
Submitted:
28 February 2024
Posted:
29 February 2024
You are already at the latest version
Abstract
Abstract: The venom-containing barb attached to their ‘whip-like’ tail provides stingrays a defensive mechanism for evading predators such as sharks. From human encounters, dermal stingray envenomation is characterized by intense pain often followed by tissue necrosis occurring over several days to several weeks. The bioactive components in stingray venoms (SRV’s) and their molecular targets and mechanisms that mediate these complex responses are not well understood. Given the utility of venom-derived proteins from other venomous species for biomedical and pharmaceutical applications, we set out to characterize the bioactivity of SRV from three local species that belong to the Dasyatoidea ‘whiptail’ superfamily. Multiple cell-based assays were used to quantify and compare the in vitro effects of these SRV’s on different cell lines. All three SRV’s demonstrated concentration-dependent growth inhibitory effects on three different human cell lines tested. In contrast, a mouse fibrosarcoma cell line was markedly resistant to all three SRV’s, indicating the molecular target(s) for mediating the SRV effects are not expressed on these cells. The multifunctional SRV responses were characterized by an acute disruption of cell adhesion leading to apoptosis. These findings aim to guide future investigations of identified SRV proteins and their molecular targets for potential use in biomedical applications.
Keywords:
apoptosis
; cell adhesion
; cell growth
; fibrosarcoma
; galectin-like proteins
; stingray
; toxin
; venom
1. Introduction
Venoms are complex mixtures of bioactive secretions that have evolved independently numerous times throughout the animal kingdom in both invertebrate and vertebrate phyla [1,2]. Each venom is composed of numerous compounds that possess a variety of bioactivities (i.e., proteolytic, hemotoxic, neurotoxic or cytotoxic activity), and collectively serve a variety of biological functions for the host animal including predation, defense from predators, and competitor deterrence [1,2]. Although several unique venom compounds have been developed into therapeutic drugs, including blood pressure regulators, blood thinners (anticoagulants), and pain relievers associated with specific ion channel blockers, none to date have originated from stingray venom (SRV) [3,4,5,6].
Unlike most venomous animals whose venom glands are distinct well-defined organs, SRV secretion emanates from diffuse masses of cells that lie between the epidermis and dermis along either side of the midventral ridge of a serrated cartilaginous barb [7,8,9,10,11]. Stingray venom glands are holocrine glands, where venom secretion results from physical rupture and disintegration of the venom-containing gland cells when the integumentary sheath is shed from the barb during the mechanical whip-like action of the tail [12,13]. Obtaining the diffuse crude venom secretions from stingray barbs is challenging and provides a likely reason why SRV’s have been less studied compared to other venomous species.
Bioassays to screen and characterize venom activity typically measure changes in cell viability (toxicity), migration, and growth using biochemical markers with colorimetric assays and impedance-based biosensing techniques [14,15]. Applying these methods to cultured cells derived from tissues to which venom compounds are typically exposed (such as skin, nerve, muscle, and blood), provides an initial assessment and characterization of SRV bioactivities. Proteomic and transcriptomic approaches are now extending these approaches by identifying the complement of expressed SRV proteins and peptides attributable to known venom functions that then holistically contribute to the physiological responses elicited by crude venom extracts [16,17,18,19].
Our study reported here characterizes the bioactive properties of SRV collected from three local species of stingrays commonly found in nearshore waters off the southwest coast of Florida; the Atlantic stingray (Hypanus sabinus, Dasyatidae family), the spotted eagle ray (Aetobatus narinari, Aetobatidae family), and the cownose ray (Rhinoptera bonasus, Rhinopteridae family). The selected target cells for characterizing the SRV effects represent cell-types derived from tissues to which these SRV’s might normally respond (i.e., skin, blood, and neural origin); specifically they include a human dermal fibroblast cell line (HDFa), a human neuroblastoma cell line (SH-SY5Y), a human T-cell leukemia cell line (Jurkat E6-1), and a murine fibrosarcoma cell line (WEHI-164) (cf. Table 1).
2. Results
2.1. Biochemical Characterization of Isolated SRV Proteins
Preparation of enriched SRV protein extracts from collected stingray spines were standardized to enable the assessment of possible species differences. Crude SRV protein samples from the Atlantic stingray, spotted eagle ray, and cownose ray were initially evaluated and compared by SDS-PAGE analysis (Figure 1). The resolved SRV proteins from each species ranged in molecular size from ~15 kD to greater than 250 kD, where the banding patterns were similar but species specific. Major protein bands corresponding to approximately 16 kD, 38 kD, and 75 kD, were present in all three venoms.
2.2. Bioactive Properties of SRV’s on Cultured HDFa and SH-SY5Y Cells
To screen the bioactivity of SRV’s on cultured HDFa and SH-SY5Y cells in vitro, cell cultures were treated with a range of SRV concentrations for a 72-hour exposure period and then assayed for cell growth using the MTT assay [15]. The concentration-dependent effects of Atlantic stingray venom and spotted eagle ray venom are shown in Figure 2 and presented as ‘percent growth inhibition’ relative to untreated control cells. Both SRV’s inhibited cell growth when compared to untreated control groups and were dependent on the concentration of SRV tested.
For HDFa cells exposed to Atlantic stingray venom proteins (Figure 2A), all concentrations caused a significant reduction in cell growth compared to the untreated control cells (P<0.001). The calculated concentration producing 50% inhibition (EC50) was 98.6 μg/mL. For HDFa cells treated with spotted eagle ray venom (Figure 2B), concentrations of 200 µg/mL and 400 µg/mL caused a significant reduction in cell growth compared to untreated control cells (P=0.011, 0.043, respectively), however the lower concentrations tested (50 and 100 µg/mL) did not produce a statistically significant difference. The calculated EC50 for spotted eagle ray venom mediated cell growth inhibition of HDFa cells was 116.8 μg/mL.
For the neuronally-derived SH-SY5Y cells (Figure 2C), the growth inhibition caused by 50, 100, 200 µg/mL spotted eagle ray venom were each significantly greater than untreated control (P=0.004, 0.002, and 0.002, respectively), where 25 µg/mL treatment was not significantly different from untreated control cells. The calculated EC50 for spotted eagle ray venom mediated cell growth inhibition of SH-SY5Y cells was 46.9 μg/mL. Taken altogether, both cell lines exhibited similar sensitivities to SRV-mediated growth inhibition as determined by the MTT assay.
2.3. Concentration Dependent Effects of SRV on Cell Morphology
The effects of SRV treatment on cell morphological features could also be readily observed under the microscope and were evident at the lowest SRV concentrations tested, 25 μg/mL (Figure 3). For both HDFa and SH-SY5Y cells, a decrease in cell density and an increase in small, round cells were readily visualized with increasing spotted eagle ray venom concentrations. At the two higher concentrations (100 and 200 µg/mL), very few intact cells were observed, and cells mainly consisted of small, rounded cells and cellular debris. These morphological features are consistent with cells undergoing apoptosis.
2.4. Time Course for SRV-Mediated Effects on HDFa Cell Cultures
To better resolve the time course for SRV mediated effects on cell growth, biosensor-based cell impedance analysis was performed on cultured HDFa cells treated with Atlantic stingray venom at varying concentrations. After initial cell seeding in the biosensor culture wells, cell growth was recorded every 15 min over a 3-day pre-treatment period. Shown in Figure 4, the near linear increase in impedance during the pre-treatment period reflects the time course for cell proliferation and an increase in adherent HDFa cells (i.e., rate of cell growth). On day 4, cells were exposed to varying concentrations of SRV and the impedance recorded for a 48-hour treatment period. Under the culturing conditions used, HDFa cells entered near log phase growth around day 3-4 when the SRV treatment was initiated (Figure 4).
Time course analysis of the 2-day SRV treatment period revealed two distinct effects caused by SRV treatment that were dependent on the concentration and exposure time. At the higher SRV concentrations tested (30-100 μg/mL), there was a distinct acute effect that was characterized by a very rapid decrease in impedance (starting immediately) that was complete after 1 hour of treatment and reflects a decrease in cell adherence (i.e., cell detachment). The rapid decrease caused by high SRV concentrations resembled the time course caused by Triton-X treatment that was used as a positive control for acute cell detachment. The EC50 for the rapid cell detachment effect cause by high SRV concentrations, measured at 24 hours of treatment, was approximately 65 μg/mL (Figure 4B).
At the lower SRV concentrations that did not produce rapid cell detachment (i.e., 3-10 μg/mL), there was a notably slower rate of cell growth over the full 48-hour SRV exposure period. This finding is consistent with the inhibition of cell growth observed in the MTT assay following 72-hour SRV exposures (cf. Figure 3). The EC50 for SRV-mediated growth inhibition at 48 hours was approximately 20 μg/mL (Figure 4B).
The bioactivity of SRV’s are known to be thermolabile, where applying hot water to the site of injury (up to 45 ˚C for 30-90 minutes) is recommended to mitigate the acute pain and tissue necrosis associated with stingray envenomation [20,21,22]. To test the thermolability of SRV in our experiments, Atlantic stingray venom was subjected to high temperature (100 ˚C) for 10 minutes to denature the venom protein components. As shown in Figure 5, heat-treatment abolished the rapid cell detachment effect caused by 100 μg/mL SRV. In contrast, the growth inhibitory effect of the SRV was still present after heat treatment (Figure 5B). These findings indicate protein component(s) in Atlantic stingray venom mediate the rapid cell detachment effect (heat-sensitive), whereas other venom components insensitive to heat can contribute towards the growth inhibitory effect.
2.5. Bioactive Effects of SRV’s on Additional Tumor Cell Types
To further explore the effects of SRV on different tumor cell types, Jurkat E6-1 cells, a non-adherent human T lymphocyte tumor cell line was examined in addition to a chemically induced mouse fibrosarcoma cell line (WEHI-164 cells). Using the MTT cell growth assay, the comparative effects of spotted eagle ray, Atlantic stingray, and cownose ray venoms are shown in Figure 6. All three SRV’s demonstrated significant growth inhibitory effects on Jurkat E6-1 cells (Figure 6A), and generally had the following rank order of potency; spotted eagle ray > Atlantic stingray > cownose ray. Venom from the Atlantic stingray, at 50 µg/mL (P=0.0122), 100 µg/mL (P=0.004), and 200 µg/mL (P=0.008) treatment concentrations, were each significantly different from the untreated control (OD comparisons). With eagle ray venom, 100 µg/mL (P=0.083) and 200 µg/mL (P=0.035) treatments were significantly different from the untreated control. Cownose ray venom caused a significant difference at 200 µg/mL (P=0.044) compared to the untreated control. The SRV concentration dependencies approximated the growth inhibition observed for both HDFa and SH-SY5Y cells (cf. Figure 2).
Interestingly, however, the mouse WEHI 164 fibrosarcoma cell line was found to be insensitive to all three SRV’s tested following the 24-hour exposure period (Figure 6B). Using the same SRV concentrations tested in the other cell lines, no venom treatments resulted in significantly different WEHI 164 cell growth when compared to untreated controls.
2.6. SRV’s Cause Apoptosis of Jurkat E6 Cells
To further explore the growth inhibitory effects of SRV’s on Jurkat E6-1 cells, these non-adherent cells were assayed for apoptosis and necrosis using flow cytometry [23]. Jurkat E6-1 cells were exposed for 24 hours to either spotted eagle ray venom or Atlantic stingray venom at varying concentrations, then assayed for apoptosis and cell death. The results shown in Figure 7 illustrate the SRV concentration-dependent decrease in healthy cells and concomitant increase in apoptosis with both SRV treatments. The percentage of cells demonstrating necrosis (dead cells) did not increase with increasing SRV concentration, indicating the 24-hour treatment period preferentially induced apoptosis. The threshold SRV concentrations producing apoptosis were similar for spotted eagle ray venom and Atlantic stingray venom (~10 μg/mL), however, Atlantic stingray venom caused significantly greater apoptosis compared to spotted eagle ray venom at 100 µg/mL (P=0.035) and 200 µg/mL (P=0.0078).
3. Discussion
3.1. The Evolution of SRV’s as a Well-Conserved Defense Response
The evolutionary emergence of stingray spines dates back to the Late Cretaceous with expansion occurring in the Late epoch, 100.5 to 66 million years ago [24]. The temporal correlation with the expansion of large sharks during this time period suggests that predatory pressures on early rays led to the adaptive development and resulting venom-laced barb that is characteristic of today’s Myliobatoidei suborder [25]. The venomous spine appendage is found worldwide on ~220 stingray species inhabiting both fresh and saltwater habitats. Since the emergence of SRV as a noxious mixture of compounds to repel predators, its conserved and non-conserved components across different stingray species are currently not well understood.
