
Submitted:
04 April 2024
Posted:
09 April 2024
You are already at the latest version
Abstract
Brain imaging studies have recently provided some evidence in favour of covert cognitive processes ongoing in patients with disorders of consciousness (DoC) (e.g., minimally consciousness states, vegetative state/unresponsive wakefulness syndrome) engaged in passive sensory stimulation or active tasks such as motor imagery. In this exploratory study, we used transcranial magnetic stimulation (TMS) of the motor cortex to assess modulations of corticospinal excitability induced by action observation in eleven patients with DoC. Action observation is known to facilitate corticospinal excitability in healthy subjects, unveiling how the observer’s motor system maps others’ actions onto her/his motor repertoire. Additional stimuli were non-biological motion and acoustic startle stimuli, provided that sudden and loud acoustic stimulation is known to lower corticospinal excitability in healthy subjects. Results indicate that some form of motor resonance is spared in a subset of patients with DoC with some significant difference between biological and non-biological motion stimuli. However, there was no covariation between corticospinal excitability and the type of DoC diagnosis (i.e., whether VS/UWS or MCS). Similarly, no covariation was detected with clinical changes between admission and discharge in clinical outcome measures. Both motor resonance and biological/non-biological motion discrimination correlated with the amplitude of the N20 somatosensory evoked potentials following the stimulation of the median nerve at the wrist (i.e., the temporal marker signalling the activation of the contralateral primary somatosensory cortex). Moreover, the startle-evoking stimulus produced an anomalous increase in corticospinal excitability, suggesting a functional dissociation between cortical and subcortical circuits in patients with DoC. Further work is needed to better comprehend the conditions in which corticospinal facilitation occurs and whether and how they may relate to individual clinical parameters.
Keywords:
Disorders of consciousness
; minimally consciousness state
; vegetative state/unresponsiveness wakefulness syndrome
; transcranial magnetic stimulation
; action observation
1. Introduction
Disorders of consciousness (DoC) include a spectrum of conditions characterized by different levels of consciousness impairment, often secondary to vascular, anoxic, metabolic, or traumatic brain injuries. Because consciousness implies both the level of arousal and the content of consciousness (e.g. functions such as attention, memory, and volition), DoC can independently affect these two domains [1,2]. For example, both arousal and the contents of consciousness are absent in a “coma”. By contrast, the “vegetative state” (VS, better known as unresponsive wakefulness syndrome (UWS) [3]), corresponds to a condition of wakeful unawareness, in which, despite spared sleep-wake cycles, patients show no conscious interaction with the surrounding environment [4]. Further along the spectrum of impaired consciousness, the “minimally consciousness state” (MCS) is defined as a condition of minimal and inconsistent – though definite – evidence of awareness, including occasional responses to external stimuli (e.g. making pursuing eye movements tracking the examiner finger, execution of basic verbal commands) [5].
In recent years, several researchers investigated the neural correlates of consciousness [6,7] to identify a neural-based definition of consciousness. Several conflicting theories about the neural correlates of consciousness have been proposed [8] and no conclusions have yet been reached. One of these theories relies on brain complexity as a critical requirement of consciousness [9,10]. According to this hypothesis, consciousness is sustained by patterns of neural activity distributed across multiple brain regions and differentiated in space and time. Support for this principle comes from studies using transcranial magnetic stimulation (TMS) coupled with electroencephalographic (EEG) recordings. In patients with VS/UWS, TMS pulses elicit only localized event-related potentials (ERPs). On the other hand, more diffused activity can be elicited in patients with MCS and fully conscious healthy subjects [11,12,13].
Beyond the debate on philosophical and neural theories of consciousness, however, compelling questions with relevant medical and ethical implications remain open when we stand at the bedside of a DoC patient: To what extent can unconscious patients perceive signals from the world surrounding them? Could some kinds of covert, though active, cognitive processing exist in these patients, escaping from behavioural and neurological assessments? The definition provided above, assuming that the contents of consciousness are absent in VS/UWS and profoundly impaired in MCS, would implicitly rule out any information processing of the external world in patients with DoC.
Over the last decades, studies employing brain imaging techniques depicted a very different scenario and provided some evidence that residual cognitive processes could be intact in patients with DoC [2]. A recent meta-analysis of studies looking for consciousness-related brain activations in patients with DoC has shown that several cortical areas become activated in some patients when they are engaged in active or passive tasks [14].
Passive paradigms are disparate, including auditory presentation of isolated words [15], and sentences [16,17,18], self-related stimuli (e.g. listening to one own name spoken by a familiar voice) [19,20,21] and emotional stimuli (e.g. pain-evoked cries) [22], as well as visual presentation of complex stimuli such as faces [23]. Overall, these studies demonstrated that residual cognitive processing is detectable at the neural level in some patients with DoC and activation patterns are very similar to those observed in healthy subjects [20,21,22,23]. However, brain activation has been often observed to scale with the severity of consciousness impairment (e.g., between VS/UWS and MCS patients) [16,22], suggesting that functional brain imaging might help classify patients with DoC.
Studies employing active paradigms showed even more striking results. A seminal work in this field reported the case of a young woman with VS/UWS who was asked to imagine playing tennis or walking through her house while her brain activity was recorded using functional magnetic resonance imaging (fMRI) [24]. The pattern of brain activity resulted indistinguishable from healthy controls, involving the supplementary motor area when the patient had to imagine playing tennis and the parahippocampal gyrus, the posterior parietal cortex, and the lateral premotor cortex when she was asked to imagine walking through her house. Similarly, when asked to move either the right or left hand, two patients diagnosed with VS/UWS showed functional changes in the contralateral premotor cortex [25]. Notably, volitional modulation of brain activity during motor imagery is present only in a subset of patients with DoC [26].
In addition to expanding our knowledge of the neural underpinnings of consciousness, studies addressing brain functioning in DoC are crucial to defining reliable prognostic criteria. Currently, the N20 component of the Somatosensory Evoked Potential (SEP) recorded from C3/C4 electrodes after electrical stimulation of the median nerve at the wrist, represents the most robust electrophysiological measure for outcome predictions in patients with DoC. Absent or low-amplitude N20 is associated with poor prognosis in patients with DoC, specifically after anoxic injury [27,28,29,30,31,32,33]. However, the presence of N20 is not as effective in predicting a favourable outcome [34,35]. Despite some evidence claiming that spared cognitive ERPs, such as mismatch negativity, could predict favourable outcomes, results are still quite heterogeneous [35].
Although still poorly employed in this field, TMS is a versatile tool that might help clarify the dynamics of residual cognitive processing in patients with DoC. By delivering TMS pulses over the primary motor cortex it is possible to probe the excitability of the corticospinal system by measuring the size of electromyographic responses known as motor evoked potentials (MEPs). Corticospinal excitability reflects the influence of the overall input to the primary motor cortex and the spinal cord [36]. Among many sources of modulation, corticospinal excitability is enhanced during action observation, due to a mirror neuron network involving frontal and parietal cortices [37] – an effect known as motor resonance [38] A crucial property of the mirror neuron network is that it shows similar activity when actions are self-generated and observed in other individuals [37,39]. Individual mirror neurons are often tuned on specific actions (both during action execution and observation) [40] and the pattern of corticospinal excitability facilitation during action observation reflects muscle activity in the observed actions [38]. Mapping observed actions into motor coordinates may underlie the ability to understand and predict what our conspecific are doing, setting the basis for any form of social interaction. In patients with DoC, motor resonance may thus provide a window into the latent processing of social cues potentially escaping clinical observation.
