Submitted:
29 February 2024
Posted:
29 February 2024
You are already at the latest version
Abstract
Isothermal Nucleic Acid Amplification based-Lateral Flow Testing (INAA-LFT) has emerged as a robust technique for on-site pathogen detection, providing a visible indication of pathogen nucleic acid amplification that rivals or even surpasses the sensitivity of real-time quantitative PCR. The isothermal nature of INAA-LFT ensures consistent conditions for nucleic acid amplification, establishing it as a crucial technology for rapid on-site pathogen detection. However, despite its considerable promise, the widespread application of isothermal INAA amplification-based lateral flow testing faces several challenges. This review provides an overview of the INAA-LFT procedure, highlighting its advancements in detecting plant viruses. Moreover, the review underscores the imperative to address existing limitations and emphasizes ongoing research efforts dedicated to enhancing the applicability and performance of this technology in the realm of rapid on-site testing.
Keywords:
Isothermal
; lateral flow
; detection
; plant virus
; on-site
; LAMP
; RPA
1. Introduction
Isothermal Nucleic Acid Amplification based-Lateral Flow Testing (INAA-LFT) has emerged as a potent technique for the detection of various pathogens in humans, animals, plants, and the environment [1,2,3]. This innovative testing method involves the amplification of genetic material, such as DNA or RNA, through isothermal amplification, followed by detection utilizing a visual readout on a lateral flow strip (LFS). Notably, INAA-LFT exhibits a considerable advantage over lateral flow immunoassay-based tests due to its heightened sensitivity [3]. The increased sensitivity can be attributed to the amplification process of the pathogen's nucleic acid within the assay [4].
The advent of the COVID-19 epidemic has underscored the critical importance of early and rapid pathogen detection as a primary measure in pathogen prevention and control [5]. INAA-LFT is being actively developed as a pivotal on-site rapid detection technology for pathogens [1,4]. Nevertheless, several key challenges currently impede the widespread utilization of INAA-LFT in on-site rapid detection. We here introduce the procedure of INAA-LFT, offering a comprehensive review of its advancements in the detection of plant viruses. Furthermore, we address the pertinent factors limiting the applicability of this technology for rapid on-site testing, and also discuss ongoing efforts and proposed solutions aimed at enhancing the efficacy of INAA-LFT.
2. Workflow of INAA-LFT
2.1. Amplification of the Target Nucleic Acid Fragments from Pathogens
Under isothermal conditions, nucleic acid amplification techniques such as loop-mediated isothermal amplification (LAMP), recombinase polymerase amplification (RPA), or recombinase-mediated chain replacement nucleic acid amplification (RAA) [3] are employed to amplify target nucleic acid fragments from pathogenic DNA or RNA. The efficient amplification for the target nucleic acid fragments constitutes a critical foundation for the high-sensitivity detection of INAA-LFTs.
2.2. Labeling of the Target Nucleic Acid Fragments from Pathogens
Various methods, including fluorescent dyes, biotin, or gold nanoparticles, are utilized to label the amplified nucleic acid fragments for the visual detection of LFT. Comparative studies of different labeling methods have shown that the modification of primers within the amplification system significantly simplifies the labeling of target nucleic acid fragments, achieving the highest detection sensitivity, which actually is used widely [6].
2.3. Visible Detection of Target Nucleic Acid Fragments from Pathogens
After labeling, the prepared target nucleic acid fragments are ready for visible detection using a paper-based strip. This strip contains specific zones with immobilized proteins capable of binding the labeled target nucleic acid amplification fragments. If the target nucleic acid fragments from pathogens are present in the sample, they will bind to the immobilized capture labels in the detection zone, forming a visible line, indicating a positive result [7]. A control line is also present on the strip to verify the correct functioning of the test. It captures excess labeled markers, and its appearance ensures the validity of the test.
3. Application of INAA-LFT in Detection of Plant Viruses
3.1. LAMP-LFT Detection of Plant Viruses
LAMP technique exhibits superior specificity, efficiency, and rapidity in DNA amplification under isothermal conditions, utilizing a DNA polymerase to amplify targeted DNA strands [8]. Unlike polymerase chain reaction (PCR), which necessitates cycling through various temperature steps, LAMP functions at a constant temperature, typically between 60°C and 65°C. This characteristic obviates the need for a thermal cycler, simplifying equipment requirements and rendering it well-suited for field applications, point-of-care diagnostics, and resource-limited settings [9,10].
The LAMP reaction employs four to six primers targeting multiple regions on the DNA, comprising two outer primers (forward and backward), two inner primers (forward inner and backward inner), and optionally, loop primers that expedite the reaction. In the LAMP reaction, the thermophilic enzyme Bst DNA polymerase, with strand displacement activity, synthesizes DNA at a constant temperature through a self-cycling amplification process. In this process, the displaced strand serves as a template for further amplification, leading to the formation of loop structures that contribute to exponential amplification of the target DNA [9,10]. The endpoint of the reaction can be visually detected through turbidity, fluorescence, or color change, depending on the chosen detection method.
Over the years, LAMP has undergone further development, integrating with other molecular approaches for plant pathogens diagnosis [1,2,8]. As an illustrative case, cassava brown streak virus (CBSV) and ugandan cassava brown streak virus (UCBSV) are causative agents of cassava brown streak disease in East Africa. These viruses, belonging to the genus Ipomovirus, family Potyviridae, possess a positive-sense (+), single-stranded (ss) RNA genome, leading to yield losses and reduced marketability of cassava roots [11]. Tomlinson et al. devised primers for the rapid detection of these viruses through reverse transcription loop-mediated isothermal amplification (RT-LAMP). To label target sequences, markers such as fluorescein isothiocyanate (FITC), biotin, or digoxigenin (DIG) were incorporated with primers. RT-LAMP achieved amplification within 40 minutes, and the products were detectable using lateral flow devices containing antibodies specific to the incorporated labels [11].
