Submitted:
24 February 2024
Posted:
01 March 2024
You are already at the latest version
Abstract
The adapted Posner spatial cue-target paradigm was used to explore the relationship between exogenous temporal attention caused by rhythm and endogenous and exogenous spatial attention. The study was divided into two experiments. In Experiment 1, 33 participants used arrow cues to orient endogenous spatial attention and rhythm to orient exogenous temporal attention. In Experiment 2, with 29 participants, exogenous spatial attention was directed by highlighting the cue box. Exogenous temporal attention was directed by rhythm. Experiment 1 indicated that RTs were faster for attended targets than the unattended. The main effect of temporal attention was significant, with RTs being faster for synchrony and late compared to early. RTs were faster for late compared to synchrony. The main effect of stimulus repetition was significant, with faster RTs for targets when the rhythmic cue consisted of five compared to four. The interaction between spatial attention and temporal attention was not significant. Experiment 2 demonstrated that exogenous spatial attention only exhibited IOR under early four stimulus repetitions. The main effect of temporal attention was significant, with RTs being faster for synchrony and late compared to early. The main effect of stimulus repetition was significant, with faster RTs for targets when the rhythmic cue consisted of five compared to four. The interaction between spatial attention and temporal attention was significant. The results indicated that rhythmic temporal attention supported foreperiod effects. Exogenous temporal attention and endogenous spatial attention independently affect behavior, while exogenous temporal attention and exogenous spatial attention mutually influenced behavior.
Keywords:
rhythm
; temporal attention
; endogenous spatial attention
; exogenous spatial attention
1. Introduction
When attention is directed towards space or time, both affect the perception of being stimulated by a target [1,2]. Posner first used the spatial cue goal paradigm to distinguish between two types of attentional orientation: endogenous and exogenous attention [3]. Studies show that endogenous attention to spatial location can enhance perceptual processing, as manifested by faster and more accurate responses to objects in the location being attended to [4]. Exogenous attention is usually guided by emergent stimuli in the peripheral space. When the time interval between the cue and the target is approximately 200-300 ms, participants respond more slowly to the attended target, which is called inhibition of return (IOR) [5], this phenomenon has been observed in detection [6,7] and discrimination tasks [8,9].
Temporal attention is the term used to describe time-based expectations of events. Several types of temporal structures, such as rhythms, can guide temporal attention [10]. Recurrent temporal structures can enable individuals to actively prepare for anticipated events, and repetitive rhythmic stimuli are often used in studies of auditory, visual, and tactile attention channels [11,12,13]. At the same time, the number of rhythmic stimuli also affects participants‘ responses. Jones adopted four and five repeated tactile cues, and found that the response was faster with five repeated tactile cues [11]. A similar pattern was found with more repetitions of rhythmic stimuli [14,15]. Similar to spatial attention, temporal attention can be divided into exogenous and endogenous attention [16]. Endogenous temporal attention can be studied using a temporal cue task employing a Posner-like cue-goal paradigm, in which temporal cues can be used to direct attention to a moment in time to predict an upcoming goal. Endogenous temporal attention can shorten reaction times and enhance the power of the alpha frequency band of the attended time [17,18]. Exogenous temporal attention is often triggered by stimuli in the environment, such as rhythms [19], which can trigger exogenous temporal attention without the need to pay attention to rhythmic events [20].
Dynamic attending theory (DAT) suggests that rhythms cause periodic fluctuations in attention, thereby regulating the gain of sensory input [21]. However, time anticipation can occur through the passage of time, even in the absence of external stimuli. The likelihood of an event occurring may change over time (e.g., the longer one waits, the more likely it is that a traffic light will change from red to green). This increasing expectation over time is called the hazard function [22]. The longer the time interval between the cue and the target, the faster the response, which is also known as the foreperiod effect. It is not clear how the time between the cue and target works [23]. Jones et al. found that when presented with regular tones, the accuracy of pitch judgments followed an inverted U-shaped pattern, with participants judging the expected tones with rhythm the most accurately, and the tones presented earlier or later than the rhythm the least accurately [24]. Herrmann et al. conducted a study using magnetoencephalography (MEG) to present participants with pitch sequences containing an intensity bias, and found that temporal attention could adjust the target detection threshold; that is, a stimulus presented in sync with the rhythm had a higher detection rate [25]. Jones adopted rhythmic tactile stimulation and found that the later the stimuli appeared, the shorter the stimulus response time, following the previous effect [11].
Spatial and temporal attention have been explored separately. However, studies on the relationship between the two have been inconsistent [2,26,27,28]. Studies on the relationship between temporal and spatial attention, originally observed by Doherty et al., manipulated temporal and spatial expectations by presenting a ball moving from left to right across an obscured area and found shorter reaction times when the ball reappeared behind the obscured area in sync with the previous rhythm [29]. Similarly, shorter reaction times were found when the ball reappeared in a spatial location predicted by the trajectory of the ball. When temporal and spatial attention were matched, the reaction time was the shortest, but there was no interaction between temporal and spatial attention in terms of accuracy and reaction time. Hence, the effects were independent. Rohenkohl et al. (2014) used arrow direction to indicate the target position and arrow color to indicate the time at which the target appeared, and found that temporal and spatial attention had a strong interaction in enhancing visual discrimination (d′), indicating that the effect of temporal attention depended on spatial position [30]. Using tactile cues, Jones (2019) found that there was no interaction between exogenous temporal and endogenous spatial attention, although there was an interaction between exogenous temporal and exogenous spatial attention [11]. Jones (2015) used the Posner cue task to explore stimulation with visual, auditory, and cross-modal audiovisual rhythms, and found that exogenous temporal attention and endogenous spatial attention did not interact [14]. Jones et al. found that temporal attention can improve recognition memory for spatially noticed objects [31].
