
Submitted:
04 March 2024
Posted:
05 March 2024
You are already at the latest version
Abstract
Drosophila melanogaster serves as a important research model both for genetic investigations and for the study of metal toxicity, facilitating the elucidation of physiological mechanisms comparable to those of human organisms. In our research, we evaluated the toxicity effect resulting from exposure to various concentrations of lead and copper on the prolificity rates throughout the life cycle (egg-adult) of four genotypes of Drosophila melanogaster: wild-type (control), white, brown, and white-vestigial. During our study, the prolificity rates were examined across three repetitions under the impact of exposure to concentrations of 0.50, 1.00, 2.00, and 4.00 mM of copper (CuSO4) and lead (Pb(C2H3O2)2). Prolificity rates throughout the life cycle exhibited variations as a direct consequence of genetic factors, the concentration of exposure, and the specific type of metal, either copper or lead. The mutant white-vestigial genotype revealed an IC50 concentration for prolificity inhibition at lower doses of 2.00 mM for copper and 4.00 mM for lead, in contrast to the control genotype (wild-type), which exhibited an inhibition concentration rate >IC50 of 4.00 mM only in the case of copper. Our results concluded that the (i) dose influences the prolificity rate in a directly proportional manner, (ii) comparative analyses between copper and lead revealed that copper displayed toxicity across all genotypes within the concentration range of 0.50 mM to 2.00 mM. In contrast, lead exhibited toxicity within the concentration range of 1.00 mM to 4.00 mM, highlighting a (iii) more acute toxicity characteristic in the case of copper. Thus, the results of this research reflect the importance of using Drosophila melanogaster as a genetic model in the comparative study of the interaction between genetic factors and the toxicity of metals, offering significant insights into monitoring their impact and defining the maximum permissible doses on organisms.
Keywords:
Drosophila melanogaster
; genotype
; prolificity
; heavy metals
; toxicology
; lead
; copper
1. Introduction
Drosophila melanogaster is a standard model for the study of various diseases, being an important bioindicator for testing the influence of chemical factors [1,2], for the development of treatments [3,4], diagnosis and understanding of toxicity phenomena in the body [5,6,7,8,9], as well as in the study of some neurodegenerative diseases [10].
Drosophila melanogaster (2n=8), exhibits a wide array of mutant genotypes alongside with a short life cycle, providing a significant advantage in the comparative study of toxicology [5,11]. Studies under the influence of heavy metals in D. melanogaster have stimulated increased numbers of reactive oxygen species (ROS) [6,12,13,14], leading to vacuolation of cells, a phenomenon associated with programmed cell death [15,16,17,18], damage to genetic material [19] and eventually apoptosis [18,20]. Numerous studies have highlighted the possibility of using antioxidants to reduce the toxic effects of heavy metals in Drosophila melanogaster [16,21,22]. Being thus an excellent bioindicator, the assay provides valuable information for the development of therapeutic strategies and personalized treatments for various diseases such as Parkinson’s [10,23,24,25,26], Alzheimer [27,28,29], cancer [30,31,32], kidney disease [33] and diabetes [34,35,36], being a model for investigating insulin action [37]. In terms of the action of metals, D. melanogaster may be an essential indicator for determining and understanding how they act on organisms beyond normal limits [6,38,39,40]. D. melanogaster is not only used for understanding the physiological aspects of essential metals [41,42,43], but also for assessing the toxic impact of heavy metals [44,45] having numerous genes and detoxification mechanisms in its composition similar to those found in humans [46,47,48,49].
Metals with metabolic implications, such as copper, although indispensable for the growth and development of organisms by participating in enzymatic reactions and maintaining homeostasis in the body [50,51], become toxic if they accumulate in high concentrations [52,53].
Copper intake in the body is provided by copper-rich foods such as vegetables, fruit [54,55,56], in meat products [57] or in cereals [1,2]. Drinking water also contributes about 0.12-0.26 mg (≈20%) [1] of copper to the average daily intake. The maximum concentration of copper contamination in drinking water is about 1.3 mg. The natural concentration of copper in soil is about 50 mg/kg. The atmospheric copper content also varies between 5 and 20 ng Cu/m3 [61].
Copper is found in nature in combinations, mainly as sulphides [62], participating in numerous biochemical processes based on oxidation-reduction reactions [63], via the various enzymes in whose composition it enters. In normal concentrations it has a positive impact on the functioning of the nervous and immune systems. In the nervous system, it facilitates the transmission of signals and the maintenance of neuronal integrity and in the immune system it contributes to the activation of defence cells and the protection of the body against pathogens.
The link between copper and these systems highlights its importance in maintaining health, with recommended doses of 1.5-3.0 mg in adults and 1 mg in children [52]. High concentrations of copper [64] act as an enzyme inhibitor and limit the activity of alkaline phosphatase [65], catalase [22], xanthine oxidase [66] and ribonuclease [67,68].
Heavy metals, such as lead, are toxic to the body [69,70,71], affecting a number of physiological and biochemical processes, such as metabolism, functioning of internal organs, and can contribute to diseases [5] and in plants affect [6,19,72], nutrition, photosynthesis [73], respiration [74], growth and development their [77,78].
Accumulation of heavy metals [78] leads to altered nutrient absorption capacity, deregulation of metabolic functions and eventually even death of organisms in case of intoxication [72,79,80]. In the context of heavy metal, pollution and climate change have a negative impact on the quality of life [81].
The maximum concentration limits for lead according to the EU Food Regulation are 0.1 mg/kg in meat products, 0.2 mg/kg in cereals and vegetables, 0.1 mg/kg and 0.02 mg/kg in milk. The normal limits for total lead in the human body in a person weighing 70 kg is on average 120 mg lead, with 0.2 mg/L in blood and between 0.2 and 3 in tissues. The U.S. Centers for Disease Control and Prevention has established standard elevated blood lead levels for adults and children (10 μg/dL and 5 μg/dL) [82].
Lead poisoning in concentrations between 0.2 mM-5 mM can manifest acute or chronic forms, and clinical symptoms usually become evident after 2-3 days [83].
Looking at the impact of lead on organisms, we see that exposure to lead can have significant health consequences. Studies show that lead can affect various organs and systems, including the nervous, cardiovascular and reproductive systems [16,84].
In the present study we aimed to monitor the effect of heavy metals on the life cycle of several mutant genotypes and the standard genotype, with a view to using them as bioindicators of comparative toxicity between copper and lead, as well as identifying toxicity limits. Drosophila melanogaster offers the possibility of establishing toxicity limits between heavy metals, their combinations, and interactions between them, which recommends it as a toxicological bioindicator for assessing impacts on ecosystems and organisms.
2. Materials and Methods
2.1. Description of biological Material
In the toxic effects research, we opted to use four genotypes of Drosophila melanogaster, including both wild and mutant variants.
The wild type genotype was used as a control variant, having a light grey body, scarlet (dull red) eyes and wings longer than the body (Figure 1a) [85].
The white mutant genotype is different from the wild type due to the presence of the recessive white gene which determines the white colour of the eyes. This gene is located on chromosome I (heterozygous X) at a distance of 1.5 centimorgans from the 0 end and is denoted by ,,w” (Figure 1b) [86].
The brown mutant genotype has as a defining feature the presence of the recessive bw gene, located on chromosome II at 104 cMo from the 0 end. This gene blocks the formation of the normal red pigment, resulting in a brown eye colour (Figure 1c) [87].
The white-vestigial mutant genotype is characterized by the white colour of the eyes, generated by the presence of the recessive gene w, located on chromosome I and the presence of reduced and rudimentary wings determined by the presence of the recessive gene ,,vg”, located on chromosome II (Figure 1d).
2.2. Preparation of culture medium and inoculation of D. melanogaster
The preparation of the culture medium consists of 200 ml of water to which 2 g of gelatine, 3.6 g of brewer’s yeast, 16.4 g of sugar and 20 g of grey are added.
The mixture obtained is boiled for 20 minutes and then 1 ml of propionic acid is added to the hot medium. The resulting composition is poured (30 ml) into culture dishes and covered with sterile stoppers until the culture medium solidifies. Subsequently, the medium is treated with a solution of yeast and distilled water, using a brush over which fine particles of dried yeast are sprinkled. Varying concentrations of lead acetate and copper sulphate were added to the prepared medium where adults of Drosophila melanogaster were inoculated. After etherization and examination, selected individuals were inoculated onto the culture media.
2.3. Growing environment and experimental metal varinates Cu2+ (CuSO4) and Pb2+ (Pb(C2H3O2)2)
Four concentrations of Cu2+ and Pb2+ were used in this study to determine the effect on lifecycle proliferation. Due to the varying toxicity limits between Cu2+ and Pb2+ according to the literature we used intermediate epidermal variants to determine the proliferation inhibition concentration (>IC50) in the four genotypes of D. melanogaster studied (wild, white, brown and white-vestigial) and to compare the toxicity levels between the two metals.
The experimental variants used were:
- -
- normal environment with grey (control) [81];
- -
- medium with grey, supplemented with copper sulphate solution CuSO4 -5H2O (0.50 mM, 1.00 mM, 2.00 mM and 4.00 mM);
- -
- medium with grey, supplemented with Pb(C2H3O2)2.·3H2O lead acetate solution (0.50 mM, 1.00 mM, 2.00 mM and 4.00 mM);
Determination of the inhibition index (CI50) of proliferation following exposure to Cu2+ (CuSO4) and Pb2+ (Pb(C2H3O2)2)
After pregrafting the experimental variants, Drosophila melanogaster adults were inoculated in a 1:1 ratio (♀ : ♂) for the wild-type and mutant genotypes: white, brown and white-vestigial.
The experiment was carried out in three replicates during the life cycle (egg-adult, Figure 2) ,during which the proliferation of different genotypes of Drosophila melanogaster was monitored under normal conditions and exposed to Cu2+ and Pb2+ concentrations of 0.50 mM, 1.00 mM, 2.00 mM and 4.00 mM.
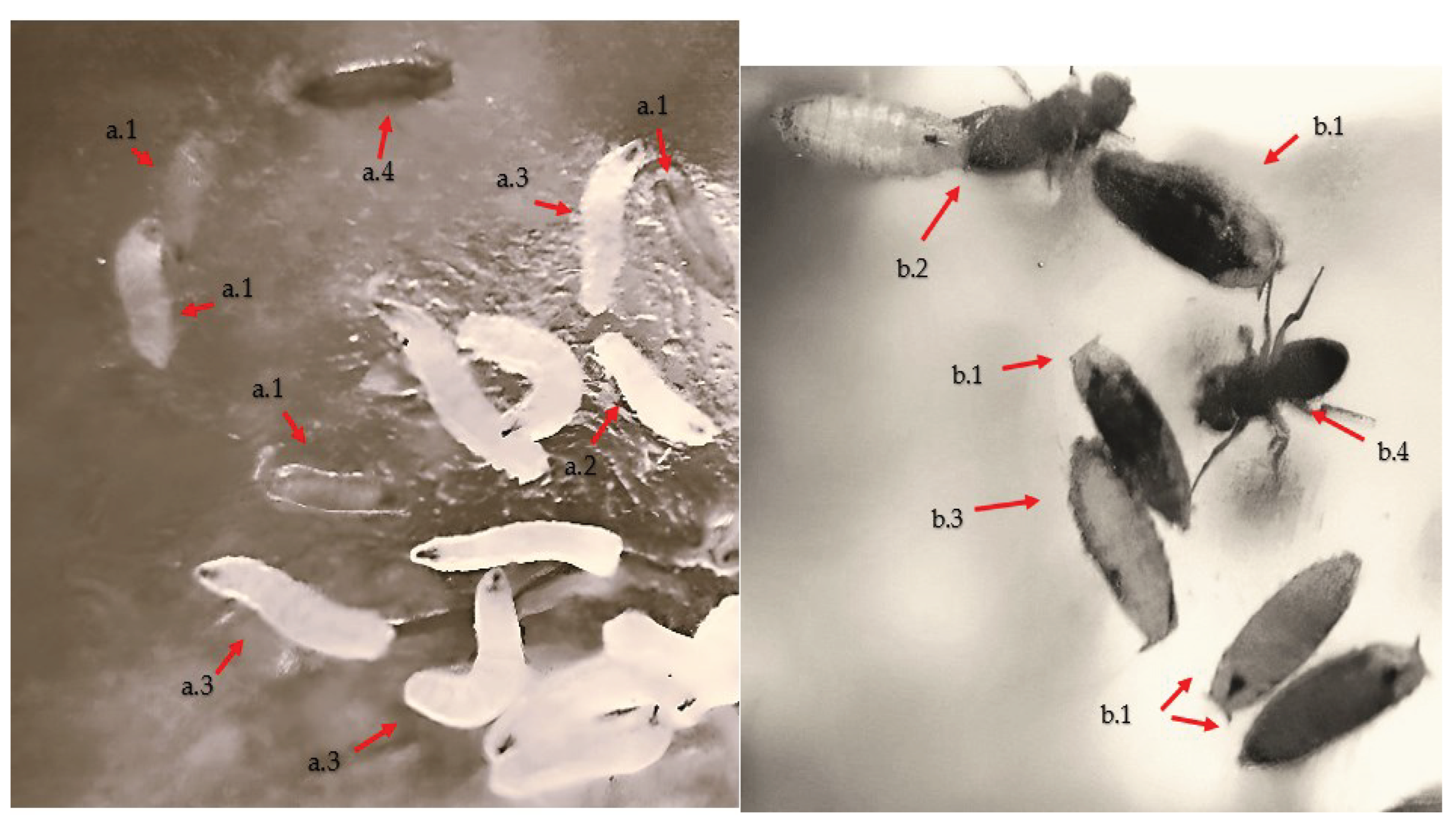
In the research, the impact of Cu2+ and Pb2+ was monitored at all developmental stages of Drosophila melanogaster (egg, larvae I, II, III, pupa and adult) (Figure 1) in order to determine the prolificacy as well as their inhibition concentrations.
