
Submitted:
04 March 2024
Posted:
05 March 2024
You are already at the latest version
Abstract
Microalgae are highly attractive to various industrial sectors and can serve as indicators of aquatic environmental health. An essential step for cultivating microalgae is assessing the cell density, which is traditionally performed through viable cell counting by optical microscopy (OM). However, this method has some limitations, such as the time involved in the process of counting, and the need for specialized training of laboratory personnel. Spectrophotometry is an affordable alternative, especially when applied to culture growth and toxicological tests. We chose the wavelength of 680 nm to detect the optical density (OD) of the green freshwater microalga Desmodesmus subspicatus and green the marine microalga Tetraselmis sp. The correlation between the number of cells and OD was suitable for both microalgae and accuracy was comparable to traditional OM counting. In toxicological tests, the OD technique allowed the monitoring of microalgal growth in the presence of the chemicals potassium dichromate, potassium chloride and sodium dodecyl sulfate (SDS), with results very close to those reported in the scientific literature. This study sheds light on this well-known technique, which is an important contribution since few articles have explored its potentialities and fragilities, showing the successful application to monitoring green microalgal cultures and toxicological assays.
Keywords:
Spectrophotometry
; Desmodesmus subspicatus
; Tetraselmis sp.
; microalgal culture
; optical density
; optical microscope counting
; toxicity test.
1. Introduction
Microalgae are a heterogeneous group of cosmopolitan eukaryotic and prokaryotic cellular organisms, ranging from 2 to 200 µm [1]. In general, they are organisms of great ecological importance since they are responsible for primary production, nutrient cycling, and energy flow [2,3,4]. Besides their important role in nature, over the years microalgal cultivation has been increasingly explored due to its attractive characteristics. Microalgae have relatively fast growth, including in culture media composed of low-cost nutrients, without the need for dry and arable land with sufficient water and space in comparison with plants, resulting in high biomass production when the cultures are in favorable conditions [5].
Biomass composition varies according to the species of the cultivated microalga, a feature that is of strong economic interest in the biotechnology field [6]. This interest is due to the possibility of controlling microalgal cultures for commercial applications such as production of fertilizers, fuels, high-value chemicals, livestock feed, aquaculture inputs, bioactive compounds for human dietary supplementation (nutraceuticals), antioxidants, and dyes [3,6,7,8].
Since microalgae are fast-growing organisms that are highly sensitive to biotic and abiotic changes, and the presence of contaminants, they are increasingly applied as test organisms in toxicity assays [9,10]. Moreover, because they are abundantly present in a wide range of ecosystems and their ecological importance, microalgae are promising organisms to indicate the health of ecosystems, in particular aquatic and marine ones [4]. Ecotoxicological assays have gained more importance and attention in recent years given the need to understand the impacts caused on the environment by human activities [11]. Marine ecosystems are among the most affected since they are the final destination of several contaminants, especially those discharged by industrial and residential effluents [12,13].
When applying microalgae as test organisms in toxicological tests, it is possible to investigate the acute, subacute or chronic effects of the tested chemical substances, with acute and chronic toxicity assays being used the most [14]. The tests are performed by exposing a microalgal culture to a contaminant at different concentrations and comparing the decrease in cell density during an exposure time, which can vary between 6 and 96 hours for algae [15], allowing plotting a dose-response curve. Usually, due to the several concentrations tested, and the requisite replicates, toxicological tests are performed with many samples, which need to be analyzed during the respective exposure time.
Assessing the cell density and culture growth is an essential step, both for microalgal cultivation for biotechnological proposes and for the assessment of toxic effects in ecotoxicity assays. Among the best-established methods to quantify cell density, dry biomass measurement and viable cell counting stand out [16]. In general, the most used method is viable cell counting. It is relatively inexpensive since it only requires an optical microscope and a counting chamber, such as a hemocytometer [17,18]. However, this method often is inefficient because it can be influenced by several factors, such as the need for manipulation, the long time spent and low reproducibility, besides the need for trained professionals.
Some techniques can be used to automate the cell counting process, such as flow cytometry and the Coulter counter system [19,20,21,22]. However, these techniques are expensive and not accessible to most laboratories [20]. For this reason, further studies need to be conducted to investigate more accessible methods with high efficiency and speed, with the ability to quantify the cell density of microalgae.
Among the techniques already used in the laboratory, a possible alternative is the use of optical devices like fluorimeters and ultraviolet-visible (UV-Vis) spectrophotometers [23]. In particular, the UV-Vis technique allows quantification through the detection of optical density (OD). The results are generally cost-effective, accurate, reproducible and obtained quickly, making the method popular [24,25,26,27]. This specific technique thus is an attractive alternative to correlate indirectly the cell density of microalgae with the absorption at specific wavelengths in a reliable way [11,28].
Therefore, this study was developed to assess the effectiveness of using OD detected by UV-Vis spectrophotometry to correlate the cell density of microalgae cultivation, using two species of green microalgae of the phylum Chlorophyta: Desmodesmus subspicatus and Tetraselmis sp., which are abundant in freshwater and marine ecosystems, respectively [29]. For this, a regression model was standardized to estimate the cell density of both microalgae cultivated in the laboratory, by measuring the spectrophotometric absorbance of culture suspensions, compared to traditional cell counting by optical microscopy (OM). To ensure this method's applicability, we also used it to measure the cell density in ecotoxicological assays using referenced chemicals (K2Cr2O7, KCl, and SDS), recommended by standard guides as highly toxic in microalgal assays.
2. Materials and Methods
2.1. Microalgae Strains, Culture Conditions, and Growth
The freshwater microalga D. subspicatus was kindly provided by the University of the Joinville Region (UNIVILLE), Brazil, and Federal University of Santa Catarina (UFSC), Brazil. Microalgal culture and maintenance were performed in CHU medium [30] according to ABNT (Brazilian Association of Technical Standards) protocol NBR 12648 [31]. The marine microalga Tetraselmis sp., strain CCMP908 (Bigelow Laboratory for Ocean Sciences) was kindly provided by the Engineering and Malacoculture Laboratory (LEMAQUI) of Federal University of Paraná (UFPR), located in Pontal do Paraná, Paraná, Brazil. Microalgal culture and maintenance were performed in Conway medium [32] according to ABNT protocol NBR 16181 [33]. Both microalgal cultures were kept at controlled temperature (20 ± 2 °C), illumination (500 lux), and aeration. The cultures were renewed weekly with the addition of 80% sterile medium. To assess D. subspicatus and Tetraselmis sp. growth, 300 mL of the culture of both microalgae was maintained under the described conditions, and growth parameters (cell density and pH) were recorded daily until cultures reached the stationary phase.
2.2. Cellular Density Determination
The cell density of D. subspicatus and Tetraselmis sp. was determined by indirectly estimating the number of cells by the optical density (OD) of the microalgal cultures, through spectrophotometry (IL-593-S, Kasuaki, China). To ensure the appropriate wavelengths for the analysis, a wavelength scan was carried out ranging from 300 to 800 nm with both microalgae in the exponential growth phase. For the indirect quantification, different dilutions of the microalgae culture samples in the exponential phase were performed, from which we measured the number of cells via OM cell counting using a Neubauer chamber, and determined OD by spectrophotometry. Both cell numbers and OD were correlated in a calibration curve to obtain equations by the linear regression model (Equation 1), necessary to infer the cell number of culture samples using only spectrophotometry.
Cell density (cells/mL) = a + b * ODXXX.
2.3. Toxicity Tests
Chronic toxicity tests were performed with both microalgae using the reference substances K2Cr2O7, KCl, and SDS, which are already known as reference chemicals due to their toxicity. Tests were developed according to the guidelines of ABNT [31,33]. The chemicals K2Cr2O7 and SDS were dissolved in stock solutions of 10 mg/L in DIN medium (the medium recommended by ABNT for toxicity tests) for D. subspicatus tests and Conway medium for Tetraselmis sp. tests. The exposures were performed at the concentrations of 0.1, 1, 10, 100, and 1,000 mg/L for K2Cr2O7 and SDS. For the KCl tests, we prepared a stock solution of 100 mg/L in DIN medium for D. subspicatus, and performed at concentrations of 1, 10, 100, 1,000, and 10,000 mg/L. For the test with Tetraselmis sp., KCl was dissolved in ultrapure water at the concentration of 300 mg/L, and the exposure concentrations were 10, 100, 1,000, 10,000, and 100,000 mg/L. Negative controls were also used for each independent assay, to assess proper microalgal growth.