In this direction are the results of a previous study, where facilitation of corticospinal excitability was detected in a subset of patients with VS/UWS who were encouraged to observe and imitate an action presented by the experimenter [41]. Remarkably, patients showing augmented corticospinal excitability during action observation also improved their consciousness level during follow-up observations after 28 weeks. These results suggest that the assessment of motor resonance in patients with DoC may convey important information for outcome prediction. However, an important limitation of this study was that patients were presented only with intransitive actions (i.e., thumb abduction). Because transitive actions are richer in terms of motor cues, involving a complex relationship between object and effector, we propose that they would constitute a more suitable approach when targeting covert processes of sensorimotor integration in patients with DoC. In addition, corticospinal facilitation during action observation in these patients might be due to non-specific arousal secondary to motion within the visual field rather than to the specific encoding of goal-directed actions. To clarify this confounding factor, one needs to also assess the effect of non-biological motion observation on corticospinal excitability.
Here, we tested corticospinal excitability in patients with DoC while they were presented with transitive actions and non-biological motion. In addition, we assessed corticospinal excitability modulation by sudden loud acoustic noises (i.e., auditory startle paradigm). Like action observation, startle stimuli are known to produce systematic changes in corticospinal excitability (mainly suppression excitability [42]). Unlike the mirror neuron system, the neural substrate of the startle response is located subcortically in the brain stem [43]. Corticospinal modulation by cortical (action observation) and subcortical (startle) neural substrates was then put with the amount of spared somatosensory input, as indexed by SEPs. Finally, we examined the relationship between the presence of detectable corticospinal excitability at least in one experimental condition and the type of DoC diagnosis (i.e., whether VS/UWS or MCS) as well as possible clinical changes between admission and discharge.
2. Materials and Methods
2.1. Participants
To be included in the study, patients had to be diagnosed with either VS/UWS or MCS due to vascular or traumatic accidents that occurred within the previous six months. Patients were classified as VS or MCS based on the JFK Coma Recovery Scale-Revised (CRS-R) and on internationally established criteria [5,44], according to which emergence from MCS is marked by the detection of a reliable yes-no communication system and/or functional object use. The CRS-R consists of 23 hierarchically organized items parceled into six subscales designed to assess auditory, visual, motor, language, and arousal functions. Weighted scores are assigned to reflect the presence or absence of specific responses, ranging from brain stem reflexes to cognitively mediated behaviours. The total score can be used to gauge the general trajectory of recovery over time as higher scores reflect progressively increasing levels of cognitive function. The transition from VS/UWS to MCS depends on the score obtained in the subtests of “communication” and “functional object use”.
VS/UWS patients fulfilled the following diagnostic criteria: a) no evidence of awareness of self or environment and an inability to interact with others; b) no evidence of sustained, reproducible, purposeful, or voluntary behavioural responses to visual, auditory, tactile, or noxious stimuli; no evidence of language comprehension or expression; c) intermittent wakefulness; d) no presence of epileptic crises and e) bowel and bladder incontinence.
The diagnostic criteria for MCS were a) ability to follow simple commands; b) presence of gestural or verbal yes/no responses (regardless of accuracy); c) presence of intelligible verbalization; and d) presence of purposeful behaviour, including movements or affective behaviours occurring in contingent relation to relevant environmental stimuli and not due to reflexive activity.
Patients with contraindications to TMS [45] were excluded from the study, as well as those fulfilling the following exclusion criteria: decompressive craniotomy, bilaterally absent brainstem auditory evoked potentials (BAEPs) or visually evoked potentials (VEPs) and unviability of the corticospinal pathway as indexed by the absence of motor evoked potentials (MEPs).
Eleven patients (7 females) aged between 46 and 81 years (mean: 70.27 years, SD: 11.21) participated in the study. Six were diagnosed with VS/UWS and 5 with MCS at the time of the inclusion in the study. Demographic and clinical information are reported in Table 1.
The study was conducted according to the Declaration of Helsinki and approved by the local Ethics Committee of the Siena Health Authority as part of a larger brain stimulation program in VS patients (protocol code: Brainsight; approval number: EME_1144_0_1) [46]. We obtained informed consent from each patient’s legal surrogate.
2.2. Clinical Assessment
Patients included in the study were assessed with the Coma Recovery Scale-Revised (CRS-R) during a 5-day monitoring period the week before they underwent the experimental procedures. The following clinical scales were also carried out to evaluate outcomes.
Functional Independence Measure (FIM) [47]. The FIM is one of the most widely used methods for assessing the basic quality of daily living activities in persons with a disability. It includes 18 items designed to determine the amount of assistance required for a person with a disability to perform basic life activities safely and effectively. Each item is rated on a scale of 1-7 (1 requires total assistance, 7 independent). The activities include a minimum set of skills related to self-care, sphincter control, transfers, locomotion, communication, and social cognition. The range of the FIM score is from 18 to 126 points. Higher scores indicate a greater level of functional independence, while lower scores indicate greater dependence.
Rancho Level of Cognitive Functioning Scale (LCF) [48]. The LCF scale is based on observations made by the clinician and is used to assess cognitive functioning in post-coma patients. It was developed for use in the planning of treatment, tracking of recovery, and classifying of outcome levels. It consists of eight levels for cognitive functions, ranging from 1 (no answer) to 8 (the patient is alert and oriented, able to recall and integrate past and recent events, and is aware of his situation).
Finally, in the years following the recordings, we also checked the computerized public health databases to verify the eventual moment of death.
2.3. Experimental Procedures
All experimental procedures were carried out at the patient’s bedside. First, we recorded BAEPs, VEPs and median nerve SEPs. BAEPs were recorded by delivering 15 Hz acoustic clicks lasting 0.1 s. The intensity of the stimuli was 100 dB in the tested ear, while 60 dB white noise was presented in the contralateral ear. The active electrode was placed on the examined ear’s lobe, while the reference electrode was placed at the Cz location (according to Jasper’s 10/20 system [49]). The ground electrode was placed at the Fz location.
VEPs were recorded using flashes delivered monocularly through LED goggles. Flashes lasted 5 ms each and were delivered at 1 Hz frequency. The VEPs recording procedure started after 5 minutes of darkness exposure. The active electrode was positioned 5 cm above the inion on the midline (Oz electrode according to the 10/20 Jasper’s system), while the reference electrode was positioned at Fz.
Finally, upper limb SEPs were recorded from C3 and C4 electrodes using electric stimulation of the median nerve at the wrist (stimulus duration, 0.3 ms; frequency, 3 Hz).
Focal TMS was delivered to the non-lesioned hemisphere using a 70 mm figure-of-eight coil connected to a Magstim 200 stimulator (Magstim Co Ltd, Whitland, UK). In the case of diffuse lesions, the less affected side was selected after clinical/neuroradiological examination. MEPs were recorded from the first dorsal interosseus (FDI) muscle contralateral to the site of stimulation using needle electrodes connected to a Neuropack S1 amplifier (Nihon Kohden Corporation, Tokyo, Japan). The sampling frequency was 5000 Hz. The optimal site of stimulation was identified as the point on the scalp where magnetic stimuli elicited the largest MEPs at the lowest stimulation intensity. The resting motor threshold (RMT) was defined as the closest 1% of the stimulator intensity that elicited 5 out of 10 MEPs of at least 50 µV of amplitude [50]. To obtain consistent responses to TMS pulses, during the experiment stimulation intensity was set at 120% RMT. During all experimental procedures, the coil was oriented with its handle pointing downward and backward, 45° away from the midline.
