
Submitted:
29 February 2024
Posted:
04 March 2024
You are already at the latest version
Abstract
Alcoholic liver disease (ALD) is a major cause of chronic liver disease. This term covers a broad spectrum of liver lesions, from simple steatosis to alcoholic hepatitis and cirrhosis. The patho-genesis of ALD is multifactorial and not fully elucidated due to complex mechanisms related to direct ethanol toxicity with subsequent hepatic and systemic inflammation. The accumulation of pro-inflammatory cytokines and the reduction of anti-inflammatory cytokines promote the development and progression of ALD. To date, there are no targeted therapies to counteract disease progression and prevent acute liver failure. Corticosteroids reduce mortality by acting on the hepatic-systemic inflammation. On the other hand, several studies analyzed the effect of the inhibition of pro-inflammatory cytokines and the stimulation of anti-inflammatory cytokines as potential therapeutic targets in ALD. This narrative review aims to clarify the role of the cy-tokines in the pathogenesis and treatment of ALD.
Keywords:
alcohol
; oxidative stress
; inflammation
; gut dysbiosis
; biological drugs
; probiotics
1. Introduction
Alcoholic liver disease (ALD) is one of the leading causes of chronic liver disease worldwide. In this way, alcohol consumption is often a cofactor in patients with hepatitis B and C and patients with non-alcoholic fatty liver disease [1]. According to the European Association for the Study of the Liver, the diagnosis of ALD should be suspected in the presence of liver damage (clinical signs and/or bio-humoral abnormalities) and a regular alcohol consumption of >30 g/day in men or >20 g/day in women [2]. ALD comprises a broad spectrum of liver lesions ranging from simple alcoholic fatty liver (AFL) to alcoholic steatohepatitis (ASH) and alcoholic hepatitis (AH) [3]. AFL diagnosis is established in patients with known alcohol use disorder (AUD) and hepatic steatosis observed on abdominal ultrasound combined with increased liver enzymes and the absence of other causes of liver disease. However, it is a serious problem due to the non-specific symptoms [4,5]. Similar to AFL, mild ASH rarely presents with clinical symptoms and can only be diagnosed by liver biopsy. AH is a clinical condition with a high mortality rate, and it is diagnosed by the presence of jaundice in the preceding eight weeks and elevated transaminase levels. Patients with AH may present signs of severe hepatic decompensation, such as ascites and hepatic encephalopathy [6]. The pathogenesis of ALD is multifactorial due to complex molecular pathways not yet fully elucidated. Certainly, a key role is played by the direct hepatotoxicity of ethanol, lipid peroxidation, stress with reactive oxygen species (ROS) production, and activation of the immune response by cytokines [7]. Although there are clear links between the amount and duration of alcohol consumption and the progression of ALD, other genetic and environmental factors are involved in the development and progression of ALD [8]. This narrative review aims to clarify the role of the cytokines in the pathogenesis and treatment of ALD.
2. Epidemiology and Social Impact of ALD
About 2 million deaths each year are linked to chronic liver disease worldwide [9]. Alcohol consumption is estimated at 43% of the global population. AUD prevalence is around 5% (283 million people), with the highest prevalence among men and women in the European region (14.8% and 3.5%, respectively) and in the Americas (11.5% and 5.1%, respectively) [10]. Alcohol is the leading cause of cirrhosis globally, responsible for almost 60% of cases in Europe, North America and Latin America, and about 35% of patients with AUD develop different forms of ALD [11]. The global prevalence of alcohol-associated cirrhosis was estimated at 23.6 million individuals with compensated cirrhosis and 2.46 million for those with decompensated cirrhosis, respectively [12]. In recent years, there has been an increase in the incidence of ALD, especially in the 15-45 age group [13]. This is a serious health problem because higher alcohol consumption in late adolescence increases the risk of developing severe liver disease, with a higher risk of death by cardiovascular diseases and cancer [14,15]. Mortality related to ALD has increased over the last decade more in developed countries, such as Europe, Asia, Latin America and the United States [16,17]. In addition, another increase in ALD deaths was observed during the Coronavirus Disease-19 pandemic due to higher alcohol consumption related to emotive stress and difficult access to treatments [18]. Especially in its final stage, represented by liver cirrhosis, ALD has a high socio-economic impact. It is estimated that in 2016, liver disease-related spending in the United States was $32.5 billion. Two-thirds of these costs are attributable to hospital or emergency room care. In 20 years, healthcare spending on liver disease has increased by 4% per year [19]. Based on ALD-related disability-adjusted life years from 2016, 21.5 million life years were lost due to ALD, with men affected significantly more than women. A significant portion of these lost life years was attributable to premature death rather than disability [20]. Based on the healthcare claims analysis report, of those who survived, more than 50% were hospitalized within one year and almost 75% in the second year, with a total cost of about $145,000 for each patient [21]. Patients with AUD have an increased risk of psychiatric comorbidities, such as anxiety, affective disorders and schizophrenia [22]. Malnutrition is frequent in ALD patients due to reduced caloric intake, abnormal digestion, increased protein catabolism and abnormal lipid metabolism [23,24]. Patients with ALD should receive appropriate nutritional assessment and support. This has a major socio-economic impact, as in many cases hospitalization is required for adequate parenteral or enteral nutrition [25]. In this way, folate deficiency is the result of reduced dietary folate intake, intestinal malabsorption, reduced hepatic absorption and deposition, and increased urinary excretion [26,27]. This event causes the megaloblastic anemia as a main complication. In addition, chronic alcoholics also have thiamine, vitamin B6 and vitamin B12 deficiency [28]. Several studies conducted in animal models have shown how these deficits contribute to the progression of ALD and the development of hepatocellular carcinoma (HCC) and colorectal cancer due to an epigenetic mechanism involving DNA methylation [29,30]. On the other hand, thiamine deficiency is associated with Wernicke encephalopathy (WE), which presents with altered mental status, gait ataxia and ophthalmoplegia. About 80% of patients with non-treated WE develop Korsakoff’s syndrome (KS), characterized by memory disturbances associated with confabulation [31]. A study conducted by Wilson et al. showed that treatment with intravenous thiamine for 5 days rather than 2 days increases the healthcare costs of acute care but reduces the expected lifecycle cost should the patient develop KS [32]. On the other hand, a study performed by Thompson et al. defined how healthcare costs for the management of ALD increased among US patients from 2006 to 2013. Total costs were nearly $145,000 per patient, decreasing from $50,000 in the first year to $10,000 per year in the last years. Liver transplanted patients averaged about $300,000 in transplant-related costs and over $1,000,000 in total healthcare costs over five years [33].
3. ALD Pathogenesis
The pathogenesis of ALD is based on multiple and complex molecular mechanisms that are not yet fully elucidated. These include direct ethanol hepatotoxicity, lipid peroxidation, oxidative stress with consequent ROS production, activation of the immune response and activation of proinflammatory cytokines [34]. Ethanol metabolism comprises three pathways: i) hepatocyte cytoplasmic alcohol dehydrogenase oxidizes ethanol into acetaldehyde, a highly toxic compound that can alter DNA synthesis; ii) the enzyme cytochrome P450 2E1 (CYP2E1), which oxidizes ethanol into acetaldehyde, generate ROS and triggers oxidative stress and inflammation; iii) heme-containing catalase in peroxisomes oxidize ethanol to acetaldehyde and subsequently the enzyme aldehyde dehydrogenase (ADH) oxidize acetaldehyde into acetate, which is released into the circulatory system, and further oxidized into carbon dioxide in various extrahepatic tissues [35,36,37,38]. Chronic alcohol use increases CYP2E1 expression, resulting in increased acetaldehyde concentration, decreased ADH activity, reduced acetaldehyde oxidation, and accumulation of acetaldehyde in hepatocytes. This explains the direct hepatotoxicity of ethanol on the liver [39]. In addition, ethanol and acetaldehyde upregulate adiponectin, signal transducer and activator of transcription 3 (STAT3) and reduce zinc levels, inhibiting 5′-AMP-activated protein kinase and peroxisome proliferator-activated receptor α, resulting in lipid peroxidation and ROS production [40]. The increase in ROS damages mitochondrial DNA and proteins, causing a reduction in mitochondrial ATP and glutathione. ROS-induced release of hepatocellular kinase 1 regulates the hepatocellular apoptosis signal through the cleavage of pro-caspase-3 into caspase-active 3 [41]. Chronic alcohol consumption is involved in the accumulation of intestinal endotoxins and increased permeability of the intestinal wall, facilitating the translocation of lipopolysaccharide (LPS) from the gut to the liver. LPS can bind to toll-like receptors (TLRs) and activate the synthesis and release of cytokines and inflammatory factors, such as tumor necrosis factor-α (TNF-α), interleukin (IL)-1β, IL-6, and platelet-derived growth factor, further stimulating the accumulation of neutrophils and macrophages and causing liver inflammation and systemic damage in liver Kupffer cells [42]. Additionally, liver damage activates the proliferation of hepatic stellate cells (HSCs), which enhance the secretion of transforming growth factor-β (TGF-β) and collagen synthesis leading to fibrogenesis [43]. In recent years, the link between the changes in the gut microbiota composition and the ALD development and progression has been investigated [44]. Several studies employing preclinical and clinical models have shown that chronic alcohol consumption induces a decrease in Bacteroidetes and Firmicutes and an increase in Enterobacteriaceae and Proteobacteria, leading to gut dysbiosis [45,46]. Gut dysbiosis is associated with increased intestinal permeability linked to altered tight junctions, with translocation of pathogen-associated molecular patterns (PAMPs) in the liver through the portal vein [47]. In addition, alcohol abuse is associated with decreased levels of butyrate-producing genera and increased levels of proinflammatory Enterobacteriaceae. All of this leads to liver damage due to PAMPs translocation process [48]. The different pathogenetic ways involved in ALD development are summarized in Figure 1.
