
Submitted:
03 March 2024
Posted:
05 March 2024
You are already at the latest version
Abstract
Antimicrobial Peptides (AMPs), as immune effectors synthesized by a variety of organisms, not only constitute a robust defense mechanism against a broad spectrum of pathogens in the host, but also show promising applications as effective antimicrobial agents. Notably, insects are significant reservoirs of natural AMPs. However, the complex array of variations in types, quantities, antimicrobial activities, production pathways of AMPs, and evolution of AMPs across insect species presents a significant challenge for immunity system understanding and AMP applications. This review covers insect AMP discoveries, classification, common properties, and mechanisms of action. Additionally, we summarize the types, quantities, and activities of im-mune-related AMPs in each model insect. We also conducted the first comprehensive investigation into the diversity, distribution, and evolution of 20 types of AMPs in model insects, employing phylogenetic analysis to describe their evolutionary relationships and shed light on conserved and distinctive AMP families. Furthermore, we summarize the regulatory pathways of AMP production through classical signaling pathways and additional pathways associated with Nitric Oxide, insulin-like signaling, and hormones. This review advances our understanding of AMP as guardians in insect immunity system and also unlocks a gateway to insect AMP resources, facilitating the use of AMPs to address food safety concerns.
Keywords:
Antimicrobial agents
; Antimicrobial Peptide evolution
; Antimicrobial Peptide regulation
; Insect
1. Introduction
Vertebrates are armed with both innate and adaptive immunity, but insects rely solely on innate immunity to navigate their complex environments with microbes, such as bacteria, fungi, and viruses [1,2]. Antimicrobial peptides (AMPs), also known as host defense peptides, are crucial and evolutionarily conserved components of the innate immune responses [3,4,5]. They are composed of a diverse group of naturally occurring molecules found in various organisms, including humans, animals, plants, insects, and microorganisms [6,7,8,9,10,11].
AMPs work by acting as the initial defense against a diverse array of pathogens such as bacteria, fungi, viruses, and certain parasites [12]. In comparison to antibiotics, AMPs exhibit a wide-ranging capacity to combat bacteria, fungi, viruses, and even cancer cells [13]. Notably, they possess the ability to kill antibiotic-resistant pathogens, making them a promising candidate for clinical applications [14]. The discovery of AMPs across diverse insect species has significantly advanced our understanding of their functions.
The history of AMPs can be traced back to the identification of gramicidins in 1939. Gramicidins belong to a class of naturally occurring peptide antibiotics produced by specific strains of soil bacteria, Bacillus brevis and Bacillus aneurinolyticus, known for their activity against Gram-positive bacteria [15,16]. Subsequently, the first plant AMP, known as purothionin, was isolated from wheat (Triticum aestivum) endosperm in 1942. As a member of the thionin family of AMPs, purothionin is a cationic peptide with antimicrobial properties, capable of targeting and eradicating various microorganisms, including bacteria and fungi [17]. In 1962, bombinin, one of the earliest animal AMPs, was discovered in the orange speckled frog [18]. Bombinin is a defense effector against pathogens, playing a crucial role in protecting frogs from bacterial, viral, and fungal infections. In 1980, the first insect AMP, cecropins, was identified in the pupae of Hyalophora cecropia (Insecta: Lepidoptera), representing a significant milestone as the first major group of α-helical AMPs [19]. Cecropins were notably found in the hemolymph and other immune tissues of insects [20]. The number of known AMPs now exceeds 3000 (detailed in the Antimicrobial Peptide Database, http://aps.unmc.edu/AP), suggesting their versatile functionalities beyond antimicrobial activity, encompassing roles in wound healing, inflammation modulation, and endotoxin neutralization. Overall, the study of AMPs has offered invaluable insights into the ancient origins of defense mechanisms and their evolution, inspiring potential applications as alternatives to conventional antibiotics and fueling biomedical research for novel therapeutic purposes.
Some reviews have summarized the AMPs from the model insect Drosophila (Insecta: Diptera), which mainly focused on the classification, activity and regulation of AMPs [21,22]. In this review, we summarize the current knowledge and recent advances on AMPs from various model insects, and highlight the regulatory pathways and evolution of insect AMPs, and present a prospective on the potential applications of insect AMPs.
2. Structures, Antimicrobial Activities, and Common Properties of Insect AMPs
2.1. Structures and Antimicrobial Activities of Insect AMPs
Different AMPs may exhibit diverse activities against bacteria, fungi, or viruses based on their structures. Understanding how AMPs function in defending against microbial pathogens relies on uncovering the relationship between their structure and activity. The quantities and types of AMPs in insects vary significantly between species (Table 1). Insect AMPs are typically categorized into four groups based on their secondary structures: α-helical AMPs, β-sheet cysteine-rich AMPs, proline-rich AMPs, and glycine-rich AMPs [23,24,25].
2.1.1. α-Helical Insect AMPs
Insect α-helical AMPs (e.g., cecropin and moricin) contain N-terminal amphiphilic α-helical and C-terminal hydrophobic α-helical [26,27]. They belong to secreted proteins, and mature active AMPs are produced following the removal of signal peptides [25]. The most abundant α-helical peptides are cecropin and cecropin-like peptides, including ceratotoxin, enbacin, hyphancin, sarcotoxins, spodopsin, and stomoxyn [28,29]. Cecropins have a broad range of activity to destroy Gram-positive, Gram-negative bacteria, and fungi [30,31,32]. Moricins are specifically found in lepidopteran insects and share similar secondary structure to cecropin, but cecropin has a hinge region to link the N and C-terminal α-helixes, which is absent in moricin [25]. Likewise, moricins can fight against Gram-positive/negative bacteria and fungi [31,32,33].
2.1.2. β-Sheet Cysteine-Rich Insect AMPs
In the β-sheet cysteine-rich subclass, AMPs (e.g., defensin and drosomycin) have conserved cysteine residues to form disulfide bonds. These AMPs are composed of α-helical and β-sheet which are stabilized by disulfide bridges [34]. Defensins are the most representative AMPs in this class. They are widely reported in insect orders, such as Coleoptera, Diptera, Hymenoptera, and Lepidoptera [35,36,37,38]. They have strong activity to kill Gram-positive and Gram-negative bacteria, and some of them such as Bombyx mori (Insecta: Lepidoptera) defensin, Galleria mellonella (Insecta: Lepidoptera) gallerimycin, and Apis mellifera (Insecta: Hymenoptera) royalisin, also defend against fungi [39,40,41,42,43]. Drosomycin and defensin have similar structures, whereas their sequences differ except for the presence of cysteine residues [34]. As known, drosomycins are antifungal peptides, they also function in killing Gram-negative bacteria [39,44,45].
2.1.3. Proline-Rich Insect AMPs
The proline-rich subclass AMPs are characterized by the presence of multiple proline residues. Some proline-rich peptides, such as lebocin, drosocin, and apidaecin, undergo O-glycosylation as a post-translational modification, which is essential for maximizing their activity [46,47]. However, proline-rich AMP abaecin from A. mellifera is not O-glycosylated [48]. Lebocins, sharing 41% sequence identity to A. mellifera abaecin, are proline-rich glycopeptides reported in lepidopteran insects. They are generated by proteolytic activation of their precursor proteins, and activated lebocins can broadly defend against Gram-positive/negative bacteria and fungi in lepidopteran insects [49,50,51]. While drosocin is a typical short-sized proline-rich glycopeptide found in D. melanogaster and its primary antimicrobial activity is targeted against Gram-negative bacteria [52,53]. Apidaecins are the major AMPs in A. mellifera humoral immunity. They share significant sequence homology with drosocin and are effective in defending against Gram-negative bacteria [54].
2.1.4. Glycine-Rich Insect AMPs
Glycine-rich AMPs have been identified in various insect orders, including Coleoptera, Diptera, Hemiptera, Hymenoptera, and Lepidoptera. The common AMPs in this subclass are attacin, coleoptericin, tenecin, diptericin, hemiptericin, hymenoptaecin, and gloverin [12,36,55,56,57]. Attacins are large glycine-rich peptides with a signal peptide, a pro-peptide region, an attactin domain and two glycine-rich domains [58,59]. Attacins have six isoforms which can be divided into two categories: basic attacins (A-D) and acidic attacins (E and F) [60]. In D. melanogaster, attacin A-D mainly kill Gram-negative bacteria. However, attacins also fight against Gram-positive bacteria and fungi in lepidopteran insects [61,62,63]. Gloverins are another group of large glycine-rich peptides identified exclusively in Lepidoptera. Although sharing a high sequence identity among lepidopteran species [64,65], gloverins show varying activities against microbes. Gloverins from B. mori mainly fight against Gram-negative bacteria like Escherichia coli [66,67]. Whereas Spodoptera exigua (Insecta: Lepidoptera) and M. sexta (Insecta: Lepidoptera) gloverins show activity against Gram-positive bacteria but no activity against E. coli [64,68]. Diptericins are glycine-rich AMPs that are mainly found in Dipteran, such as D. melanogaster, Sarcophaga peregrina (Insecta: Diptera), Mayetiola destructor (Insecta: Diptera) and Phormia terranovae (Insecta: Diptera), and have activities against Gram-negative bacteria [39,69,70,71,72].
2.2. Common Properties of Insect AMPs
Although AMPs exhibit diversity in both structure and antimicrobial activity, they share several properties, including heat stability, positive charge, hydrophobicity, and amphipathicity. Some of these properties are necessary for AMP activity and selectivity. Firstly, most of insect AMPs have a net positive charge. This cationic nature enables them to interact with negatively charged components of microbial membranes through electrostatic attraction, which is the basis for the sequent aggregation of AMPs on the microbial surface and reaching the concentration threshold for membrane rupture. Studies also show that appropriately increased positive charge enhances the antimicrobial activity of AMPs [73,74]. Secondly, hydrophobicity is a critical property that determines the extent of insertion of AMPs into microbial membrane phospholipid bilayer. AMPs exhibit hydrophobicity because they contain up to 50% hydrophobic residues, and optimal hydrophobicity is necessary for their antimicrobial activity, and higher or lower hydrophobicity results in AMP inactivation [75]. The last character is the amphiphilicity of AMPs. This refers to AMPs having a positively charged hydrophilic region that binds to negatively charged components and a hydrophobic region that binds to lipids. Amphiphilicity is the key feature for AMPs to disrupt the structure of microbial membranes [76]. These characteristics such as charge also affect the selectivity of AMPs, for example, cationic AMPs present stronger attraction to the negatively charged bacteria rather than “self”-cells, as the membranes of “self”-cells are composed of zwitterionic phospholipids [23]. It is important to note that these properties interdependently affect the activity and selectivity of AMPs, and changes in one parameter always lead to compensatory changes in others [77].
3. Evolution of Insect AMPs
To explore the evolutionary history of AMPs across model insects, as well as their diversity and function, we performed the phylogenetic relationships analysis of 20 kinds of AMPs and lysozymes across model insects. Based on our phylogenetic analysis, four major groups of AMPs are distinguished (Figure 1). Group I contains eight kinds of AMPs: moricin, cecropin, gambicin, diapausin, drosomycin, metchnikowin, cobatoxin, and bomanin; group II is clustered by lysozymes and four kinds of AMPs: defensin, gallerimycin, gloverin, and apismin; the group III is composed of only two kinds of AMPs: attacin and diptericin; the group IV includes six kinds of AMPs: lebocin, drosocin, coleoptericin, hymenoptaecin, apidaecin, and abaecin (Figure 1). We found these AMPs are broadly clustered according to their structure and activity. For example, cecropin and moricin (α-helical AMPs), attacin and diptericin (glycine-rich AMPs), drosocin and lebocin (proline-rich AMPs) cluster together based on their respective structures. Although the secondary structures are not identical, consistent antimicrobial activity (antifungal) may account for the clustering of diapausin, metchnikowin, drosomycin, and bomanin (Figure 1). This indicates that the AMPs within the same major group show close phylogenetic relationships, particularly these AMPs clustered together within smaller clusters.
The phylogenetic relationships of insect orders are well-documented [78] (Figure 2a), while our understanding of the evolutionary patterns of AMPs and lysozymes in insects remains limited. To address this knowledge gap, the distribution patterns of genes responsible for encoding AMPs and lysozymes were explored within the context of Hexapoda evolution. The homologous genes of lysozymes and the 20 kinds of AMPs were identified based on the genomic and transcriptome sequence data. The detailed methods are described in Supplementary file 1.
Lysozymes can be encoded in all insects, with the exception of Mengenilla moldrzyki (Insecta: Strepsiptera), whereas not all the kinds of AMPs can be encoded in insects (Figure 2b). Defensins are found in most insects, with over half of insect species having the ability to encode attacins. The wide distribution may be due to their broad-spectrum microbe-killing activities which fulfill the essential need of insects to destroy pathogens, allowing defensin and attacin to be largely retained over the course of evolution. In contrast, apidaecin and apisimin are uniquely present in specific bee species (Figure 2b). Bomainins can only been found in D. melanogaster, and moricins are presented in some lepidopteran insects (Figure 2b). Taken together, some AMPs are highly conserved and widely distributed across different species, while some AMPs are unique to specific species.
In the order of Lepidoptera, insects exhibit the most diverse AMP types (Figure 2), most insects produce greater number of AMPs than any other taxonomic group [24]. While some insect species have no AMP production in response to immune challenge, exemplified by some Hemiptera insects. Hemiptera are distinctly divided into two groups (Figure 2a). The first group includes Trialeurodes vaporariorum, Bemisia tabaci, Acanthocasuarina muellerianae, Planococcus citri, Essigella californica, Acyrtosiphon pisum, and Aphis gossypii (Figure 2a), where most species lack all AMP genes, whereas only one kind of AMP gene (abaecin) is found in P. citri (Figure 2b). Acanthosoma haemorrhoidale, Notostira elongate, Ranatra linearis, Velia caprai, Xenophysella greensladeae, Nilaparvata lugens, Cercopis vulnerate, and Okanagana villosa formed the second group (Figure 2a). Interestingly, most species in this group can encode one or two kinds of AMPs, at least including defensin, while only N. lugens lacks all AMP genes (Figure 2b), which is consistent with our previous study [79]. The absence of all AMPs is also observed in Protura, Ephemeroptera (such as Baetis (Insecta: Ephemeroptera), Isonychia bicolor (Insecta: Ephemeroptera), and Eurylophella (Insecta: Ephemeroptera), and Strepsiptera (like Stylops melittae) (Figure 2b). Surprisingly, Philopotamus ludificatus (Insecta: Trichoptera) lacks all the AMPs genes, while the other insects in Trichoptera show a wide variety of AMP types (Figure 2b). The shared occurrence of the absence of all AMPs across different insect orders suggests a possible result of parallel evolution in these insects. However, this could also result from the potential incompleteness of transcriptome and genome dataset, or failure in the detection method.
4. Action Mechanisms of AMPs
Bacteria can develop resistance to antibiotics in response to unreasonable antibiotic treatment [80], while AMPs have shown their attractiveness as potential antimicrobial agents [81]. AMPs play a crucial role in the insect innate immune system’s defense against pathogens such as bacteria, viruses, fungi, and even some parasites. Here are some mechanisms of how AMPs kill pathogens:
One of the most common mechanisms of action for AMPs is the disruption of the target pathogen’s cell membrane (Figure 3a). Most AMPs have both hydrophobic and hydrophilic regions. They have the capability to insert themselves into the lipid bilayer of pathogens, forming pores or disrupting the membrane’s integrity [82,83,84]. AMP-17, a novel type of AMP from Musca domestica (Insecta: Diptera), destroys 21.7% the Candida albicans (Fungi: Ascomycota) cell wall [85]. This disrupts the pathogenic ability to maintain osmotic balance, leading to cell lysis [86,87]. There are several proposed AMP-mediated disruption models such as toroidal-pore, barrel-stave, aggregated, and carpet models [88,89,90,91]. Unlike the large damage of pathogen’s cell membrane integrity, the other important AMP action is the alteration of the permeability of pathogen cell membranes through pore-forming transmembrane channel [91,92,93], allowing ions and other essential molecules to leak out and toxic molecules to enter, which in turn the disruption in ion balance can lead to cell death [94,95].
