
Submitted:
27 February 2024
Posted:
27 February 2024
You are already at the latest version
Abstract
In agriculture, soil-borne fungal pathogens, especially Fusarium oxysporum strains, are posing a serious threat to efforts to achieve global food security. In the search for safer agrochemicals, silica nanoparticles (SiO2NPs) have recently been proposed as a new tool to alleviate pathogen damage including Fusarium wilt. Hollow mesoporous silica nanoparticles (HMSNs), a unique class of SiO2NPs, have been widely accepted as a desired pesticide carrier. However, their role in enhancing the disease resistance and the specific mechanism remain unknown. In this study, three sizes of HMSNs (19, 96 and 406 nm as HMSNs–19, HMSNs–96 and HMSNs–406, respectively) were synthesized and characterized to determine their effects on seed germination, seedling growth, and Fusarium oxysporum f. sp. phaseolus (FOP) suppression in cowpea roots by foliar spray using phenotypes, fresh biomass and disease progression as indicators. The results revealed that three HMSNs exhibited no adverse impacts on seed germination and tended to improve plant growth. Also, they exert their FOP suppression with a size- and concentration-dependent manner. HMSNs–406 possessed the best control effect at a concentration of 1000 mg/L showing an upto 40.00% decline in the disease severity. The Si(OH)4 control was also effective on FOP suppression at a lower concentration of 100 mg/L, whereas its higher concentrations exhibited obviously adverse impacts on FOP control, seed germination and plant growth. Moreover, we conformed that HMSNs posed their Fusarium wilt suppression in cowpea plant by activating SA (salicylic acid)-dependent SAR (systemic acquired resistance) responses rather than directly suppressing FOP. A higher level of SA content and elevated expression of its maker genes of PR-1 and PR-5 in HMSNs–406 treated cowpea roots provided substantial evidences of this mode of action. Other resistance-related genes, as well as defense-responsive enzymes, were also involved in the HMSNs-activated SAR pathway. Overall, for the first time, our results extended a new role of HMSNs as a potent elicitor to serve as a versatile alternative for plant disease protection of low cost, highly efficiency and sustainability.
Keywords:
hollow mesoporous silica nanoparticles
; cowpea resistance
; Fusarium wilt
; salicylic acid
; systemic acquired resistance
1. Introduction
Soil-borne fungal pathogens pose a vital threat to global food security causing up to 75% yield losses of major crops [1]. Fusarium oxysporum strains, a ubiquitous class of soil-borne pathogens ranking fifth in the top 10 plant pathogens of high importance, can infect hundreds of agricultural crop species including legume crops [2]. For instance, Fusarium wilt of cowpea (Vigna unguiculata) is a severe disease caused by Fusarium oxysporum f. sp. phaseoli (FOP) and Fusarium oxysporum f.sp. tracheiphilum [3], which contributes to annual yield losses of 30–100% around the world [4]. FOP was isolated as the preponderant species from diseased cowpea plants in China [4]. Chemical approaches and cultural practices have long been used to minimize the soil-borne disease outbreak. Unfortunately, neither of these strategies give consistent and effective results for Fusarium wilt control, with no detrimental effects on the environment and humans [1]. Thus, it seems urgent to develop new sustainable methods for effective control of Fusarium wilt of cowpea and many other crops.
Plants have evolved the innate immunity system that can be activated by both pathogen attack and elicitors to fend off potential pathogens. As an especially interesting form of induced resistance, systemic acquired resistance (SAR) is characterized by the spread of locally induced disease resistance to the whole plant [5]. SAR confers a long-lasting protection mechanism on pathogen-attacked plants. A key signal molecule of salicylic acid (SA) is responsible for the activation of pathogen-related (PR) genes and thus contributes to SAR [5,6]. SAR can be initiated by using resistance-inducing compounds, avoiding application of irreversible genetic modifications or fungicides of potential environmental risks [7]. This character makes SAR a promising alternative strategy for controlling soil-borne disease including Fusarium wilt [8,9,10].
Nanoagrochemicals, recently, are emerging to improve crop yield and global food security as an alternative and complementary tool for the conventional methods of fungicides, biological control, and cultural practices [11]. This new strategy is beginning to attract significant attention due to its advantages of better efficacy, reduced input, and lower eco-toxicity [11,12,13]. Silica nanoparticles (SiO2NPs) have been a topic of recent interest demonstrating diverse functions as an antimicrobial agent to directly restrain the virulence of plant pathogens [14,15,16], as an elicitor to boost plant innate immunity to alleviate abiotic and biotic stresses [7,9,17,18], or as delivery carriers to enhance the efficacy of pesticide, micronutrients, and elicitors [9,19]. SiO2NPs have been proposed as a controlled-release pool of bioavailable silicon (orthosilicic acid) that can enhance disease resistance without negative effects on the growth and yield of plants [7,9,12]. El-Shetehy et al. (2021) described the potential of SiO2NPs in enhancing the disease resistance of Arabidopsis thaliana against Pseudomonas syringae using the SAR manner that is similar to conventional Si products [7]. Rice plants were also reported to employ SiO2NPs to decline the incidence of Magnaporthe oryzae by the same manner [12]. For Fusarium wilt control, foliar spray of SiO2NPs significantly reduced the disease severity of Fusarium oxysporum f. sp. niveum in watermelons by modulating stress-related gene expression [9,10], and soil SiO2NPs application also suppressed Fusarium oxysporum greatly in maize [20]. Yet it is also not known whether SiO2NPs can enhance the disease suppression of Fusarium wilt in many other plants. Of note, much still needs to be understood to elucidate the specific mechanism of SiO2NPs for control of Fusarium wilt.
Hollow mesoporous silica nanoparticles (HMSNs), a unique class of SiO2NPs of large hollow cavities and intact porous shells, have been accepted as a desired carrier for controlled pesticide delivery due to their robust and low-cost synthesis with tunable physicochemical properties and high loading capacity [21,22,23]. Previous studies can offer insights into the roles of solid core SiO2NPs in enhancing the disease resistance of several specific plants at a single diameter scale range [7,10,12,15,18]. It remains unclear whether HMSNs can induce disease resistance in crop plants, whether their performance is related to the diameter scale and SAR induction. To these ends, in this work, three spherical HMSNs of different diameters were synthesized and characterized. Using V. unguiculata as the target crop plant, we investigated effects of the three HMSNs on seed germination, seedling growth, and Fusarium wilt suppression through foliar treatment. HMSNs of optimal particle size and concentration were then determined and foliarly applied to cowpea to explore the supposed SAR pathway for Fusarium wilt control in terms of endogenous SA synthesis, resistance-related gene expression and variations of defense-responsive enzyme activity. This is the first example known to the authors that reveals the role of HMSNs as a valuable plant immunity nano-inducer for Fusarium wilt control by means of the SA-modulated SAR pathway.
2. Results
2.1. Characterization of Hollow Mesoporous Silica Nanoparticles (HMSNs)
Three HMSNs of different sizes were synthesized to explore their roles in control of cowpea Fusarium wilt. Transmission electron microscopy (TEM) was employed to characterize their morphologies and size distributions. As shown in Figure 1a-c, representative HMSNs morphologies of hollow cavities and intact porous shells were observed from the TEM images of three HMSNs. TEM diameter analysis (Figure 1d-f) confirmed that three HMSNs had uniform hollow sphere structures with primary particle diameters of 19 ± 3 (HMSNs–19), 96 ± 9 (HMSNs–96) and 406 ± 26 nm (HMSNs–406), respectively. Hydrodynamic diameters, as well as zeta potentials of three HMSNs were also measured with results showing larger sizes compared with the TEM data and surface electronegativity (Table S1 ), consistent with previously reported results [7,9,10]. Characters of narrow particle size distribution of each size-controlled HMSNs (12–28, 70–125, and 350–455 nm, respectively), and overall wide range scale of three HMSNs covering the existing application scope of SiO2NPs for plant disease control [7,9,12,15,24], make them desirable materials to elucidate the roles of HMSNs for Fusarium wilt control.
2.2. Effect of HMSNs on Cowpea Seed Germination and Growth
Previous studies [10,12,16] have revealed that SiO2NPs are beneficial to seed germination and plant growth within appropriate concentration ranges. Thus, we first explored effects of three synthesized HMSNs at different concentrations of 100, 500, 1000 and 1500 mg/L on seed germination and growth using germination index (GI), the root and shoot fresh biomass of seedings as the indicators. As illustrated in Figure 2a-c, HMSNs–19 were demonstrated to improve seed germination at 1000 mg/L with a higher GI value compared with the control treatment while the other two HMSNs, as well as HMSNs–19 at other concentrations exhibited no side effects on cowpea seed germination. Si(OH)4 treatment, inconsistently, posed a concentration-dependent inhibition of seed germination with declining GI values along with the increase of concentrations (Figure 2d). Compared with the control, seed germination was significantly inhibited when the concentration of Si(OH)4 exceeded 500 mg/L. Besides, three HMSNs also exhibited no adverse impacts on the seeding growth showing no significant differences in the root and shoot biomass at the test concentrations relative to the controls (Figure 3a-c). However, the root biomass was significantly decreased by Si(OH)4 treatment at a concentration exceeding 500 mg/L and the shoot biomass was also significantly decreased by Si(OH)4 treatment at a high concentration of 1000 or 1500 mg/L, as compared with those of the controls (Figure 3d). Overall, the results indicate that three HMSNs pose no adverse effects on both the cowpea seed germination and seeding growth while Si(OH)4 can produce stress effects on both the seed germination and seeding growth at higher concentrations exceeding 500 or 1000 mg/L.