4. Role of Pro-Inflammatory Cytokines
4.1. TNF-α
Alcohol-induced damage occurs at several levels, from innate immune cells to hepatocytes. Innate immune cells, including hepatic macrophages (Kupffer cells), play a key role in early alcohol-induced liver damage through the recognition of LPS in the portal circulation. This results in the production of LPS-induced inflammatory cytokines [49]. Among the pro-inflammatory cytokines that promote liver damage, a key role is played by TNF-α [50]. Recognition of LPS activates members of the mitogen-activated protein kinase family, including extracellular receptor-activated kinase 1/2 (ERK1/2), p38 and the c-jun-N-terminal kinase, resulting in TNF-α production, mediated by oxidative stress [51]. LPS stimulates TLR4 and, at the same time, NADPH oxidase (NOX) interacts with the COOH-terminal region of TLR4, resulting in the generation of ROS in neutrophils and monocytes, which directly activates nuclear factor κB (NF-κB), with increased TNF-α concentration [52]. A study conducted by Thakur et al. showed how the use of diphenyliodonium and dilinoleoyl-phosphatidylcholine in Kupffer cells of mice models, reduced ERK1/2 activation, inhibited NOX4, resulting in lower ROS and TNF-α production [53]. Convincing evidence about the key role of TNF-α in the development of ALD was obtained from transcriptome studies that confirmed its up-regulation in patients with ALD [54]. Furthermore, a higher serum concentration correlates with a worse prognosis and an advanced disease stage [55]. Gonzales-Quintela et al. showed that serum TNF-α levels were almost similar in the general population, in teetotalers and alcohol drinkers, while in chronic alcoholics were elevated [56]. Other studies clarified how chronic alcohol stimulates the liver through prolonged activation of NF-κB, resulting in TNF-α production. Similar to resident liver macrophages, monocytes from chronic alcoholics also showed increased NF-κB activation compared to the control subjects. Acute alcohol exposure inhibits NF-κB activation, leading to a reduction in TNF-α levels [57,58]. A study performed by Mookerjee et al. showed that TNF-α is an important mediator of portal and systemic hemodynamic disturbances in ALD. This was the first study in which it was observed that the use of the biological drug infliximab improved cardiovascular hemodynamics, hepatic venous pressure gradient and hepatic and renal blood flow only twenty-four hours after its use [59]. Furthermore, TNF-α plays a key role in hepatocyte proliferation and liver regeneration, as reported by in vivo studies [60]. Although TNF-α is implicated in alcohol-related liver damage, liver cell death is not the normal outcome when hepatocytes are exposed to this cytokine. This suggests that the normal responses of hepatocytes to this cytokine are subverted during liver injury [61].
4.2. IL-8 and CXCL1
Neutrophil infiltration is a key feature of ALD. Unlike other innate immune cells, neutrophils are not liver-resident immune cells [62]. IL-8 plays a key role in the recruitment of neutrophils to liver tissue involving the chemokine CXC motif chemokine ligand 1 (CXCL1). Indeed, serum and liver levels of IL-8 and CXCL1 are directly correlated with disease severity and mortality [63]. A study performed by Patel et al. showed that IL-8 serum levels were significantly higher in patients with severe AH than in those with mild AH and that these levels were better predictors of short-term mortality than conventional prognostic scores [64]. Wieser et al. demonstrated how blockade of IL-8 receptors with short lipopeptides (pepducin) in mice models reduced liver inflammation, weight loss and mortality associated with ALD [65]. Nischalke et al. also showed that the CXCL1 rs4074 single nucleotide polymorphism was associated with increased blood levels of CXCL1 and an increased risk of developing cirrhosis in alcoholics as well as the development of HCC [66]. Mice models treated with a high-fat and ethanol-rich diet significantly upregulated the hepatic expression of several chemokines, including CXCL1, with a reduction of the infiltration and hepatic damage of neutrophils [67]. Roh et al. showed that the production of cytokines, including CXCL1, is mediated by TLR-2 and TLR-9, activated by LPS translocated from the gut and expressed on Kupffer cells. This data showed that in ethanol-treated wild-type mice, there was an increase in the hepatic expression of CXCL1 and the serum level of CXCL1, while TLR2- and TLR9-deficient mice showed significantly lower levels [68].
4.3. IL-1β
As previously reported, chronic exposure to ethanol in ALD sensitizes Kupffer cells to activation by LPS through TLR-4, inducing the production of proinflammatory cytokines, including IL-1β [69]. This is produced as a result of inflammasome activation, mainly of NOD-like receptor protein 3, highly expressed in macrophages and liver monocytes but lower expressed by hepatocytes and stellate cells [70]. NLR family pyrin domain containing 3 (NLRP3) is activated upon binding of PAMPs to TLR. This results in NF-κB activation, culminating in the transcription of pro-IL-1β precursors. Subsequently, procaspase-1 binds to NLRP3, transforming into caspase-1, which is catalytically active for IL-1β processing [71]. A study by Petrasek et al. conducted in murine models found that protein levels of NLRP3 and IL-1β were significantly increased in the liver of chronic alcohol-fed mice compared to the control group. Furthermore, data showed that mice deficient in the NLRP3 and caspase-1 inflammasome components had less steatosis and liver damage than wild-type mice on an alcoholic diet. In addition, IL-1R1 knockout mice were protected against ALD [72]. Cui et al. demonstrated, in Kupffer cells of ethanol-fed mice, an up-regulation of NLRP3 inflammasome components as well as IL-1β [73]. Similarly, a study by Voican et al. showed that increased levels of IL-1β were reported in ALD patients after one week of alcohol withdrawal [74]. In vitro experiments have shown that IL-1β is able to act directly on HSCs, with a subsequent proliferation and trans-differentiation in myofibroblasts, and with an increase in collagen and TGF-β levels [75]. A further function of IL-1β is the activation of invariant natural-killer T lymphocytes (iNKT) [76]. The main role of iNKTs is to determine the hepatic infiltration of neutrophils, a hallmark in the pathogenesis of ALD [77]. iNKT cell-deficient mice were protected from hepatic infiltration of neutrophils and liver damage induced by chronic ethanol binges. In contrast, wild-type mice showed intense hepatic infiltration of neutrophils and marked upregulation of hepatic expression of several inflammation-associated genes. IL-1β also inhibits liver regeneration [78]. Finally, mice treated with an IL-1 receptor antagonist showed a better regeneration of hepatocytes and an increased rate of recovery from liver damage induced by chronic ethanol consumption than untreated mice [79]. Table 1 summarizes the different studies about the involvement of pro-inflammatory cytokines in ALD.
Legend: TNF-α, tumor necrosis factor-alpha; ERK1/2, extracellular signal-regulated kinase 1/2; NOX4, NADPH oxidase 4; ROS, reactive oxygen species; ALD, alcoholic liver disease; LPS, lipopolysaccharide; AH, alcoholic hepatitis; IL-8, interleukin-8; CXCL1, CXC motif chemokine ligand 1; TLR2, toll-like receptor 2; TLR9, toll-like receptor 9; IL-1β, interleukin-1 beta; NLRP3, NLR family pyrin domain containing 3.
5. Role of Anti-Inflammatory Cytokines
5.1. IL-6 and IL-10
To date, IL-6, IL-22, and IL-10 are the identified cytokines with anti-inflammatory and hepatoprotective roles in ALD. IL-6 and IL-10 regulate the expression of target genes involved in promoting cell proliferation, survival and differentiation through STAT3 [80]. Kupffer cells express high levels of receptors for IL-10. This binding leads to a prolonged activation of STAT3 and inhibits inflammatory responses. Conversely, IL-6 binding to its own receptor leads to transient activation of STAT3, followed by the induction of inflammatory responses. Activation of STAT3 induces the expression of the suppressor of cytokine signaling 3, which in turn inhibits STAT3 activation by IL-6 but does not inhibit IL-10 signaling, leading to an anti-inflammatory effect [81]. The role of IL-6 in ALD is complex and not entirely clear. Some studies have shown how its expression can reduce apoptosis in hepatocytes and lead to mitochondrial DNA repair [82]. In this regard, a study by Zhang et al. in mice models showed that IL-6 can activate enzymes to repair mitochondrial DNA in hepatocytes damaged by chronic alcohol consumption [83]. IL-6 promotes the differentiation of T helper 17 cells and the production of IL-17, contributing to alcohol-related inflammation. However, the activation of STAT3 by IL-6 and IL-10 inhibits the production of other proinflammatory cytokines, reducing liver damage in patients with ALD [84]. Pre-treatment with IL-6 induces hepatoprotection of steatotic liver isotransplants by preventing the apoptosis of sinusoidal endothelial cells, improving hepatic microcirculation and protecting against hepatocyte death [85]. Another study by El-Assal et al. showed that IL-6 knockout mice fed with alcohol exhibited increased liver fat accumulation, lipid peroxidation, mitochondrial DNA damage and hepatocyte sensitization to TNFα-induced apoptosis [86]. IL-10 knockout mice showed a more severe hepatic inflammatory response with higher levels of IL-6 and STAT3 activation than wild-type mice but lower liver steatosis severity and hepatocellular damage after higher alcohol levels or fatty dietary regimen [87]. Byun et al. demonstrated how treatment with polyinosinic:polycytidylic acid in vitro stimulates IL-10 production in HSCs through TLR-3 activation with reduction of alcoholic liver damage [88]. IL-10 plays an important role in the negative regulation of liver regeneration by limiting the inflammatory response and subsequently mitigating hepatic STAT3 activation [89]. A study conducted by Yang et al. on 40 ethnically Taiwanese Han patients with alcoholic liver cirrhosis showed how certain IL-10 promoter polymorphisms allow the development of severe forms of the disease. This result clarifies how a better knowledge of the genetic background of alcoholic liver cirrhosis is essential for its prevention and treatment [90].