There are some other mechanisms of how AMPs act on pathogens. Some AMPs can penetrate the pathogen’s cell membrane and interfere with essential cellular processes (Figure 3b). These AMPs can interact with pathogen macromolecules such as key enzymes related to DNA/RNA, protein, and cell wall synthesis, which in turn inhibit the growth of pathogens [96]. They may bind ribosomes, DNA, or other vital cellular components, disrupting protein synthesis, DNA replication, and other metabolic processes [97,98,99]. Biofilms are protective structures formed by some bacteria and fungi, which can make them resistant to antibiotics [100]. AMPs can disrupt biofilms by penetrating the extracellular matrix and killing the embedded pathogens [101,102,103,104] (Figure 3c).
5. Transcriptional Regulation of AMPs
As antimicrobial effectors, insect AMPs are produced in hemocytes, fat body, and epithelial cells via two major nuclear factor-κB (NF-κB) pathways during infection: the Toll and the IMD (immune deficiency) pathways [21,105,106,107]. Some AMPs are produced only upon immune stimulation, for example cecropins. While lysozyme is present at low constitutive levels and significantly enhanced transcription in response to immune challenge [108]. Although it has been well described in the model insect D. melanogaster that AMPs are transcriptional induced via Toll, IMD, and JAK-STAT pathways [107,109], the production and regulation of AMPs in other species remain largely unknown.
5.1. Insect AMPs Regulated by Toll Pathway
Toll pathway is an evolutionarily conserved signaling cascade, and is activated after detection of foreign microbial cell wall components by peptidoglycan recognition proteins (PGRPs). In D. melanogaster, PGRP-SA, PGRP-SD, and Gram-negative bacteria-binding protein 1 (GNBP1) recognize the lysine-type peptidoglycan (Lys-type PGN) of Gram-positive bacteria, and GNBP3 involves the sensing of β-glucans of fungi to activate Toll pathway via serine protease cascade [110,111]. Previous studies have widely characterized that Toll pathway responses to Gram-positive bacteria and fungi invasion. However, a recent finding in M. sexta revealed that PGRPs bound to the meso-diaminopimelic acid-peptidoglycans (DAP-type PGN) of Gram-negative bacteria also can activate Toll signaling [112]. Toll signaling is triggered when extracellular mature cytokine Spätzle binds to the transmembrane Toll receptor. However, Toll9 from B. mori can directly bind Gram-negative bacteria-associated ligand lipopolysaccharide (LPS) to initiate Toll pathway without Spätzle binding [4]. Then three intracellular proteins, MyD88, Tube adaptor, and Pelle are recruited to form a heterotrimeric complex which can phosphorylate and degrade Cactus. Activated Dorsal and/or Dif translocate into the nucleus to initiate transcription of AMP genes, such as bomanin, drosomycin, and metchnikowin (Figure 4a).
5.2. Insect AMPs Regulated by IMD Pathway
Antimicrobial peptides are also induced from another NF-κB pathway IMD pathway. The signaling is activated in the fat body together with Toll pathway during systemic infection. IMD pathway also produces AMPs in insect gut to eliminate ingested pathogens, mounting a localized immune response [113,114,115]. Studies in Drosophila show that IMD pathway is triggered by transmembrane PGRP-LC after binding with DAP-type PGN from nearly all Gram-negative bacteria and certain Gram-positive bacteria in systemic immune responses [116,117,118]. Upon binding, PGRP-LC recruits intracellular adaptor protein IMD. The downstream proteins of IMD are: Drosophila Fas-associated death domain (dFADD) adaptor, Death related ced-3/Nedd2-like caspase (Dredd), TGF-beta activated kinase 1 (TAK1), TAK1-associated binding protein 2 (TAB2), Death-associated inhibitor of apoptosis 2 (DIAP2), IκB kinase (IKK) complex (containing IRD5 and Kenny) and transcription factor Relish. Relish is cleaved into Rel domain and ankyrin-repeat domain (ANK) by Dredd once Relish is phosphorylated by IKK complex. Further, actived Rel domain translocated into the nucleus to initiate AMPs expression, such as attacin, cecropin, defensin, diptericin, drosocin and drosomycin, to combat pathogens [22,117,119] (Figure 4b).
5.3. Insect AMPs Regulated by Intestinal IMD-NF-κB Pathway
AMPs generated from intestinal IMD-NF-κB pathway has been extensively reported in insects [120,121,122,123]. In Drosophila, this local immune response is triggered by cytosolic PGRP-LE after recognition of pathogenic bacteria elicitors [116,118]. AMPs (e.g., diptericin) produced locally in the gut play critical role in defensing against foodborne pathogens [114]. Whether these AMPs also kill commensal bacteria along the gut? Study has shown that negative regulator (e.g., PGRP-SC2) of IMD pathway dampens AMPs production and maintains commensal bacteria colonies homeostasis in gut [124]. However, the molecular mechanisms that AMPs target only pathogenic bacteria while maintaining symbiotic colony homeostasis is not fully established in D. melanogaster. Recently, one study in oriental fruit fly Bactrocera dorsalis (Insecta: Diptera) has revealed this mechanism. In brief, PGRP-LC of B. dorsalis positively activates IMD pathway to generate AMPs in foregut, filtering pathogenic bacteria from entering midgut to protect symbiotic bacteria homeostasis. Furthermore, symbiotic bacteria enhance the expression of PGRP-LB and PGRP-SB which are negative regulators of IMD pathway avoiding AMPs threaten in B. dorsalis midgut. Regional production of AMPs establishes a protective region for symbiotic bacteria [125]. However, some hemipteran insects are deficient in AMPs produced by IMD pathway due to their lack of crucial compositions such as PGRPs, IMD, dFADD, Dredd, IKK, and Relish [79,126,127]. What is noteworthy is that these insects can survive normally and even exist ubiquitously in the absence of the vital IMD pathway. For example, aphid which is a hemipteran insect with IMD pathway deficiency can still ravage in nature. The reasons may be as follows: firstly, aphids are insects that adopt the R-strategy to fit environment, so they invest more energy into reproduction rather than shaping immune resistance under limit resources [128]. Secondly, the phloem sap that aphid prick-sucking is normally sterile, and it largely reduces the risk of oral infection to aphid. Moreover, endosymbiotic bacteria protect aphids against pathogens and parositoids [79,129,130,131].
5.4. Insect AMPs Regulated by JAK-STAT Pathway
The Janus kinase/signal transducers and activators of transcription (JAK-STAT) pathway is also a conserved intracellular cascade that modulates the production of AMPs [113,132,133]. This pathway also takes place in the fat body and gut, and is elicited upon damage signals, septic injury not just pathogens intrusion [39,134,135,136]. JAK-STAT pathway is initiated through binding with either of cytokines Unpaired family (Upd1, 2 and 3) to the receptor Domeless (Dome). This signal recruits the JAK kinase Hopscotch (Hop) to phosphorylate Dome and Stat92E (STAT). Then dimerized STAT translocates into the nucleus to activate transcription of target genes such as AMP gene drosomycin-like peptide [39,132,133,137] (Figure 4c).
We summarize the types and activities of AMPs which are induced by the three conserved pathways mentioned above. The data are collected from some model insects, such as D. melanogaster, M. sexta, B. mori, Helicoverpa armigera (Insecta: Lepidoptera), Tribolium castaneum (Insecta: Coleoptera), Anopheles gambiae (Insecta: Diptera), A. mellifera, basing on the available studies (Table 2). Among which the pathways of AMPs production in Drosophila are well defined, and most AMPs are induced via the IMD pathway. As the IMD pathway tightly controls immune responses in fat body, hemocytes, and gut, playing a critical and ancestral role in defending against pathogens [115]. Whereas Toll and JAK-STAT pathways have multifunctional roles and active in developmental processes. Therefore, the sole role of IMD signaling in immune system allows it to respond more rapidly and readily to invading bacteria [39].
5.5. Insect AMPs Regulated by Other Signaling Pathways
Other signaling pathways can also regulate the production of AMPs (shown in Figure 4d,e). Studies show that the Nitric Oxide (NO) signaling can trigger AMPs expression. However, the mechanisms of how NO regulate AMP production vary across different insects. In Spodoptera exigua (Insecta: Lepidoptera), NO signaling regulated AMPs production by Toll and IMD pathways together [138]. While in Drosophila, NO-induced AMPs production is dependent only on IMD pathway [139]. Similarly, a recent study showed that NO also induced AMPs expression by IMD pathway in Ostrinia furnacalis (Insecta: Lepidoptera) [140]. Apart from this, appropriate induction of c-Jun N-terminal kinase (JNK) signaling which branches out from IMD pathway at dTAK1, is required for IMD pathway to produce AMPs [141,142].
It is well established that AMPs generation is affected by insect endogenous hormones, steroid 20-hydroxyecdysone (20E) and sesquiterpenoid Juvenile hormone (JH) [143,144]. Ecdysone signaling regulates AMP production during insect metamorphosis [145,146,147]. In Drosophila, prothoracicotropic hormone (PTTH) regulates the synthesis of 20E precursor ecdysone (Figure 4e). The precursor is then released into hemolymph and converted to activate form 20E. 20E binds to the membrane receptor DoEcR and enters the cell. Subsequently 20E binds to nuclear receptor EcR/USP, initiating the expression of a series of transcription factors. Transcription factors can directly initiate AMPs expression or activate IMD pathway by up-regulating PGRP-LC expression [146]. While the JH suppressed AMP gene expression (e.g., diptericin) in Aedes aegypti (Insecta: Diptera) [148].
Under starvation stress, AMPs can be induced by insulin-like signaling (IIS). The signaling is initiated upon binding insulin-like peptide (ILP) to insulin-like receptor (InR). Energy shortage status inhibitsIIS signaling and reduces AKT kinase expression. Activated FOXO translocates into nucleus inducing the expression of AMPs and anabolic genes [149,150,151]. In addition, AMPs also can be triggered under temperature stress [152]. These pathways differ from the classical AMP signaling pathways, as they can be activated the expression of AMPs in the absence of pathogen stimulation.
6. Potential Applications of Insect AMPs
To data, AMPs have been found in almost all invertebrates examined, mainly including insects and marine invertebrates. Although some marine invertebrates, such as horseshoe carb, can produce antibacterial and antifungal peptides tachyplesin and polyphemusin [76], insects possess distinct advantages as substantial and renewable reservoirs of AMPs. Their short lifecycles, compact size, and ease of cultivation make them ideal candidates for AMP applications. Insect AMPs, with their diverse range and promising application potential, stand as potent microbial agents within the innate immune system. As a result, they have garnered substantial interest throughout the food, agriculture, and pharmaceutical industries.
With growing concerns about food safety, people prefer foods with fewer chemical preservatives and fewer processing procedures. There is a growing advocacy for the use of natural preservatives in food industry. AMPs emerge as alternatives to traditional preservatives, as they exhibit some advantages, such as broad-spectrum bactericidal ability, thermally stable, acid and alkali resistance, and can be easily degraded by human proteases [153]. AMPs have been used as bio-bacteriostatic agents to preserve meat, fruit, juice, soy milk etc., and some of these peptides have been permitted by US Food and Drug Administration as food additives [154]. AMPs also can reduce lipid oxidation which leads to the generation of harmful compounds during meat preservation [155]. Active packaging is a promising technique to ensure the quality of food products. Moreover, active packaging is used to carry AMPs in microcapsules and nano-capsules, controlling release of AMPs to combat microbes during food preservation. However, not all types of AMPs can act as potential ingredients in packaging, it depends on their effectiveness [155,156]. There is a growing demand for food supply as an increasing population. AMPs can provide solutions by reducing food waste caused by food spoilage.
Insect AMPs are also applied in agriculture as antibiotic alternatives, avoiding the development of multidrug resistance among microbes [157,158]. For example, insect AMP cecropin AD was supplemented in diets instead of antibiotics, reducing incidence of diarrhea in piglets to bacterial infection [159]. Transgenic expression of insect AMPs in plants confer resistance to pathogenic bacteria and fungi [160,161]. In the pharmaceutical industry, AMPs exhibit potential in disease treatment, owing to their ability to selectively target cancer cells and promote cell apoptosis [162,163]. For example, defensins can cooperate with Drosophila TNF-like molecule Eiger to drive tumor cell death [164]. Studies show that insect AMPs can inhibit human pathogenic bacteria, making them to be drug candidates [24]. Nano-delivery system is an effective technology to deliver AMP drugs, but such system is still at early development stage in delivering AMPs, and it will be a new research hotspot in the future [165].
7. Future Perspective
Insect AMPs are peptides with heat stability and broad-spectrum bactericidal effect which make them to be the hotspots for developing insect resources. We summarize current knowledge of insect AMPs including classification, distribution pattern in the Hexapoda evolution, mechanism of action, and regulation in this review. However, there exits some underexplored aspects of insect AMPs, and we discuss these issues, future perspectives, and challenges for AMP applications in this section.
We summarized the pathways which regulate insect AMPs expression, including Toll, IMD, and JAK-STAT pathways as well as additional NO, ecdysone, IIS, and JNK signalings. Toll and IMD pathways are two sole intracellular cascades in Drosophila and currently no evidence shows there exits cross-talk between them. Some AMPs are produced only upon one signal pathway (e.g., diptericin only for IMD), whereas some are induced from both of the two pathways, such as drosomycin, its systemic expression is induced by Toll pathway and local expression is regulated by IMD pathway in Drosophila [107]. Why the certain AMP can be induced by two independent pathways remains to be further explored. Recently, some studies have shown that AMPs can also be induced by nonconventional activation of Toll pathway in M. sexta [4,112]. This reminds us that there may exit more possibilities for activating these pathways to produce AMPs among insects. In addition to conserved NF-κB pathways, NO, and ecdysone signaling are primarily associated with the IMD pathway to induce the production of AMPs, but how these pathways activate the IMD pathway is currently unknown. So far, studies of the pathways regulating AMP production are primary focused on the insects of Diptera and Lepidoptera, which are holometabolous insect orders. While how most hemimorphic insects (e.g., crickets and locusts) regulate AMP production has been neglected. Similarly, coleopteran insects, which make up more than one-third of insects, also need more attention on their AMP production. This is conducive to the extensive development and utilization of insect AMP resources.
Although certain AMP exhibits high specificity for a particular pathogen, such as diptericin act specifically on Providencia rettgeri, drosocin defenses against Enterobacter cloacae [166,167], we found no single AMP can singularly effective against all pathogens (Table 2). This may be the reason why innate system sustains multiple AMP species. However, several AMPs show similar antimicrobial activity, for instance, attacin, cecropin, defensin, diptericin, and drosocin defense against Gram-negative bacteria in Drosophila [109]. It is not an economical strategy for the innate system to maintain the production of functionally overlapping AMPs. Understandably, this strategy prevents pathogens from developing resistant to specific AMP. This strategy also implies that there may exit synergistic effect between AMPs. While current studies mainly focus on individual AMP with its production and activity in vitro, more studies need to focus on synergistic effect among AMPs to truly reflect how AMPs work in vivo.
Current studies of evolution of AMPs mainly focused on a certain insect order or specific type of AMP [58,67]. We provide a more comprehensive study of evolutionary relationships of AMPs in this review. Understandably, these AMPs with the same structure or activity primarily cluster into one group, but that’s not the case with some of these (e.g., attacin and gloverin in Figure 1). It requires further specific sequence and structural analyses for explanation. The discrepancy may be caused by gene duplication, horizontal gene transfer, and subsequent diversification among these AMPs during the course of evolution. However, we found some insects such as Protura, Ephemeroptera, Strepasiptera, and some Hemiptera have no AMPs production (Figure 2b). We analyzed and discussed the reasons for the absence of IMD pathway and AMPs in hemipteran insects based on aphids, in terms of fitness costs [79]. Insects may redirect resources allocated to immunity towards various physiological processes such as nutrition, reproduction, and foraging due to costly immune responses [168,169]. For instance, adult honeybees prioritize phenoloxidase-based immunity over the energetically expensive cellular immunity upon entering the foraging stage [169]. We infer that the lack of AMPs in some species is due to the results of parallel evolution between these insects and adaptation to the unique developing conditions. Thus, more studies are needed to explore why the insects in Protura, Ephemeroptera, and Strepasiptera do not rely on AMPs to defend against infections.