The noxious and mostly non-lethal experience of stingray envenomation is recognized from human encounters and assumed to be a similar experience for an attacking predator. The defensive tail-whip reflex of the stingray causes two distinct events to their targets; 1) dermal laceration caused by the serrated barb that provides deep tissue access locally and to the microcirculation, and 2) envenomation of SRV components into the penetrated tissue via holocrine release from venom glands located on the barb [8,10]. Envenomation is typically localized but can be systemic depending on the anatomical location of the mechanical impalement and extent of venom deposition. Lethal outcomes are rare in humans and generally occur with systemic envenomation impacting cardiovascular system function [26].
3.2. The Bioactivity of Isolated SRV’s Is Well-Conserved across Diverse Species
We examined the properties of SRV isolated from three species belonging to three distinct stingray families (Dasyatidae, Aetobatidae, and Rhinopteridae) [27]. The bioactivity of their SRV highlights the conserved activity across this large and diverse family of whiptail stingrays. Our standardized procedures for collecting, processing, and assaying SRV bioactivity on mammalian cells in culture revealed consistent and reproducible responses produced by each venom. Based on our findings, the sequelae of events associated with SRV exposure initiates immediately with cellular events that disrupt cell adhesion leading to cell detachment, then followed by growth inhibition and apoptosis.
The biochemical profiles from SDS-PAGE separation of the crude SRV extracts indicate the presence of both conserved proteins and proteins that are species-specific. Other lower abundance venom proteins and small venom peptides are likely to be present in our samples but were not resolved in the SDS-PAGE profiles. In addition, the protein samples were derived from ventral spine tissue that includes both the venom producing secretory cells as well as other neighboring connective tissue that is unavoidably included with the collection method. We therefore cannot associate the origin of the biochemically identified proteins exclusively with venom proteins produced by the venom glands cells. Nevertheless, the SRV protein samples we examined are clearly enriched in venom proteins based on the collection method used and their demonstrated bioactivities.
3.3. SRV Proteins Identified by Recent Venomic Studies
SRV has historically been viewed as a “neglected venom” given the difficulty in obtaining significant amounts of venom for investigation. With the advent and application of multi-omic technologies, resolving the constellation of proteins in SRV tissue extracts has become increasingly feasible. Using proteomic and transcriptomic analysis of crude venom isolated from the marine, blue-spotted stingray Neotrygon kuhlii, 18 distinctive proteins were identified [16]. Two cystatin proteins (cystatin-1 and cystatin-2) represented the most abundant protein class (19% combined), followed by a galectin-like protein (12%), and then a peroxiredoxin-6 protein (10%). The other 14 proteins comprised the 59% remaining venom protein content. Cystatin proteins are known peptidase inhibitors found in many other venoms and thought to serve protective functions for other venom peptides and proteins. Interestingly, however, cystatin from snake venom has been shown to also inhibit cell growth and migration in mouse melanoma and human hepatocellular carcinoma cell lines in vitro [28]. Galectin proteins are a multifunctional class of proteins and are discussed in greater length below. Peroxiredoxin-6 is a ubiquitous bifunctional enzyme having both glutathione peroxidase and phospholipase A2 (PLA2) activities. Its PLA2 activity is of interest given the well-established toxic effects of PLA2 activity in many other well characterized venoms.
Venomic approaches have also been applied to SRV’s isolated from freshwater stingrays inhabiting South American rivers. These studies reveal a plethora of venom protein components that also identified cystatins and peroxiredoxin-6, where hyaluronidases were found in greatest abundance [17,18]. Hyaluronidases are a highly conserved component present in many venoms that promotes tissue deposition and the spread of other venom components through the degradation of hyaluronic acid, a major glycosaminoglycan component of the extracellular matrix [1,2]. A total of 111 and 115 venom transcripts were identified from two separate freshwater species, Potamotrygon amandae and Potamotrygon falkneri (Potamotrygonidae family), respectively.
Finally, a recent integrative transcriptomic study of SRV’s from three marine and two freshwater stingrays further highlight the complexity of SRV components [19]. Seventy shared venom transcripts were linked to known functional bioactivity clusters that include pain, cardiotoxicity, and hemorrhage, where venom transcripts for bibrotoxin and cholecystotoxin-like proteins were identified as candidate pain-inducing proteins in SRV, acting via G-protein coupled receptor neuronal signaling pathways. The large number of venom components and their predicted distinct functions point towards a holistic envenomation model where clustered components work synergistically to produce the physiological responses that characterize SRV injury (i.e., pain, inflammation, and tissue necrosis).
3.4. Mining SRV Proteins for Molecular Targets and Biomedical Applications
With the large number of SRV components identified by the venomic investigations, ‘next steps’ include studies investigating the bioactive contributions of individual identified SRV components to validate or invalidate their role in the stingray envenomation responses attributed to acute pain, altered hemostasis, and tissue necrosis. Interpreting our SRV bioactivity results in the context of the SRV venomic findings support multiple venom components contributing to the observed in vitro bioactivities. The potential role of SRV galectin-like proteins outlined below provides an example of future directions to functionally assess the role and contribution of individual SRV proteins and peptides in the envenomation biological response.
3.5. The SRV Galectin Toxin Hypothesis
The proteomic identification of a galectin-like protein as the second most abundant protein in N. kuhlii venom, and also reportedly present in teleost venom [11,16], is especially intriguing as a single multifunctional protein component basis on well-established and diverse functions of vertebrate galectins [29,30]. Galectins, members of the larger lectin family of proteins, are known to mediate many of the biological effects observed in our SRV experiments and therefore are poised to be an important SRV component mediating biological responses associated with envenomation [29,30].
We propose the following hypothesis to serve as a working and testable model for future investigation. This model emanates from our observations of three different SRV’s and their bioactivity profiles aligning with known galectin functions, together with the unexpected insensitivity of WEHI-164 cells to all three SRV’s tested.
The Galectin Toxin Hypothesis
- 1.)
- Galectin-like proteins expressed in venom gland cells of marine stingrays are released locally into target tissues during barb-mediated envenomation.
- 2.)
- SRV galectin-like proteins, via their carbohydrate-binding domain (CBD’s), recognize and bind to β-galactoside glycan structures present on multiple N-linked and O-linked glycosylated membrane protein targets that are present on resident cells within the affected tissue (e.g., skin fibroblasts, neurons, and blood cells).
- 3.)
-
Binding of dimerized SRV galectin-like proteins to glycosylated target cell proteins promote aberrant signaling by:
- 4.)
- Collectively these SRV galectin-mediated events either cause or contribute to the hallmark features of localized stingray injury, i.e. severe pain and dermal tissue necrosis.
- 5.)
- We further hypothesize that the SRV-resistant characteristics of WEHI-164 cells are due to a distinct glycocalyx where SRV galectin-like proteins either do not recognize glycan targets on WEHI-164 cells, and/or the recognized glycan targets on HDFa, SH-SY5Y, and Jurkat-E6-1 cells are not expressed and presented by WEHI-164 cells. Abnormal glycosylation patterns are a common feature of malignant cells.
The galectin-like transcript in N. kuhlii venom tissue (i.e., Neo galectin-1) and its encoded protein (second most abundant venom protein characterized), was originally identified by high sequence alignment with a galectin-like protein previously identified in the Japanese puffer fish (Takifugu rubripes) genome (UniprotKB ID: H2UTD9) [16,33]. A current BLAST search using the Neo galectin-1 nucleotide sequence identified a predicted transcript (XM_059969249.1) in the H. sabinus genome that encodes a 150 amino acid “galectin-12-like” protein homologue (XP_059825232.1) (Figure 8). The Neo galectin-1 protein and H. sabinus galectin-12-like protein are both protomer-type galectins (i.e., contain a single CBD) and share 90% amino acid sequence identity (Figure 8B). Given the N. kuhlii proteomic results confirmed the presence of the Neo galectin-1 protein in N. kuhlii venom, we posit that the H. sabinus galectin-12-like protein homologue is similarly present in the venom of the Atlantic stingray.
The H. sabinus galectin-12-like protein is encoded by 5 exons located on chromosome 5 (LOC132393850) based on the sHypSap1.hap1 genome assembly (GCF_030144855.1). The assembled Atlantic stingray genome revealed a total of at least 8 distinct galectin-like genes encoding 12 putative galectin-like protein isoforms (Figure 8C). Several of the H. sabinus galectin-like proteins encode protomer-type galectins (single CBD) with dimerization sequences at both N- and C-termini for homo-dimer formation. Both the H. sabinus galectin-8-like and galectin-9-like proteins are tandem repeat-type galectins, with each encoding two distinct CBD’s separated by an amino acid linker sequence [29,30]. The venom-associated galectins represent a distinct subfamily of galectin-like proteins.
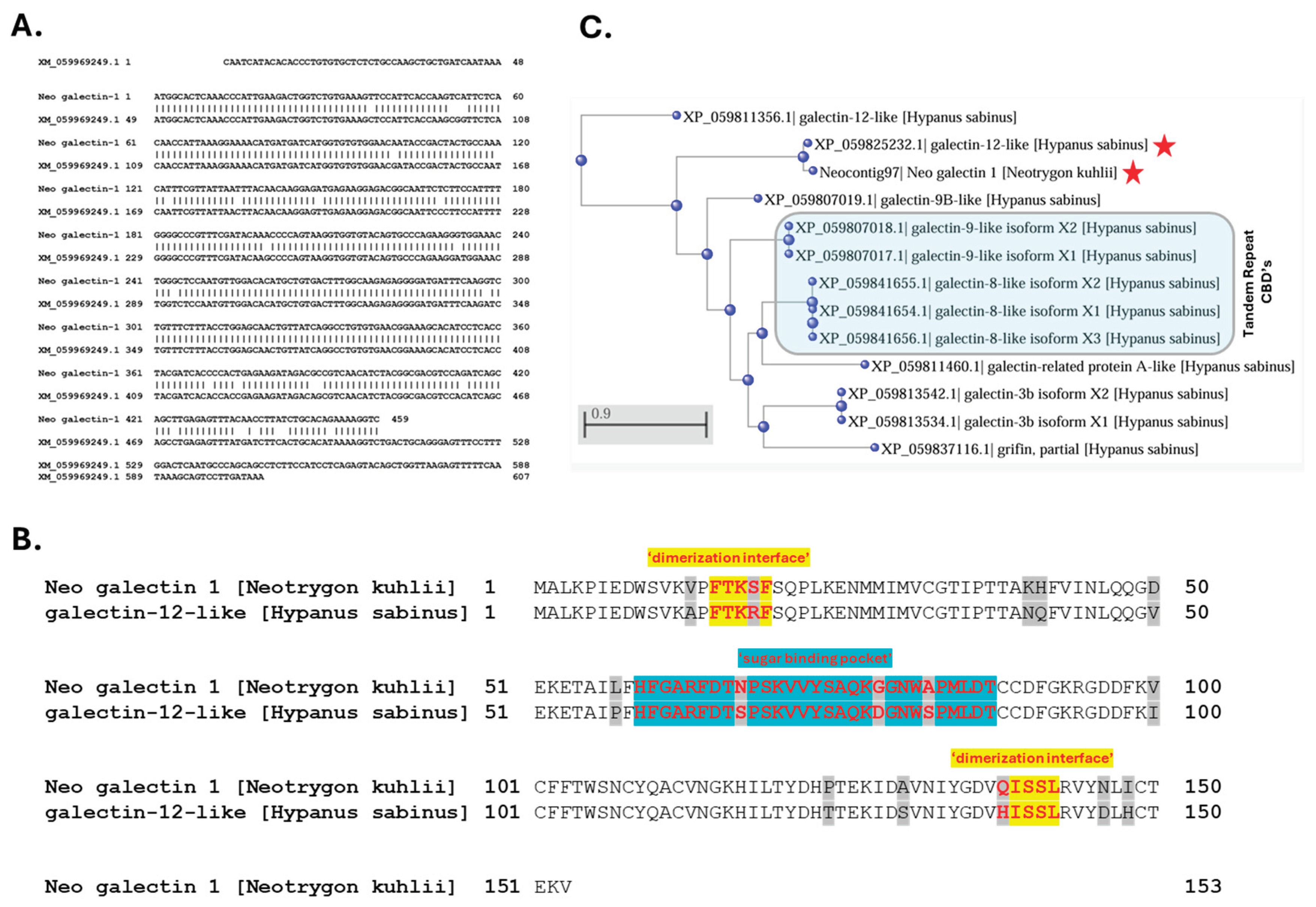
Figure 8.