In recent years, an increasing number of researchers have focused on temporal and spatial attention; however, the relationship between the two remains controversial. In conclusion, both the quantity and type of cues affect the relationship between temporal and spatial attention. Therefore, this study aimed to explore the relationship of temporal attention induced by rhythm with endogenous and exogenous spatial attention. It consisted of two experiments using the adapted Posner spatial cue target paradigm. Notably, this study did not explicitly manipulate endogenous temporal attention. Exogenous temporal attention used in this study included both the influence of rhythm, which can automatically generate the prediction of stimulus time, and the foreperiod effect, which is automatically driven by the passage of time. Experiment 1 explored the relationship between temporal and endogenous spatial attention. The clues were indicated by arrows, and the target appeared earlier, synchronously, or later than the rhythm cues. Experiment 2 explored the relationship between temporal and exogenous spatial attention. The cues were highlighted with a peripheral box, and the same target appeared earlier, synchronously, or later than the rhythm clues.
2. Experiment 1: Temporal Attention and Endogenous Spatial Attention
2.1. Participants
The required sample size was determined using G*power 3.1 [32]. A moderate effect size f = 0.25 and significance level α = 0.05 were used to determine the sample size. The results showed that 24 participants were required to achieve 95% statistical power. Thirty-three (12 female) people participated in the experiment (mean age 21.73 years; standard deviation 2.36 years); these were right-handed, and had normal or corrected vision. All participants voluntarily participated, signed informed consent forms, and received a small payment at the end of the experiment. The experiment has received a research ethics review from Tianjin Normal University.
2.2. Experimental Design
A 2 (spatial attention: attended/unattended) × 3 (temporal attention: early/synchronous/late) × 2 (number of repetitive rhythms: 4/5) three-factor within-subject design was used.
2.3. Experimental Procedure
Stimulus presentation and response data collection were performed using E-Prime 2.0.
The experiment included practice and the formal experiment the practice included 12 trials. The formal experiment consisted of 4 blocks, each containing 85 trials, for a total of 340 trials, including 240 spatially attended trials (71%), 80 spatially unattended trials (23%), and 20 capture trials (6%). Capture trials, in which no target appears after the cue, are designed to reduce the chance that if a target does not occur in sync with the rhythm, it will not necessarily appear in the latter interval and to ensure that the participant follows the instructions carefully. Excluding capture trials, the ratio of spatially attended to unattended conditions was 3:1. The early, synchronous, and late attempts were evenly distributed. There were 80 early, 80 synchronous, and 80 late spatially attended trials. For the spatially unattended trials, there were 27 early, 27 synchronous, and 26 late trials (balanced among the participants). In half of the trials, the rhythm cue consisted of four repeated stimuli, whereas in the other half it consisted of five repeated stimuli.
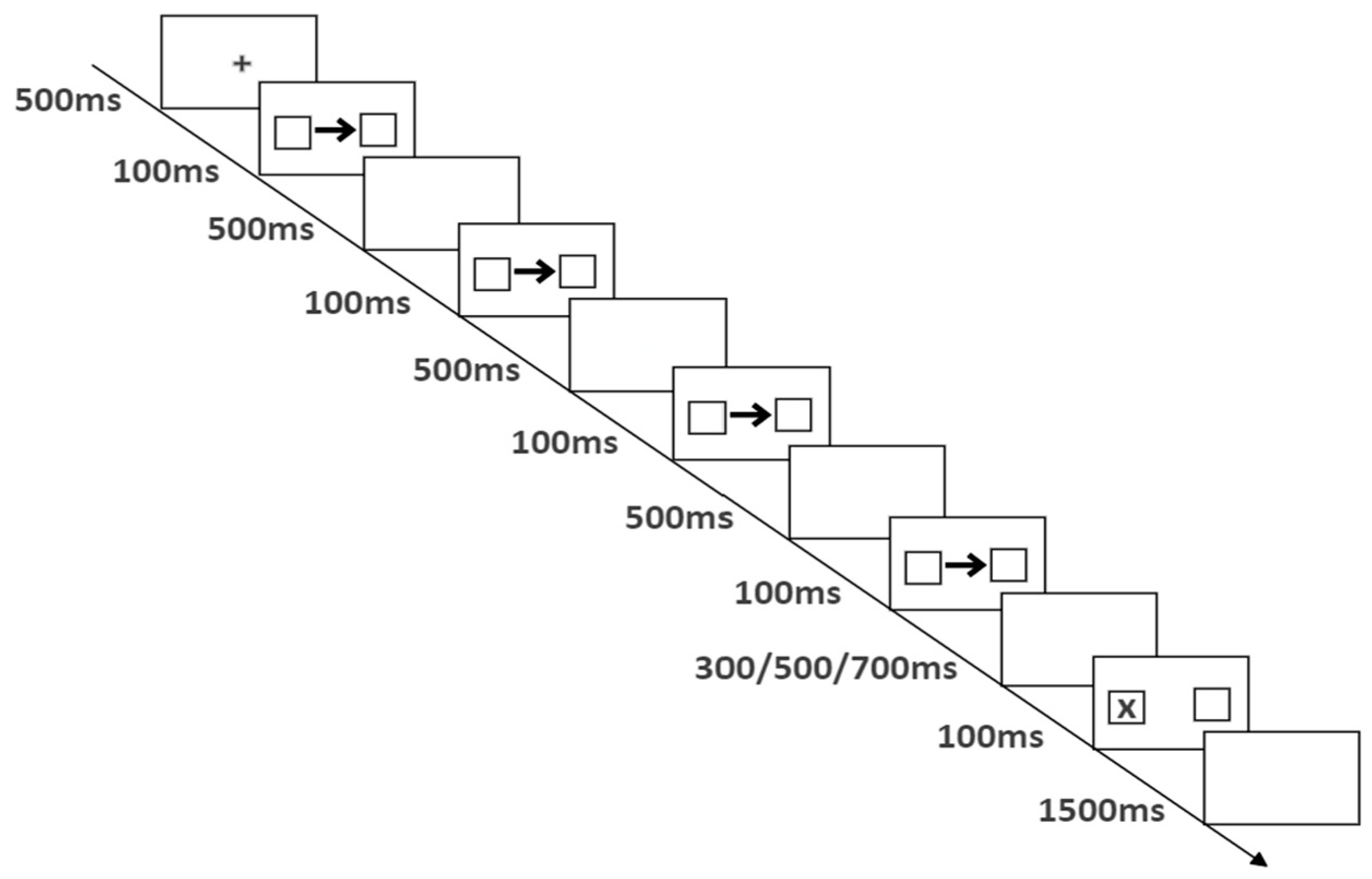
The flowchart of a single trial is shown in Figure 1. First, a 500 ms fixation point “+” was presented, followed by rhythmic cues. The cue was a left or right arrow, and there was a box with a viewing angle of 2°×2° on the left and right sides of the arrow. The first cue was presented for 100 ms, followed by a fixed stimulus interval of 500 ms, and then the cue was presented four or five times, producing a rhythm of 100 ms of stimuli every 600 ms; 300 ms (early), 500 ms (synchronous), or 700 ms (late) before the target, the target of 100 ms was presented, and the target was presented as “X” in the left or right box. The participants were asked to press the down arrow key on the keyboard to react when they saw the target, requiring them to react quickly and accurately. If there was no response, the attempt was transferred to the next attempt after 1500 ms. The trial interval varied randomly between 1000 and 2000 ms.