Tests were performed in triplicates at the initial cell concentration of 2.5x105 cells/mL, in the final volume of 20 mL, and kept in a shaker incubator (LT 600/2, LimaTec, Brazil) in the controlled conditions of 20±2 °C, 150 rpm, and 500±50 lux for 96 h. Cell density was measured every 24 hours by OD. Test acceptability criteria for OD measurement were the average OD680nm reading ranging >0.10 and <1.8 for the controls and treatment. The cell suspensions were diluted in culture media when extrapolating the absorbance of 1.8, and R2 of more than 0.8, and the best fit (highest R2 value among the exposure times) in linear regression for EC50 (half maximal effective concentration) calculation. Results were submitted to analysis of variance (ANOVA) with the RStudio software to evaluate significant differences between the cell density of the control and reference substances, allowing the definition of the values of NOEC (no observed effect concentration) and LOEC (lowest observed effect concentration). The EC50 values were calculated by the software GraphPad Prism 8.
3. Results and Discussion
3.1. Relationship between Numbers of Cells per mL and Optical Density
The absorption spectra of both microalgal cultures in the range of 300 to 800 nm are presented in Figure 1, showing several absorbance peaks, mainly from 400 to 525 nm and between 625 and 715 nm. Absorption peaks were observed for both D. subspicatus and Tetraselmis sp. in these specific ranges (Figure 1a), which are two wavelength regions related to light absorption patterns of the chlorophyll (Chl) present in both microalgae. Analysis of the absorption spectra of microalgae show absorbance peaks according to the composition of cell pigments [34].
It is well known that photosynthetic organisms have accessory pigments, which are present in the chloroplast and are essential for the photosynthesis process since they are responsible for light capture [35]. Among the accessory pigments, the most important are Chl, carotenoid, and phycobilin [35]. Besides capturing light, these pigments also impart specific colors to the organisms. Chl has different chemical structures, but all organisms that carry out oxygenic photosynthesis present Chl a [36].
Each Chl structure absorbs light and is detected at different wavelengths. The form and/or concentration of each Chl and other accessory pigment varies according to the genera and species, among plants and algae. In the case of green microalgae, Chl a and b are the main pigments present [37]. Chl a absorbs red and blue light at approximately 660 and 435 nm, respectively, and Chl b absorbs orange and blue light at 642 and 453 nm, respectively [35,36].
In this study, we chose the wavelength of 680 nm to detect the microalgal density, since it presented a clear peak, without interference from other peaks related to several cell compounds. The linear regression model was used to determine the relationship between OD680nm and density of cells per mL of both microalgae, which denoted significant (p < 0.001) and positive correlations for D. subspicatus (R2 = 0.992) and for Tetraselmis sp. (R2 = 0.987) (Table 1). Equation 2 and Equation 3, obtained with data from the two grouped microalgal cultures, are efficient for estimating the relationship between the two measurements for D. subspicatus and Tetraselmis sp., respectively.
OD680nm = 7.354 x 10-02 + 1.313 x 10-07 x Cell Density (cells/mL)
OD680nm = 1.771 x 10-02 + 3.151 x 10-07 x Cell Density (cells/mL)
In Figure 2, it is possible to visualize the growth curves from the data obtained from cell density by cell counting by OM and by OD measurements from spectrophotometry for D. subspicatus (Figure 2a) and Tetraselmis sp. (Figure 2b). Overall, both microalgae presented regular growth, showing typical phases of microalgal growth [38]. D. subspicatus (freshwater) and Tetraselmis sp. (marine) presented a two-day lag phase, and marked exponential phases (log phases) of three days for D. subspicatus and two days for Tetraselmis sp., corroborated by a pH increase. The pH level changes during the microalgal growth phases. The pH values in culture media remained stable for both microalgae during the lag phases, and increased during the log phase, explained by CO2 production from algal biomass.
Although the growth curves were similar when using the two different techniques to measure cell density, it was possible to notice some slight differences, as indicated by the error bars. These minor differences may have been caused by methodological inaccuracies in the OM counting technique, since it is done manually and can result in different values for each observer, depending on their handling of the samples. Also, despite the regular growth of both microalgae, it is important to mention the difference between cell density in growth cultures of Tetraselmis sp. and D. subspicatus (10 times more abundant) in the entire experiment. This behavior is explained by the size of these microorganisms: while the size of Tetraselmis sp. ranges from 10–25 µm long and 7-20 µm wide [39], D. subspicatus has roughly half that size, ranging from 5-13 μm long and 3–7 µm wide [40]. This cell size difference can be visualized in Figure 1b and Figure 1c.
Additionally, we estimated the specific growth rate for the microalgal cultures, indicating the relative increase in culture density over a short time. These values are listed in Table 2. In the exponential growth phase, the growth rate is described quantitatively by a simple kinetic differential equation and is therefore constant [41]. Using the initial and final cell density of the culture, as B0 and Bf respectively, for the initial and final time of this growth phase (T0 and Tf, respectively), allows obtaining the exponential growth correlation. The slope of the linear regression corresponding to the specific microalgal culture growth rate, μ (d-1), estimated in this study for both tested techniques from Equation 4, is described below:
The specific growth rates found were very close between counting techniques for the same microalga. Specific growth rate values higher than 1 indicate intense and fast growth of the culture, with more divisions per day and a faster rate of population doubling. Overall, both tested techniques for measuring the cell density of microalgae presented corroborating high values of specific growth and fast doubling time, indicating ideal cultivation conditions. In particular, the OD technique showed slightly lower values in comparison with the OM counting technique. High specific growth rates indicate a well-established culture in terms of nutrient availability and light intensity [42]; marine microalgae are also affected by salinity [43].
During the process of microalgal growth, the culture reaches dynamic equilibrium, so the culture density, concentration of cells and cell components, and their proportions are stabilized. This is known as the stationary phase. Both growth curves reached the stationary phase between the 7th and 8th day of monitoring, indicating stabilization of microalgal growth. For the marine microalga Tetraselmis sp., a decrease of pH was observed in the stationary phase (Figure 2b), while for freshwater microalga D. subspicatus, the pH stabilized at the beginning of the stationary phase (Figure 2a).
Both tested techniques (microscope counting and spectrophotometry) presented comparable data from monitoring the cell density of microalgal growth, demonstrating it is possible to track microalgal culture growth through OD measurement (Figure 2), with similar accuracy to the OM counting technique. Our result also indicates that the cell density of microalgae measured by OD did not have discrepancies in comparison with the OM technique during the different growth phases.
In the literature, other authors have reported similar wavelengths (677, 682, 684, and 688 nm) to measure cell density of different microalgal species, also showing good linear correlation fit [28,44]. In the case of applying this technique for toxicity tests, it is important to ensure that the contaminant does not interfere with the signals at 680 nm. Some contaminants are colorful or absorb at wavelengths close to those of Chl. In this case, we recommend scanning the contaminant of interest in a spectrophotometer in the range of 300-800 nm to verify there is no interference of the curves. If the contaminant has a peak close to 680 nm, then we recommend using other wavelengths related to Chl pigments (as observed in the scanning of microalgae in Figure 1) that have no interference with other chemical absorbances.
From this optical method of cell density measurement, we also performed toxicity tests with referenced chemicals to validate the method, comparing the toxicity results to the values reported in the literature. In the next phase, we used the cell density estimated by spectrophotometry to calculate important toxicological parameters, such as NOEC, LOEC, and EC50 [45].
