervation dependent expression of myosin isoforms in regeneration especially in the predominantly slow soleus muscle [33,102]. This prompted us to investigate if the slow muscle type SERCA2a level is co-regulated with that of the heavy chain of corresponding slow myosin isoform, MyHC1. In bupivacaine induced regeneration of rat soleus the MyHC1 expression is upregulated by slow type innervation [29]. The upregulation can be prevented by transfecting dominant negative Ras N17 (dnRas) into the regenerating muscle fibres [29]. Interestingly when bupivacain regeneration was induced in denervated rat soleus the transfection of Ras V12 S35, which influenced downstream the MAPK pathway, restored the MyHC1 expression in the fibres while the other Ras mutants did not [29]. This suggested that a distinct well defined pathway mediates the nerve effect on MyHC1 expression in regeneration. We wondered if the same pathway could also function in upregulation of SERCA2a. However, we found that the expression of SERCA2a was not dependent on slow type innervation like the MyHC1 neither could be influenced by transfection of Ras mutants regulating MyHC1 expression [40]. Therefore the slow type of SERCA2a and MyHC1 are not co-regulated by innervation and Ras. Similar conclusion was drawn after experiments with calcineurin [103]. The calcineurin-NFAT pathway was also found to mediate the effect of slow type innervation on the expression of MyHC1 in bupivacaine induced regeneration of rat soleus [104,105]. However, transfection with Cain, while it prevented MyHC1, it did not prevent the expression of SERCA2a in the notexin induced regeneration of rat soleus [103]. This again confirmed that the SERCA2a expression is regulated by separate pathway(s) from those controlling slow myosin expression. Later it was reported that slow myosin do not depend neither on slow type innervation nor on calcineurin in other muscle than the soleus [106]. However we have not found a similar report for SERCA2a. The question remained: what regulates then SERCA2a expression? First of all SERCA2a regulation is not exerted so much on the transcript level as the MxHC1 expression [29] but clearly has more components [107]. One of these complex effects is the passive stretch that works in selective denervation of soleus but not in ischiadic (hindlimb) denervation. To illustrate this, SERCA2a was upregulated in the selective denervated vs. ischiadic denervated regenerating soleus [40]. Similarly SERCA2a was upregulated in dnRas transfected fibres of innervated regenerating muscle which obviously were stretched by the overwhelming innervated fibres [40].
5. A novel muscle growth regulation revealed by molecular acupuncture in regeneration takes after scale-free network
The transfection of regenerating soleus with dnRas resulted in a phenomenon that according to our knowledge has not been reported yet [108]; besides inhibiting expression of slow MyHCI in the transfected fibre [29] dnRas resulted in a non-fiber-autonomous effect. Although only 20-30 fibres were transfected out of the total (approximately 2500), expressing the transgene in a few of their myonuclei [109], the fibre size (cross sectional area, CSA) was increased in the entire muscle between days 7-12 of regeneration compared to the control regenerating soleus transfected with the empty vector [108]. The size of transfected fibres was not different from the non-transfected ones within the same muscle and showed no morphological or histochemical difference. However, after 21 days when the full fiber size was nearly reached, the growth increase was attenuated therefore it did not exceeded that of the control. The augmentation of fibre growth must have started after transfection on day 4 and was finished by day 21 of regeneration. and was apparently mediated by autocrine/paracrine effect. One of the factors that might orchestrate the effect of transfection was interleukin 4 (IL4) since the perimuscular injection of its antibody stopped the augmentation of CSA increase. However, the IL4 antibody did not decreased fibre size below the level of the non-transfected regenerating soleus suggesting that the cytokine has a role in augmentation of fibre growth rather than in the normal regeneration process. Interestingly the higher IL4 level was detectable only around the transfected fibres by immunohistochemistry and not in the whole muscle by immunoblot supporting local importance instead of a wide spread systematic one. The IL4 is secreted by growing myotubes and acts on receptors of myoblast stimulating their accretion [110,111]. As the IL4 expression is regulated by the calcineurin-NFAT pathway [110,112] the calcineurin inhibitor peptide cain/cabin was co-transfected with dnRas in the regenerating soleus [108]. This prevented the augmentation of CSA increase suggesting that the calcineurin-NFAT-IL4 was activated by Ras inhibition. This revealed a scenario in which the calcineurin-NFAT and Ras pathways act in balance in controlling fibre growth of regenerating soleus muscle. According to this scene Ras kept calcineurin under inhibition but the transfected dnRas gave a way for the increase of calcineurin activity and growth augmentation.
Although Ras and calcineurin are members of two important pathways to our knowledge there are little data about their connection in skeletal muscle. However, in neonatal cardiac myocytes their interaction appears synergistic not antagonistic as in the regenerating muscle since the constitutively active Ras increased the activity of NFAT3 [113]. Ras acted upstream of calcineurin on NFAT3 activity, and similarly, dominant negative Ras inhibited the stimulated NFAT nuclear location and transcriptional activity [113]. Essentially similar coordinated functions have been found in vivo between calcineurin-NFAT and the Ras stimulated MAPK-ERK pathway in cardiac hypertrophy of transgenic mice [114]. Since this interaction has not been enlightened in muscle cells we silenced the main pump that regulates sarcoplasmic [Ca2+]i in regenerating muscle fibres. SERCA1b is a major isoform in neonatal and regenerating muscles [92,96,98]. Transfection of SERCA1b shRNA expressing vector into fibres of regenerating soleus resulted in augmentation of CSA increase in both transfected and non-transfected fibres of the whole muscle [115] in a similar manner to that of dnRAS [108]. The growth stimulation was also calcineurin and IL4 dependent as it was abolished by co-transfection with Cain and the perimuscular injection of IL4 antibody [115]. This indicated the pivotal role of the calcineurin-NFAT-IL4 pathway in both dnRas and SERCA1b silencing induced fiber growth augmentation of regenerating soleus muscle.
It is worth to emphasize that silencing of SERCA1b in a small fraction of fibers had dramatically different effect from that of knocking out of SERCA1 gene in mice as the later resulted in perinatal lethality [116]. However, it showed a somewhat similar phenomenon to the muscle specific knock out of SERCA inhibiting micropeptide myoregulin which in mice resulted in enhanced exercise performance [117].