After the establishment of the proliferation indices under normal conditions, they were compared with the experimental variants with different concentrations of Cu2+ and Pb2+ to determine the proliferation inhibition index (IC) according to formula (1):
The proliferation inhibition index >IC50 was considered the maximum toxicity level of the metal taken in the study [89,90].
Figure 2.
Developmental stages of Drosophila melanogaster larvae, (a.1)- larva stage I, (a.2)- larva stage II, (a.3)- larva stage III, (a.4)- larva pupation; Evolution of pupal to adult formation: (b.1)- pupal, (b.2)- hatching, (b.3)- hatched pupal, (b.4)- adult;.
Figure 2.
Developmental stages of Drosophila melanogaster larvae, (a.1)- larva stage I, (a.2)- larva stage II, (a.3)- larva stage III, (a.4)- larva pupation; Evolution of pupal to adult formation: (b.1)- pupal, (b.2)- hatching, (b.3)- hatched pupal, (b.4)- adult;.
2.4. Statistical calculation methods used to determine proliferation and toxicity levels between Cu2+ and Pb2+
In this study, we investigated the behavior of the genotypes concerning adaptability and prolificacy under exposure to various concentrations of heavy metals. We conducted statistical analysis using a multivariate ANOVA test, which uncovered the effects of genotype, metal type (Cu2+ or Pb2+), toxicity levels, and concentration on the developmental period from egg to adult. Significance was determined at a confidence level of p <0.05. The obtained results underwent comparative analysis using the Tukey test. The statistical interpretation was carried out using Rstudio software.
3. Results
This study determined the prolificacy of D. melanogaster genotypes following the application of different concentrations of copper and lead at different life cycle stages.
The results showed differences in the evolution of the number of larvae (I, II and III), as well as in the formation of pupae and adults, depending on the genotype, the type of metal used (Cu2+ or Pb2+) and the concentration used. Using the D. melanogaster genetic model, research has identified the influence of genotype and metal concentrations used in determining toxicity levels.
3.1. Influence of genotype, concentration and type of metal Cu2+ and Pb2+ on proliferation in Drosophila melanogaster
In the study of the influence of different wild-type genotypes (control) and variations between brown, white and white-vestigial mutant genotypes, a significant difference in the number of larvae formed in stages I, II and III under different experimental conditions was observed.
The results show a directly proportional correlation between the concentrations of heavy metals used and their type (Cu2+ and Pb2+), as well as the nature of the genotype, in terms of the number of larvae formed (p < 0.001).
There was also a significant influence of genotype on the number of individuals resulting from exposure to these metals.
Thus, from the results obtained, it can be deduced that the type of genotype studied can exert a significantly different influence on the number of individuals resulting from exposure to Cu2+ and Pb2+, thus confirming the significant role of genetic predisposition to toxicity (Table 1).
As in the larval stage, the influence of genotype and metal concentrations used was found to show significant variation in prolificacy levels throughout the life cycle, including in pupation and adult formation (p < 0.001) (Table 2).
Analysis of the interaction between the genetic factor and metal concentration levels (Cu2+ and Pb2+ ) significantly influenced proliferation in D. melanogaster throughout the life cycle, both in the larval stages (Table 1) and in the pupal and adult formation process (Table 2) (p < 0.001). Thus, as concentration increases, proliferation capacity specifically affects certain genotypes, with some being more tolerant (showing a lower inhibition concentration) and others more sensitive.
Looking at the interaction between concentration and metal type (Cu2+ or Pb2+), it had a significant impact on the number of individuals during the developmental cycle (p < 0.001). This reflects differences in the response of proliferation rates depending on the type of metal to which D. melanogaster genotypes are exposed.
The results obtained show the same impact of decreased prolificacy for both Cu2+, and Pb2+, obtaining the same effect of decreased number of inidivids in larval stages of pupal formation (Genotype*Metal, Concentration*Genotype: Metal, p >0.05). Toxicity levels (degree of inhibition of proliferation, >IC50) are influenced by genotype, concentrations and type of metal (Cu2+ or Pb2+) used in the medium (Concentration, Genotype, Metal, Concentration*Genotype, Concentration*Metal, p <0.001) (Table 1 and Table 2).

Table 2.
Analysis of factors involved in the process of inhibition of proliferation at different concentrations of copper (CuSO4) and lead (Pb(C2H3O2)2 in pupal and adult formation.
Table 2.
Analysis of factors involved in the process of inhibition of proliferation at different concentrations of copper (CuSO4) and lead (Pb(C2H3O2)2 in pupal and adult formation.
| Analysis factor | df | SS | S2 | F | Pr(>F) | |
|---|---|---|---|---|---|---|
| Pupal stage | Concentration | 4 | 24437 | 6109 | 203.090 | < 2e-16 *** |
| Genotype | 3 | 5526 | 1842 | 61.234 | < 2e-16 *** | |
| Metal | 1 | 1687 | 1687 | 56.094 | 1.45e-13 *** | |
| Concentration: Genotype | 12 | 1132 | 94 | 3.136 | 0.000211 *** | |
| Concentration:Metal | 4 | 585 | 146 | 4.861 | 0.000691 *** | |
| Genotype:Metal | 3 | 80 | 27 | 0.884 | 0.448785 Ns | |
| Concentration: Genotype: Metal | 12 | 179 | 15 | 0.496 | 0.918151 Ns | |
| Residuals | 1060 | 31886 | 30 | |||
| Adult stage | Concentration | 4 | 12209 | 3052.3 | 135.964 | < 2e-16 *** |
| Genotype | 3 | 4569 | 1522.9 | 67.837 | < 2e-16 *** | |
| Metal | 1 | 893 | 893.5 | 39.799 | 5.55e-10 *** | |
| Concentration: Genotype | 12 | 551 | 45.9 | 2.044 | 0.0189 * | |
| Concentration:Metal | 4 | 274 | 68.5 | 3.051 | 0.0166 * | |
| Genotype:Metal | 3 | 65 | 21.7 | 0.968 | 0.4075 Ns | |
| Concentration: Genotype: Metal | 12 | 89 | 7.4 | 0.331 | 0.9836 Ns | |
| Residuals | 586 | 13155 | 22.4 |
Note: SS- sum of square, dF-degrees of freedom; S2 -mean square. “Ns”-not significant, “*”- p < 0.05, “**”- p < 0.01, ***significant at p < 0.001.
3.2. Influence of copper (CuSO4) on proliferation in Drosophila melanogaster during the life cycle (egg-adult)
Investigations of the prolificacy rate of Drosophila melanogaster genotypes at various copper concentrations show a decrease directly proportional to the increase in copper concentration, both in the three larval developmental stages (stage I, II and III) and in the pupal and adult formation stages (Table 3).
Significant differences were observed in the response to copper concentrations between the genotypes used (p < 0.05). Brown and white-vestigial genotypes showed higher sensitivity to the highest Cu+2 concentrations compared to wild and white genotypes.
The wild genotype showed prolificacy rate values below 50% (>IC50 ) at copper concentrations of 4.00 mM in larval stages I, II and III. Close values of proliferation inhibition >IC50 were observed at 2.00 mM concentration of Cu+2. The same toxicity effect was observed in pupal and adult formation with >IC50 values being present at 4.00 mM concentration (Table 3).
Table 3.
Results on the proliferation of D. melanogaster genotypes following exposure to various concentrations of copper.
Table 3.
Results on the proliferation of D. melanogaster genotypes following exposure to various concentrations of copper.
| Genotype | Concentration (mM) | Copper (CuSO4) | |||||||||
| StageI | IC | StageII | IC | StageIII | IC | Pupal | IC | Adult | IC | ||
| Mean | Mean | Mean | Mean | Mean | |||||||
| wild | Control | 35.97a | 0.00 | 28.33a | 0.00 | 26.39a | 0.00 | 25.41a | 0.00 | 24.87a | 0.00 |
| 0.50 | 29.85b | 17.03 | 24.58abc | 13.24 | 22.24ab | 15.73 | 20.67ab | 18.66 | 20.27abc | 18.50 | |
| 1.00 | 25.41bcd | 29.37 | 19.58cdef | 30.88 | 17.85bcde | 32.38 | 15.67cde | 38.34 | 16.40cdef | 34.05 | |
| 2.00 | 21.56de | 40.06 | 16.58defgh | 41.47 | 15.03defg | 43.05 | 13.33def | 47.52 | 14.20def | 42.90 | |
| 4.00 | 13.23fgh | 63.22 | 10.14ijk | 64.22 | 9.24hijk | 64.98 | 7.90ghi | 68.92 | 8.39gh | 66.26 | |
| brown | Control | 27.08bc | 0.00 | 20.83cde | 0.00 | 19.52bcd | 0.00 | 18.74bc | 0.00 | 17.73bcd | 0.00 |
| 0.50 | 22.28cde | 17.71 | 17.83defgh | 14.40 | 16.39cdef | 15.99 | 14.96cdef | 20.16 | 14.47def | 18.42 | |
| 1.00 | 19.97de | 26.23 | 16.03efgh | 23.07 | 14.88efg | 23.76 | 13.04def | 30.43 | 12.73defg | 28.20 | |
| 2.00 | 16.85efg | 37.78 | 12.56hij | 39.73 | 11.67ghij | 40.22 | 10.22fgh | 45.45 | 10.80fgh | 39.10 | |
| 4.00 | 10.82hi | 60.04 | 7.03kl | 66.27 | 6.48kl | 66.77 | 6.00hi | 67.98 | 6.94hi | 60.84 | |
| white | Control | 30.54b | 0.00 | 26.64ab | 0.00 | 26.18a | 0.00 | 24.48a | 0.00 | 22.31ab | 0.00 |
| 0.50 | 28.00bc | 8.31 | 21.47bcd | 19.40 | 19.82bc | 24.31 | 17.26bcd | 29.50 | 18.33bcd | 17.83 | |
| 1.00 | 22.33cde | 26.87 | 17.14defgh | 35.66 | 16.30cdef | 37.73 | 14.07cdef | 42.51 | 14.27def | 36.06 | |
| 2.00 | 17.85ef | 41.56 | 13.53fghi | 49.22 | 12.30fghi | 53.01 | 10.85efg | 55.67 | 11.67efgh | 47.71 | |
| 4.00 | 10.46hi | 65.74 | 8.33ghij | 68.72 | 7.18jkl | 72.57 | 7.30ghi | 70.18 | 6.56hi | 70.62 | |
| white-vestigial | Control | 23.10cd | 0.00 | 21.85bcd | 0.00 | 19.70bc | 0.00 | 18.56bc | 0.00 | 16.93bcde | 0.00 |
| 0.50 | 19.79de | 14.32 | 18.42defg | 15.72 | 15.94cdefg | 19.08 | 14.63cdef | 21.16 | 13.87defg | 18.11 | |
| 1.00 | 17.05efg | 26.19 | 15.19fghi | 30.47 | 13.27efgh | 32.62 | 12.74def | 31.34 | 11.33efgh | 33.07 | |
| 2.00 | 11.51ghi | 50.17 | 9.03jkl | 58.69 | 8.45ijkl | 57.08 | 7.22ghi | 61.08 | 6.27hi | 62.99 | |
| 4.00 | 5.97i | 74.14 | 4.53l | 79.27 | 4.58l | 76.77 | 3.64i | 80.37 | 2.94i | 82.63 | |
*Superscript letters are assigned based on the results of the Tukey pairwise comparison test, indicating the significance levels of each column value obtained relative to the copper concentration and exposure time. The highest-ranked values are denoted, beginning with the first letter “a”. Similar superscript letters indicate statistically non-significant differences between the obtained values (p >0.05).**IC denotes the prolificity inhibitaion concentration of copper.
For the brown genotype, at the concentration of 4.00 mM Cu+2, the results indicate a decrease in proliferation for larval stages I, II and III (>IC50), highlighting a manifestation of increased toxicity in this genotype (p <0.05). A significant decrease in the number of pupae and adults compared to the control variant is also noted, indicating the negative impact of copper on their formation (p <0.05).
In the case of the white genotype, the concentration of 4.00 mM (>IC50) of copper shows the same toxic effect, causing a reduction in the proliferation of this genotype during the life cycle.
The highest sensitivity was recorded in the white-vestigial genotype in which the 2.00 mM Cu+2 concentration (>IC50) showed a decrease in proliferation of more than 50% compared to the control group for all developmental stages (p <0.05).
According to the results obtained (Table 3 and Figure 3), the proliferation rate of the genotypes was inhibited by the presence of copper in all larval stages, showing the significant reduction of the proliferation rate starting with the dose of 0.50 mM Cu+2(p <0.05).
In the early stages of the developmental cycle, concentrations of 0.50-1.00 mM Cu+2 resulted in decreases in the proliferation rate, which increased as the concentration of Cu+2 increased.
Pup and adult formation of the genotypes studied was influenced directly proportional to increasing concentrations for all stages followed (Table 3 and Figure 4).
Thus, from the presented results it was evident that the value of >IC50 at the concentration of 4.00 mM is present in all studied genotypes (4/4). At 2.00 mM concentration of Cu+2 the white-vestigial genotype showed the >IC50 value throughout the development cycle (1/4).
The results obtained in this study highlight the impact of copper concentrations (CuSO4) on the proliferation of Drosophila melanogaster during their life cycle, showing an inhibition with increasing copper concentration and genotype under study.
3.3. Influence of lead (Pb(C2H3O2)2 on proliferation in Drosophila melanogaster during the life cycle (egg-adult)
Analysis of the number of stage I, II and III larvae of Drosophila melanogaster genotypes exposed to varying concentrations of Pb2+ revealed significant differences in the rate of proliferation as an effect of toxicity. The results showed significant variation in the number of individuals developed under different experimental conditions.
