
Submitted:
29 February 2024
Posted:
29 February 2024
You are already at the latest version
Abstract
Pancreatic cancer (PC) is considered one of the most challenging and formidable malig-nancies to treat because it is usually diagnosed at terminal stage, thus it is characterized with a poor prognosis with 5% of patients living over 5 years. The late diagnosis leads to a lower suc-cess rate of resectability, hence chemotherapy or chemoradiotherapy are the only hopes for most PC patients. However, the available drug combinations for PC treatment did not provide satisfactory clinical outcomes. There is a pressing and urgent need to discover new chemothera-peutics which fight PC with tolerable side effects and less liability for resistance. Pentacyclic triterpenes (PTs) are privileged structures with a multitude of biological activities mitigating several ailments, particularly in neoplasm field. They are characterized by a high therapeutic window and their activity can be highly enhanced through structural modifications. Indeed, several parent PTs such as oleanolic acid, ursolic acid, lupeol, betulinic acid, and celastrol proved effective against different PC phenotypes inhibiting their proliferation, invasion, and metastasis mainly via KRAS modulation. Some PTs worked as adjuvant chemosensitizers in gemcitabine-resistant or TRAIL-resistant cells. Owing to their potential in PC treatment, we pre-sent this review article summarizing PC pathogenesis and the promising role of PTs in the man-agement of PC when used either solely or in combination with other chemotherapies such as gemcitabine and 5-Fluorouracil.
Keywords:
Pancreatic cancer
; natural products
; pentacyclic triterpenes
1. Introduction
Pancreatic ductal adenocarcinoma (PC) represents a formidable and an exceptionally devastating ailment which is ranked as the third leading cause of cancer-related death in the United States of America. PC is characterized with low survival rate compared to other cancer types (only 12%) with over 60000 new cases each year [1,2]. Several studies and meta-analysis showed that cigarette smoking, high alcohol intake, obesity, and diabetes are potential risk factors that can be associated to PC in different ratios [3,4,5].
Activating mutations of Kirsten rat sarcoma viral oncogene homolog (KRAS) were identified in around 91% of PC patients and about 30% of all cancers [6,7,8,9,10]. KRAS is one isoform of the RAS GTPase proteins in addition to HRAS, and NRAS. Further deletion or inactivating mutations of tumor-suppressor genes TP53 or SMAD4 maintain the progress of PC [10,11,12]. In the cell membrane, the inactive guanosine diphosphate (GDP)-bound KRAS is activated by guanine nucleotide exchange factor (GEF) proteins affording guanosine triphosphate (GTP)-bound KRAS through protein tyrosine kinase receptor such as epidermal growth factor receptor (EGFR) [13]. Aberrant activation of KRAS leads to inappropriate downstream signaling of different pathways implicated in cell proliferation including activation of downstream signaling molecules including RAF, Phosphoinositide 3-kinase (PI3K), and RAL [14,15].
In Ras-RAF-MEK-ERK pathway, the activated Ras recruits RAF, triggering a downstream signaling pathway activating MEK1/2 and extracellular signal-regulated kinases 1/2 (ERK1/2), consequently, which in turn, move into the nucleus and phosphorylates nuclear transcription factors implicated in cell proliferation [14]. PI3K is the second Ras effector known as Ras-PI3K-AKT-mTOR pathway [13]. Its activation transforms phosphatidylinositol (4,5)-biphosphate (PIP2) to phosphatidylinositol (3,4,5)-triphosphate (PIP3). The latter binds Akt which in turn triggers downstream signaling promoting Mammalian target of rapamycin (mTOR) which regulates gene transcription and orchestrate cell proliferation, survival, growth, and immune cell differentiation [16].
The third Ras effector is Ral-GTPases protein [17]. Indeed, GTP-bound Ral-A and Ral-B were found to be highly expressed in PC [18,19]. They activate NF-kB through TANK-binding kinase 1 (TBK1) and thus promote subsequent gene transcription, cytokine production, and contributing to oncogenesis [20,21] (Figure 1).
Depending on the severity of each case and the resectability, multidisciplinary management of PC is recommended [22]. In addition to resection and radiation, chemotherapy possesses a pivotal role in increasing the life expectancy of PC patients [6]. Nonetheless, only 15%–20% of newly-diagnosed patients are eligible for resection due to metastasis. About 75% of patients will suffer from recurrence within 2 years after resection [23]. FOLFIRINOX, a chemotherapy combination of fluorouracil, irinotecan, leucovorin, and oxaliplatin is a standard first-line therapy [6,24]. The second combination encompasses gemcitabine and paclitaxel. The currently available chemotherapy is modestly effective, encountering resistance of this recalcitrant disease and causing sever undesirable effects. Therefore, there is a continuous quest for new PC drugs to address the issues of present treatments.
Natural products presented privileged skeletons and structural basis to develop modern drugs [25,26]. Several anticancer drugs are basically natural products or semisynthetic compounds [27,28]. For example, Vincristine is a natural vinca alkaloid that is used for the treatment of leukemia, lymphoma, neuroblastoma and other cancers whereas its congener vinblastine is particularly beneficial in the treatment of Hodgkin's lymphoma [29]. Etoposide is a semisynthetic epipodophyllotoxin and the frontline chemotherapeutic drug in treatment of various cancers including non-Hodgkin lymphoma, glioblastoma, and nonlymphocytic leukemia [30]. Furthermore, bleomycin and actinomycin D are complex structures derived from bacterial source and widely used antibiotic anticancer agents [31,32]. Concomitantly, several marine derived products are used in the clinic for cancer management [33].
As we mentioned above, irinotecan is a member of FOLFIRINOX combination for PC therapy. Notably, irinotecan is a semisynthetic analogue of the pentacyclic alkaloid camptothecin that works by inhibition of topoisomerase I. Paclitaxel is a member of the second combination for PC treatment alongside with gemcitabine. The former is one of the most widely employed antineoplastic drugs of natural origin [34].
Several natural products proved effective against PC by modulating different signaling pathways [35,36,37]. For example curcumin has a synergistic effect when administered with gemcitabine [38,39]. Resveratrol induced apoptosis and autophagy in PC by upregulating the apoptotic marker Bax and inhibition of nutrient-deprivation autophagy factor-1 (NAF-1), respectively [40,41,42].
Pentacyclic triterpenes (PTs) are hydrophobic phytochemicals existing in plants and endowed with a vast array of biological activities including, anti-inflammatory [43,44], anti-atherosclerotic [45,46], antihypertensive [45,47], hepatoprotective [48,49,50], anti-viral [51,52], anti-fibrosis [53,54], and anti-ulcer effects [55]. PTs and their semisynthetic derivatives are particularly promising anticancer agents with a favorable safety profile [56,57,58,59].
In the framework of our interest in cancer chemotherapy and the tremendous biological importance of natural products especially triterpenes, we present the first review article to exhaustively explore the cytotoxic effect of PTs on different PC cells.
2. Methods
We collected the English language articles that relates triterpenes to pancreatic cancer keywords from Web of Science and PubMed database. The search resulted in around 54 articles; we excluded 3 retracted research papers of them.
3. Results
MK615 is the Japanese apricot extract which is a mixture of several triterpenes from including mainly oleanolic acid (OA) and ursolic acid (UA) [60]. The mixture exhibited antiproliferative effect in vitro and in vivo against MIAPaCa-2 PC cells in a dose-dependent manner, accompanied by an insignificant effect on human umbilical vein endothelial Cells (HUVEC) reflecting its safety. The anti-proliferative effect was observed in presence or absence of gemcitabine and ascribed to massive accumulation of ROS [61].
Triterpene mixture from the medicinal mushroom Poria cocos repressed the growth of different PC cells after 48 hours incubation; PANC-1 (IC50 = 24.5 μg/ml), MiaPaca-2 (IC50 = 23.0 μg/ml), AsPc-1 (IC50 = 11.3 μg/ml) and BxPc-3 cells (IC50 = 1.0 μg/ml). The titled extract triggered significant cell cycle arrest at G0/G1 phase and mitigated invasiveness of BxPc-3 cells via matrix metalloproteinase-9 (MMP-7) downregulation [62]. The same research group revealed the suppression of cell division cycle 20 (CDC20) by the same extract in PANC-1 cells dose-dependently [63]. Having said that, we will focus in this review on the activity of pure PTs rather than extracts. The below-mentioned bioactive PTs were classified according to their respective chemical type.
3.1. Oleanane type PTs in pancreatic cancer
Oleanolic acid (OA) (Figure 2), found in olive, jujube, ginseng, and Hawthorn berries, is the most widely studied PT in terms of structural modifications with multifaceted bioactivities [64]. OA cytotoxic effect on BxPC3 PC cells is quite strong with IC50 value 190 nM [65]. Nevertheless, IC50 of the parent β-amyrin (Figure 2) is 0.42 µM highlighting the pivotal role of -COOH group in activity [65].
Wei et al., studied the synergism between OA and 5-Fluorouracil. They found that OA potentiated 5-Fluorouracil apoptotic cell death in PANC-28 cells via increasing caspase-3 expression. The combination significantly upregulated Bax while decreasing B-cell lymphoma-2 (Bcl-2) and survivin [66]. Another synergistic effect was observed for OA/doxorubicin combination on PANC-1 cells. Indeed, the apoptotic antiproliferative potency of OA/doxorubicin combination was remarkably higher than that of each drug alone as assessed by MTT assay. The combination delayed wound closure time in the cellular wound healing assay. Concomitantly, the titled combination upregulated mRNA expression of proapoptotic genes including caspases, Bax, and p53 and downregulated the expression of Bcl-2 and survivin genes [67].
