
Submitted:
05 March 2024
Posted:
05 March 2024
You are already at the latest version
Abstract
Meloidogyne nematodes, commonly known as root-knot nematodes, pose a considerable threat to crop yields, resulting in significant economic losses due to their intricate biology and limited control methods. In line with the European Union's focus on promoting organic farming and soil health to encourage sustainable agricultural practices, this study explores the efficacy of two characterized bacterial communities obtained from compost, namely SynCom1 and SynCom2, against Meloidogyne javanica in tomato plants. Through pot experiments, it is demonstrated that both bacterial communities effectively suppress nematode reproduction and root infections while simultaneously enhancing various growth parameters in tomatoes. These findings underscore the potential of synthetic bacterial communities as promising tools for organic or integrated pest management, thereby supporting sustainable agricultural practices and contributing to improved crop yields.
Keywords:
Meloidogyne javanica
; synthetic bacterial communities
; sustainable agriculture
1. Introduction
Root knot nematodes (Nematoda: Heteroderidae: Meloidogyne spp., RKN for short) are clearly included amongst the most significant soil borne pathogens that account for highly significant yield losses per year [1]. Particularly, Meloidogyne javanica (Treub) Chitwood is one of the major pests infesting tomato, as well as many other economically important plant species, has a worldwide distribution with a preference to temperate climates and is the most abundant RKN species in Greece [2,3].
Control of these pests has always been an issue as they are considered extremely difficult to suppress, due to their soil inhabiting and endoparasitic nature, broad host range and lack of resistant varieties. Traditionally, RKN control is pursued with chemical nematicides [4], which during the past decades have unequivocally been accused with a negative environmental impact leading to almost total ban, having the global scientific committee being forced to study and develop more benign, alternative solutions. These solutions, including biological control agents should of course be incorporated into integrated pest management schemes [5].
Biological control research has revealed a number of microorganisms such as bacterial and fungal species, that have potential over RKN [6,7,8]. For example, Penicillium chrysogenum and Trichoderma spp., caused 97.67 and 95% mortality of M. javanica juveniles in vitro respectively [9]. Several soil-inhabiting fungal species, which express a nematophagous function have been considered as efficient biocontrol agents i.e. Trichoderma harzianum, Verticillium chlamydosporium and Paecilomyces lilacinus, as well as species of the Pochonia, Penicillium and Fusarium genera. These microorganisms have been reported to effectively kill eggs, juveniles or female RKN and have significantly decreased the population of parasitic nematodes in the soil [10,11]. Other fungi are capable to trap nematodes by capturing them with special hyphae forming rings or other adhesive structures where the nematodes are trapped and subsequently killed by released nematotoxic metabolites [12]. Additionally, many arbuscular mycorrhizal fungi species (AMF for short) have been shown to enhance growth and inhibit RKN activity when inoculated on the roots of plants growing in RKN infested soils [13,14,15,16,17,18]. On the other hand, several species of rhizobacteria including, Pseudomonas, Bacillus and Pasteuria have been shown to affect RKN species [19,20,21] through antibiosis, direct parasitism, competition for food or space, induction of systemic resistance of the host plant and reduction of root penetration [22,23]. Also, Streptomyces, Bacillus, Pseudomonas and Pasteuria are reported as nematicidal [24,25]. Particularly, Bacillus firmus, B. subtilis and B. coagulans have been shown to inhibit nematode infection caused by M. javanica on brinjal [26]. Bacillus subtilis has also been reported as a preventor of M. incognita egg hatching [27], and B. subtilis bioactive surfactin, lipopeptide, iturin and other compounds were reported as antagonistic to M. incognita [28].
Despite the promise of individual biocontrol agents, none have yet achieved the level of efficacy necessary to fully replace chemical nematicides. This raises an intriguing option - that these agents, when working collaboratively within a well-structured community, with each contributing to RKN control, could collectively lead to more promising outcomes.
The plant roots, namely the part of the plant that RKN invade and inhabit, are surrounded by a soil layer, the rhizosphere, that is inhabited by a vast diversity of microorganisms, collectively characterized as the microbiome. This micro-ecosystem is especially complex and affected by the plant type (annual, biannual, perennial, woody, shrubby, grassy etc.), plant species and/or plant activity (climbers, heathers, carnivorous etc.), and vice versa, the microbial species comprising the microbiome possess differential properties affecting plant growth and plant and soil health [29,30,31]. The microbiome structure’s fundamental determinant is the rhizosphere’s chemical content, primarily being defined by the root exudate composition, secondly by the organic matter content and thirdly by any organic amendment [29,30,32,33].
When RKNs enter the roots of a host plant, they release molecules (enzymes etc) that cause the root cells close to the penetration site to enlarge for the establishment of a feeding site. If these chemicals disrupt the chemical balance of the rhizosphere, the microbiome may not be capable to support plant growth sufficiently. On the other hand, it is postulated that plants, in order to support their health and growth, as well as to suppress soil-borne pathogens, actively recruit beneficial bacteria [34,35]. But if the plant’s physiology is affected by the nematode attack, this system may need to be “manually” reinforced. Considering that the microbiome is affected by the chemical status of the rhizosphere as previously described, disruption of this balance may affect microbiome composition, which subsequently may affect plant phenotype [36]. Thus, comprehension of the processes that regulate the rhizospheric microbiome structure and how particular constituents influence plant health and growth are of high importance.
For example, Bacillus rhizobacteria have been reported to possess the ability to produce antimicrobial (-bacterial and -fungal) secondary metabolites and stimulate plant growth [37,38]. Moreover, Bacillus spp. have been demonstrated as a rich source of bioactive compounds (exotoxins, enzymes, antibiotics) that give them bionematicidal attributes [26,39,40,41].
As several studies have focused on the combined control of RKN and soil borne fungi, especially Fusarium species (e.g. [42,43,44,45]), in the present work we tested the possibility to suppress RKN infection of tomato plants recruiting two synthetic bacterial communities (SynComs) that were previously successfully engaged in protecting tomato plants against Fusarium oxysporum f. sp lycopersici and promoting plant growth [36].
2. Materials and Methods
2.1. Preparation of M. javanica inoculum
The M. javanica population was reared on tomato seedlings (Solanum lycopersicum L.) cv. Belladonna in a temperature-controlled environment (27 ± 1 °C; 60% humidity; 16 h light) at the Plant Protection Laboratory of the University of Patras, Greece. Well-developed egg masses extracted from the source plant roots were placed on a Baermann funnel at room temperature until emergence of second stage juveniles (J2s). The J2s that emerged during the first three days were discarded because of age differences, then J2s collected on the 4th day were used as inoculum.
2.2. Preparation of bacterial SynComs
In this study, we employed two distinct bacterial synthetic communities with varying compositions. SynCom1 comprised a diverse array of bacterial genera, while SynCom2 exclusively consisted of Bacilli [36] (Table S1). For the preparation of the SynComs each bacterial isolate underwent a 48-hour growth period in Tryptic Soy Broth (TSB) at 25°C with agitation at 180 rpm. Following growth, cultures were subjected to two rounds of rinsing with a sterile 10 mM MgCl2 solution, followed by centrifugation at 2,600 x g for 8 minutes. After the final rinse and centrifugation, cells were resuspended in a 10 mM MgCl2 solution. The optical density at 600 nm (OD600) of each suspension was adjusted to 0.5 (~2.75 x 108 cfu ml-1). To create synthetic communities (SynComs), the bacterial isolates were mixed in equal proportions. For the tomato pot experiments, 2 ml of each SynCom suspension with an OD600 of 0.5 was added to 50 ml of sterile 10 mM MgCl2 and root-drenched in pots containing 200 grams of sterilized potting substrate, resulting in an estimated density of approximately ~2.75 x 106 cells per gr of substrate. This mixture was prepared immediately before the surface-sterilized tomato seeds were sown [36].