A similar strategy was employed for the detection of tobacco rattle virus (TRV), another plant virus with a +ssRNA genome. Edgu et al. developed and optimized a mini-LAMP-lateral flow device (LFD) approach for the sensitive and specific detection of TRV in potatoes. This approach offers an economical and efficient platform for disease management in potato breeding and cultivation [12]. Notably, viral RNA purification was circumvented, and the filtered supernatant of incubation samples was diluted 1:100 with water and directly used for amplification, simplifying sample processing without the need for sophisticated laboratory equipment [12].
Recently, Lu et al. identified a novel member of the genus Badnavirus in the family Caulimoviridae, named Chinaberry tree badnavirus 1 (ChTBV1), which harbors a single molecule of non-covalently closed circular double-stranded (ds) DNA in the Chinaberry tree. They developed a LAMP assay for viral detection and adapted it for rapid visualization of results using a lateral flow dipstick chromatographic detection method [13].
3.2. RPA-LFT Detection of Plant Viruses
RPA, an additional molecular biology technique utilized for the isothermal amplification of DNA [14], shares similarities with LAMP in its capacity to facilitate DNA amplification at a constant temperature. This feature positions RPA as well-suited for field applications and point-of-care diagnostics, addressing challenges associated with maintaining precise temperature control [15,16].
The RPA process encompasses several pivotal components, including recombinase enzymes such as recombinase A, RecA, or recombinase UvsX. These enzymes play a crucial role in facilitating strand-exchange reactions and promoting the invasion of primers into the target DNA. Additionally, the Single-stranded DNA-binding Protein (SSB) is indispensable for stabilizing single-stranded DNA regions, preventing reannealing. Two primers, each possessing homologous regions to the target DNA, recognize specific sequences and bind to opposite strands. The DNA polymerase, featuring strand displacement activity, extends the primers and synthesizes new DNA strands. Significantly, the reaction is conducted isothermally at a constant temperature, typically ranging between 37°C and 42°C.
In contrast to the LAMP-LFT amplification system, which necessitates 4~6 primers, the RPA-LFT amplification system only requires three primers for amplifying a target gene—forward and reverse primers and probes. The outer primer pair can generate specific and cloned amplification products, ensuring the accuracy of the amplification target. This streamlined primer requirement in the RPA-LFT system reduces the complexity of primer design, enhances the accuracy of detection, and lowers the detection cost compared to the more intricate primer design of the LAMP-LFT system.
RPA finds diverse applications, including molecular diagnostics, environmental monitoring, and field-based pathogen detection [15,17]. Its isothermal nature renders it suitable for resource-limited settings, and the relatively short reaction time positions it as a valuable tool for rapid DNA amplification. By incorporating specially modified probe primers into the amplification system and collaborating with LFT, RPA-LFT has emerged as a primary method for diagnosing plant viruses with varied genome types, encompassing +ssRNA, negative sense (-) ssRNA, ambisense RNA (±RNA), dsRNA, ssDNA, dsDNA, and even viroids with naked circle RNA [1,6,18,19,20,21,22,23,24,25,26,27,28,29,30] (Table 1).
Little cherry virus 2 (LChV2), belonging to the genus Ampelovirus in the family Closteroviridae with a +ssRNA genome, causes little cherry disease (LCD) in sweet cherries (Prunus avium) globally. Early detection of LChV2 is crucial for controlling LCD [31]. Mekuria et al. devised an effective diagnostic method based on RPA-LFT. They developed a simple, fast, and specific RT-RPA method utilizing LChV2 coat protein-specific primers and probes, exhibiting comparable sensitivity to RT-PCR from crude extracts. The terminally labeled amplicons were detected using a high-affinity lateral flow strip [31]. A similar approach was applied for the detection of another Closteroviridae family member, citrus tristeza virus (CTV), in the genus Closterovirus [30]. This method proves to be a powerful tool for early-stage virus detection in field samples [30,31].
Plum pox virus (PPV), a member of the genus Potyvirus in the family Potyviridae with a +ssRNA genome, causes the devastating plum pox or Sharka disease in stone fruit trees. Zhang et al. developed an efficient RPA-LFT method for PPV detection, significantly reducing the diagnostic time to as little as 20 minutes for the entire process from sample preparation to result. This innovation streamlines diagnosis, facilitating both laboratory and field applications [18].
Alfalfa mosaic virus (AMV), a plant virus belonging to the genus Alfamovirus in the family Bromoviridae with a +ssRNA genome, affects a wide range of plant species worldwide. Ivanov et al. compared two methods generating labeled RPA amplicons of AMV and found that the RPA-LFT assay based on primer labeling detected 103 copies of RNA in 30 minutes with a half-maximal binding concentration 22 times lower than the probe-dependent RPA-LFT. This indicates the simplicity and efficiency of labeling primers for RPA-LFT in viral diagnosis [6].
Recently, several other +ssRNA genome viruses have been targeted for detection by RPA-LFT, including cymbidium mosaic virus (CymMV) in the genus Potexvirus of the family Alphaflexiviridae, barley yellow dwarf virus (BYDV) in the genus Polerovirus of the family Solemoviridae, cowpea mild mottle virus (CPMMV) in the genus Carlavirus of the family Betaflexiviridae, actinidia chlorotic ringspot-associated virus (AcCRaV) in the genus Emaravirus of the family Fimoviridae, and bean pod mottle virus (BPMV) in the genus Comovirus of the family Secoviridae [21,22,23,26,28]. The developed RPA-LFT assay for these viruses exhibited 100 times more sensitivity than conventional reverse transcription polymerase chain reaction (RT-PCR), providing a simple, rapid, sensitive, and reliable method for viral diagnosis in the field.