Investigations found that genotype had a significant influence on development in D. melanogaster following exposure, confirming a higher genetic predisposition to toxicity for certain genotypes. The highest values of prolificacy were recorded for the wild and white genotypes, the average values for the brown genotype and the lowest values for the white-western genotype (Table 4). The >IC50 value at all developmental stages is present only in the white-vestigial genotype at a concentration of 4.00 mM (1/4, of genotypes).
In the early stages of the developmental cycle, larval stages exposed to concentrations ranging from 0.50 to 1.00 mM of lead exhibited a decrease in prolificacy. However, this decrease was not statistically significant concerning genotype prolificacy (p >0.05).
The wild genotype showed a significant reduction in the number of larvae in stages I. II and III at higher concentrations, especially at 4.00 mM. The same trend was observed for the white and brown genotypes, suggesting a common sensitivity to lead.
As for the white-vestigial genotype, a more pronounced sensitivity to lead was observed. with a significant reduction in larval numbers in stages I, II and III starting at 1.00 mM concentration. in contrast to the other genotypes which showed a higher tolerance at this concentration. The results suggest that genotypes show different responses to lead concentrations. influencing development in D. melanogaster to varying degrees.
Figure 5.
Effect of lead on different genotypes of Drosophila melanogaster in adult formation: (a)- wild genotype. (b)- brown genotype, (c)- white mutant genotype, (d)- white-vestigial mutant genotype.
Figure 5.
Effect of lead on different genotypes of Drosophila melanogaster in adult formation: (a)- wild genotype. (b)- brown genotype, (c)- white mutant genotype, (d)- white-vestigial mutant genotype.
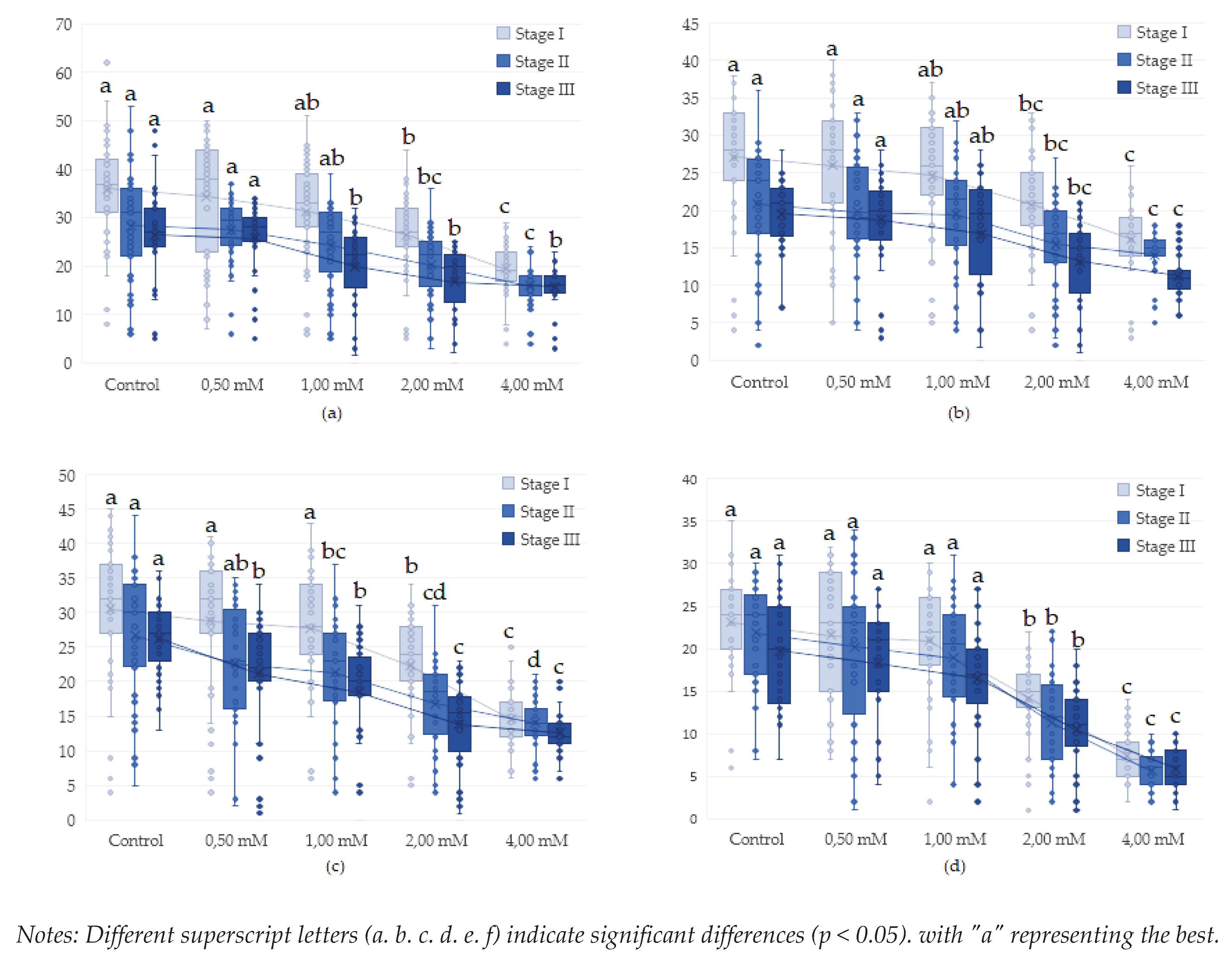
Figure 6.
Effect of lead on different genotypes of Drosophila melanogaster in adult formation: (a)- wild wild genotype (b)- brown genotype (c)- white mutant genotype (d)- white-vestigial mutant genotype.
Figure 6.
Effect of lead on different genotypes of Drosophila melanogaster in adult formation: (a)- wild wild genotype (b)- brown genotype (c)- white mutant genotype (d)- white-vestigial mutant genotype.
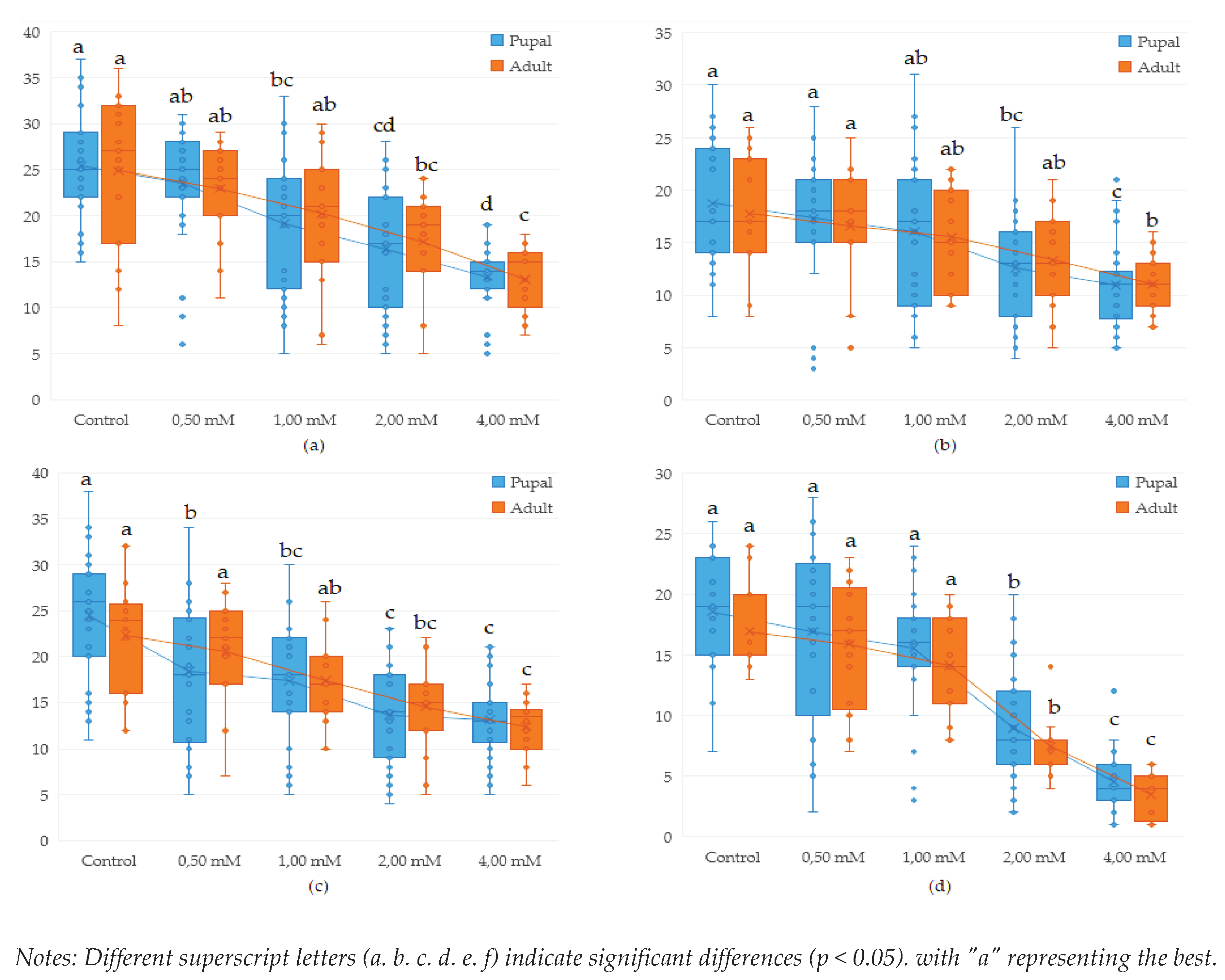
Analysing the influence of Drosophila melanogaster genotypes on the number of pupae and adults obtained we found that the diversity of genotypes had a significant impact on the results. while also influencing the number of pupae in the control samples (p <0.001). In the case of lead exposure, the results obtained indicate a significant link between lead concentration and the effects generated.
Studies on mutant genotypes revealed that the white-vestigial genotype showed significantly fewer pupae formed in most of the variants studied, compared to the wild wild genotype which showed the highest tolerance at the pupal and adult stages.
Analysis of the effect of lead in Drosophila melanogaster reveals the significant impact of this metal on the development and health of organisms highlighting the importance of further research in this field and the development of effective strategies for environmental and public health protection.
Comparative study of toxicity effects on drosophila melanogaster at various concentrations of lead and copper
The comparative study of copper and lead toxicity revealed significant variations in the formation of pupae and the emergence of adult flies. The analysis of toxicity differences demonstrated that copper exhibited higher toxicity at both low and high concentrations compared to lead. In the context of adult development, it was observed that at a concentration of 0.50 mM, no significant differences between the two metals were identified (p < 0.05).
Figure 7.
Effect of copper on various genotypes of Drosophila melanogaster in adult development.
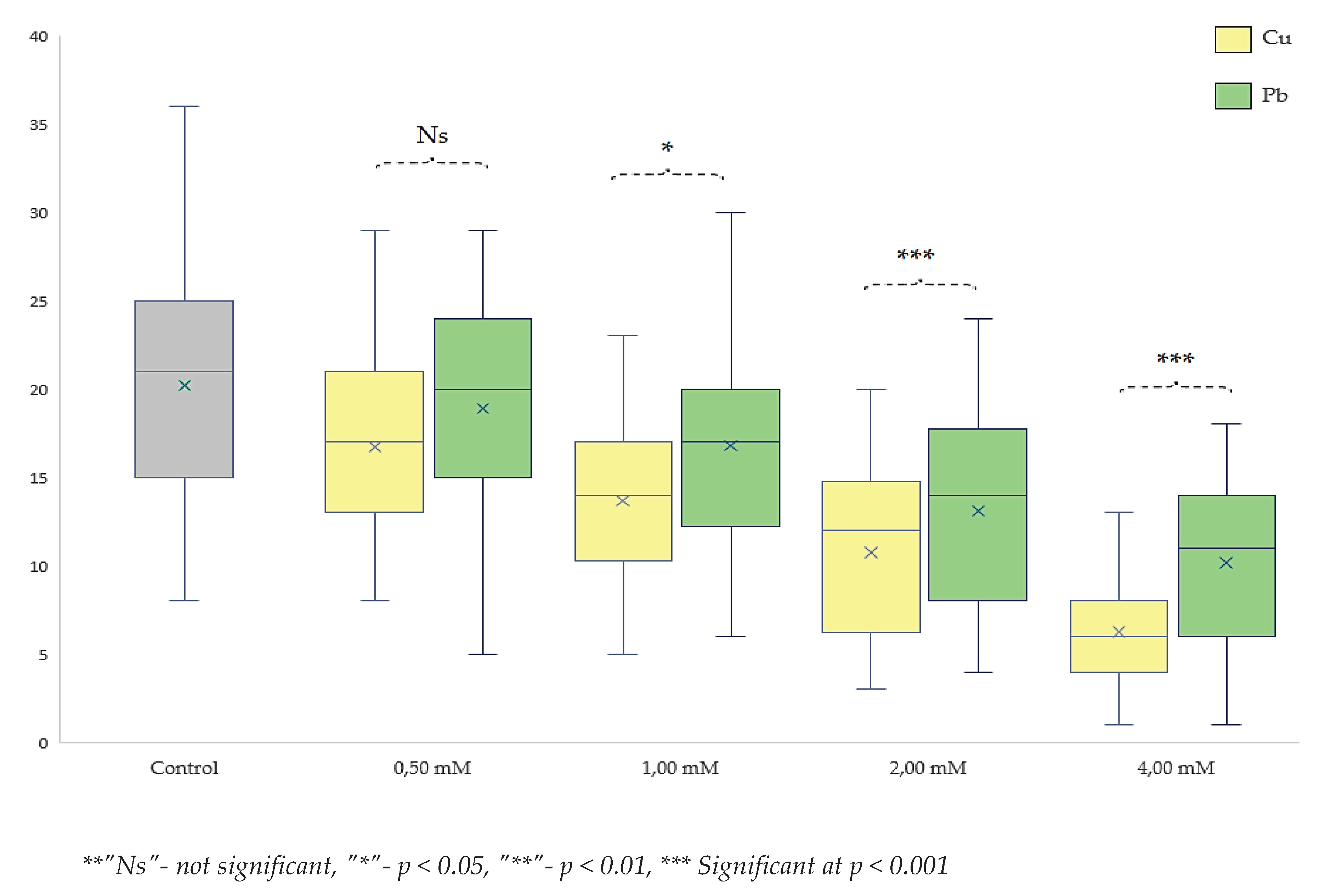
These differences persisted across other concentrations, indicating a continuous dominant effect of copper toxicity over that of lead. Consequently, the level of significance increased with rising concentrations. Therefore, at a concentration of 1.00 mM, the toxicity degree was significantly more pronounced for copper with p < 0.05 compared to lead. At concentrations of 2.00 mM and 4.00 mM, the degree of significance reached p < 0.001. The intervals were significantly different from each other (Figure 8) and also compared to the control group. These findings demonstrate a more acute toxic effect of copper compared to lead. This outcome might be inherently understandable considering that copper (CuSO4) is often used as an antimicrobial and antibacterial agent, playing a crucial regulatory role in homeostasis, whereas the accumulation of lead exhibits a significantly different toxicological profile.
