
Submitted:
29 February 2024
Posted:
29 February 2024
You are already at the latest version
Abstract
A sedentary lifestyle associated with unregulated diets rich in high-calorie foods have contributed to the great prevalence of metabolic dysfunction-associated steatotic liver disease (MASLD) latterly, with up to 60% in high-risk population and 25% in the general population. The absence of specific pharmacological strategies for this syndrome represents one of the major problems in the management of MASLD patients. Lifestyle interventions and adherence to a healthy diet are the main cornerstones of current therapies. The identification of nutraceuticals useful in the treatment of MASLD appears to be one of the most promising strategies for the development of new effective and safe treatments for this disease. The onion, one of the most widely studied foods in the field of nutraceuticals, serves as an inexhaustible reservoir of potent compounds with various beneficial effects. The following preliminary study analyses, mediating in silico studies, the iteration of a library of typical onion compounds with 3-hydroxy-3-methylglutaryl-coenzyme A reductase, Liver receptors X α and β as well as Peroxisome proliferator-activated receptors α and γ. In this study fort the first time promising smart molecules from the onion that could have a beneficial action in MASLD patients were identified.
Keywords:
MASLD
; liver
; onion
; nutraceutics
; in silico research
; molecular docking.
1. Introduction
The term non-alcoholic fatty liver disease (NAFLD) was used in 1986 by Schaffner, who observed that the inflammatory state typical of non-alcoholic steatohepatitis (NASH) was not present in NAFLD [1]. NAFLD is identified as a syndrome characterized by the fat accumulation in the liver parenchyma and encompasses a broad spectrum of pathologies including simple fatty liver, non-alcoholic steatohepatitis, cirrhosis and hepatocellular carcinoma through progressive fibrosis [2,3]. Currently, it has been proposed that the definition of fatty liver disease associated with metabolic dysfunction (MASLD) may represent a more comprehensive terminology than the acronym NAFLD since it overarches the relationship between hepatic fat deposition and metabolic dysfunction. Up to now, no specific drugs have been identified for the treatment of MASLD patients. The management of MASLD patients involves changes in lifestyle and the consumption of a healthy diet [4,5]. The Mediterranean diet is recognized for both the decrease in the cardiovascular diseases risk and for the reduction of the metabolic syndrome risk and thus it has been efficient therapeutical approach in MASLD patients [6,7]. In this regard, the scientific community has made considerable efforts in identifying nutraceutical compounds useful in MASLD patients. Onion (Allium cepa) is believed to have originated in central Asia but it is farmed globally and eaten regardless of the cuisine and ethnic group. Cooked, fired or raw, the onion is irreplaceable ingredient of the Mediterranean diet. This perennial or biennial bulbous monocotyledon which belongs to the Liliaceae family is rich in various nutraceutical compounds, such as flavonoids, anthocyanins, organosulphur compounds, saponins and phenols. Numerous beneficial properties are attributed to the nutraceutical compounds in onions, including antioxidant, antitumour, anti-diabetic and anti-inflammatory activities [8]. Thus, the nutraceutical compounds from A. cepa may be observed as potential ligands for carefully selected target proteins that interfere with MASLD development and progression. Promising targets for the identification of new pharmacological strategies useful in MASLD include 3-hydroxy-3-methylglutaryl-coenzyme A reductase (HMGCR), peroxisome proliferator-activated receptors (PPARs) and Liver X receptors (LXRs). HMGCR is an enzyme involved in the biosynthesis of cholesterol. HMGCR catalyses the reduction reaction of (S)-HMG-CoA to (R)-mevalonate, using two molecules of NADPH [9]. Statins, hypolipidemic drugs are the main inhibitors of HMGCR. Currently, the usefulness of statins in MASLD patients has been demonstrated due to their ability to reduce liver enzyme concentrations and improve the liver histology of patients. Furthermore, it has been observed that statins reduce the risk of developing MASLD [10]. On contrary, the administration of statins is related to side effects such as the increase in liver enzymes as markers of inflammation. Peroxisome proliferator-activated receptors (PPARs) comprise three subtypes: PPARα, PPARδ and PPARɣ. The PPARα is involved in the processes of fatty acid absorption and oxidation and lipoprotein metabolism and it is mainly expressed in the liver, the kidney, the heart and the muscles. PPARδ is observed in most cells, while PPARγ is expressed in macrophages, large intestine and adipose tissue, where it is involved in adipocyte differentiation processes and lipid metabolism. Fibrates by activating PPARα, promote the formation of high-density lipoprotein (HDL) and reduce the concentration of triglycerides (TG) in the blood and thus present the drugs of choice in dyslipidaemia [11]. Given the therapeutic effects of fibrates, the representatives of the new generation, saroglitizar and lanifibranor, are in clinical trials for MASLD and seem to meliorate NAFLD and NASH [12]. Thiazolidinediones (TZDs), PPARγ agonists, are mainly used in the treatment of type II diabetes due to their insulin-sensitising action and ability to reduce plasma glucose levels. TZDs, and in particular pioglitazone, improve liver histology in NAFLD patients, although their application is limited due to the side effects. LXRs (α and β isoforms) are nuclear receptors involved in many metabolic processes, such as cholesterol homeostasis, liponeogenesis and the inflammatory response. The LXRα isoform is expressed in tissues with high metabolic activity (liver, small intestine, kidney), whereas LXRβ can be found ubiquitously [13]. Recent studies suggested that LXRs play a role as gatekeepers in NAFLD/NASH progression. Activation of LXRβ in stellate cells exerts anti-inflammatory and antifibrotic activities, preventing progression to fibrosis; while activation of LXRs in hepatocytes suppressed the transactivation of genes that promote NASH. LXRs are promising molecular targets for MASLD by virtue of the numerous cellular processes in which they are involved, but the complexity of their involvement in the disease requires further study. In this study, selected molecular targets were molecularly docked using a library of naturally occurring onion compounds in order to identify promising small molecules that could provide beneficial effects in MASLD patients.
