
Submitted:
01 March 2024
Posted:
04 March 2024
You are already at the latest version
Abstract
Spodoptera frugiperda (J.E. Smith) is a global agricultural pest that poses a threat to crop production, characterized by its refractory polyphagous nature that makes it difficult to control and may lead to damage to woody plants. However, research on its adaptability to woody plants remains limited. This study compares the feeding adaptations (survival rate, pupation time, pupation rate, weight, length, and feeding volume), protective enzyme activities, and feeding preferences of S. frugiperda on leaves of six economically significant tree species (Areca catechu, Aquilaria sinensis, Cocos nucifera, Camellia oleifera, Dalbergia odorifera and Hevea brasiliensis) and maize as a control treatment. The results indicate that A. sinensis as the most vulnerable trees in high risk, H. brasiliensis and C. nucifera presented varying degrees of susceptibility, A. catechu, C. oleifera, and D. odorifera were unsuitability for S. frugiperda. In conclusion, this report extensively explores the feeding effects of S. frugiperda on six economically important tree species, and provides insights into the feeding preferences on these plants, thereby informing the potential threat posed by S. frugiperda to economically vital trees.
Keywords:
Spodoptera frugiperda
; woody plants
; economic tree species
; host plants
; adaptation
1. Introduction
Spodoptera frugiperda (Lepidoptera: Noctuidae), commonly known as the fall armyworm, is a migratory agricultural pest originating from tropical and subtropical regions of the Americas [1,2,3]. Its global significance stems from robust reproductive, adaptive, and migratory capacities, posing a formidable threat to crop production worldwide [4,5]. In January 2019, S. frugiperda infiltrated China's Yunnan Province from Myanmar for the first time, identified as the "maize type," currently posing a severe threat to Chinese food crop production safety [6,7]. Subsequently in April 2019 the pest invaded Hainan Province, which is a tropical area with the same climate with the origin of this pest, swiftly affecting nearly all maize planting areas and resulting in substantial losses [8,9].
The significant impact on agriculture has prompted comprehensive efforts to monitor, prevent, and control S. frugiperda in China. Comprehensive utilization of agricultural control technology, biological control technology, physical control technology, and chemical control technology has been made to reduce the adverse effects of this pest on Chinese agricultural development[10]. Despite its preference for Poaceae plants, this polyphagous pest has demonstrated the ability to feed on a diverse range of plant taxa, including herbs, vines, and woody vegetation, with reports of up to 353 species from 76 families as potential hosts [11,12]. Particularly concerning is the possibility that, with enhanced agricultural control measures, the pest may evolve into a major threat to forestry vegetation. Previous studies have identified various host plants, including woody species from families such as Arecaceae, Euphorbiaceae, and Fabaceae[12]. Experimental evidence, such as larvae feeding on tea plant leaves, further emphasizes the potential risk of S. frugiperda to woody plants after principal crop harvests[13]. Despite these findings, a lack of researchs remain regarding the specific adaptations of S. frugiperda to woody species.
Hainan Province is a tropical region with the same tropical and subtropical climate as the origin of this pest in the Americas. The six trees advocated by Hainan Province, including Areca catechu, Cocos nucifera, (both are Arecaceae), Hevea brasiliensis (Euphorbiaceae), Dalbergia odorifera (Fabaceae), Camellia oleifera (Theaceae) and Aquilaria sinensis (Thymelaeaceae) [14] have been expanded for planting. This not only improves the ecological and environmental quality, but also generates huge economic benefits. While possessing significant economic value, these trees have been reported to be susceptible damaged by Lepidoptera and Coleoptera pest [15,16]. Their main pests are described in Table S1. S. frugiperda as a Lepidoptera omnivorous pest, prompting concerns about the potential impact of infestation on these valuable trees.
This study aims to fill the current research gap by studying the potential threat posed by S. frugiperda to these six economically significant tree species in Hainan Province. A comprehensive understanding of the risk assessment associated with the pest's translocation among different hosts will inform Integrated Pest Management (IPM) strategies for S. frugiperda in both primary and alternate host plants.
2. Materials and Methods
2.1. Plants and Insects
Leaves from A. catechu, C. nucifera, D. odorifera and A. sinensis were sourced from the Yazhou Base of Sanya Academy of Tropical Agricultural Sciences, Sanya, Hainan Province, China (18.390246°N, 109.164020°E). Meanwhile, leaves of C.oleifera and H. brasiliensis were obtained from Danzhou Campus of Hainan University (19.507783°N, 109.495946°E). The maize (Zea mays) variety DK647, with seeds provided by Longping Biotech. Co., Ltd. (Sanya, China), served as the control treatment. The maize plants were cultivated in pots with nutrient soil (15 × 12 cm in diameter × height) in the greenhouse at the Yazhou Base of the Sanya Academy of Tropical Agricultural Sciences. All collected leaves were fresh and free from pest damage.
S. frugiperda eggs were originated from Longping Biotech. Co., Ltd (Sanya, China) and was consistently cultured in a climatic chamber (27 ± 1℃, 65 ± 3% RH, 16: 8h L:D). The eggs were placed in square plastic boxes (17.0 × 11.8 × 4.8 cm in length × width × height),covered with gauze to enhance insect respiration while prevent escape. Larvae in the 3rd instar were individually raised in cylindrical plastic boxes with lids (5.0 × 3.7 cm in diameter × height), each fitted with pinholes. The lids were punctured to facilitate adequate air circulation and simultaneously deter the larvae from escaping. After pupation, they were transferred to 100 mesh cages (Yiheng Scientific Instrument, Shanghai, China) (75 × 75 × 75 cm in length × width × height) for emergence, and a 10% honey water solution was provided to the adults for survival and reproduction. The 3rd instar larvae (7 days after hatching) were used for the experiment.
2.2. Feeding Patterns and Developmental Characteristics of S. frugiperda
The experiment encompassed seven treatments, involving the consumption of six distinct tropical tree species (A. catechu, C. nucifera, H. brasiliensis, D. odorifera, C. oleifera, A. sinensis), with Z. mays served as the control treatment. Leaves from each treatment underwent washing and cutting into 1 cm² squares for convenient measurement of leaf area according to the grid method [17]. Briefly, leaves were positioned on a transparent coordinate paper with a 1 mm2 grid. The vacant squares within a 1 cm2 area were counted, thus representing the leaf area (mm2). Third instar larvae from the same batch were individually cultured in cylindrical plastic boxes (5.0 × 3.7 cm in diameter × height), each equipped with pinholes to facilitate insect respiration and prevent escape. One larva was placed per box, and 30 replicates were conducted for each treatment. Daily replacements of leaves and cleans of insect feces were performed, feeding quantity changed as the quantity consumed increased. Mortality rates and feeding leaf areas were documented daily. Measurements of weight and length were taken by electronic scales (d = 0.001) (PL203, Metler Toledo) and industrial microscope (SZX16, Olympus Corporation) as larvae progressed to the pre-pupal stage. The eclosion time and gender of the adults were also recorded. The entire experiment was conducted in a climate chamber (27 ± 1℃, 65 ± 3% RH, 16: 8h L:D).
2.3. Determination of Enzyme Activity
Superoxide dismutase (SOD), and peroxidase (POD) activities were quantified using kits (respectively A007, A001, A084; Nanjing Jiancheng Bioengineering Institute, Nanjing, China). For enzyme activity determination, three samples were randomly selected from each of the seven treatments. Due to the low survival rate of 3rd instar larvae feeding on A. catechu, C. oleifera and D. odorifera observed in the bioassay experiment, the S. frugiperda larvae, which ate different plant leaves for 7d after the 3th instar in a climatic chamber (27 ± 1 °C, 65 ± 3 % RH, 16: 8h L:D), were collected and used for test enzyme activity. Larvae before the 3th instar were reared with maize leaves.
