
Submitted:
29 February 2024
Posted:
01 March 2024
You are already at the latest version
Abstract
Green spaces localized in cities create favourable environmental conditions for Ixodes ricinus, the most widespread and important vector of tick-borne pathogens in Central Europe. In this study, we described the prevalence of Borrelia burgdorferi s.l. and Babesia species found in mono- and double infections among I. ricinus ticks occurring in urban areas of the city of Poznań, west-central Poland. Two tick groups were examined: (i) a group of 1,029 host-seeking ticks (460 nymphs, 289 females, and 280 males and (ii) a group of 1,268 engorged female ticks removed from 1,115 tick-infested animals, dogs and cats. The overall prevalence of B. burgdorferi s.l. found in host-seeking ticks was almost two-fold higher in comparison to ticks derived from pets (8.7 vs. 4.7%, respectively, χ2 test, P = 0.001). Borrelia afzelii and B. garinii were the most prevalent species both in ticks from vegetation (3.7% and 3.7%, respectively) and from pets (3.7% and 0.6%, respectively). Babesia infections were slightly more common in feeding ticks compared to those from vegetation (6.0 vs. 4.7%, respectively), however, the difference was not significant. Three Babesia species including Ba. microti, Ba. venatorum and Ba. canis were identified. Babesia canis and Ba. microti were the most prevalent in host-seeking (2.6% and 1.4%, respectively) and feeding ticks (2.8% and 2.2%, respectively), whereas Ba. venatorum was the rarest piroplasm. All Ba microti obtained sequences (n=42) were identical to Ba. microti genotype "Jena/Germany" that is considered as pathogenic to humans. Furthermore, the sequences of Ba. venatorum (n=17) were identical with those isolated from patients in Italy, Austria and Poland. Altogether, out of 121 Babesia-positive ticks, 19.8% were simultaneously infected with B. burgdorferi s.l. spirochetes. We found similar prevalences of co-infections in host-seeking (1.1%) as well as in feeding ticks (1.0%). In the first group, Ba. canis prevailed in coinfections (73%), mostly with B. afzelii and B. garinii, in the second group, Babesia microti dominated with B. afzelii (92.7%). This is the first finding documenting the co-occurrence of both pathogens in ticks from companion animals. The detection of Ba. microti and Ba. venatorum sequences identical to strains pathogenic to humans as well as predominance of LB spirochetes, B. afzelii and B. garinii in I. ricinus from vegetation and animals indicates that there is a potential risk of acquiring human babesiosis and/or Lyme borreliosis in urban areas of the city of Poznań. Although, the overall risk of encountering a Ba. microti and B. burgdorferi s.l. co-infected tick was low (1.0%), the potential for more severe clinical symptoms and complications deserves notice. A relatively high infection rates of Ba. canis in dog-derived ticks (3.2%) and in host-seeking ticks (2.6%), suggests that I. ricinus could potentially be involved in the circulation of this piroplasm in areas, where the competent vector, Dermacentor reticulatus is absent or rare.
Keywords:
Ixodes ricinus
; Borrelia burgdorferi s.l.
; Babesia microti
; Babesia venatorum
; Babesia canis
; Borrelia miyamotoi
; coinfections
; babesiosis
; urban green areas
Introduction
Green spaces inside European towns such as parks, leisure-time areas for hiking and biking, botanic gardens, private properties with gardens, cemeteries, and urban forests, create favourable environmental (e.g., temperature and humidity) conditions for Ixodes ricinus, the most widespread and important vector of tick-borne pathogens (TBPs) in Central Europe. One of the key factors influencing the survival and maintenance of local tick populations is their access to appropriate and abundant tick bloodmeal hosts [1]. Within urban green areas, immature stages of this primarily a forest-dwelling tick species, feed mostly on small rodents, ground-feeding passerines, and hedgehogs which are additionally important hosts of adult female ticks. The latter feed also on urban pet populations represented by dogs and cats, including stray animals. The observed ongoing increase in the number of pets in towns, suggests that this group of mammals appears to be increasingly important for the persistence and size of tick populations in these ecologically altered habitats. Some of vertebrate species acting as maintenance hosts for I. ricinus, may concurrently serve as reservoir hosts of TBPs. Their high or low abundance and species composition in urban habitats, influence the level of infection of local tick populations and is critical for public health importance [1]. Therefore, investigations regarding the prevalence of TBPs infecting ticks, are necessary to establish or predict the emergence of active endemic foci of tick-borne diseases. It is particularly crucial because, there is still a lack of comprehensive knowledge on the eco-epidemiology of these infections in urban ecosystems and our understanding of how urbanization affects pathogen-host-vector relationships [2].
Ticks can carry two or more pathogenic microorganisms with a subsequent high likelihood of co-transmission to humans or animals [3,4]. Ixodes ricinus ticks infected with various bacteria, i.e., spirochetes of the Borrelia burgdorferi sensu lato (s.l.) complex, the agent of Lyme borreliosis (LB) or Anaplasma phagocytophilum, the agent of human anaplasmosis, are regularly found in urban and suburban areas across Europe [5,6,7]. Furthermore, B. burgdorferi s.l. spirochetes and intraerythrocytic parasites of the protozoan genus Babesia (Apicomplexa: Piroplasmida), including Ba. microti, Ba. divergens, Ba. venatorum, and Ba. duncani (present only in North America), can co-occur and be co-transmitted by ticks of the Ixodes ricinus species complex [8]. These protozoan pathogens are responsible for human babesiosis causing a febrile hemolytic anemia, that is generally asymptomatic or self-limiting in healthy humans, however, is a serious health concern in splenectomised, immunocompromised patients [9]. In North America, the major agent of human babesiosis is Ba. microti, a parasite associated with small mammals serving as primary reservoir hosts. There are eco-epidemiological evidence indicating that coinfections B. burgdorferi s.l. and Ba. microti among ticks and Peromyscus leucopus mice may contribute to the emergence and expansion of Ba. microti in the enzootic cycle. Ecological models demonstrated the strongest effects when the prevalence of B. burgdorferi in mice was high [10]. Recent research clearly demonstrated that infection of I. scapularis ticks with B. burgdorferi s.l. spirochetes increases the likelihood of infection with Ba. microti as well as A. phagocytophilum compared with borreliae-free ticks [11]. Furthermore, according to Zembsch et al. [12] host-seeking I. scapularis ticks that are infected with Ba. microti, are more likely to be coinfected with B. burgdorferi than expected if the pathogens were transmitted independently. This implies that such positive pathogen-vector-host interactions of both tick-borne agents may favour their emergence and maintenance in local tick populations [13]. Co-infections may also change clinical symptoms, course, severity of tick-associated disease in humans and animals compared to those induced by a single infection [14,15,16]. Patients co-infected with B. burgdorferi s.l. and Ba. microti suffer from significantly more diverse, intense, and persisting disease symptoms compared to those infected with each pathogen separately [17,18]. It has been shown that Ba. microti weakens adaptive immunity and increases the severity of LD [19]. Furthermore, human coinfection with Ba. microti and B. burgdorferi s.l. seems to be serious clinical problem because of the difficulties in diagnosis and treatment, since the antibiotics used to treat borreliae are ineffective against Ba. microti [20]. In Europe, most cases of human babesiosis are attributed to Ba. divergens, usually a cattle parasite or less frequently to Ba. venatorum for which the roe deer is the main reservoir host. Interestingly, European genotypes of Ba. microti infecting humans are not as infectious or pathogenic than those in the USA [21].
The aim of our study was to ascertain the co-occurrence of B. burgdorferi s.l. spirochetes and Babesia piroplasms in I. ricinus ticks collected from vegetation as well as from dogs and cats in urban areas of the city of Poznań, west-central Poland.
2. Materials and Methods
2.1. Tick Collection and Identification, and DNA Extraction
Host-seeking ticks were collected by sweeping up the vegetation up to 1 m with a 1 m2 flannel flag along the paths in five forested green areas used for recreational activities and as walking areas for dogs in the city of Poznań. Ticks were collected from May to September 2017 and in April 2018. The five study sites included: Morasko Adam Mickiewicz University Campus, Sołacki and Citadel city parks, two urban forests localized around Rusałka and Malta lakes, for details see [22].
Ticks feeding on dogs and cats, were collected during a three-year survey (April to October 2015, March to November 2016, and March to September 2017) in 17 veterinary clinics in Poznań (for details see [22]). In this study, we tested ticks derived from animals which did not travel outside the city, based on information from questionnaires.
All ticks were preserved in 96% ethanol until DNA extraction. Adult ticks were identified to the species level using morphological characters [23]. Ixodes nymphs were identified using DNA-barcoding based on cytochrome c oxidase subunit I (COI) amplification and next generation sequencing (NGS) of the amplicons [22,24]. Ticks were tested individually. DNA extraction from host-seeking ticks was performed with the ammonium hydroxide method, whereas DNA from pet-derived ticks were isolated using a silica-column method. Details concerning DNA extractions are described by Liberska et al. [22].
2.2. Screening for Borrelia burgdorferi s.l. DNA
Host-seeking ticks were initially screened for B. burgdorferi s.l. DNA by amplification and sequencing of the V4 hypervariable region of the 16S rRNA gene (V4 16S). Details concerning PCR primers, library construction, NGS sequencing, and sequence data analysis were described previously [25]. Host-seeking and feeding ticks positive for B. burgdorferi s.l. V4 16S were retested by amplification and sequencing the flaB gene fragment using two primer sets 132f/905r and 220f/823r [26]. Protocols for PCR reactions and Sanger sequencing followed [25].