A venom-associated galectin-like protein homologue in the Atlantic stingray. a. Nucleotide sequence alignment of Neocontig97 from Neotrygon kuhlii and the Hypanus sabinus mRNA RefSeq transcript (XM_059969249.1) identified by a BLAST database search. b. Amino acid sequence alignment of the Neotrygon kuhlii galectin 1 protein and predicted Hypanus sabinus galectin-12-like protein. The conserved domains representing the dimerization interface sequences and ‘sugar binding pocket’ (i.e., CBD) are highlighted in red font, non-identical amino acids are highlighted in grey. Both proteins were classified as GLECT domain-containing proteins using the CD-Search interface of the NCBI conserved domain database https://www.ncbi.nlm.nih.gov/cdd. The two proteins share 90% sequence identity. c. Phylogenetic representation of predicted galectin proteins encoded by galectin-like genes annotated in the sHypSap1.hap1 genome assembly. The phylogenetic tree was constructed from a multiple sequence alignment of the proteins shown using COBALT and Phylogenetic Tree View with the following parameters: Fast Minimum Evolution, Maximum Sequence Distance = 0.90, and the Grishin protein algorithm. The results were then sorted by distance in ascending order. The venom-associated galectins are indicated by the two red stars.
Figure 8.
A venom-associated galectin-like protein homologue in the Atlantic stingray. a. Nucleotide sequence alignment of Neocontig97 from Neotrygon kuhlii and the Hypanus sabinus mRNA RefSeq transcript (XM_059969249.1) identified by a BLAST database search. b. Amino acid sequence alignment of the Neotrygon kuhlii galectin 1 protein and predicted Hypanus sabinus galectin-12-like protein. The conserved domains representing the dimerization interface sequences and ‘sugar binding pocket’ (i.e., CBD) are highlighted in red font, non-identical amino acids are highlighted in grey. Both proteins were classified as GLECT domain-containing proteins using the CD-Search interface of the NCBI conserved domain database https://www.ncbi.nlm.nih.gov/cdd. The two proteins share 90% sequence identity. c. Phylogenetic representation of predicted galectin proteins encoded by galectin-like genes annotated in the sHypSap1.hap1 genome assembly. The phylogenetic tree was constructed from a multiple sequence alignment of the proteins shown using COBALT and Phylogenetic Tree View with the following parameters: Fast Minimum Evolution, Maximum Sequence Distance = 0.90, and the Grishin protein algorithm. The results were then sorted by distance in ascending order. The venom-associated galectins are indicated by the two red stars.
3.5. Galectin Inhibitors in Cancer Therapeutics
Galectins have received considerable attention in cancer research given known differences in glycosylation patterns between normal and malignant tumor cells [34,35]. The cell surface glycan differences have been attributed to changes in cell adhesion and growth that represent hallmark features in cancer biology. Fourteen known galectin genes in humans participate in a broad range of galectin-specific regulatory processes that are altered in several diseases [30]. Their utility in cancer diagnosis and prognosis has also gained considerable interest [34]. Moreover, galectin selective inhibitors have recently received FDA approval for the treatment of conditions where elevated galectin levels are linked to clinical disease, and targeting them with inhibitors has shown therapeutic potential [36].
3.6. Venoms as a Source of Potential Anti-Cancer Agents
The anti-proliferative properties of venoms extracted from vertebrates and invertebrate species have been widely reported, where the potential application of venoms as anti-cancer agents has received considerable attention during the past decade, especially with snake venoms (for reviews, see [37,38,39,40]). Anticancer activity in various snake venoms has been reported for pancreatic tumor cells [41], human breast cancer (MCF-7), human hepatocellular carcinoma (HepG2) and human prostate carcinoma (DU145) cell lines [42], human leukemic cell lines (U937 and K562) and sarcoma in a Balb C mice model [43], colorectal and breast cancer cell lines [44], human prostate adenocarcinoma (LNCaP), human breast cancer (MCF-7), human colon adenocarcinoma (HT-29) and human osteoblastic osteosarcoma (Saos-2) cells [45], and human breast carcinoma cells (MDA-MB-231 and MCF-7) [46,47].
Literature reports of anti-tumor effects from venoms derived from invertebrates are less common yet include scorpion venom effects on U251-MG glioma cells and xenograft tumors [48], as well as human brain, breast, colorectal, lungs, cervix, and larynx cancer cell lines [49]. Bee venom also has reported anti-cancer effects against MDA-MB-231 and MCF-7 human breast cancer cells [50,51] and ovarian cancers [52,53,54].
Our findings clearly revealed growth inhibitory activities that were similar among all three SRV extracts examined. The concentration dependent SRV effects were not markedly different among the three different human cell lines treated (HDFa, SH-SY5Y, and Jurkat E6 cells). Interestingly, however, the mouse fibrosarcoma WEHI 164 cell line was remarkably resistant to all three SRV’s at the relatively high exposure concentrations and durations tested.
Our live-cell time course analysis further revealed Atlantic stingray venom evoked both rapid and sustained effects, occurring within minutes after exposure (cell detachment) and persisting for days (growth inhibition and apoptosis). These findings are consistent with the high-content screening results performed with five different SRV treatments of human cervical cancer HeLa cells [19]. They also align with the antiproliferative effects on HeLa cells previously reported for the crude venom extract from the cowtail stingray (Pastinachus sephen, Dasyatidae family), that enhanced levels of reactive oxygen species, lipid peroxidation, and apoptosis [55]. Both cowtail stingray and spotted eagle ray venoms additionally exhibit anticoagulant activity [56].
4. Materials and Methods
4.1. Capture and Housing of Local Stingrays
Atlantic stingrays (Hypanus sabinus, formerly Dasyatis sabinus), spotted eagle rays (Aetobatus narinari), and cownose rays (Rhinoptera bonasus) were captured using small watercraft vessels in near shore waters off Sarasota, Florida, in Sarasota Bay, or in Terra Ceia Bay, following guidelines specified in the Special Activities Licenses issued to the Mote Marine Laboratory by the Florida Fish and Wildlife Conservation Commission (FFWCC). After visual sighting of stingrays in shallow water, capture was non-invasive, using cast nets for Atlantic stingrays, seine nets for eagle rays, and multi-panel gill nets for cownose rays. Atlantic stingrays and spotted eagle rays were placed in a live well for transport to the laboratory vivarium for later barb collection. Cownose rays were released unharmed after their barbs were collected at time of capture.
Captive Atlantic stingrays were individually housed on a 12h on/12h off photoperiod, in 340 L holding tanks containing natural seawater maintained at 23-25 ˚C. Seawater salinity was maintained between 30% and 35% by addition of water that had been de-ionized by reverse osmosis. Periodic exchanges with ozone-treated seawater were performed to supply trace elements and to keep nitrate accumulation below 20 mg/L. Stingrays were fed three times per week with a varied diet of thread herring, shrimp, and squid, supplemented with Elasmo-tabs Vitamins, (Mazuri Exotic Animal Diets). Captive eagle rays were housed in a 150,000 L tank containing natural seawater maintained at 25-28 °C and on a 12 h on/12h off photoperiod. Eagle rays were fed a variety of molluscs daily and were released within 30 days of their capture in compliance with FFWCC regulations.
4.2. Spine/Barb Removal Procedures
Following transport and arrival to the laboratory after capture, spotted eagle ray spines were immediately removed using ethanol-sterilized long nose pliers (Figure 8). Once removed, spines were taken to the laboratory where they were snap frozen and stored at -80 ˚C. The spotted eagle rays with spine removed were then maintained as described above for approved feeding behavioral studies not associated with this project.

Figure 8.
(a) Spotted eagle ray spines were removed from the dorsal surface of the tail by first grasping the spine with long nose pliers; (b) Twisting the spine dorsal and rostral away from the tail resulted in spine separation from the ray.
Figure 8.
(a) Spotted eagle ray spines were removed from the dorsal surface of the tail by first grasping the spine with long nose pliers; (b) Twisting the spine dorsal and rostral away from the tail resulted in spine separation from the ray.
Atlantic stingray spines are more firmly attached to their tail compared to eagle ray spines, and therefore were removed using ethanol-sterilized wire cutting pliers (Figure 9). The Atlantic stingrays were anesthetized prior to spine removal to eliminate undue stress on the ray and as a safety precaution for the handler(s). Rays were anesthetized by immersion in seawater containing MS-222 at a concentration of 100-200 mg/L (buffered at 1 part MS-222:2 parts sodium bicarbonate). For some rays, the removed spine was snap frozen and stored at -80 ˚C for later processing. For others, the freshly removed spine was used immediately for venom tissue collection (Figure 9).
Cownose stingray spines, which also are firmly attached to their tail, were similarly clipped using ethanol-sterilized wire cutting pliers immediately after their capture. The removed cownose spines were immediately placed in sterile 15 mL tubes and kept on ice during transit to the laboratory where they were then snap frozen and stored at -80 ˚C for later processing.
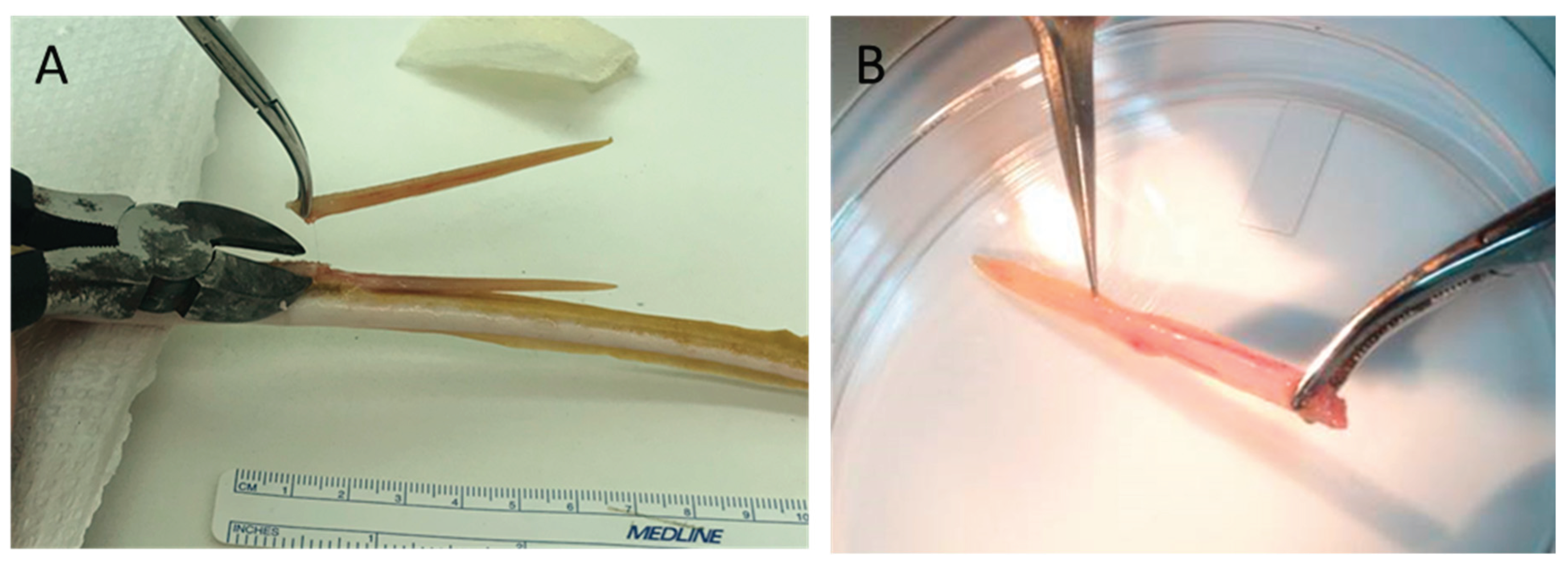
Figure 9.
Collection of venom-containing spine tissue: (a) Atlantic stingray spines were clipped near their site of attachment on the dorsal surface of the tail using ethanol-sterilized wire cutting pliers; (b) Crude venom gland tissue was then scraped from the ventral surface of the spine using sterile fine tip dissecting forceps and into a petri dish after rinsing with sterile E-PBS solution.
Figure 9.