2.3. Toxicity of HMSNs to Fusarium oxysporum f. sp. phaseolus (FOP) In Vitro
To investigate the direct virulence activity of the three HMSNs against the cowpea wilt pathogen FOP, their inhibitory activities were determined on the FOP mycelial growth with results illustrated in Figure S1. Phenotypic observations showed that the inhibitory effect of each HMSNs on FOP was very weak showing an inhibition ratio less-than 20% at 2000 mg/L. The inhibition of FOP mycelial growth is positively correlated with the size and concentration of the HMSNs (Figure S1b). Overall, none of the three HMSNs pose a strong inhibitory effect directly on FOP mycelial growth allowing a better study to explore the efficacy of HMSNs on enhancing cowpea wilt resistance in vivo.
2.4. Effect of HMSNs on the Disease Incidence
The area-under-the-disease-progress curve (AUDPC) [9,10] was determined to evaluate the role of HMSNs in suppressing Fusarium wilt of cowpea plants. As shown in Figure 4a, HMSNs exert their benefits of disease suppression in a concentration- and size-dependent manner. HMSNs–19 were observed to have little effect on alleviating the disease incidence at both low and high doses with AUDPC values comparable to the infected control. The other two HMSNs, HMSNs–96 and HMSNs–406, exhibited similar capacities for significantly suppressing the disease incidence at the high concentration of 1000 mg/L, but scarcely working on the disease suppression at the low concentration of 100 mg/L. The disease progress was reduced by 23.13% and 40.00% for HMSNs–96 and HMSNs–406 treatment at 1000 mg/L (Figure 4a), respectively. Foliar application of SiO2 nanoparticles of a high concentration (1500 mg/L) was also previously described for the suppression of Fusarium wilt of watermelon [9]. An opposite effect, however, was observed for the Si(OH)4 treatment with reduced disease incidence at the low concentration of 100 mg/L, but a detrimental effect at a higher concentration of 1000 mg/L (Figure 4a). This similar concentration-dependent effect of Si(OH)4 was also observed on Arabidopsis thaliana infected by Pseudomonas syringae [7]. The difference of HMSNs and Si(OH)4 in the appropriate concentration can be attributed to previously described slow release effect of HMSNs to generate Si(OH)4 instead of direct interaction [7].
Plant biomass was also monitored in the greenhouse as another means of assessing the overall health of plants under diseased and healthy conditions with results shown in Figure 4c. Cowpea growth was inhibited after being transplanted in the FOP infected soil for 29 days, with the fresh biomass of shoots and roots decreased by 14.63% and 20.78%, respectively. An overall trend of HMSNs was observed to promote the growth of cowpea plants showing increased fresh biomass of shoots and roots relative to the infected control plants. The three HMSNs significantly enhanced the cowpea shoot and/ or root biomass at a concentration of 100 mg/L as compared to the infected control, but did not affect disease incidence. This similar character was also previously observed for sulfur nanoparticles in tomato Fusarium wilt control [25]. Of note, treatment with a concentration of 1000 mg/L of HMSNs–406 led to significant increases of both cowpea shoot and root biomass by 31.02% and 87.67% relative to the infected control, respectively. HMSNs–96 of a concentration of 1000 mg/L only significantly improved the fresh biomass of cowpea roots. Si(OH)4 of a high concentration of 1000 mg/L significantly lowered the fresh biomass of shoots, Whereas its lower concentration of 100 mg/L improved the cowpea shoot and root biomass. This result may attribute to high-dose Si(OH)4 induced stress to plant [7]. Phenotypic images of cowpea shoots, longitudinal-cut stems and roots were presented in Figure 4b under HMSNs–406 (1000 mg/L) or Si(OH)4 (100 mg/L) treatment. Taken together, HMSNs–406 of a concentration of 1000 mg/L can be an optimal candidate for cowpea Fusarium wilt control since their both significant disease suppression and plant growth improvement.
2.5. Effect of Foliar HMSNs–406 Application on Cowpea Root Salicylic Acid (SA) Content
Since the findings that HMSNs–406 posed no obvious direct toxicity on FOP (Figure S1), whereas foliar HMSNs–406 treatment could significantly suppress the FOP incidence present in the roots (Figure 4a), we considered this role as a kind of induced resistance of systemic acquired resistance (SAR) characterized by the spread of locally induced disease resistance to the whole plant [5,7,12]. The plant hormone SA plays a core regulatory role in plant SAR as a key signaling compound [5,7,12]. Recent findings have revealed that the protective effect of SiO2NPs is mediated through activation of SA-dependent plant immune responses called SAR [7,12]. Thus, we measured SA content in cowpea roots to identify the primary pathway of SAR induction by HMSNs–406. As shown in Figure 5a, foliar application of HMSNs–406 (1000 mg/L) and Si(OH)4 (100 mg/L) increased SA content in cowpea roots by 4.3 and 3.4 folds, respectively, compared with the infected control. These findings indicate that the SAR activation in cowpea by HMSNs–406 or Si(OH)4 application is clearly SA dependent.
2.6. Gene Expression Changes after HMSNs–406 Exposure
Pathogenesis-related (PR) genes including PR-1 and PR-5 are commonly used as markers for salicylic acid-mediated activation of SAR [7,26]. To further support SA dependent SAR induced by foliar treated HMSNs–406, we next quantified the expression of PR-1 and PR-5 in FOP infected cowpea roots. Similar to treatment with Si(OH)4 (100 mg/L), treatment with HMSNs-406 (1000 mg/L) resulted in 1.97- and 9.38-fold increases of PR-1 (Figure 5b) and PR-5 (Figure 5c) in the transcript abundance, respectively, relative to the infected control. Non-expressed pathogen-related gene 1 (NPR-1) have been reported to be a receptor for salicylic acid and positively regulate the expression of PR genes [26]. Thus, NPR-1 expression was also measured to clarify whether this upstream gene participated in the HMSNs-406 activated SAR. HMSNs–406 (1000 mg/L) exposure boosted the expression level of NPR-1 by 1.62 folds in the cowpea roots compared with the infected control, so did the Si(OH)4 (100 mg/L) exposure by 1.76 folds (Figure 5d). Phenylalanine ammonia lyase gene (PAL), an antioxidant defence-related enzyme gene, is also associated with the response to pathogen-induced stress [25,27]. In cowpea infected roots, there was significant upregulation of PAL (8.54 folds) with HMSNs–406 foliar exposure (Figure S2), consistent with those reported previously [25,27]. These findings indicate that HMSNs–406, similar to Si(OH)4, can activate SA-dependent defence reactions by SAR in FOP infected cowpea plants.
2.7. Changes in Defence-Related Enzyme Activity
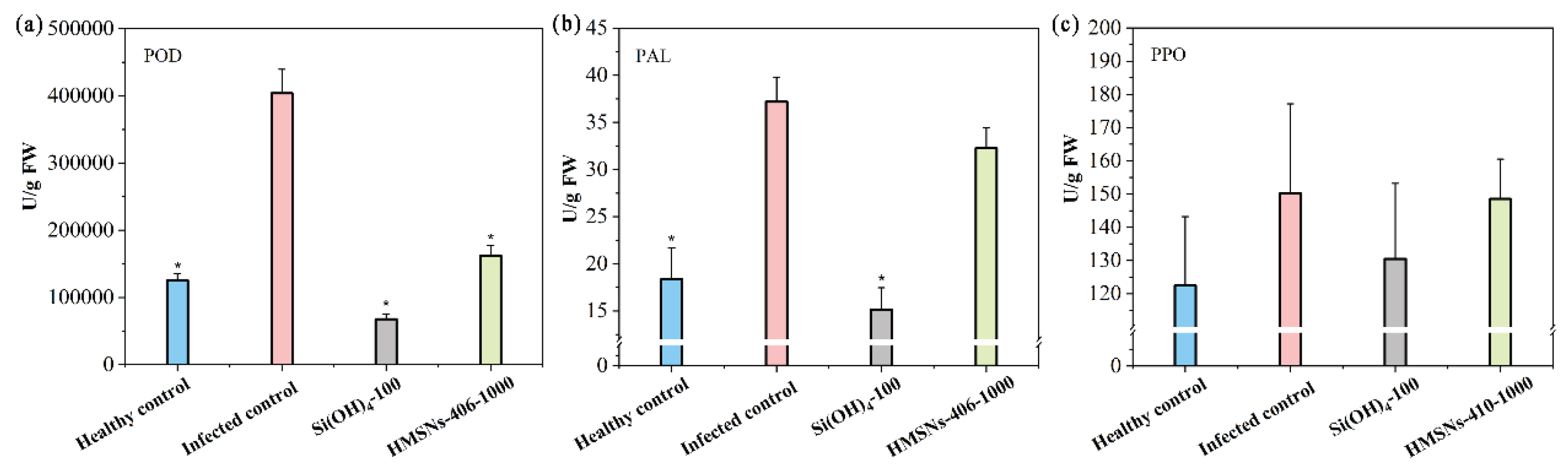
We further investigated the effects of HMSNs–406 on the levels of disease-related enzymes in cowpea roots to characterize the impact on cowpea SAR. As shown in Figure 6, FOP significantly increased the peroxidase (POD) activity by 2.22 folds in cowpea roots. The POD activity in the foliar HMSNs–406 treated cowpea roots was significantly decreased by 59.81% as compared with the infected control. Also, Si(OH)4 treatment led to a POD activity decline of 83.23%. This is likely because POD was reverted to the level of the healthy control after the suppression of pathogen activity and damage by HMSNs–406 and Si(OH)4 application [25,27]. The change trends in the activity of the other two antioxidant enzymes, PAL and PPO (polyphenol oxidase), were similar to those for POD in FOP-infected roots, despite lesser difference magnitude between the treatments (Figure 6).
