
Submitted:
06 March 2024
Posted:
07 March 2024
You are already at the latest version
Abstract
Marine microalgae are a rich reservoir of natural compounds, including bioactives. Nonetheless, these organisms remain fairly unexplored despite their potential biotechnological applications. Culture collections with diverse taxonomic groups and lifestyles are a good source to unlock this potential and discover new molecules for multiple applications such as the treatment of human pathologies or the production of aquaculture species. In the present work extracts from 33 strains (including 20 dinoflagellates, 4 diatoms and 9 strains from other 7 algal classes), cultivated under identical conditions were examined for antiviral, antibacterial, anti-inflammatory and anti-cancer activities. Among these, antiviral and anti-inflammatory activities were detected in a few strains while antibacterial tests showed positive results in most assays. In turn, most trials did not show any anti-cancer activity. Wide differences were observed between species within the same class, in particular dinoflagellates which were better represented in this study. These preliminary findings pave the way for in-depth characterization of extracts with highest signals in each test, identification of compounds responsible of the biological activities found, and a further screening of the CCVIEO culture collection.
Keywords:
Microalgae
; Bioactive compounds
; Zebrafish
; Inflammation
; Antiviral activity
; Antibacterial activity
; Anti-cancer activity
1. Introduction
Marine organisms are an important source of proteins and provide a broad range of health-promoting bioactive compounds with multiple applications in diverse biotechnological and pharmacological sectors such as human health, nutraceuticals, cosmetics, well-being, and even animal production [1,2]. Microorganisms included in the phytoplankton such as the cyanobacteria and microalgae are underexploited as source of bioactive compounds [3,4,5]. Those microorganisms are of great interest since they are highly taxonomically diverse, showing a complex evolutionary history (as compared, e.g., with land plants). Their ecophysiology diversity confers a high metabolic plasticity to respond and adapt to a changing environment by producing bioactive secondary metabolites (reviewed in Saide et al. [6]). Marine planktonic cyanobacteria and microalgae are able to synthetize a huge amount of potential bioactive compounds including pigments, lipids, glycolipids, alkaloids, terpenoids, ribosomal and non-ribosomal peptides, polyketides, phenolic acids, vitamins, flavonoids and macrolides, among others [4,6,7,8,9], but in many occasions the active principles are yet unknown (Lauritano et al. 2016; [10,11]. A great advantage of using microalgae lies on their relatively easy growing and building-up of high biomass in short generation time. The ability to produce bioactive compounds is species-specific and depends on several factors such as the environmental parameters (growth medium, temperature, light intensity, pH), length of the growth phase, and treatment of the biomass before the extraction [10,12,13,14,15].
Pharmacological and biological activities have been reported for almost all secondary metabolites produced by microalgae. Those properties included, among others, antibacterial, antiviral, antifungal and antialgal activities [16,17,18,19,20], antioxidant [21,22], anti-inflammatory and immunomodulatory activities [9,10,23,24,25,26], anti-proliferative and anticancer activity [12,27,28,29]. Moreover, compounds derived from microalgae are useful for the prevention and treatment of other human pathologies such as diabetes, hypertension, artherosclerosis and osteoporosis [6]. Microalgae derived products have also been applied to the aquaculture industry [30,31]. These organisms are included as food supplements because of their nutritional values and their immunomodulatory properties to mitigate stressful conditions in crustacean and fish production [32,33].
Bacterial and viral diseases are one of the major aquaculture challenges responsible, in some cases, of high mortalities and important economic losses. Aquaculture fish are affected by several groups of pathogenic viruses such as the birnavirus (IPNV), orthomyxovirus (ISAV), rhabdovirus (SVCV, IHNV and VHSV), adenovirus and herpesvirus (reviewed in Kim and Leong [34]). In particular, the spring viremia of carp virus (SVCV) is responsible of elevated mortalities worldwide in carp culture [35] and requires notification to the Office of International Epizootic (OIE) when detected [36]. Although it causes substantial economic losses to the aquaculture industry, there are no effective therapies for its prevention and treatment [37,38]. The major bacterial diseases in aquaculture animals include Aeromonas sp. and Pseudomonas sp. in many freshwater species, or Vibrio sp. in marine ones [39,40,41]. In particular, Aeromonas hydrophyla infection affects different freshwater fish species such as common carps, goldfish, eel, catfish and tilapia [42] and is also described as an emergent pathogen for the cultured freshwater shrimp Litopenaeus vannamei [43]. Other opportunistic bacteria such as Micrococcus luteus can also induce lethal infections when the animal is immunologically compromised [44]. In this context, bioactive compounds from microalgae emerge as a potential source of antiviral components [45,46,47] and are also a plausible alternative to antibiotics to treat bacterial infections [10,48,49,50].
Microalgae also have the ability to induce immunomodulation in fish and confer protection against bacterial infections [33,51,52,53]. The identification and isolation of compounds with anti-inflammatory activity from microalgae are of great interest for human health but also for aquaculture production [54,55,56]. The control of the inflammatory process is relevant since it is involved in many physiological processes such as inflammatory disorders, immune-related diseases and infections [57,58,59]. Microalgae compounds with anti-inflammatory properties include carotenoids, polyunsaturated fatty acids (PUFA) such as eicosapentaenoic and docosahexaenoic acids (EPA and DHA, respectively) and certain sulphated polysaccharides although other bioactive chemicals are still unknown [10,60].