3.2. Toxicity of K2Cr2O7 in Green Microalgae
From the data obtained in the toxicity assays performed with K2Cr2O7, it was possible to observe its toxic effects for both microalgae (Figure 3a and Figure 3c), with good fits of dose-response curves (Figure 3b and Figure 3d). For D. subspicatus, the toxic effect was significant at 24 and 48 h of exposure to the chemical (Figure 3a), where all tested concentrations were associated with a decrease in cell density when compared to the respective control. At exposure for 72 h, a significant decrease in cell density was observed above 10 mg/L. Therefore, for the D. subspicatus assay, the NOEC and LOEC were defined as 1 and 10 mg/L K2Cr2O7, respectively. At 72 h of exposure, the best fit of the dose-response curve was verified, resulting in EC50 of 8.1 mg/L (R2 = 0.9120) (Figure 3b, Table 3).
In Tetraselmis sp. exposure, the most toxic effects of K2Cr2O7 were significant after 48 hours of exposure, which remained throughout the assay (Figure 3c) in comparison with the control test. At the highest concentrations of K2Cr2O7, a visible decrease in microalgal growth was noted, while at lower concentrations (0.1 and 1 mg/L), there was growth stimulation at all exposure times, where the cell density achieved higher levels than the control. Due to practical issues, we did not consider this growth stimulation effect to determine NOEC and LOEC values, since we were considering only the toxic effects of decreasing cell density. For Tetraselmis sp., the EC50 was considered as 44.5 mg/L at 48 hours (best fit; R2 = 0.8967). NOEC and LOEC values were 10 and 100 mg/L, respectively, in 48 hours (Figure 3c, Table 3).
K2Cr2O7 is known as a toxic chemical, due to hexavalent chromium in its composition. This is cause for concern when it is present in water bodies, since it is highly toxic and carcinogenic to humans and aquatic organisms [46,47]. However, Cr is a widely used substance in different industrial processes [48]. Therefore, this chemical is constantly being dispersed in the various environmental compartments. Toxicity of K2Cr2O7 to Tetraselmis suecica was reported in the literature with EC50 of 8.89 mg/L at 72 h of exposition [49], a value below the toxicity results found in our experiments. Toxicity of K2Cr2O7 to D. subspicatus has been reported in the literature as EC50 of 0.357 mg/L [50], very close to the value obtained in our assay after exposure for 24 hours (EC50 of 0.422 mg/L; R2 = 0.8058), demonstrating the accuracy of the OD technique and its reliable results in toxicity assays. Other authors have also reported EC50 of K2Cr2O7 to D. subspicatus at 72 h using OM and OD techniques, obtaining values of 0.71 mg/L and 1.10 mg/L, respectively [51].
The results obtained in the toxicological assays performed with K2Cr2O7 indicated this chemical was toxic to both tested green microalgae, even in short exposures (24 and 48 h). In long exposure, low concentrations of K2Cr2O7 induced greater microalgal growth, probably due to a cell response to the stress induced by the substance. This trend was clearly noted at all exposure times of Tetraselmis sp., and slightly visible in the D. subspicatus assay (72 and 96 h), which was not confirmed by statistical difference when compared to the control test (p < 0.05). At higher concentrations, K2Cr2O7 drastically reduced the microalgal growth of both species. Comparing both microalgae, it can be observed that Tetraselmis sp. is more resistant to K2Cr2O7 than D. subspicatus. Both marine organisms presented high resistance when exposed to the same chemical in comparisons with freshwater organisms. This fact is easily ascertained by the difference in EC50 values, which were much higher for Tetraselmis sp. (44.5 mg/L) than for D. subspicatus (1.08 mg/L).
3.3. Toxicity of KCl to Green Microalgae
The toxicity tests using KCl and the OD technique to measure cell density showed evident toxic effects, mainly for D. subspicatus. The results of D. subspicatus exposed to KCl indicated toxic effects at all tested concentrations from 24-hour exposure, except for the 100 mg/L concentration (Figure 4a and Figure 4b), which showed inconsistent toxic effects and was excluded to calculate the EC50. No adjustments were observed in the exposure data of D. subspicatus in the dose-response curves. The chronic results at 48 hours indicated a significant decrease in cell density, enabling defining the LOEC as 1 mg/L when compared to the control test, while the calculated EC50 was 19.57 mg/L (R2 = 0.2277) (Table 3, Figure 4b). The large standard deviation observed in the exposures inhibited the calculation of an EC50 with adequate R2 value. Further studies need to be conducted, testing lower concentrations than 1 mg/L.
Tetraselmis sp. exposed to KCl also resulted in toxic effects at all tested concentrations from the 24-hour exposure period (Figure 4c and Figure 4d). In this test, the greatest toxic effects based on decrease in cell density were observed at a KCl concentration of 100 mg/L when compared to the control test. Good fit of dose-response curves was verified at 48 and 72 hours. Based on the best fit at 72 hours, the NOEC and LOEC values were 1,000 and 10,000 mg/L, respectively, and the calculated EC50 was 30,908 mg/L (R2 = 0.9774) (Table 3, Figure 4).
Potassium is an essential macronutrient for microalgal growth since it is important for osmotic and ionic regulation in cells, nutrient transport, and enzyme activation [52]. However, prolonged exposure to excessive concentrations of potassium can lead to ionic and osmotic imbalance, which can negatively affect the cellular structure of microalgae, also preventing their replication. The chlorine present in KCl can also be toxic to microalgae, but its concentration is relatively low. In our study, high concentrations of KCl were toxic to both D. subspicatus and Tetraselmis sp. (Figure 4). In another study, the authors observed that excess potassium was able to reduce the growth of the cyanobacterium Microcystis aeruginosa by 50% [53].
3.4. Toxicity of SDS to Green Microalgae
Examination of the exposure of D. subspicatus to SDS using the OD technique to measure cell density revealed a chemical reaction between microalgae in suspension and SDS, inducing formation of turbidity in the exposure flasks. This prevented measuring cell density in this experiment, since turbidity does not allow performing OD measurement of the cell suspension to estimate toxicity parameters like EC50, NOEC, and LOEC.
From the data obtained from the toxicity tests of Tetraselmis sp. exposed to SDS using the OD technique, it was possible to observe toxic effects at 24 hours at the concentration of 10 mg/L (Figure 5). With time, the decrease of cell density of Tetraselmis sp. was observed for the highest concentrations (100 and 1,000 mg/L), with a significant difference when compared to the control test. From this, the estimated NOEC was 10 mg/L, and LOEC was 100 mg/L. The EC50 value calculated at 72 hours was 41.6 mg/L (R2 = 0.9564) (Table 3, Figure 5).
SDS is a chemical that readily interacts with cell membranes and contents. In microalgae, the cell membranes are mainly composed of phospholipids, and contact with SDS can break these structures, negatively impacting cell survival [54]. This explains why this chemical was toxic to Tetraselmis sp. (Figure 5). Other studies also suggest a toxic effect of SDS on other marine microalgal species, such as Dunaliella salina and Chroomonas salina [55,56]. The destruction of cell membrane can also allow the release of intracellular compounds into the external environment.
In the case of D. subspicatus, which is well known for its lipid production, the disruption of the cell membrane may have released the intracellular lipids, which in contact with the external SDS may have caused the turbidity observed in the flasks. In some cases, SDS in low concentrations has been reported to stimulate the growth of different species of microalgae, since it can be used as a carbon source [55,56]. Therefore, this may also have contributed to the turbidity formation when this microalga was exposed to SDS. This result reinforces that the use of the OD technique in toxicity tests is not recommended for all chemicals, since some chemicals can cause interference when absorbing at the same wavelength used to measure cell density. In this specific situation, the OM technique is more appropriate. Additionally, this situation indicates that further investigations need to be carried out to improve the use of optical determination of D. subspicatus density in toxicity tests, especially when the investigated chemical substance has some kind of interaction with the microalgae or the culture medium, as was observed in our study with SDS. This recommendation is also valid for other substances that contain pigments that can influence the use of spectrophotometers.
4. Conclusions
The use of spectrophotometry proved to be an accurate and fast alternative to determine the density of microalgal cells, especially when compared to viable cell counting method, which requires a lot of time and training, as well as having low replicability. As observed, the number of cells per mL of both D. subspicatus and Tetraselmis sp. corresponded to the absorbance at OD680nm during all growth phases, which indicates that the Chl during all microbial growth phases was stable and well detected by spectrophotometry.