The widely known synthetic OA derivative Methyl-2-cyano-3,12-dioxooleana-1,9(11)-dien-28-oate (CDDO-Me) (Figure 2) induced apoptosis in MiaPaCa-2 and PANC-1 PC cells excessive generation of ROS including hydrogen peroxide and superoxide anions. The ROS release resulted in of hTERT telomerase inhibition. The antiproliferative and apoptotic effect of CDDO-Me were reversed by cotreatment with the antioxidant N-acetylcysteine [68].
Another synthetic OA derivative, K73-03 (Figure 2), exhibited remarkable autophagic and apoptotic antiproliferative effect on AsPC-1 cells via serine peptidase inhibitor, Kazal type 1 (SPINK1) pathway. The latter is overexpressed in different neoplasms including PC promoting proliferation, aggressiveness and chemoresistance [69]. MTT and wound healing assay revealed that K73-03 reduced AsPC-1 cell viability, size, and number of colonies in a dose-dependent manner. Indeed, K73-03 upregulated Bax, cleaved caspase-3, cleaved PARP and downregulated Bcl-2 concentration. Further mechanistic studies showed that K73-03 treatment upregulated microRNA-421 (miR-421) level and suppressed SPINK1 signaling pathway. The results were consistent in AsPC-1 cells xenografts in nude mice model [70].
The same research group indicated to the crucial role of epigenetic regulation of miR-421/SPINK1 in overcoming gemcitabine resistance. Basically, gemcitabine combined with K73-03 showed increased mitochondrial damage and apoptotic antiproliferative effect on AsPC-1 and MIA PaCa-2 cells higher than either of each drug alone. K73-03 significantly enhanced gemcitabine effect and suppressed the wound closure in gemcitabine-resistant AsPC-1 and MIA PaCa-2 cells. Noticeably, miR-421 level is lower, and SPINK1 level is higher in gemcitabine-resistant PC cells than in sensitive PC cells. The cytotoxic effect was attributed to remarkable synergistic increase in miR-421 and decrease in SPINK1 by K73-03/gemcitabine combination [71]. Additionally, another study indicated that K73-03 is a potent EGFR inhibitor as assessed by reverse pharmacophore screening and molecular dynamics simulations using EGFR crystal structure from protein data bank (PDB: 3W2S). The titled compound caused G2/M-phase arrest and suppressed ASPC-1 cells migration. Immunoblot assay exposed that K73-03 repressed the phosphorylation EGFR and Akt leading to a remarkable in vitro and in vivo anti-PC effect [72].
Maslinic acid (MA) (Figure 2), another representative of oleanane type PTs, was found to potentiate the apoptotic antiproliferative effect of tumor necrosis factor alpha (TNFα) on PC cells PANC-28. This synergism was achieved as MA enhanced TNFα-induced accumulation of caspase and cleaved PARP. The most crucial effect is that MA inhibits TNFα-induced NF-κB activation which is the main side effect of TNFα treatment. Mechanistically, MA suppressed tumor necrosis factor α (TNFα)-induced IκB degradation, p65 phosphorylation and nuclear translocation in PANC-28 cells. Consequently, MA downregulated the expression of anti-apoptosis genes, including Survivin, Bcl-2, Bcl-xl, and IAP-1, proliferation genes, including cyclin D1 and c-Myc, and metastasis/angiogenesis related genes such as vascular endothelial growth factor (VEGF), and matrix metalloproteinase-9 (MMP-9). In mouse xenograft model implanted with pancreatic cancer cells, MA reproduced the in vitro outcome by reducing tumor size and weight in a dose-dependent manner [73].
A similar outcome was obtained upon treatment of PANC-1 and other caner phenotypes with MA as reported by Lin et al. In the titled cells, MA treatment increased DNA fragmentation and upregulated the apoptotic proteins cleaved caspases, and Bax, but without significant reduction of Bcl-2 expression. At 8 and 16 µM, MA reduced PANC-1 invasion and migration by suppressing VEGF and transforming growth factor-β1 (TGF-β1) levels [74].
To improve its physicochemical properties by enhancing its solubility and bio-accessibility, Garridoa et al., formulated MA in solid lipid nanoparticles using diverse shell compositions. They tested the new formulation on PC cells BxPC3 growth. Indeed, the new formulation exhibited improved in vitro digestion, cell permeability, and crossing the gut barrier to reach cancer cells. The released MA in the basolateral compartment from models of in vitro gut barrier were sufficient to reduce BxPC3 viability [75]. Primarily, the formulation showed insignificant cytotoxicity on fibroblasts reflecting a high therapeutic index. The same research group validated their results in vivo using a xenograft mice model implanted with BxPC3 and breast cancer cells MCF-7. The used solid lipid nanoparticles were no toxic in mice either by oral or intravenous administration [76].
3.2. Ursane PTs in pancreatic cancer
Ursolic acid (UA) (Figure 3) is a common representative of the ursane type PT found in rosemary, apple, and grape skin. Li et al. found that UA has the potential to circumvent gemcitabine resistance issue. UA reduced growth and induced apoptosis in three different types of gemcitabine-resistant PC cells viz., PANC-1, MIA PaCa-2, and Capan-1 in a dose-dependent manner. The apoptosis involved increased concentrations of caspases 3/7/8/9 and activation of JNK activity. Furthermore, UA treatment suppressed total PI3K and the phosphorylated Akt levels in MIA PaCa-2 cells dose-dependently. These data were validated in vivo where administration of UA resulted in tumor regression in MIA PaCa-2 xenograft model without general toxicity [77].
Another research group found that UA apoptotic cell death mechanism in gemcitabine-resistant MIA PaCa-2, AsPC-1, and PANC-28 cells is attributed to NF-κB suppression and signal transducer and activator of transcription 3 (STAT3) activation. In mice model bearing PANC-28 cells, UA oral administration reduced tumor volume and weight, and enhanced gemcitabine effect. Surprisingly, UA/gemcitabine combination impeded PC metastasis to liver and spleen tissues via reducing Ki-67 and CD31 markers. Additionally UA treatment reduced tumor markers for cell survival (survivin) as well as Bcl-2, cell proliferation (cyclin D1), metastasis (ICAM-1 and MMP-9), and inflammation (COX-2) as assessed by immunohistochemical analysis of pancreatic tumor tissues [78].
The receptor for advanced glycation end products (RAGE) is a multiligand transmembrane receptor in the immunoglobulin (Ig) family and a key player in promoting PC and other several cancers [79,80,81]. Additionally, RAGE induces NF-kB response and several reports have showcased the presence of a positive feedback loop between RAGE and NF-kB [82,83]. In brief, stimulation of RAGE enhances angiogenesis and cancer development by upregulating PKI3/mTOR/Akt, VEGF, MMPs, MAPKs, NF-κB, downregulating p53 signaling. RAGE silencing greatly decreased IkB-α, and NF-kB/p65 protein phosphorylation and consequently mitigated multidrug resistance protein 1 (MDR1) protein expression. UA treatment suppressed RAGE, NF-kB/p65, and MDR1 protein levels in gemcitabine-resistant MIA PaCa-2 PC cells dose dependently. Conclusively, UA treatment was as efficient as RAGE silencing in abolishing MDR1 protein expression upon gemcitabine treatment environment. The chemo-sensitization effect of UA was reproduced and confirmed in mice bearing gemcitabine-resistant MIA PaCa-2 cells [82].
Ursolic acid lactone analogue, ursenolide (Figure 3), from Callistemon citrinus proved apoptotic antiproliferative against PANC-1 cells under nutrients deprived conditions (NDM). This is accompanied by less ability to metastasis as assessed by wound healing assay. Mechanistically, ursenolide suppressed the phosphorylation of Akt and mTOR under NDM [84].
3.3. Lupane type PTs in pancreatic cancer
Lupeol (Figure 4), a representative of lupane PTs class, induced apoptosis and reduced viability of AsPC-1 PC cells in a dose-dependent manner as tested by MTT and apoptosis assays. Lupeol impeded Ras-RAF-MEK-ERK and Ras-PI3K-AKT-mTOR pathways and reduced NF-kB/p65 phosphorylation in the titled cells. Concomitantly, lupeol induced poly (ADP-ribose) polymerase PARP cleavage and increased caspases and Bax protein expression. Collectively, lupeol adopted a multi-prong approach to interrupt different signaling cascades resulting in induction of apoptosis in PC cells [85].
Murtaza et al., studied the effect of lupeol on resistant tumor necrosis factor–related apoptosis-inducing ligand (TRAIL) therapy–resistant AsPC-1 cells overexpressing cellular FLICE like inhibitory protein (cFLIP). Incubation of AsPC-1 and PANC-1 cells with lupeol caused a dose-dependent antiproliferative effect with respective IC50 of 27.5 µM and 10 µM via PARP cleavage pathway. Indeed, lupeol was found to downregulate the transcriptional activation and expression of cFLIP and increase the cellular TRAIL protein level. It reduced AsPC-1 viability in either cFLIP-suppressed cells or cFLIP-overexpressing cells. Furthermore, lupeol significantly chemo-sensitized the highly resistant AsPC-1 cells to recombinant TRAIL-mediated apoptosis. These results were validated using xenograft mice model implanted with AsPC-1 cells where lupeol reduced the tumor volume and development without apparent signs of toxicity [86].
A recent study explored the effect of lupeol on KRAS signaling cascade. The authors found that lupeol snugly fits KRAS crystal structure (PDB: 4EPR) in silico and validated this outcome experimentally. Differential scanning fluorimetry (DSF) revealed that lupeol binds GDP-bound wild and mutant KRAS protein. In 2D and 3D cell culture models, lupeol exhibited significant antiproliferative effect on KRAS-induced cancer cells including PDE-KRAS, HCT116, and different National Cancer Institute (NCI)-KRAS active panel of cells with insignificant effect on normal pancreatic cells HPNE [87].