2.3. Screening for Babesia spp. DNA
Babesia spp. DNA was detected by amplification and sequencing of the 18S rRNA gene fragment using nested PCR and Sanger sequencing or by next generation sequencing (NGS) of the same target DNA. For the nested PCR, we used the RIB-19/RIB-20 primer set for the first-round reaction [27], and the P3/BabR3 primer set for the second-round reaction [22,28]. Primer sequences and protocols for nested PCR and Sanger sequencing are described by Liberska et al. [22].
The second approach for the detection of Babesia DNA was conducted with the use of the P3/BabR3 primer set fused with dual-indexed Ion Torrent adapters to generate Babesia-specific amplicons for NGS sequencing. Details concerning library construction, Ion Torrent sequencing, and sequence data analysis were described previously [25].
2.4. Statistical Analysis
Data analysis was performed using STATISTICA software version 6.0 (StatSoft Inc., Tulsa, OK, USA). Rates of infection were analysed using Chi-squared test χ2 and Fisher exact test. A p < 0.05 was considered statistically significant.
2.5. Sequence Analysis
Sanger sequence chromatograms were checked for accuracy in GeneiousR11.1.5 (Biomatters Ltd.) and contigs were assembled manually in GeneDoc sequence editing tool version 2.7 [29]. Amplicon sequence variants (ASVs) and operational taxonomic units (OTUs) obtained from NGS data were compared to those available in the GenBank using BLASTN (https://blast.ncbi.nlm.nih.gov) and the megablast algorithm. We used 100% identity threshold to determine Borrelia and Babesia species because they were determined from rDNA sequences. The identity threshold for tick species determination was lowered to 97% due to intra-species variability of the COI marker.
2.6. Phylogenetic Analysis
For phylogenetic analysis, we used 49 V4 16S sequences (Table S1) including ASVs found in this study (n=18), V4 16S sequences identical to the ASVs found in this study and sourced from reference B. burgdorferi s.l. genomes deposited in GenBank (n=18), and representants of Borrelia RF group (n=13) used to root the tree. Sequences were aligned manually in GeneDoc and a phylogenetic tree was built using FastTree 2.1.11 as implemented in Geneious Prime 2023.2.1 (Biomatters Ltd.). The tree was edited in Mega7 [30] and Corel Draw v. X5.
3. Results
3.1. Collection and Identification of Ticks
A total of 2297 I. ricinus ticks including 1029 host-seeking individuals (460 nymphs, 289 females, and 280 males) and 1268 feeding female ticks removed from 1115 tick-infested companion animals, were collected during this study (Table 1 and Table 2). In the case of pet-derived ticks sampled in veterinary clinics, 711 were removed from 609 dogs, 153 from 117 cats, and 404 from 389 undefined animals, i.e., without giving information about host species (Table 2).
3.2. Detection of Borrelia burgdorferi s.l. DNA
Based on V4 16S and flaB gene fragments, DNA of B. burgdorferi s.l. was detected in 90 (8.7%) of the 1,059 host-seeking I. ricinus ticks. The bacterium was found in nymphs, males, and females with prevalences of 6.3%, 9.6% and 11.8%, respectively (Table 1). Four out of the five collection sites yielded ticks that tested positive for Borrelia spirochetes (range between 3.0% and 40.6%). Sequencing of the amplified flaB gene fragments revealed four Borrelia species: B. afzelii, B. garinii, B. lusitaniae, and B. valaisiana (Table 1). The first two prevailed and reached the same prevalence of 3.7%, followed by B. lusitaniae (0.9%), B. valaisiana (0.3%) and Borrelia undetermined species (0.2%).
3.3. Genetic Diversity of B. burgdorferi s.l. V4 16S Amplicon Sequence Variants
In total, 18 different B. burgdorferi s.l. V4 16S ASVs were found in questing ticks (Supplementary table with GenBank acc. nos.). For most ASVs, identical sequences in species reference genomes published in GenBank could be assigned (Figure V4 Tree). Phylogenetic analysis enabled the assignment of one of the previously unknown variants, ASV05, to B. afzelii with a relatively high support (85%). Another new variant, ASV17, was reconstructed as a sister to the V4 16S sequence from the B. maritima reference genome, although with moderate support (76%). The remaining four new variants (ASV10-12 and ASV14) were reconstructed by the phylogenetic analysis as a separate, well supported clade (90%). These ASVs tended to co-occur with other B. burgdorferi s.l. V4 16S ASVs either in a relatively large number of ticks (ASV10 and ASV 14 in nine positive samples each) or in a predominant number of reads (ASV11). Amplicon sequence variant 12 was found in only one sample and in co-occurrence with ASV14; however, both sequences differed at two variable nucleotide positions, indicating that ASV12 cannot be considered an NGS artifact of the ASV14 amplicon.
Figure 1.
Phylogenetic analysis of amplicon sequence variants (ASV) of the V4 16S rRNA gene found in this study (ASV1-ASV19 ). The tree was constructed by FastTree method as described in Materials and Methods.
Figure 1.
Phylogenetic analysis of amplicon sequence variants (ASV) of the V4 16S rRNA gene found in this study (ASV1-ASV19 ). The tree was constructed by FastTree method as described in Materials and Methods.
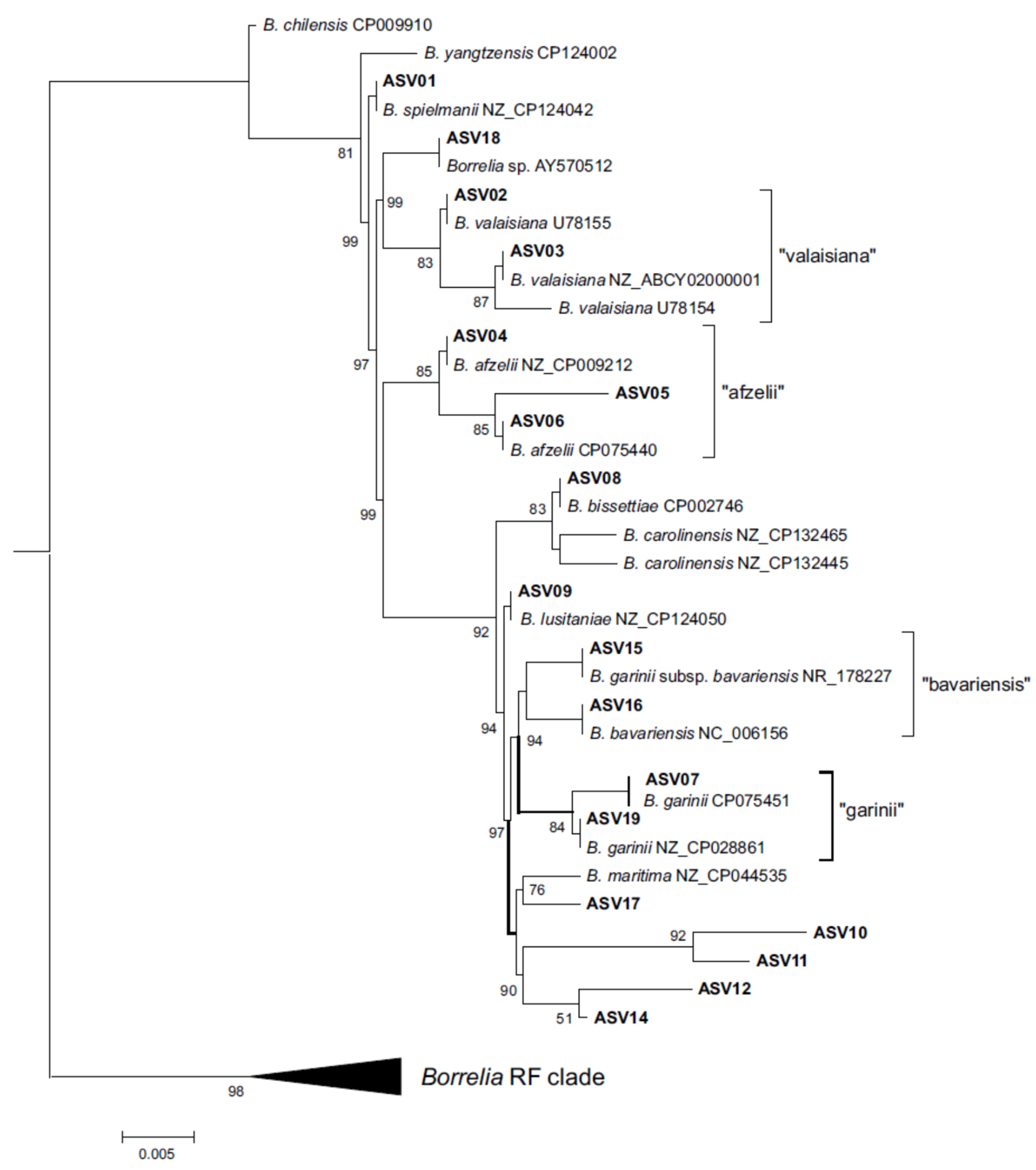
Analysis of the flaB gene sequence was unable to unambiguously assign most of the V4 16S ASVs to the species. Samples positive for B. burgdorferi s.l. V4 16S DNA that clustered basally at our phylogenetic tree (ASV01-03 and ASV18), failed in PCR amplification of the flaB gene fragment. Moreover, most of the ASVs occurred in co-infections, making it impossible to assign a specific flaB gene sequence to a specific V4 16S ASV. However, in these cases, the flaB gene results did not exclude the presence of the species identified in by the V4 16S analysis in the analyzed samples. This also applied to the clade grouping of the previously unknown V4 16S variants (ASV10-12, ASV14). In this case, the flaB gene sequences indicated the presence of B. garinii and B. lusitaniae. In this regard, due to the presence of B. burgdorferi s.l. V4 16S sequence variants that could not be unambiguously assigned to the species, further analyses were based on sequencing data from flaB amplicons.