Remarkably, only a few (1-4) myonuclei had been transfected per fiber in the muscle belly [109]. The total number of myonuclei in a regenerating fiber can be estimated as several fold higher than the ones having been transfected [109,118]. Therefore, the growth stimulation must have been initiated from a small minority of myonuclei [108,109,115]. However, the myonuclei surrounded by zones form myonuclear domain which is a functional units of gene expression within the multinucleated fiber [119]. The plasmids have to reach the nuclei therefore transfect the myonuclear domain in order to get expression. Such rare transfection of nuclei like it happened in the regenerating rat soleus is resembling to the effect of acupuncture [120]. However, this time the manipulation was made by molecular inhibitions such as the dominant negative Ras or SERCA1b silencing instead of a special needle. So this phenomenon can be described illustratively as a molecular acupuncture of regenerating skeletal muscle. Furthermore, if one wants to depict the relationship of transfected and non-transfected myonuclear domains in the above described stimulated fiber growth, it is appealing to adapt a scale-free network at least for the initial events (Figure 2) [121,122]. In this network the nodes are not identical since some of them have higher number of links than most of the rest has. These high-degree nodes become hubs compared to the other low-degree nodes which are in abundance. The hub quality is gained by transfected myonuclei because of their calcineurin activity while the non-transfected nuclei remain low-degree nodes. The regenerating muscle is probably self-organizing network and create new hubs from low-degree nodes by other features as none of the transfected fibers showed hypertrophy compared to the non-transfected others. Independent regulation of myonuclei for gene expression has been shown along multinucleated fibers and suggested to be organized in stochastic pattern [123]. However, according to my knowledge, no network like interaction between myonuclear domains in different myofibers has been reported. This also suggests that the regeneration system discussed here may fill a gap in the study of biological networks.
6. Conclusions
A synchronized in vivo muscle regeneration revealed a novel network of growth regulation. The network is stimulated by changing the equilibrium between two major signal transduction systems, controlled by Ras and calcineurin, within the same muscle fibre. Shifting this dynamic balance, by stimulating calcineurin, influences other fibers probably, in autocrine-paracrine way and augments fiber growth in the entire regenerating muscle. The reason that such an effect has not been reported before might be that the growth stimulation was not possible to detect reliably in unsynchronized and partial regenerations where the fibers did not grow evenly therefore comparison with the control was hampered. So the notexin induced soleus regeneration in rat offers a unique in vivo system for studying novel effects of signal pathway interactions and apparently represents an example of scale-free networks of signalling in skeletal muscle. It would be interesting to see whether other co-transfections with dnRAS could interfere with SERCA1b shRNA induced fibre growth amelioration and if the transfection alone with calcineurin inhibitor Cain could have any effect. Experiments in this direction are on their way.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Data sharing not applicable.
Acknowledgments
May be filled out later
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Kim, K.M.; Jang, H.C.; Lim, S. Differences among skeletal muscle mass indices derived from height-, weight-, and body mass index-adjusted models in assessing sarcopenia. Korean J Intern Med 2016, 31, 643–650. [Google Scholar] [CrossRef] [PubMed]
- Henrot, P.; Blervaque, L.; Dupin, I.; Zysman, M.; Esteves, P.; Gouzi, F.; Hayot, M.; Pomiès, P.; Berger, P. Cellular interplay in skeletal muscle regeneration and wasting: Insights from animal models. J Cachexia Sarcopenia Muscle 2023, 14, 745–757. [Google Scholar] [CrossRef] [PubMed]
- Lieber, R.L.; Roberts, T.J.; Blemker, S.S.; Lee, S.S.M.; Herzog, W. Skeletal muscle mechanics, energetics and plasticity. J Neuroeng Rehabil 2017, 14, 108. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, K.M.; Haddad, F. The Evolution of Skeletal Muscle Plasticity. in Response to Physical Activity and Inactivity; Muscle and Exercise Physiology, Zoladz, J.A., Ed.; Academic Press, 2019, 347–377. [CrossRef]
- Laumonier, T.; Menetrey, J. Muscle injuries and strategies for improving their repair. J Exp Orthop 2016, 3, 15. [Google Scholar] [CrossRef] [PubMed]