The experimental design included three conditions: (1) action observation, (2) acoustic startle, and (3) pendulum observation. In patients 1, 2 and 7, the pendulum condition was not run due to technical problems. In the action observation condition, patients observed reaching-grasping actions directed towards objects affording a precision grip (e.g., pen, banknote, eyeglass temples). Actions were presented approximately at the centre of the patient’s visual field and were performed live by one of the experimenters. The TMS pulse was delivered at the end of the reaching phase of the movement when the experimenter’s fingers started closing on the object. This timing of TMS administration during action observation generates the maximum facilitation of corticospinal excitability in healthy subjects [51]. In the acoustic startle condition, patients heard a sudden and loud bell tone, which preceded the TMS pulse by 30-60 ms [43]. As done previously [43], trials were spaced by at least 20 s to avoid habituation. In the pendulum condition, TMS was delivered while patients were presented with a pendulum oscillating within their visual field at about 2 m from their frontal plane. The TMS pulse was delivered after the pendulum had completed two or three cycles. The pendulum was a wooden sphere of ~3 cm diameter attached to a post using a string that allowed it to oscillate. The string length was adjusted to ensure the pendulum oscillated within the patient’s visual field, given that the gaze orientation was often constrained by neck posture. No other individuals were present in the patient’s visual field when the TMS pulse was delivered. One block of 12-16 trials was run for each condition. Block order was randomized across subjects. Two baseline blocks of 12-16 trials were recorded before and after the experimental blocks. During baseline recordings, TMS pulses were delivered with patients at rest without any external stimulation.
2.4. Data Analysis
All the analyses were carried out using custom software written in Matlab (Mathworks, Inc., Natick, USA). MEP onsets and offsets were assessed trial-by-trial by visual inspection (Figure 1a) and the area under curve (AUC) was taken as a measure of corticospinal excitability.
Because the patients included in the study were quite clinically heterogeneous, statistical tests were first performed at the individual subject level. Pairwise independent sample permutation tests based on a t-statistic [52] were performed in each patient to assess significant changes in corticospinal excitability across different conditions. The following comparisons were tested: Action observation vs. baseline, acoustic startle vs. baseline, pendulum observation vs. baseline and action observation vs. pendulum observation. The number of permutations was set at 5000.
For group-level analysis, the average AUC in each condition was normalized by expressing it as a percentage of baseline AUC. A one-sample permutation test was used to determine significant changes in corticospinal excitability occurring in each condition compared to the baseline. Due to the limited number of patients, permutation tests were executed with 2048 permutations in the action observation and acoustic startle conditions and 256 permutations in the pendulum observation condition (i.e., all the possible permutations). Additionally, the ratio between the average AUC during action observation and pendulum observation (AO/pendulum ratio) was calculated in each patient. A one-sample permutation test served to evaluate whether this ratio significantly differed from 1 (i.e., i.e., whether action observation determined a significant modulation of corticospinal excitability compared to pendulum observation).
Baseline-normalized AUC in each condition and AO/pendulum ratios were correlated with the N20 amplitude and the CRS-R scores measures obtained from each patient. As in previous studies, the N20 amplitude was considered from its negative peak to the subsequent positive peak (i.e., P25) (Figure 1b) [30]. The Spearman’s rank-order correlation was used for all correlation analyses.
AUC was considered the principal measure of corticospinal excitability. However, peak-to-peak amplitude data were also measured and analysed to corroborate the conclusions drawn from AUC data. Peak-to-peak amplitude is a widely used measure of corticospinal excitability in TMS studies [50]. Therefore, its use may help in framing this work within the current and future literature. All the analyses described above for the AUC data were also performed for peak-to-peak amplitude data.
To evaluate the possible relationship with coma severity, both average and normalized AUCs (in action observation vs. baseline and acoustic startle vs. baseline) were correlated with CRS-R scores. Correlations were not examined in the case of the pendulum condition due to a limited number of observations. Similar analyses were carried out on peak-to-peak amplitude data.
Finally, to verify any covariation with the results of experimental tests, we examined the individual presence of changes in the FIM and LCF scales between admission and discharge from the rehabilitation ward as a function of individual responses to the experimental manipulations. As a further measure of outcome, we report data on the patient’s lifespan.
3. Results
3.1. MEPs: AUC
In 8 patients out of 11, corticospinal excitability (MEPs), as indexed by AUC, was facilitated during action observation compared to baseline (Table 2). However, this difference was significant only in patients 1, 4, 6 and 11 (p = 0.0068, p = 0.0040, p = 0.0004, p = 0.0010, respectively). In patients 6 and 1, the AUC also resulted significantly larger during action observation than during pendulum observation (p <0.0001, p = 0.0002, respectively). Moreover, pendulum observation produced a significant increase in AUC compared to the baseline in patients 4 and 10 (p = 0.0040, p = 0.0004, respectively). In this latter patient, a significant increase of AUC was also observed in the pendulum condition compared with action observation (p = 0.003). The acoustic startle determined a significant increase in AUC compared to baseline in patients 1, 4, 6, 9 and 10 (p = 0.0232, p = 0.0142, p = 0.0104, p = 0.0038, and p <0.0001, respectively) and an opposite modulation in patient 11 (p = 0.0066).
Table 2 lists patients as a function of their CRS-R score to appreciate the relationship between AUC value in the various conditions and coma severity (and clinical diagnosis). Increased corticospinal excitability, as indexed by AUC, was present in patients with very different CRS-R values and both patients with VS/UWS and MCS. Thus, patient 1, with VS/UWS and a CRS-R score of 1, showed activation for both action observation and acoustic startle (the Pendulum condition was not run on this patient); on the other hand, patients with relatively high CRS-R values (e.g., patient 8) failed to show significant activation compared to baseline, although the direction of the data appears in the expected direction. When examined statistically, correlations between CRS-R scores and MEPs normalized AUCs in action observation and acoustic startle were negligible and not significant. In the case of average AUC values, there was a marginally significant correlation between baseline AUC values and CRS-R scores (ρ = 0.59, p = 0.0544) indicating a tendency for lower AUC values in more severe patients. All other correlations were not significant.
At the group level, baseline-normalized corticospinal excitability was facilitated both during action observation and startle response, approaching significance in both cases (p = 0.0645, p = 0.0645, respectively). Despite an average increase of MEPs AUC observed during the pendulum condition compared to baseline, there were no significant changes in corticospinal excitability in this condition. Similarly, the AO/pendulum ratio was not significant.
3.2. MEPs: Peak-to-Peak Amplitude
At the individual patient level, the analysis of MEP’s peak-to-peak amplitude showed similar results as AUC data (Table 3). MEPs peak-to-peak amplitude was significantly larger during action observation compared to baseline in patients 1, 4, 6 and 11 (p = 0.0088, p = 0.0032, p <0.0001, p = 0.0006, respectively). In patients 6 and 11, the peak-to-peak amplitude was also larger during action observation compared to pendulum observation (p <0.0001, p = 0.0004, respectively). A significant increase in peak-to-peak amplitude was observed in pendulum observation compared to baseline in patients 4 and 10 (p = 0.0042, p <0.0001, respectively). Additionally, patient 10 showed significantly higher peak-to-peak amplitude during pendulum observation compared to action observation (p = 0.0034). Finally, in the acoustic startle condition, a significant increase in peak-to-peak amplitude was detected in patients 1, 9, 4, 6 and 10 (p = 0.0204, p = 0.0124, p = 0.0074, p = 0.0032, and p < 0.0001, respectively). In contrast, an opposite modulation occurred in patient 11 (p = 0.0078).