2. Materials and Methods
The library of typical natural onion compounds used in the study was created by consulting FOODB and PubChem online databases [14,15] and the following compounds were selected: quercetin (2-(3,4-dihydroxyphenyl)-3,5,7-trihydroxychromen-4-one); kaempferol (3,5,7-trihydroxy-2-(4-hydroxyphenyl)chromen-4-one); galangin (3,5,7-trihydroxy-2-phenylchromen-4-one); baicalein (5,6,7-trihydroxy-2-phenylchromen-4-one); luteolin (2-(3,4-dihydroxyphenyl)-5,7-dihydroxychromen-4-one); myricetin (3,5,7-trihydroxy-2-(3,4,5-trihydroxyphenyl) chromen-4-one); isorhamnetin (3,5,7-trihydroxy-2-(4-hydroxy-3-methoxyphenyl)chromen-4-one) and (R)-2-(3,4-dihydroxybenzoyl)-2,4,6-trihydroxy-3(2H)-benzofuranone. In particular, the compound benzofuranone was found in “Rossa di Tropea” and “Ramata di Montoro” onion, two Italian onion varieties [16]. The 2D structures of the analyzed compounds were created using ChemDraw Professional 16.0 software (Figure 1) [17]. Optimization of the structures was carried out by applying the MMFF94 force field in Chem3D 16.0 software [18].
The X-ray crystallographic patterns of the selected targets were obtained from the Protein Data Bank (PDB): LXR-α (PDB: 1UHL); LXR-β (PBD: 1UPV); PPAR-α (PDB: 8HUK); PPAR-γ (PDB: 6D8K) and HMG-CoA reductase (PDB: 1HW8). The PDBs were optimised using the Wizard tool of the GOLD software. The optimisation enabled the addition of missing H-atoms and the removal of water molecules. The selected targets were re-docked using co-crystallised ligands. In particular, re-docking was performed with the antagonist mevastatin for HMGCR, and with the agonists T0901317, lanifibranor and GW1929 for LXR-α/β and PPARα/γ respectively. The GOLD Software’s Wizard tool enabled the identification of the binding pocket of the selected targets [19]. The chemscore kinase and slow algorithm represent the parameters applied in our analysis. The library of selected compounds was molecularly docked reporting the compounds with the best ChemPLP Fitness scores and the interactions were analysed with the academic version of Maestro, Schrödinger’s software [20].
3. Results
The PDBs used in the study were selected considering the completeness of the amino acid structure of the crystallographic model and the activity of the cocrystallised ligand. The protocol was validated by re-docking between the targets and their respective ligands. PDB 1HW8 represents the crystallographic model of HMGCR used in the study, in which the antagonist mevastatin constitutes the cocrystallised ligand. Box A in Figure 2 shows the redocking results for mevastatin, highlighting the formation of two hydrogen bonds with Glu559 and Asp690 and three salt bridges with Arg590, Lys692 and Lys735 in the catalytic site A of HMGCR. The results are in agreement with data reported in the literature that attribute a crucial role of these interactions in HMGCR inactivation [21]. Boxes B, C, D and E show the docking of benzofuranone, myricetin, quercetin and luteolin, respectively. Analyses of the molecular docking of benzofuranone (ChemPLP Fitness score 58.726) showed the formation of five hydrogen bonds with Glu559, Lys691, Ser665, Asn755, Ala751 and hydrophobic interactions with Arg590 and His752, respectively. Molecular docking of myricetin (ChemPLP Fitness score 58.348) and quercetin (ChemPLP Fitness score 57.4561) showed the formation of hydrogen bonds with Ala751, Arg590, Gly560, Cys561 and Lys735 and two hydrophobic character interactions with Arg590 and Lys691. Luteolin (ChemPLP Fitness score 54.989) showed the formation of hydrogen bonds with Glu675, Lys691, Glu559, Asn755,Ala751, Lys735 and a hydrophobic interaction with Arg590.
PDB 1UHL represents the crystallographic model of LXRα used in the study, in which the synthetic agonist T0901317 constitutes the cocrystallised ligand. Box A in Figure 3 shows the re-docking results for T0901317, highlighting the formation of a hydrogen bond with the His421 of LXRα. This interaction is in agreement with the data reported in the literature, which attribute to this bond a crucial role in LXRα activation [22]. Boxes B, C and D show the docking of myricetin (ChemPLP Fitness score 55.083), luteolin (ChemPLP Fitness score 53.643) and kaemferol (ChemPLP Fitness score 51.474), respectively. These compounds exhibit hydrogen bonding to H421, reproducing the interaction observed in T0901317; they also exhibit hydrogen bonding to Trp443.
The PDB 8HUK represents the crystallographic model of PPARα used in the study, in which the lanifibranor agonist constitutes the cocrystallised ligand. Box A in Figure 4 shows the redocking results for lanifibranor. By superimposition between the crystallographic model and the pose obtained by redocking, it was observed that the agonist retains the correct position. Boxes B and C show the docking of myricetin (PLP Fitness score 65.655) and galangin (PLP Fitness score 60.412), respectively. These compounds exhibit hydrogen bonding to Tyr464, reproducing the interaction reported in the literature responsible for PPARα activation [23]. In addition, both compounds form a hydrophobic interaction with His440.