Before collection, larvae were firstly washed by ultrapure water, the surface water were wiped with chipless paper, then the body weights were recorded. The samples were placed in centrifuge tubes and stored at -80°C until testing. Mechanical grinding on ice at 4°C was performed by mixing the insects with 0.9% saline in a 1:9 (w:v) ratio. After thorough grinding, the samples were centrifuged for 10 minutes at 2500 rpm, the resulting supernatant (10% homogenized supernatant) was used for enzyme analysis following the manufacturer's instructions. Optical density (OD) values were measured using a microplate spectrophotometer SpectraMax ABS (Molecular Devices, USA), enzyme activities were calculated using the corresponding formulas. The tests were carried out in accordance with the manufacturer's instructions [18,19,20]. Corresponding formula:
where PC = Protein concentration; ODA, ODB, and ODS represent the OD values detected by the corresponding tubes (Blank (B), standard (S), and assay (A)); Ps (Protein standard solution) = 0.524 g/l ; N = dilution times.
where SOD activity (U/mgprot) = the quantity of SOD per mg of histone corresponding to 50% SOD inhibition in 1 ml of reaction solution is one SOD viability unit (U); POD activity (U/mgprot) = amount of enzyme 1 mg of histone catalyzing 1 μg of substrate per minute at 37°C. In the formula, ODC, ODA = the OD values detected by the corresponding tubes (control (C) and assay (A)); 235.65 = the reciprocal of the slope (instructions indicate direct use); VSD, VF = Volume of SD or final reaction liquid taken for experimental use; T = reaction time.
2.4. S. frugiperda Feeding Preferences
Feeding preference experiments were conducted to further indicate the feeding selection for S. frugiperda. Six distinct tree species were assessed using the leaf disk method [17,21]. Initially, a petri dish (90 mm in diameter) was evenly divided into six sectors of equal area. Leaves from various treatments were washed and cut into 1 cm² squares, which were randomly and sequentially positioned at the end of the dividing lines within the plastic petri dish. One 3rd instar larva of S. frugiperda, starved for more than 6 hours, was introduced into the center of each dish, and the dish was covered to prevent escape. After 6 hours, the leaf disk areas consumed by larvae on different plants were assessed using the counting grid method [17]. The feeding preference of larvae on different plants was quantified using the preference index, which was calculated as the percentage of the area of a plant's leaf disk consumed by larvae, relative to the sum of the areas of all leaf disks consumed. The preference performance was evaluated with 50 insects at a time and replicated three times. Experiments were conducted in a climatic chamber set at 27 ± 1℃, 65 ± 3% RH, and 16:8h L:D.
2.5. Data Analysis
Survival ratios of S. frugiperda among various plant treatments were analyzed using the Kaplan-Meier procedure and log-rank test. A full factorial LMER analysis, incorporating random factors for different plant treatments, time (days of measurement), and replications, was employed to analyze feeding leaf area, ANOVA with type III sum of squares (car package) was used for comparisons between treatments. For growth parameters, pupation rate, ecolosion rate, females rate between different treatments were subjected to analysis and comparison using the Chi-square test. Larval development time and pupal development time did not adhere to normal distribution were tested using the Kruskal-Wallis Test with Mann-Whitney U test. Enzyme activity assay data that did not conform to normal distribution were transformed using Blom in the normal score method. The remaining data, conforming to normality and homogeneous, were analyzed using one-way ANOVA with Tukey HSD test. All data are presented as mean ± standard error (SE). A significance level of P < 0.05 was considered for determining statistical significance. Data analysis was conducted using R 4.2.3.
3. Results
3.1. Variability in S. frugiperda Survival among Various Plant Species
A comprehensive analysis of survival percentages leading to eclosion of S. frugiperda reveals significant differences (χ2 = 187, P < 0.0001) when fed on leaves from seven distinct plant species (Figure 1). While no notable variations were observed in S. frugiperda survival within the A. sinensis group (χ2 = 0, P = 1) or the C. nucifera group (χ2 = 2.2, P = 0.1) compared to the maize control group, the survival rates of the A. catechu (χ2 = 39.2, P < 0.0001), C. oleifera (χ2 = 48, P < 0.0001), D. odorifera (χ2 = 40.2, P < 0.0001), and H. brasiliensis (χ2 = 8.3, P = 0.004) groups were dramatically lower than the control group.
Specifically, survival rates of S. frugiperda fed C. oleifera leaves were significantly lower than those of other groups (P < 0.0001). Furthermore, survival rates of S. frugiperda fed A. catechu and D. odorifera leaves were notably lower than those in the A. sinensis, C. nucifera, and H. brasiliensis groups (P < 0.01). Although H. brasiliensis exhibited lower survival rates than A. sinensis (χ2 = 8.4, P = 0.004), it was not significantly different from C. nucifera (χ2 = 0.9, P = 0.30). Furthermore, all individuals of S. frugiperda died within 10 days when fed on C. oleifera leaves, whereas those in the A. catechu and D. odorifera groups succumbed after 21 days.
3.2. Leaves Consumption of S. frugiperda
The daily leaf area consumption by S. frugiperda, feeding on various plant leaves, exhibited significant variability (χ2 = 114.727, P < 0.0001) (Figure 2). The interaction of time and food treatment was also found to be significant (χ2 = 690.289, P < 0.0001). Pairwise analyses were conducted for each food treatment separately. The overall trend of leaf area among groups was as follows: Z. mays, H. brasiliensis, A. sinensis, C. nucifera, D. odorifera, A. catechu, and C. oleifera. Considering the interaction, the leaf area consumed by S. frugiperda in the Z. mays control group was significantly greater than that in the groups fed on leaves from other plants (P < 0.05). This was followed by the A. sinensis and H. brasiliensis groups, which were not significantly different from each other (χ2 = 0.4046, P =0.5247), but they exhibited significantly larger leaf areas compared to the other treatments except control (P < 0.05). Parameters of daily leaf area consumption were not significantly different between C. nucifera, D. odorifera, C. oleifera (P > 0.05) and were significantly greater than A. catechu except for C. oleifera (P < 0.05).
3.3. Performance of S. frugiperda on Different Diets
The pupation rate of S. frugiperda feeding on various leaves varied significantly (χ2 = 128.75, P < 0.001) (Table 1). The pupation rate of A. catechu, C. oleifera, D. odorifera, and H. brasiliensis groups was significantly lower than that of Z. mays and A. sinensis (P < 0.05); additionally, A. catechu, C. oleifera, and D. odorifera were also significantly lower than H. brasiliensis and C. nucifera (P < 0.05), but H. brasiliensis and C. nucifera were not significantly different (P = 0.081). The eclosion rate of S. frugiperda varied significantly (χ2 = 110.34, P < 0.001) (Table 1), with A. sinensis (83.33%) and Z. mays (83.33%) groups being identical (P = 1) and significantly higher than other groups (P < 0.05); C. nucifera (46.67%) and H. brasiliensis (46.67%) were higher than the other three groups with 0% eclosion rate (A. catechu, C. oleifera, and D. odorifera groups) (P < 0.05).
There was a significant difference (U = 59.645, P < 0.001) in larvae devolonment time (Table 1). The larval development time is significantly shortest for Z. mays group, while C. nucifera group has relatively longer larval development times. The pupal development time was significantly different between groups (U = 38.268, P < 0.001). The pupal development time was significantly shorter in the C. nucifera group compared with other groups (P < 0.006), A. sinensis and H. brasiliensis have significantly longer pupal development times compared with other groups (P < 0.001). whereas there were no significant differences among A. sinensis and H. brasiliensis (P = 0.942). There was no significant divergence was found in pupal length (F (3, 77) = 1.757, P = 0.163) when S. frugiperda consumed various leaf diets (Table 1). A significant difference was observed in the female pupal fresh weight between different groups (F (3, 34) = 13.248, P < 0.001). Significantly heavier female pupal fresh weight was observed in the H. brasiliensis when compared with control Z. mays group (P = 0.02) and C. nucifera group (P = 0.001). Nevertheless, no statistically significant variations were detected in male pupal fresh weight (F (3,42) = 2.496, P = 0.074). Results reached statistical significance with the mean fresh pupal weight (F (3,77) = 12.253, P < 0.001). The mean fresh pupal weight in the A. sinensis group and H. brasiliensis were a significant divergence from Z. mays group (P = 0.07 compared with A. sinensis group, P = 0.032 compared with H. brasiliensis group) and C. nucifera group (P = 0.008 compared with A. sinensis group, P = 0.039 compared with H. brasiliensis group).