Available studies on exploring bioactive compounds in microalgae usually employ diatoms, green algae and dinoflagellates among other groups, but often focus on a limited number of species or bioactivity tests [61]. The screening of a wide range of taxonomic groups and assays is rarely found in the literature [10,13,62]. In that sense, culture collections with different algal classes, life styles and habitats provide biological material of enormous interest to identify new sources of bioactive molecules. These resources enable to undergo different assays under the same conditions, a first step towards finding bioactive fractions and molecules with potential applications in diverse fields, such as biomedicine or aquaculture.
The present study followed this approach. Taking advantage of the microalgal strains maintained in the CCVIEO culture collection (IEO-CSIC, Spain), a number of phytoplankton groups (9 taxonomic classes) were selected, including toxic dinoflagellate species, and 33 strains were screened for their bioactivity. These tests comprised antiviral, antibacterial, anti-inflammatory and anticancer activities. To our knowledge, many of the organisms included in the present study have not been tested in previous similar works. The obtained results provided some promising findings, concerning in particular antibacterial activity that would deserve to be explored further in-depth for their potential applications in human and animal health.
2. Results
2.1. Antiviral Activity of the Extracts
A total of 17 out of the 33 extracts analyzed showed significant effects against SVCV replication. A significant reduction of the viral titer was obtained at 6 days post infection although the magnitude of decrease was always lower than 1 log (Figure 1). The increment of the extract concentration from 10 to 25 μg mL-1 did not significantly increase the antiviral activity. Extracts that showed a moderate antiviral activity (PI between 90 and 50%) were obtained from six species of dinoflagellates (genera Alexandrium (E33), Dinophysis (E5, E29), Gambierdiscus (E30, E31), and Prorocentrum (E24) and one diatom of the genus Nitzschia (E14). Also, species from other taxonomic classes such as Chlorophyceae (Tetraselmis, E20), Cryptophyceae (Guillardia E21, and Teleaulax E02) and Euglenophyceae (Eutreptiella, E26) showed moderate antiviral activity (Figure 1). Finally, extracts from the diatom genus Chaetoceros (E13), the dinoflagellates Alexandrium (E23), Gymnodinium (E12), Karlodinium (E17) and Ostreopsis (E27), and the cryptophyte Falcomonas (E22) showed a strong antiviral activity and significantly reduced the viral titer up to 90% (PI > 90%) (Figure 1).
2.2. Antibacterial Activity of the Extracts
The antibacterial activity of the extracts was assayed against both Gram (+) and (-) bacteria using 10 μg mL-1 of the extract. The treatment of bacteria with 1% DMSO did not affect the bacterial growth. The evaluation of the OD600 nm during 24 h allowed determining the times where the extracts significantly reduced the bacterial growth. The results obtained using the extract 9 isolated from the dinoflagellate Prorocentrum lima was selected as representative result (Figure 2A). The growth of the Gram (-) A. hydrophila showed logarithmic kinetics and almost reached the highest level after 16 h of incubation. In contrast, an exponential growth was observed in the Gram (+) M. luteus (Figure 2A). Only 10 out of 33 extracts significantly reduced the growth of A. hydrophila at specific time points and the percentage of reduction was always lower than 13%. The highest antibacterial activity was registered in extracts isolated from the dinoflagellates A. minutum (E01) and P. lima (E09) at 7 and 8 h of incubation, respectively (Figure 2B).
The antibacterial activity against the Gram (+) M. luteus was generalized and maintained during all the bacterial incubation (Figure 2C). A total of 31 extracts induced significant reduction of the bacterial growth with an average percentage of reduction close to 30% after 24h of incubation. The highest percentage of bacterial reduction was registered at 16h of incubation with the extract isolated from the cryptophyte Teleaulax amphioxeia (E02; Figure 2C).
2.3. Anti-Inflammatory Activity of the Extracts
The anti-inflammatory properties of the extracts (at 25 μg mL-1) was assayed in vivo using the transgenic zebrafish larvae Tg(lyz:DsRed2). This animal shows the lysozyme-expressing cells (neutrophils) marked in red allowing their detection in live animals (Figure 3A). The inflammation was induced by a transversal sectioning of the caudal fin and the number of neutrophils was measured after 2 h, 24 h, and 48 h (Figure 3B). The number of neutrophils that migrated to this site increased quickly at 2 h reaching a mean value of 7 (±3) cells (Figure 3C). The number of cells significantly decreases in the following sampling points at 24 and 48 h post-injury (Figure 3C).