Molecular docking results for LXRβ and PPARγ were not reported in the study, as none of the compounds in the considered library reproduced the crucial interactions for receptor activation.
4. Discussion
MASLD is the most prevalent liver disease reaching pandemic dimensions and the leading cause of liver-related morbidity and mortality [24]. The lack of specific drug therapy for MASLD together with the inability to achieve and access the clinical endpoints due to the slowly progression of the diseases is recognized as the main problem in the MASLD management. The ideal therapeutical approach to MASLD should also aim the metabolic risk factors that interfere with cardiovascular disease beside the mandatory inhibition of the liver fibrosis progression [25]. Healthy lifestyle and a balanced diet are the most effective strategies to improve patients’ conditions. In our work, we identified five molecular targets involved in MASLD (HMGCR, LXRα/β and PPARα/γ) that were subjected to molecular docking process, using a library of typical onion compounds. The docking poses obtained were analysed by searching for protein-ligand interactions reported in the literature, which correlated with the activation of LXRα/β and PPARα/γ and the inhibition of HMGCR. This molecular docking study has made a pioneer step in analyzing a library of typical A. cepa compounds as potential LXRα/β and PPARα/γ activators and HMGCR inhibitors.
HMGCR is an enzyme involved in cholesterol biosynthesis and is responsible for the key reaction in the biosynthetic process. HMGCR consists structurally of two portions: the N-terminal portion resides in the membrane of the endoplasmic reticulum, while the C-terminal part is responsible for catalytic activity and is immersed in the cytoplasm. These two sections are connected to each other via a linker region [26]. Specifically, the catalytic part is a homo-tetramer consisting of four identical amino acid chains. Each monomer consists of three domains: the N-domain is the smallest and forms the linker region with the portion in the endoplasmic reticulum; the L-domain is the largest and forms the binding site for HMG-CoA; while the S-domain forms the binding site for NADPH, which is necessary for catalytic activity. The four monomers unite to form dimers with two catalytic sites each. Statins are the main inhibitors of HMGCR. These drugs target the enzyme’s active sites and form a dense network of interactions with residues Arg590, Ser684, Asp690, Lys691, Lys692, Lys735, Asp755, Glu559 and Asp767 [27,28]. All analysed compounds expressed good affinity for the catalytic site of HMGCR by generating a network of interactions with the surrounding amino acid residues. In particular, best ChemPLP fitness scores and the highest number of interactions with the enzyme were observed for benzofuranone, myricetin, quercetin and luteolin. These data suggested a good stability of the compounds in the active site. Myricetin, quercetin, luteolin and benzofuranone reproduced the typical interactions of statins with Arg590 and Lys691 at the catalytic site of HMGCR. Thus, a promising inhibitory activity for benzofuranone, myricetin, quercetin and luteolin against HMGCR can be assumed. Several studies reported that quercetin, myricetin and luteolin reduced HMGCR gene expression [29]. On the contrary, quercetin had exbited cholesterolaemic activity by directly inhibiting HMGCR in animal model study [30]. LXRα is a nuclear receptor involved in the regulation of cholesterol and lipid metabolism, liponeogenesis and the inflammatory response. The structure of LXRα is characterised by five domains: the ligand-binding domain (LBD), the N-terminal activation domain, the DNA-binding domain (DBD); the hinge region; and the C-terminal domain. Specifically, the LBD consists of 10 α-helices and one β-sheet. Activation of the LXRα receptor is determined by the stabilisation of α-helix 12 in the active conformation to facilitate co-activator binding. Oxycholesterols, endogenous agonists of LXRα, interact with the LBD through the formation of two bonds with His421 and Trp443. In contrast, synthetic agonists, such as T0901317, activate the receptor through the formation of a hydrogen bond with His421 alone, resulting in the stabilisation of His421-Trp443 stacking [31]. Desirable chemical-structural affinity between the LBD of LXRα and myricetin, luteolin and kaempferol was reported in this study. All three compounds reproduced the binding mode of endogenous agonists, forming hydrogen bonds with both His421 and Trp443. In addition, each ligand was stabilised in the LBD by the formation of additional interactions that promote the stability of the receptor-ligand complex. Luteolin was already reported to improve hypercholesterolaemia and glucose intolerance through an up-regulation of LXRα in obese mice [32]. However, to the best of our knowledge there are no literature data on the direct myricetin, luteolin and kaempferol activity on LXRα. PPARα is a nuclear receptor involved in lipid metabolism in the liver. The structure of PPARα is characterised by five domains: activation function 1 (AF1) domains A and B, the DNA binding domain (DBD), the ligand binding domain (LBD) and the hinge region. Activation of PPARα involves the formation of a network of hydrogen bonds between the ligand and Tyr464, Tyr314 and Ser280 residues. These interactions stabilise the α-helix 12 facilitating coactivator binding. Fibrates are well known PPARα agonists that activate the receptor through interaction with residues Tyr464, Tyr314 and Ser280 [33]. Based on the results obtained in this study myricetin and galangin could be promising PPARα agonists, as they mimic some of the key interactions for receptor activation since both compounds form hydrogen bonds with Tyr464. Six of the compounds in the library (banzofuranone, myricetin, luteolin, quercetin, kaempferol and galangin) have promising activity for at least one of the selected targets. In particular, myricetin and luteolin appeared to be the most promising compounds from a multitarget perspective.
