
Submitted:
04 March 2024
Posted:
05 March 2024
You are already at the latest version
Abstract
The differential expression of transcription factors during embryonic development has been selected as the main feature to define the specific subclasses of spinal interneurons. However, recent studies based on single-cell RNA sequencing and transcriptomic experiments suggest that this approach could not be appropriate in the adult spinal cord, where interneurons show overlapping expression profiles, especially in the ventral region. This constitutes a major challenge for the identification and direct targeting of specific populations that could be involved in locomotor recovery after a traumatic spinal cord injury in adults. Current experimental therapies including electrical stimulation, training, pharmacological treatments, or cell implantation that resulted in improvements in locomotor behaviour, rely on the modulation of the activity and connectivity of interneurons located in the surroundings of the lesion core for the formation of detour circuits. However, very few publications clarify the specific identity of these cells. In this work, we review the studies where premotor interneurons were able to create new intraspinal circuits after different kinds of traumatic spinal cord injury, highlighting the difficulties encountered by researchers to classify these populations.
Keywords:
adult spinal cord injury
; interneuron
; V2
; sprouting
; locomotor recovery
1. Introduction
Traumatic spinal cord injury (TSCI) is a syndrome that affects thousands of people every year mainly due to traffic accidents and falls during the adulthood [1]. It substantially affects the quality of life of the patients, even reducing their lifespan, because of the apparition of severe motor, sensory and autonomic disorders (e.g., respiratory, sexual, urinary, cardiovascular, and intestinal dysfunctions) depending on the level and the extent of the [2,3]. In this review, we will focus principally on locomotor impairments. In the central nervous system, taking the spinal cord as a reference (which in its turn is divided rostro-caudally in cervical, thoracic, lumbar, and sacral regions), we can classify the structures responsible for motor control as supraspinal centers and intraspinal circuits. Supraspinal centers (brain, brainstem, cerebellum) [4,5], send the motor orders in a descending way through different tracts towards the spinal cord, being the most studied the corticospinal (CST), rubrospinal, tectospinal and reticulospinal [6,7] tracts, but also from the gigantocellular formation, the cuneiform nucleus, the pedunculopontine nucleus [8,9], and the vestibular system [10,11] (Figure 1). On the other hand, intraspinal locomotor circuits are distributed all along the spinal cord. For example, the more relevant ones for the locomotor rhythm generation are mainly located in the ventral horns (laminae VII, VIII, X) of the lower thoracic, and upper lumbar spinal cord [12]. The main components of intraspinal locomotor circuits are motor neurons (MNs), that receive inputs from supraspinal centers and sensory afferents (in a direct or indirect way) and send their output to muscles, and premotor interneurons (INs). Premotor INs, which have both their soma and prolongations inside the spinal cord, can pattern the final motor output either by connecting directly to MNs, or indirectly through connections with other INs. Some specific types of INs can be more abundant at one spinal level or another according to their function [13,14,15,16]. The production of normal locomotion also requires sensory inputs originating outside the spinal cord. In this case, sensitive (touch, thermal, and nociceptive) information, along with proprioceptive information coming from the neural receptors present in our skin, muscles, joints, and other organs will travel through the sensorimotor afferents, which have their soma inside the dorsal root ganglia (DRG) and will enter the spinal cord through the dorsal horns to reach the intraspinal circuits (Figure 2). Once integrated inside the spinal cord, these inputs will modulate the motor commands, and will in parallel be directed towards the supraspinal centers in an ascending way throughout the lemniscal, anterolateral and spinocerebellar tracts. Hence, the connection between supraspinal centers and the spinal cord is disrupted after a TSCI, while spared spinal circuits around the lesion could still receive signals from the sensory receptors [17]. Interestingly, it was demonstrated decades ago that intraspinal circuits can generate locomotion, as well as the motor reflexes, independently of the supraspinal tracts [18,19]. In this review, we will focus on the premotor INs contained in motor circuits and how they react after different kinds of TSCI. For a broader information about the spinal circuitry responsible for the locomotion in the ventral spinal cord and central pattern generators, authors strongly recommend reviewing the indicated publications [20,21,22,23,24,25].
2. Current Challenges to Characterize Spinal Premotor INTERNEURON populations in the Adult Ventral Spinal Cord
Premotor INs can be primarily classified following general neuronal features such as where they project (ipsilateral if they project to the same hemicord where their soma is located, or commissural when they cross the midline), if their axon is contained in the same spinal segment or they send their prolongations to other spinal levels (short or long; in this case the term “propriospinal INs” is also used to describe INs that connect at least two spinal segments [26,27], which neurotransmitter(s) they release (excitatory or inhibitory) and their firing properties [12,27,28,29]. However, there is still no consensus about the best way to group different subsets of INs in a meaningful manner to be able to study them separately, as well as to describe the interrelations among them, in the adult ventral spinal cord [22,30,31]. Up to date, the most used classification has been the cardinal system, where cardinal cell populations are named according to their progenitor cells and the expression of specific transcription factors during early embryological stages [32]. For instance, the ventral neural tube contains 5 interneuron progenitor domains (pd6, p0, p1, p2 and p3) that produce different interneuron populations (dI6, V0, V1, V2 and V3). These cardinal classes of INs have been further divided into main populations characterized by unique markers like Chx10 for V2a, Gata3 for V2b, Sox1 for V2c and Shox2 for V2d INs (Figure 3). This classification helped researchers to isolate these big clusters of neurons at early stages of development to properly study their contribution to the locomotor process [20,21,22,23,24,25]. However, subsets of these original main populations have been identified over time, based on the expression of other molecular markers, whose function is still unknown [33,34]. Moreover, molecular definition of cardinal neuron subtypes become less robust with time in ventral than in dorsal regions at postnatal stages. That is, dorsal INs become highly distinct over development as ventral INs present overlapping and shared molecular markers in the adult spinal cord [31]. On top of that, the expression of the embryonic markers varies according to the developmental stage and the spinal level of interest. Taken together, it constitutes a drawback for researchers working in spinal cord disorders. First, to gain insight into the general architecture and functioning of the adult spinal cord. Secondly, because in TSCI context, it makes quite difficult to describe which specific subset of spared INs (in the surroundings of the injured area) could be involved in the potential reparation of the spinal cord after a traumatic event. For example, recent outstanding work performed by Hayashi and colleagues [35] in mouse described for the first time the different expression patterns of transcription factors in V2a Chx10+ (Vsx2+) INs from the embryological to the postnatal stages along the whole spinal cord. They demonstrated that V2a INs are composed of 2 subsets that differ in their temporal expression of Chx10 transcription factor, their abundance according to the spinal level (cervical versus lumbar) and their projection targets (supraspinal centers versus neighboring neurons), suggesting that they are present in different motor circuits across the spinal cord. Moreover, Shox2, which is normally used to identify V2d INs, has been also reported as a marker of cervical spinocerebellar neurons, but cannot be used in the lumbar region to identify these cells [15]. Furthermore, in the case of V3 INs, it has been demonstrated that spatial distribution of V3 subsets is linked to their differential neuronal activity, while cell morphology relates to the temporal expression of specific transcription factors, leading to many V3 subtypes [29]. In the last few years, great effort is being done to refine IN clustering by pooling the results of different RNAseq/transcriptomics experiments obtained by many research groups [30,31,36,37]. However, there is a general shared concern about the extreme difficulty to resolve postnatal ventral cell types. It has been hypothesized that, as the cell populations located in the ventral horns belong to a more heterogeneous circuitry than cells located in the dorsal horns, the transcription factors defining them during the early stages of their differentiation are more prone to be downregulated over time to let the floor to the expression of other factors more related to their final postnatal function [31]. Methods complementary to the cardinal classification, such as the clustering according to the cell birth date or location, are being proposed to improve the way of grouping INs. Indeed, recent evidence demonstrated that cardinal categorization does not consider all the described locomotor rhythm-generating neurons (e.g., V2d and the Hb9 expressing INs) [22]. Thus, new databases are being generated to merge all the knowledge coming from different studies, and to relate the spinal information coming from early, young, or adult individuals of different species to help translational research potentially applied to spinal cord injury [38,39].