Besides the applications mentioned above, some insect AMPs exhibit resistance to parasites, and transgenic mosquitoes expressing AMPs have been generated to impede Plasmodium and nematode transmission [170,171]. Furthermore, antiviral AMPs are becoming hot research objects. However, the research on antiviral AMPs is still not in-depth. The following key issues remain further study: identification, recognition, regulation, and mechanism of action of antiviral AMPs. This research will facilitate the development of antiviral AMPs as antiviral drugs [172]. Although AMPs have been gained great attention in industrial applications, some shortcomings limit the wide application of natural AMPs, e.g., poor proteolytic stability, potential hemolysis, high production cost, low bioavailability, and unknown toxicity [76]. Many strategies have been conducted to circumvent these shortcomings by encapsulating and structurally modifying of AMPs [76,173]. Although insect cell lines have been proved to be promising systems to produce insect-derived recombinant peptides, the cost of production is still higher than conventional drugs [24]. More approaches are needed in the future to improve biological properties that make insect AMPs favorable in several industries. Facing with numerous insect AMP resources, research in Black Soldier Fly Hermetia illucens (Insecta: Diptera) provides us new techniques to predict AMP activity online, which facilitates the screening of promising AMP for further research in vitro [174]. Due to evolutionary conservatism, insect immune system has certain similarities with mammals in molecular components and signaling pathways. Insects, especially Drosophila can be used a powerful genetic tool to explore the diversity functions of AMPs, and it will provide new perspectives for determining the roles of AMPs in complex mammalian system. In addition to what we have reviewed in this paper, the design, extraction, and production processes of AMPs are also worthy of attention, which is conducive to the further application of insect AMPs resources in various fields.
8. Conclusion
AMPs are not only the effectors to eliminate the invading pathogens in insects, but also show promising application in clinic, agriculture, and food industries. In this review, the AMP discoveries, structures, mechanisms of action, antimicrobial activities, and shared characteristics in insects, and the types and quantities and activities AMPs in each model insect are summarized, this information provides references for further AMP investigations and applications. Additionally, we conducted the first comprehensive evolutionary analysis of model insect AMPs through phylogenetic analysis, unveiling four distinct groups based on structural and functional similarities. The analysis of AMP gene distribution responsible for encoding these antimicrobial effectors across diverse insect species sheds light on their conservation and uniqueness, which offers a guide on selecting the appropriate types of insects for specific applications. Moreover, the intricate network of classical signaling pathways regulating AMPs, including the classical Toll, IMD, and JAK-STAT pathways, along with additional pathways linked to Nitric Oxide, insulin-like signaling, and insect hormones such as 20-hydroxyecdysone and juvenile hormone are reviewed, suggesting the adaptability of insects in responding to diverse environmental challenges. This review not only enhances the comprehension of how AMPs serve as immunity guardians across insect species but also offers insights into using these resources in the food industry.
Supplementary Materials
The following supporting information can be downloaded at: Preprints.org, Supplementary file 1: Molecular methods; Table S1: The detailed information for the selected sequence of AMPs; Table S2: Public dataset used for AMP detection.
Author Contributions
Conceptualization, L.Z.Z., L.M. and K.K.C.; methodology, G.L.M.; validation, L.M. and K.K.C.; formal analysis, L.Z.Z., G.L.M., L.M. and K.K.C.; resources, L.M. and K.K.C.; data curation, L.Z.Z., G.L.M. and L.Z.; writing—original draft preparation, L.Z.Z. and G.L.M.; writing—review and editing, L.Z., L.M. and K.K.C.; visualization, L.Z.Z., G.L.M. and L.Z.; supervision, L.M. and K.K.C.; funding acquisition, G.L.M, L.M., and K.K.C.; All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China, grant number 31901876 and 32200391; the Natural Science Foundation of Jiangsu Province, grant number BK20190900; the Natural Science Foundation of Shanxi Province, grant number 202103021223125; the Distinguished and Excellent Young Scholar Cultivation Project of Shanxi Agricultural University, grant number 2023YQPYGC05; and the German Research Foundation, grant number DFG; project HU980/12-1.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data described in this manuscript is entirely available in this article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hoffmann, J.A. Innate immunity of insects. Curr. Opin. Immunol. 1995, 7, 4–10. [Google Scholar] [CrossRef] [PubMed]
- Hultmark, D. Immune reactions in Drosophila and other insects: a model for innate immunity. Trends Genet. TIG 1993, 9, 178–183. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Lu, Z. Immune responses to bacterial and fungal infections in the silkworm, Bombyx mori. Dev. Comp. Immunol. 2018, 83, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Tettamanti, G.; Bassal, T.; Heryanto, C.; Eleftherianos, I.; Mohamed, A. Regulators and signalling in insect antimicrobial innate immunity: Functional molecules and cellular pathways. Cellular Signal. 2021, 83, 110003. [Google Scholar] [CrossRef]
- Müller, U.; Vogel, P.; Alber, G.; Schaub, G.A. The innate immune system of mammals and insects. Contrib. Microbiol. 2008, 15, 21–44. [Google Scholar] [CrossRef]
- Zhang, L.-j.; Gallo, R.L. Antimicrobial peptides. Curr. Biol. 2016, 26, R14–R19. [Google Scholar] [CrossRef]
- Diamond, G.; Beckloff, N.; Weinberg, A.; Kisich, K.O. The roles of antimicrobial peptides in innate host defense. Curr. Pharm. Des. 2009, 15, 2377–2392. [Google Scholar] [CrossRef]
- Zasloff, M. Antimicrobial peptides of multicellular organisms. Nature 2002, 415, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Stączek, S.; Cytryńska, M.; Zdybicka-Barabas, A. Unraveling the role of antimicrobial peptides in insects. Int. J. Mol. Sci. 2023, 24, 5753. [Google Scholar] [CrossRef] [PubMed]
- Szczepanik, K.; Świątkiewicz, M. Hermetia illucens as a source of antimicrobial peptides-a review of in vitro and in vivo studies. Ann. Anim. Sci. 2024, 24, 77–88. [Google Scholar] [CrossRef]
- Dhanjal, D.S., Chopra, C., Bhardwaj, S., Sharma, P., Nepovimova, E., Singh, R., Kuca, K. Insect peptides with antimicrobial effects. In Antimicrobial Peptides. 2023.pp. 117-138. Academic Press.
- Bulet, P.; Stöcklin, R.; Menin, L. Anti-microbial peptides: from invertebrates to vertebrates. Immunol. Rev. 2004, 198, 169–184. [Google Scholar] [CrossRef] [PubMed]
- Hancock, R.E.; Diamond, G. The role of cationic antimicrobial peptides in innate host defences. Trends Microbiol. 2000, 8, 402–410. [Google Scholar] [CrossRef]
- Ageitos, J.M.; Sánchez-Pérez, A.; Calo-Mata, P.; Villa, T.G. Antimicrobial peptides (AMPs): Ancient compounds that represent novel weapons in the fight against bacteria. Biochem. Pharmacol. 2017, 133, 117–138. [Google Scholar] [CrossRef]
- Dubos, R.J. Studies on a bactericidal agent extracted from a soil bacillus: II. Protective effect of the bactericidalagent against experimental pneumococcus infections in mice. J. Exp. Med. 1939, 70, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Dubos, R.J. Studies on a bactericidal agent extracted from a soil bacillus: I. Preparation of the agent. Its activity in virto. J. Exp. Med. 1939, 70, 1–10. [Google Scholar] [CrossRef]
- Balls, A.K.; Hale, W.S.; Harris, T.H. A crystalline protein obtained from a lipoprotein of wheat flour. Cereal Chem. 1942, 19, 279–288. [Google Scholar]
- Kiss, G.; Michl, H. Uber das Giftsekret der Gelbbauchunke, Bombina variegata L. Toxicon 1962, 1, 33–39. [Google Scholar] [CrossRef]
- Hultmark, D.; Steiner, H.; Rasmuson, T.; Boman, H.G. Insect immunity. Purification and properties of three inducible bactericidal proteins from hemolymph of immunized pupae of Hyalophora cecropia. Eur. J. Biochem. 1980, 106, 7–16. [Google Scholar] [CrossRef]
- Steiner, H.; Hultmark, D.; Engström, A.; Bennich, H.; Boman, H.G. Sequence and specificity of two antibacterial proteins involved in insect immunity. Nature 1981, 292, 246–248. [Google Scholar] [CrossRef]
- Buchon, N.; Silverman, N.; Cherry, S. Immunity in Drosophila melanogaster - from microbial recognition to whole-organism physiology. Nature Rev. Immunol. 2014, 14, 796–810. [Google Scholar] [CrossRef]
- Hanson, M.A.; Lemaitre, B. New insights on Drosophila antimicrobial peptide function in host defense and beyond. Curr. Opin. Immunol. 2020, 62, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Eleftherianos, I.; Zhang, W.; Heryanto, C.; Mohamed, A.; Contreras, G.; Tettamanti, G.; Wink, M.; Bassal, T. Diversity of insect antimicrobial peptides and proteins - A functional perspective: A review. Int. J. Biol. Macromol. 2021, 191, 277–287. [Google Scholar] [CrossRef] [PubMed]
- Mylonakis, E.; Podsiadlowski, L.; Muhammed, M.; Vilcinskas, A., Diversity, evolution and medical applications of insect antimicrobial peptides. Philos. T. R. Soc. B 2016, 371, (1695). [CrossRef]
- Yi, H.Y.; Chowdhury, M.; Huang, Y.D.; Yu, X.Q. Insect antimicrobial peptides and their applications. Appl. Microbiol. Biotechnol. 2014, 98, 5807–5822. [Google Scholar] [CrossRef]
- Brady, D.; Grapputo, A.; Romoli, O.; Sandrelli, F. Insect Cecropins, Antimicrobial Peptides with Potential Therapeutic Applications. Int. J. Mol. Sci. 2019, 20, 5862. [Google Scholar] [CrossRef]
- Hemmi, H.; Ishibashi, J.; Hara, S.; Yamakawa, M. Solution structure of moricin, an antibacterial peptide, isolated from the silkworm Bombyx mori. FEBS Lett. 2002, 518, 33–38. [Google Scholar] [CrossRef]
- Sani, M.-A.; Separovic, F. How membrane-active peptides get into lipid membranes. Acc. Chem. Res. 2016, 49, 1130–1138. [Google Scholar] [CrossRef] [PubMed]
- Pöppel, A.K.; Vogel, H.; Wiesner, J.; Vilcinskas, A. Antimicrobial peptides expressed in medicinal maggots of the blow fly Lucilia sericata show combinatorial activity against bacteria. Antimicrob. Agents Chemother 2015, 59, 2508–2514. [Google Scholar] [CrossRef]
- Carboni, A.L.; Hanson, M.A.; Lindsay, S.A.; Wasserman, S.A.; Lemaitre, B.; Andrew, D., Cecropins contribute to Drosophila host defense against a subset of fungal and Gram-negative bacterial infection. Genetics 2022, 220, (1). [CrossRef]
- Wang, Q.; Liu, Y.; He, H.J.; Zhao, X.F.; Wang, J.X. Immune responses of Helicoverpa armigera to different kinds of pathogens. BMC Immunol. 2010, 11, 9. [Google Scholar] [CrossRef]
- Yang, W.; Cheng, T.; Ye, M.; Deng, X.; Yi, H.; Huang, Y.; Tan, X.; Han, D.; Wang, B.; Xiang, Z.; Cao, Y.; Xia, Q. Functional divergence among silkworm antimicrobial peptide paralogs by the activities of recombinant proteins and the induced expression profiles. PLoS One 2011, 6, e18109. [Google Scholar] [CrossRef]
- Dai, H.; Rayaprolu, S.; Gong, Y.; Huang, R.; Prakash, O.; Jiang, H. Solution structure, antibacterial activity, and expression profile of Manduca sexta moricin. J. Pept. Sci. 2008, 14, 855–863. [Google Scholar] [CrossRef]
- Imler, J.L.; Bulet, P. Antimicrobial peptides in Drosophila: structures, activities and gene regulation. Chem. Immunol. Allergy 2005, 86, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Tonk, M.; Knorr, E.; Cabezas-Cruz, A.; Valdés, J.J.; Kollewe, C.; Vilcinskas, A. Tribolium castaneum defensins are primarily active against Gram-positive bacteria. J. Inverteb. Pathol. 2015, 132, 208–215. [Google Scholar] [CrossRef] [PubMed]
- Altincicek, B.; Vilcinskas, A. Analysis of the immune-inducible transcriptome from microbial stress resistant, rat-tailed maggots of the drone fly Eristalis tenax. BMC Genomics 2007, 8, 326. [Google Scholar] [CrossRef]
- Hoffmann, J.A.; Hetru, C. Insect defensins: inducible antibacterial peptides. Immunol. Today 1992, 13, 411–415. [Google Scholar] [CrossRef] [PubMed]
- Vogel, H.; Altincicek, B.; Glöckner, G.; Vilcinskas, A. A comprehensive transcriptome and immune-gene repertoire of the lepidopteran model host Galleria mellonella. BMC Genomics 2011, 12, 308. [Google Scholar] [CrossRef] [PubMed]
- Buchon, N.; Broderick, N.A.; Poidevin, M.; Pradervand, S.; Lemaitre, B. Drosophila Intestinal Response to Bacterial Infection: Activation of Host Defense and Stem Cell Proliferation. Cell Host Microbe 2009, 5, 200–211. [Google Scholar] [CrossRef] [PubMed]
- Fujiwara, S.; Imai, J.; Fujiwara, M.; Yaeshima, T.; Kawashima, T.; Kobayashi, K. A potent antibacterial protein in royal jelly. Purification and determination of the primary structure of royalisin. J. Biol. Chem. 1990, 265, 11333–11337. [Google Scholar] [CrossRef]