5. Conclusions
Eight typical onion compounds were molecularly docked onto five targets involved in MASLD in order to identify promising small molecules. Benzofuranone, myricetin, luteolin and quercetin exhibited the most promising antagonist activity for HMGCR; myricetin, luteolin and kaempferol might have favourable agonist activity towards LXRα; and finally, myricetin and galangin seem to have good agonist activity towards PPARα. In addition, myricetin and luteolin are reconized as potential multi-target-directed ligands in MASLD. Further studies will be needed to validate these hypotheses.
Author Contributions
Conceptualization, M.R.P. and A.C.P.; methodology, M.R.P. and. A.C.P.; software, M.M.; validation, G.G.M.S. and M.R.; formal analysis, M.R.P. and A.C.P.; investigation, M.R.P. and A.C.P.; resources, M.M.; data curation, N.M. (Nataša Milošević); writing—original draft preparation, M.R.P. and A.C.P.; writing—review and editing, N.M. (Nataša Milić); visualization, L.A.; supervision, L.A.; funding acquisition, N.M (Nataša Milić). All authors have read and agreed to the published version of the manuscript.
Data Availability Statement
For the analyses performed in this paper, the software used are ChemDraw Professional 16.0 software, GOLD Software and the academic version of Maestro, Schrödinger’s software. FOOBD and PubChem online databases were consulted for ligand searching. Finally, BioRender.com was used for the creation of the graphical abstract.
Acknowledgments
This work was supported by Provincial Secretariat for Higher Education and Scientific Research, AP Vojvodina, Republic of Serbia [Grant number 142-451-3509/2023-01].
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Schaffner, F.; Thaler, H. Nonalcoholic fatty liver disease. Prog Liver Dis 1986, 8, 283–298. [Google Scholar]
- Abenavoli, L.; Giubilei, L.; Procopio, A.C.; Spagnuolo, R.; Luzza, F.; Boccuto, L.; Scarpellini, E. Gut Microbiota in Non-Alcoholic Fatty Liver Disease Patients with Inflammatory Bowel Diseases: A Complex Interplay. Nutrients 2022, 14, 5323. [Google Scholar] [CrossRef]
- Calabrò, A.; Procopio, A.C.; Primerano, F.; Larussa, T.; Luzza, F.; Di Renzo, L.; De Lorenzo, A.; Gualtieri, P.; Abenavoli, L. Beneficial effects of coffee in non-alcoholic fatty liver disease: A narrative review. Hepatoma Res 2020, 6, 69. [Google Scholar] [CrossRef]
- Elsayed, A.; Ismaiel, A.; Procopio, A.C.; Luzza, F.; Abenavoli, L.; Dumitrascu, D.L. Noninvasive biochemical markers and surrogate scores in evaluating nonalcoholic steatohepatitis. Minerva Med. 2022, 113, 864–874. [Google Scholar] [CrossRef]
- Larussa, T.; Abenavoli, L.; Procopio, A.C.; Iannelli, C.; Polimeni, N.; Spagnuolo, R.; Doldo, P.; Luzza, F. The role of gluten-free diet in nonalcoholic fatty liver disease development. Eur Rev Med Pharmacol Sci. 2021, 25, 6613–6618. [Google Scholar] [CrossRef] [PubMed]
- Abenavoli, L.; Procopio, A.C.; Paravati, M.R.; Costa, G.; Milić, N.; Alcaro, S.; Luzza, F. Mediterranean Diet: The Beneficial Effects of Lycopene in Non-Alcoholic Fatty Liver Disease. J Clin Med. 2022, 11, 3477. [Google Scholar] [CrossRef]
- Abenavoli, L.; Maurizi, V.; Boccuto, L.; Di Berardino, A.; Giostra, N.; Santori, P.; Scarcella, M.L.; Procopio, A.C.; Rasetti, C.; Scarpellini, E. Nutritional Support in Acute Liver Failure. Diseases 2022, 10, 108. [Google Scholar] [CrossRef] [PubMed]
- Kianian, F.; Marefati, N.; Boskabady, M.; Ghasemi, S.Z.; Boskabady, M.H. Pharmacological Properties of Allium cepa, Preclinical and Clinical Evidences; A Review. Iran J Pharm Res. 2021, 20, 107–134. [Google Scholar] [CrossRef]
- Istvan, E.S. Structural mechanism for statin inhibition of 3-hydroxy-3-methylglutaryl coenzyme A reductase. American heart journal 2002, 144, S27–S32. [Google Scholar] [CrossRef] [PubMed]
- Fatima, K.; Moeed, A.; Waqar, E.; Atif, A.R.; Kamran, A.; Rizvi, H.; Suri, N.F.; Haider, H.; Shuja, S.H.; Khalid, M.; Minhas, A.M.K. Efficacy of statins in treatment and development of non-alcoholic fatty liver disease and steatohepatitis: A systematic review and meta-analysis. Clin Res Hepatol Gastroenterol. 2022, 46, 101816. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.H.; Kim, S.G. Fibrates Revisited: Potential Role in Cardiovascular Risk Reduction. Diabetes Metab J. 2020, 44, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; You, H.; Qiu, S.; Yu, D.; Bai, Y.; He, J.; Cao, H.; Che, Q.; Guo, J.; Su, Z. A new perspective on NAFLD: Focusing on the crosstalk between peroxisome proliferator-activated receptor alpha (PPARα) and farnesoid X receptor (FXR). Biomed Pharmacother. 2022, 154, 113577. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Park, C.; Kim, TH. Targeting Liver X Receptors for the Treatment of Non-Alcoholic Fatty Liver Disease. Cells 2023, 12, 1292. [Google Scholar] [CrossRef] [PubMed]
- The Metabolomics Innovation Centre: FooDB (Version 1). Computer software, Canada: The Metabolomics Innovation Centre. 2017. Available online: http://foodb.ca/ (accessed on 6 November 2023).
