Submitted:
27 February 2024
Posted:
27 February 2024
You are already at the latest version
Abstract
Extensive genome structure variation, such as copy number variations (CNVs) and presence/absence variations, is the basis for the remarkable maize genetic diversity; however, the effect of CNVs on maize herbivory defense remains largely underexplored. Here, we report that the naturally occurring duplication of the maize 9-lipoxygenase gene, ZmLOX5, leads to increased resistance of maize to herbivory by fall armyworms (FAW). Previously, we showed that ZmLOX5-derived oxylipins are required for defense against chewing insect herbivores and identified several inbred lines, including Yu796, that contained duplicated CNVs of ZmLOX5, referred to as Yu796-2×LOX5. To test whether introgression of the Yu796-2×LOX5 locus into the herbivore-susceptible B73 background that contains a single ZmLOX5 gene is a feasible approach to increase resistance, we generated a series of near-isogenic lines that contained either 2, 1 or 0 copies of the Yu796-2×LOX5 locus in the B73 background by six back crosses (BC6). Droplet digital PCR (ddPCR) confirmed the successful introgression of the Yu796-2×LOX5 locus in B73. The resulting B73-2×LOX5 inbred line displayed increased resistance against FAW associated with increased expression of ZmLOX5, increased wound-induced production of its primary oxylipin product, the α-ketol, 9-hydroxy-10-oxo-12(Z),15(Z)-octadecadienoic acid (9,10-KODA), and the downstream defense hormones regulated by this molecule, 12-oxo-phytodienoic acid (12-OPDA) and abscisic acid (ABA). Surprisingly, wound-induced JA-Ile production was not increased in B73-2×LOX5, resulting from the increased JA catabolism. Furthermore, B73-2×LOX5 displayed enhanced drought tolerance likely due to increased ABA and 12-OPDA content. Taken together, this study revealed that the duplicated CNV of ZmLOX5 quantitively contributes to maize antiherbivore defense and presents proof-of-concept evidence that the introgression of naturally occurring duplicated CNVs of a defensive gene into the productive, but susceptible, crop varieties is a feasible breeding approach for enhancing plant resistance to herbivory and tolerance to abiotic stress.
Keywords:
α-ketol
; 9-hydroxy-10-oxo-12(Z)
; 15(Z)-octadecadienoic acid (9
; 10-KODA)
; copy number variations (CNVs)
; 12-oxo-phytodienoic acid (12-OPDA)
; abscisic acid (ABA)
; drought tolerance
; herbivory defense
; JA catabolism
; oxylipins
1. Introduction
Maize is a major cereal crop that serves as a significant source of food, feed, and industrial products around the world. Maize yield is significantly affected by insect herbivores, including fall armyworm (FAW), corn rootworms, aphids, and earworms [1]. Among these insects, FAW [Spodoptera frugiperda (J.E. Smith) (Insecta: Lepidoptera: Noctuidae)], is an especially highly destructive pest of not only maize but other crops like sorghum, rice, and other various grasses resulting in enormous economic losses worldwide [2,3]. Maize is one of the most widely grown staple food crops to many African communities, covering 37 million hectares in sub-Saharan Africa [4,5]. More than 300 million people in Africa are dependent on maize for food security [6]. FAW is native to tropical and subtropical Americas, suddenly first arrived to Africa in 2016 and spread rapidly through the continent [7,8]. FAW remains an important pest in Africa’s farming systems, leading to maize yield losses up to 58% [9,10] and estimated annual economic losses amounting to $9.4 billion in Africa alone [9]. Therefore, there is an urgent need to develop maize germplasm that can resist damage from pests, especially for the countries that do not rely on transgenic crops containing Bacillus thuringiensis (Bt)-genes.
The success of crop breeding programs aimed at developing varieties with superior resistance to insect herbivory is largely dependent on the identification of novel defense-related alleles, introgression of which into elite germplasm does not result in the penalty for crop productivity. While it is well documented that plant resistance to chewing insects requires activation of jasmonic acid (JA) signaling pathways [11,12], breeding for increased production of JA does not represent a feasible strategy as increased levels of this defense hormone often results in growth inhibition [13,14]. Recently, we have identified a novel signaling pathway in maize that governs resistance to insect herbivory mediated by an oxylipin α-ketol, named 9,10-KODA [9-hydroxy-10-oxo-12(Z),15(Z)-octadecadienoic acid], which promotes plant herbivory resistance through defense priming and direct toxicity to FAW [15,16,17]. This novel signaling molecule is the major product of a maize tonoplast-localized 9-lipoxygenase gene, ZmLOX5 [15,17]. Mechanical wounding and insect feeding induce ZmLOX5 expression and production of 9,10-KODA in a JA-dependent manner [15,18,19]. The transposon-insertional disruption of ZmLOX5 markedly reduced resistance to FAW and was accompanied by reduced levels of wound-induced 12-oxo-phytodienoic acid (12-OPDA), a JA-precursor and a hormone in its own right, JA, abscisic acid (ABA), green leaf volatiles (GLV) and insecticidal benzoxazinoids [15]. Exogenous treatment of maize seedlings with 9,10-KODA specifically induced production of 12-OPDA and ABA, but not jasmonoyl-L-isoleucine (JA-Ile), and resulted in increased resistance to FAW [15]. Hence, ZmLOX5 represents a valid molecular breeding target for the development of maize lines that are more resistant to herbivory by chewing insects.
In addition to the identification of herbivory defense-related genes, another molecular breeding strategy is to exploit genome structure variations to identify superior loci of those genes. Genome structure variation mainly includes copy number variation (CNV) and/or presence/absence variation (PAV). Maize genome is characterized by extraordinarily high frequency of genetic diversity as assayed at the level of single nucleotide polymorphisms (SNPs), InDel polymorphisms and CNVs. The growing evidence indicates that CNVs are among the major reasons for maize phenotypic diversity [20]. CNVs result from the genomic rearrangements that lead to gains or losses of DNA segments [21,22]. In our search for a superior ZmLOX5 locus, we have sequenced around 1.3 Kb of the ZmLOX5 gene spanning the last two exons in close to 400 inbred lines that represent maize diversity panel [23]. Sequencing results indicated the presence of multiple SNPs and indels found in the ZmLOX5 gene. Of direct releavance to this study, southern blotting analyses of 56 selected inbred lines identified several lines (including sweet, tropical, and temperate lines) that harbor duplicated copy variant of ZmLOX5. This includes Yu796 inbred that we utilized in this study as a source of the duplicated locus, designated here as Yu796-2×LOX5 [23].
In this study, we determined that Yu796-2×LOX5 segregates as a single locus indicating that ZmLOX5 CNVs are tandemly duplicated. Next, we genetically introgressed the Yu796-2×LOX5 locus into the B73 background by 6 back-crosses (BC6) to B73-lox5-3 mutant previously generated in B73 background at the BC7 stage. The resulting B73-2×LOX5 lines are near-isogenic to B73 and contain either single or duplicated ZmLOX5 locus and were used to rigorously test whether increased copy number of this gene confers increased resistance to FAW. We found that maize resistance against FAW feeding was positively correlated with the gene dosage. Specifically, the seedlings homozygous for the duplicated ZmLOX5 (2×LOX5 / 2×LOX5, containing 4 copies present on the two homologous chromosomes displayed the greatest resistance, 2×LOX5 heterozygous seedlings (2 copies) displayed intermediate resistance and mutant seedlings lacking ZmLOX5 (B73-lox5-3 mutant locus, 0 copy) showed the least resistance. B73 inbred containing 2 ZmLOX5 copies exhibited resistance level similar to 2×LOX5 heterozygous seedlings (2 copies). We further showed that the duplication of ZmLOX5 resulted in increased wound-induced expression of ZmLOX5 and other defense-associated genes including ZmLOX8, ZmLOX10 and ZmJAR1a and increased wound-induced production of 9,10-KODA, 12-OPDA and ABA. Interestingly, the levels of the biologically active jasmonate, JA-Ile, were not significantly altered in B73-2×LOX5 lines compared to B73, likely due to the increased JA catabolism.
2. Materials and Methods
2.1. Plant materials
To replace the single ZmLOX5 gene in B73 with the duplicated 2×LOX5 from the inbred line Yu796 [Ames27196 (Lot No.: 04ncai01 SD)], the Yu796-2×LOX5 locus was backcrossed into the genetic background of Mutator (Mu) transposon-insertional lox5-3 mutant in the B73 background at BC7 genetic stage (B73-lox5-3) [15] and advanced to BC6F3 stage to generate near-isogenic lines that segregate for duplicated Yu796-2×LOX5 and the mutant locus. The backcrosses were carried out with the B73-lox5-3 inbred rather than B73 itself because of the ease with which the mutant locus is routinely genotyped by PCR. Homozygous duplicated CNV (Yu796-2×LOX5 / Yu796-2×LOX5), heterozygous (Yu796-2×LOX5 / B73-lox5-3) and mutant (B73-lox5-3 / B73-lox5-3) seedlings were identified from BCnF2 segregating populations by using PCR genotyping with Mu-terminal inverted repeat-specific (9242: AGAGAAGCCAACGCCAWCGCCTCYA) or gene-specific (lox5-3F: TGCCGGACCAGTCAAGCCCATAT and lox5-3R: GGCCCCTTCCGGTTCTTCAAGTC) primers as described in the previous study [15]. Maize seeds were germinated in conical tubes (4 diameter X 20.5 height, cm) filled with commercial potting soil (Jolly Gardener Pro Line C/20 potting mix, Jolly Gardener, Atlanta, GA, USA) and grown on light shelves at room temperature (22 ± 2 °C) under 16 hours light and 8 hours dark light cycle.
2.2. Genomic DNA extraction and Droplet digital PCR (ddPCR) analyses
Individual V2-stage seedlings of B73, Yu796, and three independent B73-2×LOX5 lines at the BC6F3 genetic stage were used for genomic DNA extraction using ZR plant/seed DNA Kit (Zymo Research, Orange, CA, USA). Genomic DNA concentration was determined by SpectraMax QuickDrop UV-Vis Spectrophotometer (Molecular Devices, San Jose, CA, USA). 1 μg genomic DNA was digested by BamHI-HF (High-Fidelity, New England Biolabs, Ipswich, MA, USA) at 37 °C for one hour. Relative copy number of ZmLOX5 was quantified by droplet digital PCR (ddPCR) [24], using a QX200 Droplet Digital PCR system (Bio-Rad, Hercules, CA, USA). 22 µl of the samples were generated by mixing 2 µl of 20 ng/µl DNA template (digested genomic DNA) and primers with Bio-Rad ddPCR Evagreen Supermix. Forty µL oil emulsions of the DNA samples were generated by the Bio-Rad AutoDG automatic droplet generator (Bio-Rad, Hercules, CA, USA), and transferred to 96-well PCR plates. Plates were sealed and transferred to a Bio-Rad C1000 Touch thermocycler (Bio-Rad, Hercules, CA, USA) for PCR, using manufacturer’s recommended protocol: 95 °C for 5 minutes for polymerase activation, followed by 40 cycles at 95 °C for 30 seconds and 60 °C for one minute, then 4 °C for 5 minutes followed by 90 °C for 5 minutes to stabilize the signal. Ramp time was set to 2 °C per second for each step. 96-well plates were then moved to the QX200 droplet reader (Bio-Rad, Hercules, CA, USA), for digital classification of droplets based on fluorescence. Analysis was conducted using QX Manager Software 2.0 Standard Edition (Bio-Rad, Hercules, CA, USA). Relative copy number for ZmLOX5 was calculated as the concentration of ZmLOX5 DNA template divided by the concentration of the α-tubulin template [25,26]. Primer sequences are shown in Table S1.
2.3. Sequence analysis of 2xLOX5 CNVs
Genomic DNA was extracted from Yu796 as described above. PCR reactions were carried out using Phusion™ High-Fidelity DNA Polymerase (Thermo Fisher Scientific™, Houston, TX, USA), ZmLOX5-specific forward and reverse primers (listed in Table S1). The PCR product was loaded and separated in 1.5 % (w/v) agarose gel and purified using gel extraction kit (QIAGEN, Germantown, MA, USA). “A-Tailing” was added to 3’ blunt-ends of 100 ng purified PCR product using GoTaq™ DNA Polymerase (Promega, Madison, WI, USA) for 10 mins at 72 °C. The gel-purified DNA fragments were cloned into a TA-vector using TOPO™ TA Cloning™ Kit (Thermo Fisher Scientific™, Houston, TX, USA), and subsequently, the resulting plasmid constructs were transformed into TOP10 competent cells (Thermo Fisher Scientific™, Houston, TX, USA). The plasmids were extracted from the TOP10E. coli strains positive for ZmLOX5 fragment insertions and purified for the next step sequencing.