A remarkable difference in the sex ratio of successfully eclosed S. frugiperda was observed between treatments (χ2 = 13.874, P = 0.0031) (Table 1). The male ratio was highest in A. sinensis (76%), followed by H. brasiliensis (57.14%) and Z. mays (56%). The C. nucifera group had a significantly lower male ratio than A. sinensis (P < 0.001), H. brasiliensis (P = 0.048), and Z. mays group (P = 0.027), while the male percentage was higher in the A. sinensis (P = 0.232) and H. brasiliensis (P = 1) groups than in the Z. mays group, but the variation was not significant.
3.4. Enzyme Activity
Significant variations insuperoxide dismutase (SOD), and peroxidase (POD) activities in S. frugiperda were observed when consuming leaves from different plant species (SOD: F = 28.91, P < 0.0001; POD: F = 22.58, P < 0.0001).
For SOD activity (Figure 3A), A. catechu, A. sinensis, C. nucifera, C. oleifera, D. odorifera, and H. brasiliensis groups displayed significantly higher activity compared to the Z. mays control group. Among them, the A. catechu group exhibited the highest SOD activity, significantly surpassing other treatments (P < 0.05).
In the POD activity comparison (Figure 3B), the A. catechu group demonstrated higher activity than the control group (P < 0.0001) and other treatments (P < 0.05). Conversely, the C. nucifera group showed significantly lower activity than the Z. mays group (P < 0.0001) and other treatments (P < 0.05). Except for these two groups, no significant differences were observed between the remaining treatments in the pairwise comparisons (P > 0.05).
3.5. Feeding Preference Results of S. frugiperda
The feeding preference of 3rd instar S. frugiperda on the six plant species exhibited significant differences (F = 43.82, P < 0.0001) (Figure 4), with an overall trend from large to small observed in the following order: H. brasiliensis, C. nucifera, A. sinensis, A. catechu, C. oleifera, and D. odorifera groups. The preference index of larvae on H. brasiliensis leaves was significantly higher (P < 0.05) than that on other plants. C. nucifera showed a preference significantly larger than A. catechu, C. oleifera, and D. odorifera (P < 0.05) but did not differ significantly from A. sinensis (P > 0.05). There were no statistical differences between A. sinensis, A. catechu, C. oleifera, and D. odorifera groups (P > 0.05).
4. Discussion
The bio-experimentation on this study revealed significant differences in the performance of S. frugiperda across various tropical tree species. Notably, A. sinensis emerged as the most vulnerable to S. frugiperda infestation, followed by H. brasiliensis and C. nucifera. In contract, A. catechu, D. odorifera and C. oleifera were at low risk by the infestation of S. frugiperda.
A. sinensis is the main incense in China. Currently, the world's resource endangered species of Aquilaria have been listed in Appendix II of the Convention on International Trade in Endangered Species of Wild Fauna and Flora (CITES) [22]. It is an expensive and vital medicinal herb with an enormous financial value[23]. A. sinensis has been reported to be mainly affected by three pests: Heortia vitessoides (Lepidoptera), Dyspessa monticola (Coleoptera), and Anoplophora chinensis (Coleoptera) [24]. Host plant choice significantly influences phytophagous insect growth and development, with suitable hosts leading to higher survival rates and shorter developmental periods [25]. In current study, different host plants exerted distinct influences on the growth and development of S. frugiperda. In the experiment, the parameters of daily survival rate, pupation rate (86.67%), eclosion rate (83.33%), females rate (24%) of S. frugiperda with A. sinensis were not significantly differences compared to the control group (maize). The parameters of leaf area consumption, larval development time (14.36 ± 0.32d), pupal development time (10.72 ± 0.13d) in A. sinensis group were significantly inferior to maize group but performed well in comparison with other tree groups. Selectivity induced by plant volatiles plays a crucial role in determining feeding preference [26,27]. The observed feeding preference, as indicated by the preference index, also indicates A. sinensis as the most vulnerable trees in high risk. Therefore, we infer that A. sinensis is most likely to be victimized by S. frugiperda among the six trees.
In contrast, H. brasiliensis (Euphorbiaceae) and C. nucifera (Arecaceae) presented varying degrees of susceptibility. Despite not reaching the performance level of Z. mays, S. frugiperda feeding on the leaves of these two plants demonstrated better development. The female ratios of adults in H. brasiliensis and C. nucifera groups were not significantly different from those of the control and A. sinensis. The parameters of daily survival rate, pupation rate (50.00%), eclosion rate (46.67%) for H. brasiliensis group were significantly lower than those of Z. mays and A. sinensis, and did not differ from C. nucifera, but were higher than those of the other three treatments. The parameter of leaf area consumption, larval development time (13.27 ± 0.57d), pupal development time (10.79 ± 0.21d) in H. brasiliensis group is significantly longer than Z. mays, not different from A. sinensis, but shorter than C. nucifera. In addition to these, the preference index (63.21 ± 4.796%) of H. brasiliensis is significantly largest among the six trees. The parameters of daily survival rate, pupation rate (76.67%), eclosion rate (46.67%) in C. nucifera group are similar to the H. brasiliensis group, inferior to Z. mays and A. sinensis but superior to the other groups, but do not perform as well as H. brasiliensis on larval development time (16.13 ± 0.55d), pupal development time (9.00 ± 0.15d), preference index (34.52 ± 4.464%). They do not perform as well as the control and A. sinensis groups, but S. frugiperda can equally survive by feeding on them, so they are potentially at risk of being victimized. Besides, the pronounced feeding preference of S. frugiperda for H. brasiliensis underscores its potential as a susceptible host. These results suggest C. nucifera and H. brasiliensis also pose risks, albeit to a lesser extent.
Larval mortality, as a crucial indicator of host suitability [28], was notably higher in the A. catechu, C. oleifera, and D. odorifera groups, which had zero survival rate (0%), so there's no data on pupal mean fresh weight, pupal development time, etc. indicating their unsuitability for S. frugiperda survival. And their parameters of pupation rate (0% or 3.33%), eclosion rate (0%), and daily leaf area consumption also significantly lower, thus indicating that A. catechu, C. oleifera, and D. odorifera are unsuitable for the survival of S. frugiperda.
The activity of protective enzymes in insects is an important indicator for evaluating the physiological and biochemical responses of organisms to toxic substances [29,30,31,32]. Superoxide dismutase (SOD) and peroxidase (POD) play crucial roles as antioxidant enzymes, protecting the system from peroxidation and maintaining cellular redox balance [33]. In our study, there were significant differences in SOD and POD activities in S. frugiperda among different plant leaves. The SOD and POD enzyme activities of the A. catechu treatment were significantly higher than those of the other treatments, and the SOD enzyme activities of the other leaf treatments were significantly higher than those of maize control treatment. However, except for the significantly lower POD enzyme activity of the C. nucifera treatment group compared to the maize control treatment, there were no significant differences in POD enzyme activity among the other leaf treatments. This study further demonstrated the adaptability of S. frugiperda to different tree species by detecting the activities of SOD and POD enzyme activities in the body that feed on different leaf treatments. The results of enzyme activity indicators show that, except for the strong toxicity of A. catechu to S. frugiperda, which is not suitable for the survival, other tree species may be eaten by S. frugiperda, especially C. nucifera.