The study on D. melanogaster can provide valuable insights into both types of toxicity, considering its rapid life cycle and the ease with which multiple generations can be generated and observed.
4. Discussion
Following literature reviews the results obtained complete the explanation of the Cu2+ toxicity phenomenon. (i) The same toxic effect has been observed by other authors using wild type genotype at concentrations of 0.50 mM [61,62,63], the most damaging concentration of 3.00 mM of Cu2+, values that justify the toxicity values obtained by us (2 mM-4 mM, Table 3). (ii) In addition to the results we can say that there is a significant influence of the genotype used, as the authors also indicate an inhibition of proliferation on Cu2+ depending on the sex [93,94] or the developmental stage [2], the number of individuals starting to decrease significantly depending on the dose applied 0.40 mM for females and 0.75 mM for males [93]. (iii) Research shows the possibility of development of genotypes with increased tolerance to Cu2+ concentrations up to 50 mM of more than 75% as opposed to other genotypes with survival rates below 18%. As a result. genotype influence plays a determined role in the assessment of metalor toxicity above normal doses.
Copper is an essential element for the proper functioning of all organisms having an essential role in the composition of enzymes [95,96], antimicrobial and antiviral against Escherichia coli [97,98], Staphylococcus aureus (MRSA) [99,100,101], SARS-coronavirus [102,103,104], influencing virus A [104,105,106] and fungi [107]. However, even though the effects of copper are beneficial at high concentrations the results obtained for D. melanogaster indicate a negative effect of Cu2+, manifested by reduced numbers of larvae. pupae and adults at concentrations starting at 0.50 mM Cu2+.
Research has shown that the toxicity causes a reduction in the number of individuals and the impact is significant in all genotypes at the highest concentrations used in the experiment of 2.00-4.00 mM. Our results indicate different degrees of toxicity depending on the genotype of D. melanogaster used, which was observed for both Cu2+, and Pb2+ as well as results obtained by other authors [20].
As regards lead accumulation research has shown the appearance of oxidative stress [109] at concentrations of 0.20 mM Pb 2+ [16] from the larval stage and even the appearance of spots on the wings. showing a mutagenic effect of Pb2+ from concentrations of 0.40 mM [110].
Cumulative cytotoxic effects of Pb2+ and Cu2+, have also been shown in the organisms Ceriodaphnia dubia and Daphnia carinata [111].
Studies by other researchers on the impact of heavy metals on Emiliania huxleyi indicate a stimulation of copper uptake in the presence of lead at low concentrations [111]. However, at higher lead levelsm a reverse effect was observed. characterized by a significant reduction in copper uptake [112].
Influence on the reduction of absorption has also been identified for iron (Fe) indicating a decrease in Fe absorption under the influence of Pb2+, at a concentration of 342 µg Pb/mL [109].
These results were also obtained in mouse experiments, which indicated that DMT1, responsible for iron (Fe) uptake, facilitates lead transport across the blood-brain barrier to the brain [113] (2.00 mM Pb2+ ). This explains the toxic effect of lead on the nervous system with long-term exposure, with an even more pronounced impact on children [84].
Due to the considerable number of generations undergone by Drosophila melanogaster, this species offers a precise opportunity for localizing Quantitative Trait Loci (QTL), not only concerning genetic effects and biological impacts in heavy metal resistance, but also for analyzing the toxic effects of various chemicals and treatments. Consequently, by furnishing substantial and efficient biological support, Drosophila melanogaster emerges as a pivotal model and valuable indicator in health analysis and maintenance. This contributes significantly to the identification, monitoring, and development of solutions for various diseases, including cases of heavy metal poisoning [26].
5. Conclusions
This study unveiled the influence of genetic factors on the proliferation capacity of Drosophila melanogaster. The most sensitive genotype, white-vestigial, exhibited lower prolificacy compared to wild genotypes or other mutant variations. Prolificity varied across both control and experimental copper and lead variants throughout the life cycle.
Metals toxicity was elucidated through the proliferation inhibition index, which fluctuated according to genotype and concentration, with proliferation inhibition directly correlating with increasing concentration.
Copper displayed toxicity across all genotypes within the 0.50 mM to 2.00 mM concentration range (>IC50 valid for all genotypes), emphasizing the necessity of regulating and monitoring copper levels in the environment. Lead exhibited toxicity within the concentration range of 1.00 mM to 4.00 mM, with an inhibitory concentration exceeding IC50 solely for the white-vestigial genotype.
The comparative analysis between copper and lead highlighted that the adverse effects of copper are more enduring and acute than those of lead, suggesting the requisite for appropriate management of heavy metal exposure in natural environments. Differences in proliferation toxicity in D. melanogaster between copper and lead are substantial, escalating with increasing concentrations.
These findings underscore the significance of further research in ecotoxicology and the formulation of effective strategies for environmental protection and public health concerning heavy metal pollution, utilizing the D. melanogaster genetic model.
In conclusion, we aim to replicate the experiments with other metals and develop genotypes resistant to the most influential health-affecting factors. These research directions hold the potential to offer significant insights and practical solutions in addressing complex health issues, drawing upon the contributions of Drosophila melanogaster in the experiments.
Author Contributions
Conceptualization, Petrescu Irina; Data curation, Onisan Emilian, Tenche-Constantinescu Alina and Ganea Mariana; Formal analysis, Onisan Emilian; Funding acquisition, Morna Anamaria, Camen Droin and Ciulca Adriana; Investigation, Sarac Ioan, Petrescu Irina, Onisan Emilian, Morna Anamaria, Timar Adrian Vasile, Vicaș Laura Grațiela and Ganea Mariana; Methodology, Sarac Ioan, Petrescu Irina and Ganea Mariana; Project administration, Camen Droin; Resources, Sarac Ioan, Timar Adrian Vasile, Petolescu Cerasela, Ciulca Adriana and Ganea Mariana; Validation, Sarac Ioan, Petrescu Irina, Tenche-Constantinescu Alina, Vicaș Laura Grațiela and Stroia Ciprian; Visualization, Tenche-Constantinescu Alina, Timar Adrian Vasile, Camen Droin, Petolescu Cerasela and Ganea Mariana; Writing – original draft, Sarac Ioan, Onisan Emilian and Stroia Ciprian; Writing – review & editing, Sarac Ioan, Onisan Emilian and Stroia Ciprian.
Funding
This research paper was supported by the project “Increasing the impact of excellence research on the capacity for innovation and technology transfer within USAMVB Timișoara”. code 6PFE. submitted to the competition Program 1-Development of the national system of research-development. Subprogram 1.2-Institutional performance. Institutional development projects-Development projects of excellence in RDI.
Institutional Review Board Statement
The research conducted in this article did not involve animals or humans.
Data Availability Statement
Data are contained within the article.
Acknowledgments
The authors would like to thank the funding institutions.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Santarelli et al. ‘Drosophila melanogaster as a model to study autophagy in neurodegenerative diseases induced by proteinopathies’. Front. Neurosci. vol. 17. p. 1082047. 2023. Accessed: Dec. 01. 2023. [Online]. Available: https://www.frontiersin.org/articles/10.3389/fnins.2023.1082047/full. [CrossRef]
- S. Bahadorani and A. J. Hilliker. ‘Biological and behavioral effects of heavy metals in Drosophila melanogaster adults and larvae’ J. Insect Behav. vol. 22. pp. 399-411. 2009. Accessed: Nov. 28. 2023. [Online]. Available: https://idp.springer.com/authorize/casa?redirect_uri=https://link.springer.com/article/10.1007/s10905-009-9181-4&casa_token=OVUFcjRGCKcAAAAA:9DkjNeV35QQ7C2M4GIWsE6dzITLw1AA536l5Rc32ReOEwAbtnpIRP2iKDcOI1xtFUOkz1LzARu9PBOFY. [CrossRef]
- C. V. Klimaczewski. A. Ecker. B. Piccoli. M. Aschner. N. V. Barbosa. and J. B. T. Rocha. ‘Peumus boldus attenuates copper-induced toxicity in Drosophila melanogaster’ Biomed. Pharmacother. vol. 97. pp. 1-8. Jan. 2018. [CrossRef]
- L. K. Venkareddy and Muralidhara. ‘Potential of casein as a nutrient intervention to alleviate lead (Pb) acetate-mediated oxidative stress and neurotoxicity: First evidence in Drosophila melanogaster’. NeuroToxicology vol. 48. pp. 142-151. May 2015. [CrossRef]
- S. Saini. L. Rani. N. Shukla. M. Banerjee. D. K. Chowdhuri. and N. K. Gautam. ‘Development of a Drosophila melanogaster based model for the assessment of cadmium and mercury mediated renal tubular toxicity’. Ecotoxicol. Environ. Saf. vol. 201. p. 110811. Sep. 2020. [CrossRef]
- F. Turna Demir. G. Akkoyunlu. and E. Demir. ‘Interactions of ingested polystyrene microplastics with heavy metals (Cadmium or Silver) as environmental pollutants: A comprehensive in vivo study using Drosophila melanogaster’. Biology vol. 11. no. 10. Art. no. 10. 2022. Accessed: Nov. 28. 2023. [Online]. Available: https://www.mdpi.com/2079-7737/11/10/1470. [CrossRef]
- Calap-Quintana. Pablo. Javier González-Fernández. Noelia Sebastiá-Ortega. José Vicente Llorens. and María Dolores Moltó. ‘Drosophila melanogaster Models of Metal-Related Human Diseases and Metal Toxicity’. Int. J. Mol. Sci. vol. 18. no. 7. Art. no. 7. 2017. [CrossRef]
- ‘IJMS | Free Full-Text | Drosophila melanogaster Models of Metal-Related Human Diseases and Metal Toxicity’. Accessed: Nov. 28. 2023. [Online]. Available: https://www.mdpi.com/1422-0067/18/7/1456. [CrossRef]
- E. GÜNEȘ and E. ȘENSOY. ‘The Effect of Turkısh Coffee and Cadmıum Acetate on Drosophila melanogaster’. J. Int. Environ. Appl. Sci. vol. 15. no. 1. Accessed: Nov. 28. 2023. [Online]. Available: https://dergipark.org.tr/en/pub/jieas/issue/53199/697665.