Tomato spotted wilt virus (TSWV) and tomato chlorotic spot virus (TCSV), members of the genus Orthotospovirus in the family Tospoviridae with ambisense RNA genomes, cause significant yield loss in ornamental and vegetable crops worldwide. RPA-LFT assays for both viruses have been developed [27,32]. Furthermore, in the RPA reaction for TCSV, crude RNAs in the tube are incubated in the palm of the hand to generate sufficient heat for amplification. The detection limit is approximately 6 pg/µL of total RNA from samples, providing an equipment-free, body-heat-mediated RT-RPA-LFA technique [27]. Correspondingly, methods based on RPA-LFT were developed for the detection of plant viruses with dsRNA, ssDNA, and dsDNA genomes [19,20,24,25].
Moreover, a multiplex assay based on RPA-LFT to detect two or more kinds of plant viruses has also been developed. Ivanov et al. successfully applied this strategy for the detection of three priority potato RNA viruses: potato virus Y (PVY), potato virus S (PVS), and potato leafroll virus (PLRV). The total assay time is 30 minutes. The multiplex RPA-LFT demonstrated the capability to detect at least 4 ng of PVY per gram of plant leaves, 0.04 ng/g for PVS, and 0.04 ng/g for PLRV [33]. Multiplex assays capable of simultaneously detecting multiple plant viruses provide a more comprehensive diagnostic approach, especially in regions where multiple viral pathogens may be prevalent.
3.3. RAA-LFT Detection of Plant Viruses
RAA shares a fundamental principle with RPA. The distinguishing factor between these two methods lies in the source of the recombinase. RPA utilizes recombinase from the T4 phage, while RAA employs recombinases from diverse sources such as bacteria and fungi. RAA has been applied to detect several viruses, showcasing its versatility in molecular diagnostics. Maize chlorotic mottle virus (MCMV) has emerged as a significant threat to maize production globally, causing maize lethal necrosis in regions of East Africa, South America, and Asia [34,35]. Duan et al. combined RAA with a CRISPR/Cas12a (Clustered Regularly Interspaced Short Palindromic Repeats and CRISPR-associated proteins 12a)-based visual nucleic acid detection system for MCMV, achieving a rapid and sensitive process completed within 45 minutes [36]. Expanding on this strategy, Wang et al. developed a detection system targeting sorghum mosaic virus and rice stripe mosaic virus [37]. Recently, Zhao et al. integrated LFT with RAA, creating a visible system for MCMV detection [38]. Tomato brown rugose fruit virus (ToBRFV), a member of the Tobamovirus genus, has recently become pandemic in tomato and pepper production areas worldwide. Cao et al. devised an RAA-LFT for field detection of ToBRFV with high sensitivity, demonstrating a detection limit of 2.1 × 101 copies/50-μL reaction [39]. Subsequently, Zhao et al. combined RAA and CRISPR/Cas12a with LFT, enabling simultaneous detection of four tobamoviruses—pepper mild mottle virus (PMMoV), ToBRFV, tomato mosaic virus (ToMV), and tomato mottle mosaic virus (ToMMV) [40].
3.4. CRISPR-CAS System-Integrated LFT Detection of Plant Viruses
The CRISPR-CAS technology, renowned for its proficiency in genome editing, has found application in the INAA-LFT for plant virus detection, augmenting both specificity and sensitivity. In this context, CRISPR-CAS systems function to recognize and bind to specific amplified sequences generated by PCR, LAMP, or RPA/RAA. The CRISPR RNA (guide RNA) is meticulously designed to complement the target sequence, and the CAS protein (such as Cas12 or Cas13) undergoes activation upon binding, leading to the cleavage of the target sequence. The activation is concomitant with a detectable signal, often manifesting as a fluorescence signal. The presence of the target sequence can be identified by interpreting this signal. The integration of CRISPR-Cas with RPA confers a robust tool for nucleic acid detection, enhancing specificity through the highly precise binding of the CRISPR-Cas system to target sequences.
Marques et al. harnessed CRISPR-Cas12a and CRISPR-Cas13a/d systems for detecting viral DNA amplicons generated by PCR or isothermal amplification, focusing on three RNA viruses: tobacco mosaic virus (TMV), tobacco etch virus (TEV), and potato virus X (PVX). They innovatively adapted the detection system to circumvent the costly RNA purification step and achieve a visible readout through lateral flow strips, enabling rapid viral diagnostics within a timeframe of half an hour [41]. Addressing key viruses in rice, rice stripe virus (RSV) and rice black-streaked dwarf virus (RBSDV), Zhu et al. devised a CRISPR/Cas12a-assisted LAMP-LFT system to detect these viruses along with the bacterial pathogen Xanthomonas oryzae pv. oryzae (Xoo). The heightened sensitivity of this system reached as low as 9 or 3 copies [42]. In the LAMP-LFT assay for MCMV detection, assisted by CRISPR-Cas12a, the detection limit achieved an impressive low of 2.5 copies of the coat protein (CP) gene of MCMV [38].
Results above indicate that that the assistance of CRISPR-CAS in the detection system improve the detection limit to a very low level. Meanwhile, it should be noted that the specific implementation of CRISPR-CAS in detection may vary based on the target application and the desired detection method. It requires continued exploration and optimization of these technologies for a variety of diagnostic and research purposes.