- Mauro, A. Satellite cells of skeletal muscle fibers. J Biophys Biochem Cytol 1961, 9, 493–495. [Google Scholar] [CrossRef] [PubMed]
- Seale, P.; Rudnicki, M.A. A new look at the origin, function, and ‘‘stem-cell’’ status of muscle satellite cells. Dev Biol 2000, 218, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Morrissey, J.B.; Cheng, R.Y.; Davoudi, S.; Gilbert, P.M. Biomechanical Origins of Muscle Stem Cell Signal Transduction. J Mol Biol 2016, 428, 1441–1454. [Google Scholar] [CrossRef]
- Fukada, S.I.; Akimoto, T.; Sotiropoulos, A. Role of damage and management in muscle hypertrophy: Different behaviors of muscle stem cells in regeneration and hypertrophy. Biochim Biophys Acta Mol Cell Res 2020, 1867, 118742. [Google Scholar] [CrossRef]
- Ferrari, G.; Cusella-De Angelis, G.; Coletta, M.; Paolucci, E.; Stornaiuolo, A.; Cossu, G.; Mavilio, F. Muscle regeneration by bone marrow-derived myogenic progenitors. Science 1998, 279, 1528–1530. [Google Scholar] [CrossRef]
- Schulze, M.; Belema-Bedada, F.; Technau, A.; Braun, T. Mesenchymal stem cells are recruited to striated muscle by NFAT/IL-4-mediated cell fusion. Genes Dev 2005, 19, 1787–1798. [Google Scholar] [CrossRef]
- Andrade, B.M.; Baldanza, M.R.; Ribeiro, K.C.; Porto, A.; Peçanha, R.; Fortes, F.S.; Zapata-Sudo. G.; Campos-de-Carvalho, A.C.; Goldenberg, R.C.; Werneck-de-Castro, J.P. Bone marrow mesenchymal cells improve muscle function in a skeletal muscle re-injury model. PLoS ONE. 2015, 10, e0127561. [Google Scholar] [CrossRef]
- Gibson, A.J.; Karasinski, J.; Relvas, J.; Moss, J.; Sherratt, T.G.; Strong, P.N.; Watt, D.J. Dermal fibroblasts convert to a myogenic lineage in mdx mouse muscle. J Cell Sci 1995, 108, 207–214. [Google Scholar] [CrossRef]
- Chapman, M.A.; Meza, R.; Lieber, R. L Skeletal muscle fibroblasts in health and disease. Differentiation. 2016, 92, 108–115. [Google Scholar] [CrossRef]
- Giuliani, G.; Rosina, M.; Reggio, A. Signaling pathways regulating the fate of fibro/adipogenic progenitors (FAPs) in skeletal muscle regeneration and disease. FEBS J. 2022, 289, 6484–6517. [Google Scholar] [CrossRef]
- Molina, T.; Fabre, P.; Dumont, N.A. Fibro-adipogenic progenitors in skeletal muscle homeostasis, regeneration and diseases. Open Biol 2021, 11, 210110. [Google Scholar] [CrossRef] [PubMed]
- Carlson, B.M. Muscle regeneration in animal models. in: Skeletal Muscle Repair and Regeneration. Springer; Schiaffino, S.; Partridge, T.; 2008, pp. 163–180. [CrossRef]
- Mendler, L.; Zádor, E.; Dux, L.; Wuytack, F. mRNA levels of myogenic regulatory factors in rat slow and fast muscles regenerating from notexin-induced necrosis. Neuromusc Disorders 1998, 8, 533–541. [Google Scholar] [CrossRef] [PubMed]
- Bernard, C.; Jomard, C.; Chazaud, B.; Gondin, J. Kinetics of skeletal muscle regeneration after mild and severe muscle damage induced by electrically-evoked lengthening contractions. FASEB J. 2023, 37, e23107. [Google Scholar] [CrossRef] [PubMed]
- Tidball, J.G.; Dorshkind, K.; Wehling-Henricks, M. Shared signaling systems in myeloid cell-mediated muscle regeneration. Development 2014, 141, 1184–1196. [Google Scholar] [CrossRef] [PubMed]
- Khuu, S.; Fernandez, J.W.; Handsfield, G.G. Delayed skeletal muscle repair following inflammatory damage in simulated agent-based models of muscle regeneration. PLoS Comput Biol 2023, 19, e1011042. [Google Scholar] [CrossRef] [PubMed]
- Bordon, K.C.F.; Cologna, C.T.; Fornari-Baldo, E.C.; Pinheiro-Júnior, E.L.; Cerni, F.A.; Amorim, F.G.; Anjolette, F.A.P.; Cordeiro, F.A.; Wiezel, G.A. , Cardoso, I.A., Ferreira, I.G.; de Oliveira, I.S.; Boldrini-França, J.; Pucca, M.B., Baldo, M.A.; Arantes, E.C. From Animal Poisons and Venoms to Medicines: Achievements, Challenges and Perspectives in Drug Discovery. Front Pharmacol 2020, 11, 1132. [Google Scholar] [CrossRef]
- Gutiérrez, J.M.; Lomonte, B. Phospholipases A2: Unveiling the secrets of a functionally versatile group of snake venom toxins. Toxicon. 2013, 62, 27–39. [Google Scholar] [CrossRef]
- Benoit, P.W.; Belt, W.D. Destruction and regeneration of skeletal muscle after treatment with a local anaesthetic, bupivacaine (Marcaine). J Anat 1970, 107, 547–556. [Google Scholar] [PubMed]
- Chen, Y,; Li, X. ; Huo, Z.; Chen, H.; Zhang, L. An overview of bupivacaine-induced morphological changes: A novel animal model of skeletal muscle injury. Int J Clin Exp Med 2020, 13, 7–15. [Google Scholar]
- Harris, J.B.-; Johnson, M.A.; Karlsson, E. Pathological responses of rat skeletal muscle to a single subcutaneous injection of a toxin isolated from the venom of the Australian tiger snake, Notechis scutatus scutatus. Clin Exp Pharmacol Physiol 1975, 2, 383–404. [Google Scholar] [CrossRef]