Historically, omnivorous pests have exhibited a pattern of shifting to new host plants, eventually establishing themselves as dominant species. A case in point is the Colorado potato beetle, which originally infested wild lycophytes but swiftly transitioned to the potato as a primary host plant with the introduction of potato cultivation [34,35]. This phenomenon is not isolated, as other omnivorous pests like Polyphagotarsonemus latus, Caloptilia theory, Lopholeucaspis japonica, and Buzura suppressaria have been observed transitioning to tea trees after a period of acclimatization, becoming significant concerns for tea tree growers [36]. Notably, the mirid bug outbreak correlated with the widespread adoption of Bt cotton in China serves as an illustrative example of the potential risks associated with large-scale planting of genetically modified (GM) crops [37].
S. frugiperda, recognized as an omnivorous pest, is documented by the Center for Agriculture and Bioscience International (CABI) as a host for 353 species across 76 plant families [38]. This extensive list, compiled by Montezano and others through a thorough literature review and a survey of the Brazilian ground [12], reveals the broad range of plants susceptible to infestation by this pest. In previous results, C. nucifera and H. brasiliensis were identified as host plants in Brazil [39,40,41], aligning with the findings of our study. Up to now, these plants were not reported as host plants of S. frugiperda in China, suggesting that they may not have been scientifically investigated in the region. Notably, the specific adaptation of S. frugiperda to A. sinensis among those six trees examined in our experiment was not documented in previous surveys. The omission could be attributed to A. sinensis being an economically significant evergreen tree species native to China [42]. While there are no existing reports on whether A. sinensis has been victimized by S. frugiperda, the potential risk remains a matter of concern.
An intriguing discovery emerged during our investigation - C. oleifera demonstrated inhibitory effects on the growth and development of S. frugiperda, resulting in the mortality of all larvae within 10 days. This suggests that C. oleifera may contain toxic substances with lethal effects on insects. Notably, C. oleifera saponins are proposed as the potential agents responsible for this observed mortality, indicating their potential utility as natural plant-derived insecticides for controlling S. frugiperda [43]. The discovery of C. oleifera's inhibitory effect on S. frugiperda suggests a potential natural plant-derived insecticide. Further investigation into the toxic substances and mechanisms involved in this inhibition could contribute to the development of sustainable pest control strategies.
5. Conclusions
In conclusion, this study sheds light on the intricate interactions between S. frugiperda and various tropical tree species, offering insights into feeding preferences, adaptive responses, and potential risks. The findings underscore the importance of continued research in understanding the dynamics of herbivorous pests, especially in the context of changing agricultural practices and the introduction of genetically modified crops.
Supplementary Materials
Funding
The project was supported by the Hainan Provincial Natural Science Foundation of China (323QN196), the China Scholarship Council (No.202107565020), the Funding for Postdoctoral Research Projects in Hainan Province in 2022 (307044), the Project of Sanya Yazhou Bay Science and Technology City (SCKJ-JYRC-2023-69).
Author Contribution
Y. Y. and Y. C designed the study and J. X. conducted the experiments. Y. Y., Y. C. and J. X. analyzed the data, and drafted the original manuscript. J. X. made the figures, X. K., , R. J., X. J., J. G., and Y. G. provided the experimental materials. All authors contributed to the drafting of the final manuscript.
Data Availability
All data presented in this manuscript are available in the online supplementary materials.
Conflict of Interest
The authors declare that they have no conflict of interest.
References
- Sparks, A.N. A Review of the Biology of the Fall Armyworm. The Florida Entomologist 1979, 62, 82. [Google Scholar] [CrossRef]
- Shi, W.P. How greedy are the fall armyworm? Journal of Plant Protection 2020, 47, 687–691. [Google Scholar] [CrossRef]
- Kenis, M. Prospects for Classical Biological Control of Spodoptera Frugiperda (Lepidoptera: Noctuidae) in Invaded Areas Using Parasitoids from the Americas. J Econ Entomol 2023, 116, 331–341. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Hu, C.; Jia, H.; Wu, Q.; Shen, X.; Zhao, S.; Jiang, Y.; Wu, K. Case Study on the First Immigration of Fall Armyworm, Spodoptera Frugiperda Invading into China. Journal of Integrative Agriculture 2021, 20, 664–672. [Google Scholar] [CrossRef]
- Wu, K.M. Management strategies of fall armyworm (Spodoptera frugiperda) in China. Plant Protection 2020, 46, 1–5. [Google Scholar] [CrossRef]
- Tang, J.; Lu, B.; Lu, H.; Ji, X.; Yang, P.; Su, H.; Cai, B. Investigation and Preliminary Study of Biological Characteristic of Parasitic Wasps of Spodoptera frugiperda in Hainan. Chinese Journal of Tropical Crops 2020, 41, 1189–1195. [Google Scholar] [CrossRef]
- Zhang, L.; Liu, B.; Jiang, Y.Y.; Liu, J.; Wu, K.M.; Xiao, Y.T. Molecular characterization analysis of fall armyworm populations in China. Plant Protection 2019, 45, 20–27. [Google Scholar] [CrossRef]
- Liu, J.; Jiang, Y.Y.; Wu, Q.L.; Zhao, S.Y.; Li, H. Characteristics of noctuid moth infestation in grassland in China in winter and spring, and analysis of the occurrence trend in the second half of the year. China Plant Protection 2019, 39, 36–38+49. [Google Scholar] [CrossRef]
- Jiang, Y.Y.; Liu, J.; Xie, M.C.; Li, Y.H.; Yang, J.J.; Zhang, M.L.; Qiu, K. Observation on law of diffusion damage of Spodoptera frugiperda in China in 2019. Plant Protection 2019, 45, 10–19. [Google Scholar] [CrossRef]
- Fen, R.S. Characteristics of the Occurrence and Control Measures of Spodoptera frugiperda. Seed Science 2023, 41, 116–118. [Google Scholar] [CrossRef]
- Juárez, M.L.; Socías, M.G.; Murúa, M.G.; Prieto, S.; Medina, S.; Willink, E. Revisión de los hospederos del gusano cogollero del maíz, Spodoptera frugiperda (Lepidoptera: Noctuidae). Rev. Soc. Entomol. Arg. 2010, 69, 3–4. [Google Scholar]
- Montezano, D.G.; Specht, A.; Sosa-Gómez, D.R.; Roque-Specht, V.F.; Sousa-Silva, J.C.; Paula-Moraes, S.V.; Peterson, J.A.; Hunt, T.E. Host Plants of Spodoptera Frugiperda (Lepidoptera: Noctuidae) in the Americas. African Entomology 2018, 26, 286–300. [Google Scholar] [CrossRef]
- Sun, R.; Gao, L.; Mi, Z.; Zheng, Y.; Li, D. CnMADS1, a MADS Transcription Factor, Positively Modulates Cell Proliferation and Lipid Metabolism in the Endosperm of Coconut (Cocos Nucifera L.). Planta 2020, 252, 83. [Google Scholar] [CrossRef] [PubMed]
- Li, M. Planting the Six Trees and Finding the Power Points. Hainan Daily 2022, 5, news.hndaily.cn/html/2022–04/27/content_58468_14759402htm. [Google Scholar]
- Wu, C.B.; Ren, C.C.; Zhu, M.J.; Du, R.K.; Yan, C.Z.; Han, S.; Rui, K.; Lu, C.J. Efficacy of a combination of carbofuran and azoxystrobin against red-veined spike borer of betel nut. China Plant Protection 2022, 42, 83–85. [Google Scholar] [CrossRef]
- Meng, X.L.; Song, W.W.; Tang, Q.H.; Niu, X.Q.; Li, C.X.; Zhong, B.Z.; Lv, C.J.; Huang, S.C.; Qin, W.Q. Advances in Main Diseases and Insect Pests of Areca Palm. Chinese Journal of Tropical Crops 2021, 42, 3055–3065. [Google Scholar] [CrossRef]
- Tang, Q.B.; Wang, C.Z. Leaf disctest used in caterpillar feeding preference study. Chinese Bulletin of Entomology 2007, 912–915. [Google Scholar] [CrossRef]
- Yong, Z.; Hao-Ru, T.; Ya, L. Variation in Antioxidant Enzyme Activities of Two Strawberry Cultivars with Short-Term Low Temperature Stress. 2008, 4, 458–462, http://www.idosi.org/wjas/wjas4(4)/8.pdf. [Google Scholar]
- Loh, K.P.; Qi, J.; Tan, B.K.H.; Liu, X.H.; Wei, B.G.; Zhu, Y.Z. Leonurine Protects Middle Cerebral Artery Occluded Rats Through Antioxidant Effect and Regulation of Mitochondrial Function. Stroke 2010, 41, 2661–2668. [Google Scholar] [CrossRef]
- Li, H.-X.; Xiao, Y.; Cao, L.-L.; Yan, X.; Li, C.; Shi, H.-Y.; Wang, J.-W.; Ye, Y.-H. Cerebroside C Increases Tolerance to Chilling Injury and Alters Lipid Composition in Wheat Roots. PLoS One 2013, 8, e73380. [Google Scholar] [CrossRef]
- Su, X.N.; Li, C.Y.; Xu, Y.J.; Huang, S.H.; Liu, W.L.; Liao, Z.X.; Zhang, Y.P. Feeding preference and adaptability of fall armyworm Spodoptera frugiperda on five species of host plants and six weeds. Journal of Environmental Entomology 2022, 44, 263–272. [Google Scholar] [CrossRef]
- CITES. Convention on International Trade in Endangered Species of Wild Fauna and Flora. Available online: https://www.cites.org/eng/disc/what.php.