Collection of venom-containing spine tissue: (a) Atlantic stingray spines were clipped near their site of attachment on the dorsal surface of the tail using ethanol-sterilized wire cutting pliers; (b) Crude venom gland tissue was then scraped from the ventral surface of the spine using sterile fine tip dissecting forceps and into a petri dish after rinsing with sterile E-PBS solution.
4.3. Collection and Preparation of Crude SRV
The removed spines were initially rinsed with elasmobranch-modified PBS (E-PBS; phosphate-buffered saline modified to an osmolarity of ~970 mOsm by adding NaCl). The venom-containing tissue was then collected by mechanically scraping the ventral side of the barb using sterile forceps (Figure 9A). The collected tissue was placed in sterile Eppendorf tubes and suspended in 50 mM NH4HCO3 solution adjusted to pH 7.4. Using a hand-held tissue homogenizer, the tissue was gently homogenized and centrifuged at 10,000 x g for 10 minutes. The clear supernatant containing crude SRV was then aliquoted into small volumes (100- 250 µL). Some aliquots were used for protein quantification and biochemical analysis. Other aliquots were frozen at -80 °C and lyophilized. Lyophilized SRV was stored at -80 °C until assessed for bioactivity.
4.4. SDS Polyacrylamide Gel Electrophoresis (PAGE)
Comparative protein profiles of the different SRV samples were obtained using SDS-PAGE with 4-20% Mini-PROTEIN TGX Precast gels (BioRad). Prior to electrophoresis, protein concentrations of the SRV samples were determined using the Bradford assay (BioRad). SRV samples were loaded at 25 µg protein per lane after denaturation for 5 min at 95 °C. Samples were run at 200 volts and the gels subsequently stained using Coomassie blue for visual analysis.
4.5. Cell Culture
To test for SRV bioactivity, four different cells lines of mammalian origin were used as listed in Table 1. All cell lines were obtained from American Type Culture Collection (ATCC). Cells were maintained and sub-cultured according to instructions provided by ATCC. All cell culture media contained antibiotics (penicillin, 100U/mL/streptomycin, 10 µg/mL). Cell culture reagents were either obtained from Sigma or ATCC.
4.6. Cell Growth Assays
The comparative effects of SRV’s on cellular growth was determined against all cell types listed in Table 1. Growth inhibitory activity of cultured cells by the crude venoms was quantified using the MTT assay [15], an assay that measures the ability of live cells to convert the tetrazolium salt MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide) to a formazan product via mitochondrial enzymes.
Cells were cultured according to protocols provided by ATCC and adjusted to a concentration of 5 x 104 cells/mL in media with 20% FBS. Cells (100 µL) were added to wells of 96-well microtiter plates in triplicate for control and venom treatments. Venom solutions of varying concentrations in cell culture media were added in 100 µL volumes. For HDFa cells, final well concentrations were 2.5x104 cells/mL, 10% FBS, 0 – 400 µg/mL venom protein. SH-SY5Y cells were used at a concentration of 5x105 cells/mL, with final well concentrations of 2.5 x 105 cells/mL, 10% FBS, and 0-200 µg/mL eagle ray venom protein. Cultures were incubated at 37 °C, 5% CO2 for 72 hours. After the SRV treatment period, a 100 µL supernatant was carefully removed from each well, and 25 µL of MTT solution (5 mg/mL in PBS) added to each well. For cells in suspension (Jurkat cells only), microtiter plates were centrifuged at 300 x g for 7 minutes prior to removing supernatant. Cells were then incubated with MTT for 4 hours at 37 °C. After the 4-hour incubation period, a solubilizing solution (0.01 N HCl, 10% SDS) was added to each well and incubated overnight at 37 °C.
Using a BioTek Synergy H1 microplate reader, absorbance was read at 570 nm for each sample with background subtracted at 630 nm. The absorbance values for triplicate samples were then averaged. The percent growth inhibition (%GI) was calculated for each treatment as follows: %GI = 1 – (TreatmentAbs570-630/ControlAbs570-630) x 100.
To assess SRV effects on cell morphology, cells from different treatment groups were photographed using an Olympus CK2 inverted microscope and Infinity 2.0 digital camera.
4.7. Biosensor-Based Real-Time Cell Impedance Recordings
The time course for effects of Atlantic stingray venom were measured on HDFa cells maintained in culture for 5 days using an impedance-based biosensor system (xCELLigence Real-Time Cell Analyzer, ACEA Biosciences) [14]. Time-dependent changes in impedance values reflect changes in cell growth and adherence to the well surface over time [14]. HDFa cells were seeded in 96-well polyethylene terephthalate E-plates (ACEA Biosciences) and maintained in culture for up to 5 days at 37 °C, 5% CO2. After 3-days in culture, cells were treated with SRV at varying concentrations (0, 1, 3, 10, 30, 100 μg/mL), in triplicate wells. TritonX-100 at 0.1% was used as a positive control for detaching cells from the well surface. Impedance values for each individual well were recorded every 15 minutes over the 5-day experimental period. Analysis of the recorded impedance values, quantified as Cell Index (CI) values, was then performed using the system software and data exported to an Excel compatible file format.
4.8. Flow Cytometry Assay
Cell apoptosis and necrosis were determined using flow cytometry with an Annexin V/Dead Cell Apoptosis Kit (Invitrogen) [23]. Annexin V is a phospholipid binding protein with high affinity for phosphatidylserine. In viable healthy cells, Annexin V binding is low since phosphatidylserine is located largely on the cytoplasmic surface of cell membranes. In cells undergoing early cell membrane changes associated with apoptosis, phosphatidylserine is translocated from the inner membrane to the outer part of the plasma membrane where it can bind fluorescently labeled Annexin V conjugates. Cell death was determined using propidium iodide (PI) that binds DNA accessible in dead and dying cells having compromised cell membranes. Healthy live cells with intact cell membranes prevent PI cellular access and DNA binding.
Jurkat E6-1 cells were plated at 1 x 106 cells per well in a 24-well cell culture plate. SRV’s were included with cells in duplicate wells at final protein concentrations of 3.125, 6.25, 12.5, 25, 50, 100, 200, and 400 µg/mL. Staurosporine (2 µM, Sigma) was used as a positive control for apoptosis. Heat-killed cells (95 °C for 30 min) was used as a positive control for necrosis. For each treatment condition, cells were incubated for 24 hours at 37 °C, 5% CO2.
Following the 24-hour treatment period, Jurkat cells were transferred to Eppendorf tubes, centrifuged, and washed with ice-cold PBS. Cells were incubated for 15 minutes with 1 µg/mL PI solution and 5% FITC-labeled Annexin V in 0.1 ml annexin binding buffer. Samples were then diluted with 0.4 ml annexin binding buffer and placed on ice. Fluorescence was measured using a flow cytometer (BD Accuri C6 system) at 35 µl/min, where 10,000 events were read with 530 nm excitation and 575 nm emission wavelengths. Unstained controls and cells stained with only PI or FITC were used as instrument controls. Gating criteria for healthy, apoptotic, and necrotic cell populations were set based on positive control values. The percentage of cells counted within each category were then calculated.
4.9. Statistical Analyses
Statistical analyses on MTT assay Optical Density data were conducted using SigmaPlot v14.0. Statistical analyses conducted included one-way ANOVA and Student’s t-test. P values less than 0.05 were considered statistically significant. EC50 values were calculated using OD values and online software (AAT Bioquest EC50 Calculator).
5. Conclusions
Our comparative analysis of SRV isolated from three local species reveals species-conserved bioactive properties that could have practical biomedical or pharmacological applications. The time course analysis and sensitivity/insensitivity of different cell lines to SRV treatments sheds new light on the cellular processes and underlying molecular mechanisms that mediate their biological activities. Future studies testing the SRV galectin toxin hypothesis will fill an important gap in our current knowledge as to whether venom associated galectin-like proteins recapitulate bioactive properties of crude SRV extracts observed in vitro and with envenomation. Exploiting drug development of recent galectin inhibitors may also provide new opportunities for treating severe stingray injuries.
Author Contributions
This collaborative project was conceived and initiated by C.D. (University of South Florida) in coordination with C.L. and C.J.W. (Mote Marine Laboratory). All three authors were involved in methodology, validation, formal analysis, investigation, resources, data curation, writing—original draft preparation, writing—review and editing, and data visualization. J.R. and J.B. were involved in methodology, investigation, and data curation. Project administration was coordinated by C.D. and C.L. All authors have read and agreed to the final version of the manuscript.
Funding
This research project received no direct external funding. Partial funding was provided by NSF Award OCE-2050892 (C.J.W.) that supported undergraduate student (J.B.) research training.
Institutional Review Board Statement
The protocols for studies involving all stingrays were reviewed and approved by the Mote Marine Laboratory’s Institutional Animal Care and Use Committee (21-03-CL2, approval date 3/30/2021; 22-03-CL2, approval date 3/28/2022; 23-03-CL2, approval date 3/27/2023).
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding authors.