3. Limitations in Defining the Potential Roles of INs for Spinal Tissue Recovery after a TSCI
The most popular publications describing the potential roles of INs in spinal tissue recovery after a TSCI date from the late nineties-early two thousands, where it was described that spared supraspinal projections (in this case belonging to the corticospinal tract) that were injured due to a traumatic event, were able to create new collaterals through a process called "sprouting", in order to form new synapses with spinal INs around the lesion site as an attempt of bypassing the damage [6,40,41]. These signs of intraspinal tissue reorganization seem to be evident from three weeks after injury [42,43]. In thoracic incomplete lesions in adult animals, INs located in the segment just above the lesion can undergo plasticity processes both at the dendrites and axonal levels. For example, they can increase the number of proximal dendrites, the quantity of total dendrites that crossed the midline and the number of dendritic spines. In fact, hints of sprouting such as growth-cone like endings in these new branches were detected more than 4 months after lesion. Interestingly, injured axons create collateral branches following both ascending and descending directions, which could lead to changes in the original spinal circuitry [44]. However, in this last example as well as in many other works related to adult spinal tissue, INs were not classified as belonging to any of the cardinal subclasses. Additionally, in some cases, INs are described as "all the neuronal cells contained in the spinal cord that are not MNs" (which are clearly distinguishable thanks to their morphological features), and detected at different time points after injury in the surroundings of the lesion site, or inside the lesion core, by general neuronal markers such as β-III-Tubulin, Tau, MAP2, NeuN, or SMI311 instead of population-specific labeling [44,45,46,47]. This is not surprising since, as we still do not know which molecular markers could be specific of ventral adult populations, there is a lack of antibodies or probes to classify them, unless transgenic zebrafish or mouse lines created by using the genetic markers described for embryonic lineage populations are used to follow globally the early defined groups of INs [22]. On top of that, there is also no consensus about (1) how to compare the results regarding propriospinal IN reorganization when different TSCI models are applied (transection, hemisection, or contusion with different injury sizes), (2) whether hints of neurite sprouting can be compared when the lesion occurs at different levels (cervical, thoracic or lumbar), and (3) how to relate the results when different animal experimental models are used (although increasing comparative anatomy studies about the non-injured spinal cord in different species have been published [48,49] (Figure 3). Moreover, because of technical limitations, it is not always possible to confirm that the neuronal components that are bridging the lesion site belong to active neurons, and that they re-establish, at least to some extent, the original connections that exist in the normal non-injured spinal cord. The second and third point are related to what it is known as the neuroanatomical-functional paradox after a TSCI [50]. It has been repeatedly shown that lesions with a similar extent, at the same level, where signs of neuronal sprouting have been detected (both with supraspinal projections or between different kinds of INs), could lead to very different functional consequences in the case of injured cervical and/or thoracic tissue. Regarding animal models, in species with regenerative capacities such as zebrafish, motor restoration can happen with the regeneration of just one-third of the total amount of supraspinal descending axons and intraspinal long projecting INs (connecting the cervical and lumbar levels) present in healthy animals [51]. In small mammals (mice and rats), it has also been described that a very small percentage of fibers recovery (around the 2-7%) could be enough to obtain motor improvements [52]. In adult humans, it is well known that the central nervous system healing capacity is very limited, even if many patients living with chronic TSCI show some degree of spontaneous sensorimotor recovery below the initial spinal injury level. When it is the case, it usually happens during the first nine months after the accident, reaching a plateau around a year post-injury. Importantly, the recovery rates are better in the case of patients with incomplete lesions, thanks to the amount of preserved functional nervous tissue that can be used as a relay to bridge the lesion. Furthermore, the older a patient is, the more difficult will be the recovery, and there is currently not enough data to assess whether gender could have an impact on facing the symptoms. Also, conversion from a complete to an incomplete injury is more common in tetraplegia than paraplegia [3]. All this evidence suggests that there are still essential restorative mechanisms that need to be identified for researchers and clinicians to be able to develop better therapies [16]. However, it is critical to keep in mind that this recovery is not only dependent on neuronal damage, but also to the attenuation and stabilization of other body responses such as the immune response, the vascular damage, and the mechanical stabilization of the tissue, among others. Thus, gain of movement is not only dependent on the formation of axonal projections able to cross the lesion gap [2,5,53].
4. Ventral INs Contribution to Spinal Tissue Restoration in Different TSCI Models
Most of the research community working in TSCI has been focused either on targeting projections from supraspinal centers (mainly from the corticospinal tract), which may create new synapses with spinal INs [54,55], or on propriospinal IN circuitry modulation all along the spinal cord, because it has been demonstrated that spinal tissue reorganization can also occur below the lesion. Small mammals and zebrafish are the main experimental animal model used for INs characterization, followed by non-human primate models.
4.1. Cervical TSCI
When TSCI occurs at the cervical level, major attention has been put on studying its effects on upper limbs motor activity and respiratory circuitry [56]. For example, it seems that propriospinal neurons located in C3-C4 cervical segments can relay cortical signals after a TSCI to restore hand dexterity in non-human primate (NHP) models. Interestingly, these cells are not essential in uninjured individuals but become indispensable for the recovery process [57], as happens with V2a INs at mid-thoracic level [16]. Cervical (C6) right hemisection in adult rats also lead to impairments in the ipsilateral forelimb mobility. Injured animals received the implantation of a biocompatible scaffold placed at the lesion site as an attempt to help spinal tissue regeneration. A substantial number of newly grown vGlut2+ neurites were found at the lesion area four months after the surgery, leading to the hypothesis of a potential reorganization of preexisting reticulo-propriospinal connections coming from the brainstem, even though there were no significant motor improvements at this time point [45]. In a NHP model of incomplete cervical hemisection, electrical stimulation of dorsal roots at low cervical levels of the spinal cord during reach or grasp/pull movement phases improved arm movements likely evoked by residual descending inputs, but the involved neuronal populations remained uncharacterized [58]. Regarding breathing alterations, works have been mainly focused on the affectation of phrenic MNs, which control the movement of the diaphragm, and thus, on potential changes in pre-phrenic INs located in C3-C6 segments [59]. For example, implanted grafts of human neuronal stem cells derived from the H9-NSC line combined with neurotrophic factors (BDNF) have shown good integration in the spinal tissue when applied to cervical (C5) hemisected rat spinal cords, but the possible formation of new synapses with specific classes of interneurons was not investigated [46]. In mammals, the recruitment of developmentally defined V2a neurons during the recovery of walking and breathing suggests that these cells modify their interactions with the circuits that control these functions [57]. After demonstrating that V2a INs reorganize in the cervical tissue after a C2 hemisection [60], Zholudeva and colleagues transplanted neural progenitor cells enriched with iPSC-derived V2a after a lateralized contusion injury in the area comprising C3-C4 spinal segments. V2a-like cells contributed to diaphragm functional improvement one month after treatment [61]. Moreover, it has been shown that expiratory bulbospinal neurons boost their connections with propriospinal INs (but no MNs) in the segments above the lesion after a thoracic lesion [62]. Interestingly, neurons ipsilateral to an upper cervical hemisection (C2) do not present the same activity as those located in the contralateral hemicord, and these changes in neuronal activity can be modulated in hypoxic conditions [63]. Thus, spontaneous circuit reorganization and therapeutic interventions can collaborate to promote motor and respiratory recovery. Moreover, after a high incomplete cervical lesion, sprouting processes happening in affected ascending proprioceptive axons coming from the ipsilateral cervical DRGs, enable these afferents to create new connections in the grey matter with cuneate-projecting INs to boost tactile and proprioceptive function recovery [64]. Afterwards, in Courtine’s lab demonstrated that epidural electrical stimulation could be useful for functional recovery even applied after a cervical lesion. A device targeting the dorsal root entry zones of the lumbosacral spinal cord was implanted in a patient with a chronic TSCI due to a biking accident (incomplete cervical lesion at C5/C6 levels). Importantly, the patient was able to calibrate the device himself without extra medical assistance after the first guided manipulations. After one year of treatment combined with training routines, it allowed him to walk with crutches [65]. Currently, most influential methodologies are based on the electrical stimulation of the dorsal roots coming from the DRGs at caudal regions of the spinal cord [53]. Epidural electrical stimulation supported by training to treat injured animals or humans does not directly target spinal premotor INs, but modulates the activity of proprioceptive and sensory afferents, that in turn, will impact the activity of INs and stimulate neuronal circuit reorganization [51]. Thus, when supraspinal projections are compromised, sensory information could become the main source of control for movement [17,57]. This technology has been also applied after thoracic injuries, that corresponds to the level of choice in the majority of TSCI research approaches.