3. Discussion
HMSNs exert their Fusarium wilt suppression in cowpea plants with a size- and concentration-dependent manner. SiO2NPs have been a hot topic of recent interest as an emerging alternative for plant disease protection [14,15,17]. However, their character difference in size, shape, surface chemistry, etc. resulted in differences in findings on the effects and mechanisms in alleviating pathogen stress [15,16,17]. Particle size, a core character determining many unique properties of nanoparticles, can affect the role of SiO2NPs in plant disease control, yet scarce information is available due to different target pathogens and host plants used in previous studies [15]. SiO2NPs of the size range 30–60 nm was proved to reduce Fusarium wilt severity in watermelon [9,10]. Other size ranges of 5–100 nm were also reported to enhance disease resistance in multiple plants [15,18,28]. SiO2NPs of the size exceeding 100 nm, however, is rarely discussed, although they can also transfer in plants via leaves with size up to 300 nm [29,30,31]. HMSNs are a unique class of SiO2NPs that widely accepted as a desired pesticide carrier. It is also not known whether HMSNs can induce resistance in plants, and whether their performance will be size dependent. Thus, in this study, three sizes (19, 96, and 406 nm) of HMSNs were synthesized and characterized to determine their roles in suppressing FOP infection in cowpea by foliar application using indicators of phenotypes, fresh biomass and disease progression. The results showed that the protective effect of HMSNs was positively related their concentrations and particle sizes, where the 406-nm sized HMSNs (HMSNs–406) were the best at 1000 mg/L for cowpea Fusarium wilt control through foliar spray, with upto 40.00% reduced disease incidence (Figure 4a) and enhanced biomass of cowpea shoots and roots (Figure 4c). This optimal concentration of HMSNs is within the expected range described in previous studies [7,9]. Interestingly, HMSNs of larger size exhibited a better capacity for Fusarium wilt suppression in cowpea. This seems to be contradictory to the general knowledge of high efficacy retained by smaller nanoparticles [32,33]. El-Shetehy et al. (2021) proposed a mode of action of leaf-applied SiO2NPs: SiO2NPs can enter into the spongy mesophyll space via the stomata to activate plant immunity responses probably by slow release of Si(OH)4, closure of the stomata, and interaction with their adjacent plant cells [7]. Thus, we can deduce that variances in both size-related dissolution rate of HMSNs in the leaf tissue and interactions of intact HMSNs with mesophyll cells may explain the size effect. The specific mechanisms need to be elucidated in follow-up studies.
The resistance of cowpea roots to the FOP through foliar HMSNs treatment attributes to the SA-dependent SAR pathway. Currently, three modes of action of direct restraint [14], resistance induction [7,12,18] and the two cooperation [34] have been involved in the functions of SiO2NPs on plant disease control. Induction of resistance is the most commonly reported mechanism of SiO2NPs [15] that was also demonstrated to be suitable for HMSNs in this study since HMSNs reduced the FOP severity in cowpea plants, but exhibited no direct toxic effect on FOP in vitro. Salicylic acid (SA) is a key plant hormone for plant immunity mediation including SAR. Previous studies have found that SiO2NPs induced plant resistance to several pathogens largely rely on SA signaling since not only the enhanced level of SA content [12,18], but also the upregulation of SA-responsive genes [7,12]. Our results revealed that foliar spray of HMSNs, a unique type of SiO2NPs of 406 nm, enhanced cowpea resistance to FOP at 1000 mg/L through SA-activated SAR as well, showing higher SA content and expression levels of SA marker genes of PR-1 and PR-5 in FOP infected cowpea roots (Figure 5). El-Shetehy et al. (2021) found that SiO2NPs can induce plant SAR between local leaves and systemic leaves in A. thaliana, while Du et al. (2022) described the role of SiO2NPs on stimulating rice SAR spreading from roots to leaves [12]. In this study, we further extended the SAR range to that foliar application could enhance cowpea root leaves resistance to FOP.
HMSNs have great potential to be developed as a new type of green pesticide for plant disease protection. As a desired nanocarrier, HMSNs have been widely used to develop various delivery vehicles of pesticides, nutrients, and biomacromolecules of functional proteins and nucleic acids, to improve their efficacy [21,22,23]. Previous studies, however, mainly focused on the exceptional properties of HMSNs as nanocarriers. In this study, for the first time, we demonstrated the potential of HMSNs as a nanoscaled elicitor to enhance Fusarium wilt resistance in cowpea plants with the appropriate size and concentration. Moreover, our results also confirmed that HMSNs proved to be safer for the plant compared with direct Si(OH)4 application since HMSNs had no adverse effects on cowpea seed germination (Figure 2) and plant growth (Figure 3) consist consistent with previous studies in other crops [7,9,12,18]. We also found that the mechanism of HMSNs was involved in activating SA-dependent SAR that can not only avoid either application of irreversible genetic modifications or fungicides of potential environmental risks [7] but also remove the space limitation of pesticide application. Combining our results and results of previous studies, a new type of versatile pesticide based on HMSNs can be developed to achieve the synergistic interaction of smart delivery and resistance induction for plant disease control efficiently.
4. Materials and Methods
4.1. Materials
Tetraethyl silicate (TEOS, 99%), cetyltrimethylammonium bromide (CTAB, 98%), Pluronic®F-108 (14600 Da), homotrimethylbenzene (TMB, 98%), potassium silicate (Si(OH)4-28%), and dimethyldimethoxysilane (MSDS, 98%) were purchased from Shanghai Aladdin Biochemical Technology Co., Ltd (China). Anhydrous ethanol (99%), sodium carbonate (Na2CO3, 98%), hydrochloric acid (37 w%) and aqueous ammonia solution (25–28 wt%) were obtained from Sun Chemical Technology Co., Ltd. (Shanghai, China). Spray adjuvant Mairun was purchased from BeijingGrand AgroChem Co., Ltd. (China). All other chemicals were commercially analytical grade products, unless otherwise specified. Dialysis bags (MWCO=14000 Da) were purchased from Beijing Solarbio Science & Technology Co., Ltd. (China).
The target pathogenic fungus of Fusarium oxysporum f. sp. phaseolus (FOP) in this work was isolated from infected cowpea plant roots collected from Ledong area of Hainan Province of China, and stored in our laboratory. This FOP strain was activated on PDA (potato dextrose agar) plates for three times prior to use.
The cowpea cultivar Fengjiang1, widely cultivated in southern China, was selected as the target plant for the FOP suppression test. The potting soil mix was prepared by mixing a growth media of pindstrup plus with equal weight of cultivated soil collected from the cowpea field.
4.2. Synthesis of Hollow Mesoporous Silica Nanoparticles (HMSNs)
Two of 406 nm and 96 nm sized HMSNs were synthesized according to a previously published Na2CO3 etching method [35] with some modifications. Solid core SiO2 spheres of 250 nm (sSiO2–250) and 50 nm (sSiO2–50) were first synthesized following modified Stӧber methods [35,36] by varying the inputs of TEOS, ultrapure water, anhydrous ethanol and NH3-H2O, as well as the reaction temperature and time. 406 nm sized HMSNs (HMSNs–406) were then prepared suing sSiO2–250 as the core on which a layer of mesoporous CTAB-templated shell formed followed by selective etching off the core sSiO2 and removal of the CTAB surfactant successively. 96 nm sized HMSNs (HMSNs–96) were also obtained according to the same procedure. The detailed procedures for synthesis of HMSNs–406 and HMSNs–96 can be seen in the Supplementary Materials.
HMSNs of 19 nm size (HMSNs–19) were prepared according to a previously reported method [37] using Pluronic®F-108 as the template, and tetraethoxysilane as a silica source under acidic conditions. The detailed synthetic procedure can be found in the Supplementary Materials.
4.3. Transmission Electron Microscopy (TEM) Observation
Prior to TEM observation, each HMSNs sample was diluted to 1 mg/mL in ethanol and sonicated for 15 min to ensure dispersity. Afterward, 2 μL of the suspension was dropped onto a carbon coated copper grid (200 mesh, Beijing Zhongjingkeyi Technology Co., Ltd, China) and air-dried overnight. The morphology of three HMSNs were then observed with a JEOL JEM-2100 instrument (Tokyo, Japan) at an accelerating voltage of 120 kV. To determine the size of the HMSNs, the images were analysed using ImageJ [38] (https://imagej.nih.gov/ij/) and the diameters of at least 500 randomly selected nanoparticles were measured.
4.4. Dynamic Light Scattering (DLS) and Zeta Potential Measurements
To measure the hydrodynamic diameters and surface potentials of three HMSNs, Each nanoparticle sample was suspended in water at 0.5 mg/mL and sonicated for 15 min to ensure uniform dispersion. A Malvern Instrument of Zetasizer Nano ZS90 (Worcestershire, UK) was then used for DLS and zeta potential measurements.
4.5. Monitoring Seed Germination and Seedling Growth after HMSNs Treatment
Cowpea seeds were soaked into a 75% ethanol-water solution (v/v) for 5-min surface sterilization and washed with sterile water fully to remove the surface alcohol. Then six of the sterilized seeds were set into a 9-cm petri dish with a piece of filter paper (9 cm in diameter) immersed in 10 mL of HMSNs suspension at a given concentration of 100, 500, 1000 or 1500 mg/L. Sterile water was used as the blank control. Each treatment was repeated ten times. All the cowpea seeds were incubated under identical conditions (28 ± 1°C, 75 ± 10% relative humidity, and a12:12 light/dark photoperiod). Seeds began to sprout at 12 hours after treatment and were counted every 2 hours for 24 hours during which over 80% of seeds sprouted in the control groups. The germination index (GI) of cowpea was calculated by the equation (1) [39] as followed:
where Gt represents the number of germinations on the hour, and Dt represents the number of germination hour.
Germination index = ∑Gt/Dt
The root and shoot fresh biomass of seedlings was also measured after 7 days of treatment.
4.6. Mycelial Growth Inhibition Test
The agar dilution method [40] was used to determine the direct mycelial growth inhibition of three synthesized HMSNs against F. oxysporum f. sp. phaseoli (FOP) at concentrations of 100, 1000 and 2000 mg/L. The details for this experiment can be found in the Supplementary Materials.