- Bilikova, K.; Wu, G.; Simuth, J. Isolation of a peptide fraction from honeybee royal jelly as a potential antifoulbrood factor. Apidologie 2001, 32, 275–283. [Google Scholar] [CrossRef]
- Langen, G.; Imani, J.; Altincicek, B.; Kieseritzky, G.; Kogel, K.H.; Vilcinskas, A. Transgenic expression of gallerimycin, a novel antifungal insect defensin from the greater wax moth Galleria mellonella, confers resistance to pathogenic fungi in tobacco. Biol. Chem. 2006, 387, 549–557. [Google Scholar] [CrossRef]
- Kaneko, Y.; Tanaka, H.; Ishibashi, J.; Iwasaki, T.; Yamakawa, M. Gene expression of a novel defensin antimicrobial peptide in the silkworm, Bombyx mori. Biosci. Biotechnol. Biochem. 2008, 72, 2353–2361. [Google Scholar] [CrossRef]
- Fehlbaum, P.; Bulet, P.; Michaut, L.; Lagueux, M.; Broekaert, W.F.; Hetru, C.; Hoffmann, J.A. Insect immunity. Septic injury of Drosophila induces the synthesis of a potent antifungal peptide with sequence homology to plant antifungal peptides. J. Biol. Chem. 1994, 269, 33159–33163. [Google Scholar] [CrossRef]
- Zhang, Z.T.; Zhu, S.Y. Drosomycin, an essential component of antifungal defence in Drosophila. Insect Mol. Biol. 2009, 18, 549–556. [Google Scholar] [CrossRef]
- Lele, D.S.; Talat, S.; Kumari, S.; Srivastava, N.; Kaur, K.J. Understanding the importance of glycosylated threonine and stereospecific action of Drosocin, a Proline rich antimicrobial peptide. Eur. J. Med. Chem. 2015, 92, 637–647. [Google Scholar] [CrossRef]
- McManus, A.M.; Otvos, L., Jr.; Hoffmann, R.; Craik, D.J. Conformational studies by NMR of the antimicrobial peptide, drosocin, and its non-glycosylated derivative: effects of glycosylation on solution conformation. Biochem. 1999, 38, 705–714. [Google Scholar] [CrossRef]
- Casteels, P.; Ampe, C.; Riviere, L.; Van Damme, J.; Elicone, C.; Fleming, M.; Jacobs, F.; Tempst, P. Isolation and characterization of abaecin, a major antibacterial response peptide in the honeybee (Apis mellifera). Eur. J. Biochem. 1990, 187, 381–386. [Google Scholar] [CrossRef]
- Rao, X.-J.; Xu, X.-X.; Yu, X.-Q. Functional analysis of two lebocin-related proteins from Manduca sexta. Insect Biochem. Mol. Biol. 2012, 42, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Rayaprolu, S.; Wang, Y.; Kanost, M.R.; Hartson, S.; Jiang, H. Functional analysis of four processing products from multiple precursors encoded by a lebocin-related gene from Manduca sexta. Dev. Comp. Immunol. 2010, 34, 638–647. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.L.; Zhan, M.Y.; Zhuo, Y.L.; Dang, X.L.; Li, M.Y.; Xu, Y.; Zhou, X.H.; Yu, X.Q.; Rao, X.J. Characterization of the active fragments of Spodoptera litura Lebocin-1. Arch. Insect Biochem. Physiol. 2020, 103, e21626. [Google Scholar] [CrossRef] [PubMed]
- Bulet, P.; Urge, L.; Ohresser, S.; Hetru, C.; Otvos, L., Jr. Enlarged scale chemical synthesis and range of activity of drosocin, an O-glycosylated antibacterial peptide of Drosophila. Eur. J. Biochem. 1996, 238, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Bulet, P.; Hetru, C.; Dimarcq, J.L.; Hoffmann, D. Antimicrobial peptides in insects; structure and function. Dev. Comp. Immunol. 1999, 23, 329–344. [Google Scholar] [CrossRef]
- Casteels, P.; Ampe, C.; Jacobs, F.; Vaeck, M.; Tempst, P. Apidaecins: antibacterial peptides from honeybees. EMBO J. 1989, 8, 2387–2391. [Google Scholar] [CrossRef]
- Vilcinskas, A.; Mukherjee, K.; Vogel, H. Expansion of the antimicrobial peptide repertoire in the invasive ladybird Harmonia axyridis. Proc. R. Soc. B: Biol. Sci. 2013, 280, 20122113. [Google Scholar] [CrossRef] [PubMed]
- Imamura, M.; Wada, S.; Ueda, K.; Saito, A.; Koizumi, N.; Iwahana, H.; Sato, R. Multipeptide precursor structure of acaloleptin A isoforms, antibacterial peptides from the Udo longicorn beetle, Acalolepta luxuriosa. Dev. Comp. Immunol. 2009, 33, 1120–1127. [Google Scholar] [CrossRef] [PubMed]
- Vilcinskas, A. Anti-infective therapeutics from the Lepidopteran model host Galleria mellonella. Curr. Pharm. Des. 2011, 17, 1240–1245. [Google Scholar] [CrossRef] [PubMed]
- Hedengren, M.; Borge, K.; Hultmark, D. Expression and Evolution of the Drosophila Attacin/Diptericin Gene Family. Biochem. Biophys. Res. Commun. 2000, 279, 574–581. [Google Scholar] [CrossRef]
- Sun, S.C.; Lindström, I.; Lee, J.Y.; Faye, I. Structure and expression of the attacin genes in Hyalophora cecropia. Eur. J. Biochem. 1991, 196, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Hultmark, D.; Engström, A.; Andersson, K.; Steiner, H.; Bennich, H.; Boman, H.G. Insect immunity. Attacins, a family of antibacterial proteins from Hyalophora cecropia. EMBO J. 1983, 2, 571–576. [Google Scholar] [CrossRef] [PubMed]
- Bang, K.; Park, S.; Yoo, J.Y.; Cho, S. Characterization and expression of attacin, an antibacterial protein-encoding gene, from the beet armyworm, Spodoptera exigua (Hübner) (Insecta: Lepidoptera: Noctuidae). Mol. Biol. Rep. 2011, 39, 5151–5159. [Google Scholar] [CrossRef]
- Kwon, Y.M.; Kim, H.J.; Kim, Y.I.; Kang, Y.J.; Lee, I.H.; Jin, B.R.; Han, Y.S.; Cheon, H.M.; Ha, N.G.; Seo, S.J. Comparative analysis of two attacin genes from Hyphantria cunea. Comp. Biochem. Physiol. B: Biochem. Mol. Biol. 2008, 151, 213–220. [Google Scholar] [CrossRef]
- Nesa, J.; Sadat, A.; Buccini, D.F.; Kati, A.; Mandal, A.K.; Franco, O.L. Antimicrobial peptides from Bombyx mori: a splendid immune defense response in silkworms. RSC Adv. 2019, 10, 512–523. [Google Scholar] [CrossRef]
- Xu, X.-X.; Zhong, X.; Yi, H.-Y.; Yu, X.-Q. Manduca sexta gloverin binds microbial components and is active against bacteria and fungi. Dev. Comp. Immunol. 2012, 38, 275–284. [Google Scholar] [CrossRef]
- Axén, A.; Carlsson, A.; Engström, A.; Bennich, H. Gloverin, an antibacterial protein from the immune hemolymph of Hyalophora pupae. Eur. J. Biochem. 1997, 247, 614–619. [Google Scholar] [CrossRef]
- Kawaoka, S.; Katsuma, S.; Daimon, T.; Isono, R.; Omuro, N.; Mita, K.; Shimada, T. Functional analysis of fourGloverin-like genes in the silkworm, Bombyx mori. Arch. Insect Biochem. Physiol. 2008, 67, 87–96. [Google Scholar] [CrossRef]
- Mrinal, N.; Nagaraju, J. Intron loss is associated with gain of function in the evolution of the gloverin family of antibacterial genes in Bombyx mori. J. Biol. Chem. 2008, 283, 23376–23387. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.; Kim, Y. RNA interference of an antimicrobial peptide, gloverin, of the beet armyworm, Spodoptera exigua, enhances susceptibility to Bacillus thuringiensis. J. Inverteb. Pathol. 2011, 108, 194–200. [Google Scholar] [CrossRef]
- Cudic, M.; Bulet, P.; Hoffmann, R.; Craik, D.J.; Otvos, L., Jr. Chemical synthesis, antibacterial activity and conformation of diptericin, an 82-mer peptide originally isolated from insects. Eur. J. Biochem. 1999, 266, 549–558. [Google Scholar] [CrossRef] [PubMed]
- Dimarcq, J.L.; Keppi, E.; Dunbar, B.; Lambert, J.; Reichhart, J.M.; Hoffmann, D.; Rankine, S.M.; Fothergill, J.E.; Hoffmann, J.A. Insect immunity. Purification and characterization of a family of novel inducible antibacterial proteins from immunized larvae of the dipteran Phormia terranovae and complete amino-acid sequence of the predominant member, diptericin A. Eur. J. Biochem. 1988, 171, 17–22. [Google Scholar] [CrossRef]
- Ishikawa, M.; Kubo, T.; Natori, S. Purification and characterization of a diptericin homologue from Sarcophaga peregrina (flesh fly). Biochem. J. 1992, (Pt 2) Pt 2, 573–578. [Google Scholar] [CrossRef]
- Reichhart, J.M.; Meister, M.; Dimarcq, J.L.; Zachary, D.; Hoffmann, D.; Ruiz, C.; Richards, G.; Hoffmann, J.A. Insect immunity: developmental and inducible activity of the Drosophila diptericin promoter. EMBO J. 1992, 11, 1469–1477. [Google Scholar] [CrossRef] [PubMed]
- Gagnon, M.-C.; Strandberg, E.; Grau-Campistany, A.; Wadhwani, P.; Reichert, J.; Bürck, J.; Rabanal, F.; Auger, M.; Paquin, J.-F.; Ulrich, A.S. Influence of the length and charge on the activity of α-helical amphipathic antimicrobial peptides. Biochem. 2017, 56, 1680–1695. [Google Scholar] [CrossRef]
- Dathe, M.; Nikolenko, H.; Meyer, J.; Beyermann, M.; Bienert, M. Optimization of the antimicrobial activity of magainin peptides by modification of charge. FEBS Lett. 2001, 501, 146–150. [Google Scholar] [CrossRef]
- Chen, Y.; Guarnieri, M.T.; Vasil, A.I.; Vasil, M.L.; Mant, C.T.; Hodges, R.S. Role of peptide hydrophobicity in the mechanism of action of alpha-helical antimicrobial peptides. Antimicrob Agents Chemother 2007, 51, 1398–1406. [Google Scholar] [CrossRef]
- Kumar, P.; Kizhakkedathu, J.; Straus, S. Antimicrobial Peptides: Diversity, mechanism of action and strategies to improve the activity and biocompatibility in vivo. Biomolecules 2018, 8, 4. [Google Scholar] [CrossRef]
- Yeaman, M.R.; Yount, N.Y. Mechanisms of antimicrobial peptide action and resistance. Pharmacol. Rev. 2003, 55, 27–55. [Google Scholar] [CrossRef]
- Misof, B.; Liu, S.; Meusemann, K.; Peters, R.S.; Donath, A.; Mayer, C.; Frandsen, P.B.; Ware, J.; Flouri, T.; Beutel, R.G.; Niehuis, O.; Petersen, M.; Izquierdo-Carrasco, F.; Wappler, T.; Rust, J.; Aberer, A.J.; Aspöck, U.; Aspöck, H.; Bartel, D.; Blanke, A.; Berger, S.; Böhm, A.; Buckley, T.R.; Calcott, B.; Chen, J.; Friedrich, F.; Fukui, M.; Fujita, M.; Greve, C.; Grobe, P.; Gu, S.; Huang, Y.; Jermiin, L.S.; Kawahara, A.Y.; Krogmann, L.; Kubiak, M.; Lanfear, R.; Letsch, H.; Li, Y.; Li, Z.; Li, J.; Lu, H.; Machida, R.; Mashimo, Y.; Kapli, P.; McKenna, D.D.; Meng, G.; Nakagaki, Y.; Navarrete-Heredia, J.L.; Ott, M.; Ou, Y.; Pass, G.; Podsiadlowski, L.; Pohl, H.; von Reumont, B.M.; Schütte, K.; Sekiya, K.; Shimizu, S.; Slipinski, A.; Stamatakis, A.; Song, W.; Su, X.; Szucsich, N.U.; Tan, M.; Tan, X.; Tang, M.; Tang, J.; Timelthaler, G.; Tomizuka, S.; Trautwein, M.; Tong, X.; Uchifune, T.; Walzl, M.G.; Wiegmann, B.M.; Wilbrandt, J.; Wipfler, B.; Wong, T.K.F.; Wu, Q.; Wu, G.; Xie, Y.; Yang, S.; Yang, Q.; Yeates, D.K.; Yoshizawa, K.; Zhang, Q.; Zhang, R.; Zhang, W.; Zhang, Y.; Zhao, J.; Zhou, C.; Zhou, L.; Ziesmann, T.; Zou, S.; Li, Y.; Xu, X.; Zhang, Y.; Yang, H.; Wang, J.; Wang, J.; Kjer, K.M.; Zhou, X. Phylogenomics resolves the timing and pattern of insect evolution. Science 2014, 346, 763–767. [Google Scholar] [CrossRef]
- Ma, L.; Liu, S.; Lu, P.; Yan, X.; Hao, C.; Wang, H.; Wei, J.; Qie, X.; Lu, Z. The IMD pathway in Hemipteran: A comparative analysis and discussion. Dev. Comp. Immunol. 2022, 136, 104513. [Google Scholar] [CrossRef] [PubMed]
- Ventola, C.L. The antibiotic resistance crisis: part 2: management strategies and new agents. P. T. 2015, 40, 344–352. [Google Scholar] [PubMed]
- Li, C.; Zhu, C.; Ren, B.; Yin, X.; Shim, S.H.; Gao, Y.; Zhu, J.; Zhao, P.; Liu, C.; Yu, R.; Xia, X.; Zhang, L. Two optimized antimicrobial peptides with therapeutic potential for clinical antibiotic-resistant Staphylococcus aureus. Eur. J. Med. Chem. 2019, 183, 111686. [Google Scholar] [CrossRef] [PubMed]
- Benfield, A.H.; Henriques, S.T. Mode-of-Action of Antimicrobial Peptides: Membrane Disruption vs. Intracellular Mechanisms. Front. Med. Technol. 2020, 2. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, N.; Kahne, D.; Silhavy, T.J. Advances in understanding bacterial outer-membrane biogenesis. Nature Rev. Microbiol. 2006, 4, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Nikaido, H. Molecular basis of bacterial outer membrane permeability revisited. Microbiol. Mol. Biol. Rev. MMBR 2003, 67, 593–656. [Google Scholar] [CrossRef]
- Ma, H.; Zhao, X.; Yang, L.; Su, P.; Fu, P.; Peng, J.; Yang, N.; Guo, G. Antimicrobial peptide AMP-17 Affects Candida albicans by disrupting its cell wall and cell membrane integrity. Infect. Drug. Resist. 2020, 13, 2509–2520. [Google Scholar] [CrossRef]
- Li, J.; Koh, J.-J.; Liu, S.; Lakshminarayanan, R.; Verma, C.S.; Beuerman, R.W. Membrane Active Antimicrobial Peptides: Translating Mechanistic Insights to Design. Front. Neurosci. 2017, 11. [Google Scholar] [CrossRef]
- Meroueh, S.O.; Bencze, K.Z.; Hesek, D.; Lee, M.; Fisher, J.F.; Stemmler, T.L.; Mobashery, S. Three-dimensional structure of the bacterial cell wall peptidoglycan. Proc. Natl. Acad. Sci. U. S. A. 2006, 103, 4404–4409. [Google Scholar] [CrossRef]
- Brogden, K.A. Antimicrobial peptides: pore formers or metabolic inhibitors in bacteria? Nature Rev. Microbiol. 2005, 3, 238–250. [Google Scholar] [CrossRef]
- Le, C.-F.; Fang, C.-M.; Sekaran, S.D., Intracellular targeting mechanisms by antimicrobial peptides. Antimicrob. Agents Chemother. 2017, 61, (4). [CrossRef]
- Sengupta, D.; Leontiadou, H.; Mark, A.E.; Marrink, S.-J. Toroidal pores formed by antimicrobial peptides show significant disorder. Biophys. Acta. 2008, 1778, 2308–2317. [Google Scholar] [CrossRef] [PubMed]
- Hale, J.D.; Hancock, R.E. Alternative mechanisms of action of cationic antimicrobial peptides on bacteria. Expert Rev. Anti Infect. Ther. 2007, 5, 951–959. [Google Scholar] [CrossRef] [PubMed]