In Table 3, patients are listed as a function of their CRS-R score. No significant correlations were detected between MEPs peak-to-peak amplitudes and CRS-R scores (no ps > .10) for either average or normalized values. Furthermore, like in the case of AUCs, the pattern of data for peak-to-peak amplitudes does not show a definite relationship with the clinical diagnosis.
At the group level, only the baseline normalized peak-to-peak amplitude recorded during the acoustic startle condition significantly differed from 1 (p = 0.0391).
3.3. Relationship between the N20 Component and Corticospinal Excitability (AUC and Peak-to-Peak Amplitude)
At least in one hemisphere, all patients showed a detectable and reproducible N20 component. The visual inspection of the scatterplot representing the relationship of baseline-normalized AUC during action observation and N20 amplitude revealed that patient 1 behaved differently from the rest of the sample in this correlation. This patient was thus considered an outlier and excluded from correlation analysis. After the exclusion of patient 1, the baseline-normalized AUC during action observation was significantly correlated with the N20 amplitude (ρ = 0.83, p = 0.0056). Moreover, the correlation of the AO/pendulum ratio with the N20 amplitude approached significance (ρ = 0.69, p = 0.0694). Conversely, corticospinal excitability in the acoustic startle and pendulum conditions failed to correlate with the N20 amplitude. Moreover, no correlation analysis involving the CRS-R scores as a covariate was significant.
As for peak-to-peak amplitude, no significant correlations were detected between peak-to-peak amplitude and N20 amplitude in any condition.
4. Clinical Data at Admission and Discharge
Table 4 reports the clinical data at admission and discharge as well as lifespan data of the patients recruited for the study, as actually available. Small (1-point) improvements in LCF at discharge were present in patients 2 and 7; both patients did not show any significant sign of corticospinal excitability (MEPs), as indexed by AUC or peak-to-peak amplitude. A 2-point improvement was present in patient 4, who showed signs of corticospinal excitability across all three conditions tested (in terms of MEPs AUC and peak-to-peak amplitude.
As for FIM, only patient 4 showed a five-point improvement during recovery while all other scores were unchanged between admission and discharge.
4. Discussion
In this study, corticospinal excitability was assessed using TMS of the motor cortex in patients with DoC during observation of live-performed goal-directed actions, observation of non-biological motion (an oscillating pendulum) and loud auditory stimulation (startle paradigm). By this approach, we addressed patients’ residual ability to visually discriminate different types of motion (i.e., human motion vs. non-biological motion) and to map human motion into motor coordinates. In addition, we correlated corticospinal excitability changes with residual somatosensory processing, as expressed by the amplitude of the N20 SEP component. Finally, in addition to the modulation produced by cortical networks (i.e., mirror neuron network) we also examined the influence on corticospinal excitability of subcortical networks responsible for the startle response. Results were quite mixed across patients, likely reflecting the heterogeneity of the sample in terms of brain lesions and general clinical conditions. Due to the small number of patients recruited, the results presented here should be considered preliminary to observations with larger sample sizes.
A general increase of corticospinal excitability during action observation was observed compared to baseline recordings, approaching significance at the group level. Nevertheless, at the individual level, corticospinal excitability was facilitated by action observation compared to the baseline condition only in 4 out of 11 patients (three of whom with VS/UWS and one with MCS). In addition, corticospinal excitability was generally larger during action observation than during pendulum observation, with only one exception (Patient 10, MCS). This difference reached significance only in 2 out of 8 patients (namely one with VS/UWS and one with MCS). Pendulum observation did not produce a significant increase in corticospinal excitability at the group level and no significant difference compared to action observation. These results indicate that, in some patients, corticospinal facilitation during action observation may reflect an intact mirror neuron system operating below the level of consciousness rather than a non-specific modulation produced by the presence of motion (biological or not) within the visual field. Results on peak-to-peak amplitude generally matched those on AUCs, particularly in the case of individual data.
However, the presence of individual responders in corticospinal excitability modulation did not correlate with coma severity as well as clinical diagnosis. There were both responders and non-responders among the VS/UWS patients as well as among the MCS patients. Similarly, clinical scales (FIM and LCF) failed to detect a relationship with corticospinal excitability. Indeed, only a few patients showed improvements in these scales during recovery and again these were both responders and non-responders in terms of cortical excitability. Notably, FIM and LCF are among the most widely used scales with excellent psychometric properties [53,54], but they are still coarse, and able to identify only macroscopic changes. Finally, also data on life expectancy failed to show a detectable relationship with responses in corticospinal excitability.
In a previous study with healthy subjects, a loud and sudden noise suppressed corticospinal excitability when it preceded the TMS pulse by 30-60 ms [43]. Because the optimal auditory stimulus for MEP suppression also produced a strong startle response, authors proposed that both these phenomena might arise from the same subcortical neural network, likely located in the brainstem [55] and sending upstream projections to the motor cortex. We aimed to replicate this finding in patients with DoC by presenting a loud bell tone just before TMS delivery. Surprisingly, in our sample, we mostly observed an opposite modulation (i.e., facilitation). As such, a dissociation between cortical and subcortical networks seems to be present in patients who have lost consciousness. However, the results of the current study do not allow us to draw any conclusion about the nature of this dissociation and its clinical relevance. Therefore, further investigations are needed to test this hypothesis.
We failed to detect any correlation between corticospinal excitability and the level of consciousness, as assessed by the CRS-R or performance in clinical scales (FIM and LCF). However, baseline-normalized corticospinal excitability during action observation (and the AO/pendulum ratio) significantly correlated with the N20 amplitude, a measure of temporal activation of the primary somatosensory cortex considered to be of prognostic relevance in DoC [28]. Because we did not carry out a patient follow-up involving measures of consciousness impairment, our results are not conclusive on the prognostic value of corticospinal excitability modulation in DoC. However, recent results advocate in this direction [56]. The significant correlation of corticospinal facilitation during action observation with the amplitude of the N20 component suggests (though indirectly) that some predictive information might be extrapolated using this approach. Interestingly, patients with a larger N20 component also showed a larger AO/pendulum ratio, being thus more able to discriminate between human (biological) and non-biological motion. This result suggests that motor resonance per se but also the ability to discriminate between different types of motion might provide valuable prognostic information. Future studies should disentangle the predictive role of information provided by the modulation of corticospinal excitability across different behavioural contexts.
This work has limitations. First, we did not include a control group of age-matched healthy subjects. However, motor resonance constitutes a robust and reproducible finding in healthy subjects, as confirmed by a large amount of literature over the last two decades [37,57,58]. Similarly, both the effect of an acoustic startle on corticospinal excitability [43] and the capability of the brain to discriminate biological from non-biological motion were clarified by previous studies [58]. Second, the pendulum consisted of a graspable object (a wooden sphere of ~3 cm diameter). Although the pendulum was out of the range for reaching-grasping movements, visuomotor processing of the sphere affordance might have influenced corticospinal excitability during pendulum observation. To date, the observation of graspable objects modulated corticospinal excitability in healthy subjects [59,60]. Anecdotally, patient 10, an MCS patient, one of the two patients showing corticospinal facilitation during pendulum observation, often exhibited a gesture of attempting to catch the pendulum as she was looking at it. This suggests that, at least in this case, some visuomotor affordance might have been elicited by the mere observation of the sphere. Even though we cannot rule out the possibility that corticospinal excitability might have been affected by pendulum affordance, our results nevertheless support the hypothesis that at least some patients were able to discriminate between pendulum observation and human motion.