- Kim, S.; Thiessen, P.A.; Bolton, E.E.; Chen, J.; Fu, G.; Gindulyte, A.; Han, L.; He, J.; He, S.; Shoemaker, B.A.; Wang, J.; Yu, B.; Zhang, J.; Bryant, S.H. PubChem Substance and Compound databases. Nucleic Acids Res. 2016, 44, D1202-13. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Barbhai, M.D.; Hasan, M.; Punia, S.; Dhumal, S.; Radha; Rais, N.; Chandran, D.; Pandiselvam, R.; Kothakota, A.; Tomar, M.; Satankar, V.; Senapathy, M.; Anitha, T.; Dey, A.; Sayed, A.A.S.; Gadallah, F.M.; Amarowicz, R.; Mekhemar, M. Onion (Allium cepa L.) peels: A review on bioactive compounds and biomedical activities. Biomed Pharmacother 2022, 146, 112498. [Google Scholar] [CrossRef] [PubMed]
- ChemDraw Professional. ChemDraw Professional, Version 16 0.0.82 (68); Perkin Elmer Informatics, Inc.: Cambridge, MA, USA, 2019. [Google Scholar]
- Halgren, T.A. Merck molecular force field. I. Basis, form, scope, parameterization, and performance of MMFF94. Journal of computational chemistry 1996, 17, 490–519. [Google Scholar] [CrossRef]
- Jones, G.; Willett, P.; Glen, R.C.; Leach, A.R.; Taylor, R. Development and validation of a genetic algorithm for flexible docking. J Mol Biol. 1997, 267, 727–748. [Google Scholar] [CrossRef]
- Free Maestro Academic Version v13.4 Package; Schrödinger Release 2022-4; Maestro, Schrödinger, LLC: New York, NY, USA, 2022.
- da Costa, R.F.; Freire, V.N.; Bezerra, E.M.; Cavada, B.S.; Caetano, E.W.; de Lima Filho, J.L.; Albuquerque, E.L. Explaining statin inhibition effectiveness of HMG-CoA reductase by quantum biochemistry computations. Physical Chemistry Chemical Physics 2012, 14, 1389–1398. [Google Scholar] [CrossRef]
- Svensson, S.; Ostberg, T.; Jacobsson, M.; Norström, C.; Stefansson, K.; Hallén, D.; Johansson, I.C.; Zachrisson, K.; Ogg, D.; Jendeberg, L. Crystal structure of the heterodimeric complex of LXRalpha and RXRbeta ligand-binding domains in a fully agonistic conformation. EMBO J. 2003, 22, 4625–4633. [Google Scholar] [CrossRef]
- Kamata, S.; Honda, A.; Ishikawa, R.; Akahane, M.; Fujita, A.; Kaneko, C.; Miyawaki, S.; Habu, Y.; Shiiyama, Y.; Uchii, K.; Machida, Y.; Oyama, T.; Ishii, I. Functional and Structural Insights into the Human PPARα/δ/γ Targeting Preferences of Anti-NASH Investigational Drugs, Lanifibranor, Seladelpar, and Elafibranor. Antioxidants 2023, 12, 1523. [Google Scholar] [CrossRef]
- Riazi, K.; Azhari, H.; Charette, J.H.; Underwood, F.E.; King, J.A.; Afshar, E.E.; Swain, M.G.; Congly, S.E.; Kaplan, G.G.; Shaheen, A.A. The prevalence and incidence of NAFLD worldwide: A systematic review and meta-analysis. Lancet Gastroenterol Hepatol. 2022, 7, 851–861. [Google Scholar] [CrossRef] [PubMed]
- Machado, M.V. MASLD treatment-a shift in the paradigm is imminent. Front Med 2023, 10, 1316284. [Google Scholar] [CrossRef] [PubMed]
- Istvan, E.S.; Deisenhofer, J. The structure of the catalytic portion of human HMG-CoA reductase. Biochim Biophys Acta. 2000, 1529, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Istvan, E.S.; Deisenhofer, J. Structural mechanism for statin inhibition of HMG-CoA reductase. Science 2001, 292, 1160–1164. [Google Scholar] [CrossRef] [PubMed]
- da Silva, V.B.; Taft, C.A.; Silva, C.H. Use of virtual screening, flexible docking, and molecular interaction fields to design novel HMG-CoA reductase inhibitors for the treatment of hypercholesterolemia. J Phys Chem A. 2008, 112, 2007–2011. [Google Scholar] [CrossRef] [PubMed]
- Waiz, M.; Alvi, S.S.; Khan, M.S. Potential dual inhibitors of PCSK-9 and HMG-R from natural sources in cardiovascular risk management. EXCLI J. 2022, 21, 47–76. [Google Scholar] [CrossRef] [PubMed]
- Khamis, A.A.; Salama, A.F.; Kenawy, M.E.; Mohamed, T.M. Regulation of hepatic hydroxy methyl glutarate—CoA reductase for controlling hypercholesterolemia in rats. Biomed Pharmacother. 2017, 95, 1242–1250. [Google Scholar] [CrossRef] [PubMed]
- Tice, C.M.; Noto, P.B.; Fan, K.Y.; Zhuang, L.; Lala, D.S.; Singh, S.B. The medicinal chemistry of liver X receptor (LXR) modulators. J Med Chem. 2014, 57, 7182–7205. [Google Scholar] [CrossRef]
- Park, H.S.; Lee, K.; Kim, S.H.; Hong, M.J.; Jeong, N.J.; Kim, M.S. Luteolin improves hypercholesterolemia and glucose intolerance through LXRα-dependent pathway in diet-induced obese mice. J Food Biochem 2020, 44, e13358. [Google Scholar] [CrossRef]
- Dhoke, G.V.; Gangwal, R.P.; Sangamwar, A.T. A combined ligand and structure based approach to design potent PPAR-alpha agonists. Journal of Molecular Structure 2012, 1028, 22–30. [Google Scholar] [CrossRef]
Figure 1.