5.2. IL-22
IL-22 is a cytokine belonging to the IL-10 family due to the similarity in genetic and protein structures. It is involved in several diseases, such as inflammatory bowel diseases and skin, pancreatic, lung and liver diseases [91]. IL-22 is only produced by hematopoietic cells, while its receptor (IL-22R) is expressed in different organs. It’s a heterodimer composed of IL-10R2 and IL-22R1: while IL-10R2 is a subunit shared with various IL-10 family receptor complexes, IL-22R1 is only expressed in cells of various organs such as bronchi, liver, pancreas, and gut [92]. IL-22 plays a protective role in alcoholic liver damage through the activation of the STAT3-mediated signaling pathway. This event promotes the overexpression of anti-apoptotic and mitogenic genes, leading to tissue repair and survival of hepatocytes [93]. A recent study performed in mice models with ALD showed that four weeks of treatment with IL-22 reduced autophagy and liver fibrosis in hepatocytes [94]. In a recent prospective cohort study, IL-22 levels were significantly increased in alcohol-associated non-severe hepatitis and alcohol-associated severe hepatitis patients compared to the control group. As reported by the Authors, increased response to proinflammatory cytokines can mitigate liver damage [95]. Another investigation has clarified the role of IL-22 in liver regeneration. It was observed how IL-22 was significantly increased in murine models following liver regeneration and how this signaling blockade caused a reduction in liver regenerative capacity [96]. Table 2 summarizes the different studies on the involvement of anti-inflammatory cytokines in ALD. As previously reported, chronic alcohol consumption causes gut dysbiosis. Indeed, gut microflora imbalance reduces IL-22 production in the small intestine in mice models, which in turn leads to a decrease in antimicrobial C-type lectin regenerating islet-derived 3 gamma (REG3G). The reduction of REG3G causes an increase in bacterial translocation to the liver with consequent steatohepatitis [97].
Legend: IL-6, interleukin-6; TNF-α, tumor necrosis factor-alpha; IL-10, interleukin-10; STAT3, adiponectin, signal transducer and activator of transcription 3; ALD, alcoholic liver disease; HSCs, hepatic stellate cells; TLR3, toll-like receptor 3; IL-22, interleukin-22.
6. New Therapeutic Approaches
6.1. Biological Drugs
The treatment of ALD is based on AUD therapy and the management of severe hepatitis associated with alcohol abuse [98]. To date, the drugs approved for the AUD are disulfiram, naltrexone, nalmefene, and acamprosate. Disulfiram, naltrexone and acamprosate are used for alcohol withdrawal, while nalmefene acts to reduce alcohol consumption. These drugs act on ethanol metabolism or, at the central nervous system level, by modulating the glutaminergic and opioid pathways [99]. Two other drugs, sodium oxybate and baclofen, are used to treat AUD in some countries, such as Italy and Austria [100]. To date, treatment for severe AH benefits exclusively from the use of corticosteroids. The basic mechanism is the reduction of inflammation by cytokines modulation [101]. Corticosteroids increase survival in the short term but not in the long term and are associated with collateral effects, such as infections [102,103]. Since ALD is characterized by a cytokine storm, a potential treatment may be the inhibition of pro-inflammatory cytokines or the increase of anti-inflammatory cytokines levels. In this way, pentoxifylline, a selective phosphodiesterase inhibitor, has been tested as a therapeutic agent for the treatment of severe AH, acting with a reduction of pro-inflammatory cytokines levels [104]. The STOPAH study comparing the efficacy of pentoxifylline and prednisolone showed that pentoxifylline did not improve survival in AH patients, while prednisolone reduced 28-day mortality [105]. Other agents, such as infliximab and etanercept, which act by blocking TNF-α production, have been studied for the treatment of severe AH. A study conducted in patients with severe AH treated with infliximab and prednisolone showed an increased mortality rate compared to patients treated with prednisolone alone [106]. A further study clarified how a single dose of infliximab could improve the severity and survival rate in severe AH. However, in both studies, the most common side effect was the development of infections. For this reason, further investigations are needed in order to use them as a potential therapeutic target [107,108]. Similarly to infliximab, etanercept has also proved unsuccessful in the treatment of severe AH [109]. Recently, new therapeutic targets have emerged. Already in preclinical models, IL-22 has proven effective in reducing liver damage associated with ALD [110,111]. Phase I clinical trials have demonstrated the safety and efficacy of recombinant human IL-22 Immunoglobulin G (IgG)2-Fc, showing that the latter has good tolerance with good pharmacokinetic and pharmacodynamic properties at the intravenous dose of 45μg/kg [112,113]. Recently, an open-label phase II study was conducted on F-652, a recombinant fusion protein of human IL-22 and IgG2-Fc. Specifically, 18 patients (9 with moderate AH and 9 with severe AH) were enrolled, and three growing doses of F-652 (10, 30, 45 μg/kg) were administered. The treatment proved to be safe, and the patients showed a high rate of improvement as determined by the Lille and Model for End-Stage Liver Disease prognostic scores, with decreased levels of inflammatory biomarkers and increased levels of liver regeneration biomarkers, compared to control subjects. Despite this, the small sample number defines how multi-center, randomized, placebo-controlled studies are needed to confirm the benefits of IL-22Fc therapy in patients with moderate-severe AH [114]. IL-22 has been shown to promote the development of HCC via a STAT3-mediated mechanism [115]. However, studies performed on transgenic mouse models with high levels of IL-22 showed no higher incidence of spontaneous tumor development than in wild-type mice [116]. Another therapeutic target for the treatment of severe AH may be IL-1β inhibition. The ongoing ISAIAH study evaluates the efficacy of the monoclonal antibody canakinumab at a dose of 3 mg/kg intravenously at baseline and after 28 days. The primary endpoint obtained was the histological improvement of AH on liver biopsy after 28 days of treatment compared to baseline, defined as a reduction in lobular inflammation [117]. A study was also conducted on the use of anakinra, an interleukin-1β receptor antagonist. Therapy with anakinra 100 mg by subcutaneous injection for 14 days plus zinc sulfate 220 mg for 90 days was administered and compared to prednisone 40 mg PO daily for 30 days. As reported by the Authors, therapy with anakinra plus zinc sulfate improved survival by 90 days in patients with severe AH [118]. However, evaluating if the drug can be eliminated efficiently in patients with impaired kidney function [119] is necessary. Overall, the above-mentioned study provided better knowledge about the effects and metabolism of anakinra in patients with severe AH [118].
6.2. Gut Microbiota Modulation
Gut microbiota modulation can be a safe therapeutic strategy for improving patients’ quality of life [120]. In this way, a recent study performed on mice models with alcoholic liver damage showed that the probiotic Lactobacillus plantarum reduces the abundance of Gram-negative bacteria, with a reduction of the LPS content in the gut. Moreover, Lactobacillus plantarum J26 is able to maintain the integrity of the gut barrier, preventing bacterial translocation to the liver with a consequent reduction of the inflammation linked to alcohol consumption [121]. A further study on murine models compared the protective role of two strains of Lactobacillus plantarum, E680 and ZY08, in the ALD. Data reported that the intervention with Lactobacillus plantarum ZY08 significantly mitigated alcohol-related hepatic steatosis, liver damage, gut dysbiosis, and relieved plasma LPS levels as well as liver lipid metabolism [122]. A recent study performed in patients with severe AH showed that daily oral administration of Lactobacillus rhamnosus GG is associated with a significant reduction in liver damage after one month of treatment [123]. A double-blind, randomized, placebo-controlled, multicenter study is still ongoing with the aim of evaluating the efficacy of bovine colostrum in the treatment of AH. Bovine colostrum has two important components, lactoferrin and IgG: lactoferrin binds to lipid A to neutralize it, and IgG interacts with lymphoid tissue associated with the mucosa and reduces intestinal permeability. Indeed, this compound acts as an immunomodulator, reducing the inflammation typical of ALD [124]. Overall, further studies are required to better define the key role of gut microbiota modulation in the treatment of liver diseases [125,126].
7. Conclusions
In conclusion, pro-inflammatory and anti-inflammatory cytokines play a key role in the pathogenesis and progression of ALD. However, clarifications about the immune pathways involved in this pathology are needed. In this way, new therapeutic approaches, such as biological drugs and probiotics as targets of cytokines and the gut microbiota that modulates them, may be a good therapeutic option. Despite this, new studies are needed to evaluate the use of these promising therapies and their beneficial and adverse effects on ALD.