- Gazit, E.; Boman, A.; Boman, H.G.; Shai, Y. Interaction of the mammalian antibacterial peptide cecropin P1 with phospholipid vesicles. Biochem. 1995, 34, 11479–11488. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Harroun, T.A.; Weiss, T.M.; Ding, L.; Huang, H.W. Barrel-stave model or toroidal model? A case study on melittin pores. Biophys. J. 2001, 81, 1475–1485. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.P.; Appelberg, R.; Gama, F.M. Antimicrobial peptides as novel anti-tuberculosis therapeutics. Biotechnol. Adv. 2016, 34, 924–940. [Google Scholar] [CrossRef] [PubMed]
- Bolintineanu, D.; Hazrati, E.; Davis, H.T.; Lehrer, R.I.; Kaznessis, Y.N. Antimicrobial mechanism of pore-forming protegrin peptides: 100 pores to kill E. coli. Peptides 2010, 31, 1–8. [Google Scholar] [CrossRef]
- Shah, P.; Hsiao, F.S.; Ho, Y.H.; Chen, C.S. The proteome targets of intracellular targeting antimicrobial peptides. Proteomics 2016, 16, 1225–1237. [Google Scholar] [CrossRef]
- Mardirossian, M.; Grzela, R.; Giglione, C.; Meinnel, T.; Gennaro, R.; Mergaert, P.; Scocchi, M. The host antimicrobial peptide bac71-35 binds to bacterial ribosomal proteins and inhibits protein synthesis. Chem. Biol. 2014, 21, 1639–1647. [Google Scholar] [CrossRef]
- Chesnokova, L.S.; Slepenkov, S.V.; Witt, S.N. The insect antimicrobial peptide, l-pyrrhocoricin, binds to and stimulates the ATPase activity of both wild-type and lidless DnaK. FEBS Lett. 2004, 565, 65–69. [Google Scholar] [CrossRef] [PubMed]
- Li, W.-F.; Ma, G.-X.; Zhou, X.-X. Apidaecin-type peptides: Biodiversity, structure-function relationships and mode of action. Peptides 2006, 27, 2350–2359. [Google Scholar] [CrossRef] [PubMed]
- Donlan, R.M.; Costerton, J.W. Biofilms: Survival Mechanisms of Clinically Relevant Microorganisms. Clin. Microbiol. Rev. 2002, 15, 167–193. [Google Scholar] [CrossRef] [PubMed]
- Batoni, G.; Maisetta, G.; Esin, S. Antimicrobial peptides and their interaction with biofilms of medically relevant bacteria. Biochim. Biophys. Acta. 2016, 1858, 1044–1060. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Kamesh, A.C.; Xiao, Y.; Sun, V.; Hayes, M.; Daniell, H.; Koo, H. Topical delivery of low-cost protein drug candidates made in chloroplasts for biofilm disruption and uptake by oral epithelial cells. Biomaterials 2016, 105, 156–166. [Google Scholar] [CrossRef] [PubMed]
- Lewenza, S. Extracellular DNA-induced antimicrobial peptide resistance mechanisms in Pseudomonas aeruginosa. Front. Microbiol. 2013, 4. [Google Scholar] [CrossRef]
- Duperthuy, M.; Sjöström, A.E.; Sabharwal, D.; Damghani, F.; Uhlin, B.E.; Wai, S.N. Role of the Vibrio cholerae matrix protein Bap1 in cross-resistance to antimicrobial peptides. PLoS Pathog. 2013, 9, e1003620. [Google Scholar] [CrossRef] [PubMed]
- Imler, J.-L. Overview of Drosophila immunity: A historical perspective. Dev. Comp. Immunol. 2014, 42, 3–15. [Google Scholar] [CrossRef]
- King, J.G. Developmental and comparative perspectives on mosquito immunity. Dev. Comp. Immunol. 2020, 103, 103458. [Google Scholar] [CrossRef]
- Lemaitre, B.; Hoffmann, J. The host defense of Drosophila melanogaster. Annu. Rev. Immunol. 2007, 25, 697–743. [Google Scholar] [CrossRef]
- He, Y.; Cao, X.; Li, K.; Hu, Y.; Chen, Y.R.; Blissard, G.; Kanost, M.R.; Jiang, H. A genome-wide analysis of antimicrobial effector genes and their transcription patterns in Manduca sexta. Insect Biochem. Mol. Biol. 2015, 62, 23–37. [Google Scholar] [CrossRef]
- Yu, S.; Luo, F.; Xu, Y.; Zhang, Y.; Jin, L.H. Drosophila innate immunity involves multiple signaling pathways and coordinated communication between different tissues. Front. Immunol. 2022, 13. [Google Scholar] [CrossRef]
- Valanne, S.; Wang, J.-H.; Rämet, M. The Drosophila Toll signaling pathway. J. Immunol. 2011, 186, 649–656. [Google Scholar] [CrossRef]
- Valanne, S.; Vesala, L.; Maasdorp, M.K.; Salminen, T.S.; Rämet, M. The Drosophila Toll pathway in innate immunity: from the core pathway toward effector functions. J. Immunol. (Baltimore, Md. 1950) 2022, 209, 1817–1825. [Google Scholar] [CrossRef] [PubMed]
- Kariyawasam, U.; Gulati, M.; Wang, Y.; Bao, H.; Shan, T.; Li, X.; Cao, X.; Sumathipala, N.; Hu, Y.; Zhang, X.; Boons, G.-J.; Jiang, H. Preferential binding of DAP-PGs by major peptidoglycan recognition proteins found in cell-free hemolymph of Manduca sexta. Insect Biochem. Mol. Biol. 2022, 148, 103827. [Google Scholar] [CrossRef] [PubMed]
- Myllymäki, H.; Valanne, S.; Rämet, M. The Drosophila IMD signaling pathway. J. Immunol. 2014, 192, 3455–3462. [Google Scholar] [CrossRef]
- Liehl, P.; Blight, M.; Vodovar, N.; Boccard, F.; Lemaitre, B. Prevalence of local immune response against oral infection in a Drosophila/Pseudomonas infection model. PLoS Pathog. 2006, 2, e56. [Google Scholar] [CrossRef]
- Tzou, P.; Ohresser, S.; Ferrandon, D.; Capovilla, M.; Reichhart, J.M.; Lemaitre, B.; Hoffmann, J.A.; Imler, J.L. Tissue-specific inducible expression of antimicrobial peptide genes in Drosophila surface epithelia. Immunity 2000, 13, 737–748. [Google Scholar] [CrossRef]
- Bosco-Drayon, V.; Poidevin, M.; Boneca, Ivo G.; Narbonne-Reveau, K.; Royet, J.; Charroux, B. Peptidoglycan sensing by the receptor PGRP-LE in the Drosophila gut induces immune responses to infectious bacteria and tolerance to microbiota. Cell Host Microbe 2012, 12, 153–165. [Google Scholar] [CrossRef]
- Buchon, N.; Broderick, N.A.; Lemaitre, B. Gut homeostasis in a microbial world: insights from Drosophila melanogaster. Nature Rev. Microbiol. 2013, 11, 615–626. [Google Scholar] [CrossRef]
- Neyen, C.; Poidevin, M.; Roussel, A.; Lemaitre, B. Tissue- and ligand-specific sensing of gram-negative infection in drosophila by PGRP-LC isoforms and PGRP-LE. J. Immunol. (Baltimore, Md. 1950) 2012, 189, 1886–1897. [Google Scholar] [CrossRef]
- Marra, A.; Hanson, M.A.; Kondo, S.; Erkosar, B.; Lemaitre, B. Drosophila antimicrobial peptides and lysozymes regulate gut microbiota composition and abundance. mBio 2021, 12, e0082421. [Google Scholar] [CrossRef] [PubMed]
- Sarvari, M.; Mikani, A.; Mehrabadi, M. The innate immune gene Relish and Caudal jointly contribute to the gut immune homeostasis by regulating antimicrobial peptides in Galleria mellonella. Dev. Comp. Immunol. 2020, 110, 103732. [Google Scholar] [CrossRef] [PubMed]
- Vieira, C.S.; Figueiredo, M.B.; Moraes, C. d. S.; Pereira, S.B.; Dyson, P.; Mello, C.B.; Castro, D.P.; Azambuja, P. Azadirachtin interferes with basal immunity and microbial homeostasis in the Rhodnius prolixus midgut. Dev. Comp. Immunol. 2021, 114, 103864. [Google Scholar] [CrossRef] [PubMed]
- Zhai, Z.; Boquete, J.-P.; Lemaitre, B. Cell-specific IMD-NF-κB responses enable simultaneous antibacterial immunity and intestinal epithelial cell shedding upon bacterial infection. Immunity 2018, 48, 897–910.e7. [Google Scholar] [CrossRef] [PubMed]
- Ryu, J.H.; Ha, E.M.; Oh, C.T.; Seol, J.H.; Brey, P.T.; Jin, I.; Lee, D.G.; Kim, J.; Lee, D.; Lee, W.J. An essential complementary role of NF-kappaB pathway to microbicidal oxidants in Drosophila gut immunity. EMBO J. 2006, 25, 3693–3701. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Karpac, J.; Tran, Susan L.; Jasper, H., PGRP-SC2 promotes gut immune homeostasis to limit commensal dysbiosis and extend lifespan, Cell 2014, 156, (1-2), 109-122. [CrossRef]
- Yao, Z.; Cai, Z.; Ma, Q.; Bai, S.; Wang, Y.; Zhang, P.; Guo, Q.; Gu, J.; Lemaitre, B.; Zhang, H. Compartmentalized PGRP expression along the dipteran Bactrocera dorsalis gut forms a zone of protection for symbiotic bacteria. Cell Rep. 2022, 41, 111523. [Google Scholar] [CrossRef] [PubMed]
- Gerardo, N.M.; Altincicek, B.; Anselme, C.; Atamian, H.; Barribeau, S.M.; de Vos, M.; Duncan, E.J.; Evans, J.D.; Gabaldón, T.; Ghanim, M.; Heddi, A.; Kaloshian, I.; Latorre, A.; Moya, A.; Nakabachi, A.; Parker, B.J.; Pérez-Brocal, V.; Pignatelli, M.; Rahbé, Y.; Ramsey, J.S.; Spragg, C.J.; Tamames, J.; Tamarit, D.; Tamborindeguy, C.; Vincent-Monegat, C.; Vilcinskas, A. Immunity and other defenses in pea aphids, Acyrthosiphon pisum. Genome Biol. 2010, 11, R21. [Google Scholar] [CrossRef]
- Zumaya-Estrada, F.A.; Martínez-Barnetche, J.; Lavore, A.; Rivera-Pomar, R.; Rodríguez, M.H., Comparative genomics analysis of triatomines reveals common first line and inducible immunity-related genes and the absence of Imd canonical components among hemimetabolous arthropods. Parasit. Vectors 2018, 11, (1). [CrossRef]
- French, S.S.; DeNardo, D.F.; Moore, M.C. Trade-offs between the reproductive and immune systems: facultative responses to resources or obligate responses to reproduction? Am. Nat. 2007, 170, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Łukasik, P.; van Asch, M.; Guo, H.; Ferrari, J.; Charles, J.; Godfray, H.; van der Putten, W. Unrelated facultative endosymbionts protect aphids against a fungal pathogen. Ecology Lett. 2013, 16, 214–218. [Google Scholar] [CrossRef] [PubMed]
- Tsuchida, T.; Koga, R.; Meng, X.Y.; Matsumoto, T.; Fukatsu, T. Characterization of a facultative endosymbiotic bacterium of the pea aphid Acyrthosiphon pisum. Microb. Ecol. 2005, 49, 126–133. [Google Scholar] [CrossRef]
- Vorburger, C.; Gehrer, L.; Rodriguez, P. A strain of the bacterial symbiont Regiella insecticola protects aphids against parasitoids. Biology Lett. 2010, 6, 109–111. [Google Scholar] [CrossRef]
- Bang, I.S. JAK/STAT signaling in insect innate immunity. Entomol. Res. 2019, 49, 339–353. [Google Scholar] [CrossRef]
- Harrison, D.A. The JAK/STAT pathway. Cold Spring Harb. Perspect. Biol. 2012, 4, a011205–a011205. [Google Scholar] [CrossRef] [PubMed]
- Agaisse, H.; Petersen, U.-M.; Boutros, M.; Mathey-Prevot, B.; Perrimon, N. Signaling role of hemocytes in Drosophila JAK/STAT-dependent response to septic injury, Dev. Cell 2003, 5, 441–450. [Google Scholar] [CrossRef]
- Dong, Y.; Morton, J.C.; Ramirez, J.L.; Souza-Neto, J.A.; Dimopoulos, G. The entomopathogenic fungus Beauveria bassiana activate toll and JAK-STAT pathway-controlled effector genes and anti-dengue activity in Aedes aegypti. Insect Biochem. Mol. Biol. 2012, 42, 126–132. [Google Scholar] [CrossRef]
- Wright, V.M.; Vogt, K.L.; Smythe, E.; Zeidler, M.P. Differential activities of the Drosophila JAK/STAT pathway ligands Upd, Upd2 and Upd3. Cell Signal. 2011, 23, 920–927. [Google Scholar] [CrossRef]
- Myllymäki, H.; Rämet, M. JAK/STAT pathway in Drosophila immunity. Scand. J. Immunol. 2014, 79, 377–385. [Google Scholar] [CrossRef]
- Sadekuzzaman, M.; Kim, Y. Nitric oxide mediates antimicrobial peptide gene expression by activating eicosanoid signaling. PLoS One 2018, 13, e0193282. [Google Scholar] [CrossRef] [PubMed]
- Dijkers, P.F.; O’Farrell, P.H. Drosophila calcineurin promotes induction of innate immune responses. Curr. Biol. 2007, 17, 2087–2093. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Wang, X.; Wei, X.; Chen, J.; Wei, Y.; Jiang, H.; Lu, Z.; Feng, C. Nitric oxide-induced calcineurin a mediates antimicrobial peptide production through the IMD pathway. Front. Immunol. 2022, 13, 905419. [Google Scholar] [CrossRef] [PubMed]
- Kallio, J.; Leinonen, A.; Ulvila, J.; Valanne, S.; Ezekowitz, R.A.; Rämet, M. Functional analysis of immune response genes in Drosophila identifies JNK pathway as a regulator of antimicrobial peptide gene expression in S2 cells. Microbes Infect. 2005, 7, 811–819. [Google Scholar] [CrossRef] [PubMed]
- Delaney, J.R.; Stöven, S.; Uvell, H.; Anderson, K.V.; Engström, Y.; Mlodzik, M. Cooperative control of Drosophila immune responses by the JNK and NF-kappaB signaling pathways. EMBO J. 2006, 25, 3068–3077. [Google Scholar] [CrossRef]
- Nunes, C.; Sucena, É.; Koyama, T. Endocrine regulation of immunity in insects. The FEBS J. 2020, 288, 3928–3947. [Google Scholar] [CrossRef] [PubMed]
- Regan, J.C.; Brandão, A.S.; Leitão, A.B.; Mantas Dias, A.R.; Sucena, E.; Jacinto, A.; Zaidman-Rémy, A. Steroid hormone signaling is essential to regulate innate immune cells and fight bacterial infection in Drosophila. PLoS Pathog. 2013, 9, e1003720. [Google Scholar] [CrossRef]
- Garbuzov, A.; Tatar, M. Hormonal regulation of Drosophila microRNA let-7 and miR-125 that target innate immunity. Fly 2014, 4, 306–311. [Google Scholar] [CrossRef]
- Rus, F.; Flatt, T.; Tong, M.; Aggarwal, K.; Okuda, K.; Kleino, A.; Yates, E.; Tatar, M.; Silverman, N. Ecdysone triggered PGRP-LC expression controls Drosophila innate immunity. EMBO J. 2013, 32, 1626–1638. [Google Scholar] [CrossRef]
- Xiong, X.P.; Kurthkoti, K.; Chang, K.Y.; Li, J.L.; Ren, X.; Ni, J.Q.; Rana, T.M.; Zhou, R. miR-34 modulates innate immunity and ecdysone signaling in Drosophila. PLoS Pathog. 2016, 12, e1006034. [Google Scholar] [CrossRef]
- Chang, M.-M.; Wang, Y.-H.; Yang, Q.-T.; Wang, X.-L.; Wang, M.; Raikhel, A.S.; Zou, Z. Regulation of antimicrobial peptides by juvenile hormone and its receptor, Methoprene-tolerant, in the mosquito Aedes aegypti. Insect Biochem. Mol. Biol. 2021, 128, 103509. [Google Scholar] [CrossRef]