3.3. Presence of Babesia spp. DNA in Ixodes ricinus
The overall prevalence of Babesia spp. in host-seeking I. ricinus ticks was 4.4% (45/1,029), and the infection was noticed in all developmental stages (Table 1), with the highest prevalence found in females and the lowest in nymphs (7.3% vs. 2.4%; respectively; χ2 test: P = 0.001). Babesia infected ticks occurred in four out of five locations (range between 0.8% and 9.8%). Among three identified Babesia species, Ba. canis (2.6%) followed by Ba. microti (1.4%) were the most prevalent, and Ba. venatorum (0.4%) was the rarest species. These three species were noted in adult females and males, whereas Ba. venatorum was absent in nymphs.
In total, 6.0 % (76/1,268) feeding female I. ricinus ticks were positive for the presence of Babesia DNA (Table 2). The highest infection rate was noted in ticks collected from dogs (6.8%) and undefined animals (5.7%), and the lowest, in cat-derived ticks (3.9%), but the difference was statistically insignificant. Three Babesia species were identified: two of them, Ba. canis and Ba. microti, reached similar infection rates (2.8% and 2.2%, respectively), whereas Ba. venatorum, was the rarest species (1.0%). Ticks infected with these three species were confirmed in all animal groups. The infection levels of Ba. canis and Ba. microti in ticks derived both from dogs and undefined pets were similar (3.2%, 2.5% and 2.7%, 2.0%, respectively), with the lowest level for Ba. venatorum (1.0% and 0.7%, respectively). The latter prevailed (2.0%) in cat-derived ticks, followed by Ba. microti (1.3%) and Ba. canis (0.7%) that was found only in one female tick.
All obtained Ba. canis sequences demonstrated 100% similarity to Ba. canis isolates detected in the blood of infected dogs from Poland (e.g., GenBank: EU622793, KT844903), as well as in questing I. ricinus ticks (GenBank: MF797820). Babesia microti sequences were identical to the zoonotic Ba. microti "Jena/Germany" genotype (GenBank acc. no. EF413181), whereas Ba. venatorum sequences showed 100% homology to isolates found in patients in Italy and Austria (AY046575), and also in an asymptomatic patient in Poland (KP072001).
3.4. Co-occurrence of Borrelia burgdorferi s.l. and Babesia spp. DNA in Ixodes ricinus
Overall, of the 2,297 I. ricinus ticks, 10.7% (n=246) tested positive for a single pathogen, and 1.0% (n=24) yielded dual infections. Of the 149 positive for B. burgdorferi s.l. DNA ticks, mono-infections were identified in 83.8% (n=125) and co-occurrence with Babesia spp. DNA in 16.1% ticks. Of the 121 ticks infected with Babesia species, 80.2% (n=97) were single infections, and 19.8% were co-infected with B. burgdorferi s.l. Among host-seeking ticks, co-occurrence of Borrelia and Babesia was identified in 1.1% (11/1,029) of adult ticks. None of the 460 nymphs was concurrently infected with both pathogens (Tab. 1). Babesia canis was the most prevalent species in co-infected ticks (72.7%; 8/11), and was confirmed in four (0.4%) co-infections with B. afzelii and in four (0.4%) with B. garinii. In the remaining three female ticks, B. lusitaniae co-occurred with Ba. venatorum (n=2; 0.2%) and Ba. microti (n=1; 0.1%).
Among feeding female ticks, 1.0% (13/1,268) were co-infected with both pathogens. Babesia microti was the most prevalent species in all co-infections (92.3%; 12/13). Most co-infections of this species was recorded with B. afzelii (10/13). Furthermore, Ba. microti co-occurred with B. spielmanii and B. garinii in two other ticks, and Ba. canis co-occurred with B. garinii only in one female tick.
3.5. Co-occurrence of Borrelia burgdorferi s.l. and Borrelia miyamotoi in Ixodes ricinus
Of the 90 positive for B. burgdorferi s.l. host-seeking ticks, six (6.7%) were co-infected with B. miyamotoi spirochetes identified in our previous study [25]. Borrelia afzelii was the most common and occurred in three coinfections (2 nymphs and one male), followed by B. garinii (2 nymphs) and B. lusitaniae (one female). None of 59 B. burgdorferi s.l. positive feeding ticks, yielded concurrently B. miyamotoi.
4. Discussion
In this study, we described prevalence of B burgdorferi s.l. and Babesia spp. found in mono- and double infections among I. ricinus ticks occurring in urban areas of the city of Poznań. We focused on two tick groups: (i) a group of host-seeking collected from vegetation, and (ii) a group of feeding ticks removed from pet animals, dogs and cats.
According to a review by Hansford et al. [31], the mean Borrelia prevalence in questing ticks in urban green areas across Europe was 17.3% (range: 3.1% to 38.1%). In our study, 8.7% of ticks (range: 0.0% to 40.6%) yielded borreliae. Comparable or higher mean infection rates were found in urban I. ricinus populations of several European cities from neighboring countries. For example, in Slovakia, the prevalence of B. burgdorferi s.l. ranged from 6.8% to 15.3% in parks of Bratislava, [32,33], and in agglomerations of Košice and Bardejov reached 10.2% [34]. In the Czech Republic, 12.1% of ticks collected in parks in Brno and 13.2% in Ostrava city, yielded spirochetes [35,36]. Furthermore, in the city parks of Vilnius, Lativa, and in recreational areas of Hanover, Germany, 25% of ticks yielded B. burgdorferi s.l. [37,38]. Comprehensive studies on host-seeking ticks infected with B. burgdorferi s.l. in strictly urban agglomerations in Poland, are relatively rare. In city forests and parks of Warsaw, the mean prevalence of Borrelia spirochaetes was 10.9% [6], and in the Tri-City agglomeration area of Gdańsk, Gdynia, and Sopot, reached 12.4% [39]. To date, the highest levels of Borrelia infections have been described in green areas of the city Białystok (25.7%) and Olsztyn (27.4%) [40,41]. In our study, the highest prevalence of 40.3% (28/69) was recorded in ticks of the Citadel Park, the largest municipal park strictly in the center of the city. Although, the number of tested ticks was limited, this high infection level, may result from the lack of roe deer, that is considered to eliminate B. burgdorferi s.l. in feeding ticks [42].
The overall prevalence of B. burgdorferi s.l. found in ticks from pet animals (4.7%), was almost two-fold lower than in host-seeking ticks (8.7%). Comparable prevalences were described in the Netherlands [43,44] and Austria (4.8% and 5.2%, respectively) [45]. Higher infection rates in I. ricinus mainly from dogs, were recorded in Lativa (10.7%), Germany (11.6%), Finland (11.8%), Norway (14%), and Denmark (15%) [46,47,48,49,50]. So far, the highest prevalences in I. ricinus from dogs have been found in two city agglomerations: Wrocław (21.7%), in the southwest [51] and Olsztyn (34.4%), in the northeast of Poland [52].
Borrelia afzelii and B. garinii proved to be the most prevalent species among infected ticks both from vegetation and pets. Our results are in agreement with the frequency of main Borrelia species infecting host-seeking I. ricinus ticks in central Europe [53] and with a report by Skotarczak [54], documenting that B. afzelli and B. garinii are the most common species in dogs. Dogs can develop infection of B. burgdorferi s.l. and exhibit the presence of antibodies, but unlike humans, they rarely get sick [55]. The distribution of Borrelia species in our study, may be explained by species composition of wild hosts. The predominant B. afzelii, together with B. spielmanii, are maintained in enzootic cycles associated with small rodents and medium-sized mammals, including hedgehogs [56,57,58]. We found B. spielmanii only in three feeding females, what confirms the rarity and highly focal distribution of this spirochete. The mentioned groups of mammals were observed in our study locations, with predominance of small rodents. In the present research, B. afzelii prevailed in feeding ticks (3.7%). This spirochete was also the most prevalent (3.6%) followed by B. garinii (1.7%), B. valaisiana (1.4%) and B. spielmanii (1.4%) in ticks from dogs in Lativa [49]. A reverse pattern, with predominance of B. garinii over B. afzelii (28.1% vs. 3.2%), was observed in ticks from dogs in north-eastern Poland [52]. Avian associated B. garinii and B. valaisiana [59] were present in both infected tick groups, with almost a fourfold higher prevalence of B. garinii in ticks from vegetation in comparison to feeding ticks (42.2% vs. 11.2%). The lizard-associated B. lusitaniae [60] prevailed in host-seeking ticks compared to pet-derived ticks (10% vs. 1.7%). The finding of this spirochete only at the Rusałka lake, suggests its highly focal distribution.
In this study, the method of amplification and sequencing of the V4 region in 16S rRNA was not specific enough to clearly identify the obtained B. burgdorferi s.l. sequences to species as displayed on the phylogenetic tree. The 16S rRNA region had different sequences of V4 for some species identified by flaB gene, therefore the complex analysis, including other marker genes is necessary to correctly assign 16S rDNA sequences to the species.