5. Conclusions
In this study, we have shown that a certain number of patients with DoC, irrespective of clinical severity, can process human motion like conscious subjects and discriminate it from non-biological motion. Conversely, acoustic startle stimuli produced abnormal responses in terms of corticospinal excitability modulation, probably due to a functional dissociation between cortical and subcortical networks. There was no correlation between corticospinal excitability and level of consciousness. Similarly, no relationship was detected with outcome measures commonly used in clinical practice. Whether modulation of corticospinal excitability may convey prognostic information in patients with DoC remains a fascinating but still unconfirmed possibility. Nevertheless, at least in the case of corticospinal facilitation during action observation, the positive correlation of this measure with the amplitude of the N20 component suggests that the present approach may provide additional prognostic tools in clinical settings. However, further work is needed to better comprehend the conditions in which corticospinal facilitation occurs and whether and how it may relate to individual clinical characteristics.
Author Contributions
Conceptualization, L.F. and S.R.; methodology, L.F. and S.R.; formal analysis, M.E.; investigation, L.A, S.C., M.E., F.N. and L.M.; data curation, M.E.; writing—original draft preparation, M.E.; writing—review and editing, L.F., P.Z., B.B., M.M., S.R., and E.S.; visualization, M.E.; supervision, L.F., P.Z., B.B., M.M. and S.R.
Funding
This research received no funding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Giacino, J.T.; Fins, J.J.; Laureys, S.; Schiff, N.D. Disorders of consciousness after acquired brain injury: the state of the science. Nat. Rev. Neurol. 2014, 10, 99–114. [Google Scholar] [CrossRef] [PubMed]
- Bodart, O.; Laureys, S.; Gosseries, O. Coma and Disorders of Consciousness: Scientific Advances and Practical Considerations for Clinicians. Semin. Neurol. 2013, 33, 083–090. [Google Scholar] [CrossRef] [PubMed]
- Laureys, S.; Celesia, G.G.; Cohadon, F.; Lavrijsen, J.; León-Carrión, J.; Sannita, W.G.; Sazbon, L.; Schmutzhard, E.; von Wild, K.R.; Zeman, A.; et al. Unresponsive wakefulness syndrome: a new name for the vegetative state or apallic syndrome. BMC Med. 2010, 8, 68. [Google Scholar] [CrossRef] [PubMed]
- Jennett, B.; Plum, F. Persistent vegetative state after brain damage: A Syndrome in Search of a Name. Lancet 1972, 299, 734–737. [Google Scholar] [CrossRef] [PubMed]
- Giacino, J.T.; Ashwal, S.; Childs, N.; Cranford, R.; Jennett, B.; Katz, D.I.; Kelly, J.P.; Rosenberg, J.H.; Whyte, J.; Zafonte, R.D.; et al. The minimally conscious state: Definition and diagnostic criteria. Neurology 2002, 58, 349–353. [Google Scholar] [CrossRef] [PubMed]
- Ragazzoni, A.; Cincotta, M.; Giovannelli, F.; Cruse, D.; Young, G.B.; Miniussi, C.; Rossi, S. Clinical neurophysiology of prolonged disorders of consciousness: From diagnostic stimulation to therapeutic neuromodulation. Clin. Neurophysiol. 2017, 128, 1629–1646. [Google Scholar] [CrossRef] [PubMed]
- Rossi Sebastiano, D.; Panzica, F.; Visani, E.; Rotondi, F.; Scaioli, V.; Leonardi, M.; Sattin, D.; D’Incerti, L.; Parati, E.; Ferini Strambi, L.; et al. Significance of multiple neurophysiological measures in patients with chronic disorders of consciousness. Clin. Neurophysiol. 2015, 126, 558–564. [Google Scholar] [CrossRef] [PubMed]
- Fleming SM, Frith CD, Goodale M (2023) The integrated information theory of consciousness as pseudoscience. PsyArXiv. Advance online publication. Retrieved October 30, 2023. 10.31234. [CrossRef]
- Seth, A.K.; Izhikevich, E.; Reeke, G.N.; Edelman, G.M. Theories and measures of consciousness: An extended framework. Proc. Natl. Acad. Sci. 2006, 103, 10799–10804. [Google Scholar] [CrossRef]
- Tononi, G.; Edelman, G.M. Consciousness and Complexity. Science (80-. ). 1998, 282, 1846–1851. [Google Scholar] [CrossRef]
- Rosanova, M.; Gosseries, O.; Casarotto, S.; Boly, M.; Casali, A.G.; Bruno, M.-A.; Mariotti, M.; Boveroux, P.; Tononi, G.; Laureys, S.; et al. Recovery of cortical effective connectivity and recovery of consciousness in vegetative patients. Brain 2012, 135, 1308–1320. [Google Scholar] [CrossRef]
- Ragazzoni, A.; Pirulli, C.; Veniero, D.; Feurra, M.; Cincotta, M.; Giovannelli, F.; Chiaramonti, R.; Lino, M.; Rossi, S.; Miniussi, C. Vegetative versus Minimally Conscious States: A Study Using TMS-EEG, Sensory and Event-Related Potentials. PLoS One 2013, 8, e57069. [Google Scholar] [CrossRef] [PubMed]
- Sarasso, S.; Rosanova, M.; Casali, A.G.; Casarotto, S.; Fecchio, M.; Boly, M.; Gosseries, O.; Tononi, G.; Laureys, S.; Massimini, M. Quantifying Cortical EEG Responses to TMS in (Un)consciousness. Clin. EEG Neurosci. 2014, 45, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Berlingeri, M.; Magnani, F.G.; Salvato, G.; Rosanova, M.; Bottini, G. Neuroimaging Studies on Disorders of Consciousness: A Meta-Analytic Evaluation. J. Clin. Med. 2019, 8. [Google Scholar] [CrossRef] [PubMed]
- Owen, A.M.; Menon, D.K.; Johnsrude, I.S.; Bor, D.; Scott, S.K.; Manly, T.; Williams, E.J.; Mummery, C.; Pickard, J.D. Detecting residual cognitive function in persistent vegetative state. Neurocase 2002, 8, 394–403. [Google Scholar] [CrossRef] [PubMed]
- Crone, J.S.; Ladurner, G.; Höller, Y.; Golaszewski, S.; Trinka, E.; Kronbichler, M. Deactivation of the Default Mode Network as a Marker of Impaired Consciousness: An fMRI Study. PLoS One 2011, 6, e26373. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Espejo, D.; Junque, C.; Cruse, D.; Bernabeu, M.; Roig-Rovira, T.; Fábregas, N.; Rivas, E.; Mercader, J.M. Combination of diffusion tensor and functional magnetic resonance imaging during recovery from the vegetative state. BMC Neurol. 2010, 10, 77. [Google Scholar] [CrossRef]
- Owen, A.; Coleman, M.; Menon, D.; Johnsrude, I.; Rodd, J.; Davis, M.; Taylor, K.; Pickard, J. Residual auditory function in persistent vegetative state: a combined pet and fmri study. Neuropsychol. Rehabil. 2005, 15, 290–306. [Google Scholar] [CrossRef]
- Di, H.B.; Yu, S.M.; Weng, X.C.; Laureys, S.; Yu, D.; Li, J.Q.; Qin, P.M.; Zhu, Y.H.; Zhang, S.Z.; Chen, Y.Z. Cerebral response to patient’s own name in the vegetative and minimally conscious states. Neurology 2007, 68, 895–9. [Google Scholar] [CrossRef]