- L. Bonilla-Ramirez. M. Jimenez-Del-Rio. and C. Velez-Pardo. ‘Acute and chronic metal exposure impairs locomotion activity in Drosophila melanogaster: a model to study Parkinsonism’. Biometals vol. 24. pp. 1045-1057. 2011. Accessed: Jan. 16. 2024. [Online]. Available: https://idp.springer.com/authorize/casa?redirect_uri=https://link.springer.com/article/10.1007/s10534-011-9463-0&casa_token=tDZ6LaLxDuIAAAAA:G24wIqXMSR0Z1-tYE_ICt7bGN8h7A1w6PqX-CE7rWrzDRYe6Np_o9w4xCuWvBDgCWwI9uQIcaeQ6TCbS. [CrossRef]
- M. N. Arbeitman et al. ‘Gene Expression During the Life Cycle of Drosophila melanogaster’. Science vvol. 297. no. 5590. pp. 2270-2275. Sep. 2002. [CrossRef]
- M. Mishra and M. Panda. ‘Reactive oxygen species: the root cause of nanoparticle-induced toxicity in Drosophila melanogaster’. Free Radic. Res. vol. 55. no. 8. pp. 919-935. Aug. 2021. [CrossRef]
- M. Alaraby. A. Hernández. and R. Marcos. ‘Copper oxide nanoparticles and copper sulphate act as antigenotoxic agents in drosophila melanogaster’. Environ. Mol. Mutagen vol. 58. no. 1. pp. 46-55. 2017. [CrossRef]
- E. Baeg. K. Sooklert. and A. Sereemaspun. ‘Copper Oxide Nanoparticles Cause a Dose-Dependent Toxicity via Inducing Reactive Oxygen Species in Drosophila’. Nanomaterials vol. 8. no. 10. Art. no. 10. Oct. 2018. [CrossRef]
- et al. O. Emilian et al. ‘Cytological Applications of the Vacuolization Phenomenon as a Means of Determining Saline Cytotoxicity’. Appl. Sci. vol. 13. no. 14. p. 8461. 2023. Accessed: Dec. 01. 2023. [Online]. Available: https://www.mdpi.com/2076-3417/13/14/8461. [CrossRef]
- Shilpa. K. P. Anupama. A. Antony. and H. P. Gurushankara. ‘Lead (Pb) induced Oxidative Stress as a Mechanism to Cause Neurotoxicity in Drosophila melanogaster’. Toxicology vol. 462. p. 152959. Oct. 2021. [CrossRef]
- Y. Zheng. H. Zhang. X. Deng. J. Liu. and H. Chen. ‘The relationship between vacuolation and initiation of PCD in rice (Oryza sativa) aleurone cells’. Sci. Rep vol. 7. no. 1. Art. no. 1. Jan. 2017. [CrossRef]
- V. Shubin. I. V. Demidyuk. A. A. Komissarov. L. M. Rafieva. and S. V. Kostrov. ‘Cytoplasmic vacuolization in cell death and survival’. Oncotarget vol. 7. no. 34. pp. 55863-55889. Jun. 2016. [CrossRef]
- E. R. Carmona. A. Creus. and R. Marcos. ‘Genotoxicity testing of two lead-compounds in somatic cells of Drosophila melanogaster’. Mutat. Res. Toxicol. Environ. Mutagen. vol. 724. no. 1. Art. no. 1. Sep. 2011. [CrossRef]
- P. Yang et al. ‘Effects of cadmium on oxidative stress and cell apoptosis in Drosophila melanogaster larvae’. Sci. Rep. vol. 12. no. 1. Art. no. 1. 2022. Accessed: Nov. 28. 2023. [Online]. Available: https://www.nature.com/articles/s41598-022-08758-0. [CrossRef]
- G. J. T. Salazar. A. Ecker. S. A. Adefegha. and J. G. M. da Costa. ‘Advances in Evaluation of Antioxidant and Toxicological Properties of Stryphnodendron rotundifolium Mart. in Drosophila melanogaster Model’. Foods vol. 11. no. 15. Jan. 2022. [CrossRef]
- B. Skwarylo-Bednarz et al. ‘The impact of copper on catalase activity and antioxidant properties of soil under amaranth cultivation’. J. Elem. vol. 23. no. 3. 2018. Accessed: Dec. 01. 2023. [Online]. Available: https://bibliotekanauki.pl/articles/14416.pdf. [CrossRef]
- P. Duarte-Jurado et al. ‘Antioxidant Therapeutics in Parkinson’s Disease: Current Challenges and Opportunities’. Antioxidants vol. 10. no. 3. Art. no. 3. Mar. 2021. [CrossRef]
- V. Muñoz-Soriano and N. Paricio. ‘Drosophila Models of Parkinson’s Disease: Discovering Relevant Pathways and Novel Therapeutic Strategies’. Park. Dis vol. 2011. p. e520640. Mar. 2011. [CrossRef]
- J. Whitworth. ‘1 - Drosophila Models of Parkinson’s Disease’. in Advances in Genetics. vol. 73. T. Friedmann. J. C. Dunlap. and S. F. Goodwin. Eds. Academic Press. 2011. pp. 1-50. [CrossRef]
- M. B. Feany and W. W. Bender. ‘A Drosophila model of Parkinson’s disease’. Nature vol. 404. no. 6776. Mar. 2000. [CrossRef]
- R. Costa. E. Speretta. D. C. Crowther. and I. Cardoso. ‘Testing the Therapeutic Potential of Doxycycline in a Drosophila melanogaster Model of Alzheimer Disease *’. J. Biol. Chem vol. 286. no. 48. pp. 41647-41655. Dec. 2011. [CrossRef]
- K. Prüßing. A. Voigt. and J. B. Schulz. ‘Drosophila melanogaster as a model organism for Alzheimer’s disease’. Mol. Neurodegener. vol. 8. no. 1. p. 35. Nov. 2013. [CrossRef]
- X. Cheng. C. Song. Y. Du. U. Gaur. and M. Yang. ‘Pharmacological Treatment of Alzheimer’s Disease: Insights from Drosophila melanogaster’. Int. J. Mol. Sci. vol. 21. no. 13. Jan. 2020. [CrossRef]
- Z. Mirzoyan. M. Sollazzo. M. Allocca. A. M. Valenza. D. Grifoni. and P. Bellosta. ‘Drosophila melanogaster: A Model Organism to Study Cancer’. Front. Genet. vol. 10. 2019. Accessed: Jan. 21. 2024. [Online]. Available: https://www.frontiersin.org/articles/10.3389/fgene.2019.00051. [CrossRef]
- C. Munnik. M. P. Xaba. S. T. Malindisa. B. L. Russell. and S. A. Sooklal. ‘Drosophila melanogaster: A platform for anticancer drug discovery and personalized therapies’. Front. Genet vol. 13. 2022. Accessed: Jan. 21. 2024. [Online]. Available: https://www.frontiersin.org/articles/10.3389/fgene.2022.949241. [CrossRef]
- S. Ma et al. ‘The multisubstrate deoxyribonucleoside kinase of Drosophila melanogaster as a therapeutic suicide gene of breast cancer cells’. J. Gene Med vol. 13. no. 6. pp. 305-311. 2011. [CrossRef]
- N. A. Ramesh et al. ‘A genetic and physiological model of renal dysfunction in Lowe syndrome’. bioRxiv. p. 2024.01.15.575703. Jan. 16. 2024. [CrossRef]
- J. P. Álvarez-Rendón. R. Salceda. and J. R. Riesgo-Escovar. ‘Drosophila melanogaster as a Model for Diabetes Type 2 Progression’. BioMed Res. Int vol. 2018. p. e1417528. Apr. 2018. [CrossRef]
- N. Baenas and A. E. Wagner. ‘Drosophila melanogaster as a Model Organism for Obesity and Type-2 Diabetes Mellitus by Applying High-Sugar and High-Fat Diets’. Biomolecules vol. 12. no. 2. Art. no. 2. Feb. 2022. [CrossRef]
- Y. Miao et al. ‘Drosophila melanogaster diabetes models and its usage in the research of anti-diabetes management with traditional Chinese medicines’. Front. Med. vol. 9. 2022. Accessed: Jan. 21. 2024. [Online]. Available: https://www.frontiersin.org/articles/10.3389/fmed.2022.953490. [CrossRef]
- Y. H. Inoue. H. Katsube. and Y. Hinami. ‘Drosophila Models to Investigate Insulin Action and Mechanisms Underlying Human Diabetes Mellitus’. in Drosophila Models for Human Diseases. M. Yamaguchi. Ed. in Advances in Experimental Medicine and Biology. Singapore: Springer. 2018. pp. 235-256. [CrossRef]
- F. A. Al-Momani and A. M. Massadeh. ‘Effect of different heavy-metal concentrations on Drosophila melanogaster larval growth and development’. Biol. Trace Elem. Res. vol. 108. no. 1. pp. 271-277. Dec. 2005. [CrossRef]
- M. R. Slobodian. J. D. Petahtegoose. A. L. Wallis. D. C. Levesque. and T. J. S. Merritt. ‘The Effects of Essential and Non-Essential Metal Toxicity in the Drosophila melanogaster Insect Model: A Review’ Toxics vol. 9. no. 10. Art. no. 10. Oct. 2021. [CrossRef]
- T. Tongesayi. P. Fedick. L. Lechner. C. Brock. A. Le Beau. and C. Bray. ‘Daily bioaccessible levels of selected essential but toxic heavy metals from the consumption of non-dietary food sources’. Food Chem. Toxicol. vol. 62. pp. 142-147. Dec. 2013. [CrossRef]
- R. Burke. ‘Molecular physiology of copper in drosophila melanogaster’ Curr. Opin. Insect Sci. vol. 51. p. 100892. 2022. Accessed: Jan. 16. 2024. [Online]. Available: https://www.sciencedirect.com/science/article/pii/S221457452200027X?casa_token=pFqqOwYOLQcAAAAA:izJoBleLYyOiZxeYDZyxGkVZEIIma9lC0A2gMfRAxgxQmU3twI5i8PwuEtme4S2YKVOKWtyCew. [CrossRef]
- J. A. Navarro and S. Schneuwly. ‘Copper and zinc homeostasis: lessons from Drosophila melanogaster’. Front. Genet. vol. 8. p. 223. 2017. [CrossRef]
- Southon. R. Burke. M. Norgate. P. Batterham. and J. Camakaris. ‘Copper homoeostasis in Drosophila melanogaster S2 cells’. Biochem. J. vol. 383. no. 2. 2004. [CrossRef]
- E. Demir and F. Turna Demir. ‘Drosophila melanogaster as a dynamic in vivo model organism reveals the hidden effects of interactions between microplastic/nanoplastic and heavy metals’. J. Appl. Toxicol. vol. 43. no. 2. pp. 212-219. 2023. [CrossRef]
- X. Li. Y. Sun. S. Gao. Y. Li. L. Liu. and Y. Zhu. ‘Taste coding of heavy metal ion-induced avoidance in Drosophila’. iScience vol. 26. no. 5. p. 106607. May 2023. [CrossRef]
- L. Mo et al. ‘Hedgehog pathway is negatively regulated during the development of Drosophila melanogaster PheRS-m (Drosophila homologs gene of human FARS2) mutants’. Hum. Cell vol. 36. no. 1. pp. 121-131. Jan. 2023. [CrossRef]
- S. N. Chan and J. W. Pek. ‘Distinct biogenesis pathways may have led to functional divergence of the human and Drosophila Arglu1 sisRNA’. EMBO Rep vol. 24. no. 2. p. e54350. Feb. 2023. [CrossRef]
- L. Willoughby. H. Chung. C. Lumb. C. Robin. P. Batterham. and P. J. Daborn. ‘A comparison of Drosophila melanogaster detoxification gene induction responses for six insecticides. caffeine and phenobarbital’. Insect Biochem. Mol. Biol vol. 36. no. 12. pp. 934-942. Dec. 2006. [CrossRef]
- K. Trinh et al. ‘Induction of the Phase II Detoxification Pathway Suppresses Neuron Loss in Drosophila Models of Parkinson’s Disease’. J. Neurosci vol. 28. no. 2. pp. 465-472. Jan. 2008. [CrossRef]
- C. M. Wood. A. P. Farrell. and C. J. Brauner. Homeostasis and Toxicology of Essential Metals. Academic Press. 2012.