4.7. Pot Experiments
Pot experiments were carried out according to the previously described processes [9,25] with some modifications. FOP was inoculated into Potato Dextrose Liquid Medium (PDB) for 7 days at 25°C and 120 rpm. Afterwards, FOP conidia were harvested by filtration through three layers of sterile gauze followed by centrifugation (3000 g, 5 min). The precipitated conidia were re-suspended in sterile water and diluted to 1×106 conidia/mL. Then the conidia suspension was mixed with the potting soil mix to prepare the infected soil (1×106 conidia/g dry soil). The potting soil mix supplied with sterile water was used as the noninfected soil.
Cowpea seeds were soaked in sterile water for 24 h for pre-germination and then planted in the noninfected soils. When the plants reached the three- to four-leaf stage, seedlings of uniform size were transplanted in the infected soil for foliar nanoparticle exposure. The HMSNs dispersions for foliar spray were prepared by adding 0.1% of Pluronic®F-108 and 0.1% of Mairun as dispersive adhesives to make the final concentrations of 100 and 1000 mg/L. 5 mL of a HMSNs dispersion was sprayed on the leaf surface of each seedling and this treatment was repeated twice at 5-d intervals after the first application. The infected controls, as well as the healthy controls in the uninfected soil, were similarly established by spraying with distilled water supplied with the same amounts of dispersive adhesives. Foliarly spraying a potassium silicate solution of identical concentration to HMSNs on the seedlings in the infected soil served as the Si(OH)4 treatments. Each treatment contained 12 seedlings and was repeated for three times. All the pots were set in a greenhouse for the duration of the study.
The cowpea plants were evaluated for the severity of Fusarium wilt at 14, 21, and 29 days of post-transplanting in the infected soil, using a 0 to 4 scale monitoring both the external leaf and internal vascular symptoms modified from previous studies [41,42,43]. The detailed grading standards can be seen in the Table S2. The disease progress on the cowpea plants was exhibited by the area-under-the-disease-progress curve (AUDPC) of the cumulative severity ratings plotted as a function of time [10,44]. AUDPC was calculated using the previously described trapezoid rule (equation 2) [10,44]:
where, in this equation, Yi and Y(i+1) are the disease severity ratings at the time of ti and its adjacent time interval for rating, repectively.
After 29 days, the pot experiments were terminated and the root and shoot fresh biomass were determined.
4.8. Salicylic Acid (SA) Measurement
Cowpea plants were exposed to 1000 mg/L of HMSNs–406 by foliar application according to the procedure as described for pot experiments. Potassium silicate and water were used as the Si(OH)4 control and blank control, respectively. The plant roots of different treatments were collected at 29 days of post-transplanting in the infected soil and stored in triplicate at –80ºC for subsequent tests of salicylic acid content, RNA extraction and enzyme activity. Quantitative determination of overall SA of free and conjugated forms in cowpea roots was performed according to a previously described ultra-performance liquid chromatography (UPLC) method [45,46] using a Rigol L3000 UPLC (Bejing, China) with fluorescence detection. The detailed extraction procedures of free and hydrolysable SA, as well as the detecting parameters and quantitative method for UPLC fluorescence detection, can be found in the Supplementary Materials.
4.9. Gene Expression Variations
Quantitative real-time PCR (qRT-PCR) assay was employed to monitor changes in the expression of select resistance-related genes after HMSNs treatment. Cowpea roots collected for SA measurement were also used for total RNA extraction to observe resistance-related gene expression variations. After being grounded in liquid nitrogen, 150 mg of plant tissue homogenates were collected for total RNA extraction using a TaKaRa MiniBEST Plant RNA Extraction Kit (Dalian, China). The concentration and quality of the extracted RNA samples were assessed using a Thermo Scientific Nanodrop Lite Spectrophotometer (Wilmington, USA). The cDNA was synthesized from the extracted RNA samples using a QuantiTect Reverse Transcription kit (Qiagen, Germany). The eukaryotic elongation factor gene of EF1b was used as the internal control. The optimal primers for gene amplification and PCR programs are present in Table S3 and S4, respectively. qRT-PCR was then performed on a Bio-Rad CFX96 Touch Real-Time PCR Detection System (Hercules, USA) using SYBR Green as the fluorescent intercalating dye. The relative expression of genes was calculated by the 2-ΔΔCt method.
4.10. Defense-Responsive Enzyme Activity Measurement
Three defense-responsive enzymes of phenylalanine ammonia lyase (PAL), peroxidase (POD) and polyphenol oxidase (PPO) were selected as the target enzymes and their activities were determined according to the instructions of relevant commercial assay kits from Beijing Solarbio technology Co., Ltd. (China). Three replicates were performed for each experiment.
4.11. Statistical Analysis
Data are represented as mean ± standard deviation. In the experiments involving cowpea plants, one-way ANOVA was carried out by SPSS 23.0 software (SPSS Inc., USA) to test for statistical significance (P < 0.05) for comparing the response of plants treated with different HMSNs.
5. Conclusions
In the present study, we conformed the role of HMSNs in enhancing cowpea resistance to F. oxysporum f. sp. phaseoli with a size- and concentration-dependent manner by foliar application. The resistance mechanism induced by HMSNs attributes to the SA-dependent SAR that can be used as a promising alternative strategy for plant disease control. As a desired carrier extensively discussed previously, our results revealed the new function of HMSNs as an emerging elicitor without adverse effects on plant growth and yield to extend their application as versatile alternative for plant disease protection of low cost, highly efficiency and sustainability.
Supplementary Materials
The following supporting information can be downloaded at: www.mdpi.com/xxx/s1, Figure S1: Toxicity of three HMSNs to Fusarium oxysporum f. sp. phaseolus (FOP) in vitro; Figure S2: RT-qPCR analysis of the gene expression of an antioxidant defence-related enzyme gene PAL in cowpea roots; Table S1: Zeta potential, PDI, and average particle size of three HMSNs; Table S2: Standards of disease severity grading for Fusarium wilt in cowpea plant; Table S3: Primers for qRT-PCR assay; Table S4: Amplification program for qRT-PCR assay.
Author Contributions
Conceptualization, S.Z., Y.L., and Y.Z.; data curation, S.Z. and Y.Z.; formal analysis, Y.G. and L.W.; funding acquisition, S.Z. and Y.Z.; investigation, C.D., C.C., J.W. and M.Q.; methodology, C.D., Y.Z., and S.Z.; project administration, S.Z. and Y.Z.; resources, L.W. and Y.Z.; supervision, S.Z., Y.L., and Y.Z.; validation, C.D., L.W. and Y.L.; visualization, C.D., C.C., J.W. and M.Q.; writing—original draft, C.D., Y.Z. and Y.G.; writing—review and editing, C.D., C.C., J.W., M.Q. and L.W. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Hainan Natural Science Foundation Youth Fund Project (320QN198) and Scientific Research Foundation of Hainan University (KYQD(ZR)20024, KYQD(ZR)20025).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The datasets generated and analyzed during the current study are available from the corresponding authors upon reasonable request.
Acknowledgments
The authors would like to thank Jian Hu for his methodology assistance during this research.
Conflicts of Interest
The authors declare that there is no conflict of interest.