- Song, J.; Zhang, L.N.; Zhang, Z.H.; Zhou, Z.Z.; Liang, J.F.; Lu, J.K. High-throughput Sequencing Analysis of Fungal Diversity in Agar-wood Wound Locations. Chinese Journal of Tropical Crops 2020, 46–47. [Google Scholar] [CrossRef]
- Zhou, Y.K.; Qiao, H.L.; Zhan, Q.Q.; Zhao, X.S.; Lu, L.L.; Chen, J. Occurrence and Control of the Disease and Pests Damage on Aquilaria Siensis in Hainan. Modern Chinese Medicine 2017, 19, 1102–1105. [Google Scholar] [CrossRef]
- Silva, D.M.D.; Bueno, A.D.F.; Andrade, K.; Stecca, C.D.S.; Neves, P.M.O.J.; Oliveira, M.C.N.D. Biology and Nutrition of Spodoptera Frugiperda (Lepidoptera: Noctuidae) Fed on Different Food Sources. Scientia Agricola 2017, 74, 18–31. [Google Scholar] [CrossRef]
- Carrasco, D.; Larsson, M.C.; Anderson, P. Insect Host Plant Selection in Complex Environments. Current Opinion in Insect Science 2015, 8, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Lu, W.; Hou, M.L.; Wen, J.H.; Li, J.W. Effects of plant volatiles on herbivorous insects. Plant Protection 2007, 7–11. [Google Scholar] [CrossRef]
- Moreau, J.; Benrey, B.; Thiéry, D. Grape Variety Affects Larval Performance and Also Female Reproductive Performance of the European Grapevine Moth Lobesia Botrana (Lepidoptera: Tortricidae). Bull Entomol Res 2006, 96, 205–212. [Google Scholar] [CrossRef]
- Chen, Y.; Gao, Y.; Zhu, H.; Romeis, J.; Li, Y.; Peng, Y.; Chen, X. Effects of Straw Leachates from Cry1C-Expressing Transgenic Rice on the Development and Reproduction of Daphnia Magna. Ecotoxicology and Environmental Safety 2018, 165, 630–636. [Google Scholar] [CrossRef]
- Kumar, A.; Vajpayee, P.; Ali, M.B.; Tripathi, R.D.; Singh, N.; Rai, U.N.; Singh, S.N. Biochemical Responses of Cassia Siamea Lamk. Grown on Coal Combustion Residue (Fly-Ash). Bulletin of Environmental Contamination and Toxicology 2002, 68, 675–683. [Google Scholar] [CrossRef]
- Marciniak, B.; Grabowicz, W.; Ferenc, T. Evaluation of Micronuclei in Mice Bone Marrow and Antioxidant Systems in Erythrocytes Exposed to A-Amanitin. 2013, 63, 147–153. [Google Scholar] [CrossRef]
- Vuleta, A. Adaptive Flexibility of Enzymatic Antioxidants SOD, APX and CAT to High Light Stress: The Clonal Perennial Monocot Iris Pumila as a Study Case. Plant Physiology and Biochemistry 2016, 100, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Ismaiel, M.M.S.; El-Ayouty, Y.M.; Piercey-Normore, M. Role of pH on Antioxidants Production by Spirulina (Arthrospira) Platensis. Brazilian Journal of Microbiology 2016, 47, 298–304. [Google Scholar] [CrossRef]
- Alyokhin, A.; Benkovskaya, G.; Udalov, M. Chapter 4 - Colorado Potato Beetle. In Insect Pests of Potato (Second Edition); Alyokhin, A., Rondon, S.I., Gao, Y., Eds.; Academic Press, 2022; pp. 29–43. ISBN 978-0-12-821237-0. [Google Scholar] [CrossRef]
- Jernelöv, A. The Colorado (Potato) Beetle. In The Long-Term Fate of Invasive Species: Aliens Forever or Integrated Immigrants with Time? Jernelöv, A., Ed.; Springer International Publishing: Cham, 2017; ISBN 978-3-319-55396-2. [Google Scholar] [CrossRef]
- Chen, Z.M. Chemical Ecology of Tea Tree Pests. Shanghai Scientific & Technical Publishers: Shanghai, 2013. [Google Scholar]
- Lu, Y.; Wu, K.; Jiang, Y.; Xia, B.; Li, P.; Feng, H.; Wyckhuys, K.A.G.; Guo, Y. Mirid Bug Outbreaks in Multiple Crops Correlated with Wide-Scale Adoption of Bt Cotton in China. Science 2010, 328, 1151–1154. [Google Scholar] [CrossRef] [PubMed]
- Rwomushana, I. Spodoptera Frugiperda (Fall Armyworm). CABI Compendium 2019, CABI Compendium, 29810. [Google Scholar] [CrossRef]
- Heppner, J.B. Lepidoptera of Florida, Part 1: Introduction and Catalog. Arthropods of Florida and Neighboring Land Areas 2007, 17, 1–670. [Google Scholar]
- Casmuz, A.; Juárez, M.L.; Socías, M.G.; Murúa, M.G.; Prieto, S.; Medina, S.; Willink, E.; Gastaminza, G. Revisión de Los Hospederos Del Gusano Cogollero Del Maíz, Spodoptera Frugiperda (Lepidoptera: Noctuidae) Review of the Host Plants of Fall Armyworm, Spodoptera Frugiperda (Lepidoptera: Noctuidae). Revista de la Sociedad Entomológica Argentina 2010, 69, 209–231. [Google Scholar]