2.3. Pot experiment
Seeds of the tomato (Solanum lycopersicum) cv. Ailsa Craig were surface-sterilized following the procedure outlined by Chialva et al. (2016) [46]. Subsequently, the seeds were planted in 10.5 cm diameter pots, each filled with 200 grams of sterile peat-based potting substrate (Plantobalt substrate 2, Plantaflor) [47]. Plants were cultivated within a controlled growth chamber (25°C, 16 h light, 8 h dark, 65%–70% RH, light intensity 450 μmol m−2 s−1 at pot height). Plants were watered every other day, with each receiving an equal volume of water.
At the 3-leaf stage, the plants were subjected to a second root drenching with one of the SynCom suspensions, as previously described, or with water. Subsequently, they were placed in a growth chamber for acclimatization. After 48 hours, the plants underwent manual inoculation with a 2000-J2 nematode inoculum, which was directly applied to the roots of each plant using a pipette. Two weeks later, the plants received a third root-drenching with the respective SynCom suspensions.
To evaluate the relative effectiveness of SynComs, we introduced a commercial product in the experiments. Pochar (MS BIOTECH) is a liquid formulation containing fungal antagonists Pochonia and Arthrobotis and other microbial inocula, which serve as natural enemies against root-knot nematodes. The formulation was activated by mixing with water and Nutryaction (MS BIOTECH), an organic fertilizer containing yeast and brown algae extracts using a ratio of 1-13-2 for formulation-H2O-Nutryaction, respectively. The mixture was prepared at least 8 hours prior to application on plants. Application of the formulation was conducted through root drenching, with each plant receiving 10 ml during each application at the same time points as the application of SynComs.
The experimental treatments consisted of the following groups: (a) Plants treated with SynCom1 (SC1), (b) Plants treated with SynCom2 (SC2), (c) Plants treated with Pochar (Pochar) and (d) Plants treated with water (H2O). Within each treatment group, half of the plants were subjected to nematode inoculation (nem), while the remaining half served as controls and received only water.
2.4. Plant manipulation for nematode extraction and growth parameter assessment
Forty days after nematode inoculation, the plants were uprooted to confirm on the presence of well-developed RKN egg masses.
- Assessment of plant height: the plant height, i.e. the height from the plant collar up to its top was measured with a tape measure in cm.
- Assessment of the fresh and dry weights of the aerial parts: the aerial parts were separated from the roots, their fresh weight weighed on a 2-digit scale, then placed in a paper bag and left in an oven @50oC until completely dry, and subsequently the dry weight was recorded.
- Assessment of the fresh weight of roots: The roots of each plant were separately washed thoroughly but carefully, under running tap water, tap-dried with a paper towel and their fresh weight weighed on a 2-digit scale.
- Assessment of the total number of egg masses and eggs: The total number of egg masses per plant was counted under a stereomicroscope. Subsequently, for each plant, ten egg masses were randomly selected, placed in 1% sodium hypochlorite, shaked for 10 minutes to disperse the eggs, then the number of eggs per egg mass was estimated under the stereomicroscope.
- Assessment of the nematode population in the soil: the soil of each pot was handled following the conventional Baermann funnel technique [48] to estimate RKN soil population.
2.5. Statistical analysis
Data obtained in all experiments were analyzed by one-way ANOVA (analysis of variance, Tukey’s test with P ≤ 0.05) using GraphPad Prism 8 for Windows (GraphPad Software, La Jolla, CA, United States).
3. Results
In the pot experiments, the effects of SynCom1, SynCom2 and Pochar were assessed for various plant growth characteristics. Fresh weight and dry weight of aerial parts, plant height and fresh weight of root were measured in both control plants and plants inoculated with nematodes (Figure 1).
The results revealed notable differences in the fresh weight of plants among different treatments. Specifically, plants inoculated with nematodes exhibited a decreased fresh weight compared to their respective control plants, except for those treated with Pochar (Figure 1A). SynCom1 treatment resulted in the highest fresh weight, significantly distinct from the other treatments. SynCom2 and Pochar treatments did not significantly differ in fresh weight from water-treated plants. However, plants that were both inoculated with nematodes and treated with SynCom1, SynCom2, and Pochar exhibited a greater fresh weight compared to their counterparts treated solely with water and inoculated with nematodes (Figure 1A). The results suggest that SynCom1, SynCom2 and Pochar treatments might have a suppressive or mitigating effect on the detrimental influence of nematode infestation on plant growth. In contrast, water-treated plants inoculated with nematodes exhibited a lower fresh weight compared to the control plants, emphasizing the negative impact of nematode infestation when no treatment was applied. These findings underscore the potential effectiveness of SynCom1, SynCom2, and Pochar in promoting healthier plant growth even in the presence of nematodes.
Similar trends were observed in the results for the dry weight of plants, mirroring the patterns seen in fresh weight (Figure 1B). Significant differences in dry weight among the various treatments were consistent with the findings in fresh weight. Notably, plants inoculated with nematodes showed reduced dry weight compared to their respective controls, except for those treated with Pochar. SynCom1 treatment continued to demonstrate the most significant impact, exhibiting a dry weight distinct from the other treatments. SynCom2 also exhibited a significant difference compared to the water-treated plants, while Pochar treatments did not yield a significant difference in dry weight compared to the water-treated plants. Moreover, plants treated with SynCom1, SynCom2, and Pochar, and inoculated with nematodes, displayed a higher dry weight compared to plants treated only with water and subjected to nematode inoculation (Figure 1B). These parallel outcomes between fresh and dry weight strengthen the implication that SynCom1, SynCom2, and Pochar treatments potentially mitigate the adverse effects of nematode infestation on plant growth. Conversely, water-treated plants inoculated with nematodes consistently exhibited a lower dry weight, highlighting the detrimental impact of nematode infestation in the absence of treatment.
Regarding plant height, significant differences were observed among different treatments (Figure 1C). Specifically, in each treatment group, plants inoculated with nematodes exhibited a reduction in height compared to their corresponding control plants. SynCom1 treatment resulted in greater plant height compared to all treatments, while SynCom2 and Pochar treated plants were significantly higher compared to the water-treated plants. Moreover, plants treated with SynCom1, SynCom2, and Pochar, and inoculated with nematodes, displayed increased height compared to their counterparts treated with water alone (Figure 1C). These results align with data related to both fresh weight and dry weight highlighting the potential of SynCom1, SynCom2, and Pochar in positively influencing plant height.
In terms of the fresh weight of the roots, the application of SynCom1, SynCom2, and Pochar on plants inoculated with nematodes did not result in a significant difference compared to their respective control groups (Figure 1D). However, it is noteworthy that the fresh weight of roots in water-treated plants was significantly lower than in the other treatments. Interestingly, water-treated plants inoculated with nematodes did not show a significant difference in fresh weight of roots when compared to the corresponding control plants (Figure 1D). These findings suggest that the treatments involving SynCom1, SynCom2, and Pochar may have potential benefits in maintaining or mitigating the adverse effects on root fresh weight caused by nematode infestation, particularly when compared to untreated plants.