Arai et al., tried to isolate natural compounds from Zizyphus cambodiana targeting the hedgehog (Hh)/GLI signaling cascade which implicated in different tumors formation and development including PC [88]. Two lupeol analogues viz. colubrinic acid and betulinic acid (Figure 4) inhibited Hh/GLI1 pathway and GLI1-mediated transcriptional activity in PANC1 cells as identified by suppressing protein patched homolog (PTCH) and Bcl-2 using immunoblot assay. The cytotoxicity IC50 of colubrinic acid and betulinic acid was 43 µM and 44 µM, respectively as determined by fluorometric microculture cytotoxicity assay. Finally, colubrinic acid inhibited GLI-mediated mRNA expression of PTCH in PANC1 cells [89].
In another study, betulinic acid demonstrated antiproliferative effect on PC cells PANC-1 and SW1990 cells with respective IC50 47 and 38 µM. Wound healing test revealed that betulinic acid reduced the migration and invasion of PANC-1 and SW1990 cells dose-dependently. The proapoptotic effect is of betulinic acid belongs to mTOR pathway, increasing caspases and Bax and suppressing Bcl-2. In vivo validation was performed and confirmed that betulinic acid treatment reduced tumor volume and weight of transplanted PANC-1 tumor cells [90].
3.4. Friedelane type PTs in pancreatic cancer
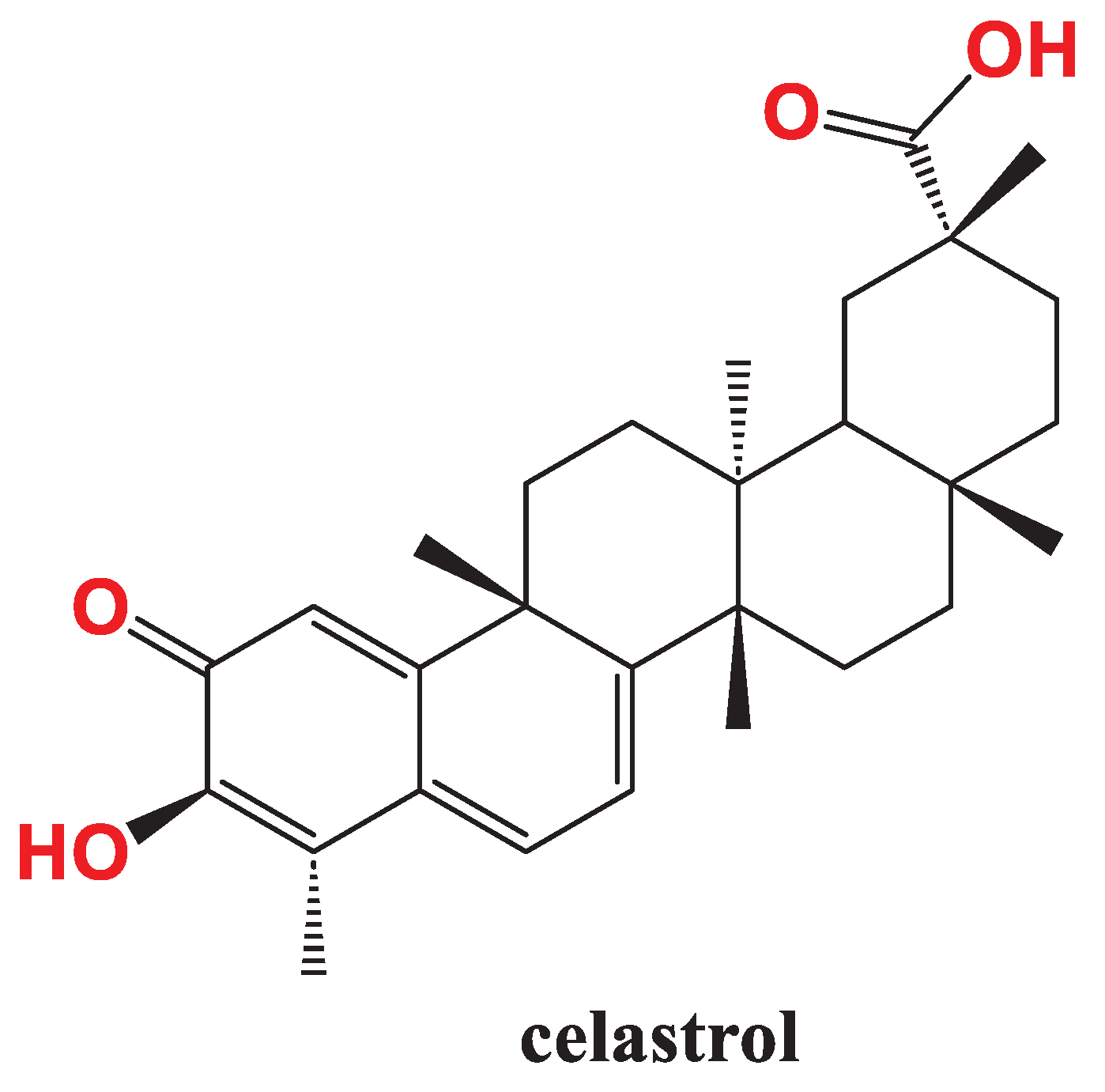
Celastrol (Figure 5) is an interesting representative of friedelane PTs with a unique quinone methide moiety and a vast array of bioactivities; it is extracted from Tripterygium wilfordii Hook F. [91,92]. Celastrol was identified as a chemokine receptor, CXCR4 inhibitor which implicated in metastasis; this effect was cell type-specific as it occurred in AsPC-1 and PANC-28 PC cells, breast cancer cells MCF-7 and MDA-MB-231, colorectal carcinoma cells HCT116, and squamous cell carcinoma SCC-4. It was found that celastrol downregulate mRNA level of CXCR4 as assessed by quantitative RTPCR [93]. Since CXCR4 is essential for CXCL12-induced invasion and metastasis [94], celastrol impeded CXCL12-induced invasion of AsPC-1 cells which indicate to the its promising role for circumvent cancer spreading [93].
Co-treatment with celastrol improved sensitivity of the resistant PANC-1 cells to TRAIL. Mechanistically, celastrol/TRAIL combination significantly upregulated 4E-binding protein 1 (4E-BP1) concentration and increased its dephosphorylation in PANC-1 cells, interrupted the formation of eIF4E/eIF4G complex and suppressed the anti-apoptotic c-FLIP and XIAP proteins synthesis. This is accompanied by enhanced generation of ROS and the activation of c-Jun N-terminal kinase (JNK). Celastrol sensitized BxPc-3 to TRAIL although BxPc-3 are irresponsive to TRAIL alone [95].
Youns et al., uncovered new genetic pathways modulated by celastrol in PC cells. Using sulforhodamine B (SRB) assay, they confirmed that celastrol suppressed the proliferation of 8 types of PC cells where Miapaca-2 and BxPc-3 were the most sensitive to celastrol treatment with respective IC50 7.30 µM and 7.79 µM with minimal effect on normal human pancreatic duct epithelial cell line (HPDE) (IC50>20µM). As anticipated, celastrol induced DNA fragmentation, activated caspases 3/7, and reduced prostaglandin E2 formation compared to untreated cells. RTPCR showed that celastrol upregulated DNA-damage-inducible transcript 4 (DDIT3) and activating transcription factor 3 (ATF3) and downregulated ribonucleotide reductase regulatory subunit M2 (RRM2) and minichromosome maintenance 4 (MCM4) genes [96].
4. Conclusions
In this review, we explored how PTs can modulate PC leading to prominent apoptotic cell and synergistic effect when combined eith other chemotherapies. OA stans out as a prominent cytotoxic agent against BxPc-3 cells (IC50 190 nM). OA proved considerable synergism when combined with 5-Fluorouracil or doxorubicin against PANC-28 or PANC-1 cells, respectively. An interesting OA derivative, K73-03 overcame gemcitabine resistance in AsPC-1 and MIA PaCa-2 cells. Furthermore, MA promoted TNFα antiproliferative effect but suppressed NF-kB activation in vitro and in vivo.
UA is an efficient chemosensitizer for gemcitabine against the resistant PANC-1, MIA PaCa-2 cells, and Capan-1 cells. It is main mechanism is suppressing KRAS downstream signaling and RAGE suppression. Other ursane type PTs such as ursenolide is an efficient suppressor of Akt/mTOR pathway in PANC-1 cells. Lupeol has a multi-prong modality against PC interrupting all KRAS downstream signaling. It acted as TRAIL sensitizer in AsPC-1 cells. Betulinic acid significantly reduced PANC-1 viability in vitro and in vivo by induction of apoptosis via Hh/GLI1 pathway inhibition.
Additionally, celastrol anticancer effect is attributed to CXCR4 inhibition. It reduced AsPC-1 viability, invasion, and metastasis. Furthermore, it sensitized PANC-1 cells to TRAIL treatment. Celastrol mechanism involves upregulation of DDIT3 and ATF3 and downregulation of MCM4 and MCM4 genes. Collectively, PTs can represent promising adjuvant chemotherapy for PC treatment inhibiting proliferation, invasion, and metastasis with a favorable safety profile.
Author Contributions
Conceptualization, M.O.R.; methodology, A.M.S.A., S.F.K., M.O.R.; software, A.M.S.A., S.F.K., M.O.R; investigation A.M.S.A, S.F.K.; resources A.M.S.A., S.F.K.; data curation A.M.S.A., S.F.K.; writing—original draft preparation, A.M.S.A., M.O.R.; writing—review and editing, A.M.S.A., M.O.R.; visualization, A.M.S.A. S.F.K.; supervision, M.O.R.; project administration, M.O.R.; funding acquisition, A.M.S.A, S.F.K. All authors have read and agreed to the published version of the manuscript.
Funding
Not applicable.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors are thankful to the Deanship of Graduate Studies and Scientific Research at University of Bisha for supporting this work through the Fast-Track Research Support Program.