2.4. Mechanical wounding of leaves
For wound-induced gene expression and metabolite analysis, the third leaves of V3- or V4-stage B73-2×LOX5 seedlings at the BC6F3 genetic stage and B73 inbred line were treated by crushing the leaf blade using a hemostat perpendicular to the main vein, but avoiding damaging of the main vein. The whole wounded leaf region was collected at designated times as described below and immediately frozen in liquid N2 and stored at 80 °C freezer. For all experiments, each replicate contained at least two seedlings, and four replicates were tested for expression and metabolite analyses for each designated time point.
2.5. RNA extraction and expression analyses using qRT-PCR
Unwounded or wounded leaves from B73-2×LOX5 and B73 seedlings were collected at 0, 1, 2 and 4 hours post wounding (hpw). Total RNA was extracted using TRIzol™ Reagent (Invitrogen, Waltham, MA, USA) and then treated with RNase-free DNase at 37 °C for 30 mins (Ambion, Corston, Bath, UK). For qRT-PCR, transcript accumulation was measured by using SYBR Green one-step qRT-PCR ROX Mix Kit (Thermo Fisher Scientific™, Waltham, MA, USA), and StepOne Real-Time PCR System (Thermo Fisher Scientific™, Waltham, MA, USA). The qPCR primers are listed in Table S2. Expression of a constitutively expressed house-keeping gene, α-Tubulin, was used as an internal control.
2.6. Fall armyworm resistance assay
The B73-2×LOX5 seedlings at BC6F2 stage, segregating for individuals homozygous for the B73-2×LOX5 locus (2×LOX5 / 2×LOX5), heterozygous (2×LOX5 / lox5-3 locus) and mutant (homozygous for the lox5-3 / lox5-3 mutant locus), were used for FAW clip-cage and 7-days continuous feeding assays. Seedlings of B73 inbred line were used as the single ZmLOX5 gene-containing controls. For the clip-cage assays, the following protocol was used. Five individual seedlings of each genotype per a replicate were grown until V3- or V4-stage. FAW eggs were hatched and fed by artificial diet as described in [15]. Insect diet contained 78.2 g/L FAW diet mix (Southland, Lake Village, AR, USA), 37 g/L casein (Muscle Feast, Hebron, OH, USA), and 13 g/L agar (BD, Sparks, MD, USA). One 2nd or 3rd instar stage larvae of FAW was confined in a “clip cage” on the third leaf for 6-8 hours feeding at room temperature. To determine the leaf area damaged by insect feeding, the infested leaf tissues were scanned and the consumed leaf surface area determined using ImageJ software (ImageJ 1.36b; Wayne Rasband, NIH, Bethesda, MD, USA). Each biological replicate contained at least five seedlings, and three replicates were tested for each designated time point. For FAW continuous infestation assay, eight FAW neonates were placed into the whorls of maize seedlings that were contained in individual plastic jars, and the neonates were allowed to freely roam and feed for seven days. The consumed leaf tissue and FAW larvae weight were measured at the end of the assay. Each replicate contained at least six seedlings, and three replicates were tested.
2.7. Quantification of metabolites
Unwounded or wounded leaves from duplicated B73-2×LOX5 and B73 seedlings were collected at 0, 1, 2 and 4 hpw. Selected oxylipins and ABA were measured by using LC-MS/MS system AB Sciex 3200 QTRAP LC/MS/MS (Sciex, Framingham, MA, USA) as previously described [27]. Four or five biological replicates were analyzed per each individual genotype, with each replicate containing leaves from at least two seedlings.
2.8. Drought stress test
V2-stage B73-2×LOX5 seedlings at the BC5F3 genetic stage and B73 inbred line were soaked in deionized water for 1 hour to achieve full water content in the soil before drought stress. Fourteen days post withholidng water, the seedling were rewatered by soaking in deionized water for 3 hours. The survival rate was calculated at 3 days post rewatering. Six biological replicates were tested per each individual genotype, with each replicate containing seven seedlings. For transpirational water loss measurements, V2-stage individual seedlings were removed from conical tubes and the soil was carefully removed from the roots to avoid causing damage to the root. The soil-free seedlings were weighted to obtain the value of fresh weight at 0 h (FW0) and wrapped with cotton at the base of the stem and then placed back in conical tubes. The measurements of fresh weight were taken at each hour post drougth stress and represented in Figure 7 as FWt. The relative water loss was calculated using the formula Δ FW (FWt-1 - FWt)/ FW0, where FWt is fresh weight at each designated time point, FWt-1 is weight at one hour before each designated time point and FW0 is weight at 0 hours post drought treatment. and the total relative water loss rate was calculated as (FW0 – FW17) / FW0. Five biological replicates were tested per each individual genotype, with each replicate consisting of eight seedlings.
2.9. Statistical analysis
Results were analyzed by Microsoft Excel. Error bars in all the figures represent standard error (SE) of the mean value. The number of biological replicates and significance thresholds are described in the figure legends. Student’s t-test was carried out for comparisons between the two groups and p values < 0.05 were marked by asterisks for pairs of groups in the Figures. For multiple-group analysis, statistical analysis was performed by one-way ANOVA (analysis of variance) with Tukey’s HSD (honestly significant difference) post hoc test. Pairwise differences between groups were summarized using letter labels: groups sharing the same letter label were not significantly different at α = 0.05, and groups with non-overlapping letter labels were significantly different at α = 0.05.
3. Results
3.1. Introgression of duplicated copy variants of ZmLOX5 from Yu796 into B73
To generate near-isogenic lines carrying duplicated copies of ZmLOX5 in the B73 genetic background, we backcrossed six times the Yu796-2×LOX5 locus into the lox5-3 mutant that was previously backcrossed seven times to B73 as described in [15] with selection carried out against the mutant locus at each back-cross stage. This resulted in the generation of B73 lines containing the Yu796-2×LOX5 locus, designated as B73-2×LOX5, that contained 2 CNV copies of ZmLOX5 instead of the single ZmLOX5 gene found in B73 inbred. To confirm the gene copy number variation, we used ddPCR to test the concentration of ZmLOX5 gene dosage. This state-of-the-art method has been successfully used in determining gene copy variations of human genomic DNA [28,29], and in testing transgene copy number in several crops, including rice, citrus, potato, maize, tomato and wheat [24,30,31]. ddPCR results showed that twice as many ZmLOX5 template molecules were found in Yu796 and the three independent near-isogenic lines of B73 carrying duplicated ZmLOX5 at the BC6F3 genetic stage as compared to B73 (Figure 1).
Representative 1D droplet plots showed the well-defined distribution of positive (shown in blue color) and negative individual droplets (in grey color), in the upper panel for ZmLOX5 and the lower panel for the reference gene, α-Tubulin (Figure 1A). As shown in Figure 1B, the copy concentration of ZmLOX5 is 977 in B73, while almost double concentration of ZmLOX5 is found in Yu796 (1769) and the three independent near-isogenic B73-2×LOX5 lines (2259, 1930 and 1923, respectively). In contrast to the target gene, ZmLOX5, almost equal copy number concentration was determined in each individual for the reference gene, α-Tubulin, representing the single copy gene. The relative copy numbers of ZmLOX5 were calculated using α-Tubulin as internal control [26,32], and the results revealed that only B73 contained one-fold change in the ZmLOX5/α-Tubulin ratio, while Yu796 and the three independent B73-ZmLOX5 lines contained two-fold changes in ZmLOX5/α-Tubulin ratios, indicating these individuals carry duplicate copies of ZmLOX5, whereas B73 encodes a single copy of ZmLOX5, as expected (Figure 1C).
3.2. CNVs of ZmLOX5 are tandemly duplicated and contain multiple SNPs and several indels when compared to the B73-ZmLOX5 locus
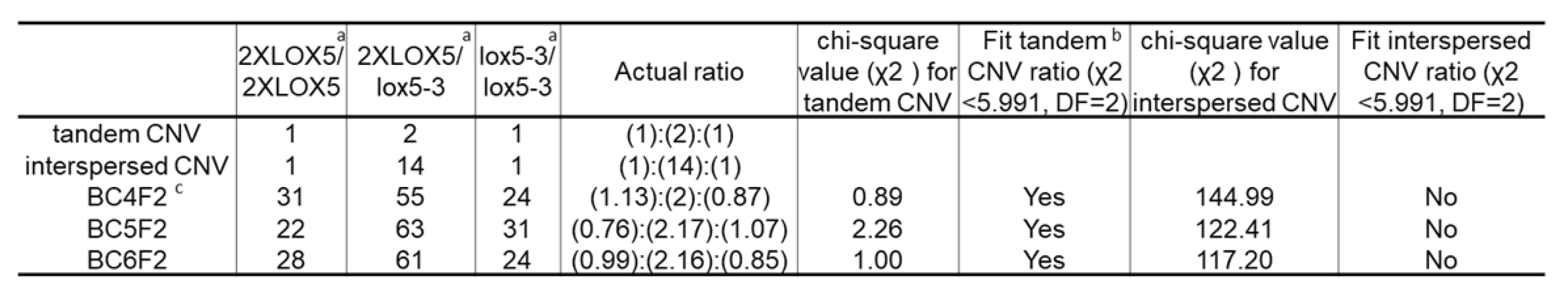
To determine whether CNVs of ZmLOX5 are tandemly duplicated (therefore, genetically linked), or interspersed (thus, not linked), we calculated segregation ratio of 2×LOX5 in the BC4F2, BC5F2 and BC6F2 populations [22]. These F2 populations segregated for individuals homozygous for the duplicated ZmLOX5 locus (2×LOX5/2×LOX5) carrying 4 copies of ZmLOX5 gene, heterozygous locus (2×LOX5/lox5-3) carrying 2 functional copies and the mutant locus (lox5-3/lox5-3) carrying 0 functional copy. The genotyping results revealed that the actual ratio of 2×LOX5/2×LOX5 : 2×LOX5/lox5-3 : lox5-3/lox5-3 individuals in the BC4F2, BC5F2 and BC6F2 populations are (1.13):(2):(0.87), (0.76):(2.17):(1.07) and (0.99):(2.16):(0.85), respectively (Table 1), which is in agreement that the 2×LOX5 CNVs are inherited as a single locus. The chi-square test showed that χ2 distributions for tandem duplication (segregation ratio is 1:2:1) are 0.89, 2.26 and 1.00, in the respective populations, which are less than χ2 (P0.05,2 = 5.99, degrees of freedom of 2 are associated with a p-value >0.05) [33], while χ2 distributions for interspersed duplication (segregation ration is 1:14:1) are 144.99, 122.41 and 117.20, respectively (Table 1). Therefore, the chi-square test agrees with the hypothesis that 2×LOX5 CNVs segregates in the normal mendelian ratio consistent with a single locus, suggesting that the two copies are linked as tandem duplicates.
Table 1a, The number of plants in each category and segregation ratios are presented for homozygous 2×LOX5 locus carrying 4 functional copies of the gene (2×LOX5/2×LOX5), heterozygous (2×LOX5/lox5-3) carrying 2 functional copies of the ZmLOX5 gene and the mutant locus (lox5-3/lox5-3), carrying 0 functional copy of the ZmLOX5 gene.
Table 1b, χ2 (P0.05,2 = 5.99, degrees of freedom of 2 are associated with a p-value >0.05); χ2 distributions for tandem duplication (segregation ratio is 1:2:1) is less than 5.99, consistant with the hypothesis that 2×LOX5 CNVs segregates in the normal mendelian ratio consistent with a single locus, suggesting that the two copies are linked as tandem duplicates.
Table 1c, 2×LOX5 locus from Yu796 was back-crossed 4, 5, or 6 times into the B73 line carrying lox5-3 mutant locus. The resulting segregating populations were PCR-genotyped and the segregation ratios were determined in BC4F2, BC5F2 and BC6F2 populations.