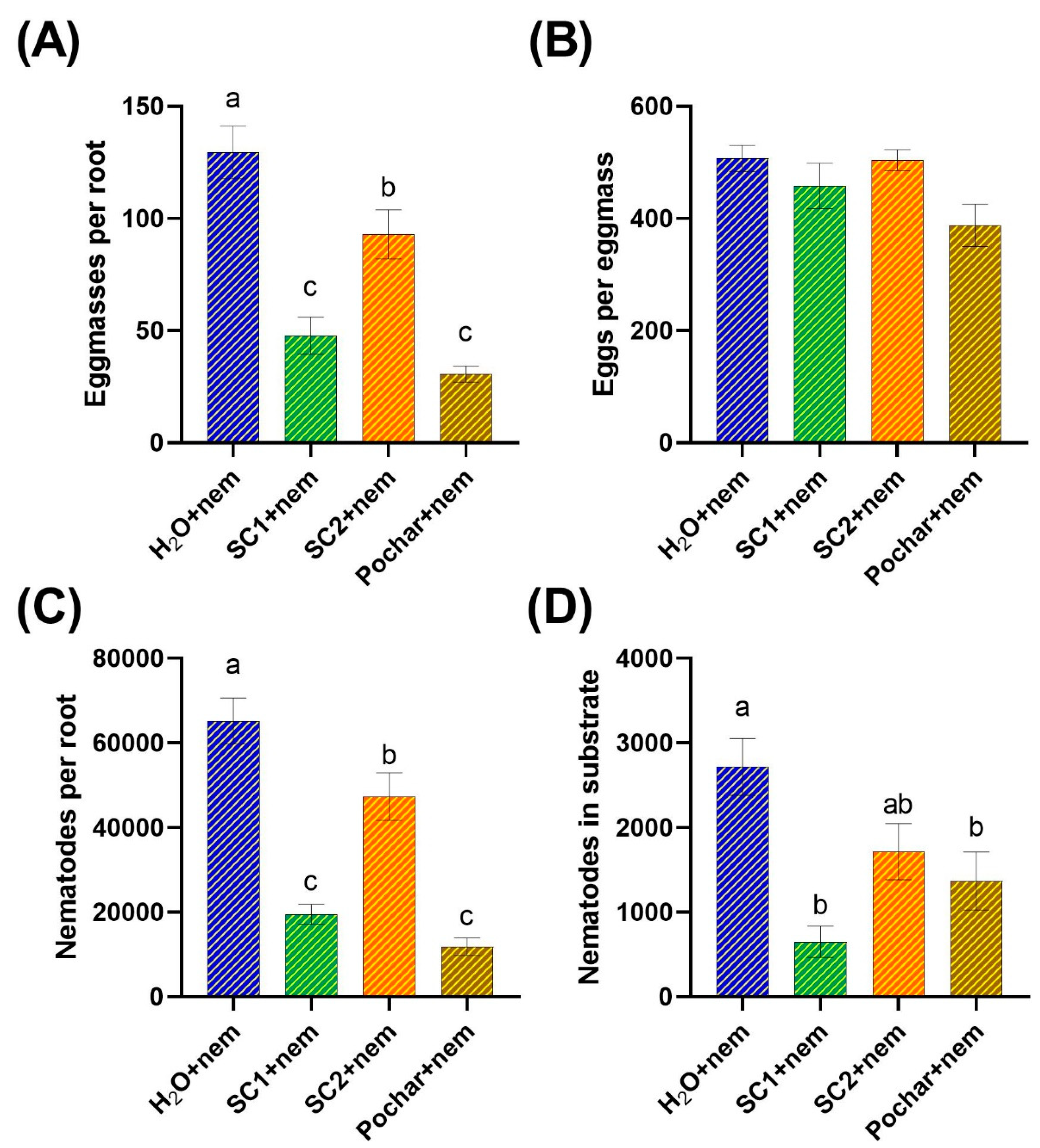
In an effort to elucidate the potential efficacy of SynCom1, SynCom2, and Pochar in alleviating the adverse effects on tomato plants caused by nematode infestation, we aimed to evaluate their impact on the reproductive capacity of Meloidogyne javanica. Forty days after nematode inoculation, the plants were uprooted, and we assessed the total number of eggmasses per root, the number of eggs per eggmass, the total number of nematodes in the substrate and the total number of nematodes per root (Figure 2).
The results revealed that water-treated plants exhibited the highest number of eggmasses and nematodes per root, followed by plants treated with SynCom2 (Figure 2A and 2C). Conversely, plants treated with Pochar and SynCom1 displayed the lowest number of eggmasses and nematodes (Figure 2A and 2C). These findings suggest that Pochar, SynCom1, and SynCom2 have a negative impact on the extent of nematode reproduction and the potential number of offspring emerging from the plant roots.
In terms of the number of nematodes in the substrate, the results revealed the highest enumeration in the substrate of water-treated plants, while the lowest count was observed in plants treated with Pochar and SynCom1. Interestingly, the number of nematodes in the substrate of plants treated with SynCom2 did not show a significant difference compared to water-treated or Pochar/SynCom1 treated plants (Figure 2D).
Regarding eggs per eggmass, no significant differences were found among the different treatments (Figure 2B), suggesting that the various treatments did not have a significant impact on the fertility or reproductive success of female Meloidogyne javanica.
Based on the results, it can be concluded that water-treated plants exhibited the highest nematode infestation, as indicated by the elevated number of eggmasses and nematodes per root. In contrast, plants treated with Pochar and SynCom1 demonstrated the lowest nematode presence, suggesting the potential suppressive effects of these treatments on nematode reproduction and population. SynCom2 treatment, while not significantly different from water-treated plants regarding nematode numbers in the substrate, showed a notable impact on reducing nematode numbers in roots. Therefore, the findings suggest that Pochar, SynCom1, and SynCom2 have the potential to negatively influence nematode reproduction and the overall nematode population in the plant's rhizosphere.
4. Discussion
Root knot nematodes (RKN), particularly Meloidogyne javanica, pose a substantial threat to agricultural productivity, causing significant yield losses globally [49,50,51]. The use of chemical nematicides, once a common strategy for RKN control, has faced environmental concerns and restrictions, necessitating the exploration of alternative, environmentally friendly approaches. Biological control agents, including bacteria, fungi, and other microorganisms, have shown promise in mitigating RKN infestations [52,53].
In our study, we investigated the potential of synthetic bacterial communities (SynCom1 and SynCom2) and the commercial product Pochar in alleviating the adverse effects of M. javanica on tomato plants. These communities were previously demonstrated to be effective against another soil-borne pathogen, Fusarium oxysporum [36].
The results indicated that SynCom1, SynCom2, and Pochar had a significant impact on various plant growth parameters in the presence of nematode infestation. Notably, plants inoculated with nematodes and treated with SynCom1, SynCom2, and Pochar exhibited higher fresh and dry weights of aerial parts, increased plant height, and enhanced fresh weight of roots compared to water-treated plants inoculated with nematodes. These findings suggest a potential suppressive effect of SynCom1, SynCom2, and Pochar on the detrimental influence of nematode infestation on plant growth.
The assessment of nematode reproductive parameters further supported the efficacy of the treatments. Water-treated plants exhibited the highest number of eggmasses and nematodes per root, while Pochar, SynCom1, and SynCom2 treatments resulted in significantly lower numbers. Additionally, the enumeration of nematodes in the substrate indicated a notable reduction in SynCom2 and Pochar-treated plants compared to water-treated plants, emphasizing their potential in limiting nematode reproduction in roots.
Surprisingly, eggs per eggmass did not show significant differences among the treatments, suggesting that while SynCom1, SynCom2, and Pochar influenced the overall nematode population, they might not have a direct impact on the fertility or reproductive success of individual females.
The apparent success of SynCom1 and SynCom2 in controlling nematodes could be attributed to several factors. First, the synthetic communities might have introduced beneficial microbial consortia into the rhizosphere, fostering a microbiome that actively contributes to plant health and simultaneously impedes nematode activity. The introduction of SynCom1 and SynCom2 might have contributed to an environment that is less hospitable for nematodes, disrupting their life cycle and reproductive success. This aligns with the findings of Zhou et al. [54], who demonstrated that soils in non-infested regions of fields under high root-knot nematode (RKN) pressure exhibit greater microbial diversity than infested areas and the inoculation of tomato roots with the microbiome from non-infested soils resulted in a reduction in the number of root galls. Consequently, this suggests that enhancing the diversity and abundance of specific microbial groups could be a viable strategy for managing RKN.