- M. A. Zoroddu. J. Aaseth. G. Crisponi. S. Medici. M. Peana. and V. M. Nurchi. ‘The essential metals for humans: a brief overview’. J. Inorg. Biochem vol. 195. pp. 120-129. 2019. Accessed: Dec. 01. 2023. [Online]. Available: https://www.sciencedirect.com/science/article/pii/S0162013418306846?casa_token=R9XvW5gvzaMAAAAA:RP8DrUsKLyWE6luXgsWTFsWOiSsDWUMttt_PJkyQR9aB6075Ig477mqdoVNJvMi2V8f-vbWxFA. [CrossRef]
- K. Jomova et al. ‘Essential metals in health and disease’. Chem. Biol. Interact. vol. 367. p. 110173. Nov. 2022. [CrossRef]
- D. Raj and S. K. Maiti. ‘Sources. bioaccumulation. health risks and remediation of potentially toxic metal (loid) s (As. Cd. Cr. Pb and Hg): an epitomised review’. Environ. Monit. Assess vol. 192. no. 2. p. 108. 2020. Accessed: Dec. 01. 2023. [Online]. Available: https://idp.springer.com/authorize/casa?redirect_uri=https://link.springer.com/article/10.1007/s10661-019-8060-5&casa_token=mGZ0TObc9usAAAAA:tWWIIF3SByFfFK4M5LN843_PkcaHv9wv9vmHn7V1rfIyWJwPDi6W_KAh4r7XZzhCEP8KT78YlNtMj1PT. [CrossRef]
- Cherfi. S. Abdoun. and O. Gaci. ‘Food survey: Levels and potential health risks of chromium. lead. zinc and copper content in fruits and vegetables consumed in Algeria’. Food Chem. Toxicol vol. 70. pp. 48-53. Aug. 2014. [CrossRef]
- N. Bagdatlioglu. C. Nergiz. and P. G. Ergonul. ‘Heavy metal levels in leafy vegetables and some selected fruits’. J. Für Verbraucherschutz Leb. vol. 5. no. 3. pp. 421-428. Sep. 2010. [CrossRef]
- Pruteanu. I. Voicea. and V. Fatu. ‘Accumulation of copper in vegetables and fruits’. presented at the 21st International Scientific Conference Engineering for Rural Development. May 2022. [CrossRef]
- D. Demirezen and K. Uruç. ‘Comparative study of trace elements in certain fish. meat and meat products’. Meat Sci. vol. 74. no. 2. pp. 255-260. Oct. 2006. [CrossRef]
- ]Ľ. Harangozo et al. ‘Copper content in cereals grown in the model condition’. Potravinarstvo Slovak J. Food Sci. vol. 11. no. 1. pp. 20-25. Jan. 2017. [CrossRef]
- S. Kashian and A. A. Fathivand. ‘Estimated daily intake of Fe. Cu. Ca and Zn through common cereals in Tehran. Iran’. Food Chem. vol. 176. pp. 193-196. Jun. 2015. [CrossRef]
- B. R. Stern et al. ‘Copper and Human Health: Biochemistry. Genetics. and Strategies for Modeling Dose-response Relationships’. J. Toxicol. Environ. Health Part B vol. 10. no. 3. pp. 157-222. Apr. 2007. [CrossRef]
- L. M. Gaetke and C. K. Chow. ‘Copper toxicity. oxidative stress. and antioxidant nutrients’. Toxicology vol. 189. no. 1-2. pp. 147-163. Jul. 2003. [CrossRef]
- J. W. Spears. E. B. Kegley. and L. A. Mullis. ‘Bioavailability of copper from tribasic copper chloride and copper sulfate in growing cattle’. Anim. Feed Sci. Technol. vol. 116. no. 1. pp. 1-13. Sep. 2004. [CrossRef]
- D. Jiang et al. ‘Redox Reactions of Copper Complexes Formed with Different β-Amyloid Peptides and Their Neuropathalogical Relevance’. Biochemistry vol. 46. no. 32. pp. 9270-9282. Aug. 2007. [CrossRef]
- M. Adrees et al. ‘The effect of excess copper on growth and physiology of important food crops: a review’. Environ. Sci. Pollut. Res. vol. 22. pp. 8148-8162. 2015. Accessed: Dec. 01. 2023. [Online]. Available: https://idp.springer.com/authorize/casa?redirect_uri=https://link.springer.com/article/10.1007/s11356-015-4496-5&casa_token=3HPXX7mXHbIAAAAA:xZOl8aFLsfXIpKE95bMZq9p9XS30aWcJUWD2GSKD_-zbsZSYp2HVK1DlbYumwaY4xLXX-m_WjWOLT7L1. [CrossRef]
- D. Huang et al. ‘The effects of rice straw biochar on indigenous microbial community and enzymes activity in heavy metal-contaminated sediment’. Chemosphere vol. 174. pp. 545-553. 2017. Accessed: Dec. 01. 2023. [Online]. Available: https://www.sciencedirect.com/science/article/pii/S0045653517301492?casa_token=_pxZ_ZOXE2wAAAAA:srx6h9_BJoLLmzQgb_LkmR2GiMNJQBDbiUAt9C3b11tcbIXiekswOWMoSgK-BgFgYikUmt42tg. [CrossRef]
- M. Zhang. D.-M. Xian. N. Zhang. H.-H. Li. and Z.-L. You. ‘The construction of versatile azido-bridged Schiff base Copper (II) complexes with xanthine oxidase inhibitory activity’. Struct. Chem. vol. 23. pp. 1489-1496. 2012. Accessed: Dec. 01. 2023. [Online]. Available: https://idp.springer.com/authorize/casa?redirect_uri=https://link.springer.com/article/10.1007/s11224-012-9956-7&casa_token=9xac46Yb7YoAAAAA:LvnT_U9do9lI_6PGPsgkNX1OV1fPfez29UfPhaNhNi3aOmIlKqf1wqn5h-DVjdPCOQ06nTILNmD1VoHr. [CrossRef]
- E. Yusefi-Tanha. S. Fallah. A. Rostamnejadi. and L. R. Pokhrel. ‘Particle size and concentration dependent toxicity of copper oxide nanoparticles (CuONPs) on seed yield and antioxidant defense system in soil grown soybean (Glycine max cv. Kowsar)’. Sci. Total Environ vol. 715. p. 136994. 2020. Accessed: Dec. 01. 2023. [Online]. Available: https://www.sciencedirect.com/science/article/pii/S0048969720305040?casa_token=igXo1RRiJQUAAAAA:I_IcHbkrWlsE-JfQjiCxSjigFojkQVcpW_WGIkFsav4P0diupz0TGRDv7u5fMakNXbk4YVXtnw. [CrossRef]
- S. I. Lavrentyeva. L. Y. Ivachenko. K. S. Golokhvast. and M. A. Nawaz. ‘Ribonuclease activity of Glycine max and Glycine soja sprouts as a marker adaptation to copper sulphate and zinc sulphate toxicity’. Biochem. Syst. Ecol. vol. 83. pp. 66-70. 2019. Accessed: Dec. 01. 2023. [Online]. Available: https://www.sciencedirect.com/science/article/pii/S0305197818305659?casa_token=rOJlpgSSphQAAAAA:Ydm7EWuB8RokVLCpjA83zLXCladUpoMjOPiVISMXXUN31t7_sOYzt515ElykfTGC5iTbH5eCpQ. [CrossRef]
- M. Jaishankar. T. Tseten. N. Anbalagan. B. B. Mathew. and K. N. Beeregowda. ‘Toxicity. mechanism and health effects of some heavy metals. Interdiscip Toxicol 7: 60-72’. 2014. [CrossRef]
- T. Jan. M. Azam. K. Siddiqui. A. Ali. I. Choi. and Q. M. R. Haq. ‘Heavy metals and human health: mechanistic insight into toxicity and counter defense system of antioxidants’. Int. J. Mol. Sci. vol. 16. no. 12. pp. 29592-29630. 2015. Accessed: Dec. 01. 2023. [Online]. Available: https://www.mdpi.com/1422-0067/16/12/29592. [CrossRef]
- G. Y. Zaghloul. H. A. Eissa. A. Y. Zaghloul. M. S. Kelany. M. A. Hamed. and K. M. E. Moselhy. ‘Impact of some heavy metal accumulation in different organs on fish quality from Bardawil Lake and human health risks assessment’ Geochem. Trans. vol. 25. no. 1. p. 1. Jan. 2024. [CrossRef]
- Duruibe. Ogwuegbu. and Egwurugwu. ‘Heavy metal pollution and human biotoxic effects’. Int. J. Phys. Sci. vol. 2. no. 5. pp. 112-118. 2007. Accessed: Dec. 01. 2023. [Online]. Available: https://academicjournals.org/journal/IJPS/article-full-text-pdf/59CA35213127.
- Giannakoula. I. Therios. and C. Chatzissavvidis. ‘Effect of lead and copper on photosynthetic apparatus in citrus (Citrus aurantium L.) plants. The role of antioxidants in oxidative damage as a response to heavy metal stress’. Plants vol. 10. no. 1. p. 155. 2021. Accessed: Dec. 01. 2023. [Online]. Available: https://www.mdpi.com/2223-7747/10/1/155. [CrossRef]
- L. Shvachiy. V. Geraldes. Â. Amaro-Leal. and I. Rocha. ‘Persistent effects on cardiorespiratory and nervous systems induced by long-term lead exposure: results from a longitudinal study’. Neurotox. Res. vol. 37. pp. 857-870. 2020. Accessed: Nov. 28. 2023. [Online]. Available: https://idp.springer.com/authorize/casa?redirect_uri=https://link.springer.com/article/10.1007/s12640-020-00162-8&casa_token=Nm8RaX-XldkAAAAA:hZWncHlznxWnNUmj10kjlzGSmkVrHAbrhHnuYU2VhPfq5LFQUat6S148LlU-OoueMLlUpAN00Scg72iD. [CrossRef]
- E. J. Morley. H. V. Hirsch. K. Hollocher. and G. A. Lnenicka. ‘Effects of chronic lead exposure on the neuromuscular junction in Drosophila larvae’. Neurotoxicology vol. 24. no. 1. pp. 35-41. 2003. Accessed: Dec. 01. 2023. [Online]. Available: https://www.sciencedirect.com/science/article/pii/S0161813X02000955. [CrossRef]
- F. A. Al-Momani and A. M. Massadeh. ‘Effect of different heavy-metal concentrations on Drosophila melanogaster larval growth and development’. Biol. Trace Elem. Res. vol. 108. pp. 271-277. 2005. Accessed: Dec. 01. 2023. [Online]. Available: https://idp.springer.com/authorize/casa?redirect_uri=https://link.springer.com/article/10.1385/BTER:108:1-3:271&casa_token=oopytYw7h_gAAAAA:Z85BaCAO0WIhan06sS0RFPOQFV8aTFiPRcwVT4bxvDOH56oURaoE31dhlkNELWNl0BfmtCRC4mkK0Ff5. [CrossRef]
- M. S. Abdullahi. ‘Toxic effects of lead in humans: an overview’. Glob. Adv. Res. J. Environ. Sci. Toxicol. vol. 2. no. 6. pp. 157-162. 2013. Accessed: Dec. 01. 2023. [Online]. Available: http://trusting-shtern.139-59-89-2.plesk.page/toxic-effects-of-lead-in-humans-an-overview_YTozNTo5.pdf.
- Q. Jamal et al. ‘Heavy metals accumulation and their toxic effects’. J. Bio-Mol. Sci. JBMS vol. 1. no. 1. pp. 27-36. 2013. Accessed: Dec. 01. 2023. [Online]. Available: https://www.researchgate.net/profile/Qaiser-Jamal/publication/276921553_Heavy_Metals_Accumulation_and_Their_Toxic_Effects_Review/links/56f12e9a08aedbe21877222d/Heavy-Metals-Accumulation-and-Their-Toxic-Effects-Review.pdf.
- R. A. Festa and D. J. Thiele. ‘Copper: an essential metal in biology’. Curr. Biol. vol. 21. no. 21. pp. R877-R883. 2011. Accessed: Dec. 01. 2023. [Online]. Available: https://www.cell.com/current-biology/pdf/S0960-9822(11)01079-7.pdf.
- L. Frat et al. ‘Single and mixed exposure to cadmium and mercury in Drosophila melanogaster: Molecular responses and impact on post-embryonic development’. Ecotoxicol. Environ. Saf.. vol. 220. p. 112377. Sep. 2021. [CrossRef]
- X. Zeng. C. Deng. Y. Liang. J. Fu. S. Zhang. and T. Ni. ‘Ecological risk evaluation and sensitivity analysis of heavy metals on soil organisms under human activities in the Tibet Plateau. China’. Plos One vvol. 18. no. 8. Accessed: Nov. 28. 2023. [Online]. Available: https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0285116. [CrossRef]
- et al. ‘Lead Toxicity: Health Hazards. Influence on Food Chain. and Sustainable Remediation Approaches’. Int. J. Environ. Res. Public. Health vol. 17. no. 7. Art. no. 7. Jan. 2020. [CrossRef]
- V. Mohod and J. Dhote. ‘Review of heavy metals in drinking water and their effect on human health’. Int. J. Innov. Res. Sci. Eng. Technol. vol. 2. no. 7. Art. no. 7. 2013. Accessed: Nov. 28. 2023. [Online]. Available: https://www.researchgate.net/profile/Jayashree-Dhote/publication/251237189_Review_of_heavy_metals_in_drinking_water_and_their_effect_on_human_health/links/00b4951ef99e1d1f97000000/Review-of-heavy-metals-in-drinking-water-and-their-effect-on-human-health.pdf.
- Y. Finkelstein. M. E. Markowitz. and J. F. Rosen. ‘Low-level lead-induced neurotoxicity in children: an update on central nervous system effects’. Brain Res. Rev vol. 27. no. 2. Art. no. 2. Jul. 1998. [CrossRef]
- C. S. Mendes. I. Bartos. T. Akay. S. Márka. and R. S. Mann. ‘Quantification of gait parameters in freely walking wild type and sensory deprived Drosophila melanogaster’elife vol. 2. p. e00231. 2013. Accessed: Dec. 01. 2023. [Online]. Available: https://elifesciences.org/articles/231. [CrossRef]
- W. Jin. R. M. Riley. R. D. Wolfinger. K. P. White. G. Passador-Gurgel. and G. Gibson. ‘The contributions of sex. genotype and age to transcriptional variance in Drosophila melanogaster’. Nat. Genet vol. 29. no. 4. pp. 389-395. 2001. Accessed: Dec. 01. 2023. [Online]. Available: https://www.nature.com/articles/ng766z. [CrossRef]
- E. H. Morrow. A. Leijon. and A. Meerupati. ‘Hemiclonal analysis reveals significant genetic. environmental and genotype × environment effects on sperm size in Drosophila melanogaster’. J. Evol. Biol vol. 21. no. 6. pp. 1692-1702. Nov. 2008. [CrossRef]
- B. Gallia. S. Ioan. P. Sorina. T. Gheorghe. and I. Stefanescu. ‘The involvement of deuterium presence in the Drosophila melanogaster evolution: diverse effects of deuterium concentrations upon the phenotype of different strains and Romanian ecotypes’. Smart Energy Sustain. Environ. vol. 16. no. 2. p. 133. 2013. Accessed: Dec. 01. 2023. [Online]. Available: https://search.proquest.com/openview/48f43fa6fdfeaf86a57bc2167b055d40/1?pq-origsite=gscholar&cbl=2045793.
- L. Everson da Silva et al. ‘In Vitro Antiprotozoal Evaluation of Zinc and Copper Complexes Based on Sulfonamides Containing 8-Aminoquinoline Ligands’. Lett. Drug Des. Discov vol. 7. no. 9. pp. 679-685. Nov. 2010. [CrossRef]
- N. Asif et al. ‘Toxicological assessment of Phormidium sp. derived copper oxide nanoparticles for its biomedical and environmental applications’. Sci. Rep. vol. 13. no. 1. Art. no. 1. Apr. 2023. [CrossRef]
- O. Abolaji. K. D. Fasae. C. E. Iwezor. M. Aschner. and E. O. Farombi. ‘Curcumin attenuates copper-induced oxidative stress and neurotoxicity in Drosophila melanogaster’. Toxicol. Rep vol. 7. pp. 261-268. 2020. Accessed: Jan. 16. 2024. [Online]. Available: https://www.sciencedirect.com/science/article/pii/S2214750019306043. [CrossRef]
- Southon. A. Farlow. M. Norgate. R. Burke. and J. Camakaris. ‘Malvolio is a copper transporter in Drosophila melanogaster’. J. Exp. Biol. vol. 211. no. 5. pp. 709-716. 2008. Accessed: Jan. 16. 2024. [Online]. Available: https://journals.biologists.com/jeb/article-abstract/211/5/709/18125. [CrossRef]
- P. T. Halmenschelager and J. B. T. da Rocha. ‘Biochemical CuSO 4 Toxicity in Drosophila melanogaster depends on sex and developmental stage of exposure’. Biol. Trace Elem. Res. vol. 189. pp. 574-585. 2019. Accessed: Nov. 28. 2023. [Online]. Available: https://idp.springer.com/authorize/casa?redirect_uri=https://link.springer.com/article/10.1007/s12011-018-1475-y&casa_token=CO_xdDrjXpQAAAAA:qWTtFM6KyMOKFrHo4FSavQgZZinXbzNtimb2Qq9q6F5heS76xjFnCvkRR-COGXC4dDtJC8vhCNJHSjgE. [CrossRef]
- M. A. Balinski and R. C. Woodruff. ‘Differential sexual survival of Drosophila melanogaster on copper sulfate’. Genetica vol. 145. pp. 131-137. 2017. Accessed: Nov. 28. 2023. [Online]. Available: https://idp.springer.com/authorize/casa?redirect_uri=https://link.springer.com/article/10.1007/s10709-017-9951-4&casa_token=lwJNGLeE7ZcAAAAA:6loOri92nj1csDLftBbi1DopNcTIbYn-Enpz1XnN6oRjmvY0gp6IQXT9eWGGc1iM0N9K92NASZTycypk. [CrossRef]
- ‘Copper and its role in the human body - the importance of establishing copper concentrations in the body’. Romanian J. Mil. Med. vol. 124. no. 2. May 2021. [CrossRef]
- M. Angelova. S. Asenova. V. Nedkova. and R. Koleva-Kolarova. ‘COPPER IN THE HUMAN ORGANISM’. Trakia J. Sci. vol. 9. no. 1. 2011.