Figure 1.
Flow chart of experiment 1 (four repeated cues).
2.4. Data Analysis
SPSS 21.0 was used for the data analysis. Responses exceeding the average response by three standard deviations were eliminated, and the Greenhouse-Geisser method was used to correct for statistical effects that did not meet the sphericity test.
2.5. Experimental Results
2.5.1. Accuracy
Participants missed an average of 0.92% (SD = 3.19) of the targets and responded to an average of 6.06% (SD = 1.24) of the capture trials.
2.5.2. Reaction Time
Table 1 lists the means and standard deviations of the reactions under different experimental conditions. The main effect of spatial attention was significant (F(1, 32) = 32.81, p<0.001, η2p = 0.51), the reaction time in the attended condition (294 ms) was significantly shorter than in the unattended condition (313 ms). The main effect of temporal attention was significant (F(2, 64) = 94.80, p<0.001, η2p = 0.75), multiple comparison results showed that reaction times were significantly longer in the early (324 ms) than in the synchronous (302 ms) condition; reaction times were significantly longer in the early (324 ms) than in the late (285 ms) condition; reaction times were significantly longer in the synchronous (302 ms) than in the late (285 ms) condition. The main effect of repetitive rhythm number was significant (F(1, 32) = 65.40, p<0.001, η2p = 0.67); the reaction time was significantly longer under four (316 ms) than under five repetitions (291 ms). The interaction between temporal attention and repetitive rhythm number was significant (F(2, 64) = 8.59, p<0.001, η2p = 0.21); simple effects analysis showed a significant difference between four and five repetitions under early conditions (p<0.001); Under synchronous conditions, there was a significant difference between four and five repetitions (p<0.001); under late conditions, there was a significant difference between four and five repetitions (p<0.05). The interaction between spatial and temporal attention was not statistically significant (p = 0.78).

Table 1.
Average reaction time and standard deviation (ms) under different conditions in Experiment 1.
Table 1.
Average reaction time and standard deviation (ms) under different conditions in Experiment 1.
| spatial attented condition | spatial unattented condition | |||
| 4 | 5 | 4 | 5 | |
| early | 330±70 | 299±69 | 348±80 | 318±71 |
| synchronous | 309±71 | 278±62 | 327±69 | 296±72 |
| late | 280±67 | 269±71 | 302±74 | 289±66 |
3. Experiment 2: Temporal Attention and Exogenous Spatial Attention
3.1. Participants
The calculation method for the sample size was the same as in experiment 1. Twenty-nine (17 female) participants participated in the experiment (mean age 20.34 years; standard deviation 1.50 years); these were right-handed and had normal or corrected vision. All participants participated voluntarily, signed an informed consent form, and received a small payment at the end of the experiment. The experiment has received a research ethics review from Tianjin Normal University.
3.2. Experimental Design
A 2 (spatial attention: attended/unattended) × 3 (temporal attention: early/synchronous/late) × 2 (number of repetitive rhythms: 4/5) three-factor within-subject design was used.
3.3. Experimental Procedure
This was similar to Experiment 1, but with the following changes: The formal experiment consisted of four blocks, each block containing 85 trials, for a total of 340 trials, including 160 spatially attended condition trials (47%), 160 spatially unattended condition trials (47%), and 20 capture attempts (6%). Excluding capture attempts, the ratio of spatially attended/unattended trials was 1:1. Early, synchronous, and late trials were evenly distributed. For spatially attended trials, there were 53 early, 53 synchronous, and 54 late trials. For the spatially unattended trials, there were 53 early, 53 synchronous, and 54 late trials (averaged between the participants). In half of the trials, the rhythm cue consisted of four repeated stimuli, while in the other half, it consisted of five repeated stimuli.