Author Contributions
Conceptualization: L.A.; formal analysis: G.G.M.S. and C.C.; data curation: G.G.M.S. and C.C.; writing original draft: G.G.M.S., C.C. and L.A.; writing, review and editing: M.L.S., M.D. and A.F.; supervision: L.A. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not. Applicable.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
| ADH | aldehyde dehydrogenase |
| AFL | alcoholic fatty liver |
| AH | alcoholic hepatitis |
| ALD | alcoholic liver disease |
| ASH | alcoholic steatohepatitis |
| AUD | alcohol use disorder |
| CXCL1 | chemokine CXC motif chemokine ligand 1 |
| CYP2E1 | cytochrome P450 2E1 |
| ERK1/2 | extracellular receptor-activated kinase 1/2 |
| HCC | hepatocellular carcinoma |
| HSCs | hepatic stellate cells |
| IgG | immunoglobulin G |
| IL | interleukin |
| iNKT | invariant natural-killer T lymphocytes |
| KS | Korsakoff’s syndrome |
| LPS | lipopolysaccharide |
| NF-κB | nuclear factor κB |
| NLRP3 | NLR family pyrin domain containing 3 |
| NOX | NADPH oxidase |
| PAMPs | pathogen-associated molecular patterns |
| REG3G | antimicrobial C-type lectin regenerating islet-derived 3 gamma |
| ROS | reactive oxygen species |
| STAT3 | signal transducer and activator of transcription 3 |
| TGF-β | transforming growth factor-β |
| TLRs | toll-like receptors |
| TNF-α | tumor necrosis factor-alpha |
| WE | Wernicke encephalopathy |
References
- Singal, A.K.; Bataller, R.; Ahn, J.; Kamath, P.S.; Shah, V.H. ACG Clinical Guideline: Alcoholic Liver Disease. Am. J. Gastroenterol. 2018, 113, 175–194. [Google Scholar] [CrossRef]
- European Association for the Study of the Liver. EASL Clinical Practice Guidelines: Management of alcohol-related liver disease. J. Hepatol. 2018, 69, 154–181. [Google Scholar] [CrossRef] [PubMed]
- Louvet, A.; Mathurin, P. Alcoholic liver disease: Mechanisms of injury and targeted treatment. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 231–242. [Google Scholar] [CrossRef]
- Abenavoli, L.; Spagnuolo, R.; Scarlata, G.G.M.; Scarpellini, E.; Boccuto, L.; Luzza, F. Ultrasound Prevalence and Clinical Features of Nonalcoholic Fatty Liver Disease in Patients with Inflammatory Bowel Diseases: A Real-Life Cross-Sectional Study. Medicina 2023, 59, 1935. [Google Scholar] [CrossRef]
- Singal, A.K.; Bataller, R.; Ahn, J.; Kamath, P.S.; Shah, V.H. ACG Clinical Guideline: Alcoholic Liver Disease. Am. J. Gastroenterol. 2018, 113, 175–194. [Google Scholar] [CrossRef] [PubMed]
- Crabb, D.W.; Bataller, R.; Chalasani, N.P.; Kamath, P.S.; Lucey, M.; Mathurin, P.; McClain, C.; McCullough, A.; Mitchell, M.C.; Morgan, T.R.; et al. Standard Definitions and Common Data Elements for Clinical Trials in Patients With Alcoholic Hepatitis: Recommendation From the NIAAA Alcoholic Hepatitis Consortia. Gastroenterology 2016, 150, 785–790. [Google Scholar] [CrossRef] [PubMed]
- Nagy, L.E.; Ding, W.X.; Cresci, G.; Saikia, P.; Shah, V.H. Linking Pathogenic Mechanisms of Alcoholic Liver Disease With Clinical Phenotypes. Gastroenterology 2016, 150, 1756–1768. [Google Scholar] [CrossRef]
- Argemi, J.; Ventura-Cots, M.; Rachakonda, V.; Bataller, R. Alcoholic-related liver disease: Pathogenesis, management and future therapeutic developments. Rev. Esp. Enferm. Dig. 2020, 112, 869–878. [Google Scholar] [CrossRef]
- Devarbhavi, H.; Asrani, S.K.; Arab, J.P.; Nartey, Y.A.; Pose, E.; Kamath, P.S. Global burden of liver disease: 2023 update. J. Hepatol. 2023, 79, 516–537. [Google Scholar] [CrossRef]
- World Health Organization. Global status report on alcohol and health 2018. Available online: https://www.who.int/publications/i/item/9789241565639 (accessed on 19 February 2024).
- Stein, E.; Cruz-Lemini, M.; Altamirano, J.; Ndugga, N.; Couper, D.; Abraldes, J.G.; Bataller, R. Heavy daily alcohol intake at the population level predicts the weight of alcohol in cirrhosis burden worldwide. J. Hepatol. 2016, 65, 998–1005. [Google Scholar] [CrossRef]
- Aslam, A.; Kwo, P.Y. Epidemiology and Disease Burden of Alcohol Associated Liver Disease. J. Clin. Exp. Hepatol. 2023, 13, 88–102. [Google Scholar] [CrossRef]
- Singal, A.K.; Arsalan, A.; Dunn, W.; Arab, J.P.; Wong, R.J.; Kuo, Y.F.; Kamath, P.S.; Shah, V.H. Alcohol-associated liver disease in the United States is associated with severe forms of disease among young, females and Hispanics. Aliment. Pharmacol. Ther. 2021, 54, 451–446. [Google Scholar] [CrossRef]
- Hagström, H.; Hemmingsson, T.; Discacciati, A.; Andreasson, A. Alcohol consumption in late adolescence is associated with an increased risk of severe liver disease later in life. J. Hepatol. 2018, 68, 505–510. [Google Scholar] [CrossRef]
- Hagström, H.; Thiele, M.; Roelstraete, B.; Söderling, J.; Ludvigsson, J.F. Mortality in biopsy-proven alcohol-related liver disease: A population-based nationwide cohort study of 3453 patients. Gut 2021, 70, 170–179. [Google Scholar] [CrossRef]
- Jyani, G.; Prinja, S.; Ambekar, A.; Bahuguna, P.; Kumar, R. Health impact and economic burden of alcohol consumption in India. Int. J. Drug Policy 2019, 69, 34–42. [Google Scholar] [CrossRef]
- Díaz, L.A.; Idalsoaga, F.; Fuentes-López, E.; Márquez-Lomas, A.; Ramírez, C.A.; Roblero, J.P.; Araujo, R.C.; Higuera-de-la-Tijera, F.; Toro, LG.; Pazmiño, G.; et al. Impact of Public Health Policies on Alcohol-Associated Liver Disease in Latin America: An Ecological Multinational Study. Hepatology 2021, 74, 2478–2490. [Google Scholar] [CrossRef]
- White, A.M.; Castle, I.P.; Powell, P.A.; Hingson, R.W; Koob, G.F. Alcohol-Related Deaths During the COVID-19 Pandemic. JAMA 2022, 327, 1704–1706. [Google Scholar] [CrossRef]
- Rehm, J.; Shield, K.D. Global Burden of Alcohol Use Disorders and Alcohol Liver Disease. Biomedicines 2019, 7, 99. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.A.; Martinson, N.; Martinson, M. Mortality and costs associated with alcoholic hepatitis: A claims analysis of a commercially insured population. Alcohol. 2018, 71, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Qian, A.S.; Nguyen, N.H.; Stukalin, I.; Congly, S.E.; Shaheen, A.A.; Swain, M.G.; Teriaky, A.; Asrani, S.K.; Singh, S. Trends in the Economic Burden of Chronic Liver Diseases and Cirrhosis in the United States: 1996-2016. Am. J. Gastroenterol. 2021, 116, 2060–2067. [Google Scholar] [CrossRef] [PubMed]
- Grant, B.F.; Goldstein, R.B.; Saha, T.D.; Chou, S.P.; Jung, J.; Zhang, H.; Pickering, R.P.; Ruan, W.J.; Smith, S.M.; Huang, B.; et al. Epidemiology of DSM-5 Alcohol Use Disorder: Results From the National Epidemiologic Survey on Alcohol and Related Conditions III. JAMA Psychiatry 2015, 72, 757–766. [Google Scholar] [CrossRef] [PubMed]
- Lucey, M.R.; Mathurin, P.; Morgan, T.R. Alcoholic hepatitis. N. Engl. J. Med. 2009, 360, 2758–2769. [Google Scholar] [CrossRef] [PubMed]
- Styskel, B.; Natarajan, Y.; Kanwal, F. Nutrition in Alcoholic Liver Disease: An Update. Clin. Liver Dis. 2019, 23, 99–114. [Google Scholar] [CrossRef] [PubMed]
- Dasarathy, S. Nutrition and Alcoholic Liver Disease: Effects of Alcoholism on Nutrition, Effects of Nutrition on Alcoholic Liver Disease, and Nutritional Therapies for Alcoholic Liver Disease. Clin. Liver Dis. 2016, 20, 535–550. [Google Scholar] [CrossRef] [PubMed]
- Medici, V.; Halsted, C.H. Folate, alcohol, and liver disease. Mol. Nutr. Food Res. 2013, 57, 596–606. [Google Scholar] [CrossRef] [PubMed]
- Pfeiffer, C.M.; Johnson, C.L.; Jain, R.B.; Yetley, E.A.; Picciano, M.F.; Rader, J.I.; Fisher, K.D. Mulinare J, Osterloh JD. Trends in blood folate and vitamin B-12 concentrations in the United States, 1988 2004. Am J Clin Nutr 2007, 86, 718–727. [Google Scholar] [CrossRef] [PubMed]
- Villanueva, J.A.; Halsted, C.H. Hepatic transmethylation reactions in micropigs with alcoholic liver disease. Hepatology 2004, 39, 1303–1310. [Google Scholar] [CrossRef] [PubMed]
- Bailey, S.M.; Robinson, G.; Pinner, A.; Chamlee, L.; Ulasova, E.; Pompilius, M.; Page, G.P.; Chhieng, D.; Jhala, N.; Landar, A.; et al. S-adenosylmethionine prevents chronic alcohol-induced mitochondrial dysfunction in the rat liver. Am. J. Physiol. Gastrointest. Liver Physiol. 2006, 291, G857–G867. [Google Scholar] [CrossRef]