Acknowledgments
The authors gratefully acknowledge the use of facilities at Mote Marine Laboratory (Sarasota, FL, United States), and thank Tonya Wiley, Krystan Wilkinson, and Kim Hull for help collecting stingray spines. The authors also acknowledge Udoka Okaro for training on the ACEA xCELLigence core facility system, and the contributions of Karlie Tischendorf who conducted preliminary studies related to this project. The authors acknowledge the support of NSF Award OCE-2050892 for both J. Brick. and K. Tischendorf.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Fry, B.G.; Roelants, K.; Champagne, D.E.; Scheib, H.; Tyndall, J.D.; King, G.F.; Nevalainen, T.J.; Norman, J.A.; Lewis, R.J.; Norton, R.S.; et al. The toxicogenomic multiverse: convergent recruitment of proteins into animal venoms. Annu Rev Genomics Hum Genet 2009, 10, 483–511. [Google Scholar] [CrossRef]
- Casewell, N.R.; Wuster, W.; Vonk, F.J.; Harrison, R.A.; Fry, B.G. Complex cocktails: the evolutionary novelty of venoms. Trends Ecol Evol 2013, 28, 219–229. [Google Scholar] [CrossRef]
- Lewis, R.J.; Garcia, M.L. Therapeutic potential of venom peptides. Nat Rev Drug Discov 2003, 2, 790–802. [Google Scholar] [CrossRef]
- King, G.F. Venoms as a platform for human drugs: translating toxins into therapeutics. Expert Opin Biol Ther 2011, 11, 1469–1484. [Google Scholar] [CrossRef]
- Zhang, Y. Why do we study animal toxins? Dongwuxue Yanjiu 2015, 36, 183–222. [Google Scholar] [CrossRef]
- Bordon, K.C.F.; Cologna, C.T.; Fornari-Baldo, E.C.; Pinheiro-Junior, E.L.; Cerni, F.A.; Amorim, F.G.; Anjolette, F.A.P.; Cordeiro, F.A.; Wiezel, G.A.; Cardoso, I.A.; et al. From Animal Poisons and Venoms to Medicines: Achievements, Challenges and Perspectives in Drug Discovery. Front Pharmacol 2020, 11, 1132. [Google Scholar] [CrossRef]
- Ocampo, R.R.; Halstead, B.W.; Modglin, F.R. The microscopic anatomy of the caudal appendage of the spotted eagleray, Aetobatus narinari (Euphrasen), with special reference to the venom apparatus. Anat Rec 1953, 115, 87–99. [Google Scholar] [CrossRef]
- Halstead, B.W.; Ocampo, R.R.; Modglin, F.R. A study on the comparative anatomy of the venom apparatus of certain north American stingrays. Journal of Morphology 1955, 97, 1–21. [Google Scholar] [CrossRef]
- Smith, D.S.; Cayer, M.L.; Russell, F.E. Membrane-limited microtubular aggregates in the venom secreting cells of a stingray. Toxicon 1974, 12, 331–335. [Google Scholar] [CrossRef] [PubMed]
- Brighton Ndandala, C.; Mustapha, U.F.; Wang, Y.; Assan, D.; Zhao, G.; Huang, C.; Mkuye, R.; Huang, H.; Li, G.; Chen, H. The perspective of fish venom: An overview of the physiology, evolution, molecular and genetics. Frontiers in Marine Science 2023, 10. [Google Scholar] [CrossRef]
- Harris, R.J. The piscine arsenal: an updated review of venomous fishes. Reviews in Fish Biology and Fisheries 2023. [Google Scholar] [CrossRef]
- Pedroso, C.M.; Jared, C.; Charvet-Almeida, P.; Almeida, M.P.; Garrone Neto, D.; Lira, M.S.; Haddad, V., Jr.; Barbaro, K.C.; Antoniazzi, M.M. Morphological characterization of the venom secretory epidermal cells in the stinger of marine and freshwater stingrays. Toxicon 2007, 50, 688–697. [Google Scholar] [CrossRef] [PubMed]
- Dehghani, H.; Sajjadi, M.M.; Parto, P.; Rajaian, H.; Mokhlesi, A. Histological characterization of the special venom secretory cells in the stinger of rays in the northern waters of Persian Gulf and Oman Sea. Toxicon 2010, 55, 1188–1194. [Google Scholar] [CrossRef] [PubMed]
- Ke, N.; Wang, X.; Xu, X.; Abassi, Y.A. The xCELLigence system for real-time and label-free monitoring of cell viability. Methods Mol Biol 2011, 740, 33–43. [Google Scholar] [CrossRef] [PubMed]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: application to proliferation and cytotoxicity assays. J Immunol Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Baumann, K.; Casewell, N.R.; Ali, S.A.; Jackson, T.N.; Vetter, I.; Dobson, J.S.; Cutmore, S.C.; Nouwens, A.; Lavergne, V.; Fry, B.G. A ray of venom: Combined proteomic and transcriptomic investigation of fish venom composition using barb tissue from the blue-spotted stingray (Neotrygon kuhlii). J Proteomics 2014, 109, 188–198. [Google Scholar] [CrossRef]
- de Oliveira Junior, N.G.; Fernandes Gda, R.; Cardoso, M.H.; Costa, F.F.; Candido Ede, S.; Garrone Neto, D.; Mortari, M.R.; Schwartz, E.F.; Franco, O.L.; de Alencar, S.A. Venom gland transcriptome analyses of two freshwater stingrays (Myliobatiformes: Potamotrygonidae) from Brazil. Sci Rep 2016, 6, 21935. [Google Scholar] [CrossRef]
- Silva, F.; Huang, Y.; Yang, V.; Mu, X.; Shi, Q.; Antunes, A. Transcriptomic Characterization of the South American Freshwater Stingray Potamotrygon motoro Venom Apparatus. Toxins (Basel) 2018, 10. [Google Scholar] [CrossRef]
- Kirchhoff, K.N.; Billion, A.; Voolstra, C.R.; Kremb, S.; Wilke, T.; Vilcinskas, A. Stingray Venom Proteins: Mechanisms of Action Revealed Using a Novel Network Pharmacology Approach. Marine Drugs 2022, 20. [Google Scholar] [CrossRef]
- Russell, F.E. Stingray injuries: a review and discussion of their treatment. Am J Med Sci 1953, 226, 611–622. [Google Scholar] [CrossRef]
- Diaz, J.H. The epidemiology, evaluation, and management of stingray injuries. J La State Med Soc 2007, 159, 198–204, quiz 204, 230. [Google Scholar] [PubMed]
- White, J. Venomous animals: clinical toxinology. In Molecular, Clinical and Environmental Toxicology: Volume 2: Clinical Toxicology, Luch, A., Ed.; Birkhäuser Basel: Basel, 2010; pp. 233–291. [Google Scholar]
- Koopman, G.; Reutelingsperger, C.P.; Kuijten, G.A.; Keehnen, R.M.; Pals, S.T.; van Oers, M.H. Annexin V for flow cytometric detection of phosphatidylserine expression on B cells undergoing apoptosis. Blood 1994, 84, 1415–1420. [Google Scholar] [CrossRef] [PubMed]
- Marmi, J.; Vila, B.; Oms, O.; Galobart, À.; Cappetta, H. Oldest records of stingray spines (Chondrichthyes, Myliobatiformes). Journal of Vertebrate Paleontology 2010, 30, 970–974. [Google Scholar] [CrossRef]
- Shea-Vantine, C.S.; Galloway, K.A.; Ingle, D.N.; Porter, M.E.; Kajiura, S.M. Caudal Spine Morphology and Puncture Performance of Two Coastal Stingrays. Integr Comp Biol 2021, 61, 749–758. [Google Scholar] [CrossRef] [PubMed]
- Russell, F.E.; Panos, T.C.; Kang, L.W.; Warner, A.M.; Colket, T.C. , 3rd. Studies on the mechanism of death from stingray venom; a report of two fatal cases. Am J Med Sci 1958, 235, 566–584 passim. [Google Scholar] [CrossRef]
- Last, P.R.; Naylor, G.J.; Manjaji-Matsumoto, B.M. A revised classification of the family Dasyatidae (Chondrichthyes: Myliobatiformes) based on new morphological and molecular insights. Zootaxa 2016, 4139, 345–368. [Google Scholar] [CrossRef]
- Xie, Q.; Tang, N.; Wan, R.; Qi, Y.; Lin, X.; Lin, J. Recombinant snake venom cystatin inhibits the growth, invasion and metastasis of B16F10 cells and MHCC97H cells in vitro and in vivo. Toxicon 2011, 57, 704–711. [Google Scholar] [CrossRef]
- Verkerke, H.; Dias-Baruffi, M.; Cummings, R.D.; Arthur, C.M.; Stowell, S.R. Galectins: An Ancient Family of Carbohydrate Binding Proteins with Modern Functions. Methods Mol Biol 2022, 2442, 1–40. [Google Scholar] [CrossRef]
- Troncoso, M.F.; Elola, M.T.; Blidner, A.G.; Sarrias, L.; Espelt, M.V.; Rabinovich, G.A. The universe of galectin-binding partners and their functions in health and disease. J Biol Chem 2023, 299, 105400. [Google Scholar] [CrossRef]
- Rabinovich, G.A.; Toscano, M.A.; Jackson, S.S.; Vasta, G.R. Functions of cell surface galectin-glycoprotein lattices. Curr Opin Struct Biol 2007, 17, 513–520. [Google Scholar] [CrossRef]
- Cha, S.K.; Ortega, B.; Kurosu, H.; Rosenblatt, K.P.; Kuro, O.M.; Huang, C.L. Removal of sialic acid involving Klotho causes cell-surface retention of TRPV5 channel via binding to galectin-1. Proc Natl Acad Sci U S A 2008, 105, 9805–9810. [Google Scholar] [CrossRef] [PubMed]
- Kai, W.; Kikuchi, K.; Tohari, S.; Chew, A.K.; Tay, A.; Fujiwara, A.; Hosoya, S.; Suetake, H.; Naruse, K.; Brenner, S.; et al. Integration of the genetic map and genome assembly of fugu facilitates insights into distinct features of genome evolution in teleosts and mammals. Genome Biol Evol 2011, 3, 424–442. [Google Scholar] [CrossRef] [PubMed]
- Thijssen, V.L.; Heusschen, R.; Caers, J.; Griffioen, A.W. Galectin expression in cancer diagnosis and prognosis: A systematic review. Biochim Biophys Acta 2015, 1855, 235–247. [Google Scholar] [CrossRef]
- Marino, K.V.; Cagnoni, A.J.; Croci, D.O.; Rabinovich, G.A. Targeting galectin-driven regulatory circuits in cancer and fibrosis. Nat Rev Drug Discov 2023, 22, 295–316. [Google Scholar] [CrossRef]
- Bertuzzi, S.; Quintana, J.I.; Arda, A.; Gimeno, A.; Jimenez-Barbero, J. Targeting Galectins With Glycomimetics. Front Chem 2020, 8, 593. [Google Scholar] [CrossRef] [PubMed]
- Vyas, V.K.; Brahmbhatt, K.; Bhatt, H.; Parmar, U. Therapeutic potential of snake venom in cancer therapy: current perspectives. Asian Pac J Trop Biomed 2013, 3, 156–162. [Google Scholar] [CrossRef]
- Calderon, L.A.; Sobrinho, J.C.; Zaqueo, K.D.; de Moura, A.A.; Grabner, A.N.; Mazzi, M.V.; Marcussi, S.; Nomizo, A.; Fernandes, C.F.; Zuliani, J.P.; et al. Antitumoral activity of snake venom proteins: new trends in cancer therapy. Biomed Res Int 2014, 2014, 203639. [Google Scholar] [CrossRef]
- Shanbhag, V.K.L. Applications of snake venoms in treatment of cancer. Asian Pacific Journal of Tropical Biomedicine 2015, 5, 275–276. [Google Scholar] [CrossRef]
- Offor, B.C.; Piater, L.A. Snake venom toxins: Potential anticancer therapeutics. J Appl Toxicol 2023. [Google Scholar] [CrossRef]
- Kerkkamp, H.; Bagowski, C.; Kool, J.; van Soolingen, B.; Vonk, F.J.; Vlecken, D. Whole snake venoms: Cytotoxic, anti-metastatic and antiangiogenic properties. Toxicon 2018, 150, 39–49. [Google Scholar] [CrossRef]
- Ebrahim, K.; Vatanpour, H.; Zare, A.; Shirazi, F.H.; Nakhjavani, M. Anticancer Activity a of Caspian Cobra (Naja naja oxiana) snake Venom in Human Cancer Cell Lines Via Induction of Apoptosis. Iran J Pharm Res 2016, 15, 101–112. [Google Scholar]
- Debnath, A.; Chatterjee, U.; Das, M.; Vedasiromoni, J.R.; Gomes, A. Venom of Indian monocellate cobra and Russell's viper show anticancer activity in experimental models. J Ethnopharmacol 2007, 111, 681–684. [Google Scholar] [CrossRef]
- Al-Asmari, A.K.; Riyasdeen, A.; Al-Shahrani, M.H.; Islam, M. Snake venom causes apoptosis by increasing the reactive oxygen species in colorectal and breast cancer cell lines. Onco Targets Ther 2016, 9, 6485–6498. [Google Scholar] [CrossRef]
- Yalcin, H.T.; Ozen, M.O.; Gocmen, B.; Nalbantsoy, A. Effect of Ottoman Viper (Montivipera xanthina (Gray, 1849)) Venom on Various Cancer Cells and on Microorganisms. Cytotechnology 2014, 66, 87–94. [Google Scholar] [CrossRef]
- Al-Sadoon, M.K.; Abdel-Maksoud, M.A.; Rabah, D.M.; Badr, G. Induction of apoptosis and growth arrest in human breast carcinoma cells by a snake (Walterinnesia aegyptia) venom combined with silica nanoparticles: crosstalk between Bcl2 and caspase 3. Cell Physiol Biochem 2012, 30, 653–665. [Google Scholar] [CrossRef] [PubMed]
- Malekara, E.; Pazhouhi, M.; Rashidi, I.; Jalili, C. Anti-proliferative and cytotoxic effect of Iranian snake (Vipera raddei kurdistanica) venom on human breast cancer cells via reactive oxygen species-mediated apoptosis. Res Pharm Sci 2020, 15, 76–86. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.X.; Ji, Y.H. Scorpion venom induces glioma cell apoptosis in vivo and inhibits glioma tumor growth in vitro. J Neurooncol 2005, 73, 1–7. [Google Scholar] [CrossRef]
- Mikaelian, A.G.; Traboulay, E.; Zhang, X.M.; Yeritsyan, E.; Pedersen, P.L.; Ko, Y.H.; Matalka, K.Z. Pleiotropic Anticancer Properties of Scorpion Venom Peptides: Rhopalurus princeps Venom as an Anticancer Agent. Drug Des Devel Ther 2020, 14, 881–893. [Google Scholar] [CrossRef]
- Ip, S.W.; Liao, S.S.; Lin, S.Y.; Lin, J.P.; Yang, J.S.; Lin, M.L.; Chen, G.W.; Lu, H.F.; Lin, M.W.; Han, S.M.; et al. The role of mitochondria in bee venom-induced apoptosis in human breast cancer MCF7 cells. In Vivo 2008, 22, 237–245. [Google Scholar]
- Kwon, N.Y.; Sung, S.H.; Sung, H.K.; Park, J.K. Anticancer Activity of Bee Venom Components against Breast Cancer. Toxins (Basel) 2022, 14. [Google Scholar] [CrossRef] [PubMed]
- Jo, M.; Park, M.H.; Kollipara, P.S.; An, B.J.; Song, H.S.; Han, S.B.; Kim, J.H.; Song, M.J.; Hong, J.T. Anti-cancer effect of bee venom toxin and melittin in ovarian cancer cells through induction of death receptors and inhibition of JAK2/STAT3 pathway. Toxicol Appl Pharmacol 2012, 258, 72–81. [Google Scholar] [CrossRef]
- Amini, E.; Baharara, J.; Nikdel, N.; Salek Abdollahi, F. Cytotoxic and Pro-Apoptotic Effects of Honey Bee Venom and Chrysin on Human Ovarian Cancer Cells. Asia Pacific Journal of Medical Toxicology 2015, 4, 68–73. [Google Scholar] [CrossRef]
- Moga, M.A.; Dimienescu, O.G.; Arvatescu, C.A.; Ifteni, P.; Ples, L. Anticancer Activity of Toxins from Bee and Snake Venom-An Overview on Ovarian Cancer. Molecules 2018, 23. [Google Scholar] [CrossRef] [PubMed]
- Rajeshkumar, R.K.; Vennila, R.; Karthikeyan, S.; Prasad, N.R.; Arumugam, M.; Velpandian, T.; Balasubramaniam, T. Antiproliferative activity of marine stingray Dasyatis sephen venom on human cervical carcinoma cell line. J Venom Anim Toxins Incl Trop Dis 2015, 21, 41. [Google Scholar] [CrossRef] [PubMed]
- Kumar, K.R.; Vennila, R.; Kanchana, S.; Arumugam, M.; Balasubramaniam, T. Fibrinogenolytic and anticoagulant activities in the tissue covering the stingers of marine stingrays Dasyatis sephen and Aetobatis narinari. J Thromb Thrombolysis 2011, 31, 464–471. [Google Scholar] [CrossRef]
Figure 1.