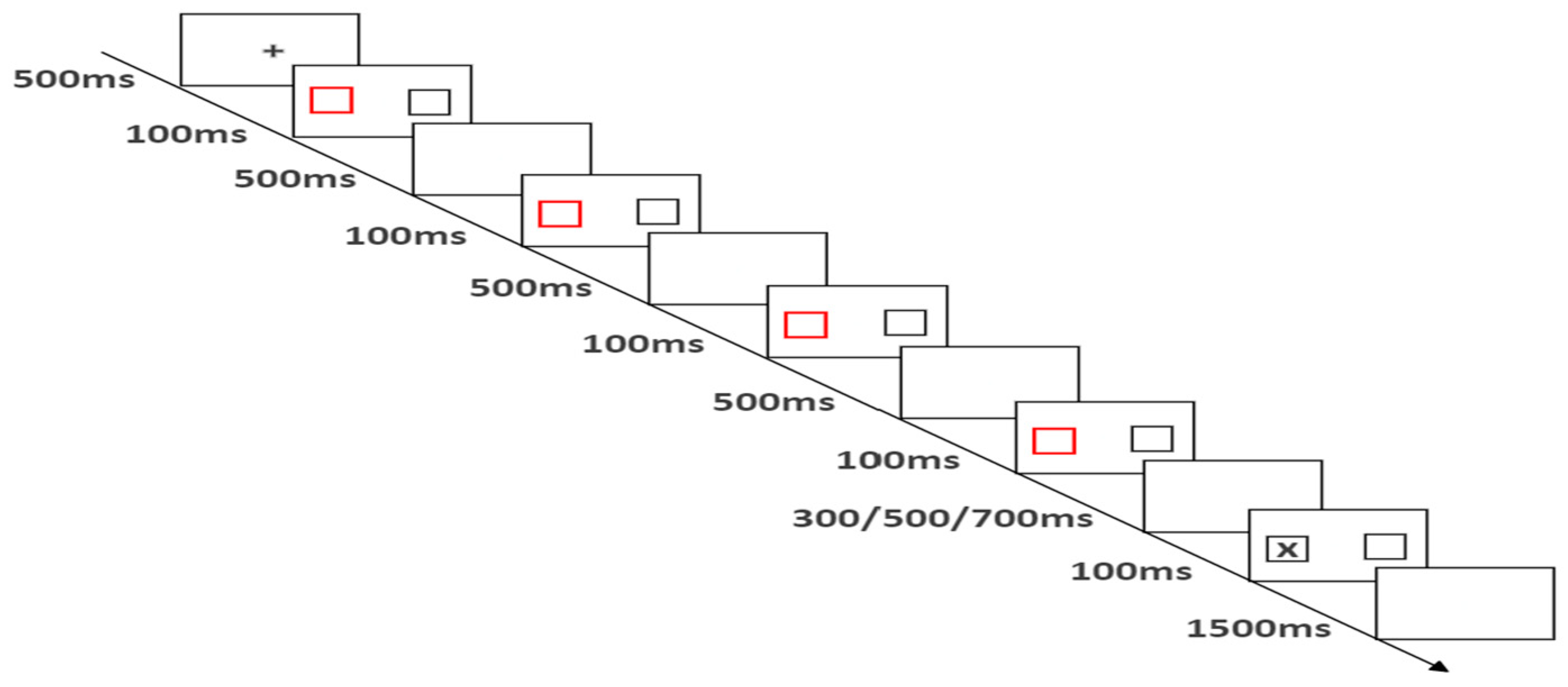
The process of a single experiment is shown in Figure 2: the same as in experiment 1, only the cue is changed to the random side of the left and right boxes and is highlighted, that is, the width becomes twice of the original, and the color is red.
Figure 2.
Flow chart of experiment 2 (four repeated cues).
3.4. Data Analysis
This was the same as in experiment 1.
3.5. Experimental Results
3.5.1. Accuracy
Participants missed an average of 1.73% (SD = 3.55) of the targets and responded to an average of 8.97% (SD = 3.49) of the capture trials.
3.5.2. Reaction Time
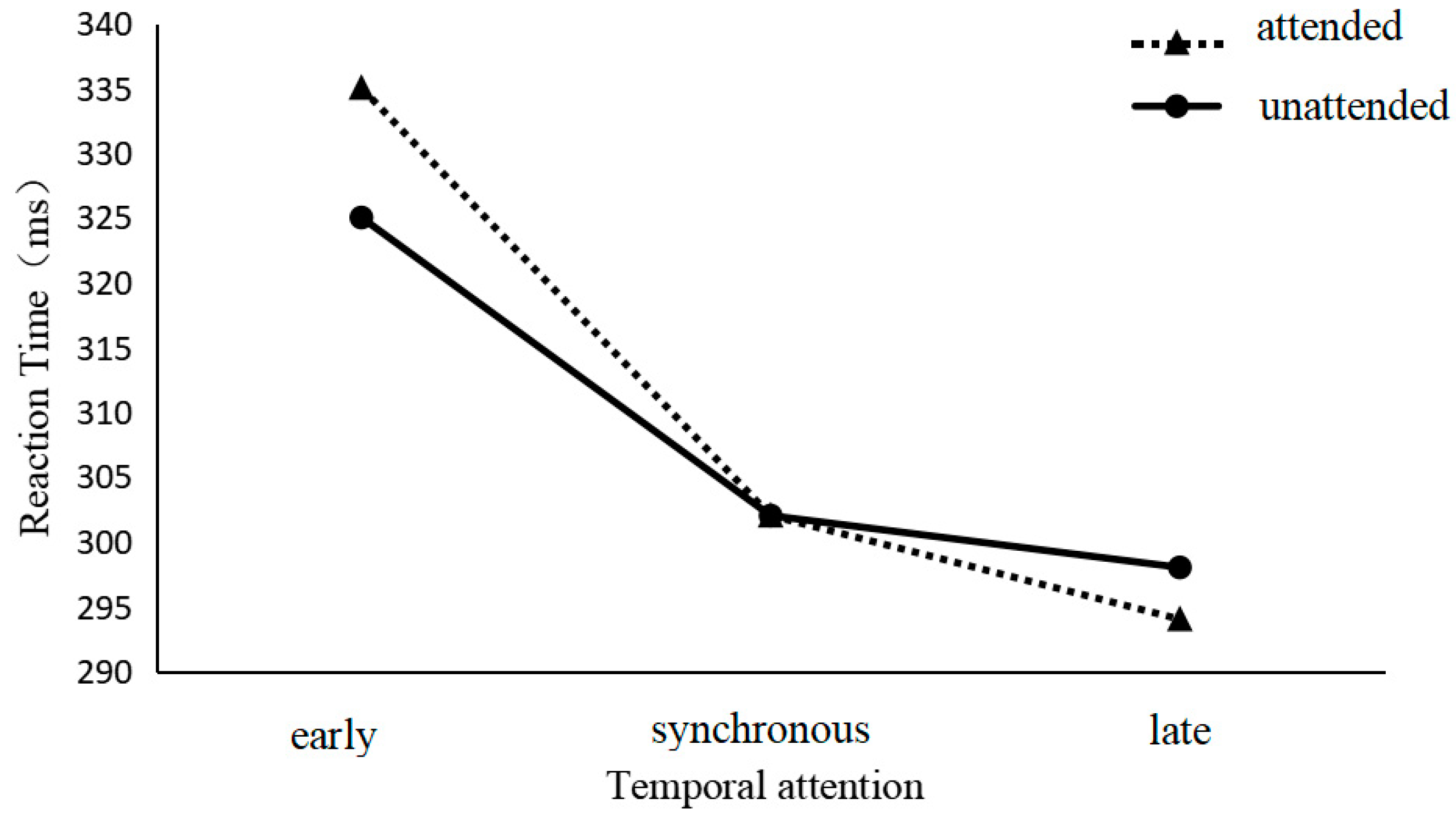
Table 2 lists the means and standard deviations of the reactions under different experimental conditions. The main effect of temporal attention was significant (F(1, 28) = 59.49, p<0.001, η2p = 0.68), the results of multiple comparisons showed that reaction times were significantly longer in the early (330 ms) than in the synchronous conditions (302 ms), and reaction times were significantly longer in the early (330 ms) than in the late conditions (296 ms). The main effect of repetitive rhythm number was significant (F(1, 28) = 96.56, p<0.001, η2p = 0.78); the reaction time was significantly longer under four (323 ms) than under five repetitions (295 ms). The interaction between spatial attention and temporal attention was significant (F(2, 56) = 4.08, p<0.05, η2p = 0.13). The interaction between spatial and temporal attention is illustrated in Figure 3. Simple effect analysis showed that there was a significant difference between spatially attended and non-attended subjects under early conditions (p<0.01), and there was no significant difference between spatially attended and non-attended trials under synchronous and late conditions (p = 0.80; p = 0.26). The interaction between temporal attention and repetitive rhythm number was significant (F(2, 56) = 33.74, p<0.001, η2p = 0.55). Simple effect analysis showed a significant difference between four and five repetitions under the early and synchronous conditions (p<0.001 and p<0.001, respectively), and there was no significant difference between four and five repetitions under the late condition (p = 0.11). The interaction of spatial attention, temporal attention, and repetitive rhythm number was significant (F(2, 56) = 4.14, p<0.05, η2p = 0.13). Simple effect analysis showed that spatial attention was significantly different only under the early-four repetitions (p<0.01); there was a significant difference in temporal attention under attended-four repetitions, attended-five repetitions, unattended-four repetitions, and unattended-five repetitions (F(2, 56) = 56.62, p<0.001, F(2, 56) = 10.45, p<0.001; F(2, 56) = 36.6, p<0.001; F(2, 56) = 7.44, p<0.01), showing the early was significantly longer than the synchronous and late conditions. The number of repetition rhythms was significantly different among the attended-early, attended-synchronous, unattended-early, and unattended-synchronous conditions (F (2, 56) = 101.23, p<0.001; F (2, 56) = 27.81, p<0.001; F (2, 56) = 61.04, p<0.001; F (2, 56) = 72.89, p<0.001).
Table 2.
Average reaction time and standard deviation (ms) under different conditions in Experiment 2.
Table 2.
Average reaction time and standard deviation (ms) under different conditions in Experiment 2.
| spatial attented condition | spatial unattented condition | |||
| 4 | 5 | 4 | 5 | |
| early | 362±61 | 307±43 | 345±45 | 306±41 |
| synchronous | 315±36 | 289±37 | 319±40 | 285±34 |
| late | 296±38 | 291±36 | 302±46 | 294±45 |
Figure 3.
Interaction between spatial and temporal attention.
4. Discussion
This study explored the relationship between temporal attention induced by rhythm and endogenous and exogenous spatial attention. The results showed that the interaction between rhythmic temporal attention and endogenous spatial attention was not significant, but the interaction between rhythmic temporal attention and exogenous spatial attention was significant.