- Qin, P.; Di, H.; Liu, Y.; Yu, S.; Gong, Q.; Duncan, N.; Weng, X.; Laureys, S.; Northoff, G. Anterior cingulate activity and the self in disorders of consciousness. Hum. Brain Mapp. 2010, 31, 1993–2002. [Google Scholar] [CrossRef]
- Staffen, W.; Kronbichler, M.; Aichhorn, M.; Mair, A.; Ladurner, G. Selective brain activity in response to one’s own name in the persistent vegetative state. J. Neurol. Neurosurg. Psychiatry 2006, 77, 1383–4. [Google Scholar] [CrossRef]
- Kotchoubey, B.; Merz, S.; Lang, S.; Markl, A.; Müller, F.; Yu, T.; Schwarzbauer, C. Global functional connectivity reveals highly significant differences between the vegetative and the minimally conscious state. J. Neurol. 2013, 260, 975–983. [Google Scholar] [CrossRef] [PubMed]
- Sharon, H.; Pasternak, Y.; Ben Simon, E.; Gruberger, M.; Giladi, N.; Krimchanski, B.Z.; Hassin, D.; Hendler, T. Emotional Processing of Personally Familiar Faces in the Vegetative State. PLoS One 2013, 8, e74711. [Google Scholar] [CrossRef] [PubMed]
- Owen, A.M.; Coleman, M.R.; Boly, M.; Davis, M.H.; Laureys, S.; Pickard, J.D. Detecting awareness in the vegetative state. Science 2006, 313, 1402. [Google Scholar] [CrossRef] [PubMed]
- Bekinschtein, T.A.; Manes, F.F.; Villarreal, M.; Owen, A.M.; Della-Maggiore, V. Functional imaging reveals movement preparatory activity in the vegetative state. Front. Hum. Neurosci. 2011, 5, 5. [Google Scholar] [CrossRef] [PubMed]
- Monti, M.M.; Vanhaudenhuyse, A.; Coleman, M.R.; Boly, M.; Pickard, J.D.; Tshibanda, L.; Owen, A.M.; Laureys, S. Willful Modulation of Brain Activity in Disorders of Consciousness. N. Engl. J. Med. 2010, 362, 579–589. [Google Scholar] [CrossRef] [PubMed]
- Wijdicks, E.F.M.; Hijdra, A.; Young, G.B.; Bassetti, C.L.; Wiebe, S. Quality Standards Subcommittee of the American Academy of Neurology Practice parameter: prediction of outcome in comatose survivors after cardiopulmonary resuscitation (an evidence-based review): report of the Quality Standards Subcommittee of the American Academy of Neurology. Neurology 2006, 67, 203–210. [Google Scholar] [PubMed]
- Cruse, D.; Norton, L.; Gofton, T.; Young, G.B.; Owen, A.M. Positive Prognostication from Median-Nerve Somatosensory Evoked Cortical Potentials. Neurocrit. Care 2014, 21, 238–244. [Google Scholar] [CrossRef] [PubMed]
- Guérit, J.-M.; Amantini, A.; Amodio, P.; Andersen, K.V.; Butler, S.; de Weerd, A.; Facco, E.; Fischer, C.; Hantson, P.; Jäntti, V.; et al. Consensus on the use of neurophysiological tests in the intensive care unit (ICU): Electroencephalogram (EEG), evoked potentials (EP), and electroneuromyography (ENMG). Neurophysiol. Clin. Neurophysiol. 2009, 39, 71–83. [Google Scholar] [CrossRef] [PubMed]
- Logi, F.; Fischer, C.; Murri, L.; Mauguière, F. The prognostic value of evoked responses from primary somatosensory and auditory cortex in comatose patients. Clin. Neurophysiol. 2003, 114, 1615–1627. [Google Scholar] [CrossRef]
- Young, G.B.; Doig, G.; Ragazzoni, A. Anoxic–Ischemic Encephalopathy: Clinical and Electrophysiological Associations With Outcome. Neurocrit. Care 2005, 2, 159–164. [Google Scholar] [CrossRef]
- Carter, B.G.; Butt, W. Are somatosensory evoked potentials the best predictor of outcome after severe brain injury? A systematic review. Intensive Care Med. 2005, 31, 765–775. [Google Scholar] [CrossRef] [PubMed]
- Amantini, A.; Grippo, A.; Fossi, S.; Cesaretti, C.; Piccioli, A.; Peris, A.; Ragazzoni, A.; Pinto, F. Prediction of “awakening” and outcome in prolonged acute coma from severe traumatic brain injury: evidence for validity of short latency SEPs. Clin. Neurophysiol. 2005, 116, 229–35. [Google Scholar] [CrossRef] [PubMed]
- Carter, B.G.; Butt, W. Review of the use of somatosensory evoked potentials in the prediction of outcome after severe brain injury. Crit. Care Med. 2001, 29, 178–186. [Google Scholar] [CrossRef]
- Vanhaudenhuyse, A.; Laureys, S.; Perrin, F. Cognitive Event-Related Potentials in Comatose and Post-Comatose States. Neurocrit. Care 2008, 8, 262–270. [Google Scholar] [CrossRef]
- Rothwell, J. C. Techniques and mechanisms of action of transcranial stimulation of the human motor cortex. J. Neurosci. Meth. 1997, 74, 113–122. [Google Scholar]
- Rizzolatti, G.; Sinigaglia, C. The mirror mechanism: a basic principle of brain function. Nat. Rev. Neurosci. 2016, 17, 757–765. [Google Scholar] [CrossRef] [PubMed]
- Fadiga, L.; Fogassi, L.; Pavesi, G.; Rizzolatti, G. Motor facilitation during action observation: a magnetic stimulation study. J. Neurophysiol. 1995, 73, 2608–11. [Google Scholar]
- Rizzolatti, G.; Craighero, L. The mirror neuron system. Annu. Rev. Neurosci. 2004, 27, 169–192. [Google Scholar] [PubMed]
- Gallese, V.; Fadiga, L.; Fogassi, L.; Rizzolatti, G. Action recognition in the premotor cortex. Brain 1996, 119, 593–609. [Google Scholar]
- Pistoia, F.; Sacco, S.; Carolei, A.; Sarà, M. Corticomotor facilitation in vegetative state: results of a pilot study. Arch. Phys. Med. Rehabil. 2013, 94, 1599–606. [Google Scholar]
- Furubayashi, T.; Ugawa, Y.; Terao, Y.; Hanajima, R.; Sakai, K.; Machii, K.; Mochizuki, H.; Shiio, Y.; Uesugi, H.; Enomoto, H.; et al. The human hand motor area is transiently suppressed by an unexpected auditory stimulus. Clin. Neurophysiol. 2000, 111, 178–83. [Google Scholar] [CrossRef] [PubMed]
- Davis, M. The Mammalian Startle Response. In Neural Mechanisms of Startle Behavior; Springer US: Boston, MA, 1984; pp. 287–351. [Google Scholar]
- Kelly, C.P.; Pothoulakis, C.; LaMont, J.T. Medical aspects of the persistent vegetative state (1). The Multi-Society Task Force on PVS. N. Engl. J. Med. 1994, 330, 1499–508. [Google Scholar]
- Rossi, S.; Hallett, M.; Rossini, P.M.; Pascual-Leone, A.; Avanzini, G.; Bestmann, S.; Berardelli, A.; Brewer, C.; Canli, T.; Cantello, R.; et al. Safety, ethical considerations, and application guidelines for the use of transcranial magnetic stimulation in clinical practice and research. Clin. Neurophysiol. 2009, 120, 2008–2039. [Google Scholar] [PubMed]
- Mancuso, M.; Abbruzzese, L.; Canova, S.; Landi, G.; Rossi, S.; Santarnecchi, E. Transcranial random noise stimulation does not improve behavioral and neurophysiological measures in patients with subacute Vegetative-Unresponsive Wakefulness State (VS-UWS). Front. Hum. Neurosci. 2017, 11. [Google Scholar] [CrossRef] [PubMed]
- Hagen C, Malkmus D, Durham P. Levels of cognitive functioning. Downey (CA): Rancho Los Amigos Hospital, 1972.