- Plant, D.R.; Colarossi, F.E.; Lynch, G.S. Notexin causes greater myotoxic damage and slower functional repair in mouse skeletal muscles than bupivacaine. Muscle Nerve 2006, 34, 577–585. [Google Scholar] [CrossRef] [PubMed]
- Zádor, E.; Mendler, L.; Ver Heyen, M.; Dux, L.; Wuytack, F. Changes in mRNA levels of the sarcoplasmic/endoplasmic-reticulum Ca(2+)-ATPase isoforms in the rat soleus muscle regenerating from notexin-induced necrosis. Biochem, J. 1996, 320, 107–113. [Google Scholar] [CrossRef]
- Murgia, M.; Serrano, A.L.; Calabria, E.; Pallafacchina, G.; Lomo, T.; Schiaffino, S. Ras is involved in nerve-activity-dependent regulation of muscle genes. Nat Cell Biol 2000, 2, 142–147. [Google Scholar] [CrossRef]
- Zhou, K.; Luo, W.; Liu, T.; Ni, Y.; Qin, Z. Neurotoxins Acting at Synaptic Sites: A Brief Review on Mechanisms and Clinical Applications. Toxins 2022, 15, 18. [Google Scholar] [CrossRef]
- Montecucco, C.; Gutiérrez, J.M.; Lomonte, B. Cellular pathology induced by snake venom phospholipase A2 myotoxins and neurotoxins: Common aspects of their mechanisms of action. Cell Mol Life Sci 2008, 65, 2897–2912. [Google Scholar] [CrossRef]
- Cintra-Francischinelli, M.; Pizzo, P.; Rodrigues-Simioni, L.; Ponce-Soto, L.A.; Rossetto, O.; Lomonte, B.; Gutiérrez, J.M.; Pozzan, T.; Montecucco, C. Calcium imaging of muscle cells treated with snake myotoxins reveals toxin synergism and presence of acceptors. Cell Mol Life Sci 2009, 66, 1718–1728. [Google Scholar] [CrossRef]
- Whalen, R.G.; Harris, J.B.; Butler-Browne, G.S.; Sesodia, S. Expression of myosin isoforms during notexin-induced regeneration of rat soleus muscles. Dev Biol 1990, 141, 24–40. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, J.M.; Rucavado, A.; Escalante, T.; Herrera, C.; Fernández, J.; Lomonte, B.; Fox, J.W. Unresolved issues in the understanding of the pathogenesis of local tissue damage induced by snake venoms. Toxicon. 2018, 148, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Harris, J.B.; Scott-Davey, T. Secreted phospholipases A2 of snake venoms: Effects on the peripheral neuromuscular system with comments on the role of phospholipases A2 in disorders of the CNS and their uses in industry. Toxins 2013, 5, 2533–2571. [Google Scholar] [CrossRef] [PubMed]
- Harris, J.B. Myotoxic phospholipases A2 and the regeneration of skeletal muscles. Toxicon 2003, 42, 933–945. [Google Scholar] [CrossRef] [PubMed]
- Gasanov, S.E.; Dagda, R.K.; Rael, E.D. Snake Venom Cytotoxins, Phospholipase A2s, and Zn2+-dependent Metalloproteinases: Mechanisms of Action and Pharmacological Relevance. J Clin Toxicol 2014, 4, 1000181. [Google Scholar] [CrossRef] [PubMed]
- Bickler, P.E. Amplification of Snake Venom Toxicity by Endogenous Signaling Pathways. Toxins. 2020, 12, 68. [Google Scholar] [CrossRef]
- Delp, M.D.; Duan, C. Composition and size of type I, IIA, IID/X, and IIB fibers and citrate synthase activity of rat muscle. J Appl Physiol 1996, 80, 261–270. [Google Scholar] [CrossRef]
- Zádor, E.; Wuytack, F. Expression of SERCA2a is independent of innervation in regenerating soleus muscle. Am J Physiol -Cell Physiol 2003, 285, C853–C861. [Google Scholar] [CrossRef]
- Zádor, E.; Szakonyi, G.; Rácz, G.; Mendler, L.; Ver Heyen, M.; Lebacq, Dux, L. ; Wuytack, F. Expression of the sarco/endoplasmic reticulum Ca2+-transport ATPase protein isoforms during regeneration from notexin induced necrosis of rat muscle. Acta Histochem 1998, 100, 355–369. [Google Scholar] [CrossRef]
- Zádor E, Mendler L, Takács V, De Bleecker J and Wuytack, F. Regenerating soleus and EDL muscles of the rat show elevated levels of TNF-α and its receptors, TNFR-60 and TNFR-80. Muscle and Nerve, 2001, 24, 1058–1067. [Google Scholar] [CrossRef]
- Tidball, J.G.; Villalta, S.A. Regulatory interactions between muscle and the immune system during muscle regeneration. Am J Physiol Regul Integr Comp Physiol 2010, 298, R1173–R1187. [Google Scholar] [CrossRef]
- Mourkioti, F.; Rosenthal, N. NF-kappaB signaling in skeletal muscle: Prospects for intervention in muscle diseases. J Mol Med 2008, 86, 747–759. [Google Scholar] [CrossRef]
- Webster, J.D.; Vucic, D. The Balance of TNF Mediated Pathways Regulates Inflammatory Cell Death Signaling in Healthy and Diseased Tissues. Front Cell Dev Biol 2020, 8, 365. [Google Scholar] [CrossRef]
- Bhatnagar, S.; Panguluri, S.K.; Gupta, S.K.; Dahiya, S.; Lundy, R.F.; Kumar, A. Tumor necrosis factor-α regulates distinct molecular pathways and gene networks in cultured skeletal muscle cells. PLoS ONE. 2010, 5, e13262. [Google Scholar] [CrossRef]
- Langen, R.C.; Van der Velden, J.L.; Schols, A.M.; Kelder, M.C.; Wouters, E.F. Janssen-Heininger, Y.M. Tumor necrosis factor-alpha inhibits myogenic differentiation through MyoD protein destabilization. FASEB J 2004, 18, 227–237. [Google Scholar] [CrossRef]