Second, the bacteria of the synthetic communities might produce bioactive compounds or exhibit antagonistic behaviors that directly or indirectly affect nematode viability. The direct mechanisms include the synthesis of lytic enzymes, antibiotics, toxins and metabolites with antagonistic effects or parasitism [55]. Conversely, indirect mechanisms encompass the release of molecules that modulate nematode behavior, the competition for nutrients, the induction of systemic resistance (ISR), and the promotion of plant growth [56,57]. These indirect strategies involve niche exclusion, nitrogen metabolism, siderophore production, phosphate solubilization, and hormonal regulation [58].
Lytic enzymes produced by bacteria can cause damage to both eggshells of nematodes which consist of a protein matrix and a chitin layer [59] and to the cuticle of the nematodes, which is composed of a proteinaceous membrane [60]. Chitinases, proteases, peptidyl-peptide hydrolases, and gelatinolytic proteins are enzymes capable of cleaving these structures [55]. In our study, both SynComs consisted of bacterial isolates, with some demonstrating the ability to produce a variety of hydrolytic enzymes including chitinases, proteases, cellulases and pectinases. More specifically, as observed in Tsolakidou et al. [36], 3 out of 25 bacterial isolates exhibited chitinase-producing capabilities within SynCom1; SynCom2 showed a higher proportion, with 10 out of 25 isolates demonstrating chitinase activity. Regarding proteases, a single isolate from SynCom1 exhibited protease activity, while SynCom2 displayed a higher activity level, with 6 out of 25 isolates showing protease activity. There is extensive literature on the pivotal role of lytic enzymes, especially chitinases and proteases, of microbial origin in combating nematodes by disrupting nematode structures, compromising their integrity, and contributing to their biocontrol in agricultural and ecological settings (reviewed in [61]).
In addition to the previously mentioned enzymes, various other lytic enzymes have been identified for their involvement in controlling the incidence of RKN infection. These include pectinases, β-glucanases, and cellulases [62]. In the study of Tsolakidou et al. [36] it was reported that, within SynCom1, 15 isolates demonstrated pectinase activity, and 2 isolates exhibited cellulase activity whereas, in SynCom2, 3 isolates showed pectinase activity, while 8 isolates demonstrated cellulase activity.
Phosphorus is an essential nutrient for plant growth and development, but it is often present in soil in insoluble forms, making it inaccessible to plants. Microorganisms capable of solubilizing phosphate play a crucial role in making this essential nutrient available to plants. Phosphate-solubilizing microorganisms also contribute to plant growth assistance by functioning as biocontrol agents against a variety of pathogenic organisms and nematodes. For example, El-Hadad et al. [63] noted that the application of Bacillus megaterium, an efficient phosphorus mobilizer, led to increased shoot length, plant biomass, NPK levels, and a reduction in the colonization of M. incognita in tomato plants. In the study of Tsolakidou et al. [36] it was reported that within SynCom1, one isolate demonstrated phosphate solubilization, while within SynCom2, eight isolates exhibited phosphate solubilization.
Apart from lytic enzymes, several studies demonstrated that rhizobacteria reduce nematodes severity by inducing plant systemic resistance. Pseudomonas spp. and Bacillus spp. have been implicated in employing induced systemic resistance (ISR) as a defense strategy against root-knot nematodes (RKNs) [64,65,66]. This induced resistance is achieved through the reinforcement of the cell wall, involving processes such as cell wall thickening, callose deposition, and the accumulation of phenolic compounds. Additionally, it encompasses the synthesis of various biochemical compounds that are up-regulated in defense reactions, including PR proteins, phytoalexins, lipopolysaccharides (LPS), siderophores, salicylic acid (SA), jasmonic acid (JA), peroxidase (PO), chitinase, and other secondary metabolites [67,68]. The induction of systemic resistance by rhizobacteria involves numerous bacterial factors, with key elements including lipopolysaccharides found in the outer membrane of bacterial cells, as well as the production of siderophores and salicylic acid [69]. Additionally, various cyclic lipopeptides (CLPs) have been demonstrated to stimulate host immune responses and trigger induced systemic resistance (ISR) in specific host plants [70,71]. In our study, both SynComs consisted of bacterial isolates harboring genes associated with the biosynthesis of cyclic lipopeptides [36]. SynCom1 comprised isolates encoding genes for bacillomycin B synthetase and surfactin A and B synthetases, while SynCom2 included bacteria with genes for bacillomycin B synthetase, surfactin A and B synthetases, fengycin D synthatase, bacilysin biosynthesis protein A, and the antibiotic subtilin [36]. The identification of cyclic lipopeptide (CLP) genes in both SynComs implies their potential involvement in stimulating host immune responses and initiating ISR. This aligns with prior research highlighting the ISR involvement against RKN.
The majority of rhizobacterial strains are able to produce plant growth-promoting substances that actively regulate plant growth and development. These plant growth-promoting rhizobacteria (PGPR) can stimulate plant growth through the production of phytohormones, including auxins, cytokinins, gibberellic acid, abscisic acid, and ethylene [72] or through enzymatic activities such as aminocyclopropane-1-carboxylate (ACC) deaminase [73]. Notably, indole acetic acid (IAA) emerges as the most prevalent phytohormone serving as a key regulator that promotes cell division, elongation, lateral and adventitious rooting in plants [74,75]. This hormone not only fosters overall plant growth but also improves nutrient and mineral uptake, contributing to increased plant vigor and yield [76,77]. Previous studies suggest that phytohormones synthesized by PGPRs may alleviate the detrimental effects of various environmental stresses [78]. Inoculating fields with IAA-producing PGPR strains through seed application resulted in enhanced plant growth and biocontrol of Meloidogyne incognita nematodes [79]. Additionally, phytohormone-producing Streptomyces strains, including IAA, have demonstrated the ability to enhance the growth of eggplants, potentially reducing the number of galls and nematode egg masses of M. incognita [80]. Similarly, Ruanpanun et al. [81] reported the nematicidal activity of IAA-producing Streptomyces sp. against M. incognita. In our study, both SynComs comprised bacterial isolates, some of which exhibited the ability to produce IAA. More specifically, within SynCom1, 10 out of 25 bacterial isolates demonstrated IAA-producing capabilities, while in SynCom2 a higher proportion was observed, with 16 out of 25 isolates producing IAA [36].