We detected Babesia DNA in 4.4% of host-seeking ticks, which were recorded in four out of five locations (range: 0.8% to 9.8%). The meta-analysis by Onyiche et al. [61] estimated the overall prevalence of babesiae in questing I. ricinus in Europe at 2.1%. Our results agree with infection rates found in the city Białystok (3.7%) [41] and within the Tri-City agglomeration (4.5%) [62]. Lower prevalences (range: 0.4% to 0.5%) were reported in ticks tested in Bavarian public parks [63] or in urban Bielański Forest (0.8%), in Warsaw [64].
Babesia DNA was identified in 6.0% of feeding ticks, with a higher prevalence in ticks from dogs (6.8%) compared to those from cats (3.9%). Stensvold et al. [48] documented a prevalence of 8.0% in dog-derived ticks in Denmark. Lower prevalences of Babesia spp. in pet-derived ticks (usually from dogs) were 0.8% in the Netherlands [44], 1.0% in Finland [50], 1.4% (62/4316) in the United Kingdom [65], 2.5% in Germany [47], and 4.7% in Latvia [49]. The highest infection rates in ticks feeding upon dogs (66.8%) and cats (15.4%) were reported in southern Poland [66].
In Europe, I. ricinus is involved in the transmission cycles of Ba. divergens, Ba. venatorum, Ba. microti, and Ba. capreoli, of which the first three are considered as human pathogens [8]. In our study, three species including Ba. microti, Ba. venatorum and Ba. canis were identified in both groups of ticks. The finding of the first two species, might be explain the availability of ticks to suitable reservoir hosts of both parasites. In Europe, Ba. microti infects small rodents [67]. The reservoir host for Ba. venatorum, is the roe deer, and this species was observed in at least four out of five study sites. However, we did not find Ba. divergens, the main etiological agent of human babesiosis in European patients [21]. Since cattle are regarded as the reservoir hosts for Ba. divergens, their absence in urban areas of Poznań, may explain that we failed to find the pathogen. In our study, Ba. canis and Ba. microti were the most prevalent species in feeding (2.6% and 1.4%, respectively) and host-seeking ticks (2.8% and 2.2%, respectively), whereas Ba. venatorum was the rarest piroplasm. Although, the role of I. ricinus in the transmission of Ba. canis remains still unclear, this parasite has already been found in questing ticks in northern Poland (range: 0.2% to 5.8%) [40,62,68], in Slovakia and the Czech Republic (range; 0.2% to 1.4%) [69,70]. In our previous study, we found that 10.2% of ticks in forest ecosystems in west-central Poland were positive for Ba. canis DNA, the highest prevalence ever reported for this tick [22]. European Ba. canis strains belong to the type A or type B, which induce different clinical symptoms of canine babesiosis [71]. In this previous and the current study, all Ba. canis obtained sequences represented the milder type A. In Europe, Ba. canis infections in I. ricinus from dogs were previously documented only in Latvia and Russia, with prevalences of 1.0% and 3.8%, respectively [49,72], and a prevalence of 3.2% in our study is comparable with those data. In Europe, most Babesia infections in I. ricinus collected from companion animals are caused by Ba. microti and Ba. venatorum. The overall prevalence of 2.2% of Ba. microti in feeding ticks in our study was lower compared to a 5.7% infection rate in dog-derived ticks in Warsaw which yielded Ba. microti Gray strain pathogenic to humans (GenBank: AY693840) [73]. In Europe, the highest prevalences of Ba. microti in this group of ticks, were reported in southeastern (42.6%) and southern (24.3%) Poland [66,74]. In the case of Ba. venatorum, the highest prevalence of 1.6% was recorded in dog-derived ticks in Lativa [49]. According to Onyiche et al. [61], Ba. microti is considered to be the most common Babesia species in questing I. ricinus in the eastern Europe. In our study, Ba. microti (1.4%) was the second most prevalent species following Ba. canis. Much higher infection rates of Ba. microti in ticks from urban areas were 2.6% in northern [39], 6.5% in central [75], and 26.4% in southern Poland [76]. Babesia venatorum that infected only 0.4% of host-seeking ticks in our study as well as in Bavarian public parks [63], reached also low infection rates of 0.8% in central [64] and 0.9% in northern Poland [68].
In Europe, more than 60 cases of human babesiosis have been caused mainly by Ba. divergens, with five cases attributed to Ba. venatorum and eleven autochthonous cases to Ba. microti, most of which were identified in Poland [21,77]. Two European genotypes of Ba. microti can infect humans, but are less infectious or pathogenic to humans as those in the United States. The first Ba. microti genotype (strain) "Jena/Germany" (EF413181) is closely related to the USA genotypes (Clade 1) including e.g., the Gray strain isolated from a patient in Massachusetts (AY693840), responsible for most cases of human babesiosis worldwide. The second Ba. microti genotype, the ‘Munich’ type (AB071177) is widely distributed in Europe and belongs to Clade 3 [78]. This genotype was originally thought to be non-zoonotic, however, it has recently been identified in six patients with mild and asymptomatic infections in Poland [79] and in one from Spain [80]. However, its pathological potential remains still uncertain [77]. It should be noted, that Ba microti sequences identified in our study, were identical to Ba. microti genotype "Jena/Germany” that is considered as pathogenic to humans [77]. In Poland, Welc-Falęciak et al. [81] found the same genotype in two asymptomatic patients. Furthermore, among 1.3% of Babesia infected I. ricinus removed from humans in Poland, most showed a high similarity (> 99%) to the Ba. microti strain Jena followed by Ba. venatorum [82]. Of note, that the sequences of Ba. venatorum found in the two mentioned study as well as in our, were identical with those isolated from splenectomised patients in Italy and Austria [83], and with an asymptomatic patient from Poland [84].
Altogether, out of 121 Babesia-positive ticks, 19.8% were simultaneously Borrelia-positive. We found almost identical prevalences of co-infections with B. burgdorferi s.l. and Babesia spp. in host-seeking (1.1%) as well as in feeding ticks (1.0%). To the best of our
knowledge, this is the first report documenting the co-occurrence of both pathogens in ticks from companion animals in Europe. In Poland, coinfection rates of both pathogens in host-seeking ticks were described in the north (range; 0.3% to 0.6%) [39,85], in the east (1.6%) [86], and in the northeast (2.8%) [41]. In the present study, Ba. canis prevailed in coinfections mostly with B. afzelii and B. garinii in host-seeking ticks, whereas Ba. microti dominated predominantly with B. afzelii, in ticks from the pets. The prevalence of 0.9% (n=9) Ba. canis and B. burgdorferi s.l. coinfections in host-seeking ticks in our study, is consistent with a previous Polish study, in which 1.0% of 104 ticks showed a coinfection of Ba. canis with B. afzelii [87]. Recently, a fatal case of a dog co-infected with Ba. canis and B. burgdorferi s.l. was diagnosed in Romania [88]. Hildebrand et al. [89], found 1.6% of coinfections Ba. microti followed by Ba. divergens, mostly with B. afzelii and B. garinii in questing ticks in Middle Germany, whereas Ba. microti and B. afzelii prevailed in co-infected ticks from humans [82].
Taking into account the mentioned studies, in host-seeking I. ricinus, Ba. microti seems to be the most prevalent species in co-infections with B. afzelii and B. garinii. Co-occurrence of these pathogens is of significant importance from a medical point of view. In humans, they may affect the clinical course of disease, especially in non-immunocompetent patients, and might be difficult to differentiate since both infections induce often nonspecific symptoms including fever, fatigue, and flu-like illness [90,91]. Jabłońska et al. [92] described a symptomatic case of babesiosis and LB in a Polish immunocompetent patient after travelling to Canada and the USA. Furthermore, in a study of 24 tick-exposed individuals from southeastern Poland, a piroplasm 98.9% homologous with Ba. divergens and Ba. venatorum was detected in one person concurrently seropositive for B. burgdorferi s.l. [93]. In another Polish report, Pańczuk et al. [94] also found a single B. burgdorferi s.l. and Ba. microti coinfection among foresters with significant levels of IgG anti-B. burgdorferi antibodies.
Using our previous results regarding the presence of B. miyamotoi spirochetes [25] in the same group of host-seeking ticks, we found that 0.6% of 1,029 individuals yielded concurrently DNA of B. burgdorferi s.l., with predominance of B. afzelii and B. garinii. Co-occurrence of both pathogens was also documented in 0.4% and 1.4% of adult I. ricinus collected from vegetation in France [95] and eastern Poland [86], respectively.
In conclusion, the finding of a predominance of two pathogenic species of spirochetes, B. afzelii and B. garinii, and the detection in both groups of ticks of the sequences Ba. microti and Ba. venatorum, which are identical to human pathogenic strains, indicate that people using urban green spaces are at risk of contracting LB and/or babesiosis. The occurrence of double infections, even though with a low prevalence of 1.0%, is both clinically and epidemiologically significant and indicates that they pose a challenge for differential diagnosis in patients with acute febrile disease after contact with a tick. Furthermore, the finding of Ba. canis in both groups of I. ricinus suggests that this species could potentially be involved in the circulation of this piroplasm in areas of western central Poland, where the competent vector, Dermacentor reticulatus is absent or rare. If this is the case, urban areas may pose a potential risk of dogs acquiring babesiosis.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Funding
This work was partially supported by the COST Action CA21170 “Prevention, anticipation and mitigation of tick-borne disease risk applying the DAMA protocol (PRAGMATICK)”.