- Li, Y.P. TNF-α is mitogen in skeletal muscle. Am J Physiol-Cell Physiol 2003, 285, C370–C376. [Google Scholar] [CrossRef]
- O'Brien, M.E.; Londino, J.; McGinnis, M.; Weathington, N.; Adair, J.; Suber, T.; Kagan, V.; Chen, K.; Zou, C.; Chen, B.; Bon, J.; Mallampalli, R.K. Tumor Necrosis Factor Alpha Regulates Skeletal Myogenesis by Inhibiting SP1 Interaction with cis-Acting Regulatory Elements within the Fbxl2 Gene Promoter. Mol Cell Biol 2020, 40, e00040–20. [Google Scholar] [CrossRef]
- Chen, S.E.; Jin, B.; Li, Y.P. TNF-alpha regulates myogenesis and muscle regeneration by activating p38 MAPK. Am J Physiol Cell Physiol 2007, 292, C1660–C1671. [Google Scholar] [CrossRef]
- Careccia, G.; Mangiavini, L.; Cirillo, F. Regulation of Satellite Cells Functions during Skeletal Muscle Regeneration: A Critical Step in Physiological and Pathological Conditions. Int J Mol Sci. 2023, 25, 512. [Google Scholar] [CrossRef]
- Ceafalan, L.C.; Popescu, B.O.; Hinescu, M.E. Cellular players in skeletal muscle regeneration. Biomed Res Int 2014, 2014, 957014. [Google Scholar] [CrossRef]
- Tu, H.; Li, Y.L. Inflammation balance in skeletal muscle damage and repair. Front Immunol 2023, 14, 1133355. [Google Scholar] [CrossRef]
- Kovács, E.; Zádor, E. The effect of a TNF-α inhibiting drug on skeletal muscle regeneration. XXXIV. European Muscle Conference, Hungary, Hortobágy J Muscle Res Cell Motil 2005, 26, p89 (Abstract). [Google Scholar]
- Ermolova, N.V.; Martinez, L.; Vetrone, S.A.; Jordan, M.C.; Roos, K.P.; Sweeney, H.L.; Spencer, M.J. Long-term administration of the TNF blocking drug Remicade (cV1q) to mdx mice reduces skeletal and cardiac muscle fibrosis, but negatively impacts cardiac function. Neuromuscul Disord. 2014, 24, 583–595. [Google Scholar] [CrossRef]
- McCroskery, S.; Thomas, M.; Maxwell, L.; Sharma, M.; Kambadur, R. Myostatin negatively regulates satellite cell activation and self-renewal. J Cell Biol 2003, 162, 1135–1147. [Google Scholar] [CrossRef] [PubMed]
- Mendler, L.; Zador, E.; Ver Heyen, M.; Dux, L.; Wuytack, F. Myostatin in regenerating rat muscles and in myogenic cell cultures. J Muscle Res and Cell Mot 2000, 21, 551–563. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Outeiriño, L.; Hernandez-Torres, F.; Ramírez-de Acuña, F.; Matías-Valiente, L.; Sanchez-Fernandez, C.; Franco, D.; Aranega, A.E. Muscle Satellite Cell Heterogeneity: Does Embryonic Origin Matter? Front Cell Dev Biol 2021, 9, 750534. [Google Scholar] [CrossRef] [PubMed]
- Vaidya, T.B.; Rhodes, S.J.; Moore, J.L.; Sharman, D.A.; Konieczny, S.F.; Taparowsky, E.J. Isolation and structural analysis of the rat myoD gene. Gene 1992, 116, 223–230. [Google Scholar] [CrossRef]
- Miller, J.B.; Everitt, E.A.; Smith, T.H.; Block, N.E.; Dominov, J.A. Cellular and Molecular Diversity in Skeletal Muscle Development: News from in vitro and in vivo. BioEssays 1993, 15, 191–195. [Google Scholar] [CrossRef]
- Olson, E.N.; Klein, W.H. bHLH factors in muscle development: Dead lines and commitments, what to leave in and what to leave out. Genes and Devel 1994, 8, 1–8. [Google Scholar] [CrossRef]
- Motohashi, N.; Asakura, A. Muscle satellite cell heterogeneity and self-renewal. Front Cell Dev Bio. 2014, 2, 1. [Google Scholar] [CrossRef]
- Ludolph, D.C.; Konieczny, S.F. Transcription factor families: Muscling in on the myogenic program. FASEB J 1995, 9, 1595–1604. [Google Scholar] [CrossRef]
- Maione, R.; Amati, P. Interdependence between muscle differentiation and cell cycle control. Biochim Biophys Acta 1997, 1322, M19–M30. [Google Scholar] [CrossRef]
- Tapscott, S.J. The circuitry of a master switch: Myod and the regulation of skeletal muscle gene transcription. Development 2005, 132, 2685–2695. [Google Scholar] [CrossRef]
- Lowe, D.A.; Lund, T.; Alway, S.E. Hypertrophy-stimulated myogenic regulatory factor mRNA increases are attenuated in fast muscle of aged quails. Am J Physiol-Cell Physiol 1998, 275, C155–C166. [Google Scholar] [CrossRef]
- Singh, K.; Dilworth, F.J. Differential modulation of cell cycle progression distinguishes members of the myogenic regulatory factor family of transcription factors. FEBS J 2013, 280, 3991–4003. [Google Scholar] [CrossRef]
- Zammit, P.S. Function of the myogenic regulatory factors Myf5, MyoD, Myogenin and MRF4 in skeletal muscle, satellite cells and regenerative myogenesis. Semin Cell Dev Biol 2017, 72, 19–32. [Google Scholar] [CrossRef]
- Rudnicki, M.A. ,.; Schnegelsberg, P.N.J.; Stead, R.H.; Braun, T.; Arnold, H.-H.,; Jaenisch, R. MyoD and myf-5 is required for the formation of skeletal muscle. Cell 1993, 75, 1351–1359. [Google Scholar] [CrossRef]