4. Factors Influencing the Implementation of INAA-LFT for On-Site Detection of Plant Viruses
The utilization of INAA-LFT for on-site detection of plant viruses is intricately influenced by several key factors, including sensitivity and specificity, rapid testing duration, portability and field-friendly device characteristics, and cost-effectiveness, among others. These factors play a pivotal role in shaping the feasibility, effectiveness, and practicality of implementing INAA-LFT in field settings.
4.1. Sensitivity and Specificity
Detection sensitivity stands as a pivotal metric when assessing a particular detection technology. Within the established INAA-LFT systems, detection limits typically fall within the range of 2.5 to 20 viral copies per reaction, often surpassing or reaching the detection threshold of real-time quantitative PCR. This level of sensitivity adequately caters to the requirements of field detection [26,28,38,41]. Notably, when diagnosing a plant sample exhibiting virus symptoms, the sample generally contains a sufficient viral load for detection by INAA-LFT, thus mitigating the necessity for exceptionally high detection sensitivity. However, in scenarios involving the early detection of samples, where virus symptoms may not be apparent, and the goal is to ascertain the presence of a specific virus, a heightened detection sensitivity becomes imperative.
The accurate detection of viral nucleic acids by INAA-LFT is pivotal, and higher specificity is instrumental in mitigating false positives, particularly in samples infected with multiple viruses. This necessitates stringent requirements for primer design during the nucleic acid amplification process, a critical consideration not only for INAA-LFT but also for other nucleic acid-based detection technologies. Furthermore, in the context of multiplex assays aiming to detect two or more distinct viruses, even within the same genus, the significance and complexity of primer design are heightened—a formidable challenge for researchers [33,39].
4.2. Detection Duration
The rapid generation of results is pivotal for on-site detection. The speed of INAA-LFT significantly influences the ability to provide timely confirmations in field settings. The current testing system typically completes the entire process, from sample collection to the presentation of test results, within one hour. The fastest recorded duration is 20 minutes, while the slowest extends to about 50 min [19,26,30,41]. Although there is potential to optimize the system framework to achieve a testing duration of 20 minutes, this timeframe still falls short of meeting the demand for rapid on-site detection of plant viruses [19,26,30].
4.3. Ease of Operation
In the reaction process of INAA-LFT, many components are required to exert biochemical activity to achieve the amplification of nucleic acids, the generation of signals, and the visualization of signals. Under laboratory conditions, these components usually participate in the reaction in order to ensure the stability of the reaction system and avoid the occurrence of various false results. In the INAA-LFT currently established detection system for plant virus, scientific researchers have tried their best to mix various ingredients into a tube (all in one) and simplify the operating steps as much as possible without affecting the activity of each ingredient. For example, reverse transcription is combined into amplification reaction, and nucleic acid amplification is combined with CRISPR/Cas12a in one mixture
In the intricate reaction process of INAA-LFT, various components must exhibit biochemical activity to achieve nucleic acid amplification, signal generation, and signal visualization. In laboratory conditions, these components are meticulously curated to ensure reaction system stability and mitigate the risk of false results. In the currently established INAA-LFT detection system for plant viruses, researchers have diligently worked towards consolidating multiple components into a single reaction tube (all in one) and simplifying operational steps without compromising the efficacy of each component. Notably, processes such as reverse transcription are amalgamated into the amplification reaction, and nucleic acid amplification is seamlessly integrated with CRISPR/Cas12a within a single mixture [28,38,39,42]. Moreover, the development of a nucleic acid extraction-free process, seamlessly incorporated into the all-in-one reaction, further streamlines the INAA-LFT steps, enhancing simplicity [25,28,31,38,39].
However, based on our practical experience, in actual field operations, operators are still required to utilize pipettes for sample addition, buffer incorporation, and subsequent product dilution. This necessity raises the basic technical threshold for operators and concurrently poses limitations on the broader application of INAA-LFT in field detection.
5. The Future Trajectory of INAA-LFT in Detection of Plant Virus
5.1. Expedited Detecting Duration
In the existing literature on the detection of plant viruses using INAA-LFT, the detection time typically spans from 20 to 50 minutes, posing a limitation for widespread on-site applications. The primary time is allocated to nucleic acid amplification, determined by the performance of the recombinase in the system. Nucleic acid amplification usually takes around 15 minutes to reach a detectable level. If this time can be reduced to less than 8-10 minutes, it would significantly advance the practical use of this detection technology in the field (Figure 1). Despite the successful integration of reverse transcription, nucleic acid amplification, and signal production into one tube, the challenge remains in simplifying or omitting operational steps. The quest for novel recombinases with higher activity holds promise for achieving faster INAA-LFT detection. Additionally, exploring alternative amplification strategies or optimizing reaction conditions may contribute to time reduction.
5.2. Operational Simplification and Automation
The envisioned scenario involves field personnel, without specialized molecular technique training, effortlessly conducting INAA-LFT procedures and interpreting results. Presently, several pipetting steps are essential, necessitating further optimization. The developmental focus on simplifying the system, enhancing user-friendliness, and potential automation of operations is a promising direction. Integrated systems managing sample preparation, testing, and result analysis could revolutionize the landscape. Moreover, coupling INAA-LFT with advanced data management systems has the potential to facilitate result collection, storage, and analysis, contributing to the establishment of a comprehensive database for monitoring and managing plant virus outbreaks (Figure 1). Additionally, exploring technologies such as robotics or microfluidics could further enhance automation and ease of use.