- Hyre. K. Casanova-Hampton. and S. Subashchandrabose. ‘Copper Homeostatic Mechanisms and Their Role in the Virulence of Escherichia coli and Salmonella enterica’. EcoSal Plus vol. 9. no. 2. p. eESP-0014-2020. Dec. 2021. [CrossRef]
- E. Santo. N. Taudte. D. H. Nies. and G. Grass. ‘Contribution of Copper Ion Resistance to Survival of Escherichia coli on Metallic Copper Surfaces’. Appl. Environ. Microbiol vol. 74. no. 4. pp. 977-986. Feb. 2008. [CrossRef]
- P. Saenkham-Huntsinger et al. ‘Copper Resistance Promotes Fitness of Methicillin-Resistant Staphylococcus aureus during Urinary Tract Infection’. mBio vol. 12. no. 5. pp. e02038-21. Oct. 2021. [CrossRef]
- W. B. Wang and J. C. Clapper. ‘Antibacterial activity of electrospun polyacrylonitrile copper nanoparticle nanofibers on antibiotic resistant pathogens and methicillin resistant Staphylococcus aureus (MRSA)’. Nanomaterials vol. 12. no. 13. p. 2139. 2022. Accessed: Jan. 16. 2024. [Online]. Available: https://www.mdpi.com/2079-4991/12/13/2139. [CrossRef]
- Y. Zhuang. L. Ren. S. Zhang. X. Wei. K. Yang. and K. Dai. ‘Antibacterial effect of a copper-containing titanium alloy against implant-associated infection induced by methicillin-resistant Staphylococcus aureus’. Acta Biomater. vol. 119. pp. 472-484. 2021. Accessed: Jan. 16. 2024. [Online]. Available: https://www.sciencedirect.com/science/article/pii/S1742706120306152?casa_token=jmF9O7-xdvsAAAAA:8rFT2nSP8lXqhxmHV20-b9HVEj38foo_kyJoTrvuYIlUBAVToHIutNWYJjnQ-PgJ0DopbPtfHA. [CrossRef]
- Andreou. S. Trantza. D. Filippou. N. Sipsas. and S. Tsiodras. ‘COVID-19: The Potential Role of Copper and N-acetylcysteine (NAC) in a Combination of Candidate Antiviral Treatments Against SARS-CoV-2’. In Vivo vol. 34. no. 3 suppl. pp. 1567-1588. 2020. [CrossRef]
- S. Raha. R. Mallick. S. Basak. and A. K. Duttaroy. ‘Is copper beneficial for COVID-19 patients?’. Med. Hypotheses vol. 142. p. 109814. Sep. 2020. [CrossRef]
- A. Cortes and J. M. Zuñiga. ‘The use of copper to help prevent transmission of SARS-coronavirus and influenza viruses. A general review’. Diagn. Microbiol. Infect. Dis vol. 98. no. 4. p. 115176. Dec. 2020. [CrossRef]
- Y. Fujimori et al. ‘Novel Antiviral Characteristics of Nanosized Copper(I) Iodide Particles Showing Inactivation Activity against 2009 Pandemic H1N1 Influenza Virus’. Appl. Environ. vol. 78. no. 4. pp. 951-955. Feb. 2012. [CrossRef]
- K. Imai et al. ‘Inactivation of high and low pathogenic avian influenza virus H5 subtypes by copper ions incorporated in zeolite-textile materials’. Antiviral Res. vol. 93. no. 2. pp. 225-233. Feb. 2012. [CrossRef]
- M. Rai et al. ‘Copper and copper nanoparticles: role in management of insect-pests and pathogenic microbes’. Nanotechnol. Rev vol. 7. no. 4. pp. 303-315. Aug. 2018. [CrossRef]
- L. Green. M. Coronado-Zamora. S. Radío. G. E. Rech. J. Salces-Ortiz. and J. González. ‘The genomic basis of copper tolerance in Drosophila is shaped by a complex interplay of regulatory and environmental factors’. BMC Biol. vol. 20. no. 1. Art. no. 1. Dec. 2022. [CrossRef]
- Z.-H. Liu et al. ‘Oxidative stress caused by lead (Pb) induces iron deficiency in Drosophila melanogaster’. Chemosphere. vol. 243. p. 125428. Mar. 2020. [CrossRef]
- D. Fatmawati. D. Khoiroh. S. Zubaidah. H. Susanto. M. Agustin. and A. Fauzi. ‘Wing morphological changes of Drosophila melanogaster exposed with lead in nine generations’. in AIP Conference Proceedings. AIP Publishing. 2023. Accessed: Nov. 28. 2023. [Online]. Available: https://pubs.aip.org/aip/acp/article-abstract/2634/1/020041/2871056ip. [CrossRef]
- N. L. Cooper. J. R. Bidwell. and A. Kumar. ‘Toxicity of copper. lead. and zinc mixtures to Ceriodaphnia dubia and Daphnia carinata’. Ecotoxicol. Environ. Saf. vol. 72. no. 5. Jul. 2009. [CrossRef]
- M. T. S. D. Vasconcelos and M. F. C. Leal. ‘Antagonistic interactions of Pb and Cd on Cu uptake. growth inhibition and chelator release in the marine algae Emiliania huxleyi’. Mar. Chem. vol. 75. no. 1. Art. no. 1. Jul. 2001. [CrossRef]
- D. Z. An et al. ‘Influence of Iron Supplementation on DMT1 (IRE)-induced Transport of Lead by Brain Barrier Systems in vivo’. Biomed. Environ. Sci. vol. 28. no. 9. Art. no. 9. Sep. 2015. [CrossRef]
- E. R. Everman. K. M. Cloud-Richardson. and S. J. Macdonald. ‘Characterizing the genetic basis of copper toxicity in Drosophila reveals a complex pattern of allelic. regulatory. and behavioral variation’. Genetics vol. 217. no. 1. Art. no. 1. Jan. 2021. [CrossRef]
Figure 1.
Genotypes of Drosophila melanogaster used, (a)-wild genotype, (b)- white mutant genotype (c)- brown mutant genotype, (d)- white-vestigial mutant genotype.
Figure 1.
Genotypes of Drosophila melanogaster used, (a)-wild genotype, (b)- white mutant genotype (c)- brown mutant genotype, (d)- white-vestigial mutant genotype.

Figure 3.
Effect of copper on different genotypes of Drosophila melanogaster in adult formation: (a)- wild wild genotype, (b)- brown genotype (c)- white mutant genotype, (d)- white-vestigial mutant genotype.
Figure 3.
Effect of copper on different genotypes of Drosophila melanogaster in adult formation: (a)- wild wild genotype, (b)- brown genotype (c)- white mutant genotype, (d)- white-vestigial mutant genotype.
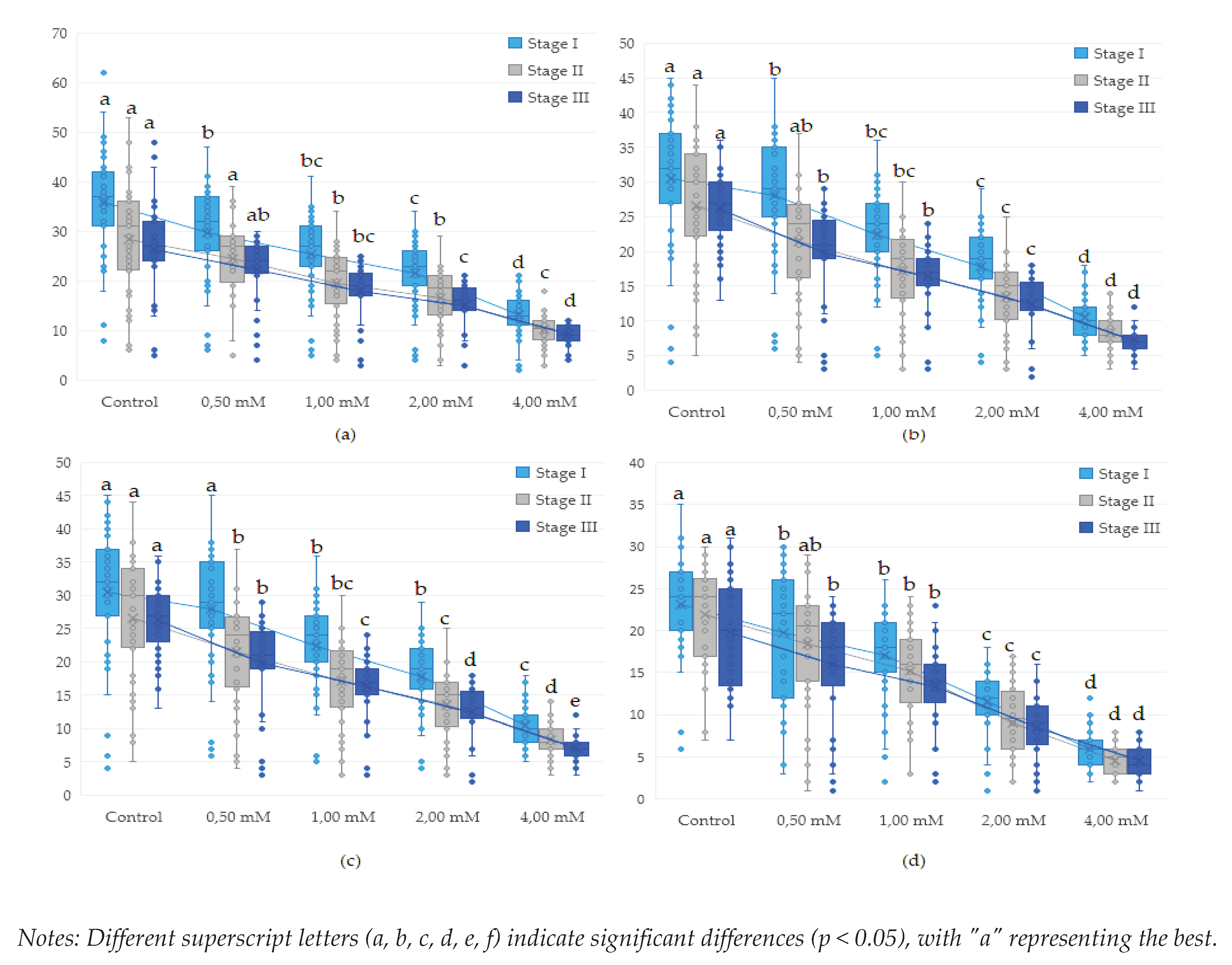
Figure 4.
Effect of copper on different genotypes of Drosophila melanogaster in adult formation: (a)- wild genotype, (b)- brown genotype (c)- white mutant genotype, (d)- white-vestigial mutant genotype.
Figure 4.
Effect of copper on different genotypes of Drosophila melanogaster in adult formation: (a)- wild genotype, (b)- brown genotype (c)- white mutant genotype, (d)- white-vestigial mutant genotype.