References
- Dutta, P.; Kumari, A.; Mahanta, M.; Upamanya, Gunadhya, K.; Heisnam, P.; Borua, S.; Kaman, P. K.; Mishra, A. K.; Mallik, M.; Muthukrishnan, G.; Sabarinathan, K. G.; Puzari, K. R.; Vijayreddy, D. Nanotechnological approaches for management of soil-borne plant pathogens. Front. Plant Sci. 2023, 14, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Sampaio, A. M.; Araújo, S. d. S.; Rubiales, D.; Vaz Patto, M. C. Fusarium Wilt Management in Legume Crops. Agronomy 2020, 10, 1073. [Google Scholar] [CrossRef]
- Cunha, W. L.; Andrade Leonel, M. R. d. S.; de Sousa, A. P. B.; Lima, M. A. d. S.; Santos, A. d. J. M.; Nascimento, G. M. d.; Mochel, G. d. M. S.; Silva, S. L. O.; Nobre, C. P.; Rodrigues, A. A. C.; Nascimento, I. d. O. Arbuscular mycorrhizal fungi can reduce severity of fusariosis (Fusarium oxysporum) in cowpea plants (Vigna unguiculata (L.) Walp.). Rhizosphere 2023, 28, 100795. [Google Scholar] [CrossRef]
- Hao, Y.; Li, Y.; Ping, X.; Yang, Q.; Mao, Z.; Zhao, J.; Lu, X.; Xie, B.; Yang, Y.; Ling, J. The Genome of Fusarium oxysporum f. sp. phaseoli Provides Insight into the Evolution of Genomes and Effectors of Fusarium oxysporum Species. Int. J. Mol. Sci. 2023, 24, 963. [Google Scholar] [CrossRef] [PubMed]
- Durrant, W. E.; Dong, X. Systemic acquired resistance. Annu. Rev. Phytopathol. 2004, 42, 185–209. [Google Scholar] [CrossRef]
- Mauch, F.; Mauch-Mani, B.; Gaille, C.; Kull, B.; Haas, D.; Reimmann, C. Manipulation of salicylate content in Arabidopsis thaliana by the expression of an engineered bacterial salicylate synthase. The Plant Journal 2001, 25, 67–77. [Google Scholar] [CrossRef]
- El-Shetehy, M.; Moradi, A.; Maceroni, M.; Reinhardt, D.; Petri-Fink, A.; Rothen-Rutishauser, B.; Mauch, F.; Schwab, F. Silica nanoparticles enhance disease resistance in Arabidopsis plants. Nat. Nanotechnol. 2021, 16, 344–353. [Google Scholar] [CrossRef]
- Ahammed, G. J.; Yang, Y. Mechanisms of silicon-induced fungal disease resistance in plants. Plant Physiol. Biochem. 2021, 165, 200–206. [Google Scholar] [CrossRef]
- Kang, H.; Elmer, W.; Shen, Y.; Zuverza-Mena, N.; Ma, C.; Botella, P.; White, J. C.; Haynes, C. L. Silica Nanoparticle Dissolution Rate Controls the Suppression of Fusarium Wilt of Watermelon (Citrullus lanatus). Environ. Sci. Technol. 2021, 55, 13513–13522. [Google Scholar] [CrossRef]
- Buchman, J. T.; Elmer, W. H.; Ma, C.; Landy, K. M.; White, J. C.; Haynes, C. L. Chitosan-Coated Mesoporous Silica Nanoparticle Treatment of Citrullus lanatus (Watermelon): Enhanced Fungal Disease Suppression and Modulated Expression of Stress-Related Genes. ACS Sustainable Chem. Eng. 2019, 7, 19649–19659. [Google Scholar] [CrossRef]
- Elmer, W.; White, J. C. The Future of Nanotechnology in Plant Pathology. Annu. Rev. Phytopathol. 2018, 56, 111–133. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Liu, B.; Zhao, T.; Xu, X.; Lin, H.; Ji, Y.; Li, Y.; Li, Z.; Lu, C.; Li, P.; Zhao, H.; Li, Y.; Yin, Z.; Ding, X. Silica nanoparticles protect rice against biotic and abiotic stresses. J. Nanobiotechnology 2022, 20, 197. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Lu, L.; Wang, A.; Zhang, H.; Huang, M.; Wu, H.; Xing, B.; Wang, Z.; Ji, R. Nano-Biotechnology in Agriculture: Use of Nanomaterials to Promote Plant Growth and Stress Tolerance. J. Agric. Food Chem. 2020, 68, 1935–1947. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Guo, X.; Zhang, B.; Nie, D.; Rao, W.; Zhang, D.; Lü, J.; Guan, X.; Chen, Z.; Pan, X. Mesoporous Silica Nanoparticles Induce Intracellular Peroxidation Damage of Phytophthora infestans: A New Type of Green Fungicide for Late Blight Control. Environ. Sci. Technol. 2023, 57, 3980–3989. [Google Scholar] [CrossRef]
- Wang, L.; Ning, C.; Pan, T.; Cai, K. Role of Silica Nanoparticles in Abiotic and Biotic Stress Tolerance in Plants: A Review. Int. J. Mol. Sci. 2022, 23, 1947. [Google Scholar] [CrossRef] [PubMed]
- Rajput, V. D.; Minkina, T.; Feizi, M.; Kumari, A.; Khan, M.; Mandzhieva, S.; Sushkova, S.; El-Ramady, H.; Verma, K. K.; Singh, A.; Hullebusch, E. D. V.; Singh, R. K.; Jatav, H. S.; Choudhary, R. Effects of Silicon and Silicon-Based Nanoparticles on Rhizosphere Microbiome, Plant Stress and Growth. Biology 2021, 10, 791. [Google Scholar] [CrossRef] [PubMed]
- Rivero-Montejo, S. D. J.; Vargas-Hernandez, M.; Torres-Pacheco, I. Nanoparticles as Novel Elicitors to Improve Bioactive Compounds in Plants. Agriculture 2021, 11, 134. [Google Scholar] [CrossRef]
- Deng, Q.; Huang, S.; Liu, H.; Lu, Q.; Du, P.; Li, H.; Li, S.; Liu, H.; Wang, R.; Huang, L.; Sun, D.; Wu, Y.; Chen, X.; Hong, Y. Silica nanoparticles conferring resistance to bacterial wilt in peanut (Arachis hypogaea L.). Sci. Total Environ. 2024, 915, 170112. [Google Scholar] [CrossRef]
- Kong, X.P.; Zhang, B.H.; Wang, J. Multiple Roles of Mesoporous Silica in Safe Pesticide Application by Nanotechnology: A Review. J. Agric. Food Chem. 2021, 69, 6735–6754. [Google Scholar] [CrossRef]
- Suriyaprabha, R.; Karunakaran, G.; Kavitha, K.; Yuvakkumar, R.; Rajendran, V.; Kannan, N. Application of silica nanoparticles in maize to enhance fungal resistance. IET Nanobiotechnol. 2014, 8, 133–137. [Google Scholar] [CrossRef]
- Gao, Y.; Xiao, Y.; Mao, K.; Qin, X.; Zhang, Y.; Li, D.; Zhang, Y.; Li, J.; Wan, H.; He, S. Thermoresponsive polymer-encapsulated hollow mesoporous silica nanoparticles and their application in insecticide delivery. Chem. Eng. J. 2020, 383, 123169. [Google Scholar] [CrossRef]
- Yang, L.; Kaziem, A. E.; Lin, Y.; Li, C.; Tan, Y.; Huang, S.; Cheng, D.; Xu, H.; Zhang, Z. Carboxylated β-cyclodextrin anchored hollow mesoporous silica enhances insecticidal activity and reduces the toxicity of indoxacarb. Carbohydr. Polym. 2021, 266, 118150. [Google Scholar] [CrossRef]
- Grunberger, J. W.; Ghandehari, H. Layer-by-Layer Hollow Mesoporous Silica Nanoparticles with Tunable Degradation Profile. Pharmaceutics 2023, 15, 832. [Google Scholar] [CrossRef] [PubMed]
- Martin-Ortigosa, S.; Valenstein, J. S.; Lin, V. S.-Y.; Trewyn, B. G.; Wang, K. Gold Functionalized Mesoporous Silica Nanoparticle Mediated Protein and DNA Codelivery to Plant Cells Via the Biolistic Method. Adv. Funct. Mater. 2012, 22, 3576–3582. [Google Scholar] [CrossRef]
- Cao, X.; Wang, C.; Luo, X.; Yue, L.; White, J. C.; Elmer, W.; Dhankher, O. P.; Wang, Z.; Xing, B. Elemental Sulfur Nanoparticles Enhance Disease Resistance in Tomatoes. ACS Nano 2021, 15, 11817–11827. [Google Scholar] [CrossRef]
- Kong, X.; Zhang, C.; Zheng, H.; Sun, M.; Zhang, F.; Zhang, M.; Cui, F.; Lv, D.; Liu, L.; Guo, S.; Zhang, Y.; Yuan, X.; Zhao, S.; Tian, H.; Ding, Z. Antagonistic Interaction between Auxin and SA Signaling Pathways Regulates Bacterial Infection through Lateral Root in Arabidopsis. Cell Rep. 2020, 32, 108060. [Google Scholar] [CrossRef]
- Ma, C.; Borgatta, J.; Hudson, B. G.; Tamijani, A. A.; De La Torre-Roche, R.; Zuverza-Mena, N.; Shen, Y.; Elmer, W.; Xing, B.; Mason, S. E.; Hamers, R. J.; White, J. C. Advanced material modulation of nutritional and phytohormone status alleviates damage from soybean sudden death syndrome. Nat. Nanotechnol. 2020, 15, 1033–1042. [Google Scholar] [CrossRef]
- Abdelrhim, A. S.; Mazrou, Y. S. A.; Nehela, Y.; Atallah, O. O.; El-Ashmony, R. M.; Dawood, M. F. A. Silicon Dioxide Nanoparticles Induce Innate Immune Responses and Activate Antioxidant Machinery in Wheat Against Rhizoctonia solani. Plants 2021, 10, 2758. [Google Scholar] [CrossRef] [PubMed]
- Zhao, P.; Yuan, W.; Xu, C.; Li, F.; Cao, L.; Huang, Q. Enhancement of Spirotetramat Transfer in Cucumber Plant Using Mesoporous Silica Nanoparticles as Carriers. J. Agric. Food Chem. 2018, 66, 11592–11600. [Google Scholar] [CrossRef]
- Pan, W.; Zhang, H.J.; Zhang, Y.F.; Wang, M.; Tsui, M. T.K.; Yang, L.; Miao, A.J. Silica nanoparticle accumulation in plants: current state and future perspectives. Nanoscale 2023, 15, 15079–15091. [Google Scholar] [CrossRef]
- Zhao, P.; Cao, L.; Ma, D.; Zhou, Z.; Huang, Q.; Pan, C. Translocation, distribution and degradation of prochloraz-loaded mesoporous silica nanoparticles in cucumber plants. Nanoscale 2018, 10, 1798–1806. [Google Scholar] [CrossRef]
- El-Shabrawy, Use Silica nanoparticles in controlling late wilt disease in maize caused by Harpophora Maydis. Egy. J. Appl. Sci. 2021, 36, 1–19.