- Silva, A.G.A.; Goncalves, C.R.; Galvão, D.M.; Gonçalves, A.J.L.; Gomes, J.; Silva, M.N.; Simoni, L.; Silva, R.; Galvao, D.; Gonçalves, M. Quarto Catálogo Dos Insetos Que Vivem Nas Plantas Do Brasil. Seus Parasitos e Predadores: Parte 2, Tomo 1o, Insetos, Hospedeiros e Inimigos Naturais – ScienceOpen. In; Ministério da Agricultura: Rio de Janeiro, Brasil, 1968. [Google Scholar]
- Wang, Z.; Cao, H. The Complete Mitochondrial Genome Sequence of Aquilaria Sinensis. Mitochondrial DNA Part B Resources 2021, 6. [Google Scholar] [CrossRef]
- Cui, C.; Yang, Y.; Zhao, T.; Zou, K.; Peng, C.; Cai, H.; Wan, X.; Hou, R. Insecticidal Activity and Insecticidal Mechanism of Total Saponins from Camellia Oleifera. Molecules 2019, 24, 4518. [Google Scholar] [CrossRef] [PubMed]
- Mohan, C.; Radhakrishnan Nair, C.P.; Nampoothiri, C.K.; Rajan, P. Leaf-Eating Caterpillar (Opisina Arenosella)-Induced Yield Loss in Coconut Palm. Int. J. Trop. Insect Sci. 2010, 30, 132–137. [Google Scholar] [CrossRef]
- Shameer, K.S.; Nasser, M.; Mohan, C.; Hardy, I.C.W. Direct and Indirect Influences of Intercrops on the Coconut Defoliator Opisina Arenosella. J Pest Sci 2018, 91, 259–275. [Google Scholar] [CrossRef]
- Wang, Y.C.; Qin, W.K. Main pests and diseases of coconut in Hainan. Chinese Journal of Tropical Agriculture 2000, 59–62. [Google Scholar] [CrossRef]
- Yan, W.; Lv, B.Q.; Li, H.; Li, C.X.; Liu, L.; Qin, W.Q.; Peng, Z.Q.; Luo, Y.Q. Risk analysis of the coconut blackheaded caterpillar, Opisina arenosella, in China and Hainan Island. Journal of Biosecurity 2013, 22, 163–168. [Google Scholar] [CrossRef]
- Zhang, Y. K.; Zhu, G. Y.; Wang, J. Q.; Wu, Z. H.; A H., C.; Duan, B. Studies on Biological Characteristics of Buzura suppressaria in Rubber Plantation. Tropical Agricultural Science and Technology 2019, 42, 6–9+12. [Google Scholar] [CrossRef]
- Zhu, G.Y.; Zhang, Y.K.; Zhang, Z.B.; Wang, J.Q.; Zhou, M.; Duan, B. Preliminary Study on a New Pest Occurred in Hevea brasiliensis in Xishuangbanna. Tropical Agricultural Science and Technology 2015, 38, 9–11. [Google Scholar] [CrossRef]
- Li, G.Y.; Wang, Q.B.; Li, Y.Y.; Zhou, S.X.; Yv, H.Y. A review of influencing factors on latex yield of Hevea brasiliensis. Chinese Journal of Ecology 2014, 33, 510–517. [Google Scholar] [CrossRef]
- Xiang, T.; Chui, L. X. A Survey of Major Pest Insects and Their Natural Enemies of Dalbergia odorifera Plantations in Hainan. Chinese Journal of Tropical Agriculture 2018, 38, 59–62. [Google Scholar] [CrossRef]
- Zhou, G. Y. Green prevention and control technology of major insect pests of Camellia oleifera (I). Forestry and Ecology 2023, 40–41. [Google Scholar] [CrossRef]
- Zhou, G. Y. Green prevention and control technology of major insect pests of Camellia oleifera (II). Forestry and Ecology 2023, 38–39. [Google Scholar] [CrossRef]
Figure 1.
The survival rate (%) of S. frugiperda when fed on leaves from seven plant species until eclosion (n = 30). The data were analyzed using the Kaplan-Meier procedure and log-rank test. Significant differences among subgroups are indicate.
Figure 1.
The survival rate (%) of S. frugiperda when fed on leaves from seven plant species until eclosion (n = 30). The data were analyzed using the Kaplan-Meier procedure and log-rank test. Significant differences among subgroups are indicate.
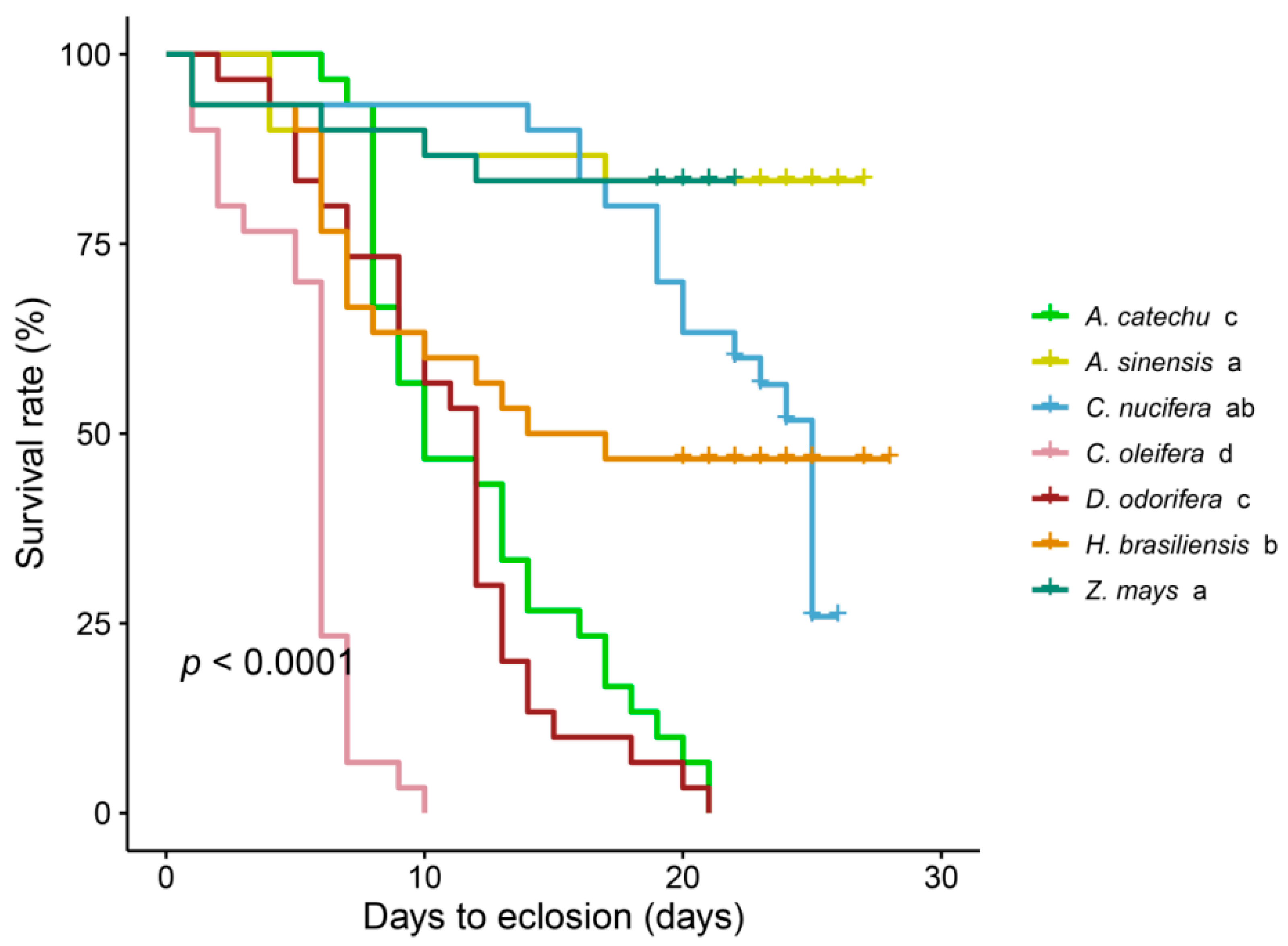
Figure 2.
The daily mean leaf area consumption (mm2) by S. frugiperda, feeding on leaves from different plants, was subjected to a comprehensive LMER analysis. Random factors were considered for distinct plant treatments, time (days), and replications. Treatment comparisons were assessed using the ANOVA function with type III sum of squares (car package). Significance levels (P < 0.05) are indicated by letters following subgroups. The gray areas in the figure delineate the upper and lower bounds of the 95% confidence intervals.