Conflicts of Interest
All the authors declare that they have no conflict of interest of any kind..
References
- Siegel, R.L.; Miller, K.D.; Wagle, N.S.; Jemal, A. Cancer Statistics, 2023. CA. Cancer J. Clin. 2023, 73, 17–48. [Google Scholar] [CrossRef] [PubMed]
- Rahib, L.; Smith, B.D.; Aizenberg, R.; Rosenzweig, A.B.; Fleshman, J.M.; Matrisian, L.M. Projecting Cancer Incidence and Deaths to 2030: The Unexpected Burden of Thyroid, Liver, and Pancreas Cancers in the United States. Cancer Res. 2014, 74, 2913–2921. [Google Scholar] [CrossRef] [PubMed]
- Bosetti, C.; Lucenteforte, E.; Silverman, D.T.; Petersen, G.; Bracci, P.M.; Ji, B.T.; Negri, E.; Li, D.; Risch, H.A.; Olson, S.H.; et al. Cigarette Smoking and Pancreatic Cancer: An Analysis from the International Pancreatic Cancer Case-Control Consortium (Panc4). Ann. Oncol. Off. J. Eur. Soc. Med. Oncol. 2012, 23, 1880–1888. [Google Scholar] [CrossRef] [PubMed]
- Genkinger, J.M.; Spiegelman, D.; Anderson, K.E.; Bergkvist, L.; Bernstein, L.; van den Brandt, P.A.; English, D.R.; Freudenheim, J.L.; Fuchs, C.S.; Giles, G.G.; et al. Alcohol Intake and Pancreatic Cancer Risk: A Pooled Analysis of Fourteen Cohort Studies. Cancer Epidemiol. Biomark. Prev. Publ. Am. Assoc. Cancer Res. Cosponsored Am. Soc. Prev. Oncol. 2009, 18, 765–776. [Google Scholar] [CrossRef]
- Johansen, D.; Stocks, T.; Jonsson, H.; Lindkvist, B.; Björge, T.; Concin, H.; Almquist, M.; Häggström, C.; Engeland, A.; Ulmer, H.; et al. Metabolic Factors and the Risk of Pancreatic Cancer: A Prospective Analysis of Almost 580,000 Men and Women in the Metabolic Syndrome and Cancer Project. Cancer Epidemiol. Biomark. Prev. Publ. Am. Assoc. Cancer Res. Cosponsored Am. Soc. Prev. Oncol. 2010, 19, 2307–2317. [Google Scholar] [CrossRef] [PubMed]
- Park, W.; Chawla, A.; O’Reilly, E.M. Pancreatic Cancer: A Review. JAMA 2021, 326, 851–862. [Google Scholar] [CrossRef]
- Tanaka, A.; Radwan, M.O.; Hamasaki, A.; Ejima, A.; Obata, E.; Koga, R.; Tateishi, H.; Okamoto, Y.; Fujita, M.; Nakao, M.; et al. A Novel Inhibitor of Farnesyltransferase with a Zinc Site Recognition Moiety and a Farnesyl Group. Bioorg. Med. Chem. Lett. 2017, 27, 3862–3866. [Google Scholar] [CrossRef]
- Klochkov, S.G.; Neganova, M.E.; Yarla, N.S.; Parvathaneni, M.; Sharma, B.; Tarasov, V.V.; Barreto, G.; Bachurin, S.O.; Ashraf, G.M.; Aliev, G. Implications of Farnesyltransferase and Its Inhibitors as a Promising Strategy for Cancer Therapy. Semin. Cancer Biol. 2019, 56, 128–134. [Google Scholar] [CrossRef]
- Aguirre, A.J.; Bardeesy, N.; Sinha, M.; Lopez, L.; Tuveson, D.A.; Horner, J.; Redston, M.S.; DePinho, R.A. Activated Kras and Ink4a/Arf Deficiency Cooperate to Produce Metastatic Pancreatic Ductal Adenocarcinoma. Genes Dev. 2003, 17, 3112–3126. [Google Scholar] [CrossRef]
- Hingorani, S.R.; Wang, L.; Multani, A.S.; Combs, C.; Deramaudt, T.B.; Hruban, R.H.; Rustgi, A.K.; Chang, S.; Tuveson, D.A. Trp53R172H and KrasG12D Cooperate to Promote Chromosomal Instability and Widely Metastatic Pancreatic Ductal Adenocarcinoma in Mice. Cancer Cell 2005, 7, 469–483. [Google Scholar] [CrossRef] [PubMed]
- Bardeesy, N.; Cheng, K.-H.; Berger, J.H.; Chu, G.C.; Pahler, J.; Olson, P.; Hezel, A.F.; Horner, J.; Lauwers, G.Y.; Hanahan, D.; et al. Smad4 Is Dispensable for Normal Pancreas Development yet Critical in Progression and Tumor Biology of Pancreas Cancer. Genes Dev. 2006, 20, 3130–3146. [Google Scholar] [CrossRef] [PubMed]
- Izeradjene, K.; Combs, C.; Best, M.; Gopinathan, A.; Wagner, A.; Grady, W.M.; Deng, C.-X.; Hruban, R.H.; Adsay, N.V.; Tuveson, D.A.; et al. Kras(G12D) and Smad4/Dpc4 Haploinsufficiency Cooperate to Induce Mucinous Cystic Neoplasms and Invasive Adenocarcinoma of the Pancreas. Cancer Cell 2007, 11, 229–243. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Lee, H.N.; Jeong, M.S.; Jang, S.B. Oncogenic KRAS: Signaling and Drug Resistance. Cancers 2021, 13, 5599. [Google Scholar] [CrossRef] [PubMed]
- Merz, V.; Gaule, M.; Zecchetto, C.; Cavaliere, A.; Casalino, S.; Pesoni, C.; Contarelli, S.; Sabbadini, F.; Bertolini, M.; Mangiameli, D.; et al. Targeting KRAS: The Elephant in the Room of Epithelial Cancers. Front. Oncol. 2021, 11. [Google Scholar] [CrossRef]
- Jancík, S.; Drábek, J.; Radzioch, D.; Hajdúch, M. Clinical Relevance of KRAS in Human Cancers. J. Biomed. Biotechnol. 2010, 2010, 150960. [Google Scholar] [CrossRef] [PubMed]
- Zou, Z.; Tao, T.; Li, H.; Zhu, X. mTOR Signaling Pathway and mTOR Inhibitors in Cancer: Progress and Challenges. Cell Biosci. 2020, 10, 31. [Google Scholar] [CrossRef] [PubMed]
- D’Adamo, D.R.; Novick, S.; Kahn, J.M.; Leonardi, P.; Pellicer, A. Rsc: A Novel Oncogene with Structural and Functional Homology with the Gene Family of Exchange Factors for Ral. Oncogene 1997, 14, 1295–1305. [Google Scholar] [CrossRef]
- Lim, K.-H.; O’Hayer, K.; Adam, S.J.; Kendall, S.D.; Campbell, P.M.; Der, C.J.; Counter, C.M. Divergent Roles for RalA and RalB in Malignant Growth of Human Pancreatic Carcinoma Cells. Curr. Biol. CB 2006, 16, 2385–2394. [Google Scholar] [CrossRef]
- Liu, P.; Wang, Y.; Li, X. Targeting the Untargetable KRAS in Cancer Therapy. Acta Pharm. Sin. B 2019, 9, 871–879. [Google Scholar] [CrossRef]
- Carbone, C.; Melisi, D. NF-κB as a Target for Pancreatic Cancer Therapy. Expert Opin. Ther. Targets 2012, 16 Suppl 2, S1-10. [Google Scholar] [CrossRef]
- Henry, D.O.; Moskalenko, S.A.; Kaur, K.J.; Fu, M.; Pestell, R.G.; Camonis, J.H.; White, M.A. Ral GTPases Contribute to Regulation of Cyclin D1 through Activation of NF-κB. Mol. Cell. Biol. 2000, 20, 8084–8092. [Google Scholar] [CrossRef]
- Kolbeinsson, H.M.; Chandana, S.; Wright, G.P.; Chung, M. Pancreatic Cancer: A Review of Current Treatment and Novel Therapies. J. Investig. Surg. Off. J. Acad. Surg. Res. 2023, 36, 2129884. [Google Scholar] [CrossRef]
- Groot, V.P.; Rezaee, N.; Wu, W.; Cameron, J.L.; Fishman, E.K.; Hruban, R.H.; Weiss, M.J.; Zheng, L.; Wolfgang, C.L.; He, J. Patterns, Timing, and Predictors of Recurrence Following Pancreatectomy for Pancreatic Ductal Adenocarcinoma. Ann. Surg. 2018, 267, 936–945. [Google Scholar] [CrossRef] [PubMed]