- JASPER, H. The ten twenty electrode system of the international federation. Electroencephalogr. Clin. Neurophysiol. 1958, 10, 371–375. [Google Scholar]
- Guide for the Uniform Data Set for Medical Rehabilitation (Adult FIM), version 4.0. Buffalo: State University of New York at Buffalo, 1993.
- Rossini, P.M.; Burke, D.; Chen, R.; Cohen, L.G.; Daskalakis, Z.; Di Iorio, R.; Di Lazzaro, V.; Ferreri, F.; Fitzgerald, P.B.B.; George, M.S.S.; et al. Non-invasive electrical and magnetic stimulation of the brain, spinal cord, roots and peripheral nerves: Basic principles and procedures for routine clinical and research application: An updated report from an I.F.C.N. Committee. Clin. Neurophysiol. 2015, 126, 1071–1107. [Google Scholar] [PubMed]
- Gangitano, M.; Mottaghy, F.M.; Pascual-Leone, A. Phase-specific modulation of cortical motor output during movement observation. Neuroreport 2001, 12, 1489–1492. [Google Scholar] [CrossRef] [PubMed]
- Shukla, D.; Devi, B.I.; Agrawal, A. Outcome measures for traumatic brain injury. Clinical neurology and neurosurgery 2011, 113, 435–441. [Google Scholar] [CrossRef] [PubMed]
- Rossato, E.; Verzini, E.; Scandola, M.; Ferrari, F.; Bonadiman, S. Role of LCF scale as an outcome prognostic index in patients with traumatic brain injury. Neurological Sciences 2021, 42, 2747–2752. [Google Scholar]
- Blair, R.C.; Karniski, W. An alternative method for significance testing of waveform difference potentials. Psychophysiology 1993, 30, 518–24. [Google Scholar]
- Leitner, D.S.; Powers, A.S.; Hoffman, H.S. The neural substrate of the startle response. Physiol. Behav. 1980, 25, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Naro, A.; Pignolo, L.; Lucca, L. F.; Calabrò, R. S. An action-observation/motor-imagery based approach to differentiate disorders of consciousness: what is beneath the tip of the iceberg? Rest. Neurol. Neurosci. 2021, 39, 181–197. [Google Scholar] [CrossRef] [PubMed]
- Keysers, C.; Fadiga, L. The mirror neuron system: New frontiers. Soc. Neurosci. 2008, 3, 193–198. [Google Scholar] [CrossRef] [PubMed]
- Dayan, E.; Sella, I.; Mukovskiy, A.; Douek, Y.; Giese, M.A.; Malach, R.; Flash, T. The Default Mode Network Differentiates Biological From Non-Biological Motion. Cereb. Cortex 2016, 26, 234–245. [Google Scholar] [CrossRef] [PubMed]
- Bartoli, E.; Maffongelli, L.; Jacono, M.; D’Ausilio, A. Representing tools as hand movements: Early and somatotopic visuomotor transformations. Neuropsychologia 2014, 61, 335–344. [Google Scholar] [CrossRef]
- Franca, M.; Turella, L.; Canto, R.; Brunelli, N.; Allione, L.; Andreasi, N.G.; Desantis, M.; Marzoli, D.; Fadiga, L. Corticospinal Facilitation during Observation of Graspable Objects: A Transcranial Magnetic Stimulation Study. PLoS One 2012, 7, e49025. [Google Scholar] [CrossRef]
Figure 1.
(a) An example MEP is plotted. AUC was calculated between the markers indicating MEP onset and offset; (b) N20 amplitude was calculated between the N20 and the subsequent P25 potential.
Figure 1.
(a) An example MEP is plotted. AUC was calculated between the markers indicating MEP onset and offset; (b) N20 amplitude was calculated between the N20 and the subsequent P25 potential.
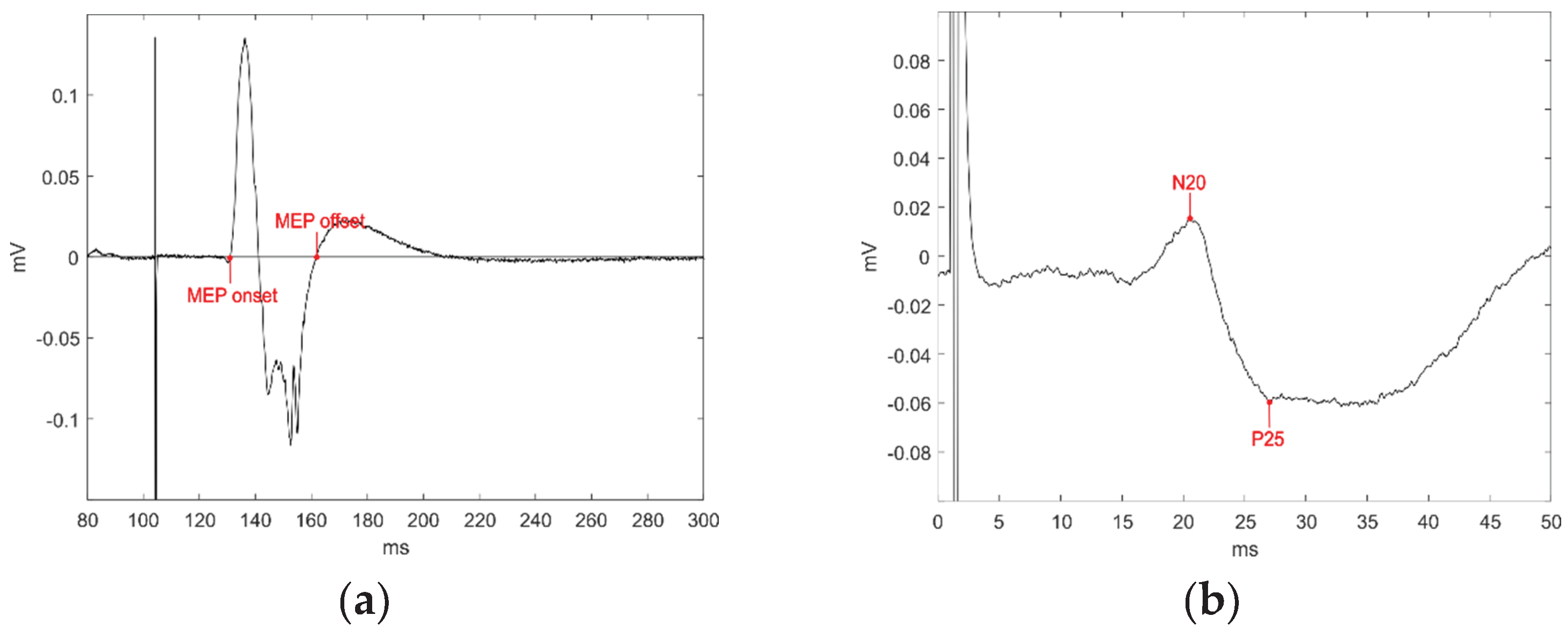

Table 1.