Experiment 1 found that the main effect of spatial attention was significant and the reaction time under the attended condition was significantly shorter than that under the unattended condition, consistent with previous studies on endogenous spatial attention [14,33,34]. The main effect of the number of repetitive rhythms was significant, and the response time with four repetitions was significantly longer than that with five repetitions, indicating that participants responded faster to more likely targets, which is consistent with relevant studies using multiple repetitive rhythm numbers [11,17]. The main effect of temporal attention was significant, and the longer the interval time, the shorter the reaction time, which was consistent with the foreperiod effect; that is, the longer the time between the cue and target, the faster the response. The results of this study did not find an advantage of synchronous presentation in response time, which is consistent with some studies [35,36], but inconsistent with others [15,37,38]. The different numbers of repetitions in this study may have disrupted this rhythm, and thus, the foreperiod effect. The interaction between endogenous spatial attention and rhythmic temporal attention was not significant, which is consistent with studies on rhythm cues using visual, auditory, cross-modal audiovisual, and tactile stimuli, suggesting that endogenous spatial attention and exogenous temporal attention can function independently [11,14,29,39,40]. Consistent with this, Coull et al. found that different neural regions were involved [41]. However, some researchers have found an interaction between endogenous spatial and exogenous temporal attention, and this interaction is regulated by the power and phase of alpha, possibly due to the different experimental paradigms, that is, the researcher used a masking paradigm to detect threshold stimuli [42].
No main effect of spatial attention was found in Experiment 2, and no return on inhibition was found in previous studies [11,43,44,45]. Previous studies found that spatial attention exists in the form of IOR and responds more slowly to cued targets than to uncued ones. In this study, a repetitive time structure was used, which may be a rhythmic cue that affects the IOR phenomenon. There was a significant difference in spatial attention only under the early condition; that is, there was an exogenous spatial attention IOR under the early condition, but the IOR was no longer effective under the synchronous and late conditions. The main effect of the number of repetitions was significant and was significantly longer with four than with five repetitions. The main effect of temporal attention was significant and reaction times were significantly longer in the early than synchronous and late conditions. No difference was found between the synchronous and late conditions, possibly because synchronous exogenous temporal attention was balanced by the foreperiod effect. However, the overall trend was that reaction time decreased, which partially supports the foreperiod effect. As time passes, the participants’ perception of time changes [46,47], and when stimuli are presented rhythmically, this effect will be weakened or even disappear. This can be explained as entrainment by the rhythm, which will further affect the behavioral response [41,48]. The interaction between temporal attention and the number of repeated rhythms was significant, and the overall trend was that the difference between four and five repetitions was less significant the later the time, and the rate of decline in the reaction for five repetitions was not as fast as that for four repetitions, indicating that the effect of rhythm declined in the late period. The interaction between exogenous spatial attention and exogenous temporal attention was significant, which is consistent with some studies [11,30,49], which suggests that the effects of temporal attention depend on other coexisting properties, such as spatial location. The difference from Experiment 1 is that spatial attention is exogenous; while some studies have shown that endogenous attention involves more higher cortical brain regions, requires more cognitive resources, and the process is relatively slow [50,51,52], exogenous attention is more rapid and automated [53], which may be caused by task resource requirements.