The optical determination of cell density of microalgae by the OD technique was suitable for monitoring cell density of green microalgae in cultures and toxicity tests. It was possible to quickly monitor the exposures every 24 h, and using the equations, to obtain the number of cells of each assay, and with this determine the cell viability of microalgae at each concentration of contaminants. Furthermore, the cell density results were not affected at different times of exposure to the investigated chemicals.
The practical application of this method was successful in the toxicological assays with both species of green microalgae, using K2Cr2O7, KCl, and SDS, reference substances known for their deleterious effects on microalgae. According to the results presented here, the method is efficient in detecting the influence of toxic substances on microalgal growth, as well as allowing the calculation of important parameters in microalgal cultures such as specific growth rates and doubling time with accuracy.
The particularities observed in the test with D. subspicatus and SDS, however, demonstrate that this technique may not be suitable for all chemicals, since optical interferences like turbidity and color of culture media need to be considered as limitations. This fact also illustrates the need to evaluate other conditions and characteristics of the studied substances, since these can influence the absorbance measurements, resulting in misinterpretation of toxicity tests.
Author Contributions
J.S.P.: Conceptualization; Methodology; Investigation; Data curation; Formal analysis; Validation; Writing - original draft preparation, review & editing. A.C.O.A.: Conceptualization; Methodology; Data curation; Formal analysis; Writing - original draft preparation. L.F.S. Data curation; Formal analysis. T.M.N.O.: Visualization; Resources. C.C.M.: Funding acquisition; Project administration; Writing - review & editing. W.G.M.: Funding acquisition; Project administration; Writing - review & editing. S.P.M.: Conceptualization; Methodology; Funding acquisition; Project administration; Supervision; Writing - original draft preparation, ; Writing - review & editing.
Funding
This work was funded in part by the CNPq [grant number 552112/2011-9] and CAPES/PNPD [grant number 2982/10]. This work is part of the EQCEP project (Historical input and future perspectives related to the occurrence of chemical stressors in the Paranaguá Estuarine System) funded by the CNPq [grant number 441265/2017-0], coordinated by Professor M. M. Mahiques.
Acknowledgments
We gratefully acknowledge the Engineering and Malacoculture Laboratory (LEMAQUI – UFPR), University of the Region of Joinville (UNIVILLE), and Federal University of Santa Catarina (UFSC) for kindly providing the microalgal strains.
Conflicts of Interest
All authors have no conflicts of interest.
References
- Hemaiswarya, S.; Raja, R.; Ravikumar, R.; Carvalho, I. Microalgae Taxonomy and Breeding. In Biofuel Crops: Production, Physiology and Genetics; CABI, 2013; pp. 44–53.
- Thoré, E.S.J.; Muylaert, K.; Bertram, M.G.; Brodin, T. Microalgae. Current Biology 2023, 33, R91–R95. [Google Scholar] [CrossRef] [PubMed]
- Singh, J.; Saxena, R.C. An Introduction to Microalgae: Diversity and Significance. Handbook of Marine Microalgae 2015, 11–24. [Google Scholar] [CrossRef]
- Lu, T.; Zhang, Q.; Zhang, Z.; Hu, B.; Chen, J.; Chen, J.; Qian, H. Pollutant Toxicology with Respect to Microalgae and Cyanobacteria. Journal of Environmental Sciences 2021, 99, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Tan, J. Sen; Lee, S.Y.; Chew, K.W.; Lam, M.K.; Lim, J.W.; Ho, S.-H.; Show, P.L. A Review on Microalgae Cultivation and Harvesting, and Their Biomass Extraction Processing Using Ionic Liquids. Bioengineered 2020, 11, 116–129. [Google Scholar] [CrossRef] [PubMed]
- Pulz, O.; Gross, W. Valuable Products from Biotechnology of Microalgae. Appl Microbiol Biotechnol 2004, 65, 635–648. [Google Scholar] [CrossRef] [PubMed]
- Becker, E.W. Micro-Algae as a Source of Protein. Biotechnol Adv 2007, 25, 207–210. [Google Scholar] [CrossRef] [PubMed]
- Torres-Tiji, Y.; Fields, F.J.; Mayfield, S.P. Microalgae as a Future Food Source. Biotechnol Adv 2020, 41, 107536. [Google Scholar] [CrossRef] [PubMed]
- Stone, S.; Adams, M.S.; Stauber, J.L.; Jolley, D.F.; Warne, M.S.J. Development and Application of a Multispecies Toxicity Test with Tropical Freshwater Microalgae. Environmental Pollution 2019, 250, 97–106. [Google Scholar] [CrossRef]
- Silva, A.; Figueiredo, S.A.; Sales, M.G.; Delerue-Matos, C. Ecotoxicity Tests Using the Green Algae Chlorella Vulgaris—A Useful Tool in Hazardous Effluents Management. J Hazard Mater 2009, 167, 179–185. [Google Scholar] [CrossRef]
- Boros, B.-V.; Ostafe, V. Evaluation of Ecotoxicology Assessment Methods of Nanomaterials and Their Effects. Nanomaterials 2020, 10, 610. [Google Scholar] [CrossRef]
- Wilkinson, J.; Hooda, P.S.; Barker, J.; Barton, S.; Swinden, J. Occurrence, Fate and Transformation of Emerging Contaminants in Water: An Overarching Review of the Field. Environmental Pollution 2017, 231, 954–970. [Google Scholar] [CrossRef]
- Schwarzenbach, R.P.; Escher, B.I.; Fenner, K.; Hofstetter, T.B.; Johnson, C.A.; von Gunten, U.; Wehrli, B. The Challenge of Micropollutants in Aquatic Systems. Science (1979) 2006, 313, 1072–1077. [Google Scholar] [CrossRef]
- Ramakrishnan, B.; Megharaj, M.; Venkateswarlu, K.; Naidu, R.; Sethunathan, N. The Impacts of Environmental Pollutants on Microalgae and Cyanobacteria. Crit Rev Environ Sci Technol 2010, 40, 699–821. [Google Scholar] [CrossRef]
- Franklin, N.M.; Stauber, J.L.; Apte, S.C.; Lim, R.P. Effect of Initial Cell Density on the Bioavailability and Toxicity of Copper in Microalgal Bioassays. Environ Toxicol Chem 2002, 21, 742. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.-K.; Shen, H. Basic Culturing Techniques. In Handbook of Microalgal Culture; Blackwell Publishing Ltd: Oxford, UK, 2003; pp. 40–56. [Google Scholar]
- Harris, E.H.; Stern, D.B.; Witman, G.B. Chlamydomonas in the Laboratory. In The Chlamydomonas Sourcebook; Harris, E.H., Stern, D.B., Witman, G.B., Eds.; Academic Press: London, 2009; pp. 241–302. ISBN 978-0-12-370873-1. [Google Scholar]
- Prieto, B.; Silva, B.; Lantes, O. Biofilm Quantification on Stone Surfaces: Comparison of Various Methods. Science of The Total Environment 2004, 333, 1–7. [Google Scholar] [CrossRef]
- Prado, R.; García, R.; Rioboo, C.; Herrero, C.; Cid, Á. Suitability of Cytotoxicity Endpoints and Test Microalgal Species to Disclose the Toxic Effect of Common Aquatic Pollutants. Ecotoxicol Environ Saf 2015, 114, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Sarrafzadeh, M.H.; La, H.-J.; Seo, S.-H.; Asgharnejad, H.; Oh, H.-M. Evaluation of Various Techniques for Microalgal Biomass Quantification. J Biotechnol 2015, 216, 90–97. [Google Scholar] [CrossRef]
- Stauber, J.L.; Franklin, N.M.; Adams, M.S. Applications of Flow Cytometry to Ecotoxicity Testing Using Microalgae. Trends Biotechnol 2002, 20, 141–143. [Google Scholar] [CrossRef] [PubMed]
- Hyka, P.; Lickova, S.; Přibyl, P.; Melzoch, K.; Kovar, K. Flow Cytometry for the Development of Biotechnological Processes with Microalgae. Biotechnol Adv 2013, 31, 2–16. [Google Scholar] [CrossRef]
- Zhou, F.; Li, C.; Zhu, H.; Li, Y. Determination of Trace Ions of Cobalt and Copper by UV–Vis Spectrometry in Purification Process of Zinc Hydrometallurgy. Optik (Stuttg) 2019, 184, 227–233. [Google Scholar] [CrossRef]
- Parmar, A.; Sharma, S. Derivative UV-Vis Absorption Spectra as an Invigorated Spectrophotometric Method for Spectral Resolution and Quantitative Analysis: Theoretical Aspects and Analytical Applications: A Review. TrAC Trends in Analytical Chemistry 2016, 77, 44–53. [Google Scholar] [CrossRef]
- Dankowska, A.; Domagała, A.; Kowalewski, W. Quantification of Coffea Arabica and Coffea Canephora Var. Robusta Concentration in Blends by Means of Synchronous Fluorescence and UV-Vis Spectroscopies. Talanta 2017, 172, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, S.L.; Hansen, B.W. Evaluation of the Robustness of Optical Density as a Tool for Estimation of Biomass in Microalgal Cultivation: The Effects of Growth Conditions and Physiological State. Aquac Res 2019, 50, 2698–2706. [Google Scholar] [CrossRef]
- Sohrabi, M.R.; Mirzabeygi, V.; Davallo, M. Use of Continuous Wavelet Transform Approach for Simultaneous Quantitative Determination of Multicomponent Mixture by UV–Vis Spectrophotometry. Spectrochim Acta A Mol Biomol Spectrosc 2018, 201, 306–314. [Google Scholar] [CrossRef] [PubMed]
- Lúcia, H.R.R.; Alexandre, A.; Maria, T.R.-R.; Nelson, F.F. Algal Density Assessed by Spectrophotometry: A Calibration Curve for the Unicellular Algae Pseudokirchneriella Subcapitata. Journal of Environmental Chemistry and Ecotoxicology 2011, 3, 225–228. [Google Scholar] [CrossRef]
- Metting, F.B. Biodiversity and Application of Microalgae. J Ind Microbiol Biotechnol 1996, 17, 477–489. [Google Scholar] [CrossRef]
- Chu, S.P. The Influence of the Mineral Composition of the Medium on the Growth of Planktonic Algae: Part I. Methods and Culture Media. J Ecol 1942, 30, 284. [Google Scholar] [CrossRef]
- ABNT ABNT NBR 12648:2018 - Ecotoxicologia Aquática - Toxicidade Crônica - Método de Ensaio Com Algas (Chlorophyceae) 2018.