Acknowledgements
We would like to thank Urszula Sobczyńska, Łukasz Broda, Weronika Pawlik, and Aneta Łyś of Adam Mickiewicz University in Poznań for their assistance in the collection of ticks from urban ecosystems. We would like to thank all the veterinary clinics involved in the study: Centrum Zdrowia Małych Zwierząt M. Majka, R. Starczewski s.c., Gabinet Weterynaryjny Katarzyna Gryglewska, Gabinet Weterynaryjny lek. wet. Dariusz Filipiński, Gabinet Weterynaryjny Marcin Konieczny, Gabinet weterynaryjny - Piotr Winiecki, Klinika Weterynaryjna Juszczak & Gorzelańczyk, Praktyka Weterynaryjna A. Golec M. Rybski, Gabinet Weterynaryjny PLUTO VET lek wet Krzysztof Kaniewski, Zwierzak Gabinet Weterynaryjny A. Adamska, AMICUS Gabinet Weterynaryjny, Usługi Weterynaryjne Jakub Golec, Przychodnia Weterynaryjna Na Polance Katarzyna Marciniak, Specjalistyczny Gabinet Weterynaryjny lek. wet. Żaneta Sokołowska, Przychodnia Weterynaryjna lek. wet. Adam Łowczak, Przychodnia Weterynaryjna Kociak lek. wet. Sylwia Łata, Klinika Weterynaryjna dr. Grzegorza Wąsiatycza.
References
- Rizzoli, A.; Silaghi, C.; Obiegala, A.; Rudolf, I.; Hubálek, Z.; Földvári, G.; Plantard, O.; Vayssier-Taussat, M.; Bonnet, S.; Śpitalská, E.; et al. 2014. Ixodes ricinus and its transmitted pathogens in urban and peri-urban areas in Europe: new hazards and relevance for public health. Front. Public Health. 2014, 2, 1–26. [Google Scholar] [CrossRef]
- Noden, B.H.; Roselli, M.A.; Los, S.R. Effect of urbanization on presence, abundance, and coinfection of bacteria and protozoa in ticks in the US Great Plains. J. Med. Entomol. 2022, 59, 957–968. [Google Scholar] [CrossRef]
- Ginsberg, H.S. Potential effects of mixed infections in ticks on transmission dynamics of pathogens: Comparative analysis of published records. Exp. Appl. Acarol. 2008, 46, 29–41. [Google Scholar] [CrossRef]
- Moutailler, S.; Valiente Moro, C.; Vaumourin, E.; Michelet, L.; Tran, FH.; Devillers, E.; et al. Co-infection of ticks: The rule rather than the exception. PLoS Negl Trop Dis. 2016, 10, e0004539. [Google Scholar] [CrossRef] [PubMed]
- Welc-Falęciak, R.; Kowalec, M.; Karbowiak, G.; Bajer, A.; Behnke, J.M.; Siński, E. Rickettsiaceae and Anaplasmataceae infections in Ixodes ricinus ticks from urban and natural forested areas of Poland. Parasites Vectors 2014, 7, 121. [Google Scholar] [CrossRef]
- Kowalec, M.; Szewczyk, T.; Welc-Falȩciak, R.; Siński, E.; Karbowiak, G.; Bajer, A. Ticks and the city—are there any differences between city parks and natural forests in terms of tick abundance and prevalence of spirochaetes? Parasites Vectors 2017, 10, 573. [Google Scholar] [CrossRef] [PubMed]
- Richtrová, E.; Míchalová, P.; Lukavská, A.; Navrátil, J.; Kybicová, K. 2022. Borrelia bugdorferi sensu lato infection in Ixodes ricinus ticks in urban green areas in Prague. Ticks Tick Borne Dis. 2022, 13, 102053. [Google Scholar] [CrossRef]
- Bajer, A.; Dwużnik-Szarek, D. The Specificity of Babesia-tick vector interactions: Recent advances and pitfalls in molecular and field studies. Parasites Vectors 2021, 14, 507. [Google Scholar] [CrossRef] [PubMed]
- Krause, P.J.; Spielman, A.; Telford, S.R., 3rd; Sikand, V.K.; McKay, K.; Christianson, D.; Pollack, R.J.; Brassard, P.; Magera, J.; Ryan, R.; et al. Persistent parasitemia after acute babesiosis. N. Engl. J. Med. 1998, 339, 160–165. [Google Scholar] [CrossRef]
- Dunn, J.M.; Krause, P.J.; Davis, S.; Vannier, E.G.; Fitzpatrick, M.C.; Rollend, L.; Belperron, A.A.; States, S.L.; Stacey, A.; Bockenstedt, L.K.; et al. Borrelia burgdorferi promotes the establishment of Babesia microti in the northeastern United States. PLoS One 2014, 9, e115494. [Google Scholar] [CrossRef]
- Little, E.A.H.; Molaei, G. Passive tick surveillance: exploring spatiotemporal associations of Borrelia burgdorferi (Spirochaetales: Spirochaetaceae), Babesia microti (Piroplasmida: Babesiidae), and Anaplasma phagocytophilum (Rickettsiales: Anaplasmataceae) infection in Ixodes scapularis (Acari: Ixodidae). Vector Borne Zoonotic Dis. 2020, 20, 177–186. [Google Scholar] [CrossRef]
- Zembsch, T.E.; Lee, X.; Bron, G.M.; Bartholomay, L.C.; Paskewitz, S.M. Coinfection of Ixodes scapularis (Acari: Ixodidae) nymphs with Babesia spp. (Piroplasmida: Babesiidae) and Borrelia burgdorferi (Spirochaetales: Spirochaetaceae) in Wisconsin. J Med Entomol. 2021, 58, 1891–1899. [Google Scholar] [CrossRef] [PubMed]
- Hersh, M.H.; Ostfeld, R.S.; McHenry, D.J.; Tibbetts, M.; Brunner, J.L.; Killilea, M.E.; LoGiudice, K.; Schmidt, K.A.; Keesing, F. Co-infection of blacklegged ticks with Babesia microti and Borrelia burgdorferi is higher than expected and acquired from small mammal hosts. PLoS ONE 2014, 9, e99348. [Google Scholar] [CrossRef]
- Diuk-Wasser, M.A.; Vannier, E.; Krause, P.J. Coinfection by Ixodes tick-borne pathogens: ecological, epidemiological, and clinical consequences. Trends Parasitol. 2015, 32, 30–42. [Google Scholar] [CrossRef]
- Jahfari, S.; Hofhuis, A.; Fonville, M.; van der Giessen, J.; van Pelt, W.; Sprong, H. Molecular detection of tick-borne pathogens in humans with tick bites and erythema migrans, in the Netherlands. PLoS Neglected Trop. Dis. 2016, 10, e0005042. [Google Scholar] [CrossRef] [PubMed]
- Cutler, S.J.; Vayssier-Taussat, M.; Estrada-Peña, A.; Potkonjak, A.; Mihalca, A.D.; Zeller, H. Tick-borne diseases and co-infection: Current considerations. Ticks Tick Borne Dis. 2021, 12, 101607. [Google Scholar] [CrossRef] [PubMed]
- Krause, P.J.; Telford, S.R., III; Spielman, A.; Sikand, V.; Ryan, R.; Christianson, D.; Burke, G.; Brassard, P.; Pollack. , et al. Concurrent Lyme disease and babesiosis: Evidence for increased severity and duration of illness. JAMA 1996, 275, 1657–1660. [Google Scholar] [CrossRef]
- Martínez-Balzano, C.; Hess, M.; Malhotra, A.; Lenox, R. Severe babesiosis and Borrelia burgdorferi co-infection. QJM Int. J. Med. 2015, 108, 141–143. [Google Scholar] [CrossRef]
- Djokic, V.; Akoolo, L.; Primus, S.; Schlachter, S.; Kelly, K.; Bhanot, P.; Parveen, N. Protozoan parasite Babesia microti subverts adaptive immunity and enhances Lyme disease severity. Front. Microbiol. 2019, 10, 1596. [Google Scholar] [CrossRef]
- Parveen, N.; Bhanot, P. Babesia microti—Borrelia burgdorferi coinfection. Pathogens 2019, 8, 117. [Google Scholar] [CrossRef]
- Hildebrandt, A. , Zintl, A., Montero, E., Hunfeld, K.P., Gray, J. 2021. Human babesiosis in Europe. Pathogens 2021, 10, 1165. [Google Scholar] [CrossRef] [PubMed]
- Liberska, J.; Michalik, J.; Pers-Kamczyc, E.; Wierzbicka, A.; Lane, R. S.; Rączka, G.; Opalińska, P.; Skorupski, M.; Dabert, M. Prevalence of Babesia canis DNA in Ixodes ricinus ticks collected in forest and urban ecosystems in west-central Poland. Ticks Tick Borne Dis. 2021, 12, 101786. [Google Scholar] [CrossRef] [PubMed]
- Siuda, K. Kleszcze (Acari: Ixodida) Polski. II. Systematyka i rozmieszczenie. Monografie Parazytologiczne vol, 12. Polish Parasitological Society, Warsaw, Poland 1993.