5.3. Portable Devices
On-site INAA-LFT detection requires specific instruments, and the absence of a commercially available all-in-one machine poses a current limitation. The ideal INAA-LFT device should be portable, tailored for field use, and resistant to environmental conditions prevalent in agricultural settings. Considerations include size, weight, durability, testing throughput, and resilience against varying environmental conditions. Recent innovations, such as our development of a portable toolbox powered by a car cigarette lighter, showcase progress in addressing portability concerns. However, the existing system still involves several pipetting steps, highlighting the need for further streamlining. Anticipating all-in-one machines that integrate sample handling, automated pipetting, result interpretation, GPS positioning, and multi-environment adaptability is a pivotal advancement eagerly awaited in the field (Figure 1). Moreover, the integration of advanced energy sources, such as solar power, could play a transformative role in enhancing the sustainability and portability of these devices, making them more practical and efficient for on-site detection applications.
5.4. Regulatory Approvals and Standardization
Regulatory considerations and adherence to standards play a pivotal role in shaping the future trajectory of a technology. Currently, fluorescence quantitative real-time polymerase chain reaction (qPCR) remains the gold standard for the sensitive and specific detection and quantification of pathogenic nucleic acids. In contrast, INAA-LFT-based assays lack standardized protocols, relying on individual laboratory conditions and procedures. A fundamental reason for this disparity is the absence of a standardized instrument. Establishing a unified protocol for INAA-LFT, grounded in standard instruments, is imperative to ensure consistency and reliability across different tests and platforms. Collaborative efforts between researchers, regulatory bodies, and industry stakeholders are essential to define and implement standardized procedures (Figure 1). Additionally, emphasizing the importance of validation studies and external quality control measures will bolster the credibility and acceptance of INAA-LFT in the scientific community and regulatory frameworks.
5.5. Integration of Artificial Intelligence (AI) for Data Analysis
With the continuous advancement of technology, the integration of Artificial Intelligence (AI) in the colorimetric data analysis of LFT can further enhance the efficiency and accuracy of result interpretation in the context of INAA-LFT. For instance, harnessing the capabilities of AI algorithms to swiftly process large datasets, AI can identify colorimetric patterns in LFT and, based on the standard colorimetric chart for absolute quantification of a pathogen in LFT, estimate the copy number of the pathogen in actual samples (Figure 1). This approach contributes to presenting more robust and reliable virus detection results. Exploring the potential synergies between INAA-LFT and AI will open new avenues for improving the overall performance and applicability of this technology in diverse settings.
Author Contributions
Conceptualization, X.S. and F.Y.; writing—original draft preparation, X.S., Y.C. and F.Y.; writing—review and editing, X.S. and F.Y.. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Chinese Agriculture Research System of MOF and MARA, grant number CARS-24-C-04.
Data Availability Statement
No new data was created in this work.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bhat, A.I.; Aman, R.; Mahfouz, M. Onsite detection of plant viruses using isothermal amplification assays. Plant Biotechnol J 2022, 20, 1859–1873. [Google Scholar] [CrossRef] [PubMed]
- Wong, Y.P.; Othman, S.; Lau, Y.L.; Radu, S.; Chee, H.Y. Loop-mediated isothermal amplification (LAMP): a versatile technique for detection of micro-organisms. J Appl Microbiol 2018, 124, 626–643. [Google Scholar] [CrossRef] [PubMed]
- Boonham, N.; Kreuze, J.; Winter, S.; van der Vlugt, R.; Bergervoet, J.; Tomlinson, J.; Mumford, R. Methods in virus diagnostics: from ELISA to next generation sequencing. Virus Res 2014, 186, 20–31. [Google Scholar] [CrossRef]
- Liu, Y.; Zhan, L.; Qin, Z.; Sackrison, J.; Bischof, J.C. Ultrasensitive and Highly Specific Lateral Flow Assays for Point-of-Care Diagnosis. ACS Nano 2021, 15, 3593–3611. [Google Scholar] [CrossRef] [PubMed]
- Lefrancois, T.; Malvy, D.; Atlani-Duault, L.; Benamouzig, D.; Druais, P.L.; Yazdanpanah, Y.; Delfraissy, J.F.; Lina, B. After 2 years of the COVID-19 pandemic, translating One Health into action is urgent. Lancet 2023, 401, 789–794. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, A.V.; Safenkova, I.V.; Zherdev, A.V.; Dzantiev, B.B. Recombinase Polymerase Amplification Assay with and without Nuclease-Dependent-Labeled Oligonucleotide Probe. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef]
- Sang, P.; Hu, Z.; Cheng, Y.; Yu, H.; Xie, Y.; Yao, W.; Guo, Y.; Qian, H. Nucleic Acid Amplification Techniques in Immunoassay: An Integrated Approach with Hybrid Performance. J Agric Food Chem 2021, 69, 5783–5797. [Google Scholar] [CrossRef]