- Gerber, A.N.; Klesert, T.R.; Bergstrom, D.A.; Tapscott, S.J. Two domains of MyoD mediate transcriptional activation of genes in repressive chromatin: A mechanism for lineage determination in myogenesis. Genes Dev 1997, 11, 436–450. [Google Scholar] [CrossRef]
- Londhe, P.; Davie, J.K. Sequential association of myogenic regulatory factors and E proteins at muscle-specific genes. Skelet Muscle 2011, 1, 14. [Google Scholar] [CrossRef]
- de Martin, X.; Sodaei, R.; Santpere, G. Mechanisms of Binding Specificity among bHLH Transcription Factors. Int. J. Mol. Sci. 2021, 22, 9150. [Google Scholar] [CrossRef]
- Sabourin, L.A.; Rudnicki, M.A. The molecular regulation of myogenesis. Clin Genet 2000, 57, 16–25. [Google Scholar] [CrossRef]
- Zammit, P.S. All muscle satellite cells are equal, but are some more equal than others? J Cell Sci 2008, 121, 2975–2982. [Google Scholar] [CrossRef]
- Günther, S.; Kim, J.; Kostin, S.; Lepper, C.; Fan, C.M.; Braun, T. Myf5-positive satellite cells contribute to Pax7-dependent long-term maintenance of adult muscle stem cells. Cell Stem Cell 2013, 13, 590–601, Erratum in: Cell Stem Cell 2013, 13, 769. [Google Scholar] [CrossRef]
- Zádor, E.; Bottka, S.; Wuytack, F. Antisense inhibition of myoD expression in regenerating rat soleus muscle is followed by an increase in the mRNA levels of myoD, myf-5 and myogenin and by a retarded regeneration. Biochim Biophys Acta Mol Cell Res 2002, 1590, 52–63. [Google Scholar] [CrossRef]
- Grubb, B.; Harris, J.; Schofield, I. Neuromuscular transmission of newly formed neuromuscular junctions in the regenerating soleus muscle. J Physiol 1991, 441, 405–421. [Google Scholar] [CrossRef]
- Abu Hatoum, O.; Gross-Mesilaty, S.; Breitschop, K.; Hoffman, A.; Gonen, H.; Ciechanover, A.; Bengal, E. Degradation of myogenic transcription factor myoD by the ubiquitin pathway in vivo and in vitro: Regulation by specific DNA binding. Mol Cell Biol 1998, 18, 5670–5677. [Google Scholar] [CrossRef]
- Bisbal, C.; Silhol, M.; Laubenthal, H.; Kaluza, T.; Carnac, G.; Milligan, L.; Le Roy, F.; Salehzada, T. The 2V–5Voligoadenylate/RNase L/RNase L inhibitor pathway regulates both myoD mRNA stability and muscle cell differentiation. Mol Cell Biol 2000, 20, 4959–4969. [Google Scholar] [CrossRef]
- Phillis, M.I.; Gyurko, R. Antisense oligonucleotides: New tools for physiology. News Physiol Sci 1997, 12, 99–105. [Google Scholar] [CrossRef]
- Tu, G.-C.; Cao, Q.-N.; Zhou, F.; Yedy, I. Tetranucleotide GGGA motif in primary RNA transcripts. Novel target site for antisense design. J Biol Chem 1998, 273, 25125–31. [Google Scholar] [CrossRef]
- Huang, Y.C.; Dennis, R.G.; Baar, K. Cultured slow vs. fast skeletal muscle cells differ in physiology and responsiveness to stimulation. Am J Physiol Cell Physiol 2006, 291, C11–C17. [Google Scholar] [CrossRef]
- Gorbe, A.; Becker, D.L.; Dux, L.; Stelkovics, E.; Krenacs, L.; Bagdi, E.; Krenacs, T. Transient upregulation of connexin43 gap junctions and synchronized cell cycle control precede myoblast fusion in regenerating skeletal muscle in vivo. Histochem Cell Biol 2005, 123, 573–583. [Google Scholar] [CrossRef] [PubMed]
- Gorbe, A.; Becker, D.L.; Dux, L.; Krenacs, L.; Krenacs, T. In differentiating prefusion myoblasts connexin43 gap junction coupling is upregulated before myoblast alignment then reduced in post-mitotic cells. Histochem Cell Biol 2006, 125, 705–716. [Google Scholar] [CrossRef] [PubMed]
- Gorbe, A.; Krenacs, T.; Cook, J.E.; Becker, D.L. Myoblast proliferation and syncytial fusion both depend on connexin43 function in transfected skeletal muscle primary cultures. Exp Cell Res. 2007, 313, 1135–1148. [Google Scholar] [CrossRef]
- Ishido, M.; Kasuga, N. Characteristics of the Localization of Connexin 43 in Satellite Cells during Skeletal Muscle Regeneration In Vivo. Acta Histochem Cytochem 2015, 48, 53–60. [Google Scholar] [CrossRef]
- Vitale, G.; Ferrantini, C.; Piroddi, N.; Scellini, B.; Pioner, J.M.; Colombini, B.; Tesi, C.; Poggesi, C. The relation between sarcomere energetics and the rate of isometric tension relaxation in healthy and diseased cardiac muscle. J Muscle Res Cell Motil 2021, 42, 47–57. [Google Scholar] [CrossRef]
- Xu, H.; Van Remmen, H. The SarcoEndoplasmic Reticulum Calcium ATPase (SERCA) pump: A potential target for intervention in aging and skeletal muscle pathologies. Skelet Muscle 2021, 11, 25. [Google Scholar] [CrossRef] [PubMed]
- Brandl, C.J.; Green, N.M.; Korczak, B.; MacLennan, D.H. Two Ca2+ ATPase genes: Homologies and mechanistic implications of deduced amino acid sequences. Cell 1986, 44, 597–607. [Google Scholar] [CrossRef]
- Brandl, C.J.; DeLeon, S. , Martin, D.R.; MacLennan, D.H. Adult forms of the Ca2+ ATPase of sarcoplasmic reticulum. Expression in developing skeletal muscle. J Biol Chem 1987, 262, 3768–3774. [Google Scholar] [CrossRef]