To characterize nucleotide sequence polymorphism between the two CNVs of ZmLOX5, hereafter named ZmLOX5-CNV1 and ZmLOX5-CNV2, we have PCR-amplified, cloned into a TOPO TA vector and sequenced a 1.3 Kb ZmLOX5 fragment containing the 3’-portion of the gene spanning the partial 8th and the entire 9th exons of the ZmLOX5 gene [23], the 8th intron, and partial 3’-UTR (Figure 2). As expected, the greatest nucleotide polymorphism was identified in the 8th intron, where sequencing results revealed the presence of four SNPs differentiating Yu796-CNV1 from Yu796-CNV2 (Figure 2). Importantly, there are two indels that differentiate the B73-LOX5 from both CNVs from Yu796, that can be exploited for PCR-based differentiation between the B73 ZmLOX5 and ZmLOX5-CNV1 and 2.
3.3. Duplication of ZmLOX5 leads to the increased expression of ZmLOX5 and other wound-inducible oxylipin biosynthesis genes
It is often reported that CNVs effect the expression level of the gene by the virtue of the difference in the number of functional copies [22,34]. Therefore, we measured ZmLOX5 transcript accumulation in B73 and the B73-2×LOX5 lines. qRT-PCR analysis showed that ZmLOX5 expression was increased by 7.8-fold at the resting stage and 2.8-fold at 1 hpw in the lines harboring duplicated ZmLOX5 compared to B73 (Figure 3), indicating that the duplication of the gene resulted in enhanced expression in both untreated and wounded leaves. Unlike ZmLOX5 at the resting stage, expression of the JA biosynthesis genes, ZmLOX8, ZmLOX10, and ZmJAR1a [16], are not significantly altered between B73-2×LOX5 and B73 lines. However, in response to mechanical wounding, ZmLOX10 and ZmJAR1a expression was increased 1.9- and 2.1-fold, respectively, at 2 hpw, and ZmLOX8 expression was increased 1.6-fold at 1 hpw (Figure 3). The increased expression of ZmLOX8, ZmLOX10 and ZmJAR1a suggested increased biosynthesis of 12-OPDA, GLV and/or possibly other jasmonates.
3.4. Duplication of ZmLOX5 conferred enhanced resistance against FAW
To test whether the duplication of ZmLOX5 increased maize resistance against insect herbivory as hypothesized, we carried out FAW resistance assays using B73-2×LOX5 population at the BC6F2 genetic stage that segregates for individuals containing either 4 copies (2×LOX5/2×LOX5), or 2 copies (heterozygous, 2×LOX5/lox5-3), or no copies of ZmLOX5 (the knock-out mutant, lox5-3/lox5-3). B73 inbred line that has 2 functional copies of ZmLOX5 (B73-LOX5/B73-LOX5) was included as a control. The individual seedlings were infested with single 2nd- or 3rd-instar FAW larvae placed in a clip-cage for feeding for the duration of 6-8 hours. As shown in Figure 4A and B, FAW consumed the least leaf tissue in the homozygous B73-2×LOX5 seedlings, the most leaf tissue in the mutant seedlings, and the intermediate area of leaf tissue in the heterozygous seedlings. Leaf tissue consumed by larvae in B73 was similar to the area consumed in the heterozygous seedlings (Figure 4A and B). Next, we tested FAW resistance in these same genotypes using 7-days continuous feeding assay. These assays clearly showed that the seedlings homozygous for the 2×LOX5 locus displayed the greatest resistance to FAW feeding, the mutants displayed the least resistance, and the heterozygous seedlings exhibited intermediate resistance. Similar to the assays measuring leaf area consumed, insect resistance in B73 was similar to the heterozygous (2×LOX5/lox5-3) seedlings (Figure 4C and D). These results were further supported by the measurements of FAW larvae weight gain, where we observed that larvae gained the least weight on the seedlings homozygous for the duplication, followed by the heterozygous individuals and the most gain weight was observed on lox5-3 mutant, as expected (Figure 4E and F). Similarly, the larvae fed on B73 seedlings gained more weight compared to those fed on the homozygous B73-2×LOX5 individuals (4 copies), but gained less weight to those on the heterozygous individuals (2 copies) (Figure 4E and 4F). Together, these results showed that duplication of ZmLOX5 enhanced maize defense against FAW attack.
3.5. Duplication of ZmLOX5 promoted wound-induced oxylipin and ABA production
Our previous study revealed that wound-triggered JA-Ile production was reduced along with 9,10-KODA, 12-OPDA and ABA in the lox5 mutants, resulting in decreased FAW defense [15]. Therefore, we assessed whether increased resistance to FAW in B73-2×LOX5 seedlings resulted from increased JA accumulation. Surprisingly, wound-induced JA-Ile production in B73-2×LOX5 was not significantly increased as compared to B73 (Figure 5), suggesting that JA-Ile is not a major reason for increased resistance. However, metabolite analyses revealed that the levels of wound-induced 9,10-KODA, the major product of ZmLOX5 [15], and ABA are significantly increased at 1 and 4 hpw, while 12-OPDA production is significantly increased at 1 hpw in the B73-2×LOX5 seedlings compared to B73. These results are consistent with our previous study that showed that exogenous treatment with 9,10-KODA strongly induced 12-OPDA and ABA, regardless of wounding, while having modest inhibitory effect on the wound-induced JA-Ile production [15].
Because JA biosynthesis genes, ZmLOX8, ZmLOX10 and ZmJAR1a, were expressed to higher levels, but JA-Ile levels were not significantly changed as the result of ZmLOX5 duplication, we measured the accumulation of the JA catabolites, which are not known to exhibit biological activity ascribed to JA-Ile [35,36]. Additional oxylipin profiling (Figure 6A) showed that JA catabolites, 12OH-JA, 12OH-JA-Ile and 12COOH-JA-Ile, were significantly increased in B73-2×LOX5 seedlings compared to B73 (Figure 6B), suggesting that JA-Ile levels were reduced due to increased JA catabolism. In summary, these results indicate that the increased gene copy number of ZmLOX5 enhanced FAW defense through increased production of 9,10-KODA, 12-OPDA and ABA, but not JA-Ile.
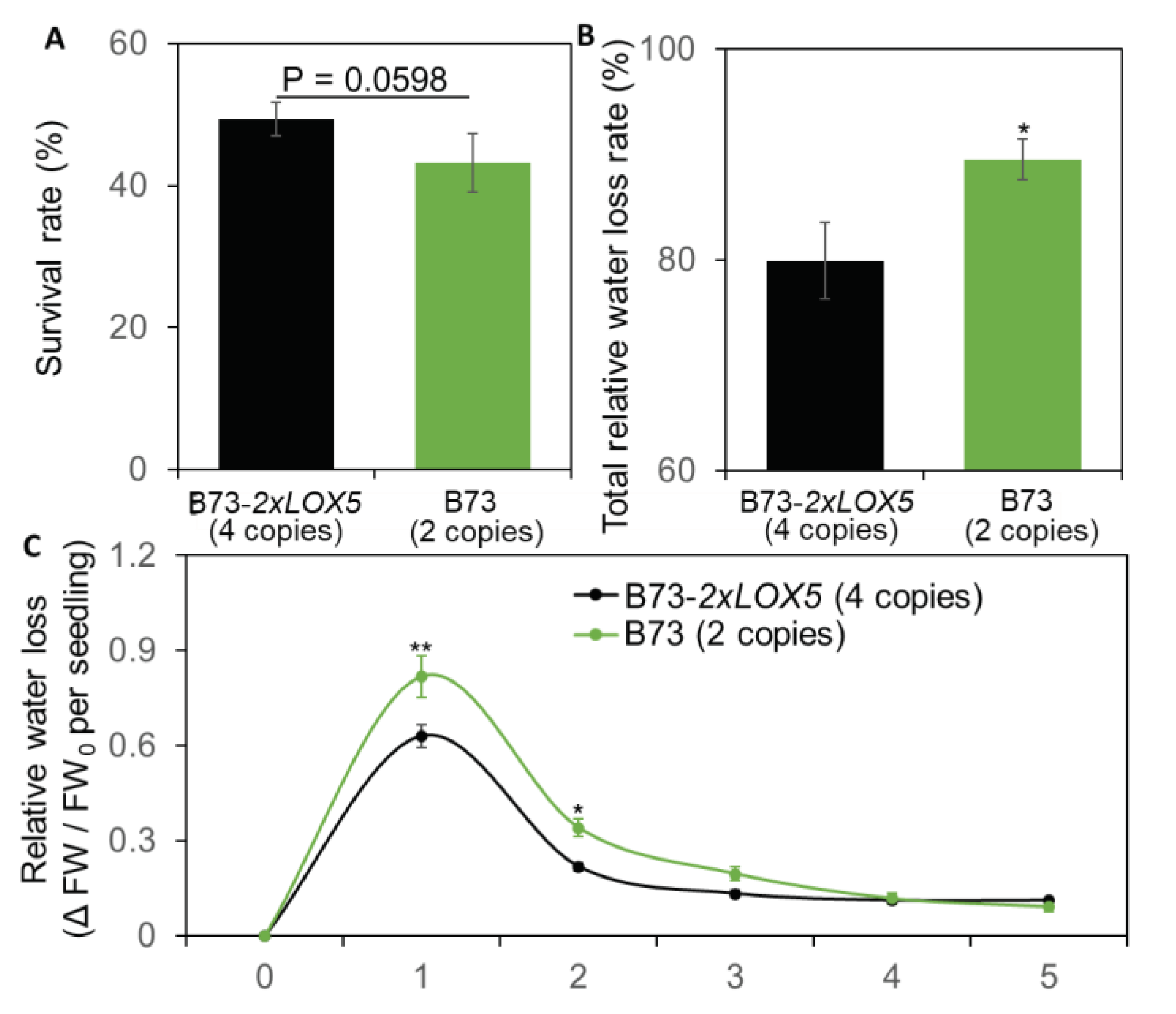
3.6. Duplication of ZmLOX5 promoted drought tolerance
ABA and 12-OPDA play a key role in the regulation of drought tolerance in plants [37]. Due to elevated levels of ABA and 12-OPDA in B73-2×LOX5 seedlings, we tested whether the duplication of ZmLOX5 altered maize response to drought. Drought stress tests revealed that B73-2×LOX5 seedlings exhibited moderately increased survival rate under 14 days drought stress followed by rewatering (Figure 7A). ABA and 12-OPDA promotes drought tolerance through the regulation of stomtal closure [38,39], hence, we assessed water loss in B73-2×LOX5 and B73 inbred line upon seedlings exposure to drought. As shown in Figure 7B, B73-2×LOX5 lost less water as compared to B73 in response to seedling drying for 17 hours. Furthermore, reduced water loss in B73-2×LOX5 was already evident at 1 and 2 hours post water deprivation (Figure 7C). These observations suggest that the duplication of ZmLOX5 increased maize tolerance to drought stress.
Figure 7.
Increased gene dosage of ZmLOX5 results in increased drought tolerance. A. Survival rates of B73-2×LOX5 (2×LOX5/2×LOX5, 4 copies) and B73 inbred line after 14-days drought stress followed by the recovery for 4 days of re-watering. Error bars indicate SE based on eight bioligical replicates. B. The relative water loss rates (%) of B73-2×LOX5 and B73 inbred line at 17 hours post drought treatment. C. The relative water loss [ΔFW (fresh weight) (FWt-1 - FWt) / FW0 (fresh weight before drought stress) per maize seedling] of B73-2×LOX5 and B73 inbred line during water deprivation. Asterisks represent significant differences between B73 -2×LOX5 and B73 lines at each time point using Student’s t-test (*p < 0.05 and **p < 0.01).
Figure 7.
Increased gene dosage of ZmLOX5 results in increased drought tolerance. A. Survival rates of B73-2×LOX5 (2×LOX5/2×LOX5, 4 copies) and B73 inbred line after 14-days drought stress followed by the recovery for 4 days of re-watering. Error bars indicate SE based on eight bioligical replicates. B. The relative water loss rates (%) of B73-2×LOX5 and B73 inbred line at 17 hours post drought treatment. C. The relative water loss [ΔFW (fresh weight) (FWt-1 - FWt) / FW0 (fresh weight before drought stress) per maize seedling] of B73-2×LOX5 and B73 inbred line during water deprivation. Asterisks represent significant differences between B73 -2×LOX5 and B73 lines at each time point using Student’s t-test (*p < 0.05 and **p < 0.01).