- Notomi, T.; Okayama, H.; Masubuchi, H.; Yonekawa, T.; Watanabe, K.; Amino, N.; Hase, T. Loop-mediated isothermal amplification of DNA. Nucleic Acids Res 2000, 28, E63. [Google Scholar] [CrossRef]
- Mori, Y.; Notomi, T. Loop-mediated isothermal amplification (LAMP): a rapid, accurate, and cost-effective diagnostic method for infectious diseases. J Infect Chemother 2009, 15, 62–69. [Google Scholar] [CrossRef]
- Gill, P.; Ghaemi, A. Nucleic acid isothermal amplification technologies: a review. Nucleosides Nucleotides Nucleic Acids 2008, 27, 224–243. [Google Scholar] [CrossRef]
- Tomlinson, J.A.; Ostoja-Starzewska, S.; Adams, I.P.; Miano, D.W.; Abidrabo, P.; Kinyua, Z.; Alicai, T.; Dickinson, M.J.; Peters, D.; Boonham, N. , et al. Loop-mediated isothermal amplification for rapid detection of the causal agents of cassava brown streak disease. J Virol Methods 2013, 191, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Edgu, G.; Freund, L.J.; Hartje, S.; Tacke, E.; Hofferbert, H.R.; Twyman, R.M.; Noll, G.A.; Muth, J.; Prufer, D. Fast, Precise, and Reliable Multiplex Detection of Potato Viruses by Loop-Mediated Isothermal Amplification. Int J Mol Sci 2020, 21. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Tang, J.; Sun, K.; Yu, X. Identification of a New Badnavirus in the Chinaberry (Melia azedarach) Tree and Establishment of a LAMP-LFD Assay for Its Rapid and Visual Detection. Viruses 2021, 13. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Easley, C.J. Isothermal DNA amplification in bioanalysis: strategies and applications. Bioanalysis 2011, 3, 227–239. [Google Scholar] [CrossRef] [PubMed]
- Daher, R.K.; Stewart, G.; Boissinot, M.; Bergeron, M.G. Recombinase Polymerase Amplification for Diagnostic Applications. Clin Chem 2016, 62, 947–958. [Google Scholar] [CrossRef] [PubMed]
- Deng, H.; Gao, Z. Bioanalytical applications of isothermal nucleic acid amplification techniques. Anal Chim Acta 2015, 853, 30–45. [Google Scholar] [CrossRef] [PubMed]
- Lau, H.Y.; Botella, J.R. Advanced DNA-Based Point-of-Care Diagnostic Methods for Plant Diseases Detection. Front Plant Sci 2017, 8, 2016. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Ravelonandro, M.; Russell, P.; McOwen, N.; Briard, P.; Bohannon, S.; Vrient, A. Rapid diagnostic detection of plum pox virus in Prunus plants by isothermal AmplifyRP((R)) using reverse transcription-recombinase polymerase amplification. J Virol Methods 2014, 207, 114–120. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Sun, F.; Li, X.; Lan, Y.; Du, L.; Zhou, T.; Zhou, Y. Reverse transcription-recombinase polymerase amplification combined with lateral flow strip for detection of rice black-streaked dwarf virus in plants. J Virol Methods 2019, 263, 96–100. [Google Scholar] [CrossRef]
- Cao, Y.; Yan, D.; Wu, X.; Chen, Z.; Lai, Y.; Lv, L.; Yan, F.; Chen, J.; Zheng, H.; Song, X. Rapid and visual detection of milk vetch dwarf virus using recombinase polymerase amplification combined with lateral flow strips. Virol J 2020, 17, 102. [Google Scholar] [CrossRef]
- Kim, D.H.; Jeong, R.D.; Choi, S.; Ju, H.J.; Yoon, J.Y. Application of Rapid and Reliable Detection of Cymbidium Mosaic Virus by Reverse Transcription Recombinase Polymerase Amplification Combined with Lateral Flow Immunoassay. Plant Pathol J 2022, 38, 665–672. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.K.; Lee, H.J.; Kim, S.M.; Jeong, R.D. Rapid and Visual Detection of Barley Yellow Dwarf Virus by Reverse Transcription Recombinase Polymerase Amplification with Lateral Flow Strips. Plant Pathol J 2022, 38, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.Q.; Zhao, X.X.; Wang, D.; Zhang, P.J.; Hu, X.N.; Wei, S.; Liu, J.Y.; Ye, Z.H.; Yu, X.P. A reverse transcription-cross-priming amplification method with lateral flow dipstick assay for the rapid detection of Bean pod mottle virus. Sci Rep 2022, 12, 681. [Google Scholar] [CrossRef]
- Zhou, Y.; Zheng, H.Y.; Jiang, D.M.; Liu, M.; Zhang, W.; Yan, J.Y. A rapid detection of tomato yellow leaf curl virus using recombinase polymerase amplification-lateral flow dipstick assay. Lett Appl Microbiol 2022, 74, 640–646. [Google Scholar] [CrossRef] [PubMed]
- Greeshma, M.; Bhat, A.I.; Jeevalatha, A. Rapid onsite detection of piper yellow mottle virus infecting black pepper by recombinase polymerase amplification-lateral flow assay (RPA-LFA). J Virol Methods 2023, 315, 114695. [Google Scholar] [CrossRef]
- Wu, X.; Chen, S.; Zhang, Z.; Zhang, Y.; Li, P.; Chen, X.; Liu, M.; Lu, Q.; Li, Z.; Wei, Z. , et al. Development of Recombinase Polymerase Amplification Combined with Lateral Flow Strips for Rapid Detection of Cowpea Mild Mottle Virus. Plant Pathol J 2023, 39, 486–493. [Google Scholar] [CrossRef]
- Yilmaz, S.; Batuman, O. Development of a reverse transcription recombinase polymerase amplification combined with lateral flow assay for equipment-free on-site field detection of tomato chlorotic spot virus. Virol J 2023, 20, 136. [Google Scholar] [CrossRef]
- Zhang, A.L.; Shi, X.; Xie, C.; Yu, F.; Gao, Z.; Xu, Y.; Liu, Z. Rapid and Visual Detection of Actinidia Chlorotic Ringspot-Associated Virus Using One-Step Reverse-Transcription Recombinase Polymerase Amplification Combined with Lateral Flow Dipstick Assay. Plant Dis, 1094. [Google Scholar] [CrossRef]