In Experiments 1 and 2, the target interval time was fixed, but was consistent with the foreperiod effect, which could also indirectly prove that temporal rhythm guided exogenous temporal attention. However, both the dynamic attention theory and foreperiod effect predicted the longest early target response time, which was confirmed in Experiments 1 and 2. Neither Experiment 1 nor 2 found a U-shaped pattern predicted by dynamic attention theory. This is contradicted by some studies [25,38], although other studies have found the opposite result; that is, the synchronous reaction time is the longest [54,55]. Therefore, rhythmic stimuli influence behavior, although different task settings may influence the effects of dynamic attention theory [56]. Rhythm not only causes exogenous temporal attention but can also be used as an endogenous time cue to accelerate target detection. When the stimulus is out of sync with the rhythm, a foreperiod effect will appear. In both Experiments 1 and 2, the main effect of the repetitive rhythm number was significant, and the response with four repetitions was significantly longer than that with five repetitions. Owing to the influence of rhythm in the task, the response with five repetitions was more conducive to the formation of a regular time rhythm by the participants, which was consistent with previous studies [11,14]. At the same time, there was a higher likelihood of rhythm entrainment under the condition of five repetitions; therefore, the response time was shorter. It is also possible that rhythm produces some form of habituation [57]. Similar results have been obtained in auditory studies [14,58]. The interaction between temporal attention and number of repeated rhythms was significant. In Experiment 1, there was a significant difference between four and five repetitions under the early, synchronous, and late conditions, and in Experiment 2, there was a significant difference between four and five repetitions under the early and synchronous conditions, which also indicated that the role of rhythm was weakened. Experiment 2 found that spatial attention had a significant difference only under the early four repetitions, and the early stage was the time window of return of inhibition, while the intensity of exogenous and endogenous temporal attention was small; therefore, the response time was longer for the attended than for the unattended condition. Under the condition of five repetitions, exogenous temporal attention was enhanced, and synchronous, late exogenous, and endogenous temporal attention were enhanced, which could offset the IOR; therefore, the difference was not significant.
Although repetitive rhythm is not directly related to the task, it still contains information that the participants may indirectly use as a processing strategy, which can be seen from the main effect of the number of repetitive rhythms in Experiments 1 and 2. Therefore, the addition of endogenous temporal attention cannot be ruled out. Even though the separation of the two has been studied [41], these inconsistencies in previous studies may be attributed to the addition of endogenous temporal attention. In future, attention should be paid to the distinction between endogenous and exogenous temporal attention. The experimental paradigm needs to be improved, and further research should be conducted from a neuroscience perspective. Possible reasons for the dynamic change in temporal attention, such as the difference between endogenous and exogenous temporal attention and the number of repeated rhythms or habituation, require further research.
5. Conclusions
In this study, the influence of rhythmic temporal attention on target response and its relationship with spatial attention are discussed. The following conclusions were drawn. First, there were foreperiod effects in both endogenous and exogenous spatial attention. Second, the response to the target stimulus was promoted in endogenous spatial attention, regardless of whether it was early, synchronous, or late; that is, exogenous temporal attention and endogenous spatial attention independently influenced behavior. Third, there was a phenomenon of return inhibition in the early condition of exogenous spatial attention, and exogenous temporal attention and spatial attention affected each other’s behaviors.
Author Contributions
Conceptualization, Xiaoxiao Li and Yong Lyu; methodology, Xiaoxiao Li; software, Xiaoxiao Li; validation, Xiaoxiao Li and Yong Lyu; formal analysis, Xiaoxiao Li; investigation, Xiaoxiao Li and Yong Lyu; resources, Xiaoxiao Li; data curation, Xiaoxiao Li and Yong Lyu; writing—original draft preparation, Xiaoxiao Li; writing—review and editing, Xiaoxiao Li and Yong Lyu; visualization, Yong Lyu; supervision, Yong Lyu; project administration, Xiaoxiao Li and Yong Lyu; funding acquisition, Yong Lyu.
Funding
This research received no external funding.
Institutional Review Board Statement
The experiment has received a research ethics review from Tianjin Normal University.
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
Original data and programs have been conducted onn Science Data Bank (https://www.scidb.cn/surl/xlxb). DOI: https://doi.org/10.57760/sciencedb.13585
Acknowledgments
In this section, you can acknowledge any support given which is not covered by the author contribution or funding sections. This may include administrative and technical support, or donations in kind (e.g., materials used for experiments).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Meehan, C.E.; Wiesman, A.I.; Spooner, R.K.; Schantell, M.; Eastman, J.A.; Wilson, T.W. Differences in rhythmic neural activity supporting the temporal and spatial cueing of attention. Cereb. Cortex. 2021, 31, 4933–4944. [Google Scholar] [CrossRef] [PubMed]
- Nobre, A.C.; Rohenkohl, G. Time for the fourth dimension in attention. In: The Oxford Handbook of Attention (Nobre, A.C.; Kastner, S. eds), 2014; pp. 676–724. Oxford: Oxford UP.
- Posner, M.A. Orienting of attention. Q. J. Exp. Psychol. 1980, 32, 3–25. [Google Scholar] [CrossRef] [PubMed]
- Carrasco, M. Spatial covert attention: Perceptual modulation. In K. Nobre (Ed.), The Oxford Handbook of Attention, 2014, Oxford University Press.