Figure 2.
The daily mean leaf area consumption (mm2) by S. frugiperda, feeding on leaves from different plants, was subjected to a comprehensive LMER analysis. Random factors were considered for distinct plant treatments, time (days), and replications. Treatment comparisons were assessed using the ANOVA function with type III sum of squares (car package). Significance levels (P < 0.05) are indicated by letters following subgroups. The gray areas in the figure delineate the upper and lower bounds of the 95% confidence intervals.
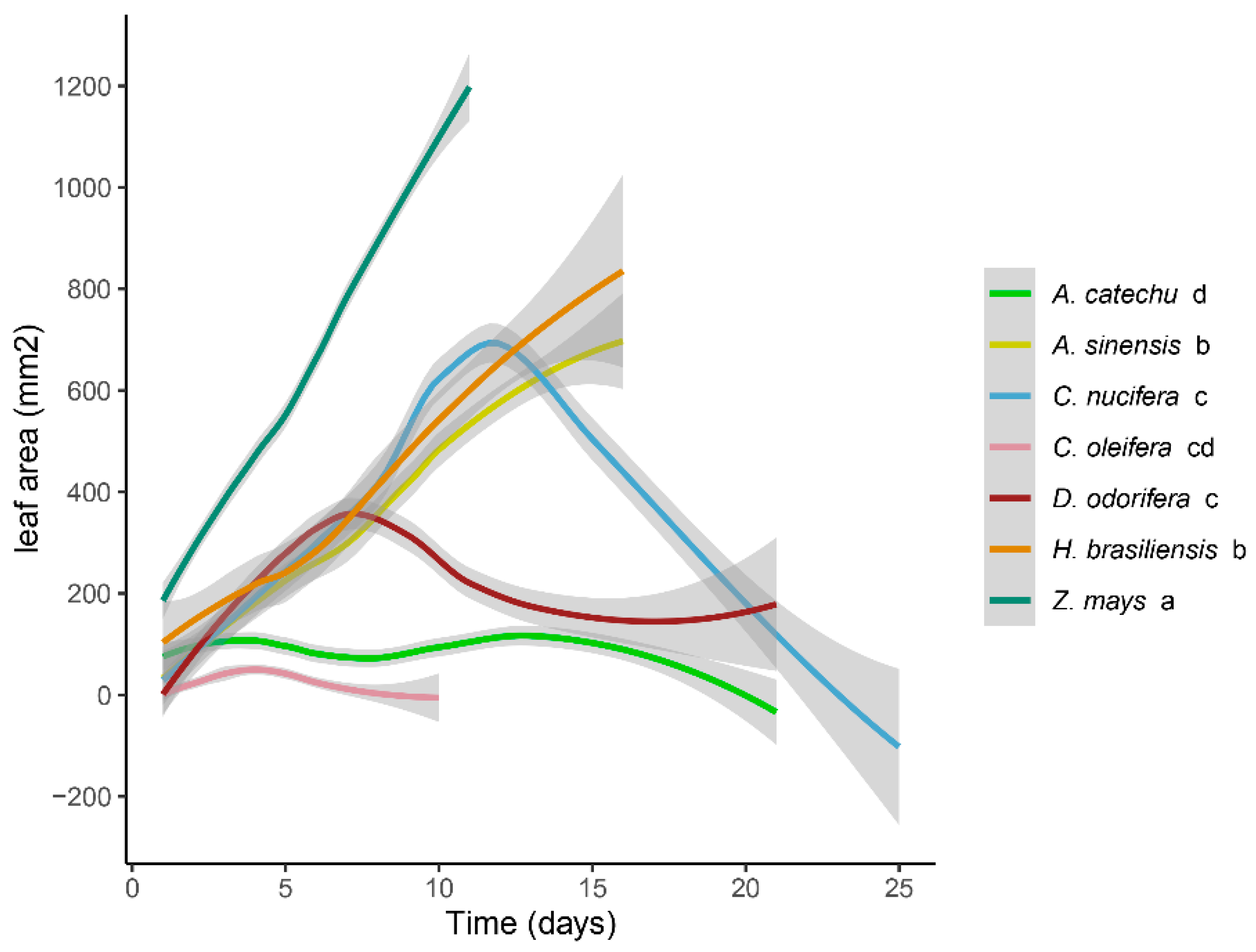
Figure 3.
(A) SOD activity; (B) POD activity. Tukey HSD test was used for analysis. Different letters indicate significant differences (P < 0.05).
Figure 3.
(A) SOD activity; (B) POD activity. Tukey HSD test was used for analysis. Different letters indicate significant differences (P < 0.05).
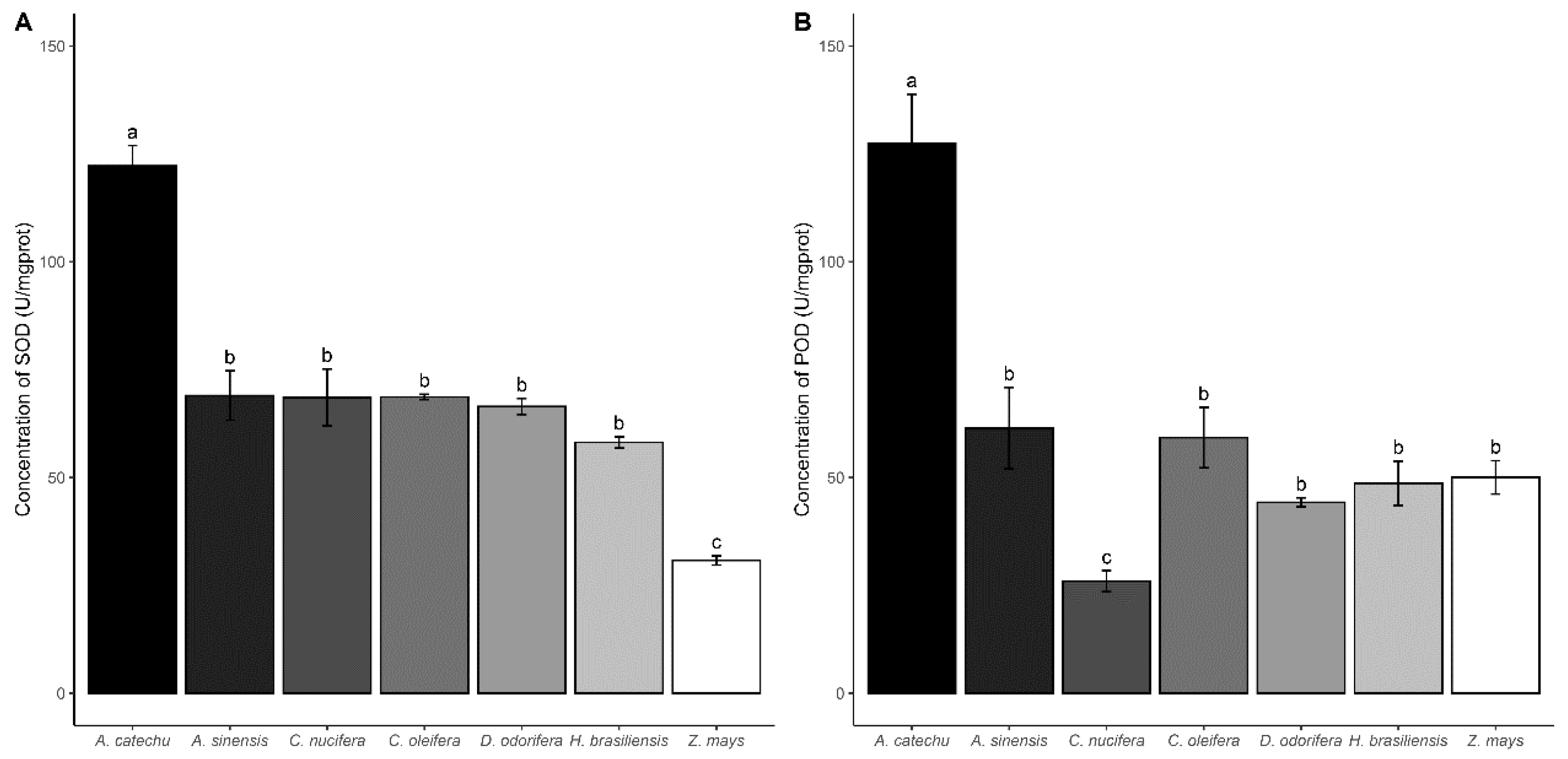
Figure 4.