4. Discussion
The high level of structural variation with frequent changes in the genome content was observed in diverse maize genomes. When two maize genomes were compared, more than three thousand CNV or PNV sequences were identified [20], while only several hundred CNV or PNV sequences were identified between the individual from human genomes using high resolution study [40]. CNVs occur widely in plant genomes, however, only a few have been associated with obvious morphological, physiological or developmental phenotypes [22,41]. One of the possible explanations is that the paralogous genes in crops usually function redundantly. As a result, the difference in copy number of one gene may lead to a change in quantitative traits, but not in the variation of qualitative traits. We observed similar results in this study. The variance of copy number of ZmLOX5 quantitatively affected maize anti-insect resistance phenotype in a step-wise gene-dosage dependent manner . Seedlings containing 4 copies of ZmLOX5 displayed the strongest insect resistance, heterozygous for 2×LOX5 (2×LOX5 / lox5-3) seedlings or B73 that contain 2 functional copies of ZmLOX5 displayed intermediate level of resistance, and the homozygous lox5-3 mutant displayed the least insect resistance. It is important to emphasize that th eB73 line harboring duplicated ZmLOX5 CNVs was sigificantly more resistant to FAW compared to B73 inbred line, which contains a single copy of ZmLOX5. The increased resistance to FAW is due to increased expression of ZmLOX5, which in turn resulted in increased production of the major ZmLOX5 product, 9,10-KODA, that we previously showed to possess a potent signaling activity in the activation of maize defense against insect herbivores [15]. Here, we showed that the increased accumulation of 9,10-KODA was accompanied by the increased content of wound-induced levels of 12-OPDA and ABA. These results agree well with our previous study that showed that the lox5 knock-out mutants, in addition to reduced 9,10-KODA levels, displayed reduced levels of 12-OPDA and ABA, the two defense hormones regulated by 9,10-KODA since both of them were strongly induced by exogenous application of this α-ketol [15].
Another consistant observation is that the increased ZmLOX5-mediated herbivore defense does not result from increased JA production. Previously, we showed that ZmLOX5 acts downstream of JA in defense since the treatment with exogenous JA did not rescue susceptibility of lox5 mutants. Moreover, exogenous 9,10-KODA suppresed wound-induced prodcution of JA-Ile. Here, we present evidence that the lack of increased wound-induced JA production in B73-2xLOX5 can be explained by increased rate of JA catabolism . The significance of JA catabolism in the dowregulation of the levels of the biologically active JA-Ile is best illustrated by the recent study of maize Tasselseed5 (Ts5) mutant. This mutant displayed a JA-deficiency phenotype, including tasselseed and reduced wound-induced JA production [35]. Positional cloning and transcriptomics analysis revealed that ZmCYP94B1 is upregulated in Ts5 and this gene regulated ω-oxidation of JA to convert JA or JA-Ile to 12OH-JA or 12OH-JA-Ile, respectively, resulting in the reduced JA accumulation [35]. The disruption of 12-oxo-phytodienoic acid reductases2 (ZmOPR2) reduced wound-induced JA and hebivory defense to FAW associated with increased JA catabolism [36]. Consitant with these studies, our data showed that the wound-induced productions of 12OH-JA, 12OH-JA-Ile and 12COOH-JA-Ile were significantly enhanced in B73-2×LOX5 seedlings compared to B73 inbred line, suggesting that ZmLOX5-mediated pathway promote JA catabolism through as yet unknown mechanism.
Another unanticipated finding in this study is that the duplication of ZmLOX5 contributes to plant drought tolerance presumably due to increased ABA and 12-OPDA production. ABA is well-documented to enhance drought tolerance in plant through the regulatory of stomatal closure to reduce water loss by reducing transpiration rate [38,42,43], and also increaing evidence revealed that 12-OPDA is involved in stomatal movement [39,44]. Our previous study revealed that exogenous apllication of 9,10-KODA resulted in increased ABA and 12-OPDA accumulation [15]. It is consistent with another study that showed 9,10-KODA treatment enhanced drought tolerance in wheat [45].
While our study provides evidence that CNVs of certain defense genes can be utilized for improving herbivory defense and drought tolerance, previous studies showed that CNVs contribute to enhancements of other plant traits. For example, increased gene copy number lead to improved resistance against soybean cyst nematodes [41]. Also, a recent review identified numerous examples of gene dosage that emerged in the course of plant adaptation to stressful environments and resulted in altered traits such as enhanced cold tolerance in grasses, and enhanced herbicide resistance in weeds [46]. In maize, gene duplication effects on shaping certain traits have also been reported [47,48,49,50,51]. For example, maize plants containing a duplicated 14.6-Mb segment of chromosome 1 showed dosage-dependent effects on ear length and flowering time [48]. CNV analysis by whole genome sequencing of the lines highly resistant or highly susceptible to Goss’ bacterial wilt revealed structural genomic differences in 141 genes including CNVs. One such CNV gene was the rp1 rust resistance locus likely involved in resistance to this pathogen [52]. Additionally, maize plants with duplicated copies of the tb1-ref gene displayed increases in crown root number, and increased density of first- and second-order lateral roots [50]. Duplication at the 27-kDa γ-zein locus resulted in enhanced expression of γ-zein protein, which in turn led to endosperm modification from chalky to vitreous, yielding quality protein maize (QPM) [51]. In addition to quantitative traits, the increased gene copy number also contributes to qualitative phenotypic traits. For example, pod corn (Tunicate maize) is the result of dominant gain-of-function mutation at the Tunicate (Tu) locus. The wild-type locus contains a single-copy gene that is only expressed in leaf tissue, however duplication of ZMM19 in Tu locus resulted in ectopic expression of the gene in the inflorescences, thereby conferring vegetative traits to the reproductive organ [53]. Overall, our study provided much needed proof-of-concept that it is possible to substantially improve maize resistance by the genetic introgression of the duplicated CNVs of ZmLOX5 into elite germplasm that is susceptible to herbivory such as B73.
An important aspect of this study is that we successfully adopted ddPCR technology for the accurate estimation of CNVs. While the original discovery of the duplicated ZmLOX5 was based on Southern blotting method, this method is not applicable for molecular breeding programs that rely on genotyping large numbers of individuals at each pollination event. Here, we used ddPCR to confirm the presence of duplicated ZmLOX5 CNVs in Yu796, the original source of the 2×ZmLOX5 locus, and in B73-2×ZmLOX5 populations. ddPCR is able to accurately estimate copy number of specific DNA fragments by dividing a PCR reaction into thousands of nanoliter-scale droplets, so that presence or absence of sequence of interest in a droplet is determined by end-point fluorescence, which can be digitally counted [28]. Previously, ddPCR has been mainly used to accurately determine the number of transgenes in plants [25,30].
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1. Primer sequences of the genes for ddPCR, genotyping and sequencing used in this study. Table S2. Primer sequences for qRT-PCR tests in this study. Table S3. Sequencing results of LOX5-CNV1 and LOX5-CNV2 showing several SNPs found in Exons 9 and 10 relative to the reference sequence of ZmLOX5 (Zm00001eb216870) of B73-REFERENCE-NAM-5.0 genome).
Author Contributions
M.V.K., P.Y., T.K.M. and T.M.C. designed the research, performed functional and biochemical analyses, and wrote the article. P.Y., M.V.K., contributed to backcrossing of Yu796 in to lox5-3 in B73 genetic background. P.Y. performed herbivory resistance test, gene expression and oxylipin analysis. T.K.M. and T.M.C. performed the ddPCR test. The article was edited and reviewed by all authors.
Funding
This work was supported by United States Department of Agriculture (USDA)-National Institute of Food and Agriculture (NIFA) 2017-67013- 26524 and 2021-67013-33568 grants awarded to M.V.K.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The datasets used and analyzed during the current study are available from the corresponding author upon reasonable request.
Acknowledgments
We thank Pei-Cheng Huang, Israel Arellano and Katherine Berg-Felloure (Texas A&M University) for their help in this study. We thank Greg Sword, Henry Fadamiro and Andie Miller, Department of Entomology at Texas A&M University for providing FAW and eggs and larvae.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Feldmann, F.; Rieckmann, U.; Winter, S. The spread of the fall armyworm Spodoptera frugiperda in Africa—What should be done next? Journal of Plant Diseases and Protection 2019, 126, 97–101. [Google Scholar] [CrossRef]
- Tambo, J.A.; Day, R.K.; Lamontagne-Godwin, J.; Silvestri, S.; Beseh, P.K.; Oppong-Mensah, B.; Phiri, N.A.; Matimelo, M. Tackling fall armyworm (Spodoptera frugiperda) outbreak in Africa: an analysis of farmers’ control actions. International Journal of Pest Management 2020, 66, 298–310. [Google Scholar] [CrossRef]
- Overton, K.; Maino, J.L.; Day, R.; Umina, P.A.; Bett, B.; Carnovale, D.; Ekesi, S.; Meagher, R.; Reynolds, O.L. Global crop impacts, yield losses and action thresholds for fall armyworm (Spodoptera frugiperda): A review. Crop Protection 2021, 145, 105641. [Google Scholar] [CrossRef]
- Clarkson, J.; Borah, J.R.; Baudron, F.; Sunderland, T.C.H. Forest Proximity Positively Affects Natural Enemy Mediated Control of Fall Armyworm in Southern Africa. Frontiers in Forests and Global Change 2022, 5. [Google Scholar] [CrossRef]
- Kassie, M.; Wossen, T.; De Groote, H.; Tefera, T.; Sevgan, S.; Balew, S. Economic impacts of fall armyworm and its management strategies: evidence from southern Ethiopia. European Review of Agricultural Economics 2020, 47, 1473–1501. [Google Scholar] [CrossRef]
- Makgoba, M.C.; Tshikhudo, P.P.; Nnzeru, L.R.; Makhado, R.A. Impact of fall armyworm (Spodoptera frugiperda) (J.E. Smith) on small-scale maize farmers and its control strategies in the Limpopo province, South Africa. Jamba (Potchefstroom, South Africa) 2021, 13, 1016. [Google Scholar] [CrossRef] [PubMed]
- De Groote, H.; Kimenju, S.C.; Munyua, B.; Palmas, S.; Kassie, M.; Bruce, A. Spread and impact of fall armyworm (Spodoptera frugiperda J.E. Smith) in maize production areas of Kenya. Agriculture, ecosystems & environment 2020, 292, 106804. [Google Scholar] [CrossRef]
- Paudel Timilsena, B.; Niassy, S.; Kimathi, E.; Abdel-Rahman, E.M.; Seidl-Adams, I.; Wamalwa, M.; Tonnang, H.E.Z.; Ekesi, S.; Hughes, D.P.; Rajotte, E.G. , et al. Potential distribution of fall armyworm in Africa and beyond, considering climate change and irrigation patterns. Scientific Reports 2022, 12, 539. [Google Scholar] [CrossRef]