- Hammond, R.W.; Zhang, S. Development of a rapid diagnostic assay for the detection of tomato chlorotic dwarf viroid based on isothermal reverse-transcription-recombinase polymerase amplification. J Virol Methods 2016, 236, 62–67. [Google Scholar] [CrossRef]
- Ghosh, D.K.; Kokane, S.B.; Gowda, S. Development of a reverse transcription recombinase polymerase based isothermal amplification coupled with lateral flow immunochromatographic assay (CTV-RT-RPA-LFICA) for rapid detection of Citrus tristeza virus. Sci Rep 2020, 10, 20593. [Google Scholar] [CrossRef]
- Mekuria, T.A.; Zhang, S.; Eastwell, K.C. Rapid and sensitive detection of Little cherry virus 2 using isothermal reverse transcription-recombinase polymerase amplification. J Virol Methods 2014, 205, 24–30. [Google Scholar] [CrossRef]
- Lee, H.J.; Cho, I.S.; Ju, H.J.; Jeong, R.D. Rapid and visual detection of tomato spotted wilt virus using recombinase polymerase amplification combined with lateral flow strips. Mol Cell Probes 2021, 57, 101727. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, A.V.; Safenkova, I.V.; Zherdev, A.V.; Dzantiev, B.B. Multiplex Assay of Viruses Integrating Recombinase Polymerase Amplification, Barcode-Anti-Barcode Pairs, Blocking Anti-Primers, and Lateral Flow Assay. Anal Chem 2021, 93, 13641–13650. [Google Scholar] [CrossRef] [PubMed]
- Redinbaugh, M.G.; Stewart, L.R. Maize Lethal Necrosis: An Emerging, Synergistic Viral Disease. Annu Rev Virol 2018, 5, 301–322. [Google Scholar] [CrossRef] [PubMed]
- Bernardo, P.; Frey, T.S.; Barriball, K.; Paul, P.A.; Willie, K.; Mezzalama, M.; Kimani, E.; Mugambi, C.; Wangai, A.; Prasanna, B.M. , et al. Detection of Diverse Maize Chlorotic Mottle Virus Isolates in Maize Seed. Plant Dis 2021, 105, 1596–1601. [Google Scholar] [CrossRef]
- Duan, X.; Ma, W.; Jiao, Z.; Tian, Y.; Ismail, R.G.; Zhou, T.; Fan, Z. Reverse transcription-recombinase-aided amplification and CRISPR/Cas12a-based visual detection of maize chlorotic mottle virus. Phytopathol Res 2022, 4, 23. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Huang, X.; Chen, S.; Chen, J.; Liang, Z.; Chen, B.; Yang, X.; Zhou, G.; Zhang, T. On-site and visual detection of sorghum mosaic virus and rice stripe mosaic virus based on reverse transcription-recombinase-aided amplification and CRISPR/Cas12a. Front Genome Ed 2023, 5, 1124794. [Google Scholar] [CrossRef]
- Lei, R.; Kuang, R.; Peng, X.; Jiao, Z.; Zhao, Z.; Cong, H.; Fan, Z.; Zhang, Y. Portable rapid detection of maize chlorotic mottle virus using RT-RAA/CRISPR-Cas12a based lateral flow assay. Front Plant Sci 2023, 14, 1088544. [Google Scholar] [CrossRef]
- Zhao, Z.; Wang, S.; Dong, Z.; Fan, Q.; Lei, R.; Kuang, R.; Zhang, Y. One-Step Reverse-Transcription Recombinase-Aided Amplification CRISPR/Cas12a-Based Lateral Flow Assay for Fast Field Screening and Accurate Differentiation of Four Major Tobamoviruses Infecting Tomato and Pepper. J Agric Food Chem, 1021. [Google Scholar] [CrossRef]
- Marques, M.C.; Sanchez-Vicente, J.; Ruiz, R.; Montagud-Martinez, R.; Marquez-Costa, R.; Gomez, G.; Carbonell, A.; Daros, J.A.; Rodrigo, G. Diagnostics of Infections Produced by the Plant Viruses TMV, TEV, and PVX with CRISPR-Cas12 and CRISPR-Cas13. ACS Synth Biol 2022, 11, 2384–2393. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Li, R.; Zhang, H.; Wang, J.; Lu, Y.; Zhang, D.; Yang, L. PAM-free loop-mediated isothermal amplification coupled with CRISPR/Cas12a cleavage (Cas-PfLAMP) for rapid detection of rice pathogens. Biosens Bioelectron 2022, 204, 114076. [Google Scholar] [CrossRef]
- Cao, Y.; Weng, H.; Rao, S.; Li, J.; Yan, F.; Song, X. Rapid and visual field diagnosis of tomato brown rugose fruit virus using reverse transcription recombinase aided amplification (RT RAA) combined with lateral flow strips (LFS). Crop Protection 2023, 173, 106355. [Google Scholar] [CrossRef]
Figure 1.
Workflow of INAA-LFT now and its future trajectory in detection of plant virus. In the current INAA-LFT system, starting with sampling, there are three subsequent processes, including reaction preparation, isothermal amplification, and product dilution and detection, which take a total of 20-60 minutes. In the future, if the detection time can be reduced to 8-10 minutes, all sample processing, reaction and detection can be completed in one device, and can be combined with artificial intelligence (AI) to achieve standardization (parts framed by yellow lines), it will open new avenues for improving the overall performance and applicability of this technology in diverse settings.
Figure 1.
Workflow of INAA-LFT now and its future trajectory in detection of plant virus. In the current INAA-LFT system, starting with sampling, there are three subsequent processes, including reaction preparation, isothermal amplification, and product dilution and detection, which take a total of 20-60 minutes. In the future, if the detection time can be reduced to 8-10 minutes, all sample processing, reaction and detection can be completed in one device, and can be combined with artificial intelligence (AI) to achieve standardization (parts framed by yellow lines), it will open new avenues for improving the overall performance and applicability of this technology in diverse settings.