2D structures of the ligands analyzed in this study.
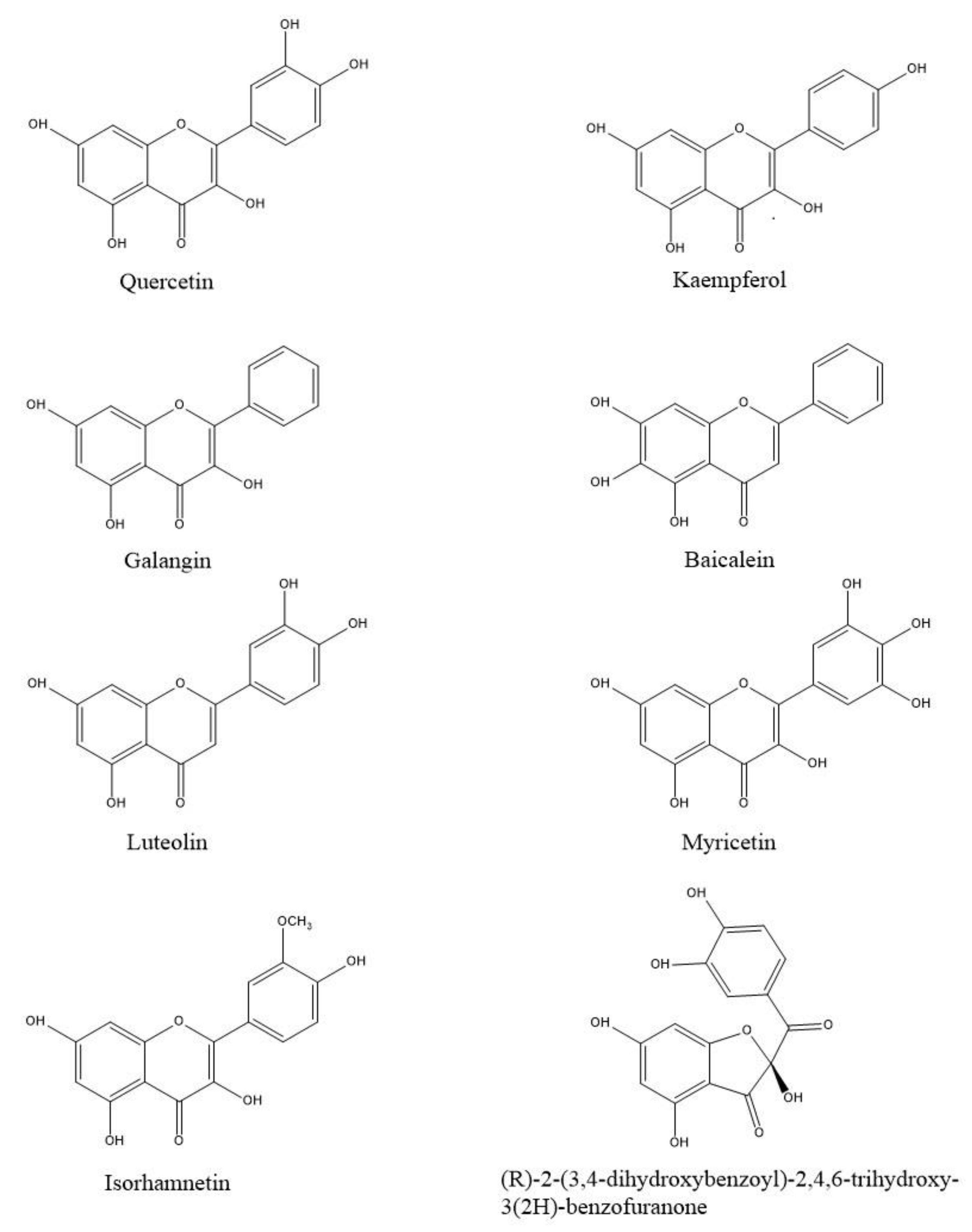
Figure 2.
Molecular docking analysis. 3D representation of HGMCR complexed with mevastatin (a); benzofuranone (b), myricetin (c), quercetin (d) and luteolin (e) docked in the active site of HMGCR, respectively. HMGCR is shown as cartoon purple (Chain A) and cyan (Chain B). Mevastatin, benzofuranone, myricetin, quercetin and luteolin are shown as light green, blue, magenta, dark green and light blue carbon sticks, respectively. The amino acid residues involved in the interactions are shown as carbon sticks purple (Chain A) and cyan (Chain B) respectively. Hydrogen bonds, π-π, cation-π interactions and salt bridges are shown as dashed lines, yellow, light blue, green and magenta respectively.
Figure 2.