- Kansiime, M.K.; Rwomushana, I.; Mugambi, I. Fall armyworm invasion in Sub-Saharan Africa and impacts on community sustainability in the wake of Coronavirus Disease 2019: reviewing the evidence. Current Opinion in Environmental Sustainability 2023, 62, 101279. [Google Scholar] [CrossRef]
- Day, R.; Abrahams, P.; Bateman, M.; Beale, T.; Clottey, V.; Cock, M.; Colmenarez, Y.; Corniani, N.; Early, R.; Godwin, J.; et al. Fall Armyworm: Impacts and Implications for Africa. Outlooks on Pest Management 2017, 28, 196–201. [Google Scholar] [CrossRef]
- McConn, M.; Creelman, R.A.; Bell, E.; Mullet, J.E.; Browse, J. Jasmonate is essential for insect defense in Arabidopsis. Proc Natl Acad Sci U S A 1997, 94, 5473–5477. [Google Scholar] [CrossRef] [PubMed]
- Koo, A.J.; Howe, G.A. The wound hormone jasmonate. Phytochemistry 2009, 70, 1571–1580. [Google Scholar] [CrossRef] [PubMed]
- Staswick, P.E.; Su, W.; Howell, S.H. Methyl jasmonate inhibition of root growth and induction of a leaf protein are decreased in an Arabidopsis thaliana mutant. Proceedings of the National Academy of Sciences 1992, 89, 6837–6840. [Google Scholar] [CrossRef] [PubMed]
- Yuan, P.; Du, L.; Poovaiah, B. Ca2+/Calmodulin-Dependent AtSR1/CAMTA3 Plays Critical Roles in Balancing Plant Growth and Immunity. International Journal of Molecular Sciences 2018, 19, 1764. [Google Scholar] [CrossRef] [PubMed]
- Yuan, P.; Borrego, E.; Park, Y.-S.; Gorman, Z.; Huang, P.-C.; Tolley, J.; Christensen, S.A.; Blanford, J.; Kilaru, A.; Meeley, R., et al. 9,10-KODA, an α-ketol produced by the tonoplast-localized 9-lipoxygenase ZmLOX5, plays a signaling role in maize defense against insect herbivory. Molecular Plant 2023, 10.1016/j.molp.2023.07.003, doi:10.1016/j.molp.2023.07.003. [CrossRef]
- Berg-Falloure, K.M.; Kolomiets, M.V. Ketols Emerge as Potent Oxylipin Signals Regulating Diverse Physiological Processes in Plants. Plants 2023, 12, 2088. [Google Scholar] [CrossRef] [PubMed]
- Jimenez-Aleman, G.H.; Jander, G. Maize defense against insect herbivory: A novel role for 9-LOX-derived oxylipins. Molecular Plant 2023, 16, 1484–1486. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.-S.; Kunze, S.; Ni, X.; Feussner, I.; Kolomiets, M.V. Comparative molecular and biochemical characterization of segmentally duplicated 9-lipoxygenase genes ZmLOX4 and ZmLOX5 of maize. Planta 2010, 231, 1425–1437. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Borrego, E.J.; Gorman, Z.; Huang, P.C.; Kolomiets, M.V. Relative contribution of LOX10, green leaf volatiles and JA to wound-induced local and systemic oxylipin and hormone signature in Zea mays (maize). Phytochemistry 2020, 174, 112334. [Google Scholar] [CrossRef] [PubMed]
- Springer, N.M.; Ying, K.; Fu, Y.; Ji, T.; Yeh, C.-T.; Jia, Y.; Wu, W.; Richmond, T.; Kitzman, J.; Rosenbaum, H. , et al. Maize Inbreds Exhibit High Levels of Copy Number Variation (CNV) and Presence/Absence Variation (PAV) in Genome Content. PLOS Genetics 2009, 5, e1000734. [Google Scholar] [CrossRef]
- Dolatabadian, A.; Yuan, Y.; Bayer, P.E.; Petereit, J.; Severn-Ellis, A.; Tirnaz, S.; Patel, D.; Edwards, D.; Batley, J. Copy Number Variation among Resistance Genes Analogues in Brassica napus. Genes (Basel) 2022, 13. [Google Scholar] [CrossRef]
- Żmieńko, A.; Samelak, A.; Kozłowski, P.; Figlerowicz, M. Copy number polymorphism in plant genomes. TAG. Theoretical and applied genetics. Theoretische und angewandte Genetik 2014, 127, 1–18. [Google Scholar] [CrossRef] [PubMed]
- De La Fuente, G.N.; Murray, S.C.; Isakeit, T.; Park, Y.-S.; Yan, Y.; Warburton, M.L.; Kolomiets, M.V. Characterization of Genetic Diversity and Linkage Disequilibrium of ZmLOX4 and ZmLOX5 Loci in Maize. PLOS ONE 2013, 8, e53973. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.M.; Dudley, Q.M.; Patron, N.J. Measurement of Transgene Copy Number in Plants Using Droplet Digital PCR. Bio Protoc 2021, 11, e4075. [Google Scholar] [CrossRef]
- Bell, A.D.; Usher, C.L.; McCarroll, S.A. Analyzing Copy Number Variation with Droplet Digital PCR. Methods Mol Biol 2018, 1768, 143–160. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Joyce, P.A. Application of droplet digital PCR to determine copy number of endogenous genes and transgenes in sugarcane. Plant Cell Reports 2017, 36, 1775–1783. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.-D.; Borrego, E.J.; Kenerley, C.M.; Kolomiets, M.V. Oxylipins Other Than Jasmonic Acid Are Xylem-Resident Signals Regulating Systemic Resistance Induced by <em>Trichoderma virens</em> in Maize. The Plant Cell 2020, 32, 166. [Google Scholar] [CrossRef] [PubMed]
- Hindson, B.J.; Ness, K.D.; Masquelier, D.A.; Belgrader, P.; Heredia, N.J.; Makarewicz, A.J.; Bright, I.J.; Lucero, M.Y.; Hiddessen, A.L.; Legler, T.C.; et al. High-Throughput Droplet Digital PCR System for Absolute Quantitation of DNA Copy Number. Analytical Chemistry 2011, 83, 8604–8610. [Google Scholar] [CrossRef] [PubMed]
- Miotke, L.; Lau, B.T.; Rumma, R.T.; Ji, H.P. High Sensitivity Detection and Quantitation of DNA Copy Number and Single Nucleotide Variants with Single Color Droplet Digital PCR. Analytical Chemistry 2014, 86, 2618–2624. [Google Scholar] [CrossRef]
- Qi, X.; Zhang, C.; Zhu, J.; Liu, C.; Huang, C.; Li, X.; Xie, C. Genome Editing Enables Next-Generation Hybrid Seed Production Technology. Molecular Plant 2020, 13, 1262–1269. [Google Scholar] [CrossRef]
- Wang, H.; Qi, X.; Zhu, J.; Liu, C.; Fan, H.; Zhang, X.; Li, X.; Yang, Q.; Xie, C. Pollen self-elimination CRISPR-Cas genome editing prevents transgenic pollen dispersal in maize. Plant communications 2023, 4, 100637. [Google Scholar] [CrossRef]
- Iwobi, A.; Gerdes, L.; Busch, U.; Pecoraro, S. Droplet digital PCR for routine analysis of genetically modified foods (GMO) – A comparison with real-time quantitative PCR. Food Control 2016, 69, 205–213. [Google Scholar] [CrossRef]
- Pires, A.M.; Branco, J.A. A Statistical Model to Explain the Mendel–Fisher Controversy. Statistical Science 2010, 25, 545–565. [Google Scholar] [CrossRef]
- Lye, Z.N.; Purugganan, M.D. Copy Number Variation in Domestication. Trends in Plant Science 2019, 24, 352–365. [Google Scholar] [CrossRef] [PubMed]
- Lunde, C.; Kimberlin, A.; Leiboff, S.; Koo, A.J.; Hake, S. Tasselseed5 overexpresses a wound-inducible enzyme, ZmCYP94B1, that affects jasmonate catabolism, sex determination, and plant architecture in maize. Communications Biology 2019, 2, 114. [Google Scholar] [CrossRef] [PubMed]
- Huang, P.-C.; Grunseich, J.M.; Berg-Falloure, K.M.; Tolley, J.P.; Koiwa, H.; Bernal, J.S.; Kolomiets, M.V. Maize OPR2 and LOX10 Mediate Defense against Fall Armyworm and Western Corn Rootworm by Tissue-Specific Regulation of Jasmonic Acid and Ketol Metabolism. Genes 2023, 14, 1732. [Google Scholar] [CrossRef] [PubMed]
- Fujita, Y.; Fujita, M.; Satoh, R.; Maruyama, K.; Parvez, M.M.; Seki, M.; Hiratsu, K.; Ohme-Takagi, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. AREB1 Is a Transcription Activator of Novel ABRE-Dependent ABA Signaling That Enhances Drought Stress Tolerance in Arabidopsis The Plant Cell 2005, 17, 3470-3488, doi:10.1105/tpc.105.035659. [CrossRef]
- Leckie, C.P.; McAinsh, M.R.; Allen, G.J.; Sanders, D.; Hetherington, A.M. Abscisic acid-induced stomatal closure mediated by cyclic ADP-ribose. Proceedings of the National Academy of Sciences 1998, 95, 15837–15842. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.; Shi, M.; Sun, Y.; Cheng, H.; Ou, X.; Zhao, Y.; Zhang, X.; Day, B.; Miao, C.; Jiang, K. Light-induced stomatal opening in Arabidopsis is negatively regulated by chloroplast-originated OPDA signaling. Current Biology 2023, 33, 1071–1081.e1075. [Google Scholar] [CrossRef] [PubMed]
- Kidd, J.M.; Cooper, G.M.; Donahue, W.F.; Hayden, H.S.; Sampas, N.; Graves, T.; Hansen, N.; Teague, B.; Alkan, C.; Antonacci, F.; et al. Mapping and sequencing of structural variation from eight human genomes. Nature 2008, 453, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Cook, D.E.; Lee, T.G.; Guo, X.; Melito, S.; Wang, K.; Bayless, A.M.; Wang, J.; Hughes, T.J.; Willis, D.K.; Clemente, T.E.; et al. Copy Number Variation of Multiple Genes at Rhg1 Mediates Nematode Resistance in Soybean. Science 2012, 338, 1206–1209. [Google Scholar] [CrossRef]
- Yuan, P.; Poovaiah, B.W. Interplay between Ca2+/Calmodulin-Mediated Signaling and AtSR1/CAMTA3 during Increased Temperature Resulting in Compromised Immune Response in Plants. International Journal of Molecular Sciences 2022, 23, 2175. [Google Scholar] [CrossRef]
- Zhu, J.-K. Abiotic Stress Signaling and Responses in Plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef]
- Simeoni, F.; Skirycz, A.; Simoni, L.; Castorina, G.; de Souza, L.P.; Fernie, A.R.; Alseekh, S.; Giavalisco, P.; Conti, L.; Tonelli, C.; et al. The AtMYB60 transcription factor regulates stomatal opening by modulating oxylipin synthesis in guard cells. Sci Rep 2022, 12, 533. [Google Scholar] [CrossRef] [PubMed]
- Haque, E.; Osmani, A.A.; Ahmadi, S.H.; Ogawa, S.; Takagi, K.; Yokoyama, M.; Ban, T. KODA, an α-ketol derivative of linolenic acid provides wide recovery ability of wheat against various abiotic stresses. Biocatalysis and Agricultural Biotechnology 2016, 7, 67–75. [Google Scholar] [CrossRef]
- Kondrashov, F.A. Gene duplication as a mechanism of genomic adaptation to a changing environment. Proceedings. Biological sciences 2012, 279, 5048–5057. [Google Scholar] [CrossRef] [PubMed]
- Wingen, L.U.; Münster, T.; Faigl, W.; Deleu, W.; Sommer, H.; Saedler, H.; Theißen, G. Molecular genetic basis of pod corn (Tunicate maize). Proceedings of the National Academy of Sciences 2012, 109, 7115–7120. [Google Scholar] [CrossRef] [PubMed]
- Zuo, T.; Zhang, J.; Lithio, A.; Dash, S.; Weber, D.F.; Wise, R.; Nettleton, D.; Peterson, T. Genes and Small RNA Transcripts Exhibit Dosage-Dependent Expression Pattern in Maize Copy-Number Alterations. Genetics 2016, 203, 1133–1147. [Google Scholar] [CrossRef] [PubMed]
- Maron, L.G.; Guimarães, C.T.; Kirst, M.; Albert, P.S.; Birchler, J.A.; Bradbury, P.J.; Buckler, E.S.; Coluccio, A.E.; Danilova, T.V.; Kudrna, D.; et al. Aluminum tolerance in maize is associated with higher MATE1 gene copy number. Proceedings of the National Academy of Sciences 2013, 110, 5241–5246. [Google Scholar] [CrossRef]
- Gaudin, A.C.M.; McClymont, S.A.; Soliman, S.S.M.; Raizada, M.N. The effect of altered dosage of a mutant allele of Teosinte branched 1 (tb1-ref) on the root system of modern maize. BMC Genetics 2014, 15, 23. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Shi, J.; Sun, C.; Gong, H.; Fan, X.; Qiu, F.; Huang, X.; Feng, Q.; Zheng, X.; Yuan, N.; et al. Gene duplication confers enhanced expression of 27-kDa γ-zein for endosperm modification in quality protein maize. Proc Natl Acad Sci U S A 2016, 113, 4964–4969. [Google Scholar] [CrossRef]
- Hu, Y.; Ren, J.; Peng, Z.; Umana, A.A.; Le, H.; Danilova, T.; Fu, J.; Wang, H.; Robertson, A.; Hulbert, S.H.; et al. Analysis of Extreme Phenotype Bulk Copy Number Variation (XP-CNV) Identified the Association of rp1 with Resistance to Goss's Wilt of Maize. Frontiers in Plant Science 2018, 9. [Google Scholar] [CrossRef]
- Wingen, L.U.; Münster, T.; Faigl, W.; Deleu, W.; Sommer, H.; Saedler, H.; Theißen, G. Molecular genetic basis of pod corn Tunicate maize). Proceedings of the National Academy of Sciences 2012, 109, 7115–7120. [Google Scholar] [CrossRef]
Figure 1.