- Tompkins, J.; DeVille, M.; Day, J.; Turner, M. Culture Collection of Algae and Protozoa: Catalogue of Strains; Ambleside, UK, 1995. [Google Scholar]
- ABNT ABNT NBR 16181:2021 - Ecotoxicologia Aquática - Toxicidade Crônica - Método de Ensaio Com Microalgas Marinhas 2021.
- Zhang, T.; Tai, F.; Hu, L.; Chen, S. Method for Extracting Pigment Characteristic Spectra from the Phytoplankton Absorption Spectrum. Opt Express 2023, 31, 22233. [Google Scholar] [CrossRef] [PubMed]
- Häder, D.-P. Photosynthesis in Plants and Algae. Anticancer Res 2022, 42, 5035–5041. [Google Scholar] [CrossRef]
- Bhuvana, P.; Sangeetha, P.; Anuradha, V.; Ali, M.S. Spectral Characterization of Bioactive Compounds from Microalgae: N. Oculata and C. Vulgaris. Biocatal Agric Biotechnol 2019, 19, 101094. [Google Scholar] [CrossRef]
- da Silva, J.C.; Lombardi, A.T. Chlorophylls in Microalgae: Occurrence, Distribution, and Biosynthesis. In Pigments from Microalgae Handbook; Jacob-Lopes, E., Queiroz, M.I., Zepka, L.Q., Eds.; Springer International Publishing: Cham, 2020; pp. 1–18. ISBN 978-3-030-50971-2. [Google Scholar]
- Lee, E.; Jalalizadeh, M.; Zhang, Q. Growth Kinetic Models for Microalgae Cultivation: A Review. Algal Res 2015, 12, 497–512. [Google Scholar] [CrossRef]
- Arora, M.; Anil, A.C.; Leliaert, F.; Delany, J.; Mesbahi, E. Tetraselmis Indica (Chlorodendrophyceae, Chlorophyta), a New Species Isolated from Salt Pans in Goa, India. Eur J Phycol 2013, 48, 61–78. [Google Scholar] [CrossRef]
- Lortou, U.; Gkelis, S. Polyphasic Taxonomy of Green Algae Strains Isolated from Mediterranean Freshwaters. Biol Res-Thessaloniki 2019, 26, 11. [Google Scholar] [CrossRef] [PubMed]
- Trenkenshu, R.P. Calculation of the Specific Growth Rate of Microalgae. Marine Biological Journal 2019, 4, 100–108. [Google Scholar] [CrossRef]
- Krzemińska, I.; Pawlik-Skowrońska, B.; Trzcińska, M.; Tys, J. Influence of Photoperiods on the Growth Rate and Biomass Productivity of Green Microalgae. Bioprocess Biosyst Eng 2014, 37, 735. [Google Scholar] [CrossRef]
- Adarme-Vega, T.C.; Thomas-Hall, S.R.; Lim, D.K.Y.; Schenk, P.M. Effects of Long Chain Fatty Acid Synthesis and Associated Gene Expression in Microalga Tetraselmis Sp. Marine Drugs 2014, Vol. 12, Pages 3381-3398 2014, 12, 3381–3398.
- Santos-Ballardo, D.U.; Rossi, S.; Hernández, V.; Gómez, R.V.; del Carmen Rendón-Unceta, M.; Caro-Corrales, J.; Valdez-Ortiz, A. A Simple Spectrophotometric Method for Biomass Measurement of Important Microalgae Species in Aquaculture. Aquaculture 2015, 448, 87–92. [Google Scholar] [CrossRef]
- Murado, M.A.; Prieto, M.A. NOEC and LOEC as Merely Concessive Expedients: Two Unambiguous Alternatives and Some Criteria to Maximize the Efficiency of Dose–Response Experimental Designs. Science of The Total Environment 2013, 461–462, 576–586. [Google Scholar] [CrossRef]
- Bagchi, D. Cytotoxicity and Oxidative Mechanisms of Different Forms of Chromium. Toxicology 2002, 180, 5–22. [Google Scholar] [CrossRef]
- Kumar, V.; Dwivedi, S.K. Hexavalent Chromium Reduction Ability and Bioremediation Potential of Aspergillus Flavus CR500 Isolated from Electroplating Wastewater. Chemosphere 2019, 237, 124567. [Google Scholar] [CrossRef]
- Shi, L.; Xue, J.; Liu, B.; Dong, P.; Wen, Z.; Shen, Z.; Chen, Y. Hydrogen Ions and Organic Acids Secreted by Ectomycorrhizal Fungi, Pisolithus Sp1, Are Involved in the Efficient Removal of Hexavalent Chromium from Waste Water. Ecotoxicol Environ Saf 2018, 161, 430–436. [Google Scholar] [CrossRef]
- Park, G.-S.; Lee, S.-H.; Lee, S.-M. Phytoplankton as Standard Test Species for Marine Ecotoxicological Evaluation. Journal of Environmental Science International 2005, 14, 1129–1139. [Google Scholar] [CrossRef]
- Bucková, M.; Hegrová, J.; Jandová, V.; Svoboda, J.; Huzlík, J.; Ličbinský, R. Study of Bioaccumulation of Cr, Ni and Zn by the Green Alga Desmodesmus Subspicatus and Assessment of the Toxic Effect of Selected Elements and Their Mixtures on Algae Cells. J Appl Phycol 2023, 35, 2241–2256. [Google Scholar] [CrossRef]
- Berden-Zrimec, M.; Drinovec, L.; Zrimec, A.; Tišler, T. Delayed Fluorescence in Algal Growth Inhibition Tests. Cent Eur J Biol 2007, 2, 169–181. [Google Scholar] [CrossRef]
- Maathuis, F.J. Physiological Functions of Mineral Macronutrients. Curr Opin Plant Biol 2009, 12, 250–258. [Google Scholar] [CrossRef] [PubMed]
- Parker, D.L.; Kumar, H.D.; Rai, L.C.; Singh, J.B. Potassium Salts Inhibit Growth of the Cyanobacteria Microcystis Spp. in Pond Water and Defined Media: Implications for Control of Microcystin-Producing Aquatic Blooms. Appl Environ Microbiol 1997, 63, 2324–2329. [Google Scholar] [CrossRef] [PubMed]
- Müller, M.T.; Zehnder, A.J.B.; Escher, B.I. Membrane Toxicity of Linear Alcohol Ethoxylates. Environ Toxicol Chem 1999, 18, 2767–2774. [Google Scholar] [CrossRef]
- Reunova, Y.A.; Ayzdaycher, N.A. Effects of Detergent on Chlorophyll a Content and Quantity Dynamics of Microalga Chroomonas Salina (Wils.) Butch. (Cryptophyta). Int J Algae 2003, 5, 90–97. [Google Scholar] [CrossRef]
- Markina, Zh. V. Effects of Sodium Dodecyl Sulfate on the Growth Dynamics and Physiological State of the Microalga Dunaliella Salina (Chlorophyta). Russ J Mar Biol 2010, 36, 191–194. [Google Scholar] [CrossRef]
Figure 1.