- Trzebny, A.; Slodkowicz-Kowalska, A.; Becnel, J.J.; Sanscrainte, N.; Dabert, M. A new method of metabarcoding Microsporidia and their hosts reveals high levels of microsporidian infections in mosquitoes (Culicidae). Mol. Ecol. Resour. 2020, 20, 1486–1504. [Google Scholar] [CrossRef]
- Liberska, J.A.; Michalik, J.F.; Dabert, M. Exposure of dogs and cats to Borrelia miyamotoi infected Ixodes ricinus ticks in urban areas of the city of Poznań, west-central Poland. Ticks Tick Borne Dis. 2023, 14, 102188. [Google Scholar] [CrossRef]
- Wodecka, B.; Leońska, A.; Skotarczak, B. A comparative analysis of molecular markers for the detection and identification of Borrelia spirochetes in Ixodes ricinus. J. Med. Microbiol. 2010, 59, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Zahler, M.; Rinder, H.; Schein, E.; Gothe, R. Detection of a new pathogenic Babesia microti-like species in dogs. Vet Parasitol. 2000, 89, 241–248. [Google Scholar] [CrossRef]
- Li, Y.; Teng, L.; Shang, L.; Ma, R.; Cai, J.; Zhao, Q.; Li, J.; Liu, Q. Detection of Theileria and Babesia sp. in Ixodid ticks from Qinghai Province, Northwestern China. J Anim Vet Adv. 2013, 12, 775–778. [Google Scholar] [CrossRef]
- Nicholas, K.B.; Nicholas, H.B. Jr. GeneDoc: A tool for editing and annotating multiple sequence alignments. Pittsburgh Supercomputing Center’s National Resource for Biomedical Supercomputing. 1997. Available online: http://www.nrbsc. org/downloads/ (accessed on 2 March 2007).
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol Biol Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Hansford, K.M.; Wheeler, B.W.; Tschirren, B.; Medlock, J.M. Questing Ixodes ricinus ticks and Borrelia spp. in urban green space across Europe: A review. Zoonoses Public Health, 2022, 69, 153–166. [Google Scholar] [CrossRef]
- Chvostáč, M.; Špitalská, E.; Václav, R.; Vaculová, T.; Minichová, L.; Derdáková, M. Seasonal patterns in the prevalence and diversity of tick-borne Borrelia burgdorferi sensu lato, Anaplasma phagocytophilum and Rickettsia spp. in an urban temperate forest in south western Slovakia. Int. J. Environ. Res. Public Health 2018, 15, 994. [Google Scholar] [CrossRef]
- Vaculová, T.; Derdáková, M.; Špitalská, E.; Václav, R.; Chvostáč, M.; Rusňáková Tarageľová, V. Simultaneous occurrence of Borrelia miyamotoi, Borrelia burgdorferi sensu lato, Anaplasma phagocytophilum and Rickettsia helvetica in Ixodes ricinus ticks in urban foci in Bratislava, Slovakia. Acta Parasitol. 2019, 64, 19–30. [Google Scholar] [CrossRef]
- Pangrácová, L.; Derdáková, M.; Pekárik, L.; Hviščová, I.; Víchová, B.; Stanko, M.; Hlavatá, H.; Peťko, B. Ixodes ricinus abundance and its infection with the tick-borne pathogens in urban and suburban areas of Eastern Slovakia. Parasites Vectors 2013, 16, 238. [Google Scholar] [CrossRef]
- Pejchalová, K.; Žakovská, A.; Mejzlíková, M.; Halouzka, J.; Dendis, M. Isolation, cultivation and identification of Borrelia burgdorferi genospecies from Ixodes ricinus ticks from the city of Brno, Czech Republic. Ann. Agric. Environ. Med. 2007, 14, 75–79. [Google Scholar]
- Venclíková, K.; Mendel, J.; Betášová, L.; Blažejová, H.; Jedličková, P.; Straková, P.; Hubálek, Z.; Rudolf, I. Neglected tick-borne pathogens in the Czech Republic, 2011-2014. Ticks Tick Borne Dis. 2016, 7, 107–112 https://101016/jttbdis201509004. [Google Scholar] [CrossRef]
- Žygutienė, M.; Alekseev, A.; Dubinina, H.; Kazlauskienė, R. Evidence for a risk of tick borne infection in the city parks of Vilnius, Lithuania. Ekologija 2008, 54, 40–43. [Google Scholar] [CrossRef]
- Glass, A.; Springer, A.; Raulf, M.K.; Fingerle, V.; Strube, C. 15-year Borrelia prevalence and species distribution monitoring in Ixodes ricinus/inopinatus populations in the city of Hanover, Germany. Ticks Tick Borne Dis. 2023, 14, 102074. [Google Scholar] [CrossRef] [PubMed]
- Stańczak, J.; Gabre, R.M.; Kruminis-Łozowska, W.; Racewicz, M.; Kubica-Biernat, B. Ixodes ricinus as a vector of Borrelia burgdorferi sensu lato, Anaplasma phagocytophilum and Babesia microti in urban and suburban forests. Ann. Agric. Environ. Med. 2004, 11, 109–114. [Google Scholar] [PubMed]
- Kubiak, K.; Dziekońska-Rynko, J.; Szymańska, H.; Kubiak, D.; Dmitryjuk, M.; Dzika, E. Questing Ixodes ricinus ticks (Acari, Ixodidae) as a vector of Borrelia burgdorferi sensu lato and Borrelia miyamotoi in an urban area of north-eastern Poland. Exp. Appl. Acarol. 2019, 78, 113–126. [Google Scholar] [CrossRef] [PubMed]
- Grochowska, A.; Dunaj-Małyszko, J.; Pancewicz, S.; Czupryna, P.; Milewski, R.; Majewski, P.; Moniuszko-Malinowska, A. Prevalence of tick-borne pathogens in questing Ixodes ricinus and Dermacentor reticulatus ticks collected from recreational areas in northeastern Poland with analysis of environmental factors. Pathogens. 2022, 11, 468. [Google Scholar] [CrossRef] [PubMed]
- Mannelli, A.; Bertolotti, L.; Gern, L.; Gray, J. Ecology of Borrelia burgdorferi sensu lato in Europe: transmission dynamics in multi-host systems, influence of molecular processes and effects of climate change. FEMS Microbiol Rev. 2015, 36, 837–861. [Google Scholar] [CrossRef] [PubMed]
- Nijhof, A.M.; Bodaan, C.; Postigo, M.; Nieuwenhuijs, H.; Opsteegh, M.; Franssen, L.; Jebbink, F.; Jongejan, F. Ticks and associated pathogens collected from domestic animals in The Netherlands. Vector Borne Zoonotic Dis. 2007, 7, 585–596. [Google Scholar] [CrossRef]
- Kooyman, F.N.J.; Zweerus, H.; Nijsse, E.R.; Jongejan, F.; Wagenaar, J.A.; Broens, E.M. Monitoring of ticks and their pathogens from companion animals obtained by the “tekenscanner” application in The Netherlands. Parasitol Res. 2022, 121, 1887–1893. [Google Scholar] [CrossRef]
- Leschnik, M.W.; Khanakah, G.; Duscher, G.; Wille-Piazzai, W.; Hörweg, C.; Joachim, A.; Stanek, G. Species, developmental stage and infection with microbial pathogens of engorged ticks removed from dogs and questing ticks. Med. Vet. Entomol. 2012, 26, 440–446. [Google Scholar] [CrossRef]
- Jenkins, A.; Hvidsten, D.; Matussek, A.; Lindgren, P-E. ; Stuen, S.; Kristiansen, B-E. Borrelia burgdorferi sensu lato in Ixodes ricinus ticks from Norway: evaluation of a PCR test targeting the chromosomal flaB gene. Exp Appl Acarol. 2012, 58, 431–439. [Google Scholar] [CrossRef]
- Schreiber, C.; Krücken, J.; Beck, S.; Maaz, D.; Pachnicke, S.; Krieger, K.; Gross, M.; Kohn, B.; von Samson-Himmelstjerna, G. Pathogens in ticks collected from dogs in Berlin/Brandenburg, Germany. Parasites Vectors 2014, 14, 535. [Google Scholar] [CrossRef]
- Stensvold, C.R.; Al Marai, D.; Andersen, L.O.; Krogfelt, K.A.; Jensen, J.S.; Larsen, K.S.; Nielsen, H.V. Babesia spp. and other pathogens in ticks recovered from domestic dogs in Denmark. Parasites Vectors 2015, 8, 262. [Google Scholar] [CrossRef] [PubMed]
- Namina, A.; Capligina,V. ; Seleznova, M.; Krumins, R.; Aleinikova, D.; Kivrane, A.; Akopjana, S.; Lazovska, M.; Berzina, I.; Ranka, R. Tick-borne pathogens in ticks collected from dogs, Latvia, 2011–2016. BMC Vet. Res. 2019, 15, 398. [Google Scholar] [CrossRef] [PubMed]
- Zakham, F.; Korhonen, E.M.; Puonti, P.T.; Castrén, R.S.; Uusitalo, R.; Smura, T.; Kant, R.; Vapalahti, O.; Sironen. ; T., Kinnunenet, P.M. Molecular detection of pathogens from ticks collected from dogs and cats at veterinary clinics in Finland. Parasites Vectors 2023, 16, 327. [Google Scholar] [CrossRef]
- Król, N.; Obiegala, A.; Pfeffer, M.; Lonc, E.; Kiewra, D. Detection of selected pathogens in ticks collected from cats and dogs in the Wroclaw agglomeration, South-West Poland. Parasites Vectors 2016, 9, 351. [Google Scholar] [CrossRef]
- Michalski, M.M.; Kubiak, K.; Szczotko, M.; Chajęcka, M.; Dmitryjuk, M. Molecular detection of Borrelia burgdorferi sensu lato and Anaplasma phagocytophilum in ticks collected from dogs in urban areas of north-eastern Poland. Pathogens 2020, 9, 455. [Google Scholar] [CrossRef] [PubMed]
- Strnad, M.; Honig, V.; Ruzek, D.; Grubhoffer, L.; Rego, R.O.M. Europe-wide meta-analysis of Borrelia burgdorferi sensu lato prevalence in questing Ixodes ricinus ticks. Appl. Environ. Microbiol. 2017, 83, e00609–17. [Google Scholar] [CrossRef] [PubMed]
- Skotarczak, B. The role of companion animals in the environmental circulation of tick-borne bacterial pathogens. Ann Agric Environ Med. 2018, 25, 473–480. [Google Scholar] [CrossRef] [PubMed]
- Vogt, NA. Lyme borreliosis in animals. In: Merck Veterinary Manual. 2022. Available online: www.merckvetmanual.com/generalized-conditions/lyme-borreliosis/lyme-borreliosis-in-animals (accessed on 31 October 2022).