Eighteen out of 33 extracts induced significant changes in the number of neutrophils in any of the sampling points (Figure 3D). At 2 h post injury only 6 extracts significantly reduced the number of neutrophils. The extract isolated from the raphidophyte Heterosigma akashiwo (E18) induced the highest reduction in the number of migrating cells (up to 70%). Moreover, the extracts isolated from P. lima (E09), Ostreopsis siamensis (E06) and Dinophysis caudata (E29) reduced the number of neutrophils around 60% (Figure 3D). At 24h, only 8 extracts modified the number of cells. The extracts isolated from O. siamensis (E06), P. lima (E09) and H. akashiwo (E18) showed a significant reduction in the number of cells that was maintained from 2h to 24h post injury. At this last point (24h) the extracts reduced the number of cells around 80% and 60% respectively (Figure 3D). Interestingly the larvae treated with the extracts 27 (O. fattorussoi), 28 (O. ovata), 29 (D. caudata), and 32 (Gambierdiscus australes) showed a significant increment in the number of neutrophils at 24 and 48h. At the end of the experiment (48h), larvae treated with the extract isolated from A. minutum (E01), T. amphioxeia (E02), O. siamensis (E06), Lingulodinium polyedrum (E07), Heterocapsa minima (E08), H. akashiwo (E18), Pyramimonas sp. (E19), and Kryptoperidinium foliaceum (E25) showed a significant reduction in the number of cells (Figure 3D).
2.4. Cytotoxic Activity of the Extracts in the Cancer Lines
The cytotoxic activity was assayed in vitro using the human cell lines HCT 116, HepG2 and MG-63. None of the extracts showed anti-cancer activity. No significant reduction in the viability of the cells was observed after a 48 h exposure to the extracts at 5, 25 and 50 μg mL-1 (Figure 4).
3. Discussion
The present study carried out four different bioactivity screenings on microalgal extracts from 33 species and 9 algal classes, chiefly dinoflagellates and diatoms, since they are by far the most abundant in the CCVIEO culture collection. Toxic strains from both taxonomic groups were also included since they represent an interesting source of bioactive compounds [10,13,62]. At least one third of the extracts showed significant effects in some bioactivity test. It is important to remind that the production of primary and secondary metabolites and their potential biological activities is species specific and depends on several factors such as the environmental parameters, as stated in the introduction. Moreover, in some cases the production and release of active metabolites occurs when microalgae are grown in mixed crops or under specific mixotrophic conditions [63,64]. In our case, different culture conditions were not examined and strains were collected at the end of the exponential phase when dense cultures were obtained. However, we cannot provide any clue about the limiting conditions, if any, in our cultures. The positive results showed by several strains in each of the four bioactivity tests deserve further exploration of the responsible fractions and compounds involved in these effects. This work could consider different culture setups, including culture media with certain nutrient limitations, to check if major differences in bioactivity are observed under stressed conditions.
3.1. Antiviral Activity of the Extracts
The antiviral effect of the extracts is related with the presence of several bioactive molecules such as phenolic and flavonoid components and sulphated polysaccharides [47,55,65,66]. It is suggested that the fatty-acid molecules penetrate the host cell and break down the lipid coat in enveloped viruses, inducing the inactivation of the viruses at their point of entry [67]. The antiviral activity of algae extracts has also been related with the inhibition of the viral RNA polymerase [68]. However, additional studies are required to analyze the mechanisms of action of the algal extracts, focusing on the specific compounds of interest.
In the present study a total of 17 extracts showed moderate to strong activity against fish SVC virus. This information is in agreement with a similar antiviral activity against fish viruses (IPNV and VHSV) registered in extracts isolated from other marine microalgae species [69]. Our results suggest that the microalgae extracts could persist on the cell surfaces interfering on the viral replication as it was previously proposed by several authors [47,69].
Antiviral activity has been observed in several species of dinoflagellates. The different composition of the extracts between three species of the same genera could explain why the extracts from Alexandrium mediterraneum (E23), A. tamarense (E33) and A. minutum (E01) showed strong, moderate and weak antiviral activity, respectively. The potential antiviral effects of bioactive compounds produced by Alexandrium on marine viruses has already been observed by a reduction of the prevalence of infection with the herpesvirus OSHV-1 µVar in Crassostrea gigas when the A. pacificum was present [70]. This differential antiviral activity between close species was also observed in Ostreopsis fattorussoi (E27) (strong activity) and O. ovata (E28) and O. siamensis (E06) (no antiviral activity). The differences in the phenolic composition of the extracts or their interaction with other compounds may affect their antiviral activity as it was previously suggested [71]. A strong anti SVCV activity was also observed in the extract obtained from Gymnodinium impudicum (E12). This dinoflagellate has previously been reported to display antiviral activity against several viruses such as the non-enveloped encephalomyocarditis (EMCV) viruses that causes death in animal production [17] and against the influenza A virus [72].
Antiviral activity has already been observed in diatoms. Previous studies reported that most species of Chaetoceros are rich in fatty acids, such as oleic acid, linoleic acid, and alpha-linolenic acid [73]. Although we did not determine the lipid composition of C. dichatoensis (E13) their most probably enriched composition in fatty acids could contribute to its strong anti SVCV activity.
Other extracts with antiviral activity belonged to the Cryptophyceae and Chlorophyceae classes. For example, Falcomonas sp. (E22) is less known and has been recently explored for bioactive compounds [74]. In contrast, the moderate anti SVCV activity obtained in T. convolutae (E20) could be expected since activity against another fish rhabdovirus (VHSV) has already been described in T. suecica [75].