(A) Scanning visible spectra of both microalgal cultures (ranging from 300 to 800 nm), used to identify and choose the maximum wavelength of 680 nm for D. subspicatus and Tetraselmis sp. Highlighted ranges (in gray) related to absorption wavelengths of Chl a (435 and 660 nm) and b (453 and 642 nm), according to the literature. Image of (B) D. subspicatus (length: 6 µm, width: 4 µm) and (C) Tetraselmis sp. (length: 14 µm, width: 9 µm) cell cultures. Images magnification: 20x.
Figure 1.
(A) Scanning visible spectra of both microalgal cultures (ranging from 300 to 800 nm), used to identify and choose the maximum wavelength of 680 nm for D. subspicatus and Tetraselmis sp. Highlighted ranges (in gray) related to absorption wavelengths of Chl a (435 and 660 nm) and b (453 and 642 nm), according to the literature. Image of (B) D. subspicatus (length: 6 µm, width: 4 µm) and (C) Tetraselmis sp. (length: 14 µm, width: 9 µm) cell cultures. Images magnification: 20x.
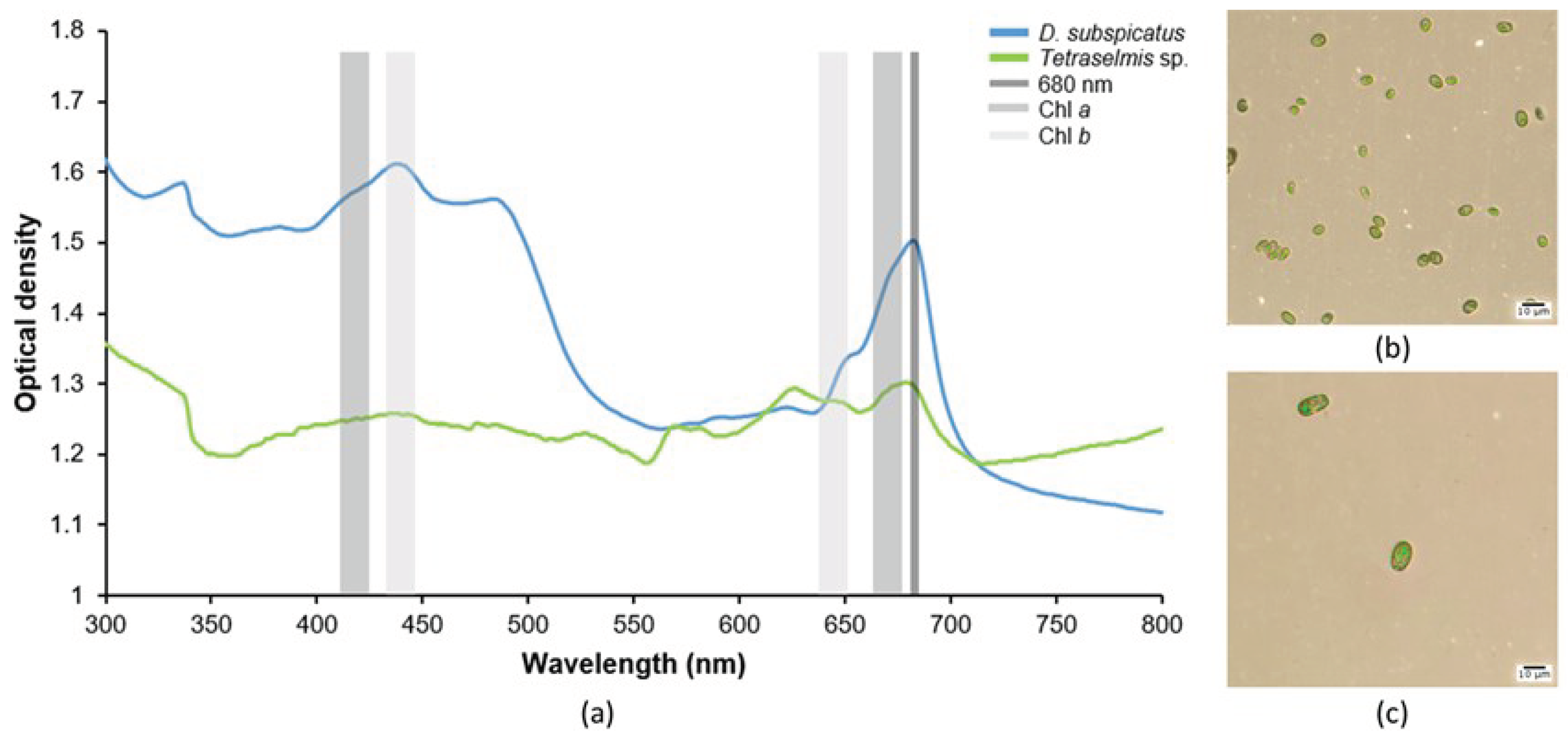
Figure 2.
Growth curves of (A) D. subspicatus and (B) Tetraselmis sp., along with the pH analysis during their growth, where the cell density of the culture was determined by OM cell counting and OD measurement at 680 nm.
Figure 2.
Growth curves of (A) D. subspicatus and (B) Tetraselmis sp., along with the pH analysis during their growth, where the cell density of the culture was determined by OM cell counting and OD measurement at 680 nm.
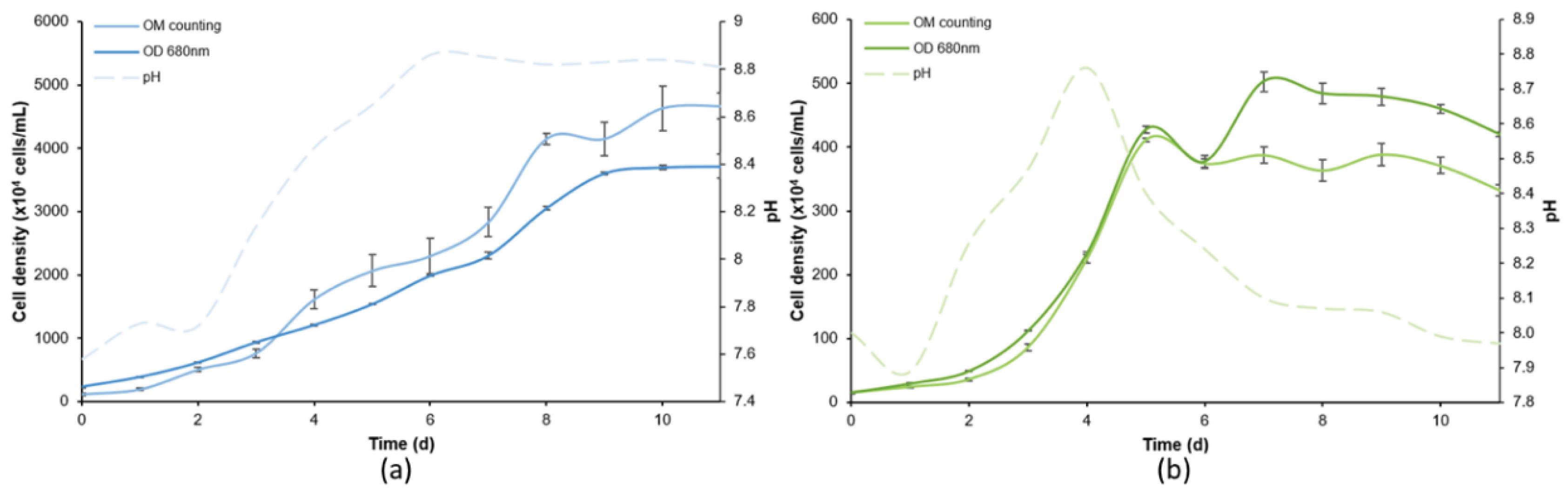
Figure 3.