- Sinha, S.; Kataria, A.; Kolla, B.P.; Thusius, N.; Loukianova, L.L. Wernicke Encephalopathy-Clinical Pearls. Mayo Clin. Proc. 2019, 94, 1065–1072. [Google Scholar] [CrossRef]
- Wilson, E.C.; Stanley, G.; Mirza, Z. The Long-Term Cost to the UK NHS and Social Services of Different Durations of IV Thiamine (Vitamin B1) for Chronic Alcohol Misusers with Symptoms of Wernicke’s Encephalopathy Presenting at the Emergency Department. Appl. Health Econ. Health Policy 2016, 14, 205–215. [Google Scholar] [CrossRef]
- Thompson, J.A.; Martinson, N.; Martinson, M. Mortality and costs associated with alcoholic hepatitis: A claims analysis of a commercially insured population. Alcohol. 2018, 71, 57–63. [Google Scholar] [CrossRef]
- Teschke, R. Alcoholic Liver Disease: Alcohol Metabolism, Cascade of Molecular Mechanisms, Cellular Targets, and Clinical Aspects. Biomedicines 2018, 6, 106. [Google Scholar] [CrossRef]
- Meroni, M.; Longo, M.; Rametta, R.; Dongiovanni, P. Genetic and Epigenetic Modifiers of Alcoholic Liver Disease. Int. J. Mol. Sci. 2018, 19, 3857. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Zhang, T.; Kusumanchi, P.; Han, S.; Yang, Z.; Liangpunsakul, S. Alcohol Metabolizing Enzymes, Microsomal Ethanol Oxidizing System, Cytochrome P450 2E1, Catalase, and Aldehyde Dehydrogenase in Alcohol-Associated Liver Disease. Biomedicines 2020, 8, 50. [Google Scholar] [CrossRef] [PubMed]
- Kong, L.Z.; Chandimali, N.; Han, Y.H.; Lee, D.H.; Kim, J.S.; Kim, S.U.; Kim, T.D.; Jeong, D.K.; Sun, H.N.; Lee, D.S.; et al. Pathogenesis, Early Diagnosis, and Therapeutic Management of Alcoholic Liver Disease. Int. J. Mol. Sci. 2019, 20, 2712. [Google Scholar] [CrossRef] [PubMed]
- Ceni, E.; Mello, T.; Galli, A. Pathogenesis of alcoholic liver disease: Role of oxidative metabolism. World J. Gastroenterol. 2014, 20, 17756–17772. [Google Scholar] [CrossRef] [PubMed]
- Boccuto, L.; Abenavoli, L. Genetic and Epigenetic Profile of Patients With Alcoholic Liver Disease. Ann. Hepatol. 2017, 16, 490–500. [Google Scholar] [CrossRef] [PubMed]
- Seitz, H.K.; Bataller, R.; Cortez-Pinto, H.; Gao, B.; Gual, A.; Lackner, C.; Mathurin, P.; Mueller, S.; Szabo, G.; Tsukamoto, H. Alcoholic liver disease. Nat. Rev. Dis. Primers 2018, 4, 16. [Google Scholar] [CrossRef] [PubMed]
- Tan, H.K.; Yates, E.; Lilly, K.; Dhanda, A.D. Oxidative stress in alcohol-related liver disease. World J. Hepatol. 2020, 12, 332–349. [Google Scholar] [CrossRef] [PubMed]
- Niederreiter, L.; Tilg, H. Cytokines and fatty liver diseases. Liver Res. 2018, 2, 14–20. [Google Scholar] [CrossRef]
- Abenavoli, L.; Scarlata, G.G.M.; Paravati, M.R.; Boccuto, L.; Luzza, F.; Scarpellini, E. Gut Microbiota and Liver Transplantation: Immune Mechanisms behind the Rejection. Biomedicines 2023, 11, 1792. [Google Scholar] [CrossRef]
- Cassard, A.M.; Ciocan, D. Microbiota, a key player in alcoholic liver disease. Clin. Mol. Hepatol. 2018, 24, 100–107. [Google Scholar] [CrossRef]
- Yan, AW.; Fouts, D.E.; Brandl, J.; Stärkel, P.; Torralba, M.; Schott, E.; Tsukamoto, H.; Nelson, K.E.; Brenner, D.A.; Schnabl, B. Enteric dysbiosis associated with a mouse model of alcoholic liver disease. Hepatology 2011, 53, 96–105. [Google Scholar] [CrossRef] [PubMed]
- Mutlu, E.A.; Gillevet, P.M.; Rangwala, H.; Sikaroodi, M.; Naqvi, A.; Engen, P.A.; Kwasny, M.; Lau, C.K.; Keshavarzian, A. Colonic microbiome is altered in alcoholism. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 302, G966–G978. [Google Scholar] [CrossRef] [PubMed]
- Szabo, G. Gut-liver axis in alcoholic liver disease. Gastroenterology 2015, 148, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Xie, G.; Zhong, W.; Zheng, X.; Li, Q.; Qiu, Y.; Li, H.; Chen, H.; Zhou, Z.; Jia, W. Chronic ethanol consumption alters mammalian gastrointestinal content metabolites. J. Proteome Res. 2013, 12, 3297–3306. [Google Scholar] [CrossRef]
- Dubinkina, V.B.; Tyakht, A.V.; Odintsova, V.Y.; Yarygin, K.S.; Kovarsky, B.A.; Pavlenko, A.V.; Ischenko, D.S.; Popenko, A.S.; Alexeev, D.G.; Taraskina, A.Y.; et al. Links of gut microbiota composition with alcohol dependence syndrome and alcoholic liver disease. Microbiome 2017, 5, 141. [Google Scholar] [CrossRef]
- Michelena, J.; Altamirano, J.; Abraldes, J.G.; Affò, S.; Morales-Ibanez, O.; Sancho-Bru, P.; Dominguez, M.; García-Pagán, J.C.; Fernández, J.; Arroyo, V.; Ginès, P.; et al. Systemic inflammatory response and serum lipopolysaccharide levels predict multiple organ failure and death in alcoholic hepatitis. Hepatology 2015, 62, 762–772. [Google Scholar] [CrossRef]
- van Loo, G.; Bertrand, M.J.M. Death by TNF: A road to inflammation. Nat. Rev. Immunol. 2023, 23, 289–303. [Google Scholar] [CrossRef]
- Mandrekar, P.; Szabo, G. Signalling pathways in alcohol-induced liver inflammation. J. Hepatol. 2009, 50, 1258–1266. [Google Scholar] [CrossRef]
- Shasthry, S.M.; Sarin, S.K. New treatment options for alcoholic hepatitis. World J. Gastroenterol. 2016, 22, 3892–3906. [Google Scholar] [CrossRef]
- Thakur, V.; Pritchard, M.T.; McMullen, M.R.; Wang, Q.; Nagy, L.E. Chronic ethanol feeding increases activation of NADPH oxidase by lipopolysaccharide in rat Kupffer cells: Role of increased reactive oxygen in LPS-stimulated ERK1/2 activation and TNF-alpha production. J. Leukoc. Biol. 2006, 79, 1348–1356. [Google Scholar] [CrossRef] [PubMed]
- Affò, S.; Dominguez, M.; Lozano, J.J.; Sancho-Bru, P.; Rodrigo-Torres, D.; Morales-Ibanez, O.; Moreno, M.; Millán, C.; Loaeza-del-Castillo, A.; Altamirano, J.; et al. Transcriptome analysis identifies TNF superfamily receptors as potential therapeutic targets in alcoholic hepatitis. Gut 2013, 62, 452–460. [Google Scholar] [CrossRef]
- Ciećko-Michalska, I.; Szczepanek, M.; Cibor, D.; Owczarek, D.; Skulina, D.; Szczepański, W.; Michalski, M. Zastosowanie oznaczania cytokin w ocenie rokowania u chorych z alkoholowa choroba watroby [Serum cytokine concentration as prognostic factor in patients with alcoholic liver disease]. Przegl Lek. 2006, 63, 249–252. [Google Scholar]
- Gonzalez-Quintela, A.; Campos, J.; Loidi, L.; Quinteiro, C.; Perez, L.F.; Gude, F. Serum TNF-alpha levels in relation to alcohol consumption and common TNF gene polymorphisms. Alcohol. 2008, 42, 513–518. [Google Scholar] [CrossRef] [PubMed]
- Mandrekar, P.; Bala, S.; Catalano, D.; Kodys, K.; Szabo, G. The opposite effects of acute and chronic alcohol on lipopolysaccharide-induced inflammation are linked to IRAK-M in human monocytes. J. Immunol. 2009, 183, 1320–1327. [Google Scholar] [CrossRef]
- Mandrekar, P.; Catalano, D.; White, B.; Szabo, G. Moderate alcohol intake in humans attenuates monocyte inflammatory responses: Inhibition of nuclear regulatory factor kappa B and induction of interleukin 10. Alcohol. Clin. Exp. Res. 2006, 30, 135–139. [Google Scholar] [CrossRef]
- Mookerjee, R.P.; Sen, S.; Davies, N.A.; Hodges, S.J.; Williams, R.; Jalan, R. Tumour necrosis factor alpha is an important mediator of portal and systemic haemodynamic derangements in alcoholic hepatitis. Gut 2003, 52, 1182–1187. [Google Scholar] [CrossRef]
- Diehl, A.M. Cytokine regulation of liver injury and repair. Immunol. Rev. 2000, 174, 160–171. [Google Scholar] [CrossRef] [PubMed]
- Yagi, S.; Hirata, M.; Miyachi, Y.; Uemoto, S. Liver Regeneration after Hepatectomy and Partial Liver Transplantation. Int. J. Mol. Sci. 2020, 21, 8414. [Google Scholar] [CrossRef]
- Jaeschke, H. Neutrophil-mediated tissue injury in alcoholic hepatitis. Alcohol. 2002, 27, 23–27. [Google Scholar] [CrossRef] [PubMed]
- Gao, B.; Tsukamoto, H. Inflammation in Alcoholic and Nonalcoholic Fatty Liver Disease: Friend or Foe? Gastroenterology 2016, 150, 1704–1709. [Google Scholar] [CrossRef] [PubMed]
- Patel, O.P.; Noor, M.T.; Kumar, R.; Thakur, B.S. Serum interleukin 8 and 12 levels predict severity and mortality in patients with alcoholic hepatitis. Indian. J. Gastroenterol. 2015, 34, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Wieser, V.; Adolph, T.E.; Enrich, B.; Kuliopulos, A.; Kaser, A.; Tilg, H.; Kaneider, N.C. Reversal of murine alcoholic steatohepatitis by pepducin-based functional blockade of interleukin-8 receptors. Gut 2017, 66, 930–938. [Google Scholar] [CrossRef] [PubMed]