- Hanincová, K; Schäfer, S.M.; Etti, S.; Sewell, H.S.; Taragelová, V., Ziak, D.; Labuda, M.; Kurtenbach, K. Association of Borrelia afzelii with rodents in Europe. Parasitology 2003, 126, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Richter, D.; Schlee, D.B.; Matuschka, F.R. Reservoir competence of various rodents for the Lyme disease spirochete Borrelia spielmanii. Appl. Environ. Microbiol. 2011, 77, 3565–3570. [Google Scholar] [CrossRef] [PubMed]
- Skuballa, J.; Petney, T.; Pfaffle, M.; Oehme, R.; Hartelt, K.; Fingerle, V.; Kimmig, P.; Taraschewski, H. Occurrence of different Borrelia burgdorferi sensu lato genospecies including B. afzelii, B. bavariensis, and B. spielmanii in hedgehogs (Erinaceus spp.) in Europe. Ticks Tick Borne Dis. 2012, 3, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Michalik, J.; Wodecka, B.; Skoracki, M.; Sikora, B.; Stańczak, J. Prevalence of avian-associated Borrelia burgdorferi s. l. genospecies in Ixodes ricinus ticks collected from blackbirds (Turdus merula) and song thrushes (T. philomelos). Int. J. Med. Microbiol. 2008, 298 (Suppl. 1), 129–138. [Google Scholar]
- Richter, D.; Matuschka, FR. Perpetuation of the Lyme disease spirochete Borrelia lusitaniae by lizards. Appl Environ Microbiol. 2006, 72, 4627–4632. [Google Scholar] [CrossRef]
- Onyiche, T.E.; Răileanu, C.; Fischer, S.; Silaghi, C. Global distribution of Babesia species in questing ticks: a systematic review and meta-analysis based on published literature. Pathogens 2021, 10, 230. [Google Scholar] [CrossRef]
- Stańczak, J.; Cieniuch, S.; Lass, A.; Biernat, B.; Racewicz, M. Detection and quantification of Anaplasma phagocytophilum and Babesia spp. in Ixodes ricinus ticks from urban and rural environment, northern Poland, by real-time polymerase chain reaction. Exp. Appl. Acarol. 2015, 66, 63–81. [Google Scholar] [CrossRef]
- Schorn, S.; Pfister, K.; Reulen, H.; Mahling, M.; Silaghi, C. Occurrence of Babesia spp., Rickettsia spp. and Bartonella spp. in Ixodes ricinus in Bavarian public parks, Germany. Parasites Vectors 2011, 4, 135. [Google Scholar] [CrossRef]
- Welc-Falęciak, R.; Bajer, A.; Paziewska-Harris, A.; Baumann-Popczyk, A.; Siński, E. Diversity of Babesia in Ixodes ricinus ticks in Poland. Adv Med Sci. 2012, 57, 364–369. [Google Scholar] [CrossRef]
- Abdullah, S.; Helps, C.; Tasker, S.; Newbury, H. , Wall, R. Prevalence and distribution of Borrelia and Babesia species in ticks feeding on dogs in the UK. Med. Vet. Entomol. 2018, 32, 14–22. [Google Scholar] [CrossRef]
- Asman, M.; Solarz, K.; Cuber, P.; Gasior, T.; Szilman, P.; Szilman, E.; Tondas, E.; Matzullok, A.; Kusion, N.; Florek, K. Detection of protozoans Babesia microti and Toxoplasma gondii and their co-existence in ticks (Acari: Ixodida) collected in Tarnogórski district (Upper Silesia, Poland). Ann Agric Environ. 2015, 22, 80–83. [Google Scholar] [CrossRef]
- Karbowiak, G. Zoonotic reservoir of Babesia microti in Poland. Pol J Microbiol. 2004, 53 (Suppl), 61–65. [Google Scholar]
- Cieniuch, S.; Stańczak, J.; Ruczaj, A. The first detection of Babesia EU1 and Babesia canis canis in Ixodes ricinus ticks (Acari, Ixodidae) collected in urban and rural areas in northern Poland. Pol J Microbiol. 2009, 58, 231–236. [Google Scholar]
- Hamšíková, Z.; Kazimírová, M.; Haruštiaková, Z.; Kazimírová, M.; Haruštiaková, D.; Mahríková, M.; Slovák, M.; Berthová, L.; Kocianová, E.; Schnittger, L. Babesia spp. in ticks and wildlife in different habitat types of Slovakia. Parasites Vectors 2016, 9, 292. [Google Scholar] [CrossRef]
- Rybářová, M.; Honsová, M.; Papoušek, I.; Široký, P. Variability of species of Babesia Starcovici, 1893 in three sympatric ticks (Ixodes ricinus, Dermacentor reticulatus and Haemaphysalis concinna) at the edge of Pannonia in the Czech Republic and Slovakia. Folia Parasitol. 2017, 64, 028. [Google Scholar] [CrossRef]
- Adaszek, Ł.; Winiarczyk, S. Molecular characterization of Babesia canis canis isolates from naturally infected dogs in Poland. Vet. Parasitol. 2008, 52, 235–241. [Google Scholar] [CrossRef]
- Livanova, N.N.; Fomenko, N.V.; Akimov, I.A.; Ivanov, M.J.; Tikunova, N.V.; Armstrong, R.; Konyaev, S.V. 2018. Dog survey in Russian veterinary hospitals: tick identification and molecular detection of tick-borne pathogens. Parasites Vectors 2018, 11, 591. [Google Scholar] [CrossRef] [PubMed]
- Zygner, W.; Bąska, P.; Wiśniewski, M.; Wędrychowicz, H. The molecular evidence of Babesia microti in hard ticks removed from dogs in Warsaw. Pol J Mocrobiol. 2010, 59, 95–7. [Google Scholar] [CrossRef]
- Kocoń, A.; Asman, M.; Nowak-Chmura, M.; Witecka, J.; Rączka, G. Exposure of domestic dogs and cats to ticks (Acari: Ixodida) and selected tick-borne diseases in urban and recreational areas in southern Poland. Sci Rep. 2022, 12, 7851. [Google Scholar] [CrossRef] [PubMed]
- Sytykiewicz, H.; Karbowiak, G. , Hapunik, J.; Szpechciński, A.; Supergan-Marwicz, M.; Goławska, S.; Sprawka, I.; Czerniewicz, P. Molecular evidence of Anaplasma phagocytophilum and Babesia microti co-infections in Ixodes ricinus ticks in central-eastern region of Poland. Ann Agric Environ Med. 2012, 19, 45–49. [Google Scholar] [PubMed]
- Asman,M. ; Solarz, K.; Szilman, E.; Szilman, P.; Sikora, B.; Jakubas-Zawalska, J. The occurrence of three tick-borne pathogens in Ixodes ricinus ticks collected from the area of the Kraków-Częstochowa Upland (Southern Poland). Acarologia 2018, 58, 967–975. [Google Scholar] [CrossRef]
- Hildebrandt, A.; Gray, J.; Montero, E. Characteristics of Human Babesiosis in Europe. Pathogens 2023, 12, 323. [Google Scholar] [CrossRef]
- Goethert, H.K. What Babesia microti is now. Pathogens 2021, 10, 1168. [Google Scholar] [CrossRef]
- Moniuszko-Malinowska, A.; Swięcicka, I.; Dunaj, J.; Zajkowska, J.; Czupryna, P.; Zambrowski, G.; Chmielewska-Badora, J.; Żukiewicz-Sobczak, W.; Swierzbińska, R.; Rutkowski, K.; et al. Infection with Babesia microti in humans with non-specific symptoms in North East Poland. Infect. Dis. Lond. Engl. 2016, 48, 537–543. [Google Scholar] [CrossRef]
- Arsuaga, M.; Gonzalez, L.M.; Lobo, C.A.; de la Calle, F.; Bautista, J.M.; Azcárate, I.G.; Puente, S.; Montero, E. First report of Babesia microti-caused babesiosis in Spain. Vector Borne Zoonotic Dis. 2016, 16, 677–679. [Google Scholar] [CrossRef]
- Welc-Falęciak, R.; Pawełczyk, A.; Radkowski, M.; Pancewicz, S.A.; Zajkowska, J.; Siński, E. First report of two asymptomatic cases of human infection with Babesia microti (Franca, 1910) in Poland. Ann. Agric. Environ. Med. 2015, 22, 51–54. [Google Scholar] [CrossRef]
- Pawełczyk, A.; Bednarska, M.; Hamera, A.; Religa, E.; Poryszewska, M.; Mierzejewska, E.J.; Welc-Falęciak, R. Long-term study of Borrelia and Babesia prevalence and co-infection in Ixodes ricinus and Dermacentor recticulatus ticks removed from humans in Poland, 2016–2019. Parasites Vectors 2021, 14, 348. [Google Scholar] [CrossRef]