Cell density of (A) D. subspicatus and (C) Tetraselmis sp. in response to K2Cr2O7 exposure, at times of 24, 48, 72, and 96 h. Dose-response curves for K2Cr2O7 according to the different exposure times for (B) D. subspicatus and (D) Tetraselmis sp. Error bars were calculated from triplicates, and the asterisks represent significant differences between the treatment and control (p < 0.05).
Figure 3.
Cell density of (A) D. subspicatus and (C) Tetraselmis sp. in response to K2Cr2O7 exposure, at times of 24, 48, 72, and 96 h. Dose-response curves for K2Cr2O7 according to the different exposure times for (B) D. subspicatus and (D) Tetraselmis sp. Error bars were calculated from triplicates, and the asterisks represent significant differences between the treatment and control (p < 0.05).
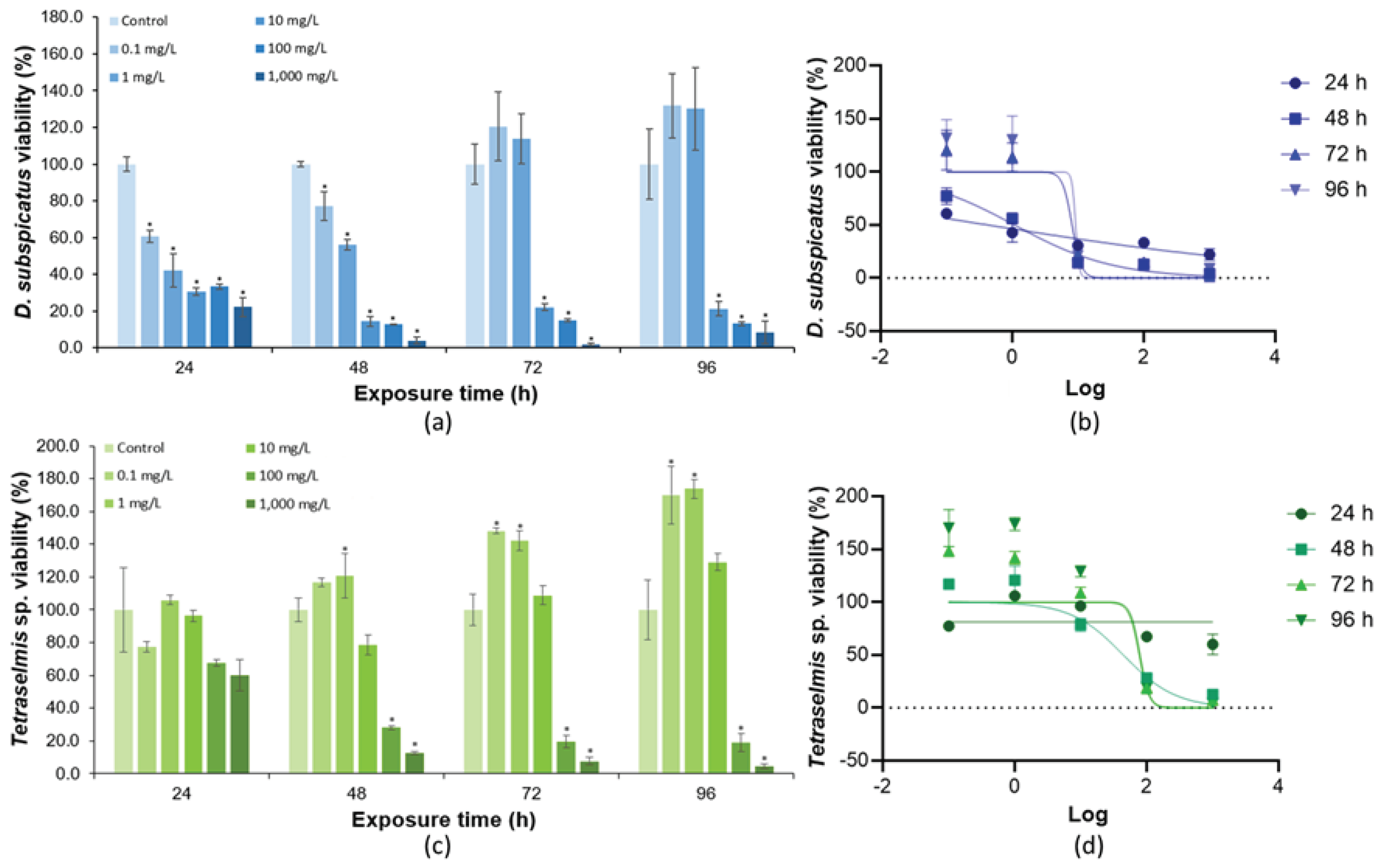
Figure 4.
Cell density of (A) D. subspicatus and (C) Tetraselmis sp. in response to KCl exposure, at times of 24, 48, 72, and 96 h. Dose-response curves for KCl according to the different exposure times for (B) D. subspicatus and (D) Tetraselmis sp. Error bars were calculated from triplicates, and the asterisks represent significant differences (p < 0.05) between the treatment and control tests.
Figure 4.
Cell density of (A) D. subspicatus and (C) Tetraselmis sp. in response to KCl exposure, at times of 24, 48, 72, and 96 h. Dose-response curves for KCl according to the different exposure times for (B) D. subspicatus and (D) Tetraselmis sp. Error bars were calculated from triplicates, and the asterisks represent significant differences (p < 0.05) between the treatment and control tests.