- Cui, J.; Liu, T.; Li, F.; Yi, J.; Liu, C.; Yu, H. Silica nanoparticles alleviate cadmium toxicity in rice cells: Mechanisms and size effects. Environ. Pollut. 2017, 228, 363–369. [Google Scholar] [CrossRef]
- Abdelrhim, A. S.; Mazrou, Y. S. A.; Nehela, Y.; Atallah, O. O.; El-Ashmony, R. M.; Dawood, M. F. A. Silicon Dioxide Nanoparticles Induce Innate Immune Responses and Activate Antioxidant Machinery in Wheat Against Rhizoctonia solani. Plants 2021, 10, 2758. [Google Scholar] [CrossRef]
- Fang, X.; Chen, C.; Liu, Z.; Liu, P.; Zheng, N. A cationic surfactant assisted selective etching strategy to hollow mesoporous silica spheres. Nanoscale 2011, 3, 1632–1639. [Google Scholar] [CrossRef] [PubMed]
- Bossert, D.; Urban, D. A.; Maceroni, M.; Ackermann-Hirschi, L.; Haeni, L.; Yajan, P.; Spuch-Calvar, M.; Rothen-Rutishauser, B.; Rodriguez-Lorenzo, L.; Petri-Fink, A.; Schwab, F. A hydrofluoric acid-free method to dissolve and quantify silica nanoparticles in aqueous and solid matrices. Sci. Rep. 2019, 9, 7938. [Google Scholar] [CrossRef]
- Zhu, J.; Tang, J.; Zhao, L.; Zhou, X.; Wang, Y.; Yu, C. Ultrasmall, Well-Dispersed, Hollow Siliceous Spheres with Enhanced Endocytosis Properties. Small 2010, 6, 276–282. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C. A.; Rasband, W. S.; Eliceiri, K. W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Wang, Y.; Zhao, C.; Wang, X.; Shen, H.; Yang, L. Exogenous Ethylene Alleviates the Inhibition of Sorbus pohuashanensis Embryo Germination in a Saline-Alkali Environment (NaHCO3). Int. J. Mol. Sci. 2023, 24, 4244. [Google Scholar] [CrossRef]
- Wang, L.Y.; Zhang, Y.F.; Yang, D.Y.; Zhang, S.J.; Han, D.D.; Luo, Y.P. Aureoverticillactam, a Potent Antifungal Macrocyclic Lactam from Streptomyces aureoverticillatus HN6, Generates Calcium Dyshomeostasis-Induced Cell Apoptosis via the Phospholipase C Pathway in Fusarium oxysporum f. sp. cubense Race 4. Phytopathology 2021, 111, 2010–2022. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Yuan, T.; Wang, Y.; Zhang, D.; Bai, T.; Xu, S.; Wang, Y.; Tang, W.; Zheng, S.J. Identification and evaluation of resistance to Fusarium oxysporum f. sp. cubense tropical race 4 in Musa acuminata Pahang. Euphytica 2018, 214, 106. [Google Scholar] [CrossRef]
- Chávez-Arias, C. C.; Gómez-Caro, S.; Restrepo-Díaz, H. Physiological Responses to the Foliar Application of Synthetic Resistance Elicitors in Cape Gooseberry Seedlings Infected with Fusarium oxysporum f. sp. physali. Plants 2020, 9, 176. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Li, P.; Zhang, M.; Li, T.; Zhang, L. The control effect of combined application of amino acid water-soluble fertilizer and microbial agents for cowpea Fusarium wilt. China plant protection 2015, 35, 52–53. [Google Scholar]
- Jeger, M. J.; Viljanen-Rollinson, S. L. H. The use of the area under the disease-progress curve (AUDPC) to assess quantitative disease resistance in crop cultivars. Theor. Appl. Genet. 2001, 102, 32–40. [Google Scholar] [CrossRef]
- Szkop, M.; Szkop, U.; Kęszycka, P.; Gajewska, D. A Simple and Robust Protocol for fast RP-HPLC Determination of Salicylates in Foods. Food Anal. Methods 2017, 10, 618–625. [Google Scholar] [CrossRef]
- Kęszycka, P. K.; Szkop, M.; Gajewska, D. Overall Content of Salicylic Acid and Salicylates in Food Available on the European Market. J. Agric. Food Chem. 2017, 65, 11085–11091. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
HMSNs under investigation. TEM images of (a) HMSNs–406, Scale bar: 1 µm; (b) HMSNs–96, Scale bar: 200 nm; and (c) HMSNs–19, Scale bar: 500 nm. Particle size distributions of (d) HMSNs–406, (e) HMSNs–96, and (f) HMSNs–19 based on the TEM image analysis. Averages ± standard deviations.
Figure 1.
HMSNs under investigation. TEM images of (a) HMSNs–406, Scale bar: 1 µm; (b) HMSNs–96, Scale bar: 200 nm; and (c) HMSNs–19, Scale bar: 500 nm. Particle size distributions of (d) HMSNs–406, (e) HMSNs–96, and (f) HMSNs–19 based on the TEM image analysis. Averages ± standard deviations.
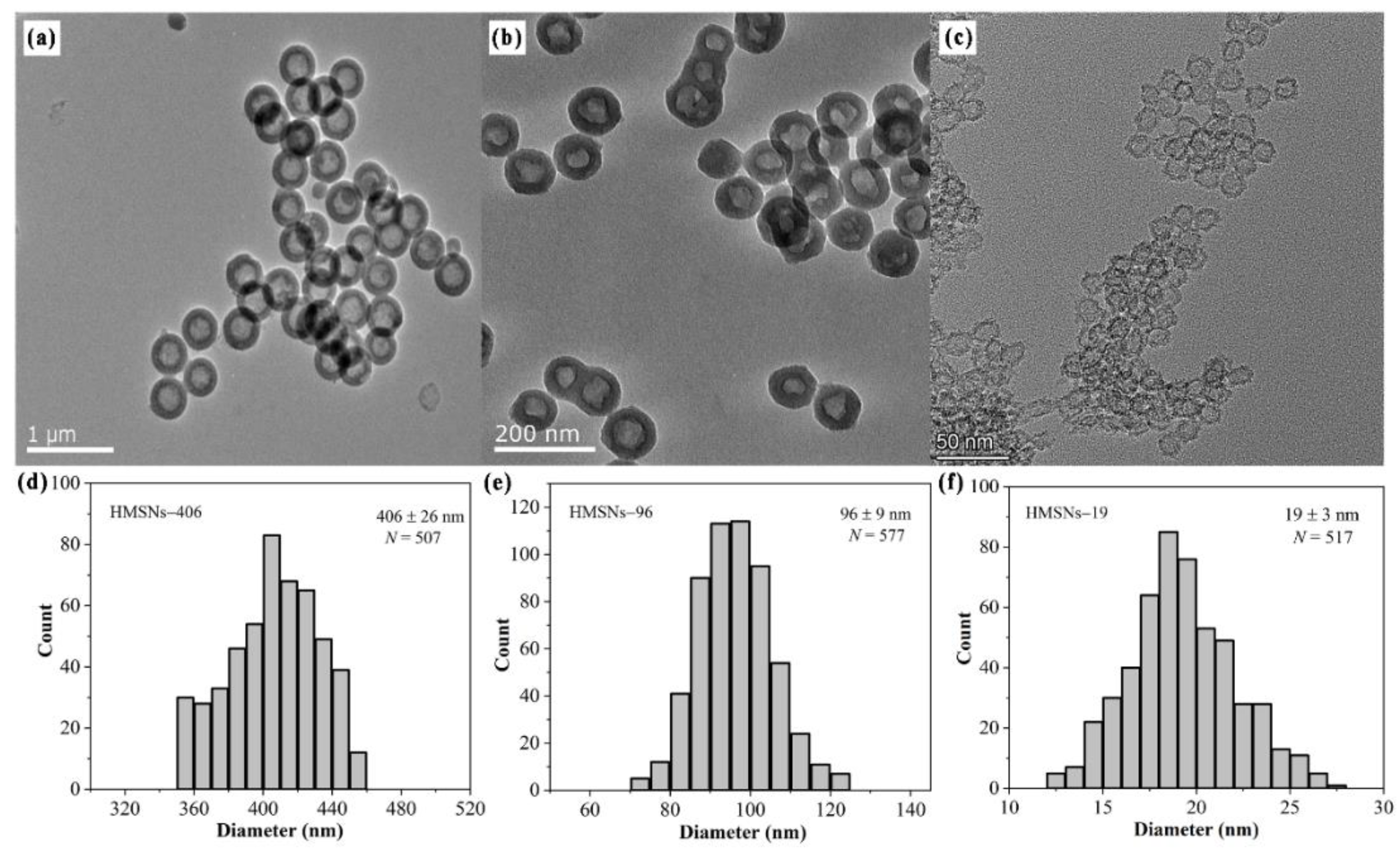
Figure 2.
Effect of HMSNs (a-c) and Si(OH)4 (d) on the germination of cowpea seeds. The concentrations were set as 100, 500, 1000 and 1500 mg/L, respectively. The error bars are averages and standard deviations of ten replicates. Asterisks (*) represent significant differences as compared with the sterile water controls (0 mg/L) using the one-way ANOVA mode for significance testing with Dunnett’s multiple comparisons test at P < 0.05.
Figure 2.
Effect of HMSNs (a-c) and Si(OH)4 (d) on the germination of cowpea seeds. The concentrations were set as 100, 500, 1000 and 1500 mg/L, respectively. The error bars are averages and standard deviations of ten replicates. Asterisks (*) represent significant differences as compared with the sterile water controls (0 mg/L) using the one-way ANOVA mode for significance testing with Dunnett’s multiple comparisons test at P < 0.05.