SDS-polyacrylamide gel electrophoretic separation of crude SRV proteins obtained from three distinct stingray species. Lane 1: Molecular weight standards. Lane 2: Cownose ray (R. bonasus). Lane 3: Atlantic stingray (H. sabinus). Lane 4: Spotted eagle ray (A. narinari). The amount of protein loaded per lane was 25 μg for each SRV.
Figure 1.
SDS-polyacrylamide gel electrophoretic separation of crude SRV proteins obtained from three distinct stingray species. Lane 1: Molecular weight standards. Lane 2: Cownose ray (R. bonasus). Lane 3: Atlantic stingray (H. sabinus). Lane 4: Spotted eagle ray (A. narinari). The amount of protein loaded per lane was 25 μg for each SRV.
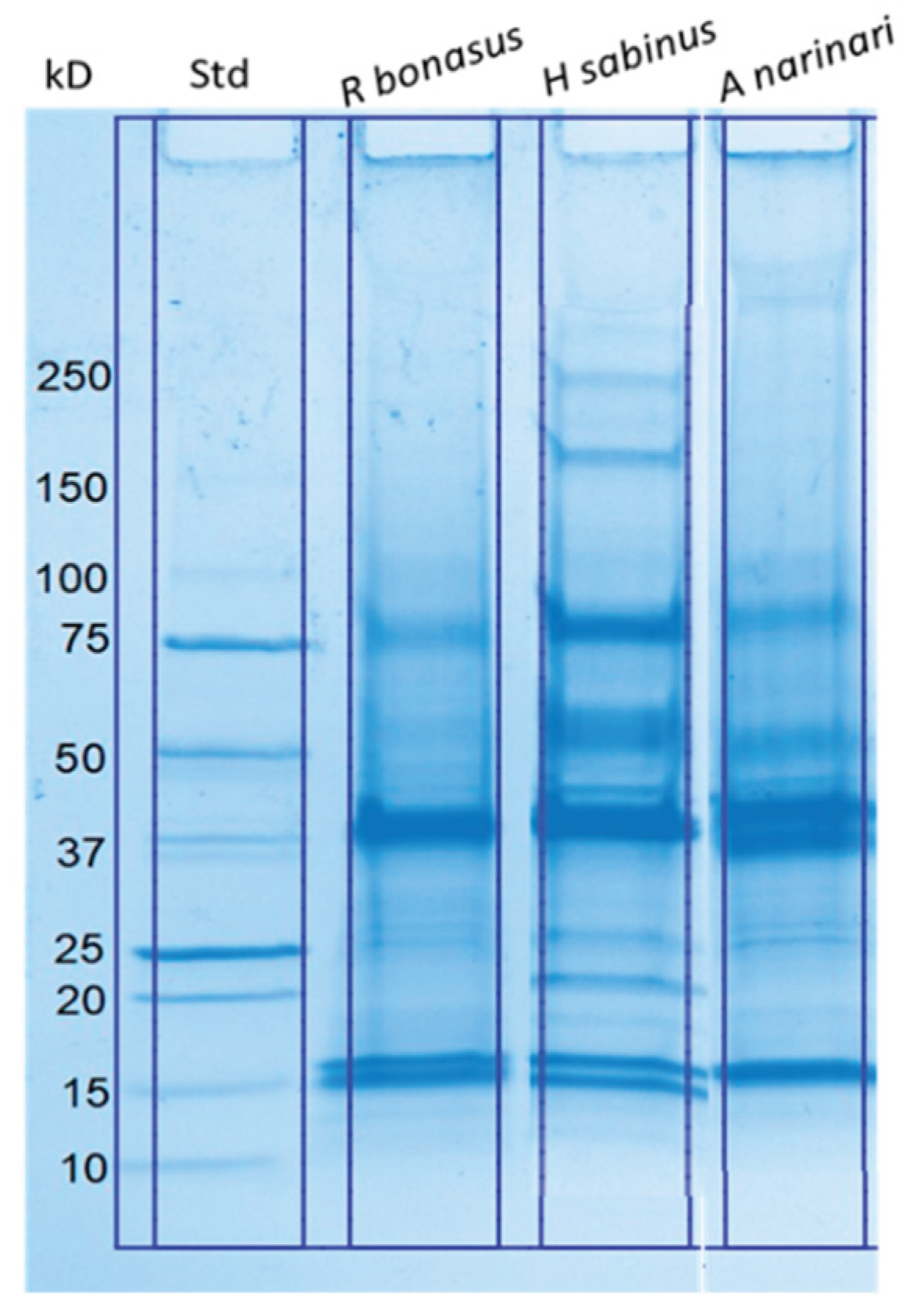
Figure 2.
Inhibition of cell growth caused by exposure to SRV’s. a. HDFa cells treated with Atlantic stingray venom (N=3) at concentrations of 50, 100, 200, and 400 µg/mL. b. HDFa cells treated with spotted eagle ray venom (N=4) at concentrations of 50, 100, 200, and 400 µg/mL. c. SH-SY5Y cells treated spotted eagle ray venom (N=6) at concentrations of 25, 50, 100, and 200 µg/mL. Percent growth inhibition was determined using the MTT assay following a 72-hour treatment period for each condition. *Significantly different (P < 0.05) from control using one-way ANOVA; **Significantly different (P < 0.05) from control using Student’s t-test.
Figure 2.
Inhibition of cell growth caused by exposure to SRV’s. a. HDFa cells treated with Atlantic stingray venom (N=3) at concentrations of 50, 100, 200, and 400 µg/mL. b. HDFa cells treated with spotted eagle ray venom (N=4) at concentrations of 50, 100, 200, and 400 µg/mL. c. SH-SY5Y cells treated spotted eagle ray venom (N=6) at concentrations of 25, 50, 100, and 200 µg/mL. Percent growth inhibition was determined using the MTT assay following a 72-hour treatment period for each condition. *Significantly different (P < 0.05) from control using one-way ANOVA; **Significantly different (P < 0.05) from control using Student’s t-test.
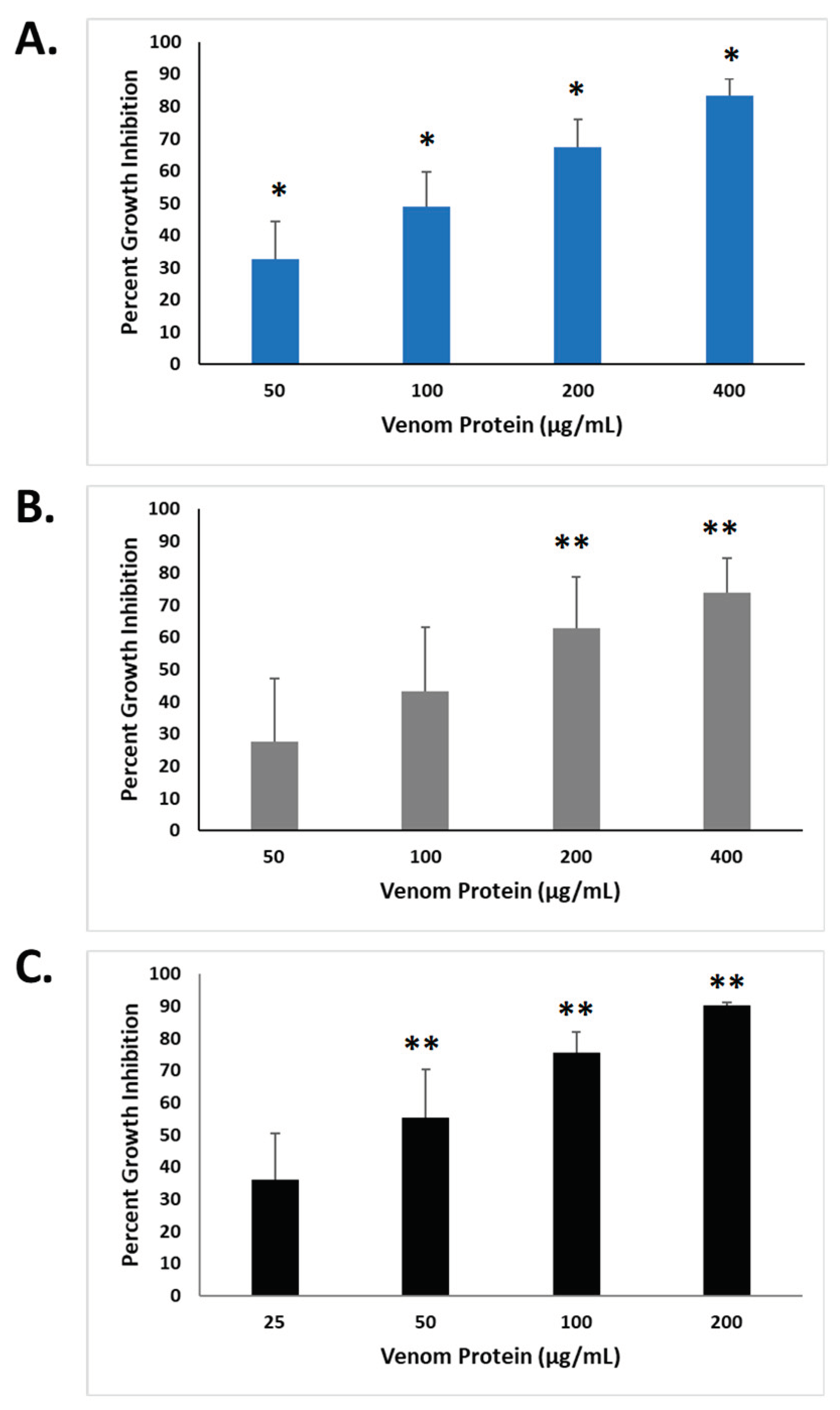
Figure 3.
Morphological effects of eagle ray venom on cultured HDFa cells (top row) compared to cultured SH-SY5Y cells (bottom row). The two cell types were each treated with a range of SRV concentrations for a 72-hour exposure period. Magnification, 10x.
Figure 3.
Morphological effects of eagle ray venom on cultured HDFa cells (top row) compared to cultured SH-SY5Y cells (bottom row). The two cell types were each treated with a range of SRV concentrations for a 72-hour exposure period. Magnification, 10x.