4.2. Thoracic TSCI
Locomotor activity was improved in mice suffering from a mid-thoracic contusion after the electrical stimulation of the ensemble of thoracic, lumbar, and sacral dorsal roots placed below the lesion, that was applied in combination with rehabilitation training [53]. It was demonstrated that the main actors in locomotor improvements were a subset of V2a cells (Hoxa10+Chx10+) located in the intermediate lamina of the ventral lumbar spinal cord. These cells receive inputs from both reticulospinal neurons coming from the ventral gigantocellular nucleus and from large-diameter sensory afferents of parvalbumin+ neurons located in the DRGs, while they exclusively project to glutamatergic, gabaergic and cholinergic neurons located in the ventral spinal cord. Optogenetic inactivation of lumbar V2a in mice that suffered from TSCI instantly suppressed walking previously enabled by electrical stimulation, while it was immediately restored if IN activity was not disrupted. In fact, animals lacking Chx10+Hoxa10+ INs that suffer from a unilateral lesion failed to fully recover locomotion [53]. Electrical stimulation was also applied in a subpopulation of V2a INs (long-propriospinal ZFH+ INs) connecting the cervical to lumbar cord to boost tissue restoration after a mid-thoracic injury (T8) [54]. However, preservation of long-connecting fibres is not beneficial in all contexts regarding functional recovery. Long ascending propriospinal INs (LAPNs) at L2 connect the lumbar and the cervical enlargements. Given the ventrolateral location of LAPN in the spinal cord white matter, they are more likely to remain uninjured after SCI, so in principle, silencing them after a thoracic TSCI (T9) could disrupt the locomotor recovery. However, silencing spared LAPNs post-SCI improved locomotor function, although the reason of this remains unclear. One possible explanation could be that silencing these LAPNs prevents maladaptive plasticity with sensory afferents, as both proprioceptive and nociceptive afferents caudal to the injury are able to sprout post injury [66].
Furthermore, after TSCI at the thoracic level, de-novo collaterals belonging to spared transected supraspinal tracts that try to bypass the lesion do not always find the correct rewiring paths. To overcome this issue, the controlled delivery of molecules involved in synapse formation have been applied in the surroundings of the injury. For instance, the overexpression of FGF22 in long propriospinal INs in the cervical spinal cord during the first day after injury (T8) reinforces the newly created CST-IN synapses by increasing the formation of excitatory contacts onto them, as well as increases motor neuron survival in the lumbar cord [55]. With the same idea, the controlled delivery of neurotrophic factors below the lesion area led to a substantial recovery of walking after a complete low thoracic injury in mice, where animals showed a gait similar to unilateral injured animals [16]. In this case, the applied molecules (a cocktail of growth and neurotrophic factors) were selected inspired by (embryonic stages when spinal neuronal circuits are being built, to mimic developmental conditions. Most of the involved neurons corresponded to a long-distance projection subtype of V2a INs (Hoxa7+Chx10+Zfhx3+) whose cell bodies are in the mid-thoracic spinal cord, but their axons reach the lumbar part. These results were confirmed through the fact that silencing regenerated Zfhx3+ neurons disrupted the regained improvements in locomotor activity after treatment. Interestingly, it has been demonstrated that the ablation of mid-thoracic INs (including Chx10 INs) has not detectable impact in locomotion in uninjured adult mice, but according to the results shown and in concordance with additional works [16,53], there is increasing evidence that they become key mediators for function recovery, especially after unilateral TSCI. They also found that stimulation of the ventral gigantocellular nucleus induced large motor evoked potentials in leg muscles, revealing that supraspinal centres regained functional access to the lumbar spinal cord thanks to INs reorganization. Finally, locomotor improvements were detected in rats, suffering from a complete transection (T8-T9), that followed a long-lasting training routine combined with a pharmacological treatment based on a 5-HT agonist applied 9 weeks after the beginning of exercise [67]. The authors suggest that this therapy prompts the reorganization of premotor INs connectivity to MNs controlling gastrocnemius and tibilialis anterior located at L4-L5 lumbar levels. Such circuit reorganization could be promoted by dynamic changes in gene expression, resulting in an increased transcription of genes related to axon guidance and regeneration [15].
Additionally, it seems that complete lower thoracic TSCI in adults promote the conversion of excitatory V2 INs (Shox2+) into inhibitory INs in the ventral spinal cord, thus inhibiting instead of activating MNs. So, preventing this phenotype switching by implementing training routines helped to obtain better locomotor outcomes after injury. This neurotransmitter switch did not happen in V3 INs, that receive less connections from propriospinal afferents than V2. Thus, it seems that the maintenance of the original glutamatergic circuitry is also dependent on the proper functioning of propriospinal afferents and in the number of connections with them [68]. In concordance with previous results, detrusor-sphincter impairments that appear in lower transected rats could be also due to alterations both in the excitatory IN activity and in dorsal root afferents happening at the lumbosacral level after injury [69]. Moreover, Shox2 IN glutamatergic/inhibitory inputs are also dysregulated after complete transection at T8-T10 thoracic levels. Although their intrinsic activity is not significantly affected, they become more sensitive to serotonin [70]. These results are in concordance with the hypersensitivity to serotonin that V2a cells located in the lumbar spinal cord presented after a complete T8-T9 transection in adult mice [71]. Other kind of INs, such as dI4 (derived from the differentiation of a hESC cell line in vitro) have been implanted in the lower thoracic hemisected spinal cord, where they survived and integrated in the surroundings of the injured tissue for up to 7 months in a nonhuman spinal cord injury model [72]. Which phenotypic changes occur in these transplanted neuronal populations after transplantation and during integration into the preserved circuits remains in most cases to be investigated. Work in zebrafish demonstrated that V2a (Chx10+) INs generated de novo from endogenous neural progenitors nine days post injury, were able to survive and synapse with motor neurons located far from the lesion site both in larvae [73] and in adult fish. Interestingly, in postnatal stages, V2a contribute to bypass the lesion site following a subtype specific chronological order [5].
4.3. Lumbar TSCI
Regarding the lumbar level, it has been demonstrated that the upper lumbar cord is more relevant for the locomotor function than lower lumbar or lower thoracic levels. So, interest about the potential role of the spared long-ascending propriospinal interneurons, that have their soma located at this region and project to lower cervical levels, in locomotor recovery has been increasing [74]. V3 propriospinal INs are implicated in spasm generation after a complete transection at the sacral level. Interestingly, in this case, cells were not suffering from a neurotransmitter phenotype switching [75], as happens with V2 after a thoracic lesion [68]. Usually, spasm apparition is described because of maladaptive sprouting (V3 creating aberrant connections that are not present in the healthy tissue with other INs and MNs), but Lin and colleagues proposed that they should be also studied as an initial point for mobility restoration, considering that spams could be a manifestation of partially recovered function in the spinal central pattern generators. Taken together, these observations suggest that a reinforcement of proprioceptive signaling to intraspinal circuits located below the lesion site in the absence of normal descending connectivity could be a key determinant modulator of the formation of detour circuits for spontaneous locomotor recovery [17].
5. Conclusions, Limitations, and Perspectives for the Study of the Role of INs in Intraspinal Tissue Remodelling after a TSCI
Currently, there is no agreement about the better way to cluster or name all the populations and subpopulations of premotor INs in the adult ventral spinal cord. It is known that the expression of specific transcription factors at well-defined developmental stages will, at least in part, set their identity and role in the spinal circuitry. For example, it is possible to predict whether a cell would project to distal levels (even to supraspinal centers) or connect locally according to a well-defined combination of active transcription factors at early stages of embryonic development. Conversely, that information is not sufficient to predict their postsynaptic targets, or the neurotransmitter that they will release [30]. This lack of specific markers in the postnatal stages means that, for the moment, researchers remain with the need of generating complex transgenic animals in which the tracing of every IN subpopulation should be designed under the control of transcription factors present at embryonic stages. This implies to verify that all kind of INs belonging to the studied population will be labeled throughout the life of the animals (that is, in the postnatal stages of interest). On the other hand, the use of lineage tracing where the conditional activation of a reporter gene to visualize a specific IN subset in the adulthood is dependent on drug administration during development (e.g., tamoxifen), could lead to undesired abortions [76]. That scenario could be at least partially solved with a broader knowledge of transcription factors or other markers that define/characterize these IN subpopulations at postnatal stages. Additionally, the generation of specific mouse transgenic lines for every IN subset would not be either economically or temporally feasible [22]. For all these reasons, it has been hypothesized that it possibly does not make sense to classify these cells in clusters in the adult spinal cord [31].