Additionally, ACC deaminase, an enzyme produced by PGPR, reduces the levels of the stress hormone ethylene by breaking down its precursor ACC. This reduction in ethylene levels is particularly beneficial for plants under stress conditions, as ethylene is associated with stress responses [74] including nematodes attack [82]. Rhizobacteria equipped with ACC deaminase have demonstrated remarkable efficacy in controlling nematode-induced diseases, as exemplified by the study using Pseudomonas putida UW4 as a model strain [83]. In the case of pine wilt disease (PWD) caused by the pinewood nematode Bursaphelenchus xylophilus, inoculation with ACC deaminase-proficient P. putida UW4 significantly reduced PWD symptoms in Pinus pinaster seedlings while the bacterial mutant lacking ACC deaminase displayed expected PWD symptoms. Moreover, seedlings inoculated with P. putida UW4 exhibited reduced nematode colonization compared to both uninoculated plants and those inoculated with the bacterial mutant. Notably, P. putida UW4 did not demonstrate direct nematicidal effects on B. xylophilus in vitro, indicating its ability to enhance plant defenses against nematodes without directly affecting nematode populations [84]. These findings highlight the potential of ACC deaminase-producing rhizobacteria as effective biocontrol agents against nematode-mediated diseases in plants. In our study, the rhizobacterial community SynCom1 was characterized by the presence of isolates exhibiting ACC deaminase activity, with 11 out of 25 isolates demonstrating this trait. In contrast, none of the isolates within SynCom2 harbored the ACC deaminase activity [36]. This distinct difference in the ACC deaminase profile between the two synthetic communities may provide insights into the observed variations in their efficiency. Notably, SynCom1 displayed a more efficient effect, evidenced by fewer eggmasses per root and a reduced nematode population per root compared to SynCom2. The absence of ACC deaminase activity in SynCom2 suggests that this particular enzymatic activity, known for its involvement in mitigating plant stress responses, could contribute to the enhanced performance of SynCom1 in promoting resistance against nematodes. This finding underscores the potential role of ACC deaminase-producing rhizobacteria in influencing the efficacy of synthetic communities in managing nematode-associated issues in plants.
5. Conclusions
The synthetic bacterial communities emerge as potent allies in the battle against nematode-induced crop losses. Their ability to enhance plant growth and concurrently suppress nematode reproduction positions them as promising components in sustainable agricultural practices. Our findings offer valuable insights into the potential mechanisms by which the synthetic communities may impact nematode susceptibility, encompassing microbial community dynamics, lytic enzyme production, cyclic lipopeptide synthesis, IAA production and ACC deaminase activity. Understanding the specific mechanisms by which SynCom1 and SynCom2 influence nematode populations will be crucial for refining and optimizing their application as biocontrol agents. Future research should delve deeper into the mechanistic aspects of their activity, the long-term stability of introduced microbial communities, and their compatibility with other elements of integrated pest management strategies. The potential of synthetic microbial communities to revolutionize nematode control opens exciting avenues for environmentally friendly and effective solutions in modern agriculture.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1: List of bacterial isolates composing the synthetic communities (SynComs).
Author Contributions
Conceptualization, E.K. and I.S.P.; methodology, E.K. and I.S.P.; formal analysis, E.K., V.K and I.S.P.; investigation, E.K., V.K and I.S.P.; resources, E.K. and I.S.P.; data curation, E.K. and I.S.P.; writing—original draft preparation, E.K. and I.S.P.; writing—review and editing, E.K. and I.S.P.; supervision, E.K. and I.S.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research was financially supported by the Research Committee of the University of Patras (Project 81883).
Data Availability Statement
The data presented in this study are available on request from the corresponding authors.
Acknowledgments
The authors would like to acknowledge the support of T.P. Agrobiotech LTD for offering the commercial products Pochar and Nutryaction (MS BIOTECH) used for experiments.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Talavera-Rubia, M.; Vela-Delgado, M.D.; Verdejo-Lucas, S. A Cost-Benefit Analysis of Soil Disinfestation Methods against Root-Knot Nematodes in Mediterranean Intensive Horticulture. Plants 2022, 11, 2774. [CrossRef]
- Gharabadiyan, F.; Jamali, S.; Komeili, R.H. Determining of root-knot nematode (Meloidogyne javanica) damage function for tomato cultivars, J Agric Sci Belgrade 2013, 58 (2), 147-157. [CrossRef]
- Tzortzakakis, E.A.; da Conceicao, I.L.P.M.; dos Santos, M.C.V.; Abrantes, I.M.D.O. Root-knot nematodes (Meloidogyne spp.) in Greece. Hell Plant Prot J 2011, 4, 25-30.
- Nyczepir, A.P.; Thomas, S.H. Current and future management strategies in intensive crop production systems. In Root-knot nematodes Wallingford UK: CABI, 2009. 412-443.
- Riyaz, M.; Mathew, P.; Zuber, S.M.; Rather, G.A. Botanical pesticides for an eco-friendly and sustainable agriculture: New challenges and prospects,” In Sustainable agriculture (Cham: Springer) 2022, 69–96.
- Abd-Elgawad, M.M.M. Biological control agents of plant-parasitic nematodes. Egypt J Biol Pest Control 2016, 26(2), 423–429.
- Almaghrabi, O.A.; Massoud, S.I.; Abdelmoneim, T.S. Influence of inoculation with plant growth promoting rhizobacteria (PGPR) on tomato plant growth and nematode reproduction under greenhouse conditions. Saudi J Biol Sci 2013, 20(1), 57–61. [CrossRef]
- Khan, F.; Asif, M.; Khan, A.; Tariq, M.; Siddiqui, M.A. Screening of carrot cultivars against root-knot nematode Meloidogyne incognita. Ind Phytopath 2018, 71, 415–421. [CrossRef]
- Ali, W.M.; Abdel-Mageed, M.A.; Hegazy, M.G.A.; Abou-Shlell, M.K.; Sultan, S.M.E.; Salama, E.A.A.; Yousef, A.F. Biocontrol agent of root-knot nematode Meloidogyne javanica and root-rot fungi, Fusarium solani in okra morphological, anatomical characteristics and productivity under greenhouse conditions. Sci Rep 2023, 13, 11103. [CrossRef]
- Khalil, M.S.E.D.H.; Allam, A.F.G.; Barakat, A.S.T. Nematicidal activity of some biopesticide agents and microorganisms against root-knot nematode on tomato plants under greenhouse conditions. J Plant Prot Res 2012, 52(1), 47-52. [CrossRef]
- Khan, R.A.A.; Najeeb, S; Mao, Z.; Ling, J.; Yang, Y.; Li, Y.; Xie, B. Bioactive secondary metabolites from Trichoderma spp. against phytopathogenic bacteria and root-knot nematode. Microorganisms 2020, 8, 401. [CrossRef]
- Naz, I.; Khan, R.A.A.; Masood, T.; Baig, A.; Siddique, I.; Haq, S. Biological control of root knot nematode, Meloidogyne incognita, in vitro, greenhouse and field in cucumber. Biocontrol 2021, 152, 104429. [CrossRef]
- Affokpon, A.; Coyne, D.L.; Lawouin, L.; Tossou, C.; Agbede, R.D.; Coosemans J. Effectiveness of native West African arbuscular mycorrhizal fungi in protecting vegetable crops against root-knot nematodes. Biol Fertil Soils 2021, 47(2), 207–217. [CrossRef]
- Campos, M.A.S. Bioprotection by arbuscular mycorrhizal fungi in plants infected with Meloidogyne nematodes: A sustainable alternative. Crop Prot 2020, 135(105203):105203. article. [CrossRef]
- Kolawole, G.O.; Haastrup, T.M.; Olabiyi, T.I. Can arbuscular mycorrhiza fungi and NPK fertilizer suppress nematodes and improve tuber yield of yam (Dioscorea rotundata ‘cv’ ewuru)? Eurasian J Soil Sci 2018, 7(2), 181–186. [CrossRef]
- Odeyemi, I.S.; Afolami, S.O.; Sosanya, O.S. Effect of Glomus mosseae (arbuscular mycorrhizal fungus) on host-parasite relationship of Meloidogyne incognita (Southern root-knot nematode) on four improved cowpea varieties. J Plant Prot Res 2010, 50(3), 320–325. [CrossRef]
- Rouphael, Y.; Franken, P. ; Schneider, C. ; Schwarz, D.; Giorannetti, M.; Agnolucci, M.; Pascale, S.; Bonini, P.; Colla, G. Arbuscular mycorrhizal fungi act as biostimulants in horticultural crops. Sci Hortic 2015, 196, 91–108. [CrossRef]
- Udo, I.A.; Uguru, M.I.; Ogbuji; R.O. Pathogenicity of Meloidogyne incognita race 1 on tomato as influenced by different arbuscular mycorrhizal fungi and bioformulated Paecilomyces lilacinus in a dysteric cambisol soil. J Plant Prot Res 2013, 53(1), 71–78. [CrossRef]
- Ahmad, G.; Nishat, Y.; Ansari, M.; Khan, A.; Haris, M.; Khan, A.A. Eco-friendly approaches for the alleviation of root-knot nematodes. In: Plant growth-promoting microbes for sustainable biotic and abiotic stress management. Springer, Cham, 2021, 557–575.