- Zhang, J.; Yang, W.; Xu, J.; Yang, W.; Li, Q.; Zhong, Y.; Cao, Y.; Yu, X.-Q.; Deng, X. Regulation of antimicrobial peptide genes via insulin-like signaling pathway in the silkworm Bombyx mori. Insect Biochem. Mol. Biol. 2018, 103, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Becker, T.; Loch, G.; Beyer, M.; Zinke, I.; Aschenbrenner, A.C.; Carrera, P.; Inhester, T.; Schultze, J.L.; Hoch, M. FOXO-dependent regulation of innate immune homeostasis. Nature 2010, 463, 369–373. [Google Scholar] [CrossRef] [PubMed]
- Nässel, D.R.; Liu, Y.; Luo, J. Insulin/IGF signaling and its regulation in Drosophila. Gen. Comp. Endocrinol. 2015, 221, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Tang, T.; Song, Q.; Wang, Z.; He, K.; Liu, X.; Song, J.; Wang, L.; Yang, Y.; Feng, C. Transcription analysis of the stress and immune response genes to temperature stress in Ostrinia furnacalis. Front. Physiol. 2019, 10, 1289. [Google Scholar] [CrossRef] [PubMed]
- Hou, L.; Shi, Y.; Zhai, P.; Le, G. Inhibition of foodborne pathogens by Hf-1, a novel antibacterial peptide from the larvae of the housefly (Musca domestica) in medium and orange juice. Food Control 2007, 18, 1350–1357. [Google Scholar] [CrossRef]
- Rai, M.; Pandit, R.; Gaikwad, S.; Kövics, G. Antimicrobial peptides as natural bio-preservative to enhance the shelf-life of food. J. Food Sci. Technol. 2016, 53, 3381–3394. [Google Scholar] [CrossRef]
- Liu, Y.; Sameen, D.E.; Ahmed, S.; Dai, J.; Qin, W. Antimicrobial peptides and their application in food packaging. Trends Food Sci. Technol. 2021, 112, 471–483. [Google Scholar] [CrossRef]
- Xu, K.; Zhao, X.; Tan, Y.; Wu, J.; Cai, Y.; Zhou, J.; Wang, X. A systematical review on antimicrobial peptides and their food applications. Biomater. Adv. 2023, 155, 213684. [Google Scholar] [CrossRef]
- Patyra, E.; Kwiatek, K. Insect meals and insect antimicrobial peptides as an alternative for antibiotics and growth promoters in livestock production. Pathogens 2023, 12, 854. [Google Scholar] [CrossRef]
- Sibinga, N.A., M.T. Lee, N. Buchon, E.L. Johnson, V. Selvaraj, H. Marquis., Do antimicrobial peptide levels alter performance of insect-based aquaculture feeds-a study using genetic models of insect immune activation. J. Insects Food Feed 2023, 9, 919-934. [CrossRef]
- Wu, S.; Zhang, F.; Huang, Z.; Liu, H.; Xie, C.; Zhang, J.; Thacker, P.A.; Qiao, S. Effects of the antimicrobial peptide cecropin AD on performance and intestinal health in weaned piglets challenged with Escherichia coli. Peptides 2012, 35, 225–230. [Google Scholar] [CrossRef] [PubMed]
- Jan, P.-S.; Huang, H.-Y.; Chen, H.-M. Expression of a synthesized gene encoding cationic peptide cecropin B in transgenic tomato plants protects against bacterial diseases. Appl. Environ. Microb. 2010, 76, 769–775. [Google Scholar] [CrossRef]
- Rahnamaeian, M.; Vilcinskas, A. Defense gene expression is potentiated in transgenic barley expressing antifungal peptide metchnikowin throughout powdery mildew challenge. J. Plant Res. 2011, 125, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Hilchie, A.L.; Hoskin, D.W.; Power Coombs, M.R. Anticancer activities of natural and synthetic peptides. Adv. Exp. Med. Biol. 2019, 1117, 131–147. [Google Scholar] [CrossRef]
- Jin, X.; Mei, H.; Li, X.; Ma, Y.; Zeng, A.-h.; Wang, Y.; Lu, X.; Chu, F.; Wu, Q.; Zhu, J. Apoptosis-inducing activity of the antimicrobial peptide cecropin of Musca domestica in human hepatocellular carcinoma cell line BEL-7402 and the possible mechanism. Acta Biochim. Biophys. Sin. 2010, 42, 259–265. [Google Scholar] [CrossRef] [PubMed]
- Parvy, J.P.; Yu, Y.; Dostalova, A.; Kondo, S.; Kurjan, A.; Bulet, P.; Lemaître, B.; Vidal, M.; Cordero, J.B. The antimicrobial peptide defensin cooperates with tumour necrosis factor to drive tumour cell death in Drosophila. Elife 2019, 8, e45061. [Google Scholar] [CrossRef]
- Biswaro, L.S.; da Costa Sousa, M.G.; Rezende, T.M.B.; Dias, S.C.; Franco, O.L. Antimicrobial peptides and nanotechnology, recent advances and challenges. Front. Microbiol. 2018, 9, 855. [Google Scholar] [CrossRef]
- Hanson, M.A.; Dostálová, A.; Ceroni, C.; Poidevin, M.; Kondo, S.; Lemaitre, B. Synergy and remarkable specificity of antimicrobial peptides in vivo using a systematic knockout approach. eLife 2019, 8. [Google Scholar] [CrossRef]
- Unckless, Robert L.; Howick, Virginia M.; Lazzaro, Brian P., Convergent balancing selection on an antimicrobial peptide in Drosophila. Curr. Biol. 2016, 26, 257-262. [CrossRef]
- Krams, I.; Daukšte, J.; Kivleniece, I.; Kaasik, A.; Krama, T.; Freeberg, T.M.; Rantala, M.J. Trade-off between cellular immunity and life span in mealworm beetles Tenebrio molitor. Curr. Zool. 2013, 59, 340–346. [Google Scholar] [CrossRef]
- Bedick, J.C.; Tunaz, H.; Nor Aliza, A.R.; Putnam, S.M.; Ellis, M.D.; Stanley, D.W. Eicosanoids act in nodulation reactions to bacterial infections in newly emerged adult honey bees, Apis mellifera, but not in older foragers. Comp. Biochem. Physiol. Part C 2001, 130, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.; Koo, H.; Richman, A.M.; Seeley, D.; Vizioli, J.; Klocko, A.D.; O’Brochta, D.A. Ectopic expression of a cecropin transgene in the human malaria vector mosquito Anopheles gambiae (Diptera: Culicidae): effects on susceptibility to Plasmodium. J. Med. Entomol. 2004, 41, 447–455. [Google Scholar] [CrossRef] [PubMed]
- Kokoza, V.; Ahmed, A.; Woon Shin, S.; Okafor, N.; Zou, Z.; Raikhel, A.S. Blocking of Plasmodium transmission by cooperative action of Cecropin A and Defensin A in transgenic Aedes aegypti mosquitoes. Proc. Natl. Acad. Sci. U. S. A. 2010, 107, 8111–8116. [Google Scholar] [CrossRef] [PubMed]
- Feng, M.; Fei, S.; Xia, J.; Labropoulou, V.; Swevers, L.; Sun, J. Antimicrobial peptides as potential antiviral factors in insect antiviral immune response. Front. Immunol. 2020, 11, 2030. [Google Scholar] [CrossRef]
- Malheiros, P. da. S.; Daroit, D.J.; Brandelli, A. Food applications of liposome-encapsulated antimicrobial peptides. Trends Food Sci. Technol. 2010, 21, 284–292. [Google Scholar] [CrossRef]
- Moretta, A.; Salvia, R.; Scieuzo, C.; Di Somma, A.; Vogel, H.; Pucci, P.; Sgambato, A.; Wolff, M.; Falabella, P. A bioinformatic study of antimicrobial peptides identified in the Black Soldier Fly (BSF) Hermetia illucens (Diptera: Stratiomyidae). Sci. Rep. 2020, 10, 16875. [Google Scholar] [CrossRef]
- Christophides, G.K.; Zdobnov, E.; Barillas-Mury, C.; Birney, E.; Blandin, S.; Blass, C.; Brey, P.T.; Collins, F.H.; Danielli, A.; Dimopoulos, G.; Hetru, C.; Hoa,; Hoffmann, J.A.; Kanzok, S.M.; Letunic, I.; Levashina, E.A.; Loukeris, T.G.; Lycett, G.; Meister, S.; Michel, K.; Moita, L.F.; Müller, H.M.; Osta, M.A.; Paskewitz, S.M.; Reichhart, J.M.; Rzhetsky, A.; Troxler, L.; Vernick, K.D.; Vlachou, D.; Volz, J.; von Mering, C.; Xu, J.; Zheng, L.; Bork, P.; Kafatos, F.C., Immunity-related genes and gene families in Anopheles gambiae. Science 2002, 298, 159-165. [CrossRef]
- Evans, J.D.; Aronstein, K.; Chen, Y.P.; Hetru, C.; Imler, J.L.; Jiang, H.; Kanost, M.; Thompson, G.J.; Zou, Z.; Hultmark, D. Immune pathways and defence mechanisms in honey bees Apis mellifera. Insect Mol. Biol. 2006, 15, 645–656. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, H.; Ishibashi, J.; Fujita, K.; Nakajima, Y.; Sagisaka, A.; Tomimoto, K.; Suzuki, N.; Yoshiyama, M.; Kaneko, Y.; Iwasaki, T.; Sunagawa, T.; Yamaji, K.; Asaoka, A.; Mita, K.; Yamakawa, M. A genome-wide analysis of genes and gene families involved in innate immunity of Bombyx mori. Insect Biochem. Mol. Biol. 2008, 38, 1087–1110. [Google Scholar] [CrossRef]
- Xiong, G.H.; Xing, L.S.; Lin, Z.; Saha, T.T.; Wang, C.; Jiang, H.; Zou, Z. High throughput profiling of the cotton bollworm Helicoverpa armigera immunotranscriptome during the fungal and bacterial infections. BMC Genom. 2015, 16, 321. [Google Scholar] [CrossRef]
- Zou, Z.; Evans, J.D.; Lu, Z.; Zhao, P.; Williams, M.; Sumathipala, N.; Hetru, C.; Hultmark, D.; Jiang, H. Comparative genomic analysis of the Tribolium immune system. Genome Biol. 2007, 8, R177. [Google Scholar] [CrossRef]
- An, C.; Kanost, M.R. Manduca sexta serpin-5 regulates prophenoloxidase activation and the Toll signaling pathway by inhibiting hemolymph proteinase HP6. Insect Biochem. Mol. Biol. 2010, 40, 683–689. [Google Scholar] [CrossRef] [PubMed]
- Rao, X.J.; Yu, X.Q. Lipoteichoic acid and lipopolysaccharide can activate antimicrobial peptide expression in the tobacco hornworm Manduca sexta. Dev. Comp. Immunol. 2010, 34, 1119–1128. [Google Scholar] [CrossRef] [PubMed]
- Clemmons, A.W.; Lindsay, S.A.; Wasserman, S.A. An effector peptide family required for Drosophila toll-mediated immunity. PLoS Pathog. 2015, 11, e1004876. [Google Scholar] [CrossRef] [PubMed]
- Gregory, S.M.; Cavenaugh, A.; Journigan, V.; Pokorny, A.; Almeida, P.F. A quantitative model for the all-or-none permeabilization of phospholipid vesicles by the antimicrobial peptide cecropin A. Biophys. J. 2008, 94, 1667–1680. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.L.; Zheng, A.L. Genomic organization and regulation of three cecropin genes in Anopheles gambiae. Insect Mol. Biol. 2002, 11, 517–525. [Google Scholar] [CrossRef]
- Lourenço, A.P.; Florecki, M.M.; Simões, Z.L.P.; Evans, J.D. Silencing of Apis mellifera dorsal genes reveals their role in expression of the antimicrobial peptide defensin-1. Insect Mol. Biol. 2018, 27, 577–589. [Google Scholar] [CrossRef]
- Al Souhail, Q.; Hiromasa, Y.; Rahnamaeian, M.; Giraldo, M.C.; Takahashi, D.; Valent, B.; Vilcinskas, A.; Kanost, M.R. Characterization and regulation of expression of an antifungal peptide from hemolymph of an insect, Manduca sexta. Dev. Comp. Immunol. 2016, 61, 258–268. [Google Scholar] [CrossRef]
- Vizioli, J.; Bulet, P.; Hoffmann, J.A.; Kafatos, F.C.; Müller, H.M.; Dimopoulos, G. Gambicin: a novel immune responsive antimicrobial peptide from the malaria vector Anopheles gambiae. Proc. Natl. Acad. Sci. U. S. A. 2001, 98, 12630–12635. [Google Scholar] [CrossRef]
- Schlüns, H.; Crozier, R.H. Relish regulates expression of antimicrobial peptide genes in the honeybee, Apis mellifera, shown by RNA interference. Insect Mol. Biol. 2007, 16, 753–759. [Google Scholar] [CrossRef] [PubMed]
- Levashina, E.A.; Ohresser, S.; Bulet, P.; Reichhart, J.M.; Hetru, C.; Hoffmann, J.A. Metchnikowin, a novel immune-inducible proline-rich peptide from Drosophila with antibacterial and antifungal properties. Eur. J. Biochem. 1995, 233, 694–700. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Larsson, A. AliView: a fast and lightweight alignment viewer and editor for large datasets. Bioinformatics 2014, 30, 3276–3278. [Google Scholar] [CrossRef] [PubMed]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New models and efficient methods for phylogenetic inference in the genomic era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [PubMed]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef]
- Junier, T.; Zdobnov, E.M. The Newick utilities: high-throughput phylogenetic tree processing in the UNIX shell. Bioinformatics 2010, 26, 1669–1670. [Google Scholar] [CrossRef]
- Yu, G.; Smith, D.K.; Zhu, H.; Guan, Y.; Lam, T.T.-Y. ggtree: an r package for visualization and annotation of phylogenetic trees with their covariates and other associated data. Methods Ecol. Evol. 2017, 8, 28–36. [Google Scholar] [CrossRef]
- Durbin, R.; Eddy, S.R.; Krogh, A.; Mitchison, G., Biological sequence analysis: probabilistic models of proteins and nucleic acids, Cambridge University Press, Cambridge, 1998.
- Krogh, A.; Brown, M.; Mian, I.S.; Sjolander, K.; Haussler, D. Hidden Markov models in computational biology. Applications to protein modeling. J. Mol. Biol. 1994, 235, 1501–1531. [Google Scholar] [CrossRef]
- Shen, W.; Le, S.; Li, Y.; Hu, F. SeqKit: A cross-platform and ultrafast toolkit for FASTA/Q file manipulation. PLoS One 2016, 11, e0163962. [Google Scholar] [CrossRef]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef]
Figure 1.
The unrooted Maximum-likelihood tree of different AMP genes. The tree was constructed with IQ-TREE and published sequences, with the optimal evolutionary model (Q.pfam+R4) determined by the ModelFinder algorithm. The sequences are colored by genes (i.e., the names on the outer circle), while the tip labels of the tree are the sequences names. The AMPs are tentatively classified into four groups (I, II, III and IV). SH-aLRT supports from node 1 through 5 (i.e., the inner node labels near the root) are: 71, 93.2,64, 86.9, 53.9.
Figure 1.
The unrooted Maximum-likelihood tree of different AMP genes. The tree was constructed with IQ-TREE and published sequences, with the optimal evolutionary model (Q.pfam+R4) determined by the ModelFinder algorithm. The sequences are colored by genes (i.e., the names on the outer circle), while the tip labels of the tree are the sequences names. The AMPs are tentatively classified into four groups (I, II, III and IV). SH-aLRT supports from node 1 through 5 (i.e., the inner node labels near the root) are: 71, 93.2,64, 86.9, 53.9.