- Nischalke, H.D.; Berger, C.; Lutz, P.; Langhans, B.; Wolter, F.; Eisenhardt, M.; Krämer, B.; Kokordelis, P.; Glässner, A.; Müller, T.; et al. Influence of the CXCL1 rs4074 A allele on alcohol induced cirrhosis and HCC in patients of European descent. PLoS ONE 2013, 8, e80848. [Google Scholar] [CrossRef] [PubMed]
- Chang, B.; Xu, M.J.; Zhou, Z.; Cai, Y.; Li, M.; Wang, W.; Feng, D.; Bertola, A.; Wang, H.; Kunos, G.; et al. Short- or long-term high-fat diet feeding plus acute ethanol binge synergistically induce acute liver injury in mice: An important role for CXCL1. Hepatology 2015, 62, 1070–1085. [Google Scholar] [CrossRef] [PubMed]
- Roh, Y.S.; Zhang, B.; Loomba, R.; Seki, E. TLR2 and TLR9 contribute to alcohol-mediated liver injury through induction of CXCL1 and neutrophil infiltration. Am. J. Physiol. Gastrointest. Liver Physiol. 2015, 309, G30–G41. [Google Scholar] [CrossRef]
- Tilg, H.; Moschen, A.R.; Szabo, G. Interleukin-1 and inflammasomes in alcoholic liver disease/acute alcoholic hepatitis and nonalcoholic fatty liver disease/nonalcoholic steatohepatitis. Hepatology 2016, 64, 955–965. [Google Scholar] [CrossRef] [PubMed]
- Knorr, J.; Wree, A.; Tacke, F.; Feldstein, A.E. The NLRP3 Inflammasome in Alcoholic and Nonalcoholic Steatohepatitis. Semin. Liver Dis. 2020, 40, 298–306. [Google Scholar] [CrossRef]
- He, Y.; Hara, H.; Núñez, G. Mechanism and Regulation of NLRP3 Inflammasome Activation. Trends Biochem. Sci. 2016, 41, 1012–1021. [Google Scholar] [CrossRef]
- Petrasek, J.; Bala, S.; Csak, T.; Lippai, D.; Kodys, K.; Menashy, V.; Barrieau, M.; Min, S.Y.; Kurt-Jones, E.A.; Szabo, G. IL-1 receptor antagonist ameliorates inflammasome-dependent alcoholic steatohepatitis in mice. J. Clin. Invest. 2012, 122, 3476–3489. [Google Scholar] [CrossRef]
- Cui, K.; Yan, G.; Xu, C.; Chen, Y.; Wang, J.; Zhou, R.; Bai, L.; Lian, Z.; Wei, H.; Sun, R.; et al. Invariant NKT cells promote alcohol-induced steatohepatitis through interleukin-1β in mice. J. Hepatol. 2015, 62, 1311–1318. [Google Scholar] [CrossRef] [PubMed]
- Voican, C.S.; Njiké-Nakseu, M.; Boujedidi, H.; Barri-Ova, N.; Bouchet-Delbos, L.; Agostini, H.; Maitre, S.; Prévot, S.; Cassard-Doulcier, A.M.; Naveau, S.; et al. Alcohol withdrawal alleviates adipose tissue inflammation in patients with alcoholic liver disease. Liver Int. 2015, 35, 967–978. [Google Scholar] [CrossRef] [PubMed]
- Alegre, F.; Pelegrin, P.; Feldstein, A.E. Inflammasomes in Liver Fibrosis. Semin. Liver Dis. 2017, 37, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V. NKT-cell subsets: Promoters and protectors in inflammatory liver disease. J. Hepatol. 2013, 59, 618–620. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.J.; Gao, B.; Zakhari, S.; Nagy, L.E. Inflammation in alcoholic liver disease. Annu. Rev. Nutr. 2012, 32, 343–368. [Google Scholar] [CrossRef] [PubMed]
- Mathews, S.; Feng, D.; Maricic, I.; Ju, C.; Kumar, V.; Gao, B. Invariant natural killer T cells contribute to chronic-plus-binge ethanol-mediated liver injury by promoting hepatic neutrophil infiltration. Cell Mol. Immunol. 2016, 13, 206–216. [Google Scholar] [CrossRef] [PubMed]
- Iracheta-Vellve, A.; Petrasek, J.; Gyogyosi, B.; Bala, S.; Csak, T.; Kodys, K.; Szabo, G. Interleukin-1 inhibition facilitates recovery from liver injury and promotes regeneration of hepatocytes in alcoholic hepatitis in mice. Liver Int. 2017, 37, 968–973. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Lafdil, F.; Kong, X.; Gao, B. Signal transducer and activator of transcription 3 in liver diseases: A novel therapeutic target. Int. J. Biol. Sci. 2011, 7, 536–550. [Google Scholar] [CrossRef]
- Ansari, R.A.; Husain, K.; Rizvi, S.A. Role of Transcription Factors in Steatohepatitis and Hypertension after Ethanol: The Epicenter of Metabolism. Biomolecules 2016, 6, 29. [Google Scholar] [CrossRef]
- Hong, F.; Kim, W.H.; Tian, Z.; Jaruga, B.; Ishac, E.; Shen, X.; Gao, B. Elevated interleukin-6 during ethanol consumption acts as a potential endogenous protective cytokine against ethanol-induced apoptosis in the liver: Involvement of induction of Bcl-2 and Bcl-x(L) proteins. Oncogene 2002, 21, 32–43. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Tachibana, S.; Wang, H.; Hisada, M.; Williams, G.M.; Gao, B.; Sun, Z. Interleukin-6 is an important mediator for mitochondrial DNA repair after alcoholic liver injury in mice. Hepatology 2010, 52, 2137–2147. [Google Scholar] [CrossRef]
- Kawaratani, H.; Moriya, K.; Namisaki, T.; Uejima, M.; Kitade, M.; Takeda, K.; Okura, Y.; Kaji, K.; Takaya, H.; Nishimura, N.; et al. Therapeutic strategies for alcoholic liver disease: Focusing on inflammation and fibrosis (Review). Int. J. Mol. Med. 2017, 40, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Klein, A.S.; Radaeva, S.; Hong, F.; El-Assal, O.; Pan, H.N.; Jaruga, B.; Batkai, S.; Hoshino, S.; Tian, Z.; et al. In vitro interleukin-6 treatment prevents mortality associated with fatty liver transplants in rats. Gastroenterology 2003, 125, 202–215. [Google Scholar] [CrossRef] [PubMed]
- El-Assal, O.; Hong, F.; Kim, W.H.; Radaeva, S.; Gao, B. IL-6-deficient mice are susceptible to ethanol-induced hepatic steatosis: IL-6 protects against ethanol-induced oxidative stress and mitochondrial permeability transition in the liver. Cell Mol. Immunol. 2004, 1, 205–211. [Google Scholar] [PubMed]
- Miller, A.M.; Wang, H.; Bertola, A.; Park, O.; Horiguchi, N.; Ki, SH.; Yin, S.; Lafdil, F.; Gao, B. Inflammation-associated interleukin-6/signal transducer and activator of transcription 3 activation ameliorates alcoholic and nonalcoholic fatty liver diseases in interleukin-10-deficient mice. Hepatology 2011, 54, 846–856. [Google Scholar] [CrossRef] [PubMed]
- Byun, J.S.; Suh, Y.G.; Yi, H.S.; Lee, Y.S.; Jeong, W.I. Activation of toll-like receptor 3 attenuates alcoholic liver injury by stimulating Kupffer cells and stellate cells to produce interleukin-10 in mice. J. Hepatol. 2013, 58, 342–349. [Google Scholar] [CrossRef] [PubMed]
- Yin, S.; Wang, H.; Park, O.; Wei, W.; Shen, J.; Gao, B. Enhanced liver regeneration in IL-10-deficient mice after partial hepatectomy via stimulating inflammatory response and activating hepatocyte STAT3. Am. J. Pathol. 2011, 178, 1614–1621. [Google Scholar] [CrossRef]
- Yang, A.M.; Wen, L.L.; Yang, C.S.; Wang, S.C.; Chen, C.S.; Bair, M.J. Interleukin 10 promoter haplotype is associated with alcoholic liver cirrhosis in Taiwanese patients. Kaohsiung J. Med. Sci. 2014, 30, 291–298. [Google Scholar] [CrossRef]
- Dudakov, J.A.; Hanash, A.M.; van den Brink, M.R. Interleukin-22: Immunobiology and pathology. Annu. Rev. Immunol. 2015, 33, 747–785. [Google Scholar] [CrossRef]
- Gao, B.; Xiang, X. Interleukin-22 from bench to bedside: A promising drug for epithelial repair. Cell Mol. Immunol. 2019, 16, 666–667. [Google Scholar] [CrossRef] [PubMed]
- Park, O.; Wang, H.; Weng, H.; Feigenbaum, L.; Li, H.; Yin, S.; Ki, S.H.; Yoo, S.H.; Dooley, S.; Wang, F.S.; et al. In vivo consequences of liver-specific interleukin-22 expression in mice: Implications for human liver disease progression. Hepatology 2011, 54, 252–261. [Google Scholar] [CrossRef] [PubMed]
- Meng, Y.X.; Zhao, R.; Huo, L.J. Interleukin-22 alleviates alcohol-associated hepatic fibrosis, inhibits autophagy, and suppresses the PI3K/AKT/mTOR pathway in mice. Alcohol. Clin. Exp. Res. (Hoboken) 2023, 47, 448–458. [Google Scholar] [CrossRef] [PubMed]
- Sagaram, M.; Frimodig, J.; Jayanty, D.; Hu, H.; Royer, A.J.; Bruner, R.; Kong, M.; Schwandt, M.L.; Vatsalya, V. One-month assessment of Th-cell axis related inflammatory cytokines, IL-17 and IL-22 and their role in alcohol-associated liver disease. Front. Immunol. 2023, 14, 1202267. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Li, J.; Li, Q.; Liang, Y.; Gao, J.; Meng, Z.; Li, P.; Yao, M.; Gu, J.; Tu, H.; et al. Environmental eustress promotes liver regeneration through the sympathetic regulation of type 1 innate lymphoid cells to increase IL-22 in mice. Hepatology 2023, 78, 136–149. [Google Scholar] [CrossRef] [PubMed]
- Hendrikx, T.; Duan, Y.; Wang, Y.; Oh, J.H.; Alexander, L.M.; Huang, W.; Stärkel, P.; Ho, S.B.; Gao, B.; Fiehn, O.; et al. Bacteria engineered to produce IL-22 in intestine induce expression of REG3G to reduce ethanol-induced liver disease in mice. Gut 2019, 68, 1504–1515. [Google Scholar] [CrossRef] [PubMed]