3.2. Antibacterial Activity
The use of microalgae as sources of natural antibiotics has been extensively explored as an alternative to conventional ones to limit microbial infections in aquaculture. Marine and freshwater microalgae included in different taxonomic groups showed a potent bactericidal activity against both Gram (+) and Gram (-) bacteria [10,48,50]. This activity has been associated with the presence of several metabolites, including sulfated polysaccharides, fatty acids, alkaloids, and phenolic compounds [69,76]. For example, extracts containing high amount of polyunsaturated fatty acids (PUFAs) or saturated fatty acids with 14, 16, or 18 carbons (SFAs) are reported to be more effective against Gram (+) and Gram (-) bacteria, respectively [77]. However, other studies using medicinal plant extracts enriched in SFAs described an antibacterial activity against both bacterial types [78]. In this study, a limited effect against the Gram (-) A. hydrophyla and a generalized antibacterial activity against the Gram (+) M. luteus were observed. It is necessary to analyze the composition of the extracts to understand the observed results, although there is not a sole responsible compound for the biological activity of the extracts.
The antimicrobial activity of several groups of marine diatoms has been extensively reported [10,18,79,80]. In the present work, the extracts from all the selected diatoms (C. dichatoensis (E13), Nitzschia sp. (E14), Pseudo-nitzschia australis (E15) and Thalassiosira delicatula (E04)) presented activity only against the Gram (+) M. luteus. This lack of bactericidal effect against Gram (-) bacteria was also described in Chaetoceros gracilis (Jusidin et al. 2022). In contrast, other species such as C. muelleri, C. affinis, Nitzschia sp., P. pseudodelicatissima, T. rotula and T. weissflogii showed activity against both bacterial groups [10,79,80,81,82].
Antimicrobial activities of bioactive metabolites, including shellfish toxins of dinoflagellate origins have also been reported [10,80,83,84]. Only the extracts obtained from the dinoflagellates A. minutum (E1) and P. lima (E9) showed bactericidal effects against both bacteria (M. luteus and A. hydrophyla). This result is in line with a strong antibacterial activity of extracts reported for other dinoflagellates (P. lima, Dinophysis fortii and Gambierdiscus sp.) towards several bacteria and fungi (Aspergillus niger, Penicillium funiculosum, Candida rugosa, Escherichia coli, Bacillus megaterium and Staphylococcus aureus) [84,85]. The extracts obtained from H. minima (E08) and Karlodinium veneficum (E17) also showed a higher activity against the Gram (+) M. luteus than the Gram (-) A. hydrophyla, as it was previously found in H. circularisquama and K. micrum using other bacterial species[86,87]. Only bactericidal activity against M. luteus was observed in the extract isolated from L. polyedrum (E07), although an extensive activity against both Gram (+) and Gram (-) bacteria has been reported [88].
In this study, antibacterial activity has also been observed in other taxonomic groups such as Chlorophyceae (T. convolutae, E20) and Prymnesiophyceae (Emiliania huxleyi, E16) in line with previous publications [89,90]. Interestingly, our result suggests the interest of strains from other classes such as Dictyochophyceae, Euglenophyceae, Prasinophyceae and Raphidophyceae as potential reservoirs for antibacterial compounds, as it is suggested for the class Cryptophyceae [74].
3.3. Anti-Inflammatory Activity
The anti-inflammatory activity of microalgae extracts has been classically measured in vitro using cell cultures [10,62]. Nevertheless, in the last decade, zebrafish (Danio rerio) has emerged as an in vivo model for the screening of anti-inflammatory natural products [91,92]. This in vivo model has proved to be suitable for the investigation of the kinetics of inflammation [93,94,95]. In the present study a mechanical injury in the fin was conducted in zebrafish larvae to attract leukocytes to a damaged area. To our knowledge, this is the first work using this procedure on zebrafish model to evaluate in vivo the anti-inflammatory activity of several marine microalgae extracts.
Several species of diatoms are known to produce molecules with anti-inflammatory properties [10,11,13,60,81]. In this study only the extract isolated from T. delicatula (E04) decreased the number of neutrophils at 24h. Although previous information about this species was not available, Asha shalini et al. [81] reported a potent anti-inflammatory activity of a close specie (T. weissflogii) when assayed in vitro.
The anti-inflammatory activity of dinoflagellates was also described on the literature [10,62], but this activity was previously reported in only two species out of 20 tested in this study (A. minutum E01 and Ostreopsis ovata E28). Extracts from A. tamarense (E33); D. caudata (E29); O. fattorussoi (E27); O. siamensis (E06) and P. lima (E09) showed an anti-inflammatory activity as early as 2h post-injury. Those last two extracts (extracts 6 and 9) also maintained the anti-inflammatory state at 24h post injury. In this context, Lauritano et al. [10] and Asha shalini et al. [81] reported anti-inflammatory activity in closely related dinoflagellate species such as Coolia malayensis, Heterocapsa psammophila, Prorocentrum rhathymum and P. gracile. In this study, the anti-inflammatory activity was also observed in extracts from other groups such as the raphidophyte H. akashiwo (E18), already described as a beneficial anti-inflammatory agent [92], and the class Cryptophyceae which has recently been targeted in the search for bioactive compounds [74].