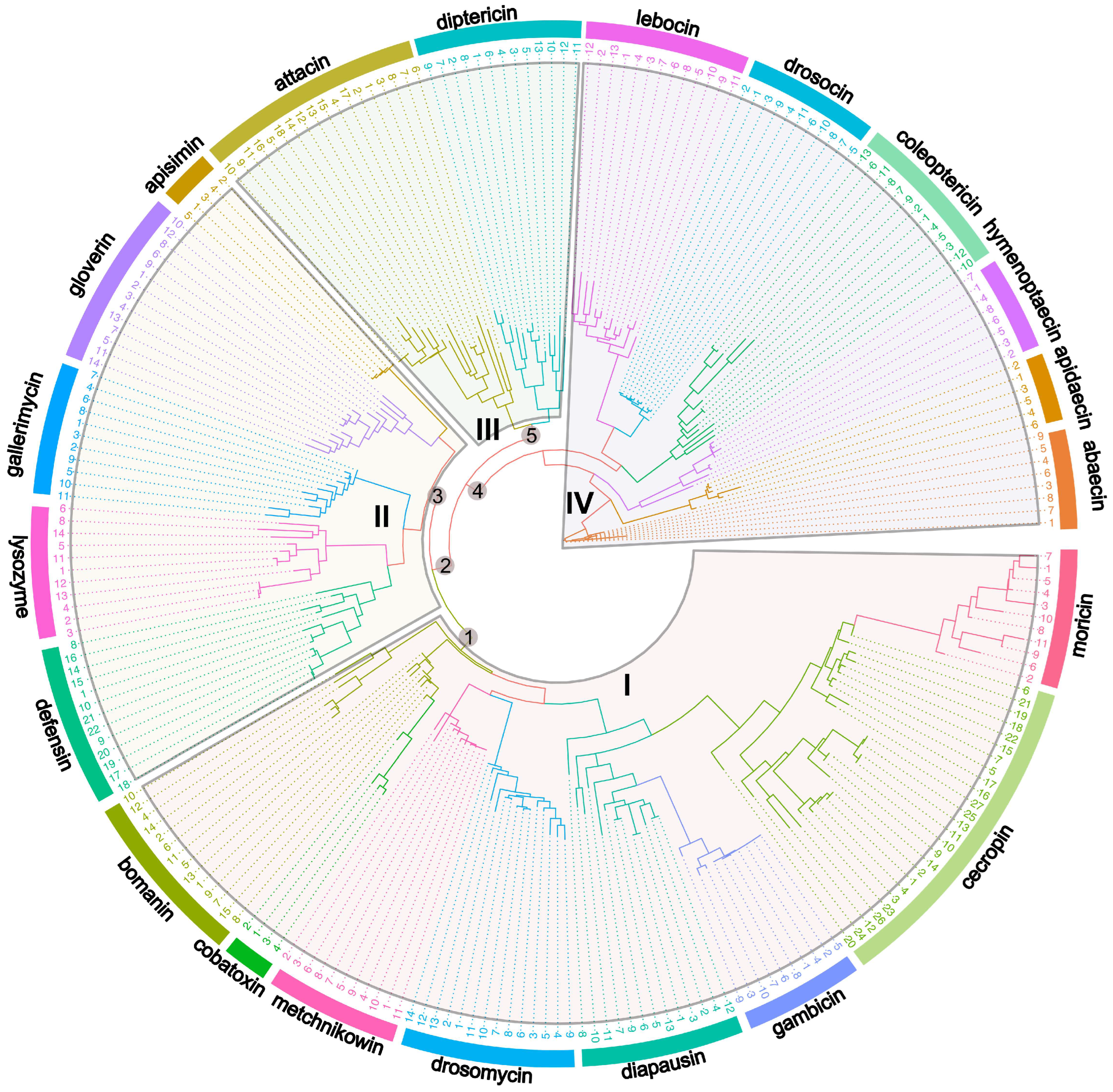
Figure 2.
The phylogenetic distribution patterns of AMPs across insect orders. (a) The phylogenetic relationships of different insect orders, adapted from Misof et al. 2014. The chronostratigraphic scale at the bottom shows the divergence times of different insects. (b) The distribution pattern of 20 AMPs and lysozyme. Light blue and dark blue, presence of AMPs. White, absence of AMPs. Gray background, data not available. Light blue indicates that the determination of AMPs was based on RNA-seq data, while dark blue indicates the determination of AMPs was based on the protein dataset of the corresponding genomes.
Figure 2.
The phylogenetic distribution patterns of AMPs across insect orders. (a) The phylogenetic relationships of different insect orders, adapted from Misof et al. 2014. The chronostratigraphic scale at the bottom shows the divergence times of different insects. (b) The distribution pattern of 20 AMPs and lysozyme. Light blue and dark blue, presence of AMPs. White, absence of AMPs. Gray background, data not available. Light blue indicates that the determination of AMPs was based on RNA-seq data, while dark blue indicates the determination of AMPs was based on the protein dataset of the corresponding genomes.
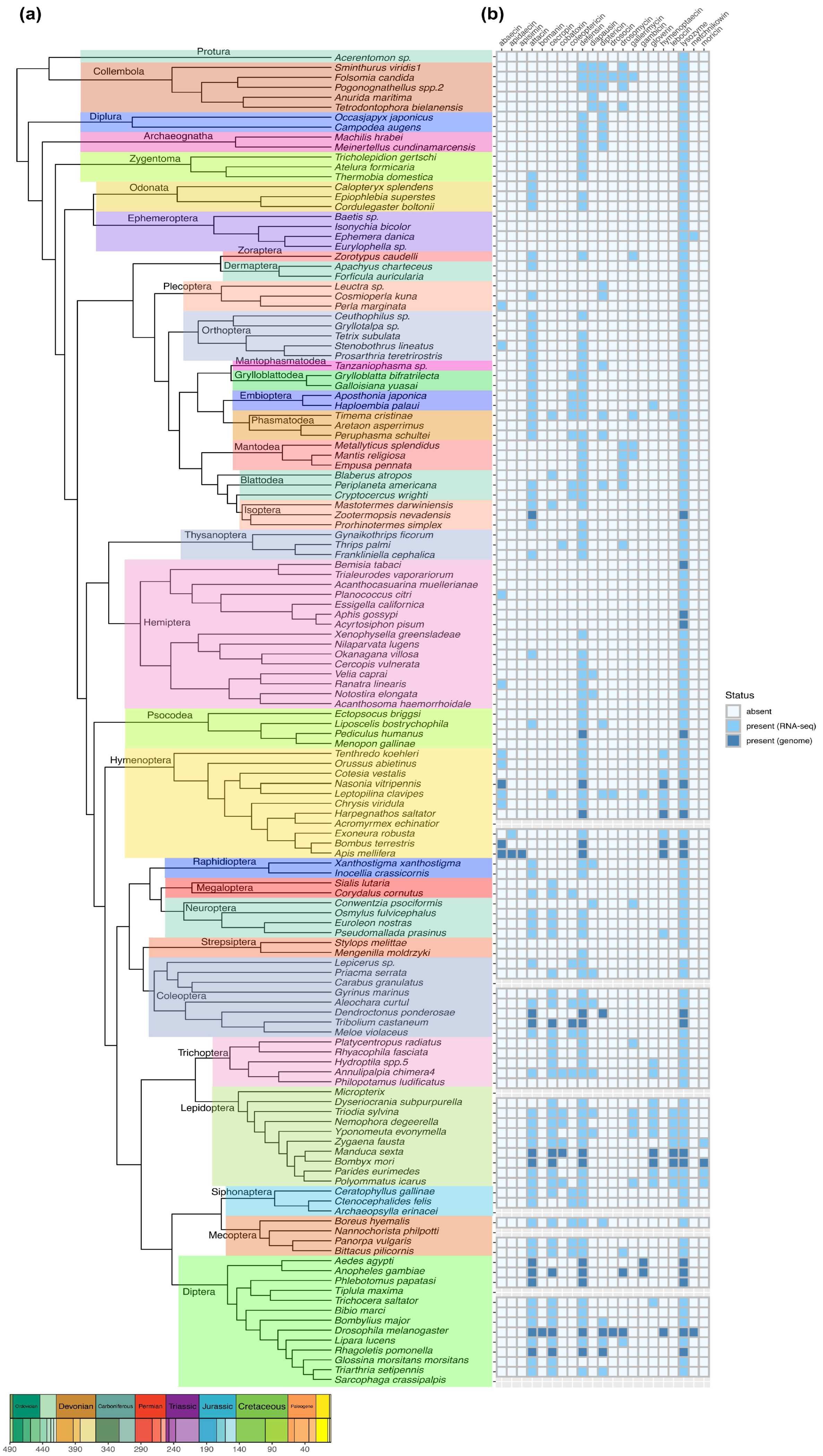
Figure 3.
Mechanisms of action of AMPs. These mechanisms are (a) Bilayer disruption. AMPs insert into the microbial membrane disrupting membrane integrity, (b) Biofilm disruption. AMPs penetrate the biofilm matrix, which is produced by microbes, and enter to kill the embedded pathogens, and (c) Targeting internal components. AMPs target intracellular macromolecules that participate in nucleic acid, protein, and cell wall synthesis, to block cell physiological process.
Figure 3.
Mechanisms of action of AMPs. These mechanisms are (a) Bilayer disruption. AMPs insert into the microbial membrane disrupting membrane integrity, (b) Biofilm disruption. AMPs penetrate the biofilm matrix, which is produced by microbes, and enter to kill the embedded pathogens, and (c) Targeting internal components. AMPs target intracellular macromolecules that participate in nucleic acid, protein, and cell wall synthesis, to block cell physiological process.
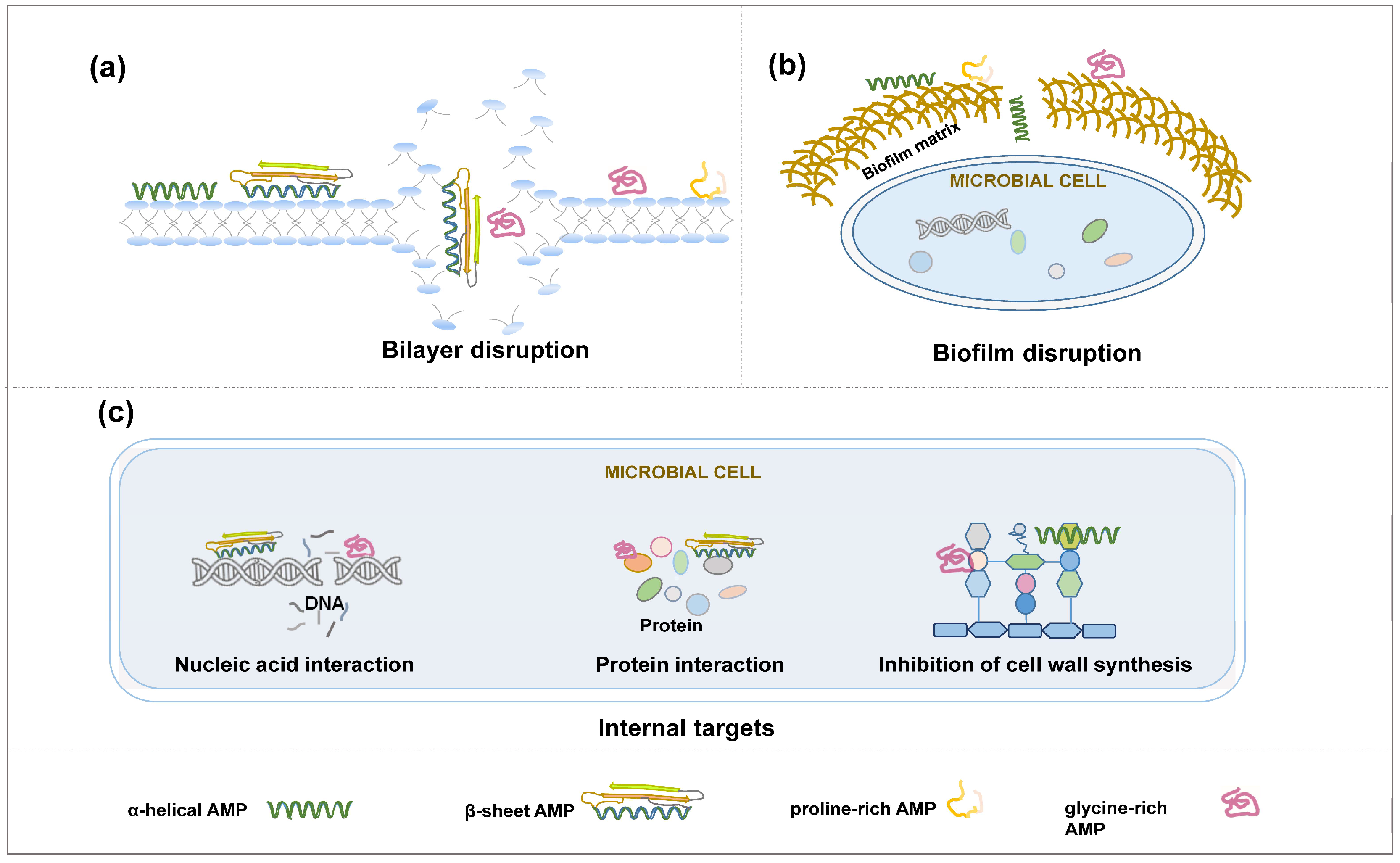
Figure 4.
An overview of signal pathways for producing AMPs in insects. The model is mainly based on the immune pathways of Drosophila. NF-κB pathways are activated upon binding with bacteria and fungi cell wall components by recognition modules. (a) In Toll pathway, cell surface recognition molecules PGRP-SA, PGRP-SD and GNBP1 target Lys-type PGN of Gram-positive bacteria, and GNBP3 recognizes β-glucan of yeast and entomopathogenic fungi to activate Toll signaling. Contrary to common belief, MsPGRPs (from M. sexta) bind to DAP-type PGN of Gram-negative bacteria to activate the Toll pathway. While Toll9 from B. mori act as pattern recognition receptor and directly binds lipopolysaccharide (LPS) to initiate Toll pathway without Spätzle binding. Eventually transcription factor Dorsal and/or Dif translocate into nucleus to induce the expression of AMP genes (bomanin, drosomycin, and metchnikowin). (b) In IMD pathway, pathogen recognition receptors PGRP-LC and PGRP-LE recognize DAP-type PGN of Gram-negative bacteria and some Gram-positive bacteria. These recognition receptors recruit the IMD adaptor to finally activate the transactivator Rel, and Rel translocates into the nucleus to initiate transcription of specific AMP genes (attacin, cecropin, defensin, diptericin, drosocin, and drosomycin). (c) In JAK-STAT pathway, damage signals/septic injury or pathogens induce Unpaired (Upd) expression, and the pathway is activated through binding of Upd to Dome. STAT, which is phosphorylated by JAK, then separates from Dome, dimerizes and enters into nucleus to induce AMPs transcription (drosomycin). Besides, AMPs can be regulated by other signaling pathways including insulin-like signaling, ecdysone signaling, NO signaling, and JNK pathway. In brief, (d) starvation stress triggers FOXO activation by insulin-like signaling, and FOXO translocates into nucleus initiating the expression of AMPs without microbe challenge. (e) Gram-negative bacteria activate NO signaling (NOS oxidizes Arginine to generate NO) in hemocyte, and the released X-factor triggers AMPs production by IMD pathways in Drosophila. JNK pathway consists of TAK1, Hep, JNK, Jun/Fox, transcription factors FOXO and AP-1. Appropriate activation of JNK signaling which shares a kinase dTAK1 with IMD pathway contributes to AMPs production. Ecdysone signaling also regulates AMPs production by IMD pathway in Drosophila. Prothoracicotropic hormone (PTTH) promotes the synthesis of ecdysone (e). Active 20E binds to nuclear receptor EcR/USP, initiating the expression of a series of transcription factors. Subsequently transcription factors up-regulate PGRP-LC expression to activate IMD pathway or directly initiate AMPs expression by IMD pathway. Dashed arrows indicate that the path is in deduced.