In the context of TSCI, it seems that a diversity of mechanisms involved in spinal tissue restoration have not been discovered yet, because, even in the cases where multidisciplinary approaches have been used, results regarding spinal tissue restoration remain puzzling. The sprouting of supraspinal axons to create new durable connections with spinal INs [54,55], along with propriospinal interneurons reorganization both allowing the reconnection of distal parts of the spinal cord after lesion or at a more local level, are crucial for locomotor recovery. Regarding intraspinal circuits, V2a INs are frequently identified as, either autonomously or in response to therapeutic interventions, contributing to functional recovery after TSCI (refs of V2a-related papers). However, this seems due to the higher abundance of markers and tools to identify these cells, including the persistence of Chx10 in a subset of them [14], rather than to a specific propensity of V2a for axonal reorganization or response to stimuli, as other INs display similar organization or function in spinal motor circuits [20,21,22,23,24,25]. Accordingly, advances in IN characterization highlighted other neurons which belong to supraspinal centres that share some genetic features with propriospinal INs, and whose activation impact on the activity of ventral horn neurons [9]. Thus, a better understanding of the identity of spared IN able to sprout and to reconstitute functional circuits in the different injury models at different levels of the spinal cord will be critical [77] to design better therapies to support their survival, and to boost their growth and maintenance for the recovery of spinal tissue functionality [16]. In parallel, being able to isolate and silence the INs forming aberrant connections that result in detrimental outcomes and poorer life quality to the patients, such as the appearance of pain, spasms, or impaired mobility amelioration due to maladaptive adaptations, would also be of great importance [66]. Finally, one group recently suggested that, after a cervical TSCI, injured but spared axons coming from supraspinal centres, such as the cortico-spinal tract, could somehow revert to an embryonic transcriptional state, which would enable them to create collateral neurites for the regeneration of the damaged nervous tissue [78]. If in the coming years it is demonstrated that this reversion also applies to spared premotor INs, along with a better understanding of which subtypes are implicated in this process, this will open great path for the development of more directed and efficient translational strategies for the recovery of locomotor activity.
Acknowledgments
Research in this field in the FC laboratory is supported by grants from the « Fonds spéciaux de recherche » (FSR) of the Université catholique de Louvain, by « Projet de recherche (PDR) » #T.0039.21 and “Crédit de recherche (CDR)” #J.0032.24 fundings of the Fonds de la Recherche Scientifique (F.R.S.-FNRS, Belgium) and by the Association Belge contre les Maladies neuro-Musculaires (ABMM). ADB is a Postdoctoral Researcher and FC is a Research Director of the F.R.S.-FNRS, respectively.
Conflicts of Interest
authors declare that they have no competing interests that could influence the work reported in this paper.
References
- National Spinal Cord Injury Statistical Center, Facts and figures at a glance. Birmingham, AL: University of Alabama at Birmingham, 2023. https://www.nscisc.uab.edu/public/Facts%20and%20Figures%202023%20-%20Final.pdf.
- Ahuja CS, Wilson JR, Nori S, Kotter MRN, Druschel C, Curt A, Fehlings MG. Traumatic spinal cord injury. Nat Rev Dis Primers. 2017 Apr 27;3:17018. PMID: 28447605. [CrossRef]
- Kirshblum S, Snider B, Eren F, Guest J. Characterizing Natural Recovery after Traumatic Spinal Cord Injury. J Neurotrauma. 2021 May 1;38(9):1267-1284. Epub 2021 Jan 22. PMID: 33339474; PMCID: PMC8080912. [CrossRef]
- Guérout N. Plasticity of the Injured Spinal Cord. Cells. 2021 Jul 26;10(8):1886. PMID: 34440655; PMCID: PMC8395000. [CrossRef]
- Huang CX, Wang Z, Cheng J, Zhu Z, Guan NN, Song J. De novo establishment of circuit modules restores locomotion after spinal cord injury in adult zebrafish. Cell Rep. 2022 Oct 25;41(4):111535. PMID: 36288693. [CrossRef]
- Filli L, Engmann AK, Zörner B, et al. Bridging the gap: a reticulo-propriospinal detour bypassing an incomplete spinal cord injury. J Neurosci. 2014;34(40):13399-13410. [CrossRef]
- Asboth L, Friedli L, Beauparlant J, Martinez-Gonzalez C, Anil S, Rey E, Baud L, Pidpruzhnykova G, Anderson MA, Shkorbatova P, Batti L, Pagès S, Kreider J, Schneider BL, Barraud Q, Courtine G. Cortico-reticulo-spinal circuit reorganization enables functional recovery after severe spinal cord contusion. Nat Neurosci. 2018 Apr;21(4):576-588. Epub 2018 Mar 19. PMID: 29556028. [CrossRef]
- Zörner B, Bachmann LC, Filli L, Kapitza S, Gullo M, Bolliger M, Starkey ML, Röthlisberger M, Gonzenbach RR, Schwab ME. Chasing central nervous system plasticity: the brainstem’s contribution to locomotor recovery in rats with spinal cord injury. Brain. 2014 Jun;137(Pt 6):1716-32. Epub 2014 Apr 15. PMID: 24736305. [CrossRef]
- Roussel M, Lafrance-Zoubga D, Josset N, Lemieux M, Bretzner F. Functional contribution of mesencephalic locomotor region nuclei to locomotor recovery after spinal cord injury. Cell Rep Med. 2023 Feb 21;4(2):100946.PMID: 36812893; PMCID: PMC9975330. [CrossRef]
- Kiehn O. Decoding the organization of spinal circuits that control locomotion. Nat Rev Neurosci. 2016 Apr;17(4):224-38. Epub 2016 Mar 3. PMID: 26935168; PMCID: PMC4844028. [CrossRef]
- Leiras R, Cregg JM, Kiehn O. Brainstem Circuits for Locomotion. Annu Rev Neurosci. 2022 Jul 8;45:63-85. Epub 2022 Jan 5. PMID: 34985919. [CrossRef]
- Zholudeva LV, Abraira VE, Satkunendrarajah K, McDevitt TC, Goulding MD, Magnuson DSK, Lane MA. Spinal Interneurons as Gatekeepers to Neuroplasticity after Injury or Disease. J Neurosci. 2021 Feb 3;41(5):845-854. Epub 2021 Jan 20. PMID: 33472820; PMCID: PMC7880285. [CrossRef]
- Francius C, Harris A, Rucchin V, Hendricks TJ, Stam FJ, Barber M, Kurek D, Grosveld FG, Pierani A, Goulding M, Clotman F. Identification of multiple subsets of ventral interneurons and differential distribution along the rostrocaudal axis of the developing spinal cord. PLoS One. 2013 Aug 15;8(8):e70325. PMID: 23967072; PMCID: PMC3744532. [CrossRef]
- Hayashi M, Hinckley CA, Driscoll SP, Moore NJ, Levine AJ, Hilde KL, Sharma K, Pfaff SL. Graded Arrays of Spinal and Supraspinal V2a Interneuron Subtypes Underlie Forelimb and Hindlimb Motor Control. Neuron. 2018 Feb 21;97(4):869-884.e5. Epub 2018 Feb 1. PMID: 29398364; PMCID: PMC8601153. [CrossRef]