3.4. Anti-Cancer Activity
Marine microalgae are a reservoir for the discovery of new anti-cancer drugs [12,96,97,98,99]. A preliminary evaluation of the anti-cancer activity of natural extracts is frequently done by the colorimetric based MTT/MTS in vitro cell proliferation assay since this is a reliable and economic method to evaluate the whole cell cytotoxicity. However, additional and specific anti-cancer assays are conducted on positive extracts [100,101]. This methodology has been widely used in the analysis of extracts obtained from several microalgae groups [10,13,62,96,102,103,104,105].
In this work, anti-cancer activity was assessed in vitro on 11 extracts isolated from dinoflagellates and diatoms using different cell lines of human cancers, including colorectal cancer (HCT116), hepatocellular carcinoma (HepG2) and osteosarcoma (MG63). Negative results in all these assays evidenced the lack of any anti-cancer activity, at least under our culture settings and concentrations. This outcome is strictly different from previous publications where anti-cancer activity has been reported in diatoms and dinoflagellates in general [12,97] and particularly in some species closely related to those in our study, such as Chaetoceros calcitrans [104], C. furcellatus, and C. socialis (Ingebrigtsen et al. 2016), Nitzschia palea [106], Pseudo-nitzschia delicatissima, Thalassiosira rotula [97,107], Coolia malayensis, Heterocapsa psammophila, Prorocentrum rhathymum, and P. gracile [10,62]. In addition to the composition of the extracts on bioactive compounds, it is reported that the anti-cancer activity is dose-dependent and specific for a particular cancer cell line [10]. All those factors could partially explain the discrepancy between the negative results in this work and those from the literature.
4. Materials and Methods
4.1. The CCVIEO Collection of Harmful Marine Microalgae
The Spanish Institute of Oceanography (IEO-CSIC) maintains a collection of marine microalgae (CCVIEO culture collection) constituted by about 250 strains and more than 80 species of microalgae isolated from the Spanish coasts but also from other parts of the world. The collection includes several species associated with harmful algal blooms (Fraga et al. 2010; https://vgohab.com/en/coleccion-de-cultivos/).
A total of 33 species from 9 different classes were selected in the present study, including strains isolated from the Atlantic coasts (eg. Chaetoceros dichatoensis and Tetraselmis convolutae), some of them responsible for harmful algal blooms (e.g. Alexandrium minutum and Dinophysis acuminata). The distribution of strains in each class was as follows: Bacillariophyceae (4), Chlorophyceae (1), Cryptophyceae (3), Dictyochophyceae (1), Dinophyceae (20), Euglenophyceae (1), Prasinophyceae (1), Prymnesiophyceae (1), and Raphidophyceae (1). The information about species and the strain codes used in the manuscript are specified in Table 1.
4.2. Culture and Preparation of Microalgae Extracts
The selected species were grown under specific culture parameters. The cultures were scaled up to a final volume of 1,600 mL. The culture parameters were optimized (temperature, light and composition of culture medium) for each species (Table 1). Cultures were grown in three culture chambers kept at 16 °C, 19 °C and 25 °C, under different light intensities depending on the requirements of each strain. A photon irradiance between 80-150 μE m2s-1 of PAR (LED illumination), measured with a QSL-100 irradiometer (Biospherical Instruments Inc., San Diego, CA, USA) and at a 12:12 L:D photoperiod was used (Table 1). The cultures were centrifuged (17,000 g for 10 min) to obtain the entire microalgae biomass. The protocol for organic extraction in freeze dried biomass from the studied strains followed that used for cyanobacteria by Edwards et al. [108].
Some modifications were introduced due to the lower biomass collected in the case of dinoflagellates. The bioactive extracts were obtained from the pellets following a protocol based on H2O-methanol and CH2Cl2-methanol extraction. Briefly, cultures were centrifuged (3,000 g, 20 min, 4 °C) and the resulting pellets frozen at -20 °C. Samples were lyophilized during 48 h, and final weights were recorded in each case. Afterwards, 20 mL of methanol were added to the extracts on 50 mL tubes, sonicated on an ultrasonic bath (3 min) and centrifuged again (7,500 g, 10 min, 4 °C), these steps being repeated three times. Methanol was fully evaporated on a rotary evaporator (Büchi R-200; Flawil, CH). Then, extracts were filtered to eliminate salts using reverse phase columns (Phenomenex, 500 mg 6 mL-1, Strata; Torrance, CA). Columns were activated adding 6 mL of methanol 5%. The non-polar fraction was collected as follows: 6 mL methanol 100% were added twice and a final rinsing step with 6 mL CH2CL2 100%, to render a final volume of 18 mL. Then, non-polar extracts were concentrated on a Speedvac (35 °C, 3 h). Dried extracts were weighted again, immediately frozen (-20 °C) and on the day after solubilized in DMSO to get a stock solution of 1 mg mL-1, kept at -80 °C until use.