Molecular docking analysis. 3D representation of HGMCR complexed with mevastatin (a); benzofuranone (b), myricetin (c), quercetin (d) and luteolin (e) docked in the active site of HMGCR, respectively. HMGCR is shown as cartoon purple (Chain A) and cyan (Chain B). Mevastatin, benzofuranone, myricetin, quercetin and luteolin are shown as light green, blue, magenta, dark green and light blue carbon sticks, respectively. The amino acid residues involved in the interactions are shown as carbon sticks purple (Chain A) and cyan (Chain B) respectively. Hydrogen bonds, π-π, cation-π interactions and salt bridges are shown as dashed lines, yellow, light blue, green and magenta respectively.
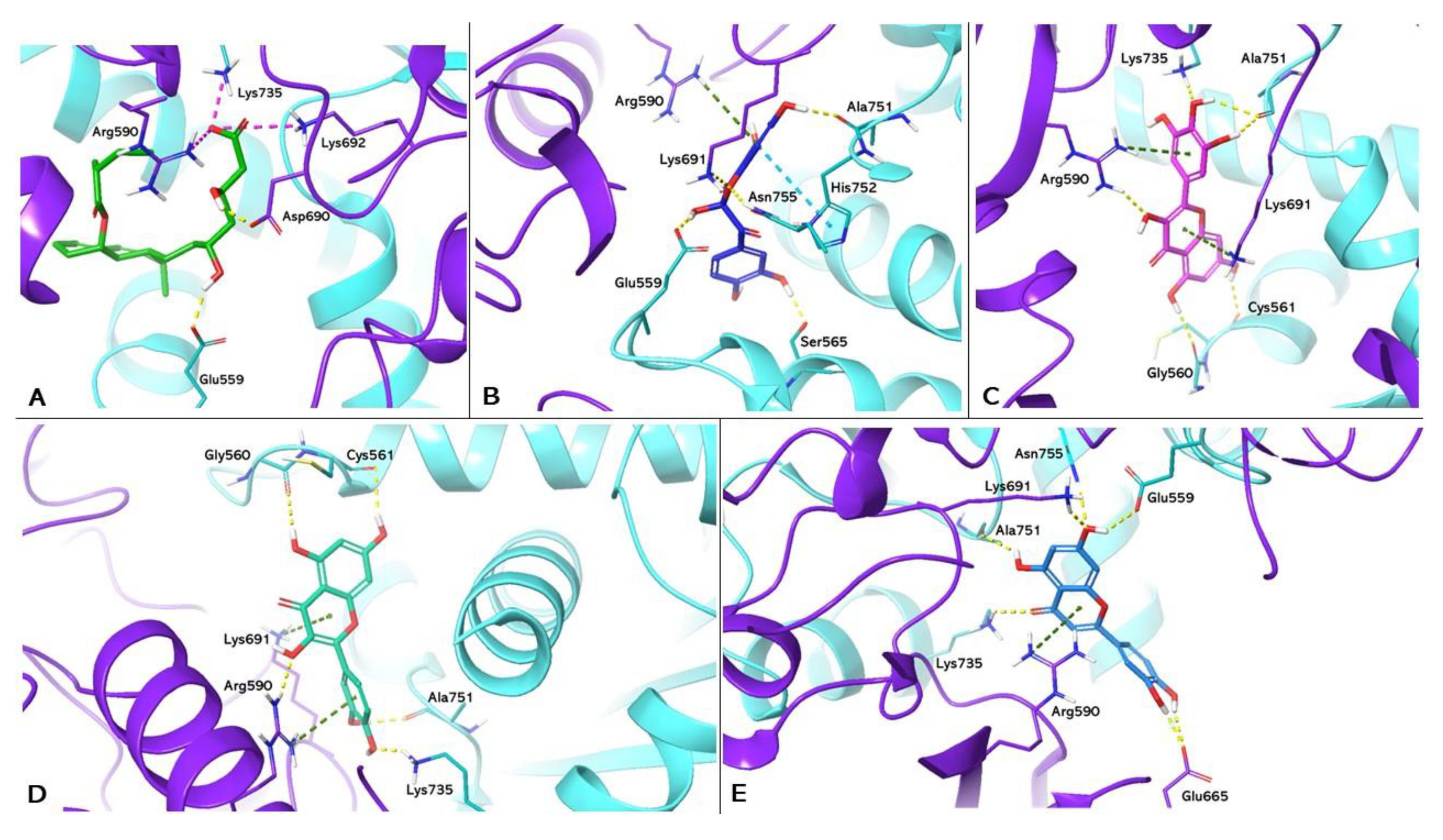
Figure 3.
Molecular docking analysis. 3D representation of LXRα complexed with T0901317 (a); myricetin (b), luteolin (c) and kaempferol (d) docked in the LBD of LXRα, respectively. HMGCR is shown as cyan cartoon; T0901317, myricetin, luteolin and kaempferol are represented as green, magenta, blue and red carbon sticks, respectively. The amino acid residues involved in the interactions are shown as cyan carbon sticks. Hydrogen bonds and π-π interactions are shown as dashed lines, yellow and light blue respectively.
Figure 3.
Molecular docking analysis. 3D representation of LXRα complexed with T0901317 (a); myricetin (b), luteolin (c) and kaempferol (d) docked in the LBD of LXRα, respectively. HMGCR is shown as cyan cartoon; T0901317, myricetin, luteolin and kaempferol are represented as green, magenta, blue and red carbon sticks, respectively. The amino acid residues involved in the interactions are shown as cyan carbon sticks. Hydrogen bonds and π-π interactions are shown as dashed lines, yellow and light blue respectively.