- Matson KJE, Russ DE, Kathe C, Hua I, Maric D, Ding Y, Krynitsky J, Pursley R, Sathyamurthy A, Squair JW, Levi BP, Courtine G, Levine AJ. Single cell atlas of spinal cord injury in mice reveals a pro-regenerative signature in spinocerebellar neurons. Nat Commun. 2022 Sep 26;13(1):5628. PMID: 36163250; PMCID: PMC9513082. [CrossRef]
- Squair JW, Milano M, de Coucy A, Gautier M, Skinnider MA, James ND, Cho N, Lasne A, Kathe C, Hutson TH, Ceto S, Baud L, Galan K, Aureli V, Laskaratos A, Barraud Q, Deming TJ, Kohman RE, Schneider BL, He Z, Bloch J, Sofroniew MV, Courtine G, Anderson MA. Recovery of walking after paralysis by regenerating characterized neurons to their natural target region. Science. 2023 Sep 22;381(6664):1338-1345. Epub 2023 Sep 21. PMID: 37733871. [CrossRef]
- Takeoka A, Arber S. Functional Local Proprioceptive Feedback Circuits Initiate and Maintain Locomotor Recovery after Spinal Cord Injury. Cell Rep. 2019 Apr 2;27(1):71-85.e3. PMID: 30943416. [CrossRef]
- Lovely RG, Gregor RJ, Roy RR, Edgerton VR. Effects of training on the recovery of full-weight-bearing stepping in the adult spinal cat. Exp Neurol. 1986 May;92(2):421-35. PMID: 3956672. [CrossRef]
- Sherrington CS, Laslett EE. Observations on some spinal reflexes and the interconnection of spinal segments. J Physiol. 1903 Feb 23;29(1):58-96. PMID: 16992657; PMCID: PMC1540608. [CrossRef]
- Côté MP, Murray LM, Knikou M. Spinal Control of Locomotion: Individual Neurons, Their Circuits and Functions. Front Physiol. 2018 Jun 25;9:784. PMID: 29988534; PMCID: PMC6026662. [CrossRef]
- Ronzano R, Skarlatou S, Barriga BK, Bannatyne BA, Bhumbra GS, Foster JD, Moore JD, Lancelin C, Pocratsky AM, Özyurt MG, Smith CC, Todd AJ, Maxwell DJ, Murray AJ, Pfaff SL, Brownstone RM, Zampieri N, Beato M. Spinal premotor interneurons controlling antagonistic muscles are spatially intermingled. Elife. 2022 Dec 13;11:e81976. PMID: 36512397; PMCID: PMC9844990. [CrossRef]
- Dougherty KJ. Distinguishing subtypes of spinal locomotor neurons to inform circuit function and dysfunction. Curr Opin Neurobiol. 2023 Oct;82:102763. Epub 2023 Aug 21. PMID: 37611531; PMCID: PMC10578609. [CrossRef]
- El Manira A. Modular circuit organization for speed control of locomotor movements. Curr Opin Neurobiol. 2023 Oct;82:102760. Epub 2023 Aug 17. PMID: 37597455. [CrossRef] [PubMed]
- Gosgnach S. Spinal inhibitory interneurons: regulators of coordination during locomotor activity. Front Neural Circuits. 2023 Apr 20;17:1167836. PMID: 37151357; PMCID: PMC10159059. [CrossRef]
- Sengupta M, Bagnall MW. Spinal Interneurons: Diversity and Connectivity in Motor Control. Annu Rev Neurosci. 2023 Jul 10;46:79-99. Epub 2023 Feb 28. PMID: 36854318. [CrossRef]
- Flynn JR, Graham BA, Galea MP, Callister RJ. The role of propriospinal interneurons in recovery from spinal cord injury. Neuropharmacology. 2011 Apr;60(5):809-22. Epub 2011 Jan 18. PMID: 21251920. [CrossRef]
- Laliberte AM, Goltash S, Lalonde NR, et al. Propriospinal neurons: Essential elements of locomotor control in the intact and possibly the injured spinal cord. Front Cell Neurosci. 2019;13:512. [CrossRef]
- Deska-Gauthier D, Zhang Y. The functional diversity of spinal interneurons and locomotor control. Current Opinion in Physiology. 8; 2019, 99-108. [CrossRef]
- Deska-Gauthier D, Borowska-Fielding J, Jones C, Zhang H, MacKay CS, Michail R, Bennett LA, Bikoff JB, Zhang Y. Embryonic temporal-spatial delineation of excitatory spinal V3 interneuron diversity. Cell Rep. 2024 Jan 23;43(1):113635. Epub 2023 Dec 29. PMID: 38160393; PMCID: PMC10877927. [CrossRef]
- Osseward PJ 2nd, Amin ND, Moore JD, Temple BA, Barriga BK, Bachmann LC, Beltran F Jr, Gullo M, Clark RC, Driscoll SP, Pfaff SL, Hayashi M. Conserved genetic signatures parcellate cardinal spinal neuron classes into local and projection subsets. Science. 2021 Apr 23;372(6540):385-393. PMID: 33888637; PMCID: PMC8612134. [CrossRef]
- Roome RB, Levine AJ. The organization of spinal neurons: Insights from single cell sequencing. Curr Opin Neurobiol. 2023 Oct;82:102762. Epub 2023 Aug 30. PMID: 37657185; PMCID: PMC10727478. [CrossRef]
- Lu DC, Niu T, Alaynick WA. Molecular and cellular development of spinal cord locomotor circuitry. Front Mol Neurosci. 2015 Jun 16;8:25. PMID: 26136656; PMCID: PMC4468382. [CrossRef]
- Bikoff JB, Gabitto MI, Rivard AF, Drobac E, Machado TA, Miri A, Brenner-Morton S, Famojure E, Diaz C, Alvarez FJ, Mentis GZ, Jessell TM. Spinal Inhibitory Interneuron Diversity Delineates Variant Motor Microcircuits. Cell. 2016 Mar 24;165(1):207-219. Epub 2016 Mar 3. PMID: 26949184; PMCID: PMC4808435. [CrossRef]
- Delile J, Rayon T, Melchionda M, Edwards A, Briscoe J, Sagner A. Single cell transcriptomics reveals spatial and temporal dynamics of gene expression in the developing mouse spinal cord. Development. 2019 Mar 27;146(12):dev173807. PMID: 30846445; PMCID: PMC6602353. [CrossRef]
- Hayashi M, Hinckley CA, Driscoll SP, Moore NJ, Levine AJ, Hilde KL, Sharma K, Pfaff SL. Graded Arrays of Spinal and Supraspinal V2a Interneuron Subtypes Underlie Forelimb and Hindlimb Motor Control. Neuron. 2018 Feb 21;97(4):869-884.e5. Epub 2018 Feb 1. PMID: 29398364; PMCID: PMC8601153. [CrossRef]
- Rosenberg AB, Roco CM, Muscat RA, Kuchina A, Sample P, Yao Z, Graybuck LT, Peeler DJ, Mukherjee S, Chen W, Pun SH, Sellers DL, Tasic B, Seelig G. Single-cell profiling of the developing mouse brain and spinal cord with split-pool barcoding. Science. 2018 Apr 13;360(6385):176-182. Epub 2018 Mar 15. PMID: 29545511; PMCID: PMC7643870. [CrossRef]
- Sathyamurthy A, Johnson KR, Matson KJE, Dobrott CI, Li L, Ryba AR, Bergman TB, Kelly MC, Kelley MW, Levine AJ. Massively Parallel Single Nucleus Transcriptional Profiling Defines Spinal Cord Neurons and Their Activity during Behavior. Cell Rep. 2018 Feb 20;22(8):2216-2225. PMID: 29466745; PMCID: PMC5849084. [CrossRef]
- Nielson JL, Guandique CF, Liu AW, Burke DA, Lash AT, Moseanko R, Hawbecker S, Strand SC, Zdunowski S, Irvine KA, Brock JH, Nout-Lomas YS, Gensel JC, Anderson KD, Segal MR, Rosenzweig ES, Magnuson DS, Whittemore SR, McTigue DM, Popovich PG, Rabchevsky AG, Scheff SW, Steward O, Courtine G, Edgerton VR, Tuszynski MH, Beattie MS, Bresnahan JC, Ferguson AR. Development of a database for translational spinal cord injury research. J Neurotrauma. 2014 Nov 1;31(21):1789-99. Epub 2014 Jul 31. PMID: 25077610; PMCID: PMC4186058. [CrossRef]