4.3. Antiviral and Antibacterial Activity of the Extracts
The antiviral activity of the extracts against the spring viraemia of carp virus (SVCV) was assayed using the ZF4 cell line. The ZF4 cells (ATCC CRL-2050) were cultured at 28 ºC in DMEM/F-12, HEPES (Invitrogen, GIBCO) supplemented with 10% fetal bovine serum (FBS Invitrogen, GIBCO), penicillin (100 IU mL-1) (Invitrogen, GIBCO), and streptomycin (100g mL-1) (Invitrogen, GIBCO), and buffered with 7.5% sodium bicarbonate (Invitrogen, GIBCO). The SVCV isolate 56/70 was previously propagated on ZF4 cells and titrated in 96-well plates. The effect of the extracts on the viral replication was assayed by mixing the SVC virus with a non-toxic final concentration of 10 and 25 μg mL-1 of each extract (1% and 2.5% final concentration of DMSO, respectively). The plates were incubated at 28 °C for 6 days and examined for cytopathic effects. The virus dilution that causes an infection of 50% of the cell line (TCID50) was determined using the Reed-Müench [109] method. Each extract was titrated 4 times. The percentage of inhibition (PI) was calculated to present the antiviral activity of the extracts using the values of TCID50/mL, as follows: PI = [1 − (T/C)] × 100, with T being the viral titer of treated cells and C the viral titer of the positive control. The antiviral activity was scored as strong (PI>90%), moderate (PI between 50 and 90%) and weak (PI<50%) [69].
The antibacterial activity of the different extracts was assayed against both Gram (-) and Gram (+) bacteria. Briefly, The Gram (-) Aeromonas hydrophyla AH-1 strain (Yu et al., 2004) and the Gram (+) Micrococcus luteus (aka lysodeikticus) were grown on Tryptic Soy Agar (TSA, Scharlab) plates at 25 °C. Two bacterial suspensions were prepared in Tryptic Soy Broth (TSB, Scharlab) (at a final concentration of 5x106 and 106 CFUs/mL for A. hydrophyla and M. luteus, respectively) and mixed with the different extracts at a final concentration of 10 μg mL-1 (1% final concentration of DMSO). The bacteria were also treated with 1% DMSO and used as growth control. The bacterial growth was analyzed in a 96 well plate by measuring the OD at 600 nm during 24h in the GloMax reader (Promega). The percentage of bacterial growth reduction was calculated by using the values obtained in the control wells as 100% of bacterial growth. The experiment was conducted 4 times.
For both assays, the normality of the data was evaluated by a Kolmogorov-Smirnov test and a T-test was used to determinate significant differences at p>0.05 by using the GraphPad Prism V7 software.
4.4. Anti-Inflammatory Activity of the Extracts
The anti-inflammatory activity of the extracts was assayed in vivo using the transgenic zebrafish larvae Tg(lyz:DsRed2) showing red fluorescent neutrophils [110]. Eggs were obtained by natural spawning and reared at 28 °C. Transection of the caudal fin was performed in three days post-fertilization (3 dpf) larvae using a sapphire single edge lancet (WPI instrument) at the boundary of the notochord without injury the notochord and the vascular tissue. Animals were previously anesthetized by immersion in water containing 70 mg mL-1 tricaine (ethyl 3-aminobenzoate, Sigma-Aldrich). Animals were distributed in a 96 well plate (one animal per well) and immersed in water containing the different extracts at a final concentration of 25 μg mL-1. Control animals were treated with water containing the same concentration of DMSO (2.5%). Ten animals were used for each extract. Images of the injured animals were taken at 2h, 24h, and 48h using a fluorescent microscope DMi8 (LEICA). The sampling points were selected to analyze the initial cell migration and accumulation (2h), the resolution of the inflammatory process (24h) and the late rearrange of neutrophils (84h) according to the bibliography [93,94,95].
The number of neutrophils in the tail was counted in a 100 μm section anterior to the injury. Multiple focal planes were manually acquired when several neutrophils were overlapped to obtain images at single cell resolution. The z-stacks ensure an accurate determination of the number of neutrophils in those thick areas of the tail. The percentage of neutrophils was calculated by considering the number of cells observed in the control animals at 2h post injury as the 100% of neutrophil migration. All results were expressed as the mean and SD. Significant differences between the data were determined by performing a non-parametric Kruskal-Wallis test with a Dunns post-test using the GraphPad Prism V7 software. A p-value of less than 0.05 was considered statistically significant.
4.5. Cytotoxic Activity of the Extracts in Cancer Cell Lines
The human colon carcinoma cell line HCT 116 was obtained from Sigma-Aldrich (St. Louis, Missouri, USA). The human hepatocellular carcinoma and osteosarcoma cell lines, HepG2 and MG-63, respectively, were obtained from the American Culture Collection (ATCC) (Manassas, Virginia, EUA). The cell lines were maintained in an incubator with an atmosphere of 5% CO2 at 37 °C. HCT 116 was cultured in McCoy′s 5A medium (Sigma-Aldrich, Missouri, EUA), while HepG2 and MG-63 were cultured in Dulbecco’s modified Eagle medium (DMEM) (Biowest SAS, Nuaillé, France). Both media were supplemented with 10% fetal bovine serum (FBS) (Biochrom, Berlin, Germany), 1% of penicillin/streptomycin (Biochrom, Berlin, Germany), and 0.1% of amphotericin (GE Healthcare, Little Chafont, Buckinghamshire, UK).