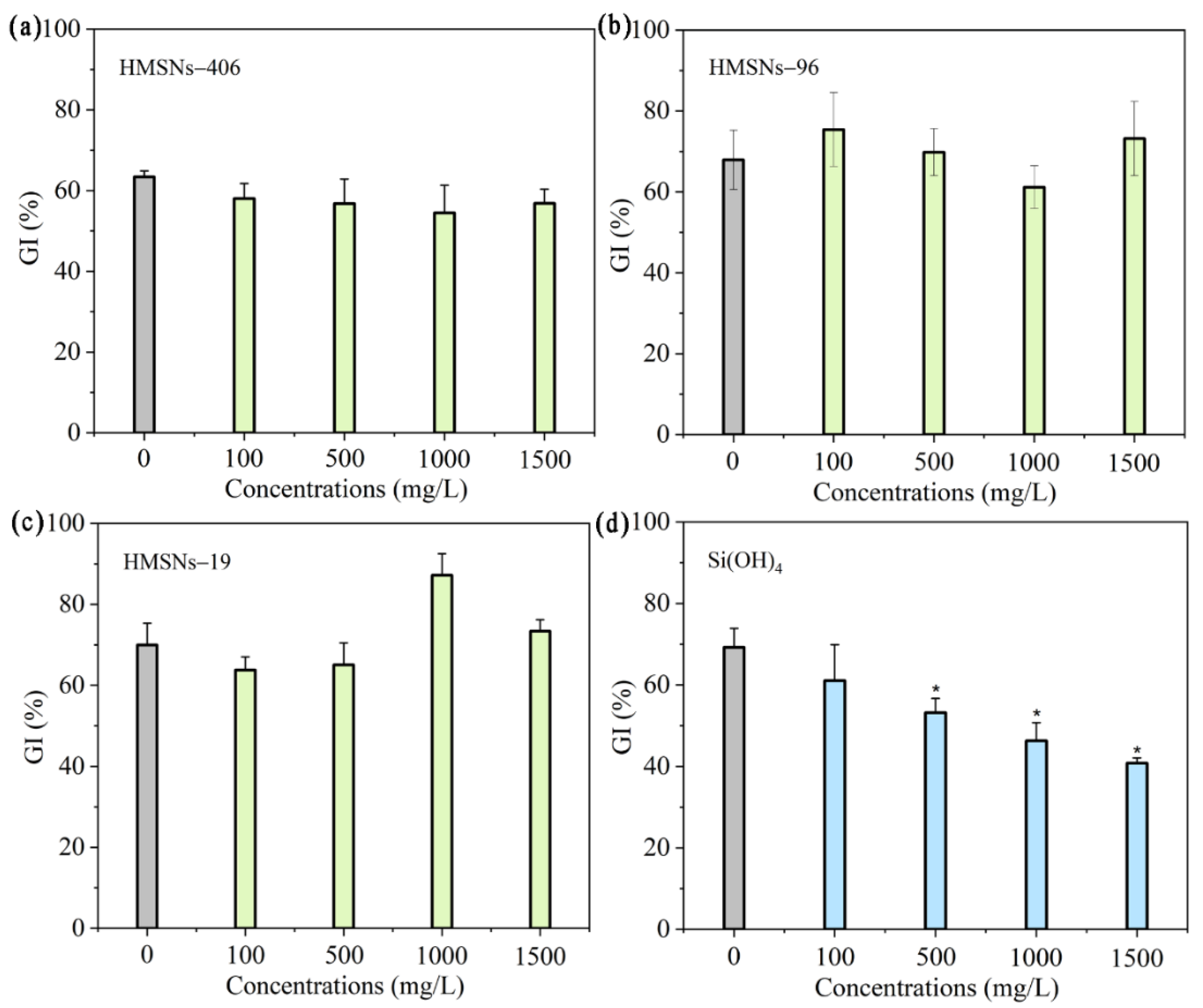
Figure 3.
Effect of HMSNs (a-c) and Si(OH)4 (d) on the cowpea seeding growth. The concentrations were set as 100, 500, 1000 and 1500 mg/L, respectively. The error bars are averages and standard deviations of ten replicates. Asterisks (*) represent significant differences as compared with the sterile water controls (0 mg/L) using the one-way ANOVA mode for significance testing with Dunnett’s multiple comparisons test at P < 0.05.
Figure 3.
Effect of HMSNs (a-c) and Si(OH)4 (d) on the cowpea seeding growth. The concentrations were set as 100, 500, 1000 and 1500 mg/L, respectively. The error bars are averages and standard deviations of ten replicates. Asterisks (*) represent significant differences as compared with the sterile water controls (0 mg/L) using the one-way ANOVA mode for significance testing with Dunnett’s multiple comparisons test at P < 0.05.
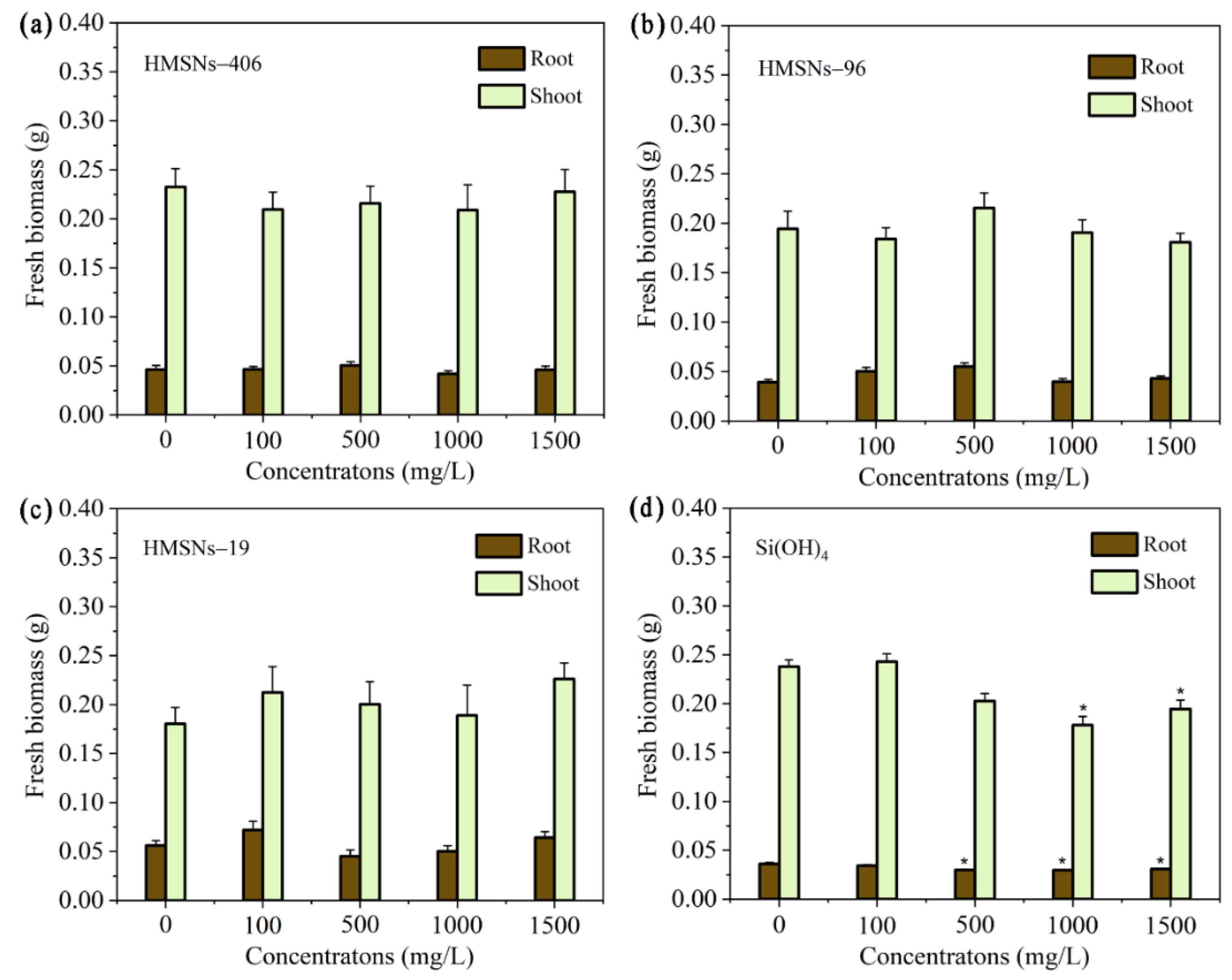
Figure 4.
HMSNs enhance the disease resistance in a concentration- and size-dependent manner. (a) Effects of HMSNs and Si(OH)4 on the cowpea Fusarium wilt incidence by foliar application using AUDPC as the indicator. (b) Phenotypic images of cowpea shoots, longitudinal-cut stems and roots by HMSNs–406 (1000 mg/L) or Si(OH)4 (100 mg/L) foliar treatment for 29 days. (c) Changes in the shoot and root biomass of cowpea foliarly treated by HMSNs and Si(OH)4 at different concentrations of 100 and 1000 mg/L for 29 days. The healthy control represents cowpea plants growing in the noninfected soil and treated with water. Other treatments represent cowpea plants growing in the infected soil and foliarly treated with water (the infected control), Si(OH)4 or HMSNs–406. The error bars are averages and standard deviations of three replicates. Asterisks (*) in (a) and (c) represent significant differences as compared with the infected controls using the one-way ANOVA mode for significance testing with Dunnett’s multiple comparisons test at P < 0.05.
Figure 4.
HMSNs enhance the disease resistance in a concentration- and size-dependent manner. (a) Effects of HMSNs and Si(OH)4 on the cowpea Fusarium wilt incidence by foliar application using AUDPC as the indicator. (b) Phenotypic images of cowpea shoots, longitudinal-cut stems and roots by HMSNs–406 (1000 mg/L) or Si(OH)4 (100 mg/L) foliar treatment for 29 days. (c) Changes in the shoot and root biomass of cowpea foliarly treated by HMSNs and Si(OH)4 at different concentrations of 100 and 1000 mg/L for 29 days. The healthy control represents cowpea plants growing in the noninfected soil and treated with water. Other treatments represent cowpea plants growing in the infected soil and foliarly treated with water (the infected control), Si(OH)4 or HMSNs–406. The error bars are averages and standard deviations of three replicates. Asterisks (*) in (a) and (c) represent significant differences as compared with the infected controls using the one-way ANOVA mode for significance testing with Dunnett’s multiple comparisons test at P < 0.05.