- Korczak, B.; Zarain-Herzberg, A.; Brandl, C.J.; Ingles, C.J.; Green, M.N.; MacLennan, D.H. Structure of the rabbit fast-twitch skeletal muscle Ca2+ ATPase gene. J Biol Chem 1988, 263, 4813–4819. [Google Scholar] [CrossRef]
- Zádor, E.; Vangheluwe, P.; Wuytack, F. The expression of the neonatal sarcoplasmic reticulum Ca2+ pump (SERCA1b) hints to a role in muscle growth and development. Cell Calcium. 2007, 41, 379–388. [Google Scholar] [CrossRef]
- Zador, E.; Dux, L.; Wuytack, F. Prolonged passive stretch of rat soleus muscle provokes an increase in the mRNA levels of the muscle regulatory factors distributed along the entire length of the fibers. J. Muscle Res. and Cell Mot 1999, 20, 395–402. [Google Scholar] [CrossRef]
- Szabó, A.; Wuytack, F.; Zádor, E. The effect of passive movement on denervated soleus highlights a differential nerve control on SERCA and MyHC isoforms. J Histochem Cytochem. 2008, 56, 1013–1022. [Google Scholar] [CrossRef]
- Zádor, E.; Kósa, M. The neonatal sarcoplasmic/endoplasmic reticulum calcium ATPase (SERCA1b): A neglected pump in scope. Pflugers Arch 2015, 467, 1395–1401. [Google Scholar] [CrossRef]
- Kósa, M.; Brinyiczki, K.; van Damme, P.; Goemans, N.; Hancsák, K.-; Mendler, L.; Zádor, E. The neonatal sarcoplasmic reticulum Ca(2+)-ATPase gives a clue to development and pathology in human muscles. J Muscle Res Cell Motil 2015, 36, 195–203. [Google Scholar] [CrossRef]
- Fodor, J.; Gomba-Tóth, A.; Oláh, T.; Zádor, E.; Tóth, Z.C.; Ioannis, I.; Molnár, B.; Kovács, I.; Csernoch, L. Alteration of sarcoplasmic reticulum Ca(2+) ATPase expression in lower limb ischemia caused by atherosclerosis obliterans. Physiol Int 2017, 104, 183–192. [Google Scholar] [CrossRef]
- Zhao, Y.; Ogawa, H.; Yonekura, S.; Mitsuhashi, H.; Mitsuhashi, S.; Nishino, I.; Toyoshima, C.; Ishiura, S. Functional analysis of SERCA1b, a highly expressed SERCA1 variant in myotonic dystrophy type 1 muscle. Biochim Biophys Acta 2015, 1852, 2042–2047. [Google Scholar] [CrossRef] [PubMed]
- Guglielmi, V.; Oosterhof, A.; Voermans, N.C.; Cardani, R.; Molenaar, J.P.; van Kuppevelt, T.H.; Meola, G.; van Engelen, B.G.; Tomelleri, G.; Vattemi, G. Characterization of sarcoplasmic reticulum Ca(2+) ATPase pumps in muscle of patients with myotonic dystrophy and with hypothyroid myopathy. Neuromuscul Disord 2016, 26, 378–385. [Google Scholar] [CrossRef] [PubMed]
- Mendler, L.; Szakonyi, G.; Zádor, E.; Görbe, A.; Dux, L.; Wuytack, F. Expression of sarcoplasmic/endoplasmic reticulum Ca2+ ATPases in the rat extensor digitorum longus (EDL) muscle regenerating from notexin-induced necrosis. J Muscle Res Cell Mot 1998b, 19, 777–785, *Authors contributing equally. [Google Scholar] [CrossRef]
- Kiss, G.; Zádor, E.; Szalay, J.; Somogyi, F.; Vér, A. Molecular forms of acetylcholinesterase in rat extensor digitorum longus and soleus muscles regenerating from notexin induced necrosis. J Muscle Res Cell Motil 2004, 25, 509–514. [Google Scholar] [CrossRef] [PubMed]
- Fenyvesi, R.; Rácz, G.; Wuytack, F.; Zádor, E. The calcineurin activity and MCIP1.4 mRNA levels are increased by innervation in regenerating soleus muscle. Biochem Biophys Res Com 2004, 320, 599–605. [Google Scholar] [CrossRef] [PubMed]
- Zádor, E.; Fenyvesi, R.; Wuytack, F. Expression of SERCA2a is not regulated by calcineurin or upon mechanical unloading in skeletal muscle regeneration. FEBS Letters 2005, 579, 749–752. [Google Scholar] [CrossRef]
- Serrano, A.L.; Murgia, M.; Pallafacchina, G.; Calabria, E.; Coniglio, P.; Lømo, T.; Schiaffino, S. Calcineurin controls nerve activity-dependent specification of slow skeletal muscle fibers but not muscle growth. Proc Natl Acad Sci U S A 2001, 98, 13108–13113. [Google Scholar] [CrossRef] [PubMed]
- McCullagh, K.J.; Calabria, E.; Pallafacchina, G.; Ciciliot, S.; Serrano, A.L.; Argentini, C.; Kalhovde, J.M.; Lømo, T.; Schiaffino, S. NFAT is a nerve activity sensor in skeletal muscle and controls activity-dependent myosin switching. Proc Natl Acad Sci USA 2004, 101, 10590–10595. [Google Scholar] [CrossRef]
- Launay, T.; Noirez, P.; Butler-Browne, G.; Agbulut, O. Expression of slow myosin heavy chain during muscle regeneration is not always dependent on muscle innervation and calcineurin phosphatase activity. Am J Physiol-Regul Integr Comp Physiol. 2006, 290, R1508–R1514. [Google Scholar] [CrossRef]
- Misquitta, C.M.; Chen, T.; Grover, A.K. Control of protein expression through mRNA stability in calcium signalling. Cell Calcium 2006, 40, 329–346. [Google Scholar] [CrossRef] [PubMed]
- Zádor, E. dnRas stimulates autocrine-paracrine growth of regenerating muscle via calcineurin-NFAT-IL-4 pathway. Biochem Biophys Res Com 2008, 375, 265–270. [Google Scholar] [CrossRef] [PubMed]