- Halbrook, C.J.; Lyssiotis, C.A.; Pasca di Magliano, M.; Maitra, A. Pancreatic Cancer: Advances and Challenges. Cell 2023, 186, 1729–1754. [Google Scholar] [CrossRef] [PubMed]
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs from 1981 to 2014. J. Nat. Prod. 2016, 79, 629–661. [Google Scholar] [CrossRef] [PubMed]
- Koehn, F.E.; Carter, G.T. The Evolving Role of Natural Products in Drug Discovery. Nat. Rev. Drug Discov. 2005, 4, 206–220. [Google Scholar] [CrossRef] [PubMed]
- Cragg, G.M.; Newman, D.J. Plants as a Source of Anti-Cancer Agents. J. Ethnopharmacol. 2005, 100, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.; Lu, J.-J.; Ding, J. Natural Products in Cancer Therapy: Past, Present and Future. Nat. Prod. Bioprospecting 2021, 11, 5–13. [Google Scholar] [CrossRef] [PubMed]
- Moudi, M.; Go, R.; Yien, C.Y.S.; Nazre, M. Vinca Alkaloids. Int. J. Prev. Med. 2013, 4, 1231–1235. [Google Scholar] [PubMed]
- Shah, Z.; Gohar, U.F.; Jamshed, I.; Mushtaq, A.; Mukhtar, H.; Zia-UI-Haq, M.; Toma, S.I.; Manea, R.; Moga, M.; Popovici, B. Podophyllotoxin: History, Recent Advances and Future Prospects. Biomolecules 2021, 11, 603. [Google Scholar] [CrossRef]
- Ali, T.F.S.; Ciftci, H.I.; Radwan, M.O.; Koga, R.; Ohsugi, T.; Okiyama, Y.; Honma, T.; Nakata, A.; Ito, A.; Yoshida, M.; et al. New SIRT2 Inhibitors: Histidine-Based Bleomycin Spin-Off. Bioorg. Med. Chem. 2019, 27, 1767–1775. [Google Scholar] [CrossRef] [PubMed]
- Koba, M.; Konopa, J. [Actinomycin D and its mechanisms of action]. Postepy Hig. Med. Doswiadczalnej Online 2005, 59, 290–298. [Google Scholar]
- Alves, C.; Silva, J.; Pinteus, S.; Gaspar, H.; Alpoim, M.C.; Botana, L.M.; Pedrosa, R. From Marine Origin to Therapeutics: The Antitumor Potential of Marine Algae-Derived Compounds. Front. Pharmacol. 2018, 9, 777. [Google Scholar] [CrossRef] [PubMed]
- Gallego-Jara, J.; Lozano-Terol, G.; Sola-Martínez, R.A.; Cánovas-Díaz, M.; de Diego Puente, T. A Compressive Review about Taxol®: History and Future Challenges. Molecules 2020, 25, 5986. [Google Scholar] [CrossRef] [PubMed]
- Yue, Q.; Gao, G.; Zou, G.; Yu, H.; Zheng, X. Natural Products as Adjunctive Treatment for Pancreatic Cancer: Recent Trends and Advancements. BioMed Res. Int. 2017, 2017, e8412508. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Wang, N.; Zhang, Y.; Huang, X.; Wang, Y. The Therapeutic Potential of Natural Products for Treating Pancreatic Cancer. Front. Pharmacol. 2022, 13. [Google Scholar] [CrossRef] [PubMed]
- Kim, A.; Ha, J.; Kim, J.; Cho, Y.; Ahn, J.; Cheon, C.; Kim, S.-H.; Ko, S.-G.; Kim, B. Natural Products for Pancreatic Cancer Treatment: From Traditional Medicine to Modern Drug Discovery. Nutrients 2021, 13, 3801. [Google Scholar] [CrossRef]
- Hosseini, M.; Hassanian, S.M.; Mohammadzadeh, E.; ShahidSales, S.; Maftouh, M.; Fayazbakhsh, H.; Khazaei, M.; Avan, A. Therapeutic Potential of Curcumin in Treatment of Pancreatic Cancer: Current Status and Future Perspectives. J. Cell. Biochem. 2017, 118, 1634–1638. [Google Scholar] [CrossRef]
- Rambam Health Care Campus Phase II Trial of Gemcitabine and Curcumin in Patients With Advanced Pancreatic Cancer; clinicaltrials.gov, 2010.
- Yang, L.; Yang, L.; Tian, W.; Li, J.; Liu, J.; Zhu, M.; Zhang, Y.; Yang, Y.; Liu, F.; Zhang, Q.; et al. Resveratrol Plays Dual Roles in Pancreatic Cancer Cells. J. Cancer Res. Clin. Oncol. 2014, 140, 749–755. [Google Scholar] [CrossRef]
- Qin, T.; Cheng, L.; Xiao, Y.; Qian, W.; Li, J.; Wu, Z.; Wang, Z.; Xu, Q.; Duan, W.; Wong, L.; et al. NAF-1 Inhibition by Resveratrol Suppresses Cancer Stem Cell-Like Properties and the Invasion of Pancreatic Cancer. Front. Oncol. 2020, 10. [Google Scholar] [CrossRef]
- Ratajczak, K.; Glatzel-Plucińska, N.; Ratajczak-Wielgomas, K.; Nowińska, K.; Borska, S. Effect of Resveratrol Treatment on Human Pancreatic Cancer Cells through Alterations of Bcl-2 Family Members. Molecules 2021, 26, 6560. [Google Scholar] [CrossRef]
- Radwan, M.O.; Ismail, M.A.H.; El-Mekkawy, S.; Ismail, N.S.M.; Hanna, A.G. Synthesis and Biological Activity of New 18β-Glycyrrhetinic Acid Derivatives. Arab. J. Chem. 2016, 9, 390–399. [Google Scholar] [CrossRef]
- Tsai, S.-J.; Yin, M.-C. Antioxidative and Anti-Inflammatory Protection of Oleanolic Acid and Ursolic Acid in PC12 Cells. J. Food Sci. 2008, 73, H174-178. [Google Scholar] [CrossRef] [PubMed]
- Somova, L.I.; Shode, F.O.; Ramnanan, P.; Nadar, A. Antihypertensive, Antiatherosclerotic and Antioxidant Activity of Triterpenoids Isolated from Olea Europaea, Subspecies Africana Leaves. J. Ethnopharmacol. 2003, 84, 299–305. [Google Scholar] [CrossRef] [PubMed]
- Kadasah, S.F.; Radwan, M.O. Overview of Ursolic Acid Potential for the Treatment of Metabolic Disorders, Autoimmune Diseases, and Cancers via Nuclear Receptor Pathways. Biomedicines 2023, 11, 2845. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Liu, Y.; Wang, X.; Tian, Z.; Qi, D.; Li, Y.; Jiang, H. Antihypertensive Activity of Oleanolic Acid Is Mediated via Downregulation of Secretory Phospholipase A2 and Fatty Acid Synthase in Spontaneously Hypertensive Rats. Int. J. Mol. Med. 2020, 46, 2019–2034. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Rebolledo, G.A.; Siordia-Reyes, A.G.; Meckes-Fischer, M.; Jiménez-Arellanes, A. Hepatoprotective Properties of Oleanolic and Ursolic Acids in Antitubercular Drug-Induced Liver Damage. Asian Pac. J. Trop. Med. 2016, 9, 644–651. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.-B.; Xiao, Y.-H.; Zhang, Q.-Y.; Zhou, M.; Liao, S.-G. Hepatoprotective Natural Triterpenoids. Eur. J. Med. Chem. 2018, 145, 691–716. [Google Scholar] [CrossRef]
- Radwan, M.O.; Kadasah, S.F.; Aljubiri, S.M.; Alrefaei, A.F.; El-Maghrabey, M.H.; El Hamd, M.A.; Tateishi, H.; Otsuka, M.; Fujita, M. Harnessing Oleanolic Acid and Its Derivatives as Modulators of Metabolic Nuclear Receptors. Biomolecules 2023, 13, 1465. [Google Scholar] [CrossRef]
- Pompei, R.; Laconi, S.; Ingianni, A. Antiviral Properties of Glycyrrhizic Acid and Its Semisynthetic Derivatives. Mini Rev. Med. Chem. 2009, 9, 996–1001. [Google Scholar] [CrossRef]
- Sun, Z.-G.; Zhao, T.-T.; Lu, N.; Yang, Y.-A.; Zhu, H.-L. Research Progress of Glycyrrhizic Acid on Antiviral Activity. Mini Rev. Med. Chem. 2019, 19, 826–832. [Google Scholar] [CrossRef]