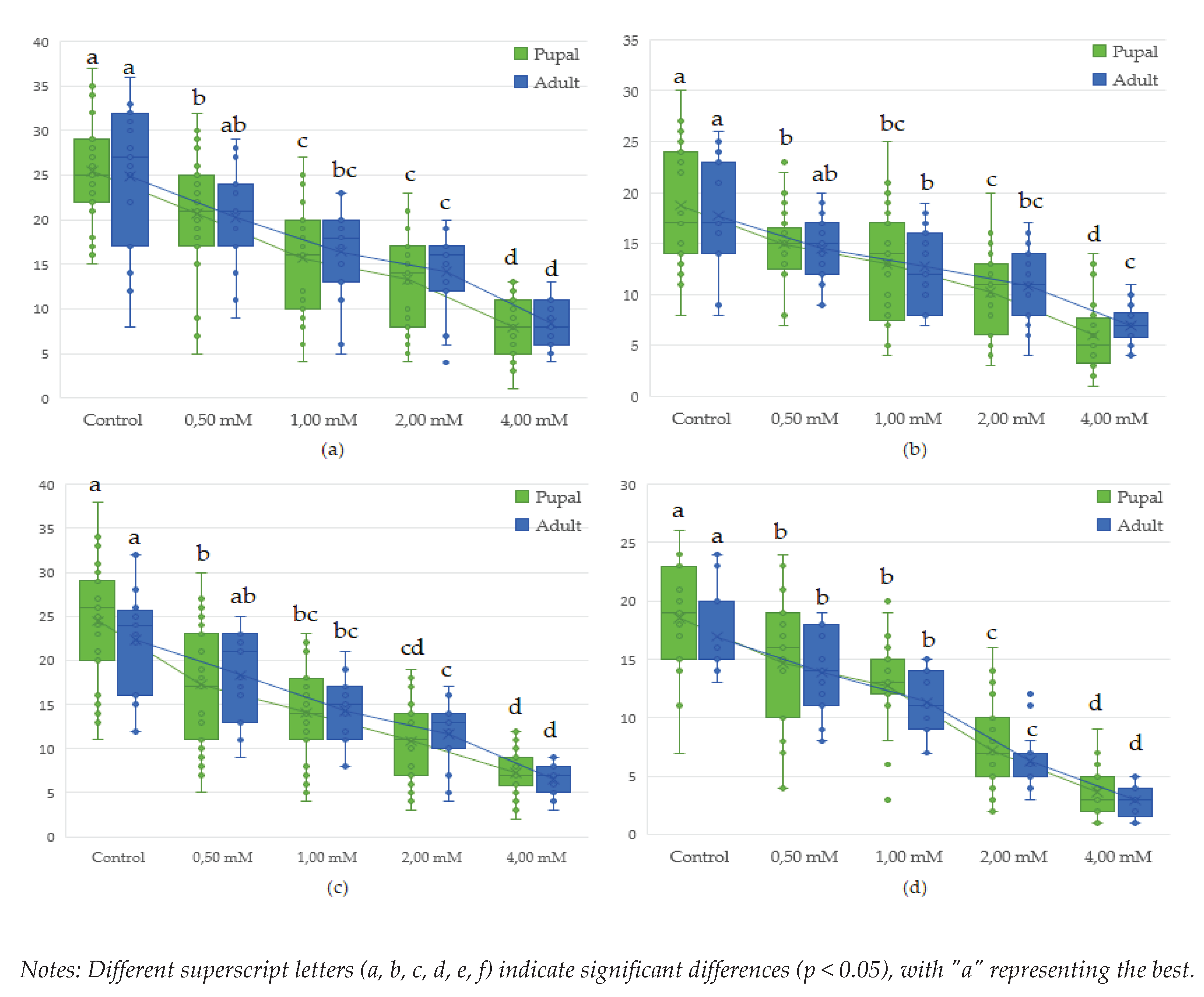
Figure 8.
Polar histrogram of the effect of copper on various genotypes of Drosophila melanogaster in adult development.
Figure 8.
Polar histrogram of the effect of copper on various genotypes of Drosophila melanogaster in adult development.
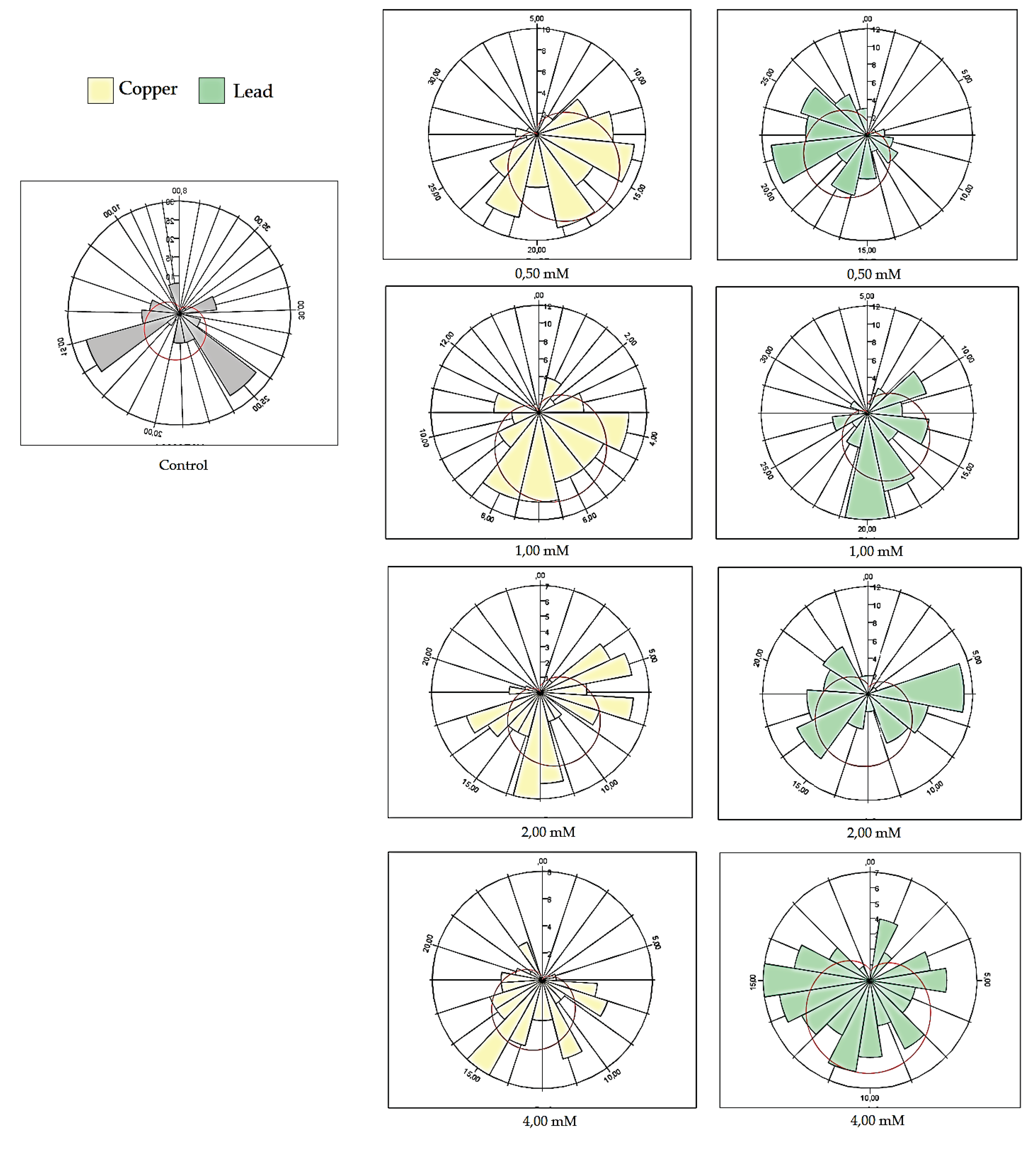
Table 1.
Analysis of factors involved in the process of inhibition of proliferation at different concentrations of copper (CuSO4 ) and lead Pb(C2H3O2)2 in larval stages.
Table 1.
Analysis of factors involved in the process of inhibition of proliferation at different concentrations of copper (CuSO4 ) and lead Pb(C2H3O2)2 in larval stages.
| Analysis factor | df | SS | S2 | F | Pr(>F) | |
|---|---|---|---|---|---|---|
| Larval stage I | Concentration | 4 | 55264 | 13816 | 240.429 | < 2e-16 *** |
| Genotype | 3 | 24097 | 8032 | 139.782 | < 2e-16 *** | |
| Metal | 1 | 3936 | 3936 | 68.499 | 2.74e-16 *** | |
| Concentration: Genotyp | 12 | 1240 | 103 | 1.798 | 0.043 * | |
| Concentration:Metal | 4 | 1200 | 300 | 5.219 | 0.000356 *** | |
| Genotype:Metal | 3 | 290 | 97 | 1.680 | 0.169 Ns | |
| Concentration: Genotip: Metal | 12 | 232 | 19 | 0.337 | 0.982 Ns | |
| Residuals | 1520 | 87344 | 57 | |||
| Larvar stage II | Concentration | 4 | 37452 | 9363 | 200.948 | < 2e-16 *** |
| Genotype | 3 | 9527 | 3176 | 68.159 | < 2e-16 *** | |
| Metal | 1 | 2763 | 2763 | 59.292 | 2.56e-14 *** | |
| Concentration: Genotyp | 12 | 1617 | 135 | 2.892 | 0.000583 *** | |
| Concentration:Metal | 4 | 1037 | 259 | 5.563 | 0.000192 *** | |
| Genotype:Metal | 3 | 130 | 43 | 0.933 | 0.423 Ns | |
| Concentration: Genotip: Metal | 12 | 284 | 24 | 0.508 | 0.910 Ns | |
| Residuals | 1392 | 64859 | 47 | |||
| Larvar stage III | Concentration | 4 | 31284 | 7821 | 248.496 | < 2e-16 *** |
| Genotype | 3 | 8277 | 2759 | 87.665 | < 2e-16 *** | |
| Metal | 1 | 2266 | 2266 | 71.992 | < 2e-16 *** | |
| Concentration: Genotyp | 12 | 1084 | 90 | 2.870 | 0.000646 *** | |
| Concentration:Metal | 4 | 748 | 187 | 5.943 | 9.72e-05 *** | |
| Genotype:Metal | 3 | 114 | 38 | 1.209 | 0.304 Ns | |
| Concentration: Genotip: Metal | 12 | 199 | 17 | 0.526 | 0.899 Ns | |
| Residuals | 1278 | 40223 | 31 |
SS- sum of square, dF-Degrees of freedom; S2 -Mean square. “Ns”-not significant, “*”- p < 0.05, “**”- p < 0.01, *** Significant at p < 0.001.
Table 4.
Results on D. melanogaster genotype proliferation and viability following exposure to different lead concentrations Pb(C2H3O2)2.
Table 4.
Results on D. melanogaster genotype proliferation and viability following exposure to different lead concentrations Pb(C2H3O2)2.
| Genotype | Concentration (mM) | Lead (C2H3O2)2 | |||||||||
| StageI | IC | StageII | IC | StageIII | IC | Pupal | IC | Adult | IC | ||
| Mean | Mean | Mean | Mean | Mean | |||||||
| wild | Control | 35.9a | 0.00 | 28.33a | 0.00 | 26.39a | 0.00 | 25.41a | 0.00 | 24.87a | 0.00 |
| 0.50 | 34.3ab | 4.63 | 27.47ab | 3.04 | 25.76a | 2.41 | 23.48abc | 7.58 | 22.93ab | 7.77 | |
| 1.00 | 31.3abc | 12.97 | 24.19abcd | 14.61 | 19.92b | 24.54 | 19.11bcd | 24.78 | 20.20abc | 18.77 | |
| 2.00 | 26.4cdefg | 26.37 | 20.31defg | 28.33 | 16.72bcde | 36.64 | 16.37defg | 35.57 | 17.13bcde | 31.10 | |
| 4.00 | 19.4hijk | 46.04 | 15.89fghi | 43.92 | 15.70bcdef | 40.53 | 13.66defgh | 46.26 | 13.28def | 46.60 | |
| brown | Control | 27.08cdefg | 0.00 | 20.83cdefg | 0.00 | 19.52b | 0.00 | 18.74cd | 0.00 | 17.73bcd | 0.00 |
| 0.50 | 25.97cdefg | 4.07 | 19.83defgh | 4.80 | 18.70bc | 4.19 | 17.30def | 7.71 | 16.60bcde | 6.39 | |
| 1.00 | 24.6defgh | 8.90 | 19.44defgh | 6.67 | 16.81bcde | 13.60 | 16.07defg | 14.23 | 15.53cde | 12.41 | |
| 2.00 | 20.7ghij | 23.30 | 15.47ghi | 25.73 | 13.17cdef | 32.53 | 12.59fgh | 32.81 | 13.33def | 24.81 | |
| 4.00 | 16.18ijk | 40.25 | 14.06hi | 32.53 | 11.24efg | 42.39 | 10.97gh | 41.48 | 11.06ef | 37.66 | |
| white | Control | 30.54abcd | 0.00 | 26.64abc | 0.00 | 26.18a | 0.00 | 24.48ab | 0.00 | 22.31ab | 0.00 |
| .50 | 28.7bcde | 5.71 | 22.61abcde | 15.12 | 21.12ab | 19.33 | 18.42cde | 24.75 | 20.53abc | 7.97 | |
| 1.00 | 27.72cdef | 9.24 | 21.28cdefg | 20.13 | 18.42bc | 29.66 | 17.33def | 29.20 | 17.40bcd | 22.02 | |
| 2.00 | 22.26fghi | 27.12 | 16.75efghi | 37.12 | 13.83cdef | 47.16 | 13.59defgh | 44.48 | 14.53cde | 34.86 | |
| 4.00 | 14.62jk | 52.14 | 14.03hi | 47.34 | 12.67def | 51.62 | 13.13efgh | 46.35 | 12.39def | 44.48 | |
| white-vestigial | Control | 23.10efgh | 0.00 | 21.85bcdef | 0.00 | 19.70b | 0.00 | 18.56cde | 0.00 | 16.93bcde | 0.00 |
| 0.50 | 21.54fghi | 6.77 | 20.22defg | 7.46 | 18.29bcd | 7.14 | 16.97def | 8.57 | 15.88cde | 6.21 | |
| 1.00 | 20.92ghij | 9.43 | 18.89defgh | 13.56 | 16.42bcde | 16.62 | 15.56defg | 16.17 | 14.13cde | 16.54 | |
| 2.00 | 14.13k | 38.85 | 11.19ij | 48.77 | 10.48fg | 46.77 | 8.93hi | 51.90 | 7.47fg | 55.91 | |
| 4.00 | 7.41ll | 67.92 | 5.65j | 74.16 | 5.82g | 70.46 | 4.69i | 74.73 | 3.59g | 78.81 | |
*Superscript letters are assigned based on the results of the Tukey pairwise comparison test. indicating the significance levels of each column value obtained relative to the copper concentration and exposure time. The highest-ranked values are denoted. beginning with the first letter “a”. Similar superscript letters indicate statistically non-significant differences between the obtained values (p > 0.05). **IC denotes prolificity inhibition concentration of lead.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