- Ehrmann, J.; Aiglová, K.; Urban, O.; Cveková, S.; Dvoran, P. Alcohol-related liver diseases (ALD). Vnitr. Lek. 2020, 66, 39–51. [Google Scholar] [CrossRef]
- Burnette, E.M.; Nieto, S.J.; Grodin, E.N.; Meredith, L.R.; Hurley, B.; Miotto, K.; Gillis, A.J.; Ray, L.A. Novel Agents for the Pharmacological Treatment of Alcohol Use Disorder. Drugs 2022, 82, 251–274. [Google Scholar] [CrossRef]
- Leggio, L.; Litten, R.Z. The GABA-B receptor agonist baclofen helps patients with alcohol use disorder: Why these findings matter. Neuropsychopharmacology 2021, 46, 2228–2229. [Google Scholar] [CrossRef]
- Lieber, S.R.; Rice, J.P.; Lucey, M.R.; Bataller, R. Controversies in clinical trials for alcoholic hepatitis. J. Hepatol. 2018, 68, 586–592. [Google Scholar] [CrossRef]
- Louvet, A.; Thursz, M.R.; Kim, D.J.; Labreuche, J.; Atkinson, S.R.; Sidhu, S.S.; O’Grady, J.G.; Akriviadis, E.; Sinakos, E.; Carithers, R.L. Jr.; et al. Corticosteroids Reduce Risk of Death Within 28 Days for Patients With Severe Alcoholic Hepatitis, Compared With Pentoxifylline or Placebo-a Meta-analysis of Individual Data From Controlled Trials. Gastroenterology 2018, 155, 458–468. [Google Scholar] [CrossRef]
- Dao, A.; Rangnekar, A.S. Steroids for Severe Alcoholic Hepatitis: More Risk Than Reward? Clin Liver Dis (Hoboken) 2019, 12, 151–153. [Google Scholar] [CrossRef]
- Lee, Y.S.; Kim, H.J.; Kim, J.H.; Yoo, Y.J.; Kim, T.S.; Kang, S.H.; Suh, S.J.; Joo, M.K.; Jung, Y.K.; Lee, B.J.; et al. Treatment of Severe Alcoholic Hepatitis With Corticosteroid, Pentoxifylline, or Dual Therapy: A Systematic Review and Meta-Analysis. J. Clin. Gastroenterol. 2017, 51, 364–377. [Google Scholar] [CrossRef] [PubMed]
- Thursz, M.R.; Richardson, P.; Allison, M.; Austin, A.; Bowers, M.; Day, C.P.; Downs, N.; Gleeson, D.; MacGilchrist, A.; Grant, A.; et al. Prednisolone or pentoxifylline for alcoholic hepatitis. N. Engl. J. Med. 2015, 372, 1619–1628. [Google Scholar] [CrossRef] [PubMed]
- Naveau, S.; Chollet-Martin, S.; Dharancy, S.; Mathurin, P.; Jouet, P.; Piquet, M.A.; Davion, T.; Oberti, F.; Broët, P.; Emilie, D.; et al. A double-blind randomized controlled trial of infliximab associated with prednisolone in acute alcoholic hepatitis. Hepatology 2004, 39, 1390–1397. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Kumar, A.; Sharma, B.C.; Sarin, S.K. Infliximab monotherapy for severe alcoholic hepatitis and predictors of survival: An open label trial. J. Hepatol. 2009, 50, 584–591. [Google Scholar] [CrossRef] [PubMed]
- Mackowiak, B.; Fu, Y.; Maccioni, L.; Gao, B. Alcohol-associated liver disease. J. Clin. Invest. 2024, 134, e176345. [Google Scholar] [CrossRef] [PubMed]
- Boetticher, N.C.; Peine, C.J.; Kwo, P.; Abrams, G.A.; Patel, T.; Aqel, B.; Boardman, L.; Gores, G.J.; Harmsen, W.S.; McClain, C.J.; et al. A randomized, double-blinded, placebo-controlled multicenter trial of etanercept in the treatment of alcoholic hepatitis. Gastroenterology 2008, 135, 1953–1960. [Google Scholar] [CrossRef] [PubMed]
- Scheiermann, P.; Bachmann, M.; Goren, I.; Zwissler, B.; Pfeilschifter, J.; Mühl, H. Application of interleukin-22 mediates protection in experimental acetaminophen-induced acute liver injury. Am. J. Pathol. 2013, 182, 1107–1113. [Google Scholar] [CrossRef]
- Xiang, X.; Feng, D.; Hwang, S.; Ren, T.; Wang, X.; Trojnar, E.; Matyas, C.; Mo, R.; Shang, D.; He, Y.; et al. Interleukin-22 ameliorates acute-on-chronic liver failure by reprogramming impaired regeneration pathways in mice. J. Hepatol. 2020, 72, 736–745. [Google Scholar] [CrossRef]
- Tang, K.Y.; Lickliter, J.; Huang, Z.H.; Xian, Z.S.; Chen, H.Y.; Huang, C.; Xiao, C.; Wang, Y.P.; Tan, Y.; Xu, L.F.; et al. Safety, pharmacokinetics, and biomarkers of F-652, a recombinant human interleukin-22 dimer, in healthy subjects. Cell Mol. Immunol. 2019, 16, 473–482. [Google Scholar] [CrossRef]
- Rothenberg, M.E.; Wang, Y.; Lekkerkerker, A.; Danilenko, D.M.; Maciuca, R.; Erickson, R.; Herman, A.; Stefanich, E.; Lu, T.T. Randomized Phase I Healthy Volunteer Study of UTTR1147A (IL-22Fc): A Potential Therapy for Epithelial Injury. Clin. Pharmacol. Ther. 2019, 105, 177–189. [Google Scholar] [CrossRef]
- Arab, J.P.; Sehrawat, T.S.; Simonetto, D.A.; Verma, V.K.; Feng, D.; Tang, T.; Dreyer, K.; Yan, X.; Daley, W.L.; Sanyal, A.; et al. An Open-Label, Dose-Escalation Study to Assess the Safety and Efficacy of IL-22 Agonist F-652 in Patients With Alcohol-associated Hepatitis. Hepatology 2020, 72, 441–453. [Google Scholar] [CrossRef]
- Jiang, R.; Tan, Z.; Deng, L.; Chen, Y.; Xia, Y.; Gao, Y.; Wang, X.; Sun, B. Interleukin-22 promotes human hepatocellular carcinoma by activation of STAT3. Hepatology 2011, 54, 900–909. [Google Scholar] [CrossRef]
- Park, O.; Wang, H.; Weng, H.; Feigenbaum, L.; Li, H.; Yin, S.; Ki, S.H.; Yoo, S.H.; Dooley, S.; Wang, F.S.; et al. In vivo consequences of liver-specific interleukin-22 expression in mice: Implications for human liver disease progression. Hepatology 2011, 54, 252–261. [Google Scholar] [CrossRef]
- Vergis, N.; Patel, V.; Bogdanowicz, K.; Czyzewska-Khan, J.; Fiorentino, F.; Day, E.; Cross, M.; Foster, N.; Lord, E.; Goldin, R.; et al. IL-1 Signal Inhibition In Alcoholic Hepatitis (ISAIAH): A study protocol for a multicentre, randomised, placebo-controlled trial to explore the potential benefits of canakinumab in the treatment of alcoholic hepatitis. Trials 2021, 22, 792. [Google Scholar] [CrossRef] [PubMed]
- Tu, W.; Gawrieh, S.; Dasarathy, S.; Mitchell, M.C.; Simonetto, D.A.; Patidar, K.R.; McClain, C.J.; Bataller, R.; Szabo, G.; Tang, Q.; et al. Design of a multicenter randomized clinical trial for treatment of Alcohol-Associated Hepatitis. Contemp. Clin. Trials Commun. 2023, 32, 101074. [Google Scholar] [CrossRef] [PubMed]
- Urien, S.; Bardin, C.; Bader-Meunier, B.; Mouy, R.; Compeyrot-Lacassagne, S.; Foissac, F.; Florkin, B.; Wouters, C.; Neven, B.; Treluyer, J.M.; et al. Anakinra pharmacokinetics in children and adolescents with systemic-onset juvenile idiopathic arthritis and autoinflammatory syndromes. BMC Pharmacol. Toxicol. 2013, 14, 40. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Liu, Z.; Bai, F. Roles of Gut Microbiota in Alcoholic Liver Disease. Int. J. Gen. Med. 2023, 16, 3735–3746. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Cheng, S.; Huo, J.; Dong, K.; Ding, Y.; Man, C.; Zhang, Y.; Jiang, Y. Lactobacillus plantarum J26 Alleviating Alcohol-Induced Liver Inflammation by Maintaining the Intestinal Barrier and Regulating MAPK Signaling Pathways. Nutrients 2022, 15, 190. [Google Scholar] [CrossRef] [PubMed]
- Ding, Q.; Cao, F.; Lai, S.; Zhuge, H.; Chang, K.; Valencak, T.G.; Liu, J.; Li, S.; Ren, D. Lactobacillus plantarum ZY08 relieves chronic alcohol-induced hepatic steatosis and liver injury in mice via restoring intestinal flora homeostasis. Food Res. Int. 2022, 157, 111259. [Google Scholar] [CrossRef]
- Vatsalya, V.; Feng, W.; Kong, M.; Hu, H.; Szabo, G.; McCullough, A.; Dasarathy, S.; Nagy, L.E.; Radaeva, S.; Barton, B.; et al. The Beneficial Effects of Lactobacillus GG Therapy on Liver and Drinking Assessments in Patients with Moderate Alcohol-Associated Hepatitis. Am. J. Gastroenterol. 2023, 118, 1457–1460. [Google Scholar] [CrossRef] [PubMed]
- Sidhu, S.S.; Dusseja, A.; Shalimar; Nijhawan, S.; Kapoor, D.; Goyal, O.; Kishore, H. A multicenter double-blind, placebo-controlled randomized trial to evaluate the safety and efficacy of bovine colostrum in the treatment of severe alcoholic hepatitis (SAH). Trials 2023, 24, 515. [Google Scholar] [CrossRef] [PubMed]
- Abenavoli, L.; Scarlata, G.G.; Scarpellini, E.; Procopio, A.C.; Ponziani, F.R.; Boccuto, L.; Cetkovic, N.; Luzza, F. Therapeutic success in primary biliary cholangitis and gut microbiota: A safe highway? Minerva Gastroenterol (Torino) 2024. [Google Scholar] [CrossRef] [PubMed]
- Abenavoli, L.; Montori, M.; Svegliati Baroni, G.; Argenziano, M.E.; Giorgi, F.; Scarlata, G.G.M.; Ponziani, F.; Scarpellini, E. Perspective on the Role of Gut Microbiome in the Treatment of Hepatocellular Carcinoma with Immune Checkpoint Inhibitors. Medicina (Kaunas) 2023, 59, 1427. [Google Scholar] [CrossRef]
Figure 1.