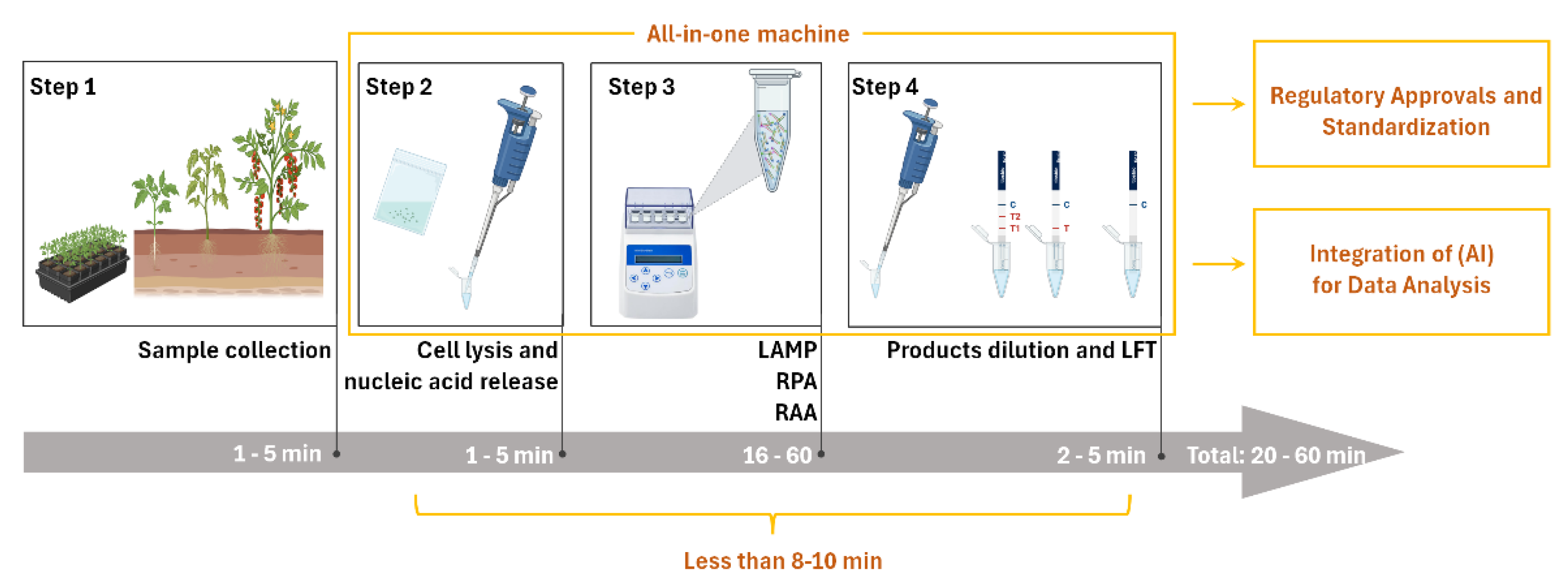

Table 1.
Plant viruses detected through INAA-LFT.
| Type of Amplification | Virus | Type of Viral Genome | Testing Duration | Ref |
|---|---|---|---|---|
| LAMP | cassava brown streak virus | +ssRNA | 40 min | [11] |
| ugandan cassava brown streak virus | +ssRNA | 40 min | [11] | |
| tobacco rattle virus | +ssRNA | <50 min | [12] | |
| Chinaberry tree badnavirus 1 | dsRNA | 45 min | [13] | |
| rice stripe virus | ±RNA | 50 min | [42] | |
| rice black-streaked dwarf virus | dsRNA | 50 min | [42] | |
| RPA | alfalfa mosaic virus | +ssRNA | 30 min | [6] |
| plum pox virus | +ssRNA | 20 min | [18] | |
| rice black-streaked dwarf virus | dsRNA | 20 min | [19] | |
| milk vetch dwarf virus | ssDNA | 30 min | [20] | |
| cymbidium mosaic virus | +ssRNA | 30 min | [21] | |
| barley yellow dwarf virus | +ssRNA | 20 min | [22] | |
| bean pod mottle virus | +ssRNA | <90 min | [23] | |
| tomato yellow leaf curl virus | ssDNA | 30 min | [24] | |
| piper yellow mottle virus | dsDNA | 30 min | [25] | |
| tomato chlorotic spot virus | ±RNA | 15 min | [27] | |
| actinidia chlorotic ringspot-associated virus | +ssRNA | <40 min | [28] | |
| citrus tristeza virus | +ssRNA | 15-20 min | [30] | |
| little cherry virus 2 | +ssRNA | - | [31] | |
| tomato spotted wilt virus | ±RNA | 15 min | [32] | |
| potato virus Y (PVY) | +ssRNA | 30 min | [33] | |
| potato virus S (PVS) | +ssRNA | 30 min | [33] | |
| potato leafroll virus (PLRV) | +ssRNA | 30 min | [33] | |
| tobacco mosaic virus | +ssRNA | 40 min | [41] | |
| tobacco etch virus | +ssRNA | 40 min | [41] | |
| potato virus X | +ssRNA | 40 min | [41] | |
| RAA | maize chlorotic mottle virus | +ssRNA | 45 min | [36,38] |
| sorghum mosaic virus | +ssRNA | 30 min | [37] | |
| rice stripe mosaic virus | -ssRNA | 30 min | [37] | |
| tomato brown rugose fruit virus | +ssRNA | 20 min | [39] | |
| pepper mild mottle virus | +ssRNA | <1 h | [40] | |
| tomato mosaic virus | +ssRNA | < 1 h | [40] | |
| tomato mottle mosaic virus | +ssRNA | < 1 h | [40] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