Feeding preference of 3rd instar S. frugiperda on six plant species, the preference index is the percentage of the larval feeding area on the leaf discs from one plant to the sum of the feeding area from all leaf discs, and the index will range from 0 to 100%, with ≤1/6 indicating no preference for feeding, using Tukey HSD test. Different letters indicate significant differences (P < 0.05).
Figure 4.
Feeding preference of 3rd instar S. frugiperda on six plant species, the preference index is the percentage of the larval feeding area on the leaf discs from one plant to the sum of the feeding area from all leaf discs, and the index will range from 0 to 100%, with ≤1/6 indicating no preference for feeding, using Tukey HSD test. Different letters indicate significant differences (P < 0.05).
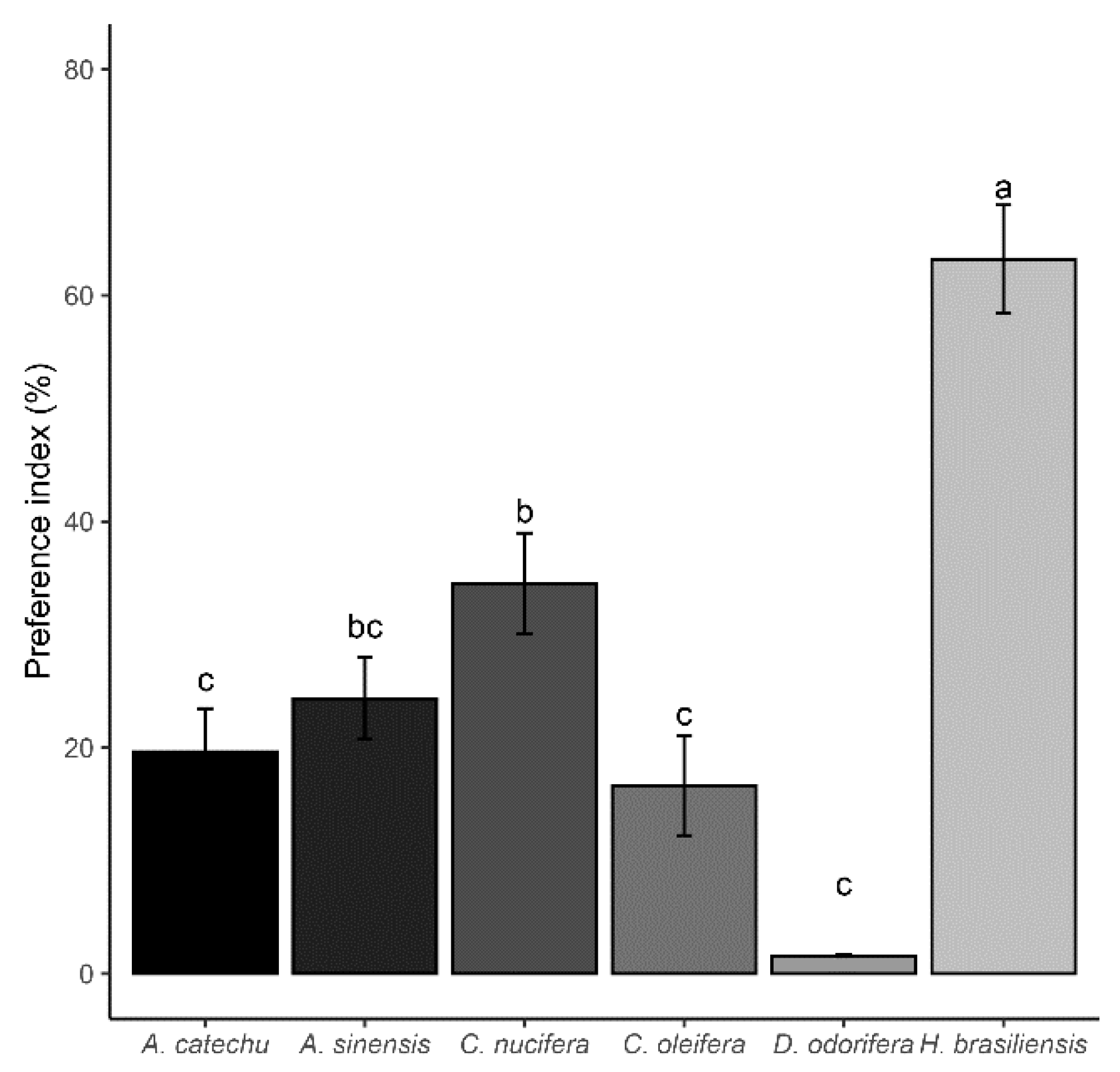

Table 1.
Performance of S. frugiperda on different diets.
| Parameter | Z. mays | H. brasiliensis | A. sinensis | C. nucifera | D.odorifera | A.catechu | C. oleifer |
|---|---|---|---|---|---|---|---|
| Pupation rate (%) a | 90.00 (30) a | 50.00 (30) b | 86.67 (30) a | 76.67 (30) ab | 0 (30) c | 3.33 (30) c | 0 (30) c |
| Eclosion rate (%) a | 83.33 (30) a | 46.67 (30) b | 83.33 (30) a | 46.67 (30) b | 0 (30) c | 0 (30) c | 0 (30) c |
| Larval development time (d) b | 10.18±0.21 (27) a | 13.27±0.57 (15) b | 14.36±0.32 (26) bc | 16.13±0.55 (23) c | - | 14 (1) | - |
| Pupal development time (d) b | 10.04±0.12 (25) a | 10.79±0.21 (14) b | 10.72±0.13 (25) b | 9.00±0.15 (14) c | - | - | - |
| Pupal length (mm) c | 15.34±0.20 (25) a | 16.11±0.38 (14) a | 15.81±0.23 (25) a | 15.38±0.27 (14) a | - | 12.00 (1) | - |
| Female pupal fresh weight (mg) c | 120.72±2.84 (11) a | 150.00±6.06 (6) b | 130.50±5.08 (6) ab | 111.67±4.00 (12) a | - | - | - |
| Male pupal fresh weight (mg) c | 131.00±3.18 (14) a | 133.00±7.94 (8) a | 143.47±3.00 (19) a | 127.00±3.00 (2) a | - | - | - |
| Pupal mean fresh weight (mg) | 126.48±3.17 (25) a | 140.29±5.56 (14) b | 140.36±3.70 (25) b | 113.86±0.3.73 (14) a | |||
| Females rate (%) a | 44.00 (25) a | 42.86 (14) a | 24.00 (25) a | 85.71 (14) a | - | - | - |
Life-table parameters (±SE) of S. frugiperda larvae when fed exclusively with different tree leaves. Number of replicates is given in parentheses. The means in the same rows followed by the same letters denoted no significant difference between the treatments. Due to limitations in the sample size, larval development time and pupal length for the A. catechu group were not compared with those of other groups. a χ2 test (Significance values have been adjusted by the Bonferroni correction for multiple tests). b Kruskal-Wallis Test with Mann-Whitney U-test (Significance values have been adjusted by the Bonferroni correction for multiple tests). c one-way ANOVA with Tukey's Honestly Significant Difference (HSD) test.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