- Klein, R.M. Inhibition of return. Trends. Cogn. Sci. 2000, 4, 138–147. [Google Scholar] [CrossRef]
- Chao, H.F.; Hsiao, F.S. Location-response binding and inhibition of return in a detection task. Atten. Percept. Psychophys. 2021, 83, 1992–2001. [Google Scholar] [CrossRef]
- Jones, A.; Forster, B. Reflexive attention in touch: An investigation of event related potentials and behavioural responses. Bio. Psychol. 2012, 89, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhang, M.; Wu, L.; Zhang, Q.; Wei, P. Neural mechanisms of reward-by-cueing interactions: ERP evidence. Front. Hum. Neurosci. 2021, 15, 608427. [Google Scholar] [CrossRef]
- Martín-Arévalo, E.; Lupiáñez, J.; Narganes-Pineda, C.; Marino, G.; Colás, I.; Chica, A.B. The Causal Role of the Left Parietal Lobe in Facilitation and Inhibition of Return. Cortex. 2019, 117, 311–322. [Google Scholar] [CrossRef]
- Nobre, A.C.; van Ede, F. Anticipated moments: Temporal structure in attention. Nat. Rev. Neurosc. 2018, 19, 34–48. [Google Scholar] [CrossRef]
- Jones, A. Temporal expectancies and rhythmic cueing in touch: The influence of spatial attention. Cognition. 2019, 182, 140–150. [Google Scholar] [CrossRef] [PubMed]
- Nozaradan, S.; Zerouali, Y.; Peretz, I.; Mouraux, A. Capturing with EEG the neural entrainment and coupling underlying sensorimotor synchronization to the beat. Cereb. Cortex. 2015, 25, 736–747. [Google Scholar] [CrossRef] [PubMed]
- Ongchoco, J.D.K.; Scholl, B.J. Hallucinating visual structure: Individual differences in ‘scaffolded attention’. Cognition. 2022, 225. [Google Scholar] [CrossRef]
- Jones, A. Independent effects of bottom-up temporal expectancy and top-down spatial attention. An audiovisual study using rhythmic cueing. Front. Integr. Neurosci. 2015, 8. [Google Scholar] [CrossRef]
- Mathewson, K.E.; Fabiani, M.; Gratton, G.; Beck, D.M.; Lleras, A. Rescuing stimuli from invisibility: Inducing a momentary release from visual masking with pre-target entrainment. Cognition. 2010, 115, 186–191. [Google Scholar] [CrossRef]
- Rohenkohl, G.; Coull, J.T.; Nobre, A.C. Behavioural dissociation between exogenous and endogenous temporal orienting of attention. PLoS. One. 2011, 6, e14620. [Google Scholar] [CrossRef]
- Pomper, U.; Keil, J.; Foxe, J.J.; Senkowski, D. Intersensory selective attention and temporal orienting operate in parallel and are instantiated in spatially distinct sensory and motor cortices. Hum. Brain. Mapp. 2015, 36, 3246–3259. [Google Scholar] [CrossRef]
- Rohenkohl, G.; Nobre, A.C. Alpha Oscillations Related to Anticipatory Attention Follow Temporal Expectations. J. Neurosci. 2011, 31, 14076–14084. [Google Scholar] [CrossRef]
- Lawrence, M.A.; Klein, R.M. Isolating exogenous and endogenous modes of temporal attention. J. Exp. Psychol. Gen. 2013, 142, 560–572. [Google Scholar] [CrossRef] [PubMed]
- Ball, F.; Michels, L.E.; Thiele, C.; Noesselt, T. The role of multisensory interplay in enabling temporal expectations. Cognition. 2018, 170, 130–146. [Google Scholar] [CrossRef]
- Large, E.W.; Jones, M.R. The dynamics of attending: How people track time-varying events. Psychol. Rev. 1999, 106, 119–159. [Google Scholar] [CrossRef]
- Nobre, A.C.; Correa, A.; Coull, J.T. The hazards of time. Curr. Opin. Neurobiol. 2007, 17, 465–470. [Google Scholar] [CrossRef] [PubMed]
- Jones, A.; Hsu, Y.F.; Granjon, L.; Waszak, F. Temporal expectancies driven by self- and externally generated rhythms. Neuroimage. 2017, 156, 352–362. [Google Scholar] [CrossRef]
- Jones, M.R.; Moynihan, H.; Mackenzie, N.; Puente, J. Temporal Aspects of Stimulus-Driven Attending in Dynamic Arrays. Psychol. Sci. 2002, 13, 313–319. [Google Scholar] [CrossRef]
- Herrmann, B.; Henry, M.J.; Haegens, S.; Obleser, J. Temporal expectations and neural amplitude fluctuations in auditory cortex interactively influence perception. NeuroImage. 2016, 124, 487–497. [Google Scholar] [CrossRef]
- MacKay, A.; Juola, J.F. Are spatial and temporal attention independent? Percept. Psychophys. 2007, 69, 972–979. [Google Scholar] [CrossRef]
- Nobre, A.C. How can temporal expectations bias perception and action. In: Attention and Time, eds A. C. Nobre and J. T. Coull (Oxford, UK: Oxford University Press), 2010; pp. 137–330.