Digital droplet PCR identification of copy number variation of ZmLOX5 in B73 (1×LOX5), Yu796 (2×LOX5), B73-2×LOX5 genetic backgrounds of maize. B73 inbred line carries 2 functional copies of the ZmLOX5 gene on the two homologous chromosomes, Yu796 inbred line carries 4 functional copies of ZmLOX5 (designated as 2×LOX5) and three independent near-isogenic lines of B73 carrying 4 functional copies of ZmLOX5 gene from Yu796, designated B73-2×LOX5, at the BC6F3 genetic stage. House-keeping single-copy gene, α-Tubulin (Zm00001eb215710), was chosen as an internal reference. A, one-dimensional plots of droplets measured for fluorescence signal (amplitude indicated on y-axis) emitted from the droplets containing either ZmLOX5 or α-Tubulin from each individual. Evergreen™-bound gene-positive droplets are shown in blue, while negative droplets are shown in grey. Top panel, fluorescence amplitudes of the target gene, ZmLOX5; bottom panel, amplitudes of the reference gene, α-Tubulin. B, template concentration (copies / μl) of the target gene, ZmLOX5 (black bars), and reference gene, α-Tubulin (grey bars) in B73 and Yu796 inbred lines, and in three independent near-isogenic lines B73-2×LOX5, determined by ddPCR. C, calculated ZmLOX5 copy numbers using the ratio of the concentrations of the target gene and reference gene (ZmLOX5/α-Tubulin).
Figure 1.
Digital droplet PCR identification of copy number variation of ZmLOX5 in B73 (1×LOX5), Yu796 (2×LOX5), B73-2×LOX5 genetic backgrounds of maize. B73 inbred line carries 2 functional copies of the ZmLOX5 gene on the two homologous chromosomes, Yu796 inbred line carries 4 functional copies of ZmLOX5 (designated as 2×LOX5) and three independent near-isogenic lines of B73 carrying 4 functional copies of ZmLOX5 gene from Yu796, designated B73-2×LOX5, at the BC6F3 genetic stage. House-keeping single-copy gene, α-Tubulin (Zm00001eb215710), was chosen as an internal reference. A, one-dimensional plots of droplets measured for fluorescence signal (amplitude indicated on y-axis) emitted from the droplets containing either ZmLOX5 or α-Tubulin from each individual. Evergreen™-bound gene-positive droplets are shown in blue, while negative droplets are shown in grey. Top panel, fluorescence amplitudes of the target gene, ZmLOX5; bottom panel, amplitudes of the reference gene, α-Tubulin. B, template concentration (copies / μl) of the target gene, ZmLOX5 (black bars), and reference gene, α-Tubulin (grey bars) in B73 and Yu796 inbred lines, and in three independent near-isogenic lines B73-2×LOX5, determined by ddPCR. C, calculated ZmLOX5 copy numbers using the ratio of the concentrations of the target gene and reference gene (ZmLOX5/α-Tubulin).
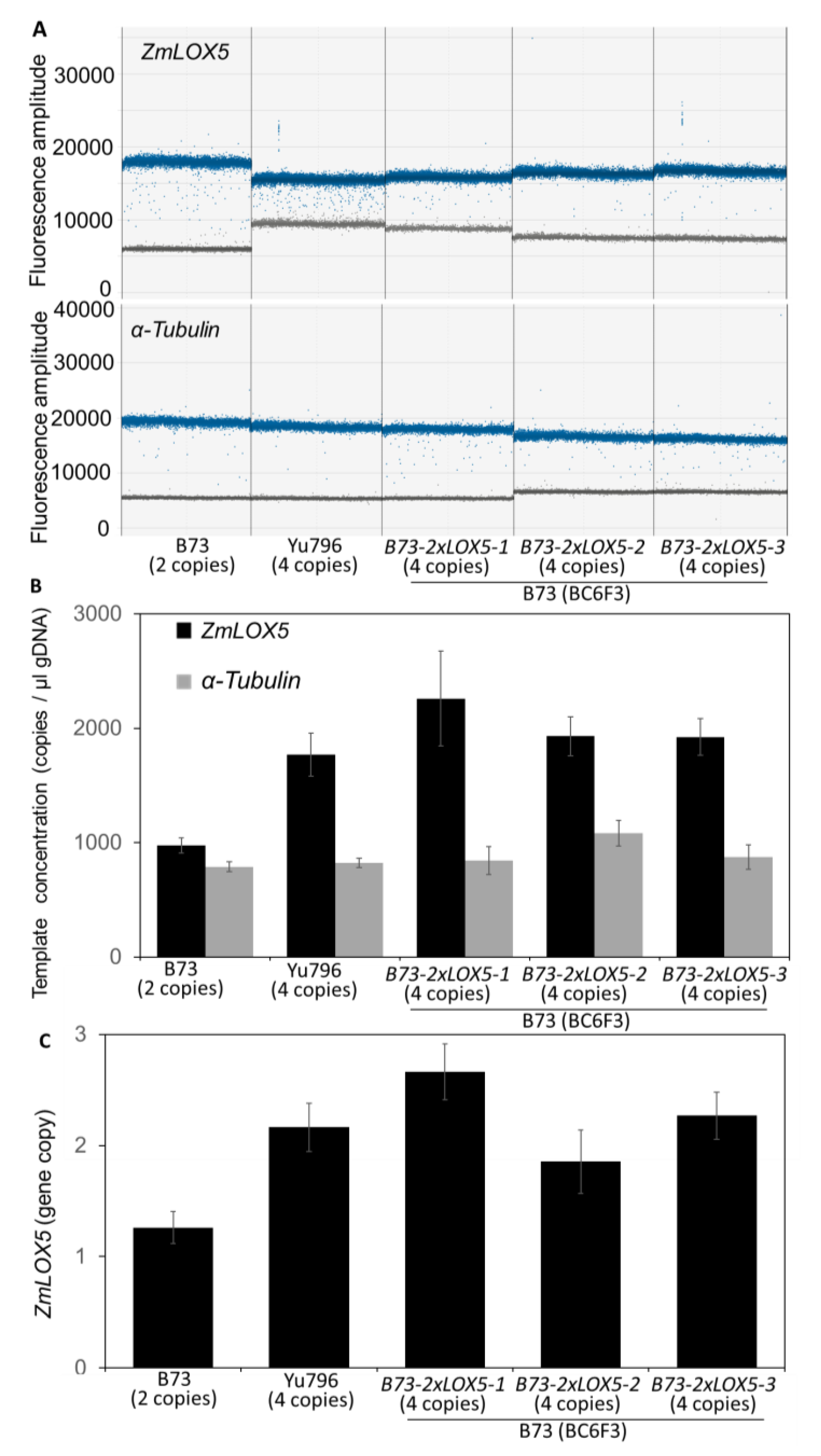
Figure 2.
Alignment of the nucleotide sequences of the 8th intron of the ZmLOX5 gene in B73 and the copy number variants (CNV) of ZmLOX5 in Yu796 (Yu796-CNV1 and Yu796-CNV2). Upper panel: genomic structure of the ZmLOX5 gene containing 9 exons (shown in blue color), 8 introns (shown in black color), and 5’-UTR and 3’-UTR (shown in green color). ZmLOX5-specific primers gDL-F and gDL-R (shown in red color) cover partially exon VIII, complete exon IX and 3’UTR, and complete 8th intron. Sequences highlighted in yellow color represent the nucleotide differences between the B73-LOX5 and Yu796-2×LOX5 locus. Nucleotides shown in red color represents the difference between the Yu796-CNV1 and Yu796-CNV2 locus.
Figure 2.
Alignment of the nucleotide sequences of the 8th intron of the ZmLOX5 gene in B73 and the copy number variants (CNV) of ZmLOX5 in Yu796 (Yu796-CNV1 and Yu796-CNV2). Upper panel: genomic structure of the ZmLOX5 gene containing 9 exons (shown in blue color), 8 introns (shown in black color), and 5’-UTR and 3’-UTR (shown in green color). ZmLOX5-specific primers gDL-F and gDL-R (shown in red color) cover partially exon VIII, complete exon IX and 3’UTR, and complete 8th intron. Sequences highlighted in yellow color represent the nucleotide differences between the B73-LOX5 and Yu796-2×LOX5 locus. Nucleotides shown in red color represents the difference between the Yu796-CNV1 and Yu796-CNV2 locus.
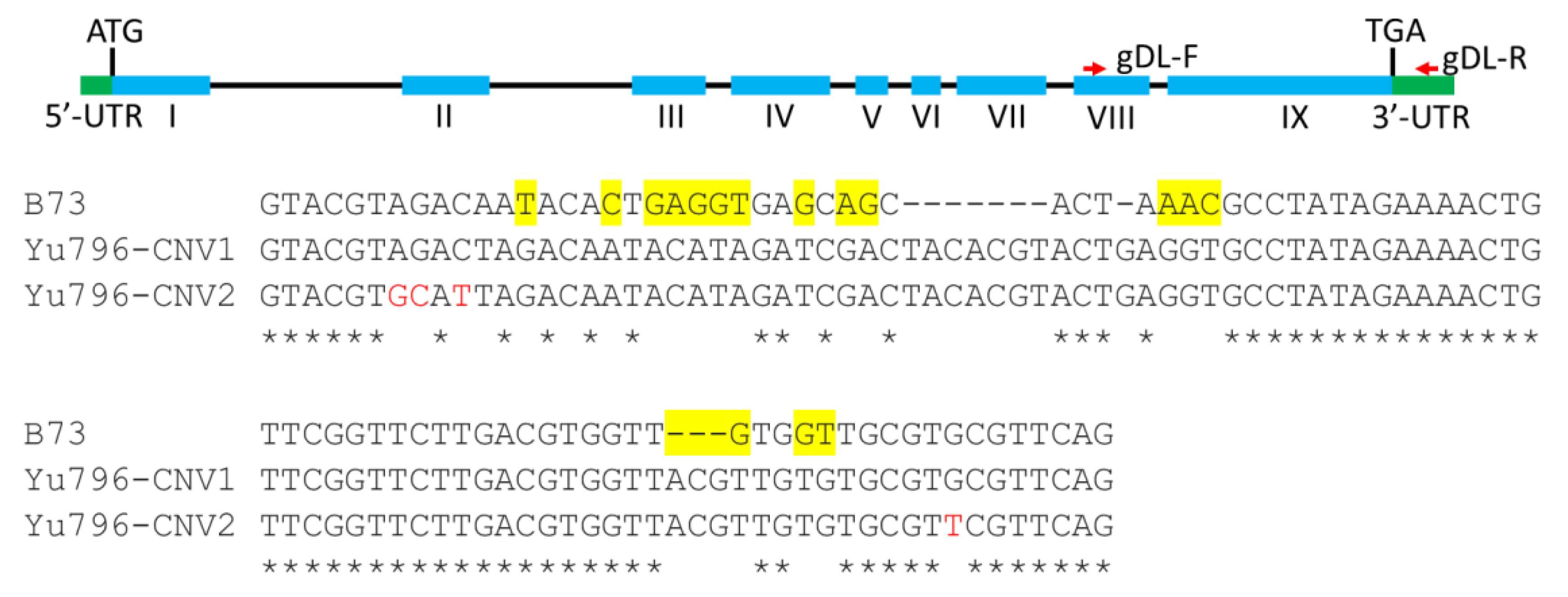
Figure 3.