Figure 4.
An overview of signal pathways for producing AMPs in insects. The model is mainly based on the immune pathways of Drosophila. NF-κB pathways are activated upon binding with bacteria and fungi cell wall components by recognition modules. (a) In Toll pathway, cell surface recognition molecules PGRP-SA, PGRP-SD and GNBP1 target Lys-type PGN of Gram-positive bacteria, and GNBP3 recognizes β-glucan of yeast and entomopathogenic fungi to activate Toll signaling. Contrary to common belief, MsPGRPs (from M. sexta) bind to DAP-type PGN of Gram-negative bacteria to activate the Toll pathway. While Toll9 from B. mori act as pattern recognition receptor and directly binds lipopolysaccharide (LPS) to initiate Toll pathway without Spätzle binding. Eventually transcription factor Dorsal and/or Dif translocate into nucleus to induce the expression of AMP genes (bomanin, drosomycin, and metchnikowin). (b) In IMD pathway, pathogen recognition receptors PGRP-LC and PGRP-LE recognize DAP-type PGN of Gram-negative bacteria and some Gram-positive bacteria. These recognition receptors recruit the IMD adaptor to finally activate the transactivator Rel, and Rel translocates into the nucleus to initiate transcription of specific AMP genes (attacin, cecropin, defensin, diptericin, drosocin, and drosomycin). (c) In JAK-STAT pathway, damage signals/septic injury or pathogens induce Unpaired (Upd) expression, and the pathway is activated through binding of Upd to Dome. STAT, which is phosphorylated by JAK, then separates from Dome, dimerizes and enters into nucleus to induce AMPs transcription (drosomycin). Besides, AMPs can be regulated by other signaling pathways including insulin-like signaling, ecdysone signaling, NO signaling, and JNK pathway. In brief, (d) starvation stress triggers FOXO activation by insulin-like signaling, and FOXO translocates into nucleus initiating the expression of AMPs without microbe challenge. (e) Gram-negative bacteria activate NO signaling (NOS oxidizes Arginine to generate NO) in hemocyte, and the released X-factor triggers AMPs production by IMD pathways in Drosophila. JNK pathway consists of TAK1, Hep, JNK, Jun/Fox, transcription factors FOXO and AP-1. Appropriate activation of JNK signaling which shares a kinase dTAK1 with IMD pathway contributes to AMPs production. Ecdysone signaling also regulates AMPs production by IMD pathway in Drosophila. Prothoracicotropic hormone (PTTH) promotes the synthesis of ecdysone (e). Active 20E binds to nuclear receptor EcR/USP, initiating the expression of a series of transcription factors. Subsequently transcription factors up-regulate PGRP-LC expression to activate IMD pathway or directly initiate AMPs expression by IMD pathway. Dashed arrows indicate that the path is in deduced.
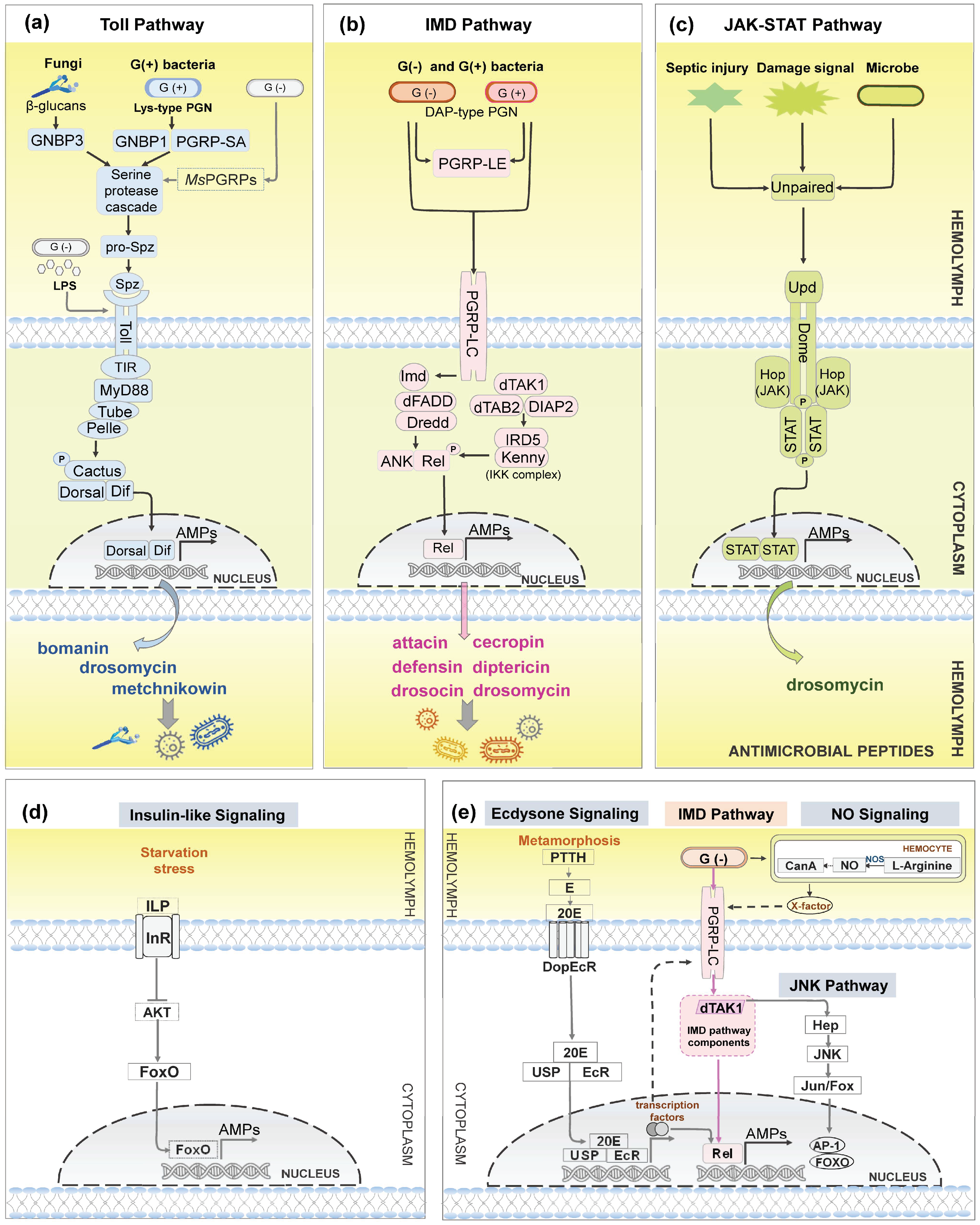

Table 1.
Gene names and number of immune related AMPs from model insects.
| AMP genes | Dm | Ms | Bm | Ha | Tc | Ag | Am |
|---|---|---|---|---|---|---|---|
| abaecin | - | - | - | - | - | - | 1 |
| apidaecin | - | - | - | - | - | - | 2 |
| apisimin | - | - | - | - | - | - | 1 |
| attacin | 4 | 11 | 2 | 1 | 3 | 1 | - |
| bomanin | 3 | - | - | - | - | - | - |
| cecropin | 4 | 15 | 13 | 5 | 3 | 4 | - |
| cobatoxin | - | - | - | 1 | - | - | - |
| coleoptericin | - | - | - | - | 2 | - | - |
| defensin | 1 | 6 | 2 | 1 | 4 | 4 | 2 |
| diapausin | - | 14 | - | - | - | - | - |
| diptericin | 2 | - | - | - | - | - | - |
| drosocin | 1 | - | - | - | - | - | - |
| drosomycin | 7 | - | - | - | - | - | - |
| gambicin | - | - | - | - | - | - | 1 |
| gloverin | - | 1 | 4 | 3 | - | - | - |
| hymenoptaecin | - | - | - | - | - | - | 1 |
| lebocin | - | 4 | 1 | 1 | - | - | - |
| metchnikowin | 1 | - | - | - | - | - | - |
| moricin | - | 6 | 9 | 4 | - | - | - |
Dm, Drosophila melanogaster (Insecta: Diptera); Ms, Manduca sexta (Insecta: Lepidoptera), Bm, Bombyx mori (Insecta: Lepidoptera), Ha, Helicoverpa armigera (Insecta: Lepidoptera); Tc, Tribolium castaneum (Insecta: Coleoptera); Ag, Anopheles gambiae (Insecta: Diptera); Am, Apis mellifera (Insecta: Hymenoptera). “-”, not found. The counts of AMPs listed in this table are mainly from references [31,108,175,176,177,178], and [179].
Table 2.
Model insect antimicrobial peptides and main activity.
| AMP family | Species | Accession number |
Gene name | Main activity |
Immune pathway |
References |
|---|---|---|---|---|---|---|
| abaecin | Apis mellifera | NP_001011617.1 | abaecin | G+, G— | Imd | [48] |
| apidaecin | Apis mellifera | NP_001011642.1 | apidaecin I | G— | nd | [54] |
| apidaecin | Apis mellifera | NP_001011613.1 | apidaecin II | G— | nd | [54] |
| attacin | Drosophila melanogaster | NP_523745.1 | attA | G— | Imd | [39] |
| attacin | Drosophila melanogaster | NP_523746.1 | attB | G— | Imd | [39] |
| attacin | Drosophila melanogaster | NP_523729.3 | attC | G— | Imd | [39] |
| attacin | Drosophila melanogaster | NP_524391.2 | attD | G— | Imd | [39,58] |
| attacin | Bombyx mori | ADB08384.1 | attacin | G+, G— | nd | [63] |
| attacin | Helicoverpa armigera | ADR51155.1 | Haatt | G+, F | nd | [31] |
| attacin | Tribolium castaneum | XP_001809637.1 | Tc-attacin 2 | G+, G— | nd | [179] |
| attacin | Manduca sexta | AAY82587.1 | attacin-1 | G+, | nd | [180] |
| attacin | Manduca sexta | CAL25130.1 | attacin-2 | G+, G— | nd | [181] |
| bomanin | Drosophila melanogaster | NP_611319.1 | IM1-type | G+, F | Toll | [182] |
| bomanin | Drosophila melanogaster | NP_001262823.1 | CG5778-type | G+, F | Toll | [182] |
| bomanin | Drosophila melanogaster | NP_611318.2 | IM23-type | G+, F | Toll | [182] |
| cecropin | Drosophila melanogaster | NP_524588.1 | cecA1 | G— | Imd | [30,39] |
| cecropin | Drosophila melanogaster | NP_524589.1 | cecA2 | G— | Imd | [30,39] |
| cecropin | Drosophila melanogaster | NP_524590.1 | cecB | G— | Imd | [30,39] |
| cecropin | Drosophila melanogaster | NP_524591.1 | cecC | G— | Imd | [30,39] |
| cecropin | Bombyx mori | NP_001037462.1 | Bmcec A1 | G+, G— | nd | [183] |
| cecropin | Bombyx mori | NP_001037460.1 | BmcecB6 | G+, G— | nd | [32] |
| cecropin | Bombyx mori | BAL70382.1 | BmcecD | G+, G— | nd | [32] |
| cecropin | Bombyx mori | NP_001037392.1 | BmcecE | G— | nd | [32] |
| cecropin | Helicoverpa armigera | ADR51154.1 | cecropin-1 | F | nd | [31] |
| cecropin | Helicoverpa armigera | ADR51147.1 | cecropin-2 | G+, G— | nd | [31] |
| cecropin | Helicoverpa armigera | ADR51148.1 | cecropin-3 | F | nd | [31] |
| cecropin | Anopheles gambiae | AAF22649.1 | cecropin A | G+, G— | nd | [184] |
| cecropin | Anopheles gambiae | XP_040173530.1 | cecropin B | G+, G— | nd | [184] |
| cecropin | Manduca sexta | AAO74638.1 | cecropin-6 | G+ | nd | [180] |
| cobatoxin | Helicoverpa armigera | ADR51150.1 | Hacob | G+, G—,F | nd | [31] |
| defensin | Drosophila melanogaster | NP_523672.1 | def | G+, G— | Imd | [39] |
| defensin | Apis mellifera | NP_001011616.1 | Royalisin | G+, F | Toll | [185] |
| defensin | Bombyx mori | NP_001037370.1 | Bmdef | G+, G—,F | Toll,Imd | [43] |
| defensin | Tribolium castaneum | XP_973575.3 | Tcdefensin1 | G+, G—,F | nd | [179] |
| defensin | Tribolium castaneum | XP_968237.2 | Tcdefensin2 | G+, G—,F | nd | [179] |
| diapausin | Manduca sexta | ALP00204.1 | diapausin-1 | F | nd | [186] |
| diptericin | Drosophila melanogaster | NP_476808.1 | dptA | G— | Imd | [39,69] |
| diptericin | Drosophila melanogaster | NP_523787.2 | dptB | G— | Imd | [39,69] |
| drosocin | Drosophila melanogaster | NP_523744.1 | dro | G— | Imd | [52] |
| drosomycin | Drosophila melanogaster | NP_523901.1 | drs | G—, F | Toll, Imd | [44],[45] |
| drosomycin | Drosophila melanogaster | NP_728872.1 | drs-like1 | G— | JAK-STAT | [39] |
| drosomycin | Drosophila melanogaster | AAF47756.2 | drs-like2 | G— | JAK-STAT | [39] |
| drosomycin | Drosophila melanogaster | NP_728861.1 | drs-like3 | G— | JAK-STAT | [39] |
| drosomycin | Drosophila melanogaster | NP_728862.1 | drs-like4 | G— | JAK-STAT | [39] |
| drosomycin | Drosophila melanogaster | AAF47757.1 | drs-like5 | G— | JAK-STAT | [39] |
| drosomycin | Drosophila melanogaster | AAF47765.1 | drs-like6 | G— | JAK-STAT | [39] |
| gambicin | Anopheles gambiae | ACA05604.1 | gambicin | G+, G— | nd | [187] |
| gloverin | Manduca sexta | CAL25129.1 | Msglv | G+, G—,F | nd | [64] |
| gloverin | Bombyx mori | NP_001036930.1 | Bmglv1 | G+, G— | nd | [32],[69] |
| gloverin | Bombyx mori | NP_001037683.1 | Bmglv2 | G+, G— | nd | [32],[69] |
| gloverin | Bombyx mori | NP_001093312.1 | Bmglv3 | G+, G— | nd | [32] |
| gloverin | Bombyx mori | NP_001093312.1 | Bmglv4 | G+, G— | nd | [32] |
| gloverin | Helicoverpa armigera | ADR51146.1 | Haglo | G+, G—,F | nd | [31] |
| hymenoptaecin | Apis mellifera | NP_001011615.1 |
hymenoptaecin | G+, G— | Imd | [188] |
| lebocin | Manduca sexta | ADE20197.1 | lebocin B | G+, G—,F | nd | [49] |
| lebocin | Manduca sexta | XP_030038912.2 | lebocin C | G+, G—,F | nd | [49] |
| lebocin | Bombyx mori | sp|P54684.1| | lebocin 1/2 | G+, G— | nd | [51] |
| lebocin | Bombyx mori | NP_001119732.2 | lebocin 3 | G+, G— | nd | [51] |
| moricin | Manduca sexta | sp|Q86MA1.1| | moricin 1 | G+, G— | nd | [32,33] |
| moricin | Bombyx mori | NP_001036829.2 | Bmmor | G+, G— | nd | [32] |
| moricin | Bombyx mori | pdb|1KV4| | morLA1 | G+, G— | nd | [32] |
| moricin | Helicoverpa armigera | ADR51149.1 | Hamor | G+, G—,F | nd | [31] |
| metchnkowin | Drosophila melanogaster | NP_523752.1 | mtk | G+, G—,F | Toll, Imd | [39,189] |
G+, Gram-positive bacteria; G—, Gram-negative bacteria; F, Fungi; nd, not determined.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