- Yang, Y.; Huang, Y.; Huang, C.; Lv, X.; Liu, L.; Wang, Y.; Li, J. Antifibrosis Effects of Triterpene Acids of Eriobotrya Japonica (Thunb.) Lindl. Leaf in a Rat Model of Bleomycin-Induced Pulmonary Fibrosis. J. Pharm. Pharmacol. 2012, 64, 1751–1760. [Google Scholar] [CrossRef]
- Lee, M.K.; Lee, K.Y.; Jeon, H.Y.; Sung, S.H.; Kim, Y.C. Antifibrotic Activity of Triterpenoids from the Aerial Parts of Euscaphis Japonica on Hepatic Stellate Cells. J. Enzyme Inhib. Med. Chem. 2009, 24, 1276–1279. [Google Scholar] [CrossRef]
- Somensi, L.B.; Costa, P.; Boeing, T.; Mariano, L.N.B.; Longo, B.; Magalhães, C.G.; Duarte, L.P.; Maciel E Silva, A.T.; de Souza, P.; de Andrade, S.F.; et al. Gastroprotective Properties of Lupeol-Derived Ester: Pre-Clinical Evidences of Lupeol-Stearate as a Potent Antiulcer Agent. Chem. Biol. Interact. 2020, 321, 108964. [Google Scholar] [CrossRef]
- Chudzik, M.; Korzonek-Szlacheta, I.; Król, W. Triterpenes as Potentially Cytotoxic Compounds. Molecules 2015, 20, 1610–1625. [Google Scholar] [CrossRef]
- I. Ciftci, H.; O. Radwan, M.; E. Ozturk, S.; Ulusoy, N.G.; Sozer, E.; E. Ellakwa, D.; Ocak, Z.; Can, M.; F.S. Ali, T.; I. Abd-Alla, H.; et al. Design, Synthesis and Biological Evaluation of Pentacyclic Triterpene Derivatives: Optimization of Anti-ABL Kinase Activity. Molecules 2019, 24, 3535–3535. [Google Scholar] [CrossRef]
- Tang, Z.-Y.; Li, Y.; Tang, Y.-T.; Ma, X.-D.; Tang, Z.-Y. Anticancer Activity of Oleanolic Acid and Its Derivatives: Recent Advances in Evidence, Target Profiling and Mechanisms of Action. Biomed. Pharmacother. Biomedecine Pharmacother. 2022, 145, 112397. [Google Scholar] [CrossRef] [PubMed]
- Radwan, M.O.; Abd-Alla, H.I.; Alsaggaf, A.T.; El-Mezayen, H.; Abourehab, M.A.S.; El-Beeh, M.E.; Tateishi, H.; Otsuka, M.; Fujita, M. Gypsogenin Battling for a Front Position in the Pentacyclic Triterpenes Game of Thrones on Anti-Cancer Therapy: A Critical Review—Dedicated to the Memory of Professor Hanaa M. Rady. Molecules 2023, 28, 5677. [Google Scholar] [CrossRef] [PubMed]
- Inoue, M.; Honma, Y.; Urano, T.; Suzumiya, J. Japanese Apricot Extract (MK615) Potentiates Bendamustine-Induced Apoptosis via Impairment of the DNA Damage Response in Lymphoma Cells. Oncol. Lett. 2017, 14, 792–800. [Google Scholar] [CrossRef] [PubMed]
- Hattori, M.; Kawakami, K.; Akimoto, M.; Takenaga, K.; Suzumiya, J.; Honma, Y. Antitumor Effect of Japanese Apricot Extract (MK615) on Human Cancer Cells in Vitro and in Vivo through a Reactive Oxygen Species-Dependent Mechanism. Tumori 2013, 99, 239–248. [Google Scholar] [CrossRef]
- Cheng, S.; Eliaz, I.; Lin, J.; Sliva, D. Triterpenes from Poria Cocos Suppress Growth and Invasiveness of Pancreatic Cancer Cells through the Downregulation of MMP-7 Erratum in /Ijo/44/5/1781. Int. J. Oncol. 2013, 42, 1869–1874. [Google Scholar] [CrossRef]
- Cheng, S.; Castillo, V.; Sliva, D. CDC20 Associated with Cancer Metastasis and Novel Mushroom-derived CDC20 Inhibitors with Antimetastatic Activity. Int. J. Oncol. 2019, 54, 2250–2256. [Google Scholar] [CrossRef] [PubMed]
- Castellano, J.M.; Ramos-Romero, S.; Perona, J.S. Oleanolic Acid: Extraction, Characterization and Biological Activity. Nutrients 2022, 14, 623. [Google Scholar] [CrossRef]
- Dall’Acqua, S.; Peron, G.; Ferrari, S.; Gandin, V.; Bramucci, M.; Quassinti, L.; Mártonfi, P.; Maggi, F. Phytochemical Investigations and Antiproliferative Secondary Metabolites from Thymus Alternans Growing in Slovakia. Pharm. Biol. 2017, 55, 1162–1170. [Google Scholar] [CrossRef]
- Wei, J.; Liu, H.; Liu, M.; Wu, N.; Zhao, J.; Xiao, L.; Han, L.; Chu, E.; Lin, X. Oleanolic Acid Potentiates the Antitumor Activity of 5-Fluorouracil in Pancreatic Cancer Cells. Oncol. Rep. 2012, 28, 1339–1345. [Google Scholar] [CrossRef]
- Esmaeili, H.; Nasrollahzadeh Sabet, M.; Mosaed, R.; Chamanara, M.; Hadi, S.; Hazrati, E.; Farhadi, A.; Heidari, M.F.; Behroozi, J. Oleanolic Acid Increases the Anticancer Potency of Doxorubicin in Pancreatic Cancer Cells. J. Biochem. Mol. Toxicol. 2023, 37, e23426. [Google Scholar] [CrossRef] [PubMed]
- Deeb, D.; Gao, X.; Liu, Y.; Varma, N.; Arbab, A.; Gautam, S. Inhibition of Telomerase Activity by Oleanane Triterpenoid CDDO-Me in Pancreatic Cancer Cells Is ROS-Dependent. Molecules 2013, 18, 3250–3265. [Google Scholar] [CrossRef]
- Chen, F.; Long, Q.; Fu, D.; Zhu, D.; Ji, Y.; Han, L.; Zhang, B.; Xu, Q.; Liu, B.; Li, Y.; et al. Targeting SPINK1 in the Damaged Tumour Microenvironment Alleviates Therapeutic Resistance. Nat. Commun. 2018, 9, 4315. [Google Scholar] [CrossRef] [PubMed]
- Shopit, A.; Li, X.; Tang, Z.; Awsh, M.; Shobet, L.; Niu, M.; Wang, H.; Mousa, H.; Alshwmi, M.; Tesfaldet, T.; et al. miR-421 up-Regulation by the Oleanolic Acid Derivative K73-03 Regulates Epigenetically SPINK1 Transcription in Pancreatic Cancer Cells Leading to Metabolic Changes and Enhanced Apoptosis. Pharmacol. Res. 2020, 161, 105130. [Google Scholar] [CrossRef]
- Shopit, A.; Li, X.; Wang, S.; Awsh, M.; Safi, M.; Chu, P.; Jia, J.; Al-Radhi, M.; Baldi, S.; Wang, F.; et al. Enhancement of Gemcitabine Efficacy by K73-03 via Epigenetically Regulation of miR-421/SPINK1 in Gemcitabine Resistant Pancreatic Cancer Cells. Phytomedicine Int. J. Phytother. Phytopharm. 2021, 91, 153711. [Google Scholar] [CrossRef]
- Zhou, Z.; Dong, Y.; Li, N.; Niu, M.; Wang, S.; Zhou, Y.; Sun, Z.; Chu, P.; Tang, Z. An Oleanolic Acid Derivative, K73-03, Inhibits Pancreatic Cancer Cells Proliferation in Vitro and in Vivo via Blocking EGFR/Akt Pathway. Cell Biol. Int. 2022, 46, 1801–1813. [Google Scholar] [CrossRef]
- Li, C.; Yang, Z.; Zhai, C.; Qiu, W.; Li, D.; Yi, Z.; Wang, L.; Tang, J.; Qian, M.; Luo, J.; et al. Maslinic Acid Potentiates the Anti-Tumor Activity of Tumor Necrosis Factor a by Inhibiting NF- B Signaling Pathway. 2010. [Google Scholar] [CrossRef]
- Lin, C.; Yan, S.; Yin, M. Inhibitory Effects of Maslinic Acid upon Human Esophagus, Stomach and Pancreatic Cancer Cells. J. Funct. Foods 2014, 11, 581–588. [Google Scholar] [CrossRef]
- Aguilera-Garrido, A.; Arranz, E.; Gálvez-Ruiz, M.J.; Marchal, J.A.; Galisteo-González, F.; Giblin, L. Solid Lipid Nanoparticles to Improve Bioaccessibility and Permeability of Orally Administered Maslinic Acid. Drug Deliv. 2022, 29, 1971–1982. [Google Scholar] [CrossRef] [PubMed]