Schematic representation of the different pathogenetic ways involved in ALD development.
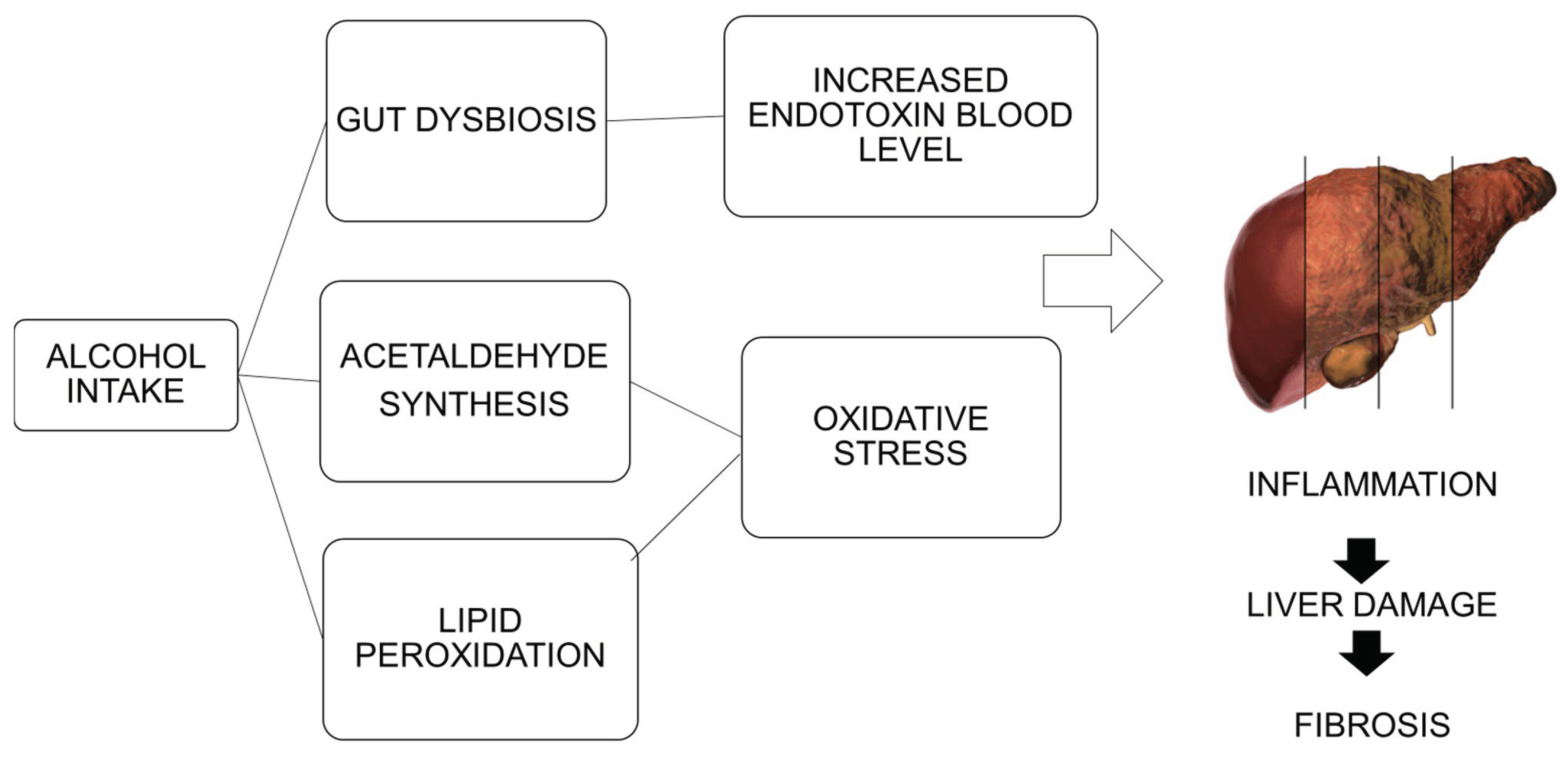

Table 1.
Summary of different studies about the involvement of pro-inflammatory cytokines in ALD.
| Cytokine evaluated | Reference | Study design | Outcome |
|---|---|---|---|
| TNF-α | Thakur et al., 2006 [53] | Pre-clinical study | Reduction of ERK1/2 activation and inhibition of NOX4 resulting in lower ROS and TNF-α production after the use of diphenyl-iodonium and dilinoleoyl-phosphatidylcholine in Kupffer cells of mice models |
| Ciećko-Michalska et al., 2006 [55] | Case-control study | Higher concentrations of TNF-α in ALD patients were correlated with poor prognosis | |
| Mandrekar et al., 2006 [58] | Pre-clinical study | Significant reduction in monocyte production of TNF-α in response to LPS or staphylococcal enterotoxin B stimulation eighteen hours after moderate alcohol consumption | |
| Gonzales-Quintela et al., 2008 [56] | Case-control study | TNF-α levels were almost similar in the general population, in teetotalers and alcohol drinkers, while elevated in chronic alcoholics | |
| Affò et al., 2013 [54] | Translational study | TNF superfamily receptors are overexpressed in AH humans and mice models | |
| IL-8 | Patel et al., 2015 [64] | Case-control study | Serum IL-8 predicts severity and mortality in patients with AH |
| Wieser et al., 2017 [65] | Pre-clinical study | Blockade of IL-8 receptors with pepducin reduced liver inflammation, weight loss and mortality associated with ALD in mice models | |
| CXCL1 | Nischalke et al., 2013 [66] | Case-control study | CXCL1 rs4074 single nucleotide polymorphism was associated with increased blood levels of CXCL1 and an increased risk of developing liver cirrhosis and HCC |
| Chang et al., 2015 [67] | Pre-clinical study | Mice models treated with a high-fat and ethanol-rich diet markedly upregulated the hepatic expression of CXCL1, with a reduction of the infiltration and hepatic damage of neutrophils | |
| Roh et al., 2015 [68] | Pre-clinical study | Ethanol-treated wild-type mice showed an increased hepatic expression of CXCL1 and serum level of CXCL1, while TLR2- and TLR9-deficient mice showed significantly lower levels | |
| IL-1β | Petrasek et al., 2012 [72] | Pre-clinical study | IL-1β levels were significantly increased in the liver of chronic alcohol-fed mice compared to controls |
| Cui et al., 2015 [73] | Pre-clinical study | Up-regulation of NLRP3 inflammasome components and IL-1β in Kupffer cells of ethanol-fed mice | |
| Voican et al., 2015 [74] | Prospective cohort study | Significantly increased levels of IL-1β in ALD patients after one week of alcohol withdrawal | |
| Mathews et al., 2016 [78] | Pre-clinical study | A loss of function of IL-1β protected mice models from hepatic infiltration of neutrophils and liver damage induced by chronic ethanol binges |
Table 2.
Summary of different studies about the involvement of anti-inflammatory cytokines in ALD.
| Cytokine evaluated | Reference | Study design | Outcome |
|---|---|---|---|
| IL-6 | Sun et al., 2003 [85] | Pre-clinical study | Pre-treatment with IL-6 induces hepatoprotection of steatotic liver isotransplants by preventing the apoptosis of sinusoidal endothelial cells, improving hepatic microcirculation and protecting against hepatocyte death in mice models |
| El-Assal et al., 2004 [86] | Pre-clinical study | IL-6 knockout mice fed with alcohol exhibited increased liver fat accumulation, lipid peroxidation, mitochondrial DNA damage and hepatocyte sensitization to TNFα-induced apoptosis | |
| Zhang et al., 2010 [83] | Pre-clinical study | IL-6 activates enzymes to repair mitochondrial DNA in hepatocytes damaged by chronic alcohol consumption in mice models | |
| Miller et al., 2011 [87] | Pre-clinical study | IL-10 knockout mice showed a more severe hepatic inflammatory response with higher levels of IL-6 and STAT3 activation, compared to wild-type mice, but lower liver steatosis severity and hepatocellular damage after higher alcohol levels or fatty dietary regimen | |
| IL-10 | Byun et al., 2013 [88] | Pre-clinical study | Treatment with polyinosinic:polycytidylic acid in vitro stimulates IL-10 production in HSCs through TLR-3 activation with reduction of alcoholic liver damage |
| Yang et al., 2014 [90] | Case-control study | IL-10 promoter polymorphisms allow the development of severe forms of alcoholic liver cirrhosis | |
| IL-22 | Meng et al., 2023 [94] | Pre-clinical study | Four-week treatment with IL-22 reduced liver fibrosis in mice models with ALD |
| Sagaram et al., 2023 [95] | Prospective cohort study | IL-22 levels were significantly increased in alcohol-associated non-severe hepatitis and alcohol-associated severe hepatitis patients compared to controls | |
| Liu et al., 2023 [96] | Pre-clinical study | IL-22 levels were significantly increased in murine models following liver regeneration |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