- Li, J.; Zou, C.; Xu, J.; Ji, X.; Niu, X.; Yang, J.; Zhang, K.Q. Molecular mechanisms of nematode-nematophagous microbe interactions: basis for biological control of plant-parasitic nematodes. Annu Rev Phytopathol 2015, 53, 67–95. [CrossRef]
- Tiwari, S.; Pandey, S.; Singh Chauhan, P.; Pandey, R. Biocontrol agents in co-inoculation manages root knot nematode [Meloidogyne incognita (Kofoid & White) Chitwood] and enhances essential oil content in Ocimum basilicum L. J Ind Crop Prod 2017, 97, 292–301. [CrossRef]
- Lee, Y.S.; Kim, K.Y. Antagonistic potential of Bacillus pumilus L1 against root-knot nematode, Meloidogyne arenaria. J Phytopathol 2016, 164, 29–39. [CrossRef]
- Mendoza, A.R.; Kiewnick, S.; Sikora, R.A. In vitro activity of Bacillus firmus against the burrowing nematode Radopholus similis, the root-knot nematode Meloidogyne incognita and the stem nematode Ditylenchus dipsaci. Biocontrol Sci Technol 2008, 18, 377–389. [CrossRef]
- Turatto, M.F.; Dourado, F.D.S.; Zilli, J.E.; Botelho, G.R. Control potential of Meloidogyne javanica and Ditylenchus spp. using fluorescent Pseudomonas and Bacillus spp. Braz J Microbiol 2018, 49, 54–59. [CrossRef]
- Xiang, N.; Lawrence, K.S.; Kloepper, J.W.; Donald, P.A.; McInroy, J.A.; Lawrence, G.W. Biological control of Meloidogyne incognita by spore-forming plant growth-promoting rhizobacteria on cotton. Plant Dis 2017, 101, 774–784. [CrossRef]
- Abbasi, M.; Ahmed, N.; Zaki, M.; Shuakat, S.; Khan, D. Potential of Bacillus species against Meloidogyne javanica parasitizing eggplant (Solanum melongena L.) and induced biochemical changes. Plant Soil 2014, 375, 159–173. [CrossRef]
- Metwally, W.E.; Mostafa, F.A.M.; Refaei, A.R. In vitro study on the antagonistic activity of different native isolates of rhizobacteria against Meloidogyne incognita. Egypt J Agronematol 2015, 14, 1–9. [CrossRef]
- Mnif, I.; Ghribi, D. Review lipopeptides biosurfactants: Mean classes and new insights for industrial, biomedical, and environmental applications. Biopolymers 2015, 104, 129-47. [CrossRef]
- Bais, H.P.; Weir, T.L.; Perry, L.G.; Gilroy, S.; Vivanco, J.M. The Role of root exudates in rhizosphere interactions with plants and other organisms. Annu Rev Plant Biol 2006, 57, 233–66. [CrossRef]
- Berendsen, R.L.; Pieterse, C.M.J.; Bakker, P.A.H.M. The rhizosphere microbiome and plant health. Trends Plant Sci 2012, 17, 478–86. [CrossRef]
- Raaijmakers, J.M.; De Bruijn, I.; Nybroe, O.; Ongena, M. Natural functions of lipopeptides from Bacillus and Pseudomonas: more than surfactants and antibiotics. FEMS Microbiol Rev 2010, 34, 1037–62. [CrossRef]
- Ling, N.; Zhu, C.; Xue, C.; Chen, H.; Duan, Y.; Peng, C.; Guo, S.; Shen, Q. Insight into how organic amendments can shape the soil microbiome in long-term field experiments as revealed by network analysis. Soil Biol Biochem 2016, 99, 137–49. [CrossRef]
- Mendes, L.W.; Tsai, S.M.; Navarrete, A.A.; de Hollander, M.; van Veen, J.A.; Kuramae, E.E. Soil-borne microbiome: Linking diversity to function. Microb Ecol 2015, 70, 255–65. [CrossRef]
- Berendsen, R.L.; Vismans, G.; Yu, K.; Song, Y.; de Jonge, R.; Burgman, W.P.; Burmølle, M.; Herschend, J.; Bakker, P.A.H.M.; Pieterse, C.M.J. Disease-induced assemblage of a plant-beneficial bacterial consortium. ISME J 2018, 12, 1496–507. [CrossRef]
- Stringlis, I.A.; Proietti, S.; Hickman, R.; Van Verk, M.C.; Zamioudis, C.; Pieterse, C.M.J. Root transcriptional dynamics induced by beneficial rhizobacteria and microbial immune elicitors reveal signatures of adaptation to mutualists. Plant J 2018, 93, 166–80. [CrossRef]
- Tsolakidou, M-D.; Stringlis, I.A.; Fanega-Sleziak, N.; Papageorgiou, S.; Tsalakou, A.; Pantelides, I.S. Rhizosphere-enriched microbes as a pool to design synthetic communities for reproducible beneficial outputs, FEMS Microbiology Ecology 2019, 95(10), 138. [CrossRef]
- Oliveira, D.F.; Santos Júnior, H.M.; Nunes, A.S.; Campos, V.P.; Pinho, R.S.; Gajo, G.C. Purification and identification of metabolites produced by Bacillus cereus and B. subtilis active against Meloidogyne exigua, and their in silico interaction with a putative phosphoribosyltransferase from M. incognita. An Acad Bras Cienc 2014, 86(2), 525-538. [CrossRef]
- Cao, G.T.; Dai, B.; Wang, K.L.; Yan, Y.; Xu, Y.L.; Wang, Y.X.; Yang, C.M. Bacillus licheniformis, a potential probiotic, inhibits obesity by modulating colonic microflora in C57BL/6J mice model. J Appl Microbiol 2019, 127(3), 880-888. [CrossRef]
- Engelbrecht, G.; Claassens, S.; Mienie, C.M.S.; Fourie, H. Filtrates of mixed Bacillus spp. inhibit second-stage juvenile motility of root-knot nematodes Rhizosphere 2022, 22, 10052. [CrossRef]
- Moghaddam, R.M.; Moghaddam, M.E.; Ravari, B.S.; Rouhani, H. The first report of Bacillus pumilus influence against Meloidogyne javanica in Iran. JCP 2014, 3(1), 105-112. [CrossRef]
- Sansinenea, E.; Ortiz, A. Secondary metabolites of soil Bacillus spp. Biotechnol Lett 2011, 33(8), 1523-38. [CrossRef]
- El-Nagdi, W.M.A.; Abd-El-Khair, H. Biological control of Meloidogyne incognita and Fusarium solani in dry common bean in the field Arch. Phytopath Plant Prot 2014, 47(4), 388-397. [CrossRef]
- Munawar, M.; Khan, S.A.; Javed, N.; Ul Haq, I.; Gondal, A.S. Bio-management of tomato wilt complex caused by Meloidogyne incognita and Fusarium oxysporum f. sp. lycopersici. Nematology 2015, 17(4),479-485. [CrossRef]
- Patil, J.A.; Yadav, S.; Kumar, A. Management of root-knot nematode, Meloidogyne incognita and soil borne fungus, Fusarium oxysporum in cucumber using three bioagents under polyhouse conditions, Saudi J Biol Sci 2021, 28(12), 7006-7011. [CrossRef]
- Shi, X .; Qiao, K.; Li, B.; Zhang, S. Integrated management of Meloidogyne incognita and Fusarium oxysporum in cucumber by combined application of abamectin and fludioxonil Crop Protection 2019, 126, 104922. [CrossRef]
- Chialva, M.; Zouari, I.; Salvioli, A.; Novero, M.; Vrebalov, J.; Giovannoni, J.J.; Bonfante, P. Gr and hp-1 tomato mutants unveil unprecedented interactions between arbuscularmycorrhizal symbiosis and fruit ripening. Planta 2016, 244, 155–65. [CrossRef]
- Antoniou, A.; Tsolakidou, M-D.; Stringlis, I.A.; Pantelides, I.S. Rhizosphere microbiome recruited from a suppressive compost improves plant fitness and increases protection against vascular wilt pathogens of tomato. Front Plant Sci 2017, 8, 2022. [CrossRef]
- Baermann, G. Eine einfache Methode zur Auffindung von Anklostomum (Nematoden) Larven in Erdproben. – Tijdschr Diergeneeskd 1917, 57, 131–137.