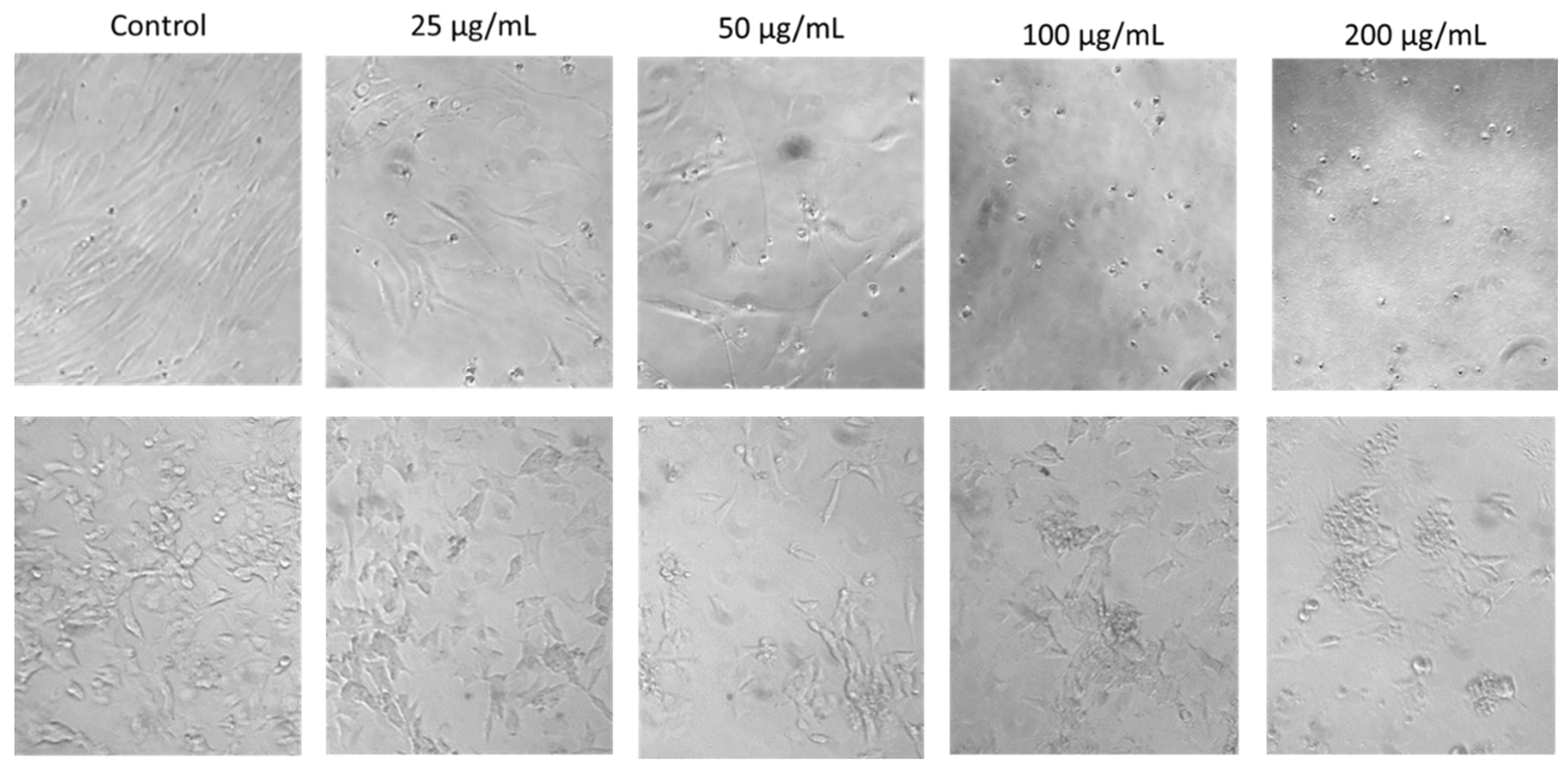
Figure 4.
Time course for SRV-mediated effects on HDFa cells in culture. a. The Cell Index (normalized impedance) was recorded in 96-well E-plates every 15 minutes and plotted over a 5-day time period. The slow near linear increase in Cell Index that occurred during the 3-day pretreatment period reflects the proliferation and adherence of HDFa cells that result in an increase in impedance. b. Time course for changes in Cell Index caused by the different experimental groups over the 2-day treatment period; sham (green trace), SRV treatments (red traces), TritonX-100 (black trace). c. The Cell Index values measured at increasing durations of SRV exposure (1, 12, 24, 36, 48 hours) are plotted for each concentration of SRV tested. d. The derived SRV concentration-response effects are shown for acute SRV exposure (1 hour) and chronic SRV exposure (48 hours). The Cell Index values from SRV treatment groups were normalized to the sham control value.
Figure 4.
Time course for SRV-mediated effects on HDFa cells in culture. a. The Cell Index (normalized impedance) was recorded in 96-well E-plates every 15 minutes and plotted over a 5-day time period. The slow near linear increase in Cell Index that occurred during the 3-day pretreatment period reflects the proliferation and adherence of HDFa cells that result in an increase in impedance. b. Time course for changes in Cell Index caused by the different experimental groups over the 2-day treatment period; sham (green trace), SRV treatments (red traces), TritonX-100 (black trace). c. The Cell Index values measured at increasing durations of SRV exposure (1, 12, 24, 36, 48 hours) are plotted for each concentration of SRV tested. d. The derived SRV concentration-response effects are shown for acute SRV exposure (1 hour) and chronic SRV exposure (48 hours). The Cell Index values from SRV treatment groups were normalized to the sham control value.
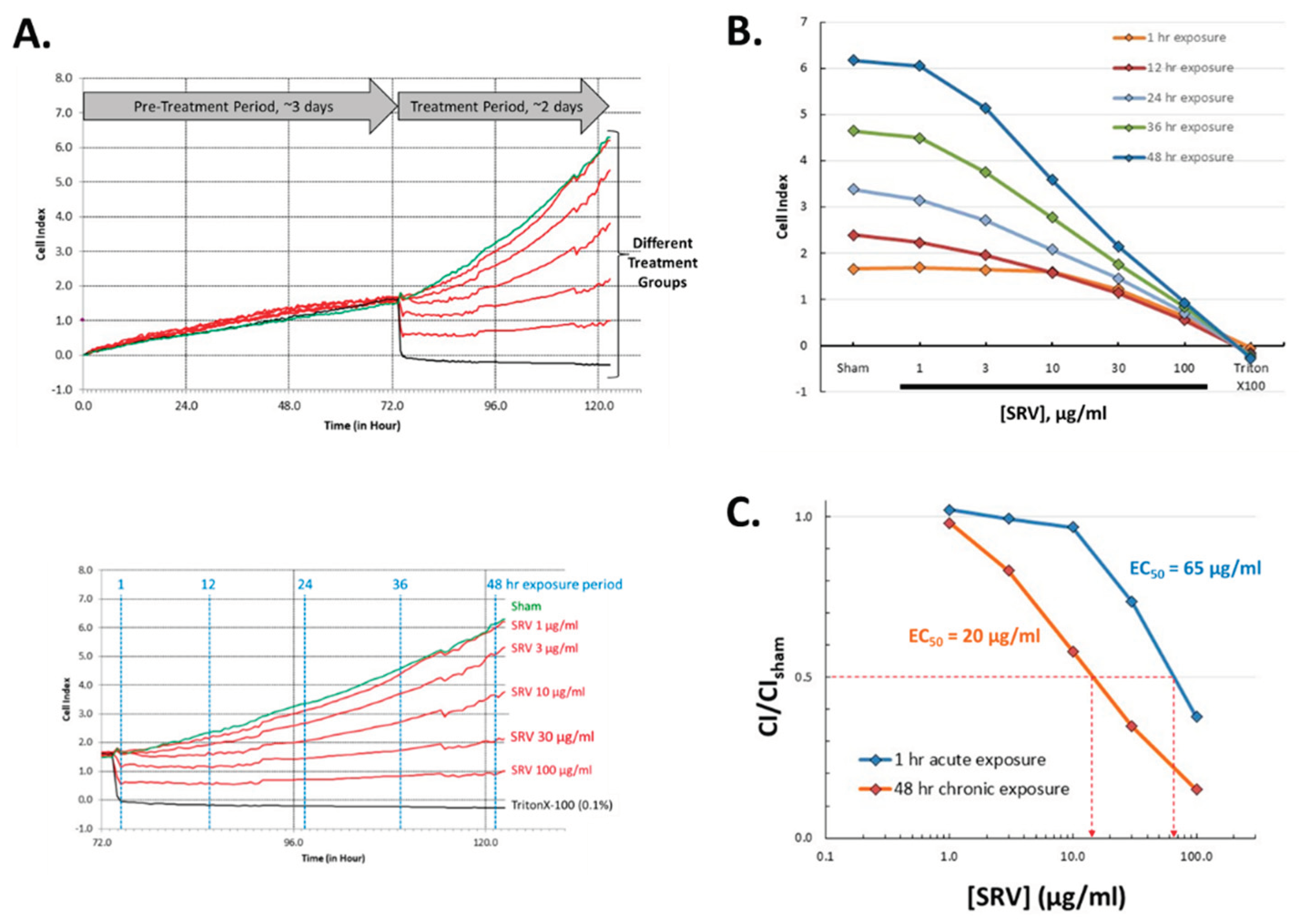
Figure 5.
Heat denaturation of SRV abolishes the rapid cellular detachment effect. a. HDFa cells in log phase growth were either untreated (sham, green trace), treated with 100 μg/mL Atlantic stingray (red trace), or treated with 100 μg/mL Atlantic stingray venom subjected to high heat (purple trace). TritonX-100 treatment (black trace) served as a positive control for cell detachment. b. HDFa cell adherence measured 1 hour after the start of the treatment periods is shown for each experimental group (mean + S.D., n=3, N.S. denotes not statistically significant, *denotes P<0.05).
Figure 5.
Heat denaturation of SRV abolishes the rapid cellular detachment effect. a. HDFa cells in log phase growth were either untreated (sham, green trace), treated with 100 μg/mL Atlantic stingray (red trace), or treated with 100 μg/mL Atlantic stingray venom subjected to high heat (purple trace). TritonX-100 treatment (black trace) served as a positive control for cell detachment. b. HDFa cell adherence measured 1 hour after the start of the treatment periods is shown for each experimental group (mean + S.D., n=3, N.S. denotes not statistically significant, *denotes P<0.05).
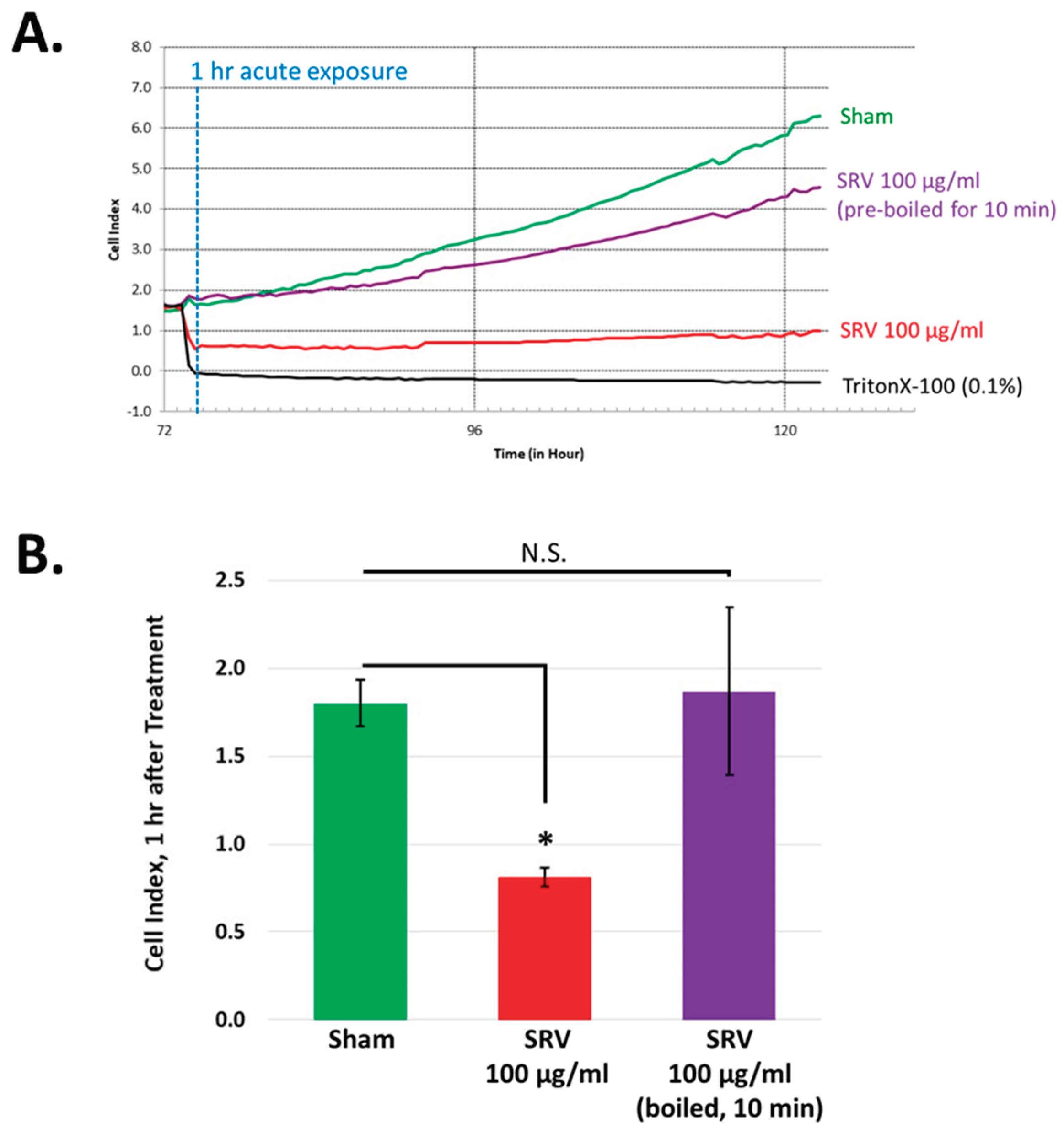
Figure 6.