- Kósa, M.; Zádor, E. Transfection efficiency along the regenerating soleus muscle of the rat. Mol Biotechnol 2013, 54, 220–227. [Google Scholar] [CrossRef]
- Horsley, V.; Jansen, K.M.; Mills, S.T.; Pavlath, G.K. IL-4 acts as a myoblast recruitment factor during mammalian muscle growth. Cell 2003, 113, 483–494. [Google Scholar] [CrossRef]
- Shaikh, S. , Lee, E.; Ahmad, K.; Ahmad, S.S.; Chun, H.; Lim, J.; Lee, Y.; Choi, I. Cell Types Used for Cultured Meat Production and the Importance of Myokines. Foods 2021, 10, 2318. [Google Scholar] [CrossRef]
- Horsley, V.; Friday, B.B.; Matteson, S.; Kegley, K.M.; Gephart, J.; Pavlath, G.K. Regulation of the growth of multinucleated muscle cells by an NFATC2-dependent pathway. J Cell Biol 2001, 153, 329–338. [Google Scholar] [CrossRef]
- Ichida, M.; Finkel, T. Ras regulates NFAT3 activity in cardiac myocytes. J Biol Chem 2001, 276, 3524–3530. [Google Scholar] [CrossRef]
- Sanna, B.; Bueno, O.F.; Dai, Y.S.; Wilkins, B.J.; Molkentin, J.D. Direct and indirect interactions between calcineurin-NFAT and MEK1-extracellular signal-regulated kinase 1/2 signaling pathways regulate cardiac gene expression and cellular growth. Mol Cell Biol 2005, 25m, 865–78. [Google Scholar] [CrossRef]
- Zádor, E.; Owsianik, G.; Wuytack, F. Silencing SERCA1b in a few fibers stimulates growth in the entire regenerating soleus muscle. Histochem Cell Biol 2011, 135, 11–20. [Google Scholar] [CrossRef]
- Pan, Y.; Zvaritch, E.; Tupling, A.R.; Rice, W.J.; de Leon, S.; Rudnicki, M.; McKerlie, C.; Banwell, B.L.; MacLennan, D.H. Targeted disruption of the ATP2A1 gene encoding the sarco(endo)plasmic reticulum Ca2+ ATPase isoform 1 (SERCA1) impairs diaphragm function and is lethal in neonatal mice. The Journal of biological chemistry, 2003, 278, 13367–13375. [Google Scholar] [CrossRef]
- Anderson, D.M.; Anderson, K.M.; Chang, C.L.; Makarewich, C.A.; Nelson, B.R.; McAnally, J.R.; Kasaragod, P.; Shelton, J.M.; Liou, J.; Bassel-Duby, R.; Olson, E.N. A micropeptide encoded by a putative long noncoding RNA regulates muscle performance. Cell 2015, 160, 595–606. [Google Scholar] [CrossRef]
- Hansson, K.A.; Eftestøl, E.; Bruusgaard, J.C.; Juvkam, I.; Cramer, A.W.; Malthe-Sørenssen, A.; Millay, D.P.; Gundersen, K. Myonuclear content regulates cell size with similar scaling properties in mice and humans. Nature communications 2020, 11, 6288. [Google Scholar] [CrossRef]
- Bagley, J.R.; Denes, L.T.; McCarthy, J.J.; Wang, E.T.; Murach, K.A. The myonuclear domain in adult skeletal muscle fibres: Past, present and future. The Journal of physiology 2023, 601, 723–741. [Google Scholar] [CrossRef]
- Lin, J.G.; Kotha, P.; Chen, Y.H. ; Understandings of acupuncture application and mechanisms. American journal of translational research 2022, 14, 1469–1481. [Google Scholar]
- Rajula, H.S.R. , Mauri, M., & Fanos, V. Scale-free networks in metabolomics. Bioinformation, 2018, 14, 140–144. [Google Scholar] [CrossRef]
- Barabasi, A.L.; Albert, R. Emergence of scaling in random networks. Science, 1999, 286, 509–512. [Google Scholar] [CrossRef]
- Newlands, S. , Levitt, L.K., Robinson, C.S., Karpf, A.B., Hodgson, V.R., Wade, R.P., & Hardeman, E.C. Transcription occurs in pulses in muscle fibers. Genes & development, 1998, 12, 2748–2758. [Google Scholar] [CrossRef]
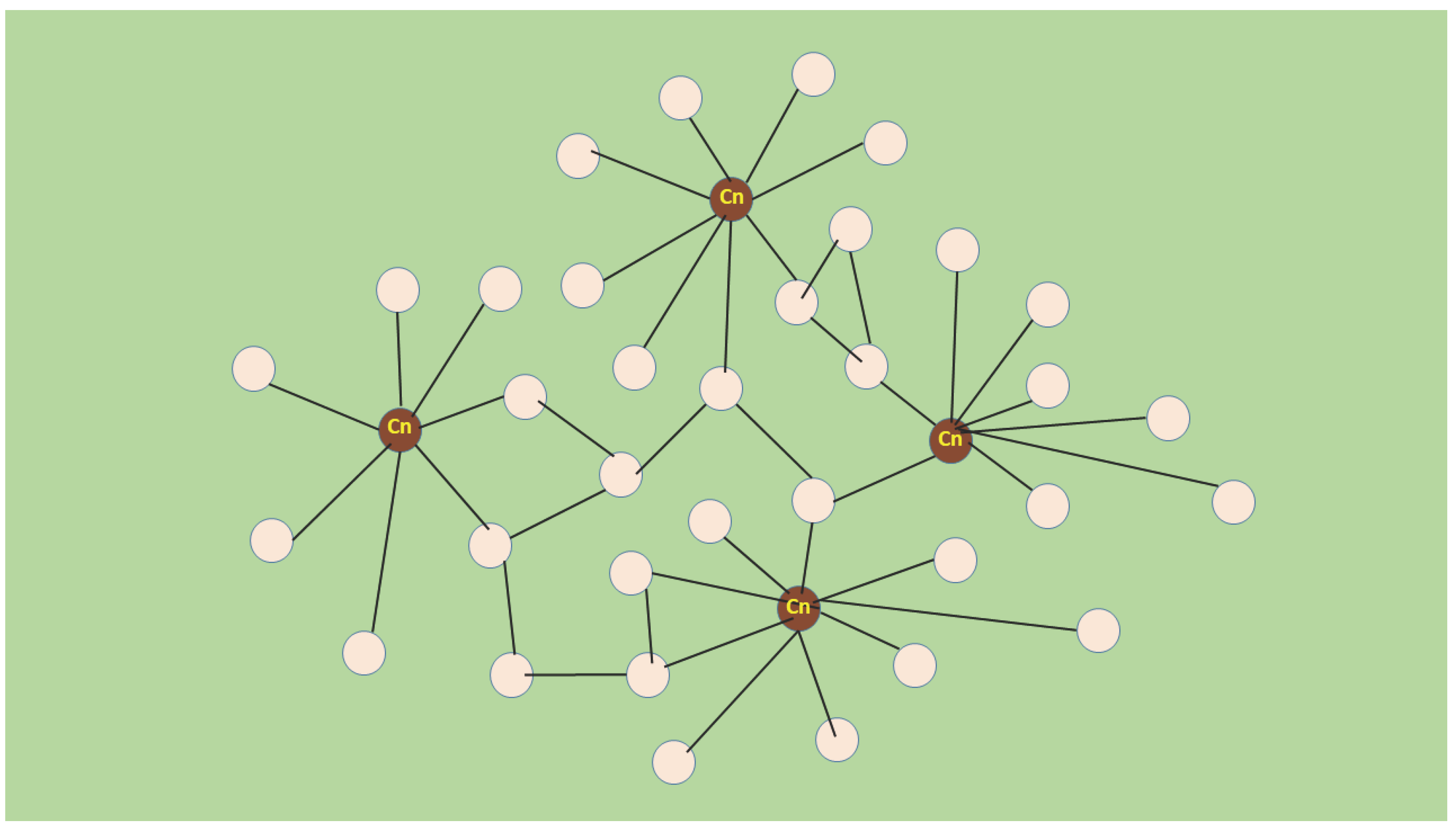
Figure 2.
Scale-free network in molecular-acupuncture-stimulated muscle regeneration. Myonuclear domains (Md) are depicted as nodes (illustrated with circles) and lines indicate links between the nodes. Md transforms into „hub” (a high grade node with many links) when transfected with dnRas or SERCA1b shRNA probably because of its increased calcineurin (Cn) activity. The links between hubs and nontransfected Mds might be formed by autocrine-paracrine factors like IL4. Cn indicates Md with hub-creating calcineurin activity. This representation does not intend to show the entire operation of the growth network, it only meant to reflect the driving process.
Figure 2.
Scale-free network in molecular-acupuncture-stimulated muscle regeneration. Myonuclear domains (Md) are depicted as nodes (illustrated with circles) and lines indicate links between the nodes. Md transforms into „hub” (a high grade node with many links) when transfected with dnRas or SERCA1b shRNA probably because of its increased calcineurin (Cn) activity. The links between hubs and nontransfected Mds might be formed by autocrine-paracrine factors like IL4. Cn indicates Md with hub-creating calcineurin activity. This representation does not intend to show the entire operation of the growth network, it only meant to reflect the driving process.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