Duplication of ZmLOX5 in the B73 background results in enhanced expression of ZmLOX5 and selected 12-OPDA and JA biosynthesis genes in response to wounding compared to the B73 near isogenic line containing a single gene. RT-qPCR analysis of wound-induced gene expression of ZmLOX5, ZmLOX8, ZmLOX10 and ZmJAR1a in B73 line and in the near-isogenic lines containing duplicated ZmLOX5 CNVs in the B73 genetic background (B73-2×LOX5) at the BC6F3 stage at 0, 1, 2 and 4 hours post wounding (hpw). The 2-ΔΔCt method was used with α-tubulin utilized as the house-keeping gene and compared to the mean at 0 h. For each graph, the X-axis represents hpw while the Y-axis represents expression fold change; black line represents the B73-2×LOX5 line (4 copies of ZmLOX5) and green line represents B73 (2 copies of ZmLOX5), bars are means ± SE; n=4. Asterisks represent significant differences between B73-2×LOX5 and B73 lines at each time point using Student’s t-test (*p < 0.05 and **p < 0.01).
Figure 3.
Duplication of ZmLOX5 in the B73 background results in enhanced expression of ZmLOX5 and selected 12-OPDA and JA biosynthesis genes in response to wounding compared to the B73 near isogenic line containing a single gene. RT-qPCR analysis of wound-induced gene expression of ZmLOX5, ZmLOX8, ZmLOX10 and ZmJAR1a in B73 line and in the near-isogenic lines containing duplicated ZmLOX5 CNVs in the B73 genetic background (B73-2×LOX5) at the BC6F3 stage at 0, 1, 2 and 4 hours post wounding (hpw). The 2-ΔΔCt method was used with α-tubulin utilized as the house-keeping gene and compared to the mean at 0 h. For each graph, the X-axis represents hpw while the Y-axis represents expression fold change; black line represents the B73-2×LOX5 line (4 copies of ZmLOX5) and green line represents B73 (2 copies of ZmLOX5), bars are means ± SE; n=4. Asterisks represent significant differences between B73-2×LOX5 and B73 lines at each time point using Student’s t-test (*p < 0.05 and **p < 0.01).
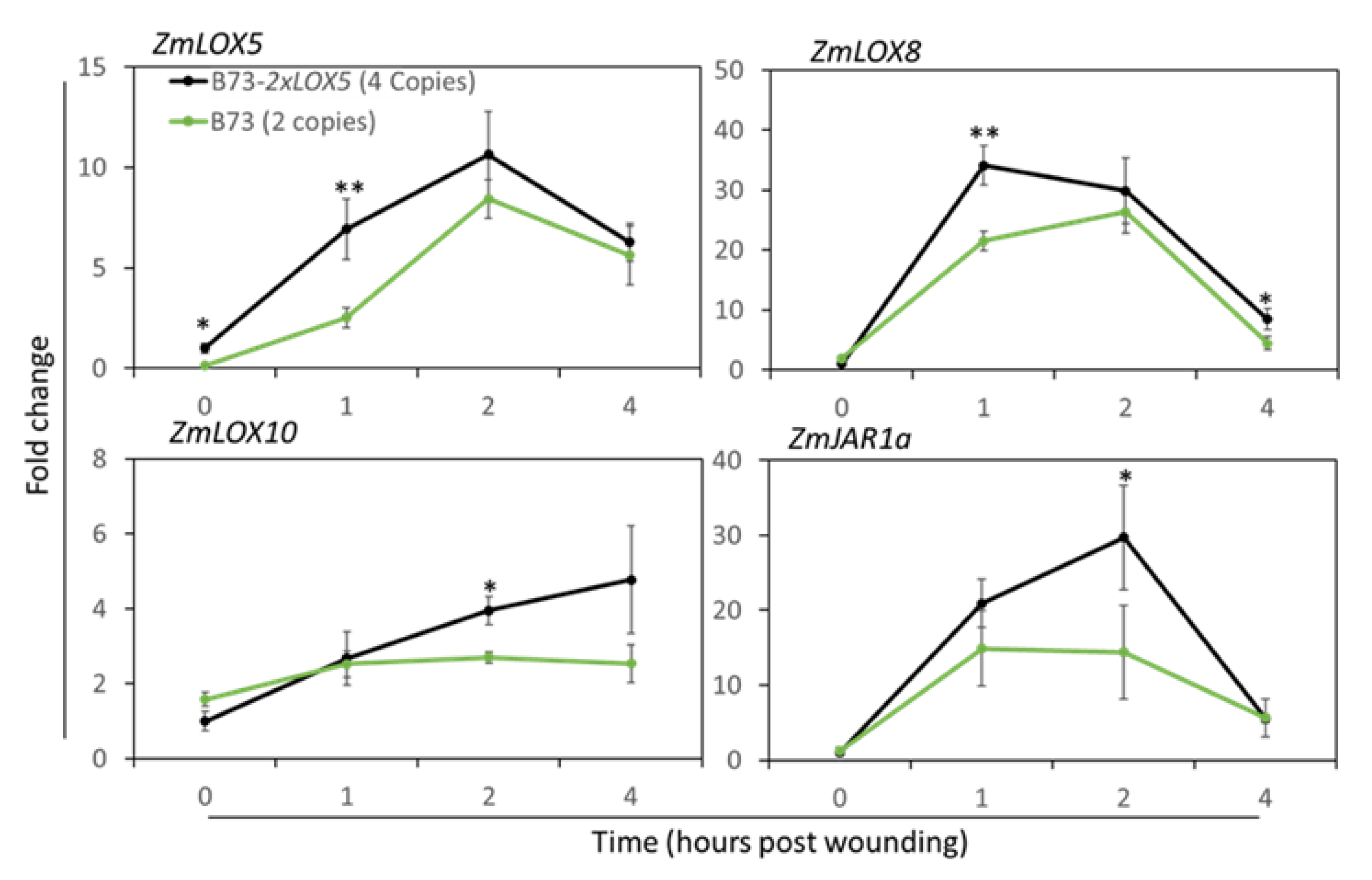
Figure 4.
Increased gene dosage of ZmLOX5 results in increased resistance to fall armyworms (FAW). A and B. Effect of ZmLOX5 duplication on FAW resistance during localized herbivory in the homozygous seedlings (2×LOX5/2×LOX5, 4 copies), heterozygous seedlings (2×LOX5/lox5-3, 2 copies), knockout mutants (lox5-3/lox5-3, 0 copy) and B73 (2 copies). Second to third instar larvae were confined to clip-cages positioned on the maize leaf and allowed to feed for 6-8 h. Area consumed was determined by ImageJ analysis of scanned leaves. (C-F). The individual seedlings as in A and B were exposed to continued feeding by 8 neonates per seedling for 7 days. Afterwards, seedlings were photographed (C) and weighed (D), bars are means ± SE; n=6-7; the recovered larvae were photographed (E), and the fresh weight of recovered larvae were weighed (F). Bars are means ± SE; n=16-20. Groups with non-overlapping letters represent statistically significance differences (p<0.05) by one-way ANOVA with Tukey HSD post-hoc test.
Figure 4.
Increased gene dosage of ZmLOX5 results in increased resistance to fall armyworms (FAW). A and B. Effect of ZmLOX5 duplication on FAW resistance during localized herbivory in the homozygous seedlings (2×LOX5/2×LOX5, 4 copies), heterozygous seedlings (2×LOX5/lox5-3, 2 copies), knockout mutants (lox5-3/lox5-3, 0 copy) and B73 (2 copies). Second to third instar larvae were confined to clip-cages positioned on the maize leaf and allowed to feed for 6-8 h. Area consumed was determined by ImageJ analysis of scanned leaves. (C-F). The individual seedlings as in A and B were exposed to continued feeding by 8 neonates per seedling for 7 days. Afterwards, seedlings were photographed (C) and weighed (D), bars are means ± SE; n=6-7; the recovered larvae were photographed (E), and the fresh weight of recovered larvae were weighed (F). Bars are means ± SE; n=16-20. Groups with non-overlapping letters represent statistically significance differences (p<0.05) by one-way ANOVA with Tukey HSD post-hoc test.
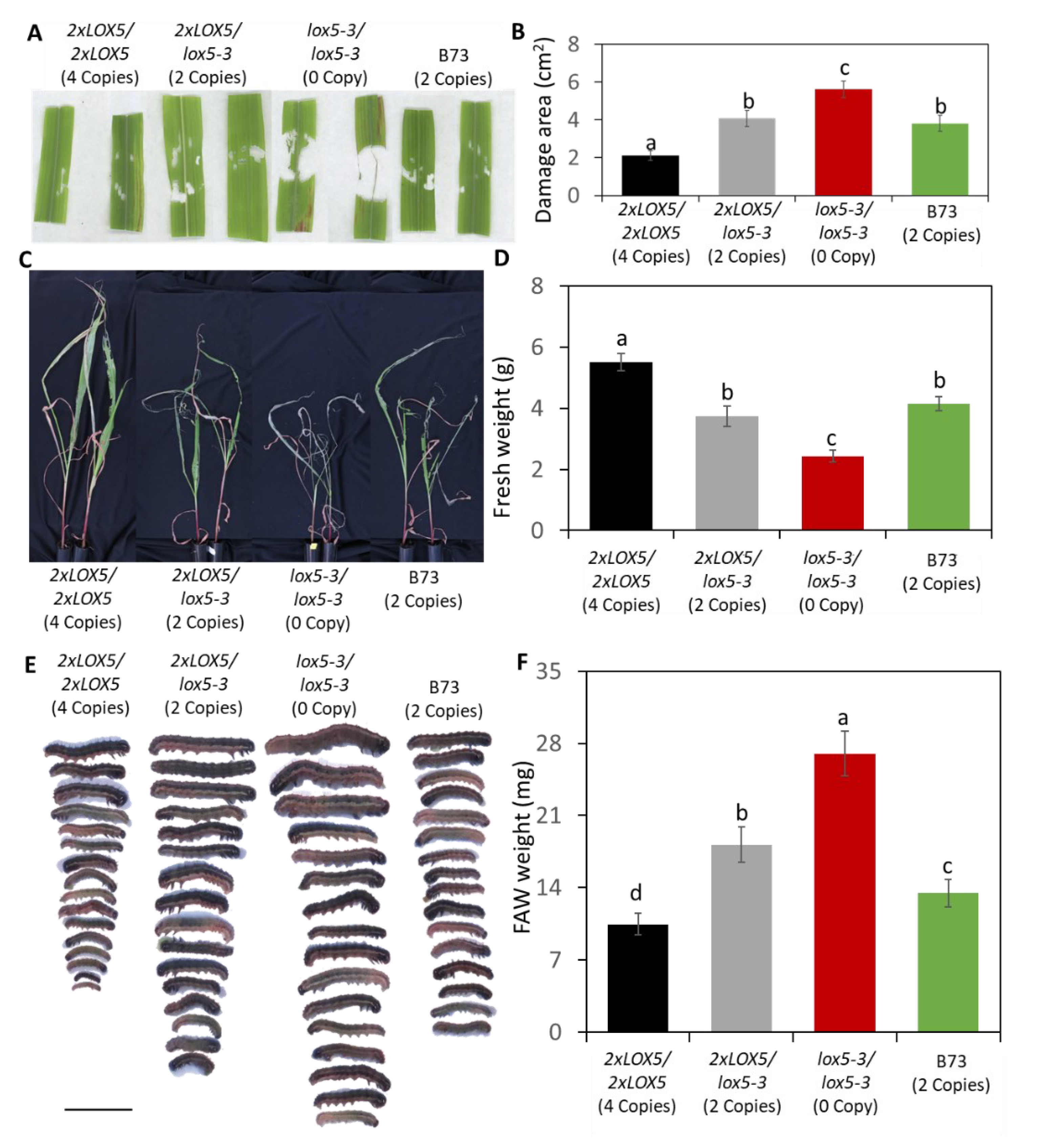
Figure 5.