SRV-mediated inhibition of Jurkat E6-1 cell growth and insensitivity of WEHI 164 cells. a. SRV from cownose rays (R. bonasus), spotted eagle rays (A. narinari) and Atlantic stingrays (H. sabinus) were used to treat Jurkat E6-1 cells for a 24-hour exposure period at varying concentrations. The percent growth inhibition was determined using the MTT assay. b. WEHI 164 fibrosarcoma cells treated with SRV’s under the same conditions described for Jurkat cells. *Significantly different (P<0.05) than untreated control as determined Student’s t-test of OD values of control vs treatments.
Figure 6.
SRV-mediated inhibition of Jurkat E6-1 cell growth and insensitivity of WEHI 164 cells. a. SRV from cownose rays (R. bonasus), spotted eagle rays (A. narinari) and Atlantic stingrays (H. sabinus) were used to treat Jurkat E6-1 cells for a 24-hour exposure period at varying concentrations. The percent growth inhibition was determined using the MTT assay. b. WEHI 164 fibrosarcoma cells treated with SRV’s under the same conditions described for Jurkat cells. *Significantly different (P<0.05) than untreated control as determined Student’s t-test of OD values of control vs treatments.
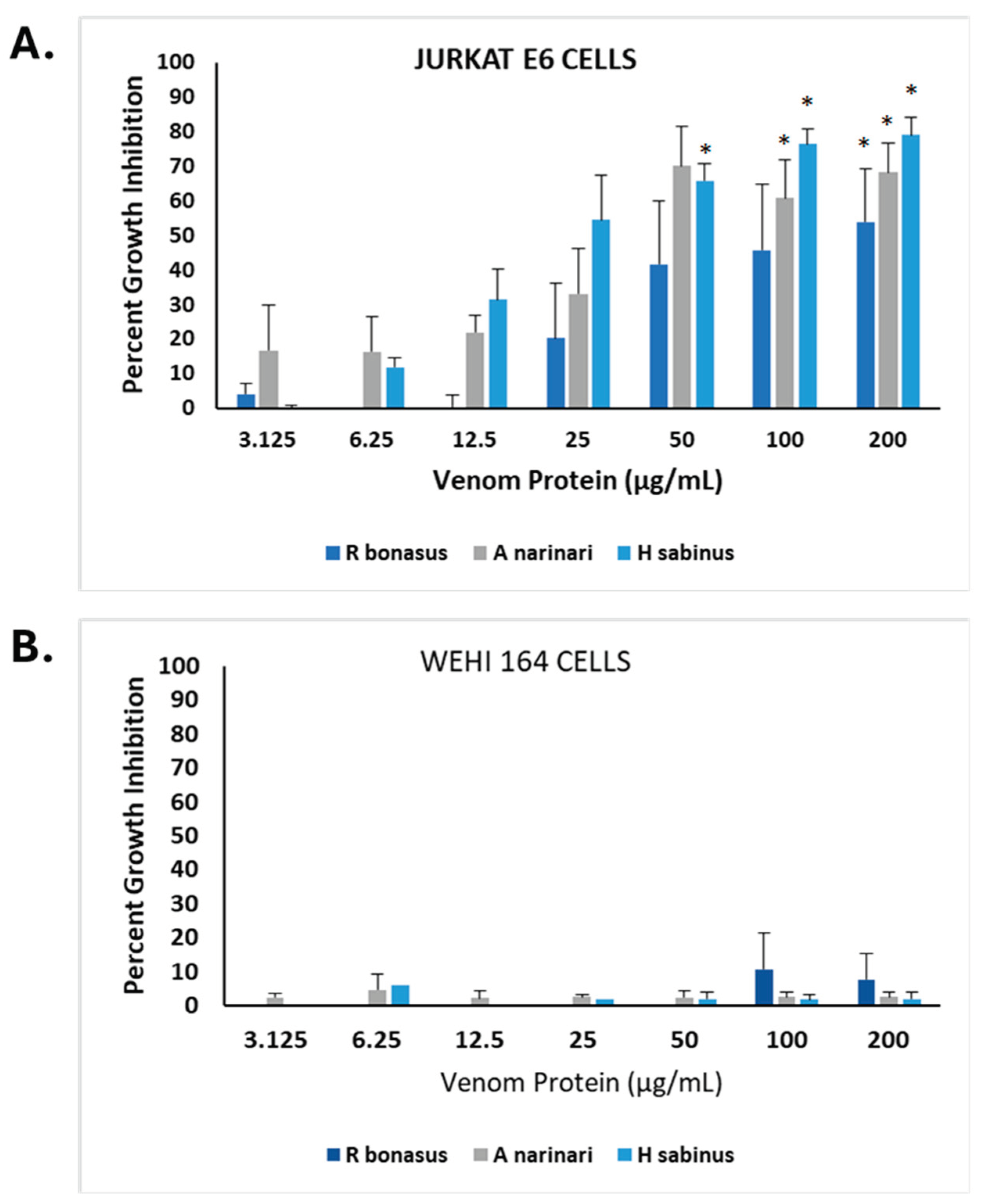
Figure 7.
SRV-mediated apoptosis of Jurkat E6 cells. a. Representative flow cytometry analysis of Jurkat E6-1 cells assessed with FITC-labeled Annexin V binding (FL1-A channel) and propidium iodide staining (FL2-A channel). Healthy viable cells cluster in Quadrant 2 (Q2), apoptotic cells cluster in Quadrant 3 (Q3), and dead or necrotic cells cluster in Quadrant 4 (Q4). b, c. The percentage of Jurkat E6 cells characterized as ‘healthy’, ‘apoptotic’, or ‘necrotic’ following a 24-hour exposure to eagle ray venom (b) or Atlantic stingray venom (c) is plotted over the range of concentrations tested (0 – 200 μg/mL). Staurosporine treatment (Stauro) was used as a positive control for apoptosis, and heat-killing (HK) of cells was used as a positive control for cell necrosis (95 °C for 30 min).
Figure 7.
SRV-mediated apoptosis of Jurkat E6 cells. a. Representative flow cytometry analysis of Jurkat E6-1 cells assessed with FITC-labeled Annexin V binding (FL1-A channel) and propidium iodide staining (FL2-A channel). Healthy viable cells cluster in Quadrant 2 (Q2), apoptotic cells cluster in Quadrant 3 (Q3), and dead or necrotic cells cluster in Quadrant 4 (Q4). b, c. The percentage of Jurkat E6 cells characterized as ‘healthy’, ‘apoptotic’, or ‘necrotic’ following a 24-hour exposure to eagle ray venom (b) or Atlantic stingray venom (c) is plotted over the range of concentrations tested (0 – 200 μg/mL). Staurosporine treatment (Stauro) was used as a positive control for apoptosis, and heat-killing (HK) of cells was used as a positive control for cell necrosis (95 °C for 30 min).
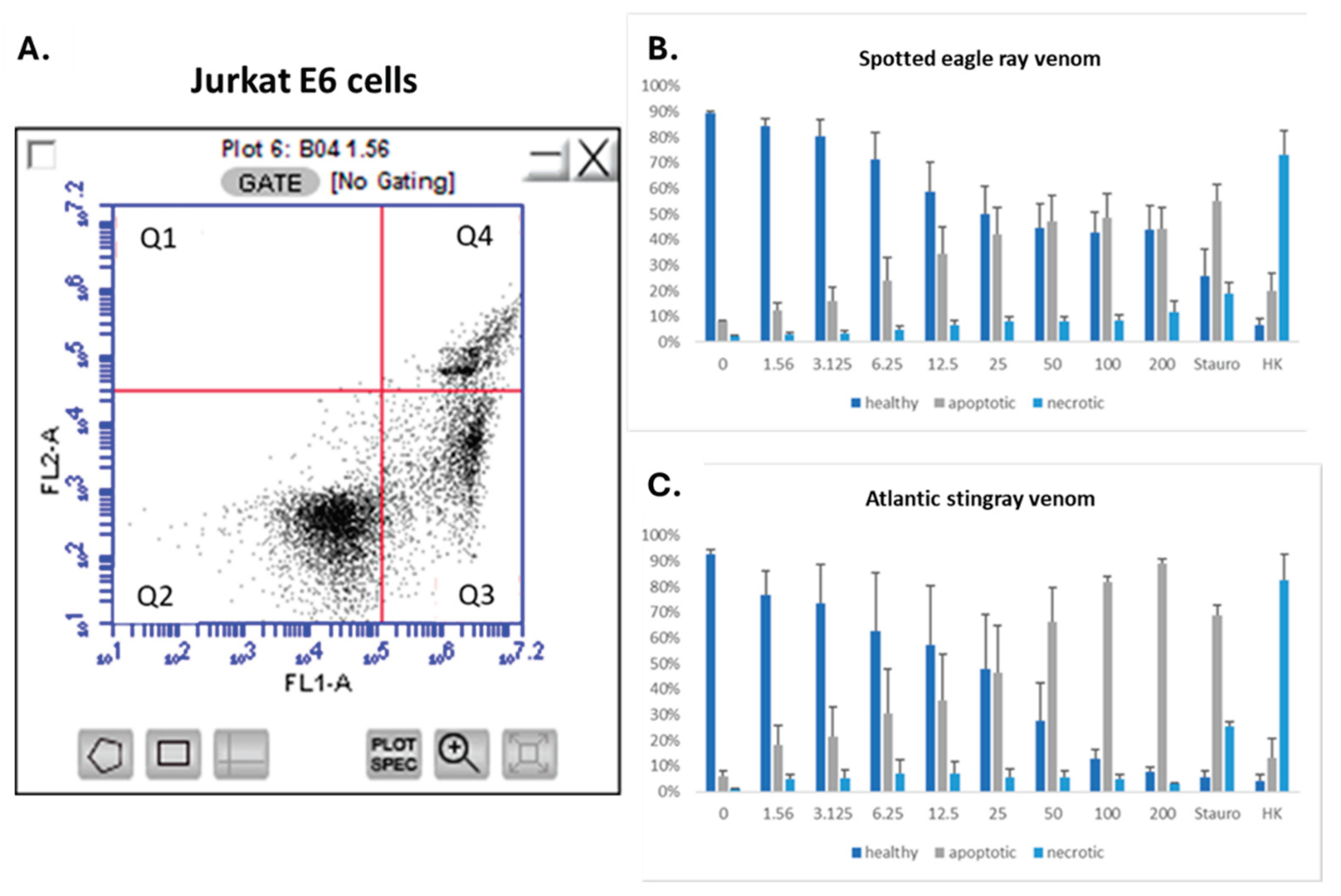

Table 1.
Cell lines used in bioactivity assays.
| Cell Line | ATCC # | Origin | Culture Media | Subculture |
|---|---|---|---|---|
| HDFa | PCS-201-012 | Human, primary dermal fibroblast, normal | Fibroblast Basal Medium (ATCC PCS-201-030); Fibroblast Growth Kit-Low Serum (ATCC PCS-201-041) | At 80-100% confluence |
| SH-SY5Y | CRL-2266 | Human, neuroblastoma | 1:1 Eagle’s Minimum Essential Medium:F12 Medium; 10% FBS | 4-7 days |
| Jurkat E6-1 | TIB-152 | Human, acute T-cell leukemia | RPMI + 10% FBS | 2-3 days |
| WEHI 164 | CRL-1751 | Mouse, fibrosarcoma | RPMI + 10% FBS | 2-3 days |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