- Russ DE, Cross RBP, Li L, Koch SC, Matson KJE, Yadav A, Alkaslasi MR, Lee DI, Le Pichon CE, Menon V, Levine AJ. A harmonized atlas of mouse spinal cord cell types and their spatial organization. Nat Commun. 2021 Sep 29;12(1):5722. Erratum in: Nat Commun. 2022 Feb 18;13(1):1033. Erratum in: Nat Commun. 2022 Oct 19;13(1):6184. PMID: 34588430; PMCID: PMC8481483. [CrossRef]
- Alstermark B, Lundberg A, Pettersson LG, et al. Motor recovery after serial spinal cord lesions of defined descending pathways in cats. Neurosci Res. 1987;5(1):68-73. [CrossRef]
- Bareyre FM, Kerschensteiner M, Raineteau O, et al. The injured spinal cord spontaneously forms a new intraspinal circuit in adult rats. Nat Neurosci. 2004;7(3):269-277. [CrossRef]
- Courtine G, Song B, Roy RR, et al. Recovery of supraspinal control of stepping via indirect propriospinal relay connections after spinal cord injury. Nat Med. 2008;14(1):69-74. [CrossRef]
- May Z, Fenrich KK, Dahlby J, et al. Following spinal cord injury transected reticulospinal tract axons develop new collateral inputs to spinal interneurons in parallel with locomotor recovery. Neural Plast. 2017;2017:1932875. [CrossRef]
- Meehan CF, Ford TW, Kirkwood PA. Plasticity of thoracic interneurones rostral to a lateral spinal cord lesion. Exp Neurol. 2020 Sep;331:113361. Epub 2020 May 26. PMID: 32464119. [CrossRef]
- Domínguez-Bajo A, González-Mayorga A, Guerrero CR, Palomares FJ, García R, López-Dolado E, Serrano MC. Myelinated axons and functional blood vessels populate mechanically compliant rGO foams in chronic cervical hemisected rats. Biomaterials. 2019 Feb;192:461-474. Epub 2018 Nov 19. PMID: 30502723. [CrossRef]
- Li Y, Tran A, Graham L, Brock J, Tuszynski MH, Lu P. BDNF guides neural stem cell-derived axons to ventral interneurons and motor neurons after spinal cord injury. Exp Neurol. 2023 Jan;359:114259. Epub 2022 Oct 26. PMID: 36309123 Wang et al., 2023. [CrossRef]
- Wang Z, Duan H, Hao F, Hao P, Zhao W, Gao Y, Gu Y, Song J, Li X, Yang Z. Circuit reconstruction of newborn neurons after spinal cord injury in adult rats via an NT3-chitosan scaffold. Prog Neurobiol. 2023 Jan;220:102375. Epub 2022 Nov 21. PMID: 36410665. [CrossRef]
- Lemon RN. Descending pathways in motor control. Annu Rev Neurosci. 2008;31:195-218. PMID: 18558853. [CrossRef]
- Wilson AC, Sweeney LB. Spinal cords: Symphonies of interneurons across species. Front Neural Circuits. 2023 Apr 26;17:1146449. PMID: 37180760; PMCID: PMC10169611. [CrossRef]
- Fouad K, Popovich PG, Kopp MA, Schwab JM. The neuroanatomical-functional paradox in spinal cord injury. Nat Rev Neurol. 2021 Jan;17(1):53-62. Epub 2020 Dec 11. Erratum in: Nat Rev Neurol. 2023 Oct;19(10):635. PMID: 33311711; PMCID: PMC9012488. [CrossRef]
- Cheng J, Guan NN. A fresh look at propriospinal interneurons plasticity and intraspinal circuits remodeling after spinal cord injury. IBRO Neurosci Rep. 2023 Apr 3;14:441-446. PMID: 37388491; PMCID: PMC10300475. [CrossRef]
- Bradbury EJ, McMahon SB. Spinal cord repair strategies: why do they work? Nat Rev Neurosci. 2006 Aug;7(8):644-53. PMID: 16858392. [CrossRef]
- Kathe C, Skinnider MA, Hutson TH, Regazzi N, Gautier M, Demesmaeker R, Komi S, Ceto S, James ND, Cho N, Baud L, Galan K, Matson KJE, Rowald A, Kim K, Wang R, Minassian K, Prior JO, Asboth L, Barraud Q, Lacour SP, Levine AJ, Wagner F, Bloch J, Squair JW, Courtine G. The neurons that restore walking after paralysis. Nature. 2022 Nov;611(7936):540-547. Epub 2022 Nov 9. PMID: 36352232; PMCID: PMC9668750. [CrossRef]
- Van Steenbergen V, Burattini L, Trumpp M, Fourneau J, Aljović A, Chahin M, Oh H, D’Ambra M, FM. Coordinated neurostimulation promotes circuit rewiring and unlocks recovery after spinal cord injury. J Exp Med. 2023 Mar 6;220(3):e20220615. Epub 2022 Dec 26. PMID: 36571760; PMCID: PMC9794600. [CrossRef]
- Aljović A, Jacobi A, Marcantoni M, Kagerer F, Loy K, Kendirli A, Bräutigam J, Fabbio L, Van Steenbergen V, Pleśniar K, Kerschensteiner M, Bareyre FM. Synaptogenic gene therapy with FGF22 improves circuit plasticity and functional recovery following spinal cord injury. EMBO Mol Med. 2023 Feb 8;15(2):e16111. Epub 2023 Jan 5. PMID: 36601738; PMCID: PMC9906383. [CrossRef]
- Satkunendrarajah K, Karadimas SK, Laliberte AM, Montandon G, Fehlings MG. Cervical excitatory neurons sustain breathing after spinal cord injury. Nature. 2018 Oct;562(7727):419-422. Epub 2018 Oct 10. PMID: 30305735. [CrossRef]
- Anderson MA, Squair JW, Gautier M, Hutson TH, Kathe C, Barraud Q, Bloch J, Courtine G. Natural and targeted circuit reorganization after spinal cord injury. Nat Neurosci. 2022 Dec;25(12):1584-1596. Epub 2022 Nov 17. PMID: 36396975. [CrossRef]
- Barra B, Conti S, Perich MG, Zhuang K, Schiavone G, Fallegger F, Galan K, James ND, Barraud Q, Delacombaz M, Kaeser M, Rouiller EM, Milekovic T, Lacour S, Bloch J, Courtine G, Capogrosso M. Epidural electrical stimulation of the cervical dorsal roots restores voluntary upper limb control in paralyzed monkeys. Nat Neurosci. 2022 Jul;25(7):924-934. Epub 2022 Jun 30. PMID: 35773543. [CrossRef]
- Vandeweerd JM, Hontoir F, De Knoop A, De Swert K, Nicaise C. Retrograde Neuroanatomical Tracing of Phrenic Motor Neurons in Mice. J Vis Exp. 2018 Feb 22;(132):56758. PMID: 29553523; PMCID: PMC5931327. [CrossRef]
- Zholudeva LV, Karliner JS, Dougherty KJ, Lane MA. Anatomical Recruitment of Spinal V2a Interneurons into Phrenic Motor Circuitry after High Cervical Spinal Cord Injury. J Neurotrauma. 2017 Nov 1;34(21):3058-3065. Epub 2017 Jun 29. PMID: 28548606; PMCID: PMC5661864. [CrossRef]
- Zholudeva LV, Iyer N, Qiang L, Spruance VM, Randelman ML, White NW, Bezdudnaya T, Fischer I, Sakiyama-Elbert SE, Lane MA. Transplantation of Neural Progenitors and V2a Interneurons after Spinal Cord Injury. J Neurotrauma. 2018 Dec 15;35(24):2883-2903. Epub 2018 Aug 10. PMID: 29873284; PMCID: PMC6306689. [CrossRef]
- Ford TW, Anissimova NP, Meehan CF, Kirkwood PA. Functional plasticity in the respiratory drive to thoracic motoneurons in the segment above a chronic lateral spinal cord lesion. J Neurophysiol. 2016 Jan 1;115(1):554-67. Epub 2015 Oct 21. PMID: 26490290; PMCID: PMC4760466. [CrossRef]
- Streeter KA, Sunshine MD, Patel SR, Gonzalez-Rothi EJ, Reier PJ, Baekey DM, Fuller DD. Mid-cervical interneuron networks following high cervical spinal cord injury. Respir Physiol Neurobiol. 2020 Jan;271:103305. Epub 2019 Sep 22. PMID: 31553921; PMCID: PMC6864252. [CrossRef]
- Granier C, Schwarting J, Fourli E, Laage-Gaupp F, Hennrich AA, Schmalz A, Jacobi A, Wesolowski M, Conzelmann KK, Bareyre FM. Formation of somatosensory detour circuits mediates functional recovery following dorsal column injury. Sci Rep. 2020 Jul 2;10(1):10953. PMID: 32616790; PMCID: PMC7331809. [CrossRef]