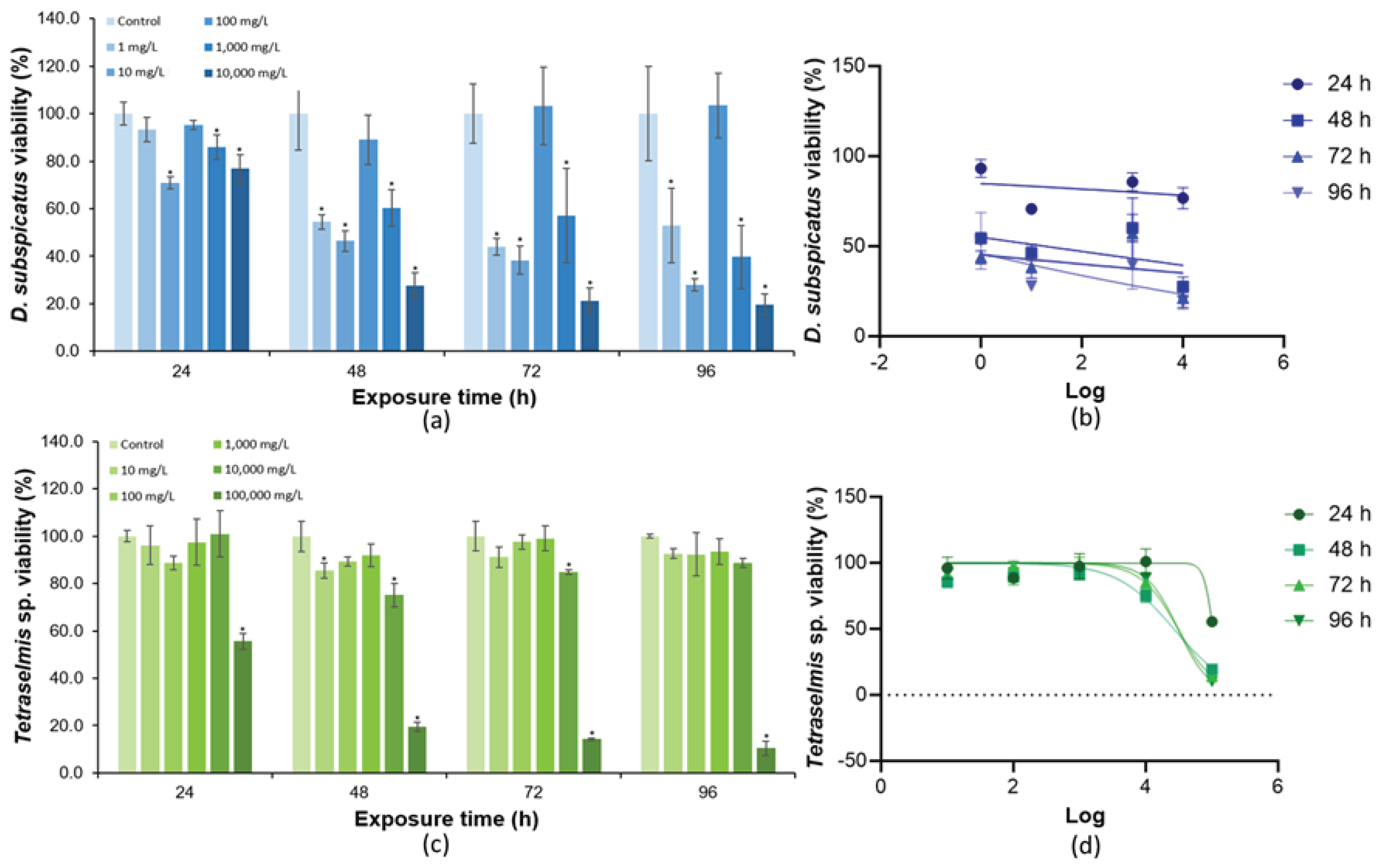
Figure 5.
(A) Cell density of Tetraselmis sp. in response to SDS exposure. (B) Dose-response for SDS curves according to the different exposure times for Tetraselmis sp. Error bars were calculated from triplicates and the asterisks represent significant differences between the control and exposure tests (p < 0.05).
Figure 5.
(A) Cell density of Tetraselmis sp. in response to SDS exposure. (B) Dose-response for SDS curves according to the different exposure times for Tetraselmis sp. Error bars were calculated from triplicates and the asterisks represent significant differences between the control and exposure tests (p < 0.05).
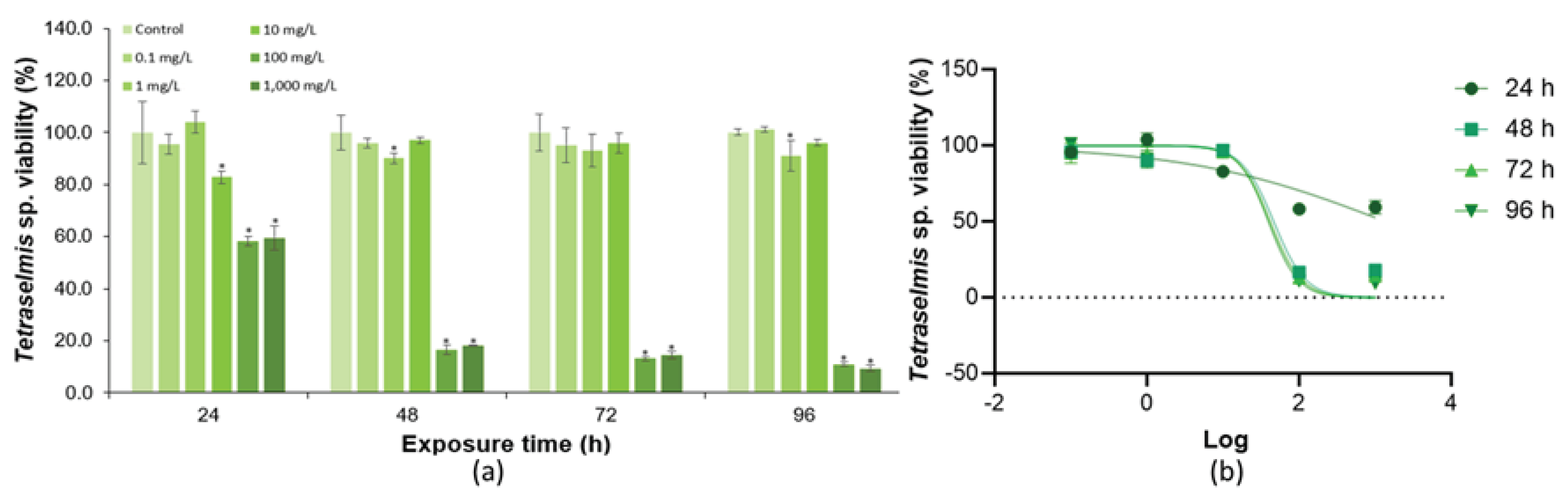

Table 1.
Linear regression model parameters for isolated and grouped cultures of D. subspicatus and Tetraselmis sp. Significant codes: p < 0.0001 ‘***’, p < 0.001 ‘**’, p < 0.01 ‘*’.
Table 1.
Linear regression model parameters for isolated and grouped cultures of D. subspicatus and Tetraselmis sp. Significant codes: p < 0.0001 ‘***’, p < 0.001 ‘**’, p < 0.01 ‘*’.
| Specie | Culture | a | b | R2 | F |
|---|---|---|---|---|---|
| D. subspicatus | 1 | 1.573 x 10-01 | 1.252 x 10-07 | 0.996 | 766.7*** |
| 2 | 3.640 x 10-03 | 1.362 x 10-07 | 0.999 | 3280*** | |
| Pooled | 7.354 x 10-02 | 1.313 x 10-07 | 0.980 | 399.1*** | |
| Tetraselmis sp. | 3 | 3.592 x 10-02 | 2.918 x 10-07 | 0.997 | 1113*** |
| 4 | -3.144 x 10-04 | 3.359 x 10-07 | 0.998 | 1345*** | |
| Pooled | 1.771 x 10-02 | 3.151 x 10-07 | 0.992 | 1038*** |
Table 2.
Specific growth rates (μ d-1) for the microalgal cultures estimated by both cell density measurement techniques.
Table 2.
Specific growth rates (μ d-1) for the microalgal cultures estimated by both cell density measurement techniques.
| Microalgae | Cell density technique | μ (d-1) | Doubling time (h) |
|---|---|---|---|
| D. subspicatus | OM | 1.00 | 16.56 |
| OD | 0.71 | 23.52 | |
| Tetraselmis sp. | OM | 3.13 | 5.28 |
| OD | 2.35 | 6.96 |
Table 3.
EC50, NOEC, and LOEC values, obtained in the toxicity tests performed with the microalgae D. subspicatus and Tetraselmis sp. and the chemicals K2Cr2O7, KCl, and SDS on the best fit on exposure times.
Table 3.
EC50, NOEC, and LOEC values, obtained in the toxicity tests performed with the microalgae D. subspicatus and Tetraselmis sp. and the chemicals K2Cr2O7, KCl, and SDS on the best fit on exposure times.
|
Toxicity Data |
D. subspicatus | Tetraselmis sp. | ||||
|---|---|---|---|---|---|---|
| K2Cr2O7 | KCl | SDS | K2Cr2O7 | KCl | SDS | |
| Exposure time | 72 h | 48 h | 72 h | 48 h | 72 h | 72 h |
| EC50 (mg/L) | 8.1 | 19.57 | - | 44.5 | 30,908 | 41.6 |
| R2 | 0.9120 | 0.2277 | - | 0.8967 | 0.9774 | 0.9564 |
| NOEC (mg/L) | 1 | <1 | 100 | 0.1 | 1,000 | 10 |
| LOEC (mg/L) | 10 | 1 | 1,000 | 1 | 10,000 | 100 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