For the assays, the selected extracts were resuspended in dimethyl sulfoxide (DMSO) and the cell lines were seeded at 3.3x104 cells mL-1 in 96 wells plates and incubated for 24 h. After, the cells were exposed to 0.5% of DMSO as a solvent control (maximum solvent concentration used), 1 μM of staurosporine (positive control) and to the concentrations 5, 25 and 50 μg mL-1 of the extracts, for 48 h.
Cell viability was evaluated by a colorimetric assay with 3-(4,5-Dimethylthiazol-2-yl)-2,5-Diphenyltetrazolium Bromide (MTT reagent). MTT was added to each well, to a final concentration of 200 μg mL-1, and incubated for 4 h. The formed formazan crystals were dissolved in 100 μL of DMSO and the absorbance was read in a multi-detection microplate reader (Synergy HT, Biotek, Bart Frederick Shahr, Germany) at 570 nm. Each cell line was used in three independent assays. For each assay, the values of each treatment were used to calculate the media and then normalized to the solvent control. Then the data was analyzed using the following calculus:
A T-test was used to determinate significant differences at p>0.05 by using the GraphPad Prism V7 software.
5. Conclusions
The results in this study show that extracts isolated from different groups of microalgae have interesting bioactivities and therefore additional efforts should be done to test their potential use in the production of aquaculture species. This is a first set of results that would lead in the future to a larger screening of strains in the CCVIEO culture collection. Promising results have been registered not only in dinoflagellates and diatoms but also in species belonging to some other groups like cryptophytes and raphidophytes.
Future work should continue this approach towards a more exhaustive exploitation of this culture collection, selecting strains with multiple bioactivities and manipulating the culture conditions to obtain novel bioactive metabolites in the seek of biotechnological and pharmacological applications.
Author Contributions
Conceptualization, J.H.U., A.R., A.F., B.N. and F.R.; methodology, J.H.U., F.R., A.R., P.C.; formal analysis, J.H.U., A.R.; writing—original draft preparation, J.H.U., A.R., F.R.; writing—review and editing, J.H.U., A.R., F.R., B.N., P.C.; funding acquisition, A.F., B.N., F.R., V.V. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by BLUEBIOLAB (0474 BLUEBIOLAB 1E, Interreg V (Spain-Portugal, POCTEP) 2014-2020), Spanish Ministry of Science and Innovation (project PID2020-119532RB-I00), CCVIEO projects (IEO-CSIC) and CIIMAR FCT projects UIDB/04423/2020 and UIDP/04423/2020. JHU was partially supported by a Juan de la Cierva’s postdoctoral research grant (IJC2020-043701-I; Ministry of Science and Innovation, Spain).
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Acknowledgments
The authors would like to thank Patricia Loures for technical assistance with CCVIEO cultures and Dr. María García-Portela for help with the extraction protocol.
Conflicts of Interest
“The authors declare no conflict of interest.”.
References
- Ghosh, S.; Sarkar, T.; Pati, S.; Kari, Z.A.; Edinur, H.A.; Chakraborty, R. Novel Bioactive Compounds From Marine Sources as a Tool for Functional Food Development. Front Mar Sci 2022, 9. [Google Scholar] [CrossRef]
- Øverland, M.; Mydland, L.T.; Skrede, A. Marine Macroalgae as Sources of Protein and Bioactive Compounds in Feed for Monogastric Animals. J Sci Food Agric 2019, 99, 13–24. [Google Scholar] [CrossRef]
- Kini, S.; Divyashree, M.; Mani, M.K.; Mamatha, B.S. Algae and Cyanobacteria as a Source of Novel Bioactive Compounds for Biomedical Applications. In Advances in Cyanobacterial Biology; Elsevier, 2020; pp. 173–194. [Google Scholar]
- Saha, S.; Shukla, S.K.; Singh, H.R.; Singh, B.; Jha, S.K. Bioactive Compounds from Microalgae. In An Integration of Phycoremediation Processes in Wastewater Treatment; Elsevier, 2022; pp. 337–358. [Google Scholar]
- Żak, A.; Kosakowska, A. Cyanobacterial and Microalgal Bioactive Compounds – the Role of Secondary Metabolites in Allelopathic Interactions. Oceanol Hydrobiol Stud 2016, 45, 131–143. [Google Scholar] [CrossRef]
- Saide, A.; Martínez, K.A.; Ianora, A.; Lauritano, C. Unlocking the Health Potential of Microalgae as Sustainable Sources of Bioactive Compounds. Int J Mol Sci 2021, 22, 4383. [Google Scholar] [CrossRef] [PubMed]
- Fu, W.; Nelson, D.R.; Yi, Z.; Xu, M.; Khraiwesh, B.; Jijakli, K.; Chaiboonchoe, A.; Alzahmi, A.; Al-Khairy, D.; Brynjolfsson, S.; et al. Bioactive Compounds From Microalgae: Current Development and Prospects. In; 2017; pp. 199–225.