Demographic and clinical information.
| No. | Primitive Pathology | Age (Years) | Diagnosis | CRS-R | Time from Injury (Days) |
|---|---|---|---|---|---|
| 1 | Subarachnoid haemorrhage | 76 | VS/UWS | 1 | 26 |
| 2 | Ischemic stroke | 59 | VS/UWS | 5 | 33 |
| 3 | Intraparenchymal haemorrhage | 72 | VS/UWS | 5.6 | 75 |
| 4 | Subarachnoid haemorrhage | 62 | VS/UWS | 6.4 | 96 |
| 5 | Subarachnoid haemorrhage | 46 | VS/UWS | 6.4 | 60 |
| 6 | Ischemic stroke | 64 | VS/UWS | 6.8 | 80 |
| 7 | Subarachnoid haemorrhage | 81 | VS/UWS | 7.8 | 121 |
| 8 | Ischemic stroke | 74 | MCS | 8.4 | 51 |
| 9 | Subarachnoid haemorrhage | 79 | MCS | 8.4 | 130 |
| 10 | Intraparenchymal haemorrhage | 79 | MCS | 8.4 | 24 |
| 11 | Intraparenchymal haemorrhage | 81 | MCS | 9.2 | 61 |
Table 2.
Mean MEPs AUC (mV*ms) across patients and conditions. Patients are listed for their CRS-R to appreciate the relationship with coma severity. Individual data significantly different from the baseline are presented in bold. See the text for additional significant comparisons. The standard deviations are shown in brackets.
Table 2.
Mean MEPs AUC (mV*ms) across patients and conditions. Patients are listed for their CRS-R to appreciate the relationship with coma severity. Individual data significantly different from the baseline are presented in bold. See the text for additional significant comparisons. The standard deviations are shown in brackets.
| No. | Baseline | Action Observation | Acoustic Startle | Pendulum | Diagnosis | CRS-R |
|---|---|---|---|---|---|---|
| 1 | 13.58 (6.63) | 28.62 (18.52) | 25.47 (16.12) | VS/UWS | 1 | |
| 2 | 11.18 (2.30) | 15.09 (8.28) | 13.69 (5.04) | VS/UWS | 5 | |
| 3 | 20.07 (20.83) | 21.54 (18.94) | 20.37 (16.48) | 19.20 (21.67) | VS/UWS | 5.6 |
| 4 | 28.65 (3.29) | 31.32 (1.60) | 31.01 (2.10) | 31.32 (1.86) | VS/UWS | 6.4 |
| 5 | 91.82 (20.86) | 81.01 (18.40) | 95.42 (22.11) | 92.00 (17.28) | VS/UWS | 6.4 |
| 6 | 20.21 (11.23) | 31.45 (2.92) | 28.18 (3.14) | 25.84 (4.31) | VS/UWS | 6.8 |
| 7 | 43.93 (29.64) | 35.66 (23.95) | 41.81 (8.91) | VS/UWS | 7.8 | |
| 8 | 32.63 (12.36) | 47.94 (42.04) | 46.96 (47.29) | 39.80 (26.64) | MCS | 8.4 |
| 9 | 26.60 (18.64) | 20.15 (8.75) | 46.71 (16.90) | 25.23 (18.84) | MCS | 8.4 |
| 10 | 35.90 (24.73) | 38.02 (27.13) | 80.32 (44.35) | 84.18 (46.62) | MCS | 8.4 |
| 11 | 34.16 (24.72) | 59.53 (21.13) | 15.29 (8.67) | 27.36 (19.30) | MCS | 9.2 |
Table 3.
Average MEPs peak-to-peak amplitude (mV) across patients and conditions. Patients are listed for their CRS-R to appreciate the relationship with coma severity. Individual data significantly different from the baseline are presented in bold. See the text for additional significant comparisons. The standard deviations are shown in brackets.
Table 3.
Average MEPs peak-to-peak amplitude (mV) across patients and conditions. Patients are listed for their CRS-R to appreciate the relationship with coma severity. Individual data significantly different from the baseline are presented in bold. See the text for additional significant comparisons. The standard deviations are shown in brackets.
| No. | Baseline | Action Observation | Acoustic Startle | Pendulum | Diagnosis | CRS-R |
|---|---|---|---|---|---|---|
| 1 | 0.38 (0.19) | 0.82 (0.55) | 0.68 (0.44) | VS/UWS | 1 | |
| 2 | 0.50 (0.10) | 0.77 (0.38) | 0.68 (0.23) | VS/UWS | 5 | |
| 3 | 1.18 (0.86) | 1.20 (0.99) | 1.12 (0.80) | 1.15 (1.15) | VS/UWS | 5.6 |
| 4 | 1.03 (0.11) | 1.11 (0.07) | 1.10 (0.11) | 1.13 (0.07) | VS/UWS | 6.4 |
| 5 | 4.15 (0.84) | 3.67 (0.82) | 3.92 (1.02) | 4.51 (0.66) | VS/UWS | 6.4 |
| 6 | 0.72 (0.41) | 1.08 (0.16) | 1.02 (0.10) | 0.93 (0.16) | VS/UWS | 6.8 |
| 7 | 1.11 (0.76) | 0.98 (0.74) | 1.46 (0.63) | VS/UWS | 7.8 | |
| 8 | 1.08 (0.43) | 1.12 (0.40) | 1.56 (1.64) | 1.32 (0.95) | MCS | 8.4 |
| 9 | 0.85 (0.56) | 0.61 (0.26) | 1.37 (0.55) | 0.80 (0.55) | MCS | 8.4 |
| 10 | 1.42 (0.92) | 1.60 (0.91) | 2.83 (1.49) | 3.09 (1.67) | MCS | 8.4 |
| 11 | 1.44 (0.84) | 2.18 (0.81) | 0.80 (0.39) | 1.20 (0.81) | MCS | 9.2 |
Table 4.
Clinical data ad admission and discharge and lifespan data. Patients are listed for their CRS-R to appreciate the relationship with coma severity.
Table 4.
Clinical data ad admission and discharge and lifespan data. Patients are listed for their CRS-R to appreciate the relationship with coma severity.
| LCF | FIM | Lifespan (months) |
||||
|---|---|---|---|---|---|---|
| No. | CRS-R | Admission | Discharge | Admission | Discharge | |
| 1 | 1 | 2 | - | 18 | - | 1 |
| 2 | 5 | 2 | 3 | 18 | 18 | Alive |
| 3 | 5.6 | 2 | 2 | 18 | 18 | 4 |
| 4 | 6.4 | 2 | 4 | 18 | 23 | 41 |
| 5 | 6.4 | 2 | 2 | 18 | 18 | 8 |
| 6 | 6.8 | 2 | 2 | 18 | 18 | 6 |
| 7 | 7.8 | 1 | 2 | 18 | 18 | 35 |
| 8 | 8.4 | 2 | 2 | 18 | 18 | - |
| 9 | 8.4 | 2 | deceased | 18 | deceased | 4 |
| 10 | 8.4 | 2 | 2 | 18 | 18 | 6 |
| 11 | 9.2 | 3 | 3 | 21 | - | 15 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