- Aguilera-Garrido, A.; Graván, P.; Navarro-Marchal, S.A.; Medina-O’Donnell, M.; Parra, A.; Gálvez-Ruiz, M.J.; Marchal, J.A.; Galisteo-González, F. Maslinic Acid Solid Lipid Nanoparticles as Hydrophobic Anticancer Drug Carriers: Formulation, in Vitro Activity and in Vivo Biodistribution. Biomed. Pharmacother. 2023, 163, 114828. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Liang, X.; Yang, X. Ursolic Acid Inhibits Growth and Induces Apoptosis in Gemcitabine-Resistant Human Pancreatic Cancer via the JNK and PI3K/Akt/NF-κB Pathways. Oncol. Rep. 2012, 28, 501–510. [Google Scholar] [CrossRef] [PubMed]
- Prasad, S.; Yadav, V.R.; Sung, B.; Gupta, S.C.; Tyagi, A.K.; Aggarwal, B.B. Ursolic Acid Inhibits the Growth of Human Pancreatic Cancer and Enhances the Antitumor Potential of Gemcitabine in an Orthotopic Mouse Model through Suppression of the Inflammatory Microenvironment. Oncotarget 2016, 7, 13182–13196. [Google Scholar] [CrossRef] [PubMed]
- Shahab, U.; Ahmad, M.K.; Mahdi, A.A.; Waseem, M.; Arif, B.; Moinuddin, null; Ahmad, S. The Receptor for Advanced Glycation End Products: A Fuel to Pancreatic Cancer. Semin. Cancer Biol. 2018, 49, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Swami, P.; Thiyagarajan, S.; Vidger, A.; Indurthi, V.S.K.; Vetter, S.W.; Leclerc, E. RAGE Up-Regulation Differently Affects Cell Proliferation and Migration in Pancreatic Cancer Cells. Int. J. Mol. Sci. 2020, 21, 7723. [Google Scholar] [CrossRef] [PubMed]
- Olaoba, O.T.; Kadasah, S.; Vetter, S.W.; Leclerc, E. RAGE Signaling in Melanoma Tumors. Int. J. Mol. Sci. 2020, 21, 8989. [Google Scholar] [CrossRef]
- Li, Z.-Y.; Chen, S.-Y.; Weng, M.-H.; Yen, G.-C. Ursolic Acid Restores Sensitivity to Gemcitabine through the RAGE/NF-κB/MDR1 Axis in Pancreatic Cancer Cells and in a Mouse Xenograft Model. J. Food Drug Anal. 2021, 29, 262–274. [Google Scholar] [CrossRef]
- Kang, R.; Hou, W.; Zhang, Q.; Chen, R.; Lee, Y.J.; Bartlett, D.L.; Lotze, M.T.; Tang, D.; Zeh, H.J. RAGE Is Essential for Oncogenic KRAS-Mediated Hypoxic Signaling in Pancreatic Cancer. Cell Death Dis. 2014, 5, e1480. [Google Scholar] [CrossRef]
- Tawila, A.M.; Sun, S.; Kim, M.J.; Omar, A.M.; Dibwe, D.F.; Awale, S. A Triterpene Lactone from Callistemon Citrinus Inhibits the PANC-1 Human Pancreatic Cancer Cells Viability through Suppression of Unfolded Protein Response. Chem. Biodivers. 2020, 17, e2000495. [Google Scholar] [CrossRef]
- Saleem, M.; Kaur, S.; Kweon, M.-H.; Adhami, V.M.; Afaq, F.; Mukhtar, H. Lupeol, a Fruit and Vegetable Based Triterpene, Induces Apoptotic Death of Human Pancreatic Adenocarcinoma Cells via Inhibition of Ras Signaling Pathway. Carcinogenesis 2005, 26, 1956–1964. [Google Scholar] [CrossRef] [PubMed]
- Murtaza, I.; Saleem, M.; Adhami, V.M.; Hafeez, B.B.; Mukhtar, H. Suppression of cFLIP by Lupeol, a Dietary Triterpene, Is Sufficient to Overcome Resistance to TRAIL-Mediated Apoptosis in Chemoresistant Human Pancreatic Cancer Cells. Cancer Res. 2009, 69, 1156–1165. [Google Scholar] [CrossRef] [PubMed]
- Ganaie, A.A.; Siddique, H.R.; Sheikh, I.A.; Parray, A.; Wang, L.; Panyam, J.; Villalta, P.W.; Deng, Y.; Konety, B.R.; Saleem, M. A Novel Terpenoid Class for Prevention and Treatment of KRAS -driven Cancers: Comprehensive Analysis Using in Situ, in Vitro, and in Vivo Model Systems. Mol. Carcinog. 2020, 59, 886–896. [Google Scholar] [CrossRef] [PubMed]
- Thayer, S.P.; di Magliano, M.P.; Heiser, P.W.; Nielsen, C.M.; Roberts, D.J.; Lauwers, G.Y.; Qi, Y.P.; Gysin, S.; Fernández-del Castillo, C.; Yajnik, V.; et al. Hedgehog Is an Early and Late Mediator of Pancreatic Cancer Tumorigenesis. Nature 2003, 425, 851–856. [Google Scholar] [CrossRef] [PubMed]
- Arai, M.A.; Tateno, C.; Hosoya, T.; Koyano, T.; Kowithayakorn, T.; Ishibashi, M. Hedgehog/GLI-Mediated Transcriptional Inhibitors from Zizyphus Cambodiana. Bioorg. Med. Chem. 2008, 16, 9420–9424. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Zhu, H.; Weng, M.; Wang, C.; Sun, L. Chemopreventive Effect of Betulinic Acid via mTOR -Caspases/Bcl2/Bax Apoptotic Signaling in Pancreatic Cancer. BMC Complement. Med. Ther. 2020, 20, 178. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Hu, T.; Gao, L.; Su, P.; Zhang, Y.; Zhao, Y.; Chen, S.; Tu, L.; Song, Y.; Wang, X.; et al. Friedelane-Type Triterpene Cyclase in Celastrol Biosynthesis from Tripterygium Wilfordii and Its Application for Triterpenes Biosynthesis in Yeast. New Phytol. 2019, 223, 722–735. [Google Scholar] [CrossRef] [PubMed]
- Venkatesha, S.H.; Moudgil, K.D. Celastrol and Its Role in Chronic Diseases. Adv. Exp. Med. Biol. 2016, 928, 267–289. [Google Scholar] [CrossRef] [PubMed]
- Yadav, V.R.; Sung, B.; Prasad, S.; Kannappan, R.; Cho, S.-G.; Liu, M.; Chaturvedi, M.M.; Aggarwal, B.B. Celastrol Suppresses Invasion of Colon and Pancreatic Cancer Cells through the Downregulation of Expression of CXCR4 Chemokine Receptor. J. Mol. Med. 2010, 88, 1243–1253. [Google Scholar] [CrossRef]
- Marchesi, F.; Monti, P.; Leone, B.E.; Zerbi, A.; Vecchi, A.; Piemonti, L.; Mantovani, A.; Allavena, P. Increased Survival, Proliferation, and Migration in Metastatic Human Pancreatic Tumor Cells Expressing Functional CXCR4. Cancer Res. 2004, 64, 8420–8427. [Google Scholar] [CrossRef]
- Chakravarthy, R.; Clemens, M.J.; Pirianov, G.; Perdios, N.; Mudan, S.; Cartwright, J.E.; Elia, A. Role of the eIF4E Binding Protein 4E-BP1 in Regulation of the Sensitivity of Human Pancreatic Cancer Cells to TRAIL and Celastrol-induced Apoptosis. Biol. Cell 2013, 105, 414–429. [Google Scholar] [CrossRef] [PubMed]
- Youns, M.; Askoura, M.; Abbas, H.A.; Attia, G.H.; Khayyat, A.N.; Goda, R.M.; Almalki, A.J.; Khafagy, E.-S.; Hegazy, W.A.H. Celastrol Modulates Multiple Signaling Pathways to Inhibit Proliferation of Pancreatic Cancer via DDIT3 and ATF3 Up-Regulation and RRM2 and MCM4 Down-Regulation. OncoTargets Ther. 2021, 14, 3849–3860. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
KRAS signaling cascade implicated in pancreatic cancer through three effector pathways.
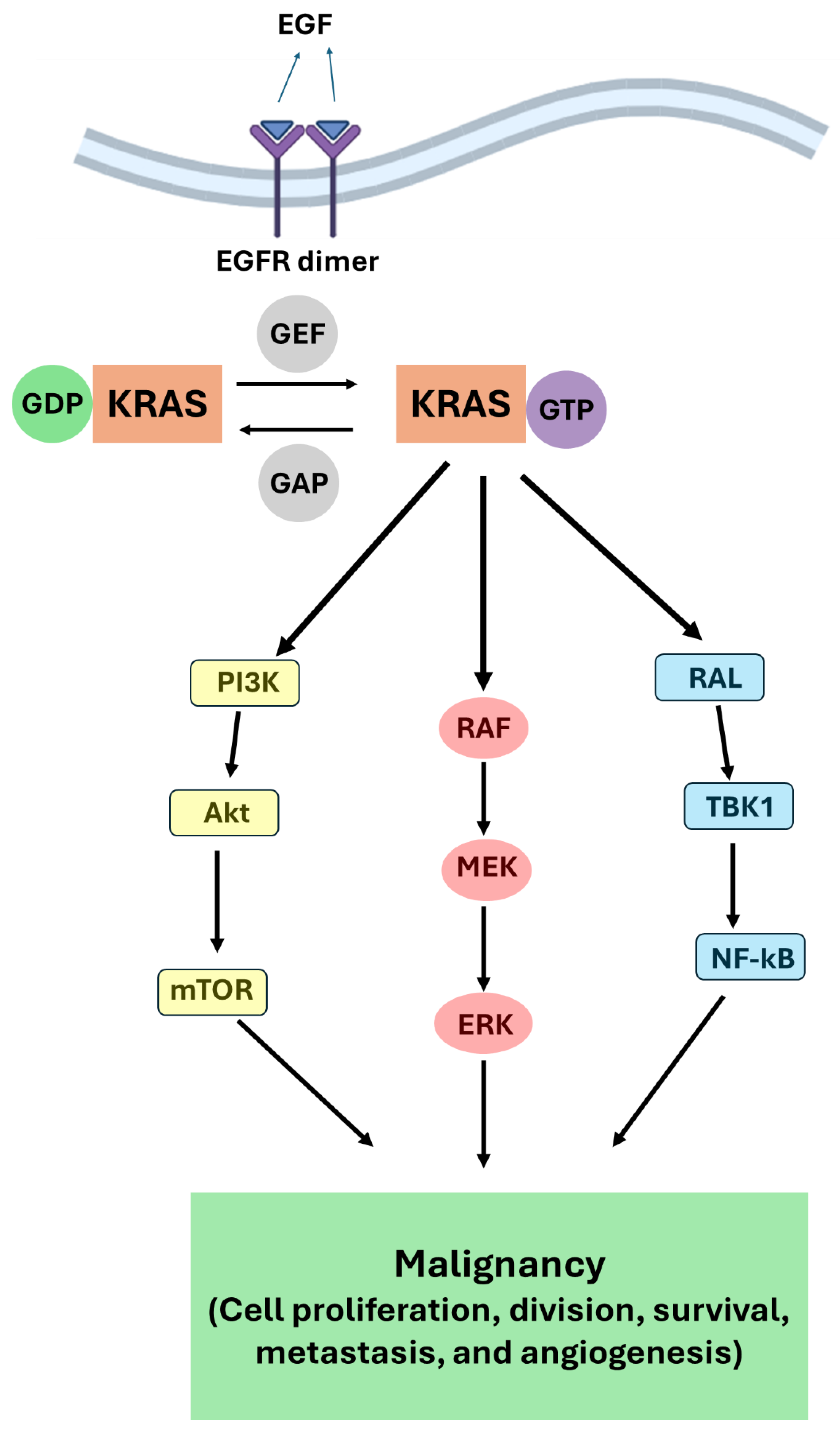
Figure 2.
Oleanane type pentacyclic triterpenes (PT) active against pancreatic cancer (PC).