- Abd-Elgawad, M.; Askary, T. Impact of Phytonematodes on Agriculture Ecology. In Biocontrol Agents of Phytonematodes. Askary T.H., Martinelli P.R.P., editors. CAB International Wallingford; Wallingford, UK 2015, 3–49.
- Phani, V.; Khan, M.R.; Dutta, T.K. Plant-parasitic nematodes as a potential threat to protected agriculture: Current status and management options. Crop Prot 2021, 144, 105573. [CrossRef]
- Sikora, R.A.; Fernandez, E. Nematode parasites of vegetables. In Plant Parasitic Nematodes in Subtropical and Tropical Agriculture. CABI Publishing; Wallingford, UK 2005, 319–392.
- Abd-Elgawad, M.M. Optimizing safe approaches to manage plant-parasitic nematodes. Plants 2021, 10,1911. [CrossRef]
- Poveda, J.; Abril-Urias, P.; Escobar, C. Biological control of plant-parasitic nematodes by filamentous fungi inducers of resistance: Trichoderma, mycorrhizal and endophytic fungi. Front Microbiol 2020, 11,992. [CrossRef]
- Zhou, D.; Feng, H.; Schuelke, T.; De Santiago, A.; Zhang, Q.; Zhang, J.; Luo, C.; Wei, L. Rhizosphere Microbiomes from Root Knot Nematode Non-infested Plants Suppress Nematode Infection. Microb Ecol 2019, 78, 470–481. [CrossRef]
- Gamalero, E.; Glick, B.R. The Use of Plant Growth-Promoting Bacteria to Prevent Nematode Damage to Plants. Biology (Basel) 2020, 9(11), 381. [CrossRef]
- Kamalanathan, V.; Sevugapperumal, N.; Nallusamy, S. Antagonistic Bacteria Bacillus velezensis VB7 Possess Nematicidal Action and Induce an Immune Response to Suppress the Infection of Root-Knot Nematode (RKN) in Tomato. Genes (Basel) 2023, 25;14(7), 1335. [CrossRef]
- Mhatre, P.H.; Karthik, C.; Kadirvelu, K.; Divya, K.; Venkatasalam, E.; Srinivasan, S.; Ramkumar, G.; Saranya, C.; Shanmuganathan, R. Plant growth promoting rhizobacteria (PGPR): A potential alternative tool for nematodes bio-control. Biocat Agricul Biotechnol 2019, 17, 119–128. [CrossRef]
- Sidhu, H.S. Potential of plant growth-promoting rhizobacteria in the management of nematodes: a review. J Entomol Zool Stud 2018, 6(3), 1536-1545.
- Wharton, D. Nematode eggshells. Parasitology 1980, 81, 447–463. [CrossRef]
- Ray, S.; Reddigarim, S.R.; Jansma, P.L.; Allen, R.; Hussey, R.S. Immunocytochemical analysis of the stage-specific distribution of collagen in the cuticle of Meloidogyne incognita. Fund Appl Nematol 1996, 19, 71–78.
- Khanna, K.; Kohli, S.K.; Ohri, P.; Bhardwaj, R. Plants-nematodes-microbes crosstalk within soil: A trade-off among friends or foes, Microbiological Research 2021, 248, 126755, ISSN 0944-5013. [CrossRef]
- Krechel, A.; Faupel, A.; Hallmann, J.; Ulrich, A.; Berg, G. Potato-associated bacteria and their antagonistic potential towards plant-pathogenic fungi and the plant-parasitic nematode Meloidogyne incognita (Kofoid & White) Chitwood Can J Microbiol 2002, 48(9), 772-786. [CrossRef]
- El-Hadad, M.E.; Mustafa, M.I.; Selim, S.M.; El-Tayeb, T.S.; Mahgoob, A.E.A.; Aziz, N.H.A. The nematicidal effect of some bacterial biofertilizers on Meloidogyne incognita in sandy soil. Braz J Microbiol 2011, 42(1), 105-113. [CrossRef]
- Siddiqui, I.A.; Shaukat, S.S. Rhizobacteria-mediated induction of systemic resistance (ISR) in tomato against Meloidogyne javanica. J Phytopathol 2002, 150, 469–473. [CrossRef]
- Sikora, R.A. Interrelationship between plant health promoting rhizobacteria, plant parasitic nematodes and soil microorganisms. Med Fac Landbouww Rijksuniv Gent 1988, 53/2b, 867-878.
- Kloepper, J.W.; Ryu, C.-M.; Zhang, S. Induced systemic resistance and promotion of plant growth by Bacillus spp. Phytopathology 2004, 94, 1259-1266. [CrossRef]
- Siddiqui, Z.A.; Mahmood I. Role of bacteria in the management of plant parasitic nematodes: a review Bioresour Technol 1999, 69, 167-179. [CrossRef]
- Ramamoorthy, V.; Viswanathan, R.; Raguchander, T.; Prakasam, V.; Samiyappan R. Induction by systemic resistance by plant growth promoting rhizobacteria in crop plants against pests and diseases Crop Prot 2001, 20, 1-11. [CrossRef]
- Van Loon, L.C.; Geraats, B.P.J.; Linthorst, H.J.M. Ethylene as a modulator of disease resistance in plants. Trends Plant Sci 2006, 11, 184–191. [CrossRef]
- Raaijmakers, J.M.; De Bruijn, I.; Nybroe, O.; Ongena, M. Natural functions of lipopeptides from Bacillus and Pseudomonas: more than surfactants and antibiotics. FEMS Microbiol Rev 2010, 34, 1037–62. [CrossRef]
- Falardeau, J.; Wise, C.; Novitsky, L.; Avis, T.J. Ecological and mechanistic insights into the direct and indirect antimicrobial properties of Bacillus subtilis lipopeptides on plant pathogens. J Chem Ecol 2013, 39, 869–78. [CrossRef]
- Gopalakrishnan, S.; Sathya, A.; Vijayabharathi, R.; Varshney, R.K.; Gowda, C.L.L.; Krishnamurthy, L. Plant growth promoting rhizobia: challenges and opportunities 3 Biotech, 2015, 355-377. [CrossRef]
- Penrose, D.M.; Glick, B.R. Methods for isolating and characterizing ACC deaminase-containing plant growth-promoting rhizobacteria. Physiol Plant 2003, 118, 10–5. [CrossRef]
- Zamioudis, C.; Mastranesti, P.; Dhonukshe, P.; Blilou, I.; Pieterse, C.M.J. Unraveling root developmental programs initiated by beneficial Pseudomonas spp. bacteria. Plant Physiol 2013, 162, 304–18. [CrossRef]
- Asari, S.; Tarkowská, D.; Rolčík, J.; Novák, O.; Palmero, D.V.; Bejai, S.; Meijer, J. Analysis of plant growth-promoting properties of Bacillus amyloliquefaciens UCMB5113 using Arabidopsis thaliana as host plant. Planta 2017, 245(1), 15-30. [CrossRef]
- Arora, N.K.; Tewari, S.; Singh R. Multifaceted plant-associated microbes and their mechanisms diminish the concept of direct and indirect PGPRs. In N.K. Arora (Ed.), Plant Microbe Symbiosis: Fundamentals and Advances Springer 2013, 411-449.