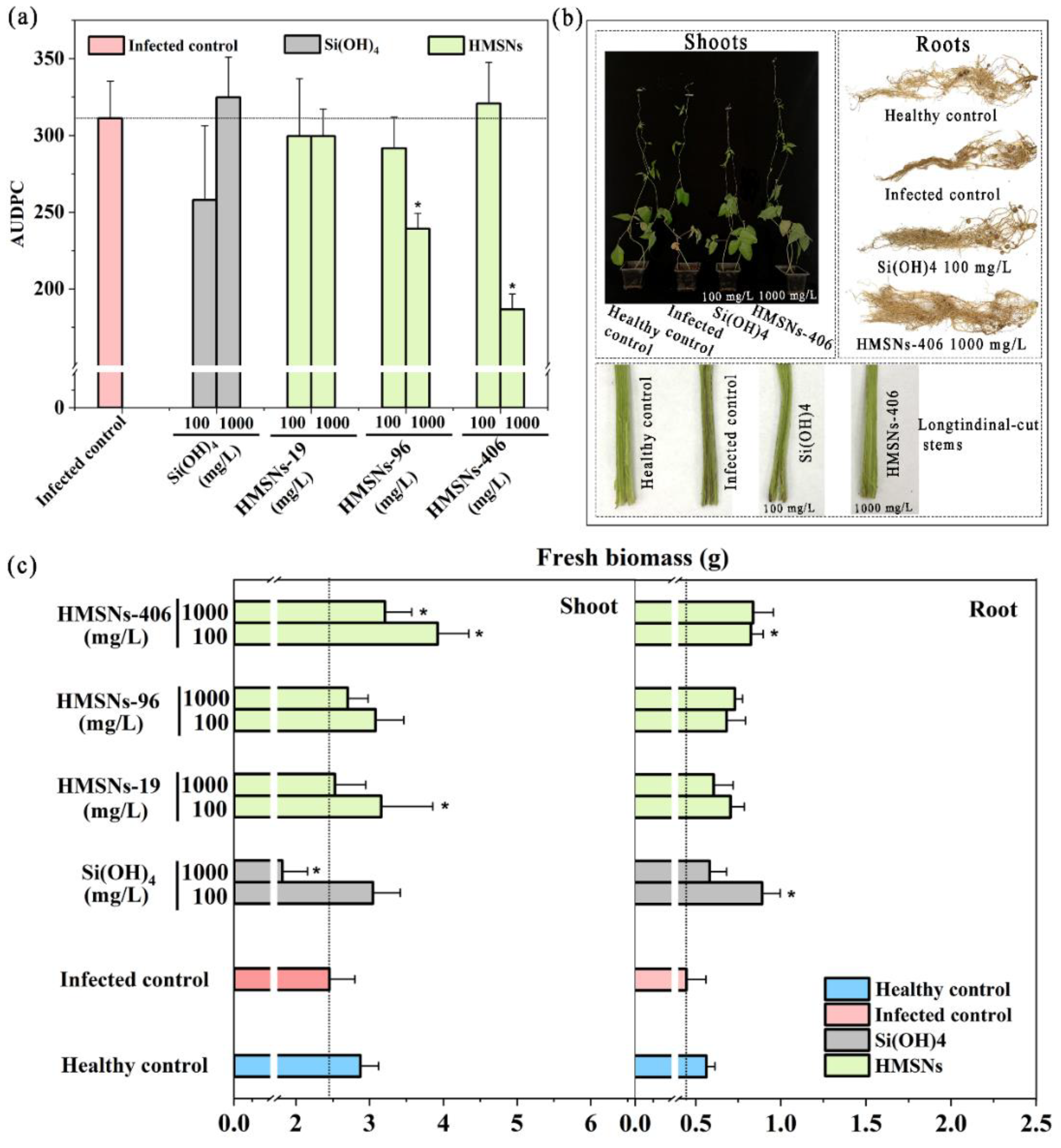
Figure 5.
HMSNs–406 induce Fusarium wilt resistance based on SA-dependent SAR pathway in cowpea. (a) Changes in salicylic acid (SA) content in cowpea roots infected by FOP. (b-c) RT-qPCR analysis of the gene expression of the SAR-related genes PR-1 (b), PR-5 (c) and NPR-1 (d) in cowpea roots. Root samples were collected 29 days after FOP infection. EF1b was used as the reference gene. The healthy control represents cowpea plants growing in the noninfected soil and treated with water. Other treatments represent cowpea plants growing in the infected soil and foliarly treated with water (the infected control), Si(OH)4 (100 mg/L) or HMSNs–406 (1000 mg/L). The error bars are averages and standard deviations of three replicates. Asterisks (*) represent significant differences as compared with the infected controls using the one-way ANOVA mode for significance testing with Dunnett’s multiple comparisons test at P < 0.05.
Figure 5.
HMSNs–406 induce Fusarium wilt resistance based on SA-dependent SAR pathway in cowpea. (a) Changes in salicylic acid (SA) content in cowpea roots infected by FOP. (b-c) RT-qPCR analysis of the gene expression of the SAR-related genes PR-1 (b), PR-5 (c) and NPR-1 (d) in cowpea roots. Root samples were collected 29 days after FOP infection. EF1b was used as the reference gene. The healthy control represents cowpea plants growing in the noninfected soil and treated with water. Other treatments represent cowpea plants growing in the infected soil and foliarly treated with water (the infected control), Si(OH)4 (100 mg/L) or HMSNs–406 (1000 mg/L). The error bars are averages and standard deviations of three replicates. Asterisks (*) represent significant differences as compared with the infected controls using the one-way ANOVA mode for significance testing with Dunnett’s multiple comparisons test at P < 0.05.
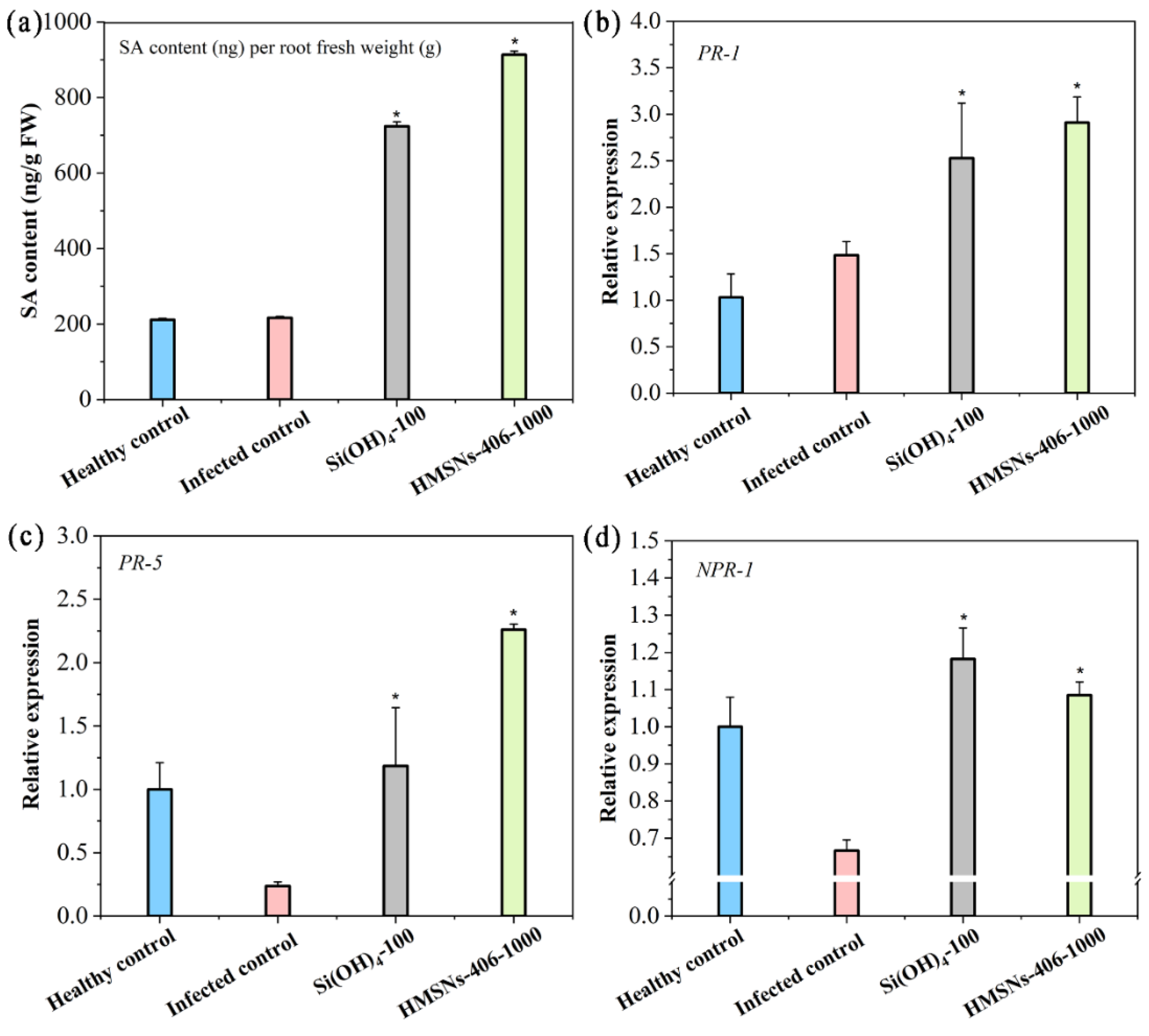
Figure 6.
HMSNs–406 induced changes in the activity of disease-related enzymes of (a) peroxidase (POD), (b) phenylalanine ammonia lyase (PAL) and (c) polyphenol oxidase (PPO) in cowpea roots infected by FOP. Root samples were collected 29 days after FOP infection. The healthy control represents cowpea plants growing in the noninfected soil and foliarly treated with water. Other treatments represent cowpea plants growing in the infected soil and foliarly treated with water (the infected control), Si(OH)4 (100 mg/L) or HMSNs–406 (1000 mg/L). The error bars are averages and standard deviations of three replicates. Asterisks (*) represent significant differences as compared with the infected controls using the one-way ANOVA mode for significance testing with Dunnett’s multiple comparisons test at P < 0.05.
Figure 6.
HMSNs–406 induced changes in the activity of disease-related enzymes of (a) peroxidase (POD), (b) phenylalanine ammonia lyase (PAL) and (c) polyphenol oxidase (PPO) in cowpea roots infected by FOP. Root samples were collected 29 days after FOP infection. The healthy control represents cowpea plants growing in the noninfected soil and foliarly treated with water. Other treatments represent cowpea plants growing in the infected soil and foliarly treated with water (the infected control), Si(OH)4 (100 mg/L) or HMSNs–406 (1000 mg/L). The error bars are averages and standard deviations of three replicates. Asterisks (*) represent significant differences as compared with the infected controls using the one-way ANOVA mode for significance testing with Dunnett’s multiple comparisons test at P < 0.05.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