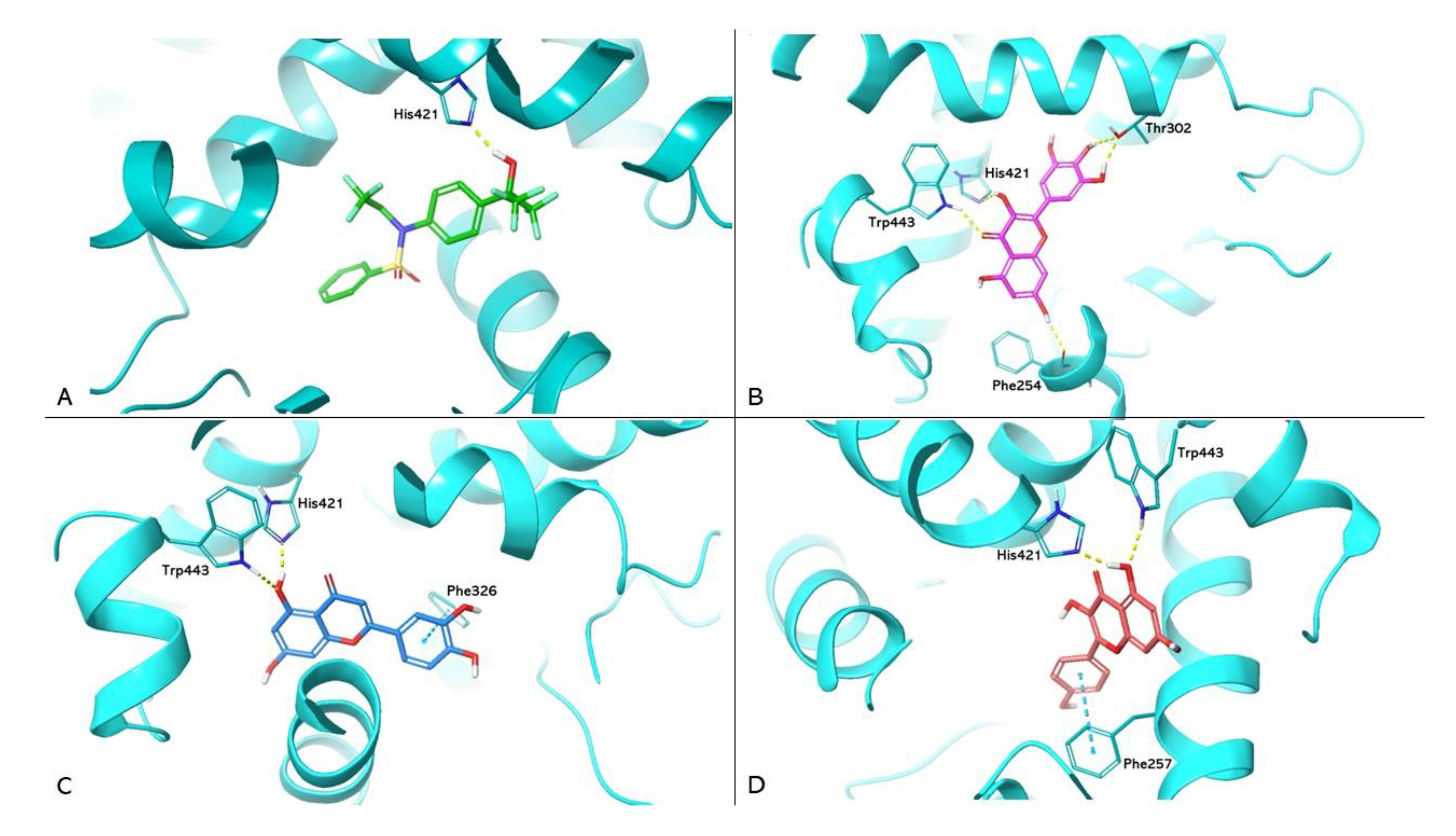
Figure 4.
Molecular docking analysis. (a) Superimposition of the lanifibranor complexed with PPARα of the crystallographic model and the redocking pose. PPARα is shown as magenta cartoons for the crystallographic model and cyan for the redocking pose, respectively. Lanifibranor is shown as orange carbon sticks for the crystallographic model and green for the redocking pose, respectively. (b, c) 3D representation of PPARα complexed with myricetin (magenta carbon sticks) and galangin (yellow carbon sticks) docked in the LBD of PPARα, respectively. PPARα is shown as cyan cartoon and the amino acid residues involved in the interactions are shown as cyan carbon sticks. Hydrogen bonds and π-π, cation-π interactions are shown as dashed lines, yellow, blue and green respectively.
Figure 4.
Molecular docking analysis. (a) Superimposition of the lanifibranor complexed with PPARα of the crystallographic model and the redocking pose. PPARα is shown as magenta cartoons for the crystallographic model and cyan for the redocking pose, respectively. Lanifibranor is shown as orange carbon sticks for the crystallographic model and green for the redocking pose, respectively. (b, c) 3D representation of PPARα complexed with myricetin (magenta carbon sticks) and galangin (yellow carbon sticks) docked in the LBD of PPARα, respectively. PPARα is shown as cyan cartoon and the amino acid residues involved in the interactions are shown as cyan carbon sticks. Hydrogen bonds and π-π, cation-π interactions are shown as dashed lines, yellow, blue and green respectively.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