- Lorach H, Galvez A, Spagnolo V, Martel F, Karakas S, Intering N, Vat M, Faivre O, Harte C, Komi S, Ravier J, Collin T, Coquoz L, Sakr I, Baaklini E, Hernandez-Charpak SD, Dumont G, Buschman R, Buse N, Denison T, van Nes I, Asboth L, Watrin A, Struber L, Sauter-Starace F, Langar L, Auboiroux V, Carda S, Chabardes S, Aksenova T, Demesmaeker R, Charvet G, Bloch J, Courtine G. Walking naturally after spinal cord injury using a brain-spine interface. Nature. 2023 Jun;618(7963):126-133. Epub 2023 May 24. PMID: 37225984; PMCID: PMC10232367. [CrossRef]
- Shepard CT, Brown BL, Van Rijswijck MA, Zalla RM, Burke DA, Morehouse JR, Riegler AS, Whittemore SR, Magnuson DSK. Silencing long-descending inter-enlargement propriospinal neurons improves hindlimb stepping after contusive spinal cord injuries. Elife. 2023 Dec 15;12:e82944. PMID: 38099572; PMCID: PMC10776087. [CrossRef]
- Khalki L, Sadlaoud K, Lerond J, Coq JO, Brezun JM, Vinay L, Coulon P, Bras H. Changes in innervation of lumbar motoneurons and organization of premotor network following training of transected adult rats. Exp Neurol. 2018 Jan;299(Pt A):1-14. Epub 2017 Sep 14. PMID: 28917641. [CrossRef]
- Bertels H, Vicente-Ortiz G, El Kanbi K, Takeoka A. Neurotransmitter phenotype switching by spinal excitatory interneurons regulates locomotor recovery after spinal cord injury. Nat Neurosci. 2022 May;25(5):617-629. Epub 2022 May 6. PMID: 35524138; PMCID: PMC9076533. [CrossRef]
- Sartori AM, Hofer AS, Scheuber MI, Rust R, Kessler TM, Schwab ME. Slow development of bladder malfunction parallels spinal cord fiber sprouting and interneurons’ loss after spinal cord transection. Exp Neurol. 2022 Feb;348:113937. Epub 2021 Nov 24. PMID: 34826427. [CrossRef]
- Garcia-Ramirez DL, Ha NTB, Bibu S, Stachowski NJ, Dougherty KJ. Spinal cord injury alters spinal Shox2 interneurons by enhancing excitatory synaptic input and serotonergic modulation while maintaining intrinsic properties in mouse. J Neurosci. 2021 May 13;41(27):5833–48. Epub ahead of print. PMID: 34006587; PMCID: PMC8265802. [CrossRef]
- Husch A, Van Patten GN, Hong DN, Scaperotti MM, Cramer N, Harris-Warrick RM. Spinal cord injury induces serotonin supersensitivity without increasing intrinsic excitability of mouse V2a interneurons. J Neurosci. 2012 Sep 19;32(38):13145-54. PMID: 22993431; PMCID: PMC3506248. [CrossRef]
- Zheng X, Zhu B, Xu J, Liu D, Huang Y, Chen D, Liu Z, Guo F, Dong Y, Zhu W, Pan D, Zhang SC, Chen H, Wang W. Human spinal GABA neurons survive and mature in the injured nonhuman primate spinal cord. Stem Cell Reports. 2023 Feb 14;18(2):439-448. Epub 2023 Jan 19. PMID: 36669493; PMCID: PMC9969075. [CrossRef]
- Vasudevan D, Liu YC, Barrios JP, Wheeler MK, Douglass AD, Dorsky RI. Regenerated interneurons integrate into locomotor circuitry following spinal cord injury. Exp Neurol. 2021 Aug;342:113737. Epub 2021 May 4. PMID: 33957107. [CrossRef]
- Danner SM, Shepard CT, Hainline C, Shevtsova NA, Rybak IA, Magnuson DSK. Spinal control of locomotion before and after spinal cord injury. bioRxiv [Preprint]. 2023 Jun 1:2023.03.22.533794. Update in: Exp Neurol. 2023 Jul 25;:114496. PMID: 36993490; PMCID: PMC10055332. [CrossRef]
- Lin S, Li Y, Lucas-Osma AM, Hari K, Stephens MJ, Singla R, Heckman CJ, Zhang Y, Fouad K, Fenrich KK, Bennett DJ. Locomotor-related V3 interneurons initiate and coordinate muscles spasms after spinal cord injury. J Neurophysiol. 2019 Apr 1;121(4):1352-1367. Epub 2019 Jan 9. PMID: 30625014; PMCID: PMC6485742. [CrossRef]
- Lizen B, Claus M, Jeannotte L, Rijli FM, Gofflot F. Perinatal induction of Cre recombination with tamoxifen. Transgenic Res. 2015 Dec;24(6):1065-77. Epub 2015 Sep 22. PMID: 26395370. [CrossRef]
- Punjani N, Deska-Gauthier D, Hachem LD, Abramian M, Fehlings MG. Neuroplasticity and regeneration after spinal cord injury. N Am Spine Soc J. 2023 Jun 8;15:100235. PMID: 37416090; PMCID: PMC10320621. [CrossRef]
- Poplawski GHD, Kawaguchi R, Van Niekerk E, Lu P, Mehta N, Canete P, Lie R, Dragatsis I, Meves JM, Zheng B, Coppola G, Tuszynski MH. Injured adult neurons regress to an embryonic transcriptional growth state. Nature. 2020 May;581(7806):77-82. Epub 2020 Apr 15. PMID: 32376949. [CrossRef]
- Saliani A, Perraud B, Duval T, Stikov N, Rossignol S, Cohen-Adad J. Axon and Myelin Morphology in Animal and Human Spinal Cord. Front Neuroanat. 2017 Dec 22;11:129. PMID: 29311857; PMCID: PMC5743665. [CrossRef]
- Meltzer S, Santiago C, Sharma N, Ginty DD. The cellular and molecular basis of somatosensory neuron development. Neuron. 2021 Dec 1;109(23):3736-3757. Epub 2021 Sep 29. PMID: 34592169; PMCID: PMC8639614. [CrossRef]
Figure 1.
Schemes representing the major white tracts in the human (top; cervical region as example) and in the mouse (bottom) spinal cord. 1Sp, lamina 1; 2Spl, lamina 2 inner part; 2SpO, lamina 2 outer part; 3Sp, lamina 3; 7Sp, lamina 7; CC, central canal; crts, caudal reticulospinal tract; cu, cuneate fasciculus; dcs, dorsal corticospinal tract; dsc, dorsal spinocerebellar tract; dl, dorsolateral fasciculus; dlst, dorsolateral spinothalamic tract; gr, gracile fasciculus; IML, intermediolateral column; lst, lateral spinothalamic tract; LSp, lateral spinal nucleus; lvs, lateral vestibulospinal tract; mvs, medial vestibulospinal tract; pdsc, postsynaptic dorsal column pathway; rrts, rostral reticulospinal tract; rs, rubrospinal tract; SPSy, sacral parasympathic nucleus; vsc, ventral spinocerebellar tract; vst, ventral spinothalamic tract Adapted with permission from [79].
Figure 1.
Schemes representing the major white tracts in the human (top; cervical region as example) and in the mouse (bottom) spinal cord. 1Sp, lamina 1; 2Spl, lamina 2 inner part; 2SpO, lamina 2 outer part; 3Sp, lamina 3; 7Sp, lamina 7; CC, central canal; crts, caudal reticulospinal tract; cu, cuneate fasciculus; dcs, dorsal corticospinal tract; dsc, dorsal spinocerebellar tract; dl, dorsolateral fasciculus; dlst, dorsolateral spinothalamic tract; gr, gracile fasciculus; IML, intermediolateral column; lst, lateral spinothalamic tract; LSp, lateral spinal nucleus; lvs, lateral vestibulospinal tract; mvs, medial vestibulospinal tract; pdsc, postsynaptic dorsal column pathway; rrts, rostral reticulospinal tract; rs, rubrospinal tract; SPSy, sacral parasympathic nucleus; vsc, ventral spinocerebellar tract; vst, ventral spinothalamic tract Adapted with permission from [79].
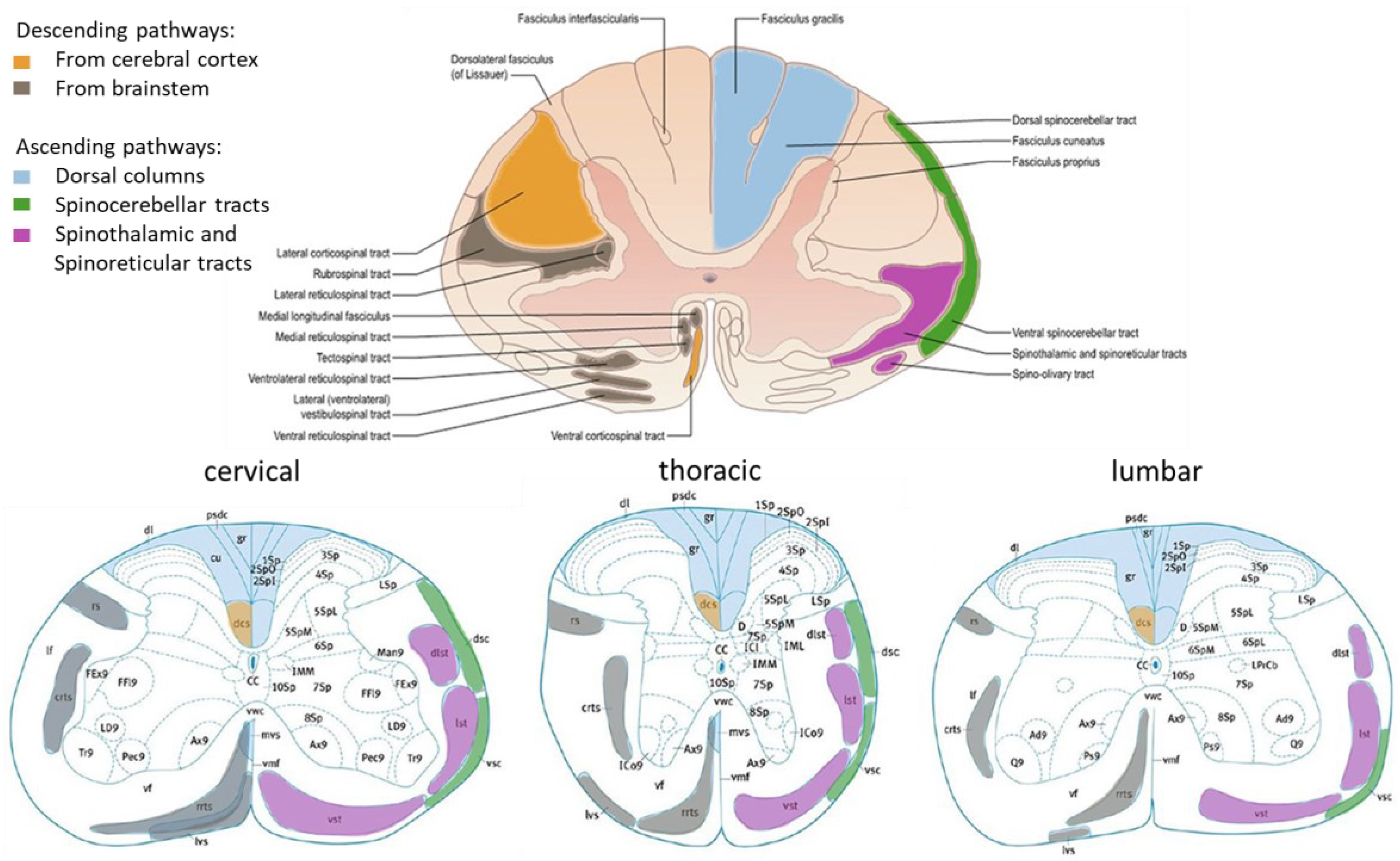
Figure 2.