- Shaikh, S.; Saraf M. Biofortification of Triticum aestivum through the inoculation of zinc solubilizing plant growth promoting rhizobacteria in field experiment. Biocatal Agric Biotechnol 2016, 9, 120-126. [CrossRef]
- Glick, B.R. Using soil bacteria to facilitate phytoremediation. Biotechnol Adv 2010, 367-374. [CrossRef]
- Khan, M.R.; Khan, M.M.; Anwer, M.A.; Haque Z. Laboratory and field performance of some soil bacteria used as seed treatments on Meloidogyne incognita in chickpea. Nematol Mediterr 2012, 40, 143-151.
- Rashad, F.M.; Fathy, H.M.; El-Zayat, A.S.; Elghonaimy A.M. Isolation and characterization of multifunctional Streptomyces species with antimicrobial, nematicidal and phytohormone activities from marine environments. Egypt Microbiol Res 2015, 175, 34-47. [CrossRef]
- Ruanpanun, P.; Tangchitsomkid, N.; Hyde, K.D.; Lumyong S. Actinomycetes and fungi isolated from plant-parasitic nematode infested soils: screening of the effective biocontrol potential, indole-3-acetic acid and siderophore production World J Microbiol Biotechnol 2010, 1569-1578. [CrossRef]
- Van Loon, L.C.; Bakker, P.A.H.M.; Pieterse C.M.J. Systemic resistance induced by rhizosphere bacteria Annu Rev Phytopathol 1988, 453-483.
- Nascimento, F.X.; Vicente, C.S.; Barbosa, P.; Espada, M.; Glick, B.R.; Mota, M.; Oliveira, S. Evidence for the involvement of ACC deaminase from Pseudomonas putida UW4 in the biocontrol of pine wilt disease caused by Bursaphelenchus xylophilus. Biol Control 2013, 58, 427–433. [CrossRef]
Figure 1.
Impact of SynComs and Pochar on tomato growth and nematode protection. (A), Fresh weight of aerial parts (gr). (B), Dry weight of aerial parts (gr). (C), Plant height (cm). (D), Fresh weight of root (gr). Vertical bars indicate the SE of mean (n = 10 replicates). Different letters indicate statistically significant differences: P < 0.05; one-way ANOVA, Tukey’s test.
Figure 1.
Impact of SynComs and Pochar on tomato growth and nematode protection. (A), Fresh weight of aerial parts (gr). (B), Dry weight of aerial parts (gr). (C), Plant height (cm). (D), Fresh weight of root (gr). Vertical bars indicate the SE of mean (n = 10 replicates). Different letters indicate statistically significant differences: P < 0.05; one-way ANOVA, Tukey’s test.
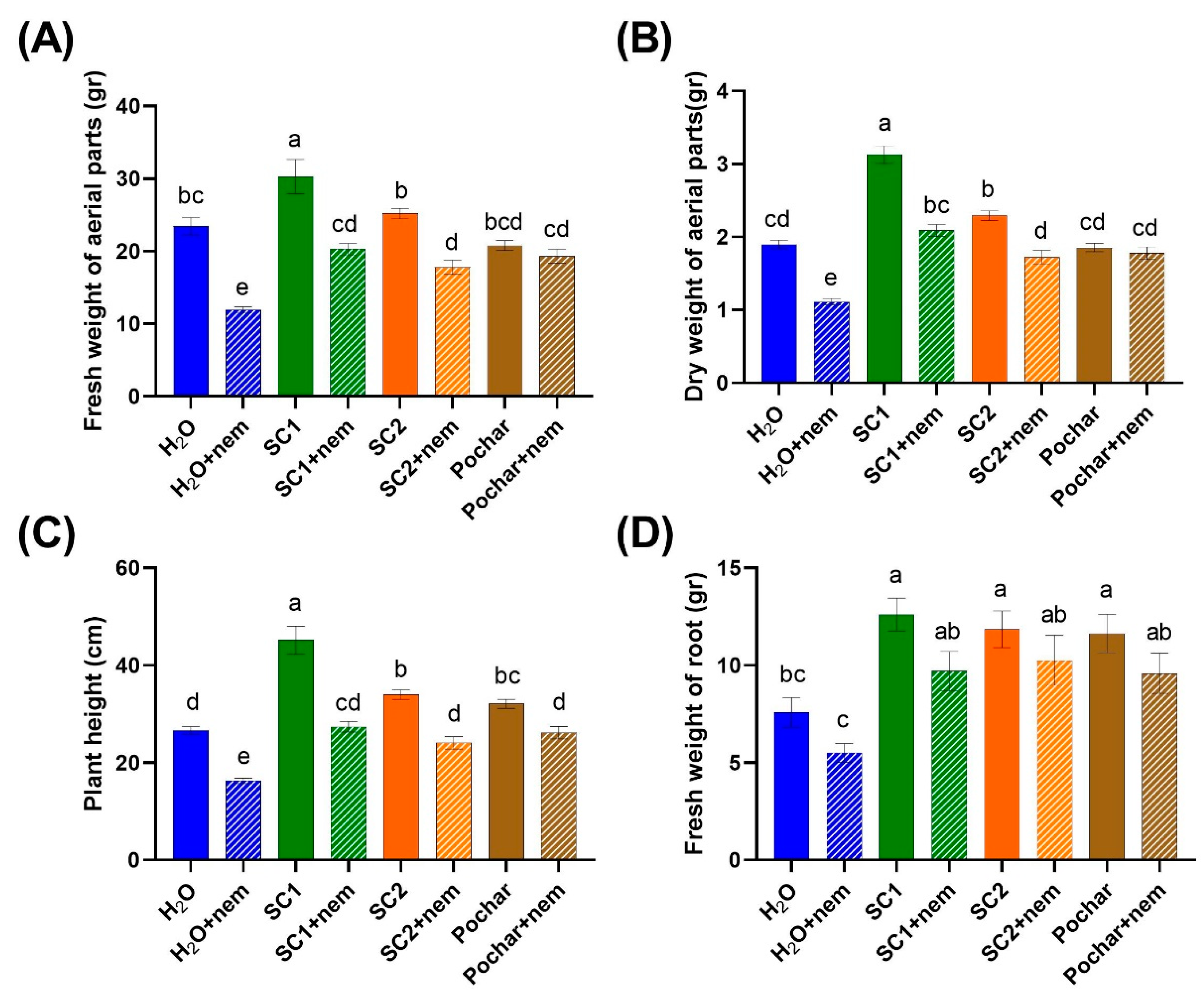
Figure 2.
This is a figure. Schemes follow another
format. If there are multiple panels, they should be listed as: (a)
Description of what is contained in the first panel; (b) Description of
what is contained in the second panel. Figures
should be placed in the main text near to the first time they are cited.
Figure 2.
This is a figure. Schemes follow another
format. If there are multiple panels, they should be listed as: (a)
Description of what is contained in the first panel; (b) Description of
what is contained in the second panel. Figures
should be placed in the main text near to the first time they are cited.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