Duplication of ZmLOX5 enhanced 9,10-KODA, 12-OPDA, and ABA, but not JA-Ile, accumulation in response to mechanical wounding. For each graph, the X-axis represents hpw and the Y-axis represents metabolite concentration in pmol/g FW for each metabolite (JA-Ile, 12-OPDA, 9,10-KODA and ABA) indicated within the graphs. Black line represents duplicated copy variant (2×ZmLOX5/2×ZmLOX5, 4 copies of ZmLOX5 in the B73 genetic background at the BC6F3 genetic stage) and green line represents B73 inbred line (2 copies of ZmLOX5); bars are means ± SE; n=4-5; asterisks represent significant differences near-isogenic 2×LOX5 and B73-LOX5 at each time point using Student’s t-test (*p < 0.05 and **p < 0.01).
Figure 5.
Duplication of ZmLOX5 enhanced 9,10-KODA, 12-OPDA, and ABA, but not JA-Ile, accumulation in response to mechanical wounding. For each graph, the X-axis represents hpw and the Y-axis represents metabolite concentration in pmol/g FW for each metabolite (JA-Ile, 12-OPDA, 9,10-KODA and ABA) indicated within the graphs. Black line represents duplicated copy variant (2×ZmLOX5/2×ZmLOX5, 4 copies of ZmLOX5 in the B73 genetic background at the BC6F3 genetic stage) and green line represents B73 inbred line (2 copies of ZmLOX5); bars are means ± SE; n=4-5; asterisks represent significant differences near-isogenic 2×LOX5 and B73-LOX5 at each time point using Student’s t-test (*p < 0.05 and **p < 0.01).
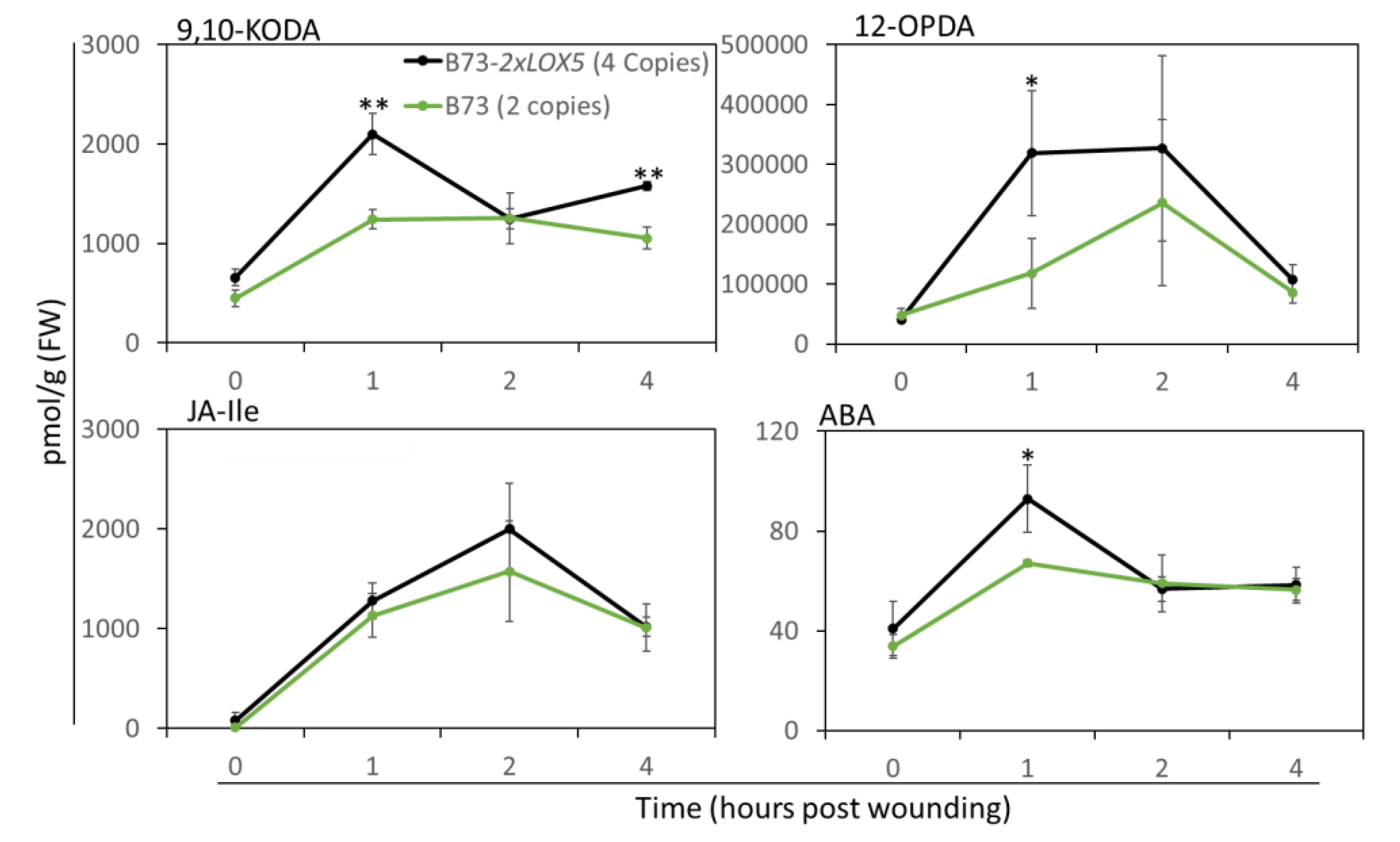
Figure 6.
JA catabolites, 12OH-JA, 12OH-JA-Ile and 12COOH-JA-Ile, were significantly increased in the B73-2×LOX5 seedlings in response to mechanical wounding. A. (A) Heatmap showing relative accumulation of 9-oxylipins, 13-oxylipins and ABA in B73-2×LOX5, duplicated copy variant (2×ZmLOX5/2×ZmLOX5, 4 copies of ZmLOX5 in the B73 genetic background at the BC6F3 genetic stage) and B73 inbred line (2 copies of ZmLOX5) at 1, 2 and 4 h post wounding (hpw) compared to unwounded B73. Cells are shaded by Z-score scaling within columns, with the higher-abundant metabolite in red and the lower-abundant metabolite in blue; B. For each graph, the X-axis represents hpw and the Y-axis represents metabolite concentration in pmol / g FW for each JA catabolites (12OH-JA, 12OH-JA-Ile and 12COOH-JA-Ile) indicated within the graphs. Black line represents duplicated copy variant (2×ZmLOX5/2×ZmLOX5, 4 copies of ZmLOX5 in the B73 genetic background at the BC6F3 genetic stage) and green line represents B73 inbred line (2 copies of ZmLOX5); bars are means ± SE; n=4-5; asterisks represent significant differences near-isogenic 2xLOX5 and B73-LOX5 at each time point using Student’s t-test (*p < 0.05 and **p < 0.01).
Figure 6.
JA catabolites, 12OH-JA, 12OH-JA-Ile and 12COOH-JA-Ile, were significantly increased in the B73-2×LOX5 seedlings in response to mechanical wounding. A. (A) Heatmap showing relative accumulation of 9-oxylipins, 13-oxylipins and ABA in B73-2×LOX5, duplicated copy variant (2×ZmLOX5/2×ZmLOX5, 4 copies of ZmLOX5 in the B73 genetic background at the BC6F3 genetic stage) and B73 inbred line (2 copies of ZmLOX5) at 1, 2 and 4 h post wounding (hpw) compared to unwounded B73. Cells are shaded by Z-score scaling within columns, with the higher-abundant metabolite in red and the lower-abundant metabolite in blue; B. For each graph, the X-axis represents hpw and the Y-axis represents metabolite concentration in pmol / g FW for each JA catabolites (12OH-JA, 12OH-JA-Ile and 12COOH-JA-Ile) indicated within the graphs. Black line represents duplicated copy variant (2×ZmLOX5/2×ZmLOX5, 4 copies of ZmLOX5 in the B73 genetic background at the BC6F3 genetic stage) and green line represents B73 inbred line (2 copies of ZmLOX5); bars are means ± SE; n=4-5; asterisks represent significant differences near-isogenic 2xLOX5 and B73-LOX5 at each time point using Student’s t-test (*p < 0.05 and **p < 0.01).
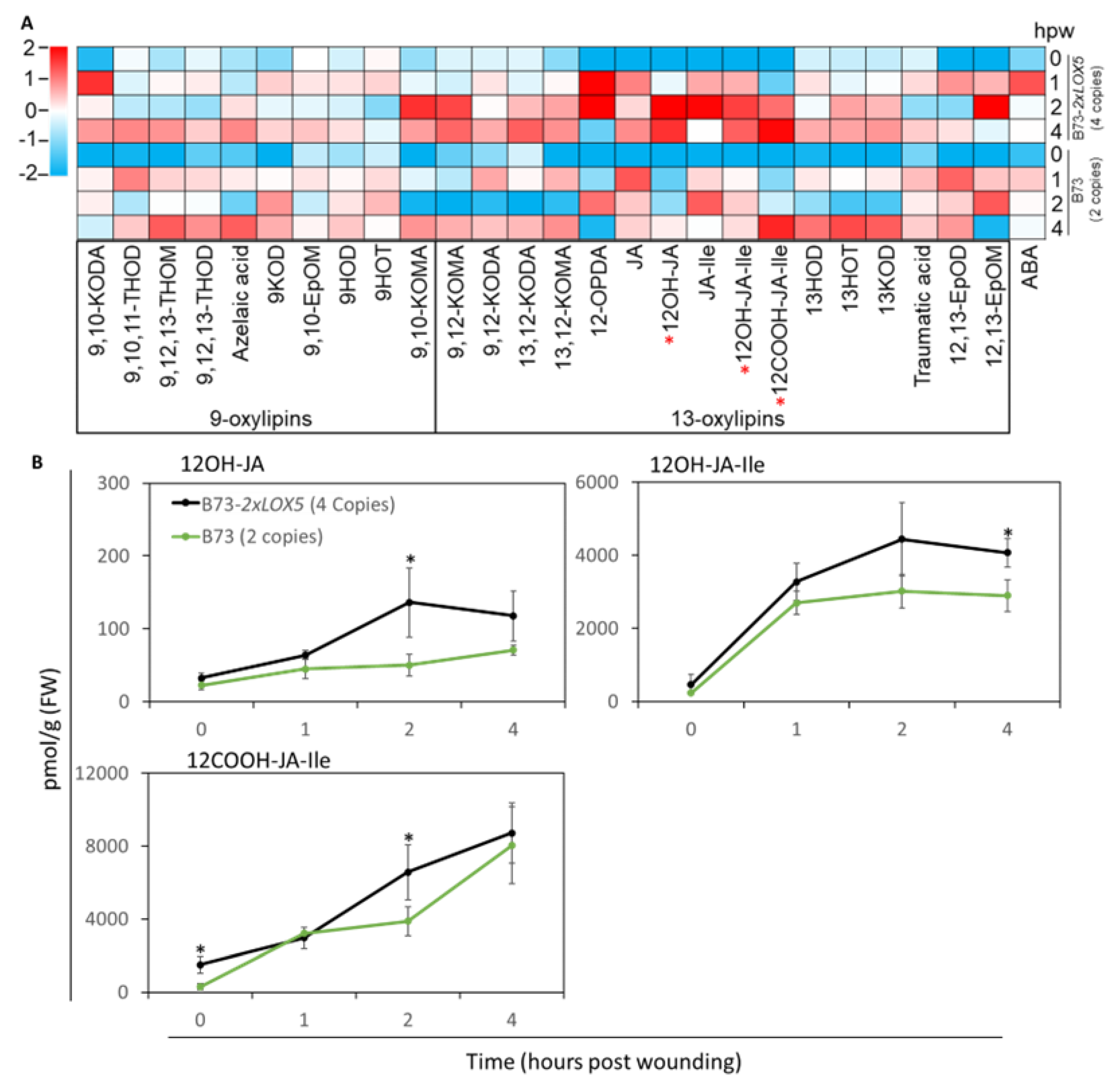

Table 1.
Duplicated copy variant of ZmLOX5 (2×LOX5) segregates in the mendelian ratio consistent with a single locus, confirming that the two copies are linked as tandem duplicates, designated as copy number variant 1 (LOX5-CNV1) and 2 (LOX5-CNV2).
Table 1.
Duplicated copy variant of ZmLOX5 (2×LOX5) segregates in the mendelian ratio consistent with a single locus, confirming that the two copies are linked as tandem duplicates, designated as copy number variant 1 (LOX5-CNV1) and 2 (LOX5-CNV2).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