Left: Schemes of propriospinal INs having their cell body at the cervical (green), thoracic (red) or lumbar levels (blue) of the spinal cord (reproduced from [27]). Right: Schematic representation of sensory afferents which their soma inside the DRGs, transmitting the information to INs and MNs through the dorsal horn (adapted from [80]).
Figure 2.
Left: Schemes of propriospinal INs having their cell body at the cervical (green), thoracic (red) or lumbar levels (blue) of the spinal cord (reproduced from [27]). Right: Schematic representation of sensory afferents which their soma inside the DRGs, transmitting the information to INs and MNs through the dorsal horn (adapted from [80]).
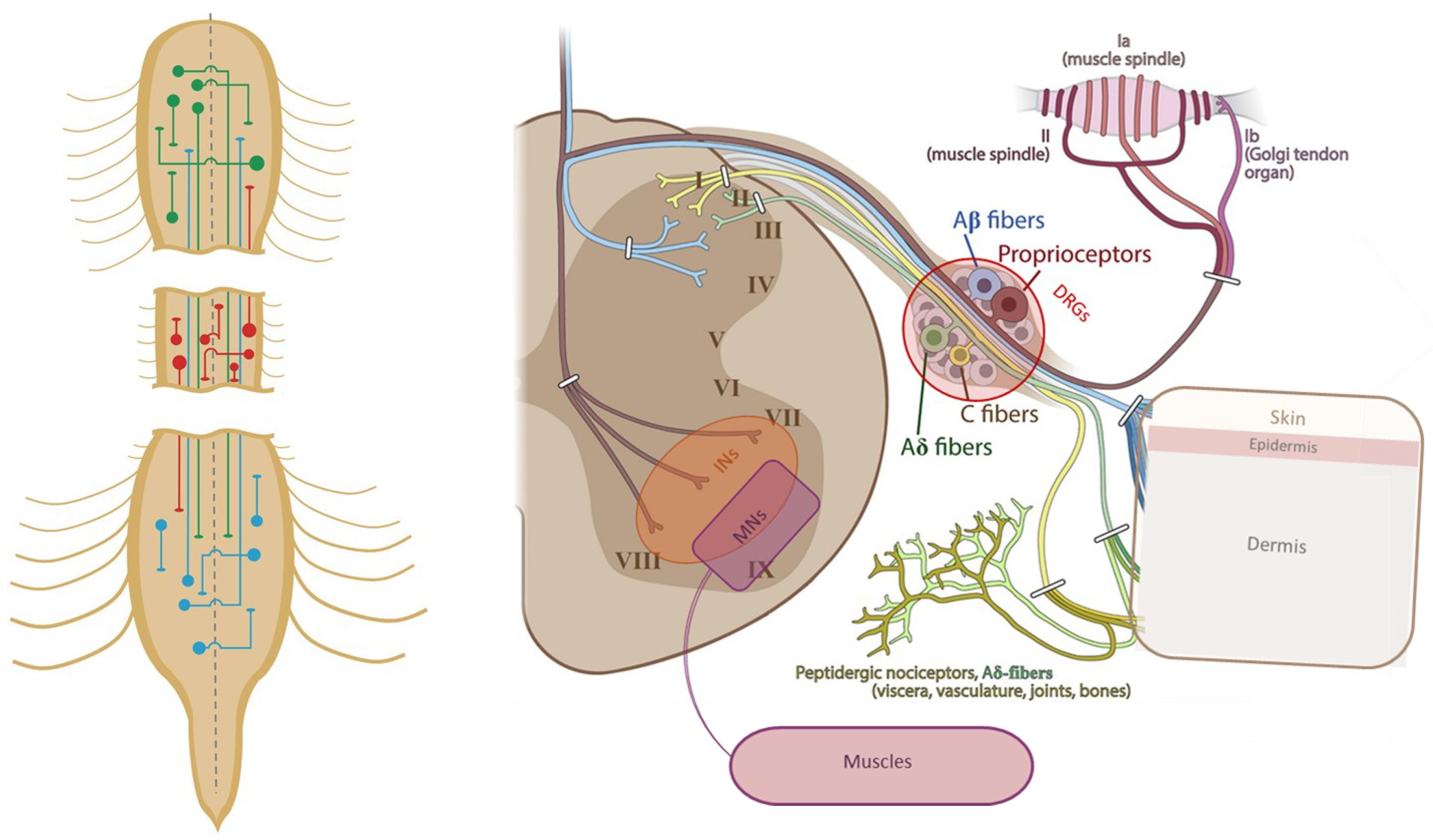
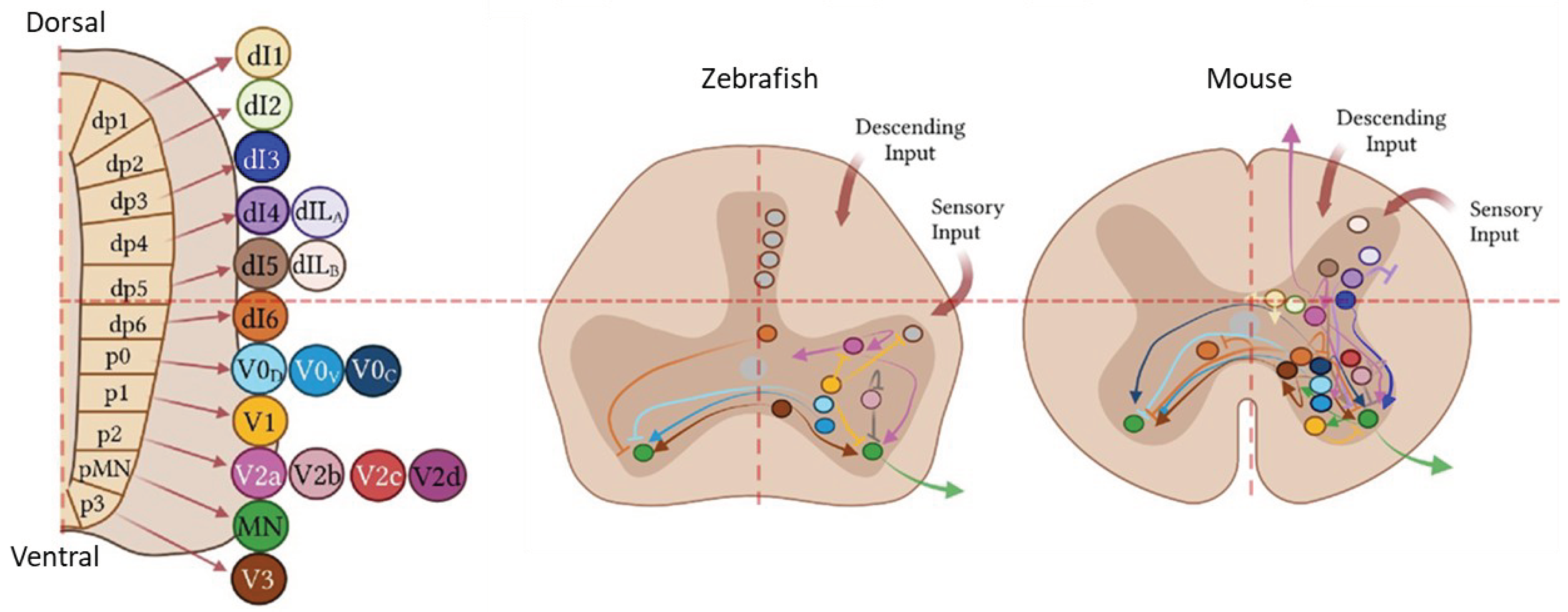
Figure 3.
Left: Eleven IN progenitor domains produce different IN populations (ventral ones correspond to dI6, V0, V1, V2 and V3). Right: comparison between IN connectivity between zebrafish and mouse. Adapted from 49.
Figure 3.
Left: Eleven IN progenitor domains produce different IN populations (ventral ones correspond to dI6, V0, V1, V2 and V3). Right: comparison between IN connectivity between zebrafish and mouse. Adapted from 49.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