- Herwaldt, B.L.; Cacció, S.; Gherlinzoni, F.; Aspöck, H.; Slemenda, S.B.; Piccaluga, P.; Martinelli, G.; Edelhofer, R.; Hollenstein, U.; Poletti, G.; Pampiglione, S. , Löschenberger, K., Tura, S.,Pieniazek, N.J. Molecular characterization of a non-Babesia divergens organism causing zoonotic babesiosis in Europe. Emerg. Infect. Dis. 2003, 9, 942–948. [Google Scholar] [CrossRef]
- Rozej-Bielicka, W.; Masny, A.; Gołąb, E. High-resolution melting PCR assay, applicable for diagnostics and screening studies, allowing detection and differentiation of several Babesia spp. infecting humans and animals. Parasitol Res. 2017, 116, 2671–2681. [Google Scholar] [CrossRef]
- Skotarczak, B.; Wodecka, B.; Cichocka, A. Coexistence DNA of Borrelia burgdorferi sensu lato and Babesia microti in Ixodes ricinus ticks from north-western Poland. Ann Agric Environ Med. 2002, 9, 25–8. [Google Scholar]
- Sawczyn-Domańska, A.; Zwoliński, J.; Kloc, A.; Wójcik-Fatla, A. Prevalence of Borrelia, Neoehrlichia mikurensis and Babesia in ticks collected from vegetation in Eastern Poland. Exp. App. Acarol. 2023, 90, 409–428. [Google Scholar] [CrossRef]
- Kubiak, K; Dmitryjuk, M. ; Dziekońska-Rynko, J., Siejwa, P.; Dzika, E. The risk of exposure to ticks and tick-borne pathogens in a spa town in northern Poland. Pathogens 2022, 11, 542. [Google Scholar] [CrossRef]
- Miron, L.D.; Ciuca, L.; Ilie, C.; Potoroaca, A.; Lazăr, C.; Martinescu, G.-V. Co-Infection with Babesia canis and Borrelia burgdorferi s.l. in a dog from Northeastern Romania: A case report. JALSE. 2022, 54, 439–449. [Google Scholar] [CrossRef]
- Hildebrandt, A.; Pauliks, K.; Sachse, S.; Straube, E. Coexistence of Borrelia spp. and Babesia spp. in Ixodes ricinus Ticks in Middle Germany. Vector Borne Zoonotic Dis. 2010, 10, 831–837. [Google Scholar] [CrossRef]
- Meer-Scherrer, L.; Adelson, M.; Mordechai, E.; Lottaz, B.; Tilton Meer-Scherrer, R. Babesia microti Infection in Europe. Curr Microbiol. 2004, 48, 435–437. [Google Scholar] [CrossRef]
- Dunaj, J.; Moniuszko-Malinowska, A.; Swiecicka, I.; Andersson, M.; Czupryna, P.; Rutkowski, K.; Zambrowski, G.; Zajkowska, J.; Grygorczuk, S.; Kondrusik, M.; et al. Tick-borne infections and co-infections in patients with non-specific symptoms in Poland. Adv Med Sci. 2018, 63, 167–172. [Google Scholar] [CrossRef]
- Jabłońska, J.; Żarnowska-Prymek, H.; Stańczak, J.; Kozłowska, J.; Wiercińska-Drapało, A. Symptomatic coinfection with Babesia microti and Borrelia burgdorferi in patient after international exposure; a challenging case in Poland. Ann Agric Environ Med. 2016, 23, 387–389. [Google Scholar] [CrossRef]
- Welc-Falęciak, R.; Hildebrandt, A.; Siński, E. Coinfection with Borrelia species and other tick-borne pathogens in humans: two cases from Poland. Ann Agric Environ Med. 2010, 17, 309–313. [Google Scholar] [PubMed]
- Pańczuk, A.; Tokarska-Rodak, M.; Kozioł-Montewka, M.; Plewik, D. The incidence of Borrelia burgdorferi, Anaplasma phagocytophilum and Babesia microti coinfections among foresters and farmers in eastern Poland. J Vector Borne Dis. 2016, 53, 348–354. [Google Scholar]
- Cosson, J.F.; Michelet, L.; Chotte, J.; Le Naour, E.; Cote, M.; Devillers, E.; Poulle, L.M.; Huet, D.; Galan, M.; Geller, J.; et al. Genetic characterization of the human relapsing fever spirochete Borrelia miyamotoi in vectors and animal reservoirs of Lyme disease spirochetes in France. Parasit. Vectors 2014, 7, 233. [Google Scholar] [CrossRef]

Table 1.
The prevalence of Borrelia burgdorferi s.l. and Babesia spp. in single and double infections found in 1029 host-seeking I. ricinus ticks collected in urban areas of the city of Poznań.
Table 1.
The prevalence of Borrelia burgdorferi s.l. and Babesia spp. in single and double infections found in 1029 host-seeking I. ricinus ticks collected in urban areas of the city of Poznań.
| Females | Males | Nymphs | TOTAL | |
|---|---|---|---|---|
| Borrelia spp. | ||||
| B. afzelii | 10 | 17 | 11 | 38 (3.7) |
| B. garinii | 19 | 8 | 11 | 38 (3.7) |
| B. lusitaniae | 2 | 0 | 7 | 9 (0.9) |
| B. valaisianae | 2 | 1 | 0 | 3 (0.3) |
| Borrelia spp. | 1 | 1 | 0 | 2 (0.2) |
| Total | 34/289 (11.8) | 27/280 (9.6) | 29/460 (6.3) | 90/1029 (8.7) |
| Babesia spp. | ||||
| Babesia microti | 10 | 9 | 8 | 27 (2.6) |
| Babesia canis | 8 | 3 | 3 | 14 (1.4) |
| Babesia venatorum | 3 | 1 | 0 | 4 (0.4) |
| Total | 21/289 (7.3) | 13/280 (4.6) | 11/460 (2.4) | 45/1029 (4.4) |
| co-infections | ||||
| B. afzelii + Ba. canis | 1 | 3 | 0 | 4 (0.4) |
| B. garinii + Ba. canis | 1 | 3 | 0 | 4 (0.4) |
| B. lusitaniae + Ba. venatorum | 2 | 0 | 0 | 2 (0.2) |
| B. lusitaniae + Ba. microti | 1 | 0 | 0 | 1 (0.1) |
| Total | 5 | 6 | 0 | 11/1029 (1.1) |
Among the 1,268 feeding I. ricinus females, DNA B. burgdorferi s.l. was detected in 4.7% (n=59) individuals, and this prevalence was almost two-fold lower in comparison to host-seeking ticks (8.7%, χ2 test, P = 0.001). Infected ticks were found in all animal groups: dogs (4.8%), cats (3.3%) and undefined pets (5.0%). Five spirochete species were identified with a clear predominance of B. afzelii (3.7%; n=47) followed by B. garinii (0.6%; n=7), B. spielmani (0.2%; n=3), B. lusitaniae (0.1%; n=1) and B. valaisianae (0.1%). Except of B. valaisiana, the remaining four species were detected in dog-derived ticks. The five infected ticks collected from cats, harboured only B. afzelii, whereas 20 individuals from the group of undefined pets were infected with B. afzelii (n=16), B. garinii (n=3) and B. valaisiana.
Table 2.
The prevalence of Borrelia burgdorferi s.l. and Babesia spp. in single and double infections found in 1268 feeding I. ricinus female ticks removed from three groups of pet animals surveyed in veterinary clinics of the city of Poznań.
Table 2.
The prevalence of Borrelia burgdorferi s.l. and Babesia spp. in single and double infections found in 1268 feeding I. ricinus female ticks removed from three groups of pet animals surveyed in veterinary clinics of the city of Poznań.
| Dogs (n= 609) |
Cats (n=117) |
Undefined hosts (n=389) |
TOTAL | |
|---|---|---|---|---|
| Borrelia spp. | ||||
| B. afzelii | 26 (3,7) | 5 (3,3) | 16 (4,0) | 47 (3,7) |
| B. garinii | 4 (0,6) | 0 | 3 (0,7) | 7 (0,6) |
| B. spielmanii | 3 (0,4) | 0 | 0 | 3 (0,2) |
| B. lusitaniae | 1 (0,1) | 0 | 0 | 1 (0,1) |
| B. valaisianae | 0 | 0 | 1 | 1 (0,1) |
| Total | 34/711 (4.8) | 5/153 (3,3) | 20/404 (5,0) | 59/1268 (4,7) |
| Babesia spp. | ||||
| Babesia canis | 23 (3.2) | 1 (0.7) | 11 (2.7) | 35 (2.8) |
| Babesia microti | 18 (2.5) | 2 (1.3) | 8 (2.0) | 28 (2.2) |
| Babesia venatorum | 7 (1.0) | 3 (2.0) | 3 (0.7) | 13 (1.0) |
| Total | 48/711 (6,8) | 6/153 (3,9) | 22/404 (5,7) | 76/1268 (6,0) |
| co-infections | ||||
| B. afzelii + Ba. microti | 5 | 0 | 5 | 10 (0.8) |
| B. spielmanii + Ba. microti | 1 | 0 | 0 | 1 (0.1) |
| B. garinii + Ba. microti | 0 | 0 | 1 | 1 (0.1) |
| B. garinii + Ba. canis | 0 | 1 | 0 | 1 (0.1) |
| Total | 13/1268 (1.0) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