- Sharp, P.; Melcher, D.; Hickey, C. Different effects of spatial and temporal attention on the integration and segregation of stimuli in time. Atten. Percept. Psychophys. 2019, 81, 433–441. [Google Scholar] [CrossRef]
- Doherty, J.R.; Rao, A.; Mesulam, M.M.; Nobre, A.C. Synergistic Effect of Combined Temporal and Spatial Expectations on Visual Attention. J. Neurosci. 2005, 25, 8259–8266. [Google Scholar] [CrossRef] [PubMed]
- Rohenkohl, G.; Gould, I.C. , Pessoa, J.; Nobre, A.C. Combining spatial and temporal expectations to improve visual perception. J. Vis. 2014, 14, 8. [Google Scholar] [CrossRef] [PubMed]
- Jones, A.; Ward, E.V.; Csiszer, E.L.; Szymczak, J. Temporal Expectation Improves Recognition Memory for Spatially Attended Objects. J. Cogn. Neurosci. 2022, 34, 1616–1629. [Google Scholar] [CrossRef] [PubMed]
- Faul, F.; Erdfelder, E.; Buchner, A.; Lang, A.G. Statistical power analyses using G*Power 3.1: Tests for correlation and regression analyses. Behav. Res. Methods 2009, 41, 1149–1160. [Google Scholar] [CrossRef] [PubMed]
- Jones, A.; Forster, B. Neural correlates of endogenous attention, exogenous attention and inhibition of return in touch. Eur. J. Neurosci. 2014, 40, 2389–2398. [Google Scholar] [CrossRef] [PubMed]
- Peelen, M.V.; Heslenfeld, D.J.; Theeuwes, J. Endogenous and exogenous attention shifts are mediated by the same large-scale neural network. NeuroImage. 2004, 22, 822–830. [Google Scholar] [CrossRef]
- Cravo, A.M.; Rohenkohl, G.; Wyart, V.; Nobre, A.C. Endogenous modulation of low frequency oscillations by temporal expectations. J. Neurophysiol. 2011, 106, 2964–2972. [Google Scholar] [CrossRef]
- van Ede, F.; de Lange, F.; Jensen, O.; Maris, E. Orienting attention to an upcoming tactile event involves a spatially and temporally specific modulation of sensorimotor alpha- and beta-band oscillations. J. Neurosci. 2011, 31, 2016–2024. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.R.; Johnston, H.M.; Puente, J. Effects of auditory pattern structure on anticipatory and reactive attending. Cogn. Psychol. 2006, 53, 59–96. [Google Scholar] [CrossRef]
- Lawrance, E.L.A.; Harper, N.S.; Cooke, J.E.; Schnupp, J.W.H. Temporal predictability enhances auditory detection. J. Acoust. Soc. Am. 2014, 135, EL357–EL363. [Google Scholar] [CrossRef]
- Rimmele, J.; Jolsvai, H.; Sussman, E. Auditory Target Detection Is Affected by Implicit Temporal and Spatial Expectations. J. Cogn. Neurosci. 2011, 23, 1136–1147. [Google Scholar] [CrossRef] [PubMed]
- Weinbach, N.; Shofty, I.; Gabay, S.; Henik, A. Endogenous temporal and spatial orienting: Evidence for two distinct attentional mechanisms. Psychon. Bull. Rev. 2015, 22, 967–973. [Google Scholar] [CrossRef]
- Coull, J.T.; Cotti, J.; Vidal, F. Differential roles for parietal and frontal cortices in fixed versus evolving temporal expectations: Dissociating prior from posterior temporal probabilities with fMRI. NeuroImage. 2016, 141, 40–51. [Google Scholar] [CrossRef]
- Kizuk, S.A.D.; Mathewson, K.E. Power and Phase of Alpha Oscillations Reveal an Interaction between Spatial and Temporal Visual Attention. J. Cogn. Neurosci. 2017, 29, 480–494. [Google Scholar] [CrossRef] [PubMed]
- Smith, T.J.; Henderson, J.M. Facilitation of return during scene viewing. Vis. Cogn. 2009, 17, 1083–1108. [Google Scholar] [CrossRef]
- Tang, X.; Gao, Y.; Yang, W.; Ren, Y.; Wu, J.; Zhang, M.; Wu, Q. Bimodal-divided attention attenuates visually induced inhibition of return with audiovisual targets. Exp. Brain. Res. 2019, 237, 1093–1107. [Google Scholar] [CrossRef]
- Tang, X.; Wang, X.; Peng, X.; Li, Q.; Zhang, C.; Wang, A.; Zhang, M. Electrophysiological evidence of different neural processing between visual and audiovisual inhibition of return. Sci. Rep. 2021, 11, 8056. [Google Scholar] [CrossRef] [PubMed]
- Muller-Gethmann, H.; Ulrich, R.; Rinkenauer, G. Locus of the effect of temporal preparation: Evidence from the lateralized readiness potential. Psychophysiology. 2003, 40, 597–611. [Google Scholar] [CrossRef] [PubMed]
- Schroter, H.; Birngruber, T.; Bratzke, D.; Miller, J.; Ulrich, R. Task predictability influences the variable foreperiod effect: Evidence of task-specific temporal preparation. Psychol. Res. 2015, 79, 230–237. [Google Scholar] [CrossRef] [PubMed]
- Ellis, R.J.; Jones, M.R. Rhythmic context modulates foreperiod effects. Atten. Percept. Psychophys. 2010, 72, 2274–2288. [Google Scholar] [CrossRef] [PubMed]
- Seibold, V.C.; Stepper, M.Y.; Rolke, B. Temporal attention boosts perceptual effects of spatial attention and feature-based attention. Brain. Cogn. 2020, 142, 105570. [Google Scholar] [CrossRef] [PubMed]
- Krauzlis. R.J.; Lovejoy, L.P.; Zenon, A. Superior colliculus and visual spatial attention. Ann. Rev. Neurosci. 2013, 36, 165–182. [Google Scholar] [CrossRef]
- Serences, J.T.; Yantis, S. Selective visual attention and perceptual coherence. Trends. Cogn. Sci. 2006, 10, 38–45. [Google Scholar] [CrossRef]
- Zhang, X.; Japee, S.; Safiullah, Z.; Mlynaryk, N.; Ungerleider, L.G. A normalization framework for emotional attention. PloS. Biol. 2016, 14. [Google Scholar] [CrossRef]
- Anton-Erxleben, K.; Carrasco, M. Attentional enhancement of spatial resolution: Linking behavioural and neurophysiological evidence. Nat. Rev. Neurosci. 2013, 14, 188–200. [Google Scholar] [CrossRef]
- Hickok, G.; Farahbod, H.; Saberi, K. The rhythm of perception: Entrainment to acoustic rhythms induces subsequent perceptual oscillation. Psychol. Sci. 2015, 26, 1006–1013. [Google Scholar] [CrossRef] [PubMed]
- Spaak, E.; de Lange, F.P.; Jensen, O. Local entrainment of alpha oscillations by visual stimuli causes cyclic modulation of perception. J. Neurosci. 2014, 34, 3536–3544. [Google Scholar] [CrossRef] [PubMed]
- Bauer, A.K.R.; Jaeger, M.; Thorne, J.D.; Bendixen, A.; Debener, S. The auditory dynamic attending theory revisited: A closer look at the pitch comparison task. Brain. Res. 2015, 1626, 198–210. [Google Scholar] [CrossRef] [PubMed]
- Dukewich, K.R. Reconceptualizing inhibition of return as habituation of the orienting response. Psychon. Bull. Rev. 2009, 16, 238–251. [Google Scholar] [CrossRef]
- McAuley, J.D.; Fromboluti, E.K. Attentional entrainment and perceived event duration. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2014, 369, 20130401. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