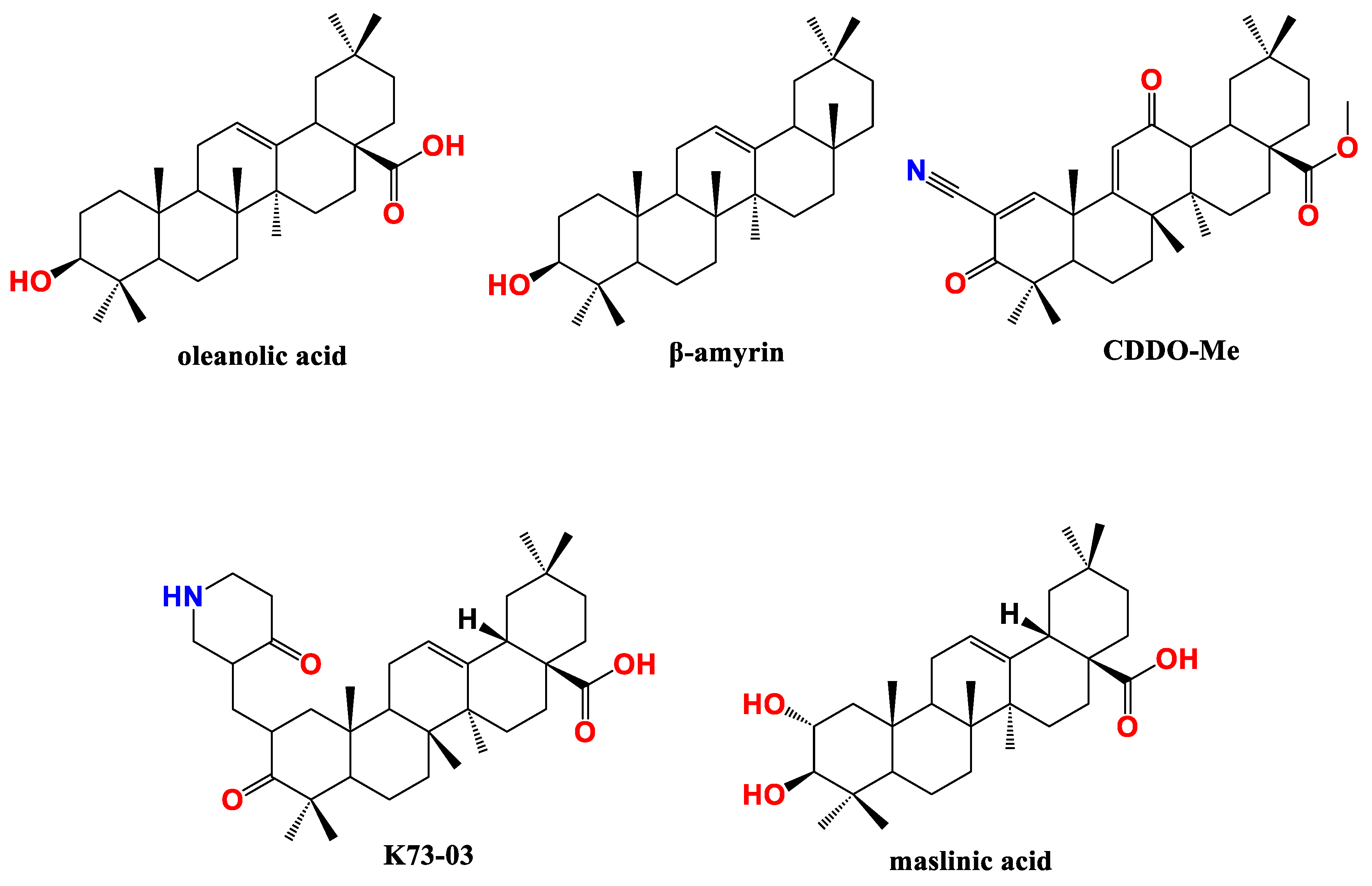
Figure 3.
Ursane type pentacyclic triterpenes (PT) active against pancreatic cancer (PC).
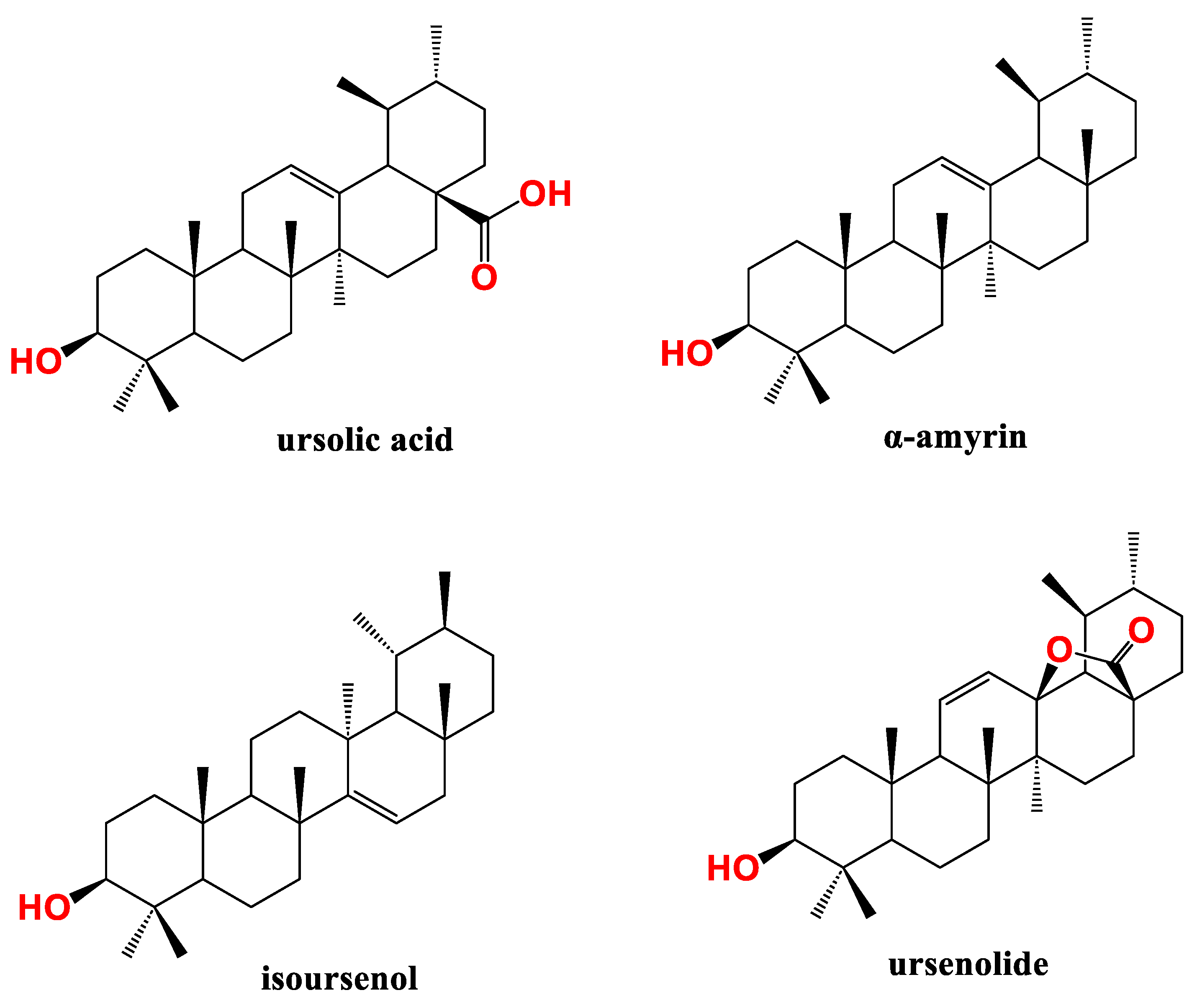
Figure 4.
Lupane type pentacyclic triterpenes (PT) active against pancreatic cancer (PC).
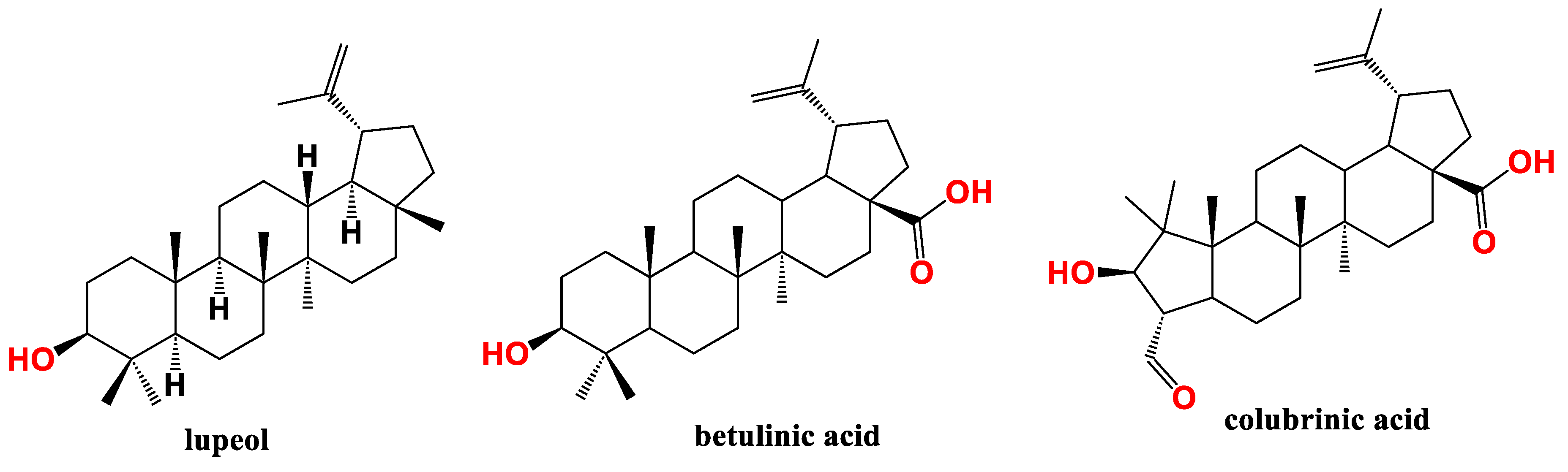
Figure 5.
Structural formula of celastrol.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