- Heydarizadeh, P.; Poirier, I.; Loizeau, D.; Ulmann, L.; Mimouni, V.; Schoefs, B.; Bertrand, M. Plastids of Marine Phytoplankton Produce Bioactive Pigments and Lipids. Mar Drugs 2013, 11, 3425–3471. [Google Scholar] [CrossRef] [PubMed]
- Nandagopal, P.; Steven, A.N.; Chan, L.-W.; Rahmat, Z.; Jamaluddin, H.; Mohd Noh, N.I. Bioactive Metabolites Produced by Cyanobacteria for Growth Adaptation and Their Pharmacological Properties. Biology (Basel) 2021, 10, 1061. [Google Scholar] [CrossRef] [PubMed]
- Lauritano, C.; Andersen, J.H.; Hansen, E.; Albrigtsen, M.; Escalera, L.; Esposito, F.; Helland, K.; Hanssen, K.Ø.; Romano, G.; Ianora, A. Bioactivity Screening of Microalgae for Antioxidant, Anti-Inflammatory, Anticancer, Anti-Diabetes, and Antibacterial Activities. Front Mar Sci 2016, 3. [Google Scholar] [CrossRef]
- Samarakoon, K.W.; Ko, J.-Y.; Shah, Md.M.R.; Lee, J.-H.; Kang, M.-C.; Kwon, O.-N.; Lee, J.-B.; Jeon, Y.-J. In Vitro Studies of Anti-Inflammatory and Anticancer Activities of Organic Solvent Extracts from Cultured Marine Microalgae. ALGAE 2013, 28, 111–119. [Google Scholar] [CrossRef]
- Martínez Andrade, K.; Lauritano, C.; Romano, G.; Ianora, A. Marine Microalgae with Anti-Cancer Properties. Mar Drugs 2018, 16, 165. [Google Scholar] [CrossRef]
- Ingebrigtsen, R.A.; Hansen, E.; Andersen, J.H.; Eilertsen, H.C. Light and Temperature Effects on Bioactivity in Diatoms. J Appl Phycol 2016, 28, 939–950. [Google Scholar] [CrossRef]
- Prestegard, S.K.; Knutsen, G.; Herfindal, L. Adenosine Content and Growth in the Diatom Phaeodactylum Tricornutum (Bacillariophyceae): Effect of Salinity, Light, Temperature and Nitrate. Diatom Research 2014, 29, 361–369. [Google Scholar] [CrossRef]
- Varfolomeev, S.D.; Wasserman, L.A. Microalgae as Source of Biofuel, Food, Fodder, and Medicines. Appl Biochem Microbiol 2011, 47, 789–807. [Google Scholar] [CrossRef]
- Carbone, D.A.; Pellone, P.; Lubritto, C.; Ciniglia, C. Evaluation of Microalgae Antiviral Activity and Their Bioactive Compounds. Antibiotics 2021, 10, 746. [Google Scholar] [CrossRef]
- Kim, M.; Yim, J.H.; Kim, S.-Y.; Kim, H.S.; Lee, W.G.; Kim, S.J.; Kang, P.-S.; Lee, C.-K. In Vitro Inhibition of Influenza A Virus Infection by Marine Microalga-Derived Sulfated Polysaccharide p-KG03. Antiviral Res 2012, 93, 253–259. [Google Scholar] [CrossRef]
- Lauritano, C.; Martín, J.; de la Cruz, M.; Reyes, F.; Romano, G.; Ianora, A. First Identification of Marine Diatoms with Anti-Tuberculosis Activity. Sci Rep 2018, 8, 2284. [Google Scholar] [CrossRef] [PubMed]
- Moore, R.E.; Cheuk, C.; Yang, X.Q.G.; Patterson, G.M.L.; Bonjouklian, R.; Smitka, T.A.; Mynderse, J.S.; Foster, R.S.; Jones, N.D.; Swartzendruber, J.K.; et al. Hapalindoles, Antibacterial and Antimycotic Alkaloids from the Cyanophyte Hapalosiphon Fontinalis. J Org Chem 1987, 52, 1036–1043. [Google Scholar] [CrossRef]
- Ribeiro, M.C.M.; Salles, T.S.; Moreira, M.F.; Barbarino, E.; do Valle, A.F.; Couto, M.A.P.G. Antiviral Activity of Microalgae Extracts against Mayaro Virus. Algal Res 2022, 61, 102577. [Google Scholar] [CrossRef]
- Giordano, D.; Costantini, M.; Coppola, D.; Lauritano, C.; Núñez Pons, L.; Ruocco, N.; di Prisco, G.; Ianora, A.; Verde, C. Biotechnological Applications of Bioactive Peptides From Marine Sources. In; 2018; pp. 171–220.
- Guedes, A.; Gião, M.; Seabra, R.; Ferreira, A.; Tamagnini, P.; Moradas-Ferreira, P.; Malcata, F. Evaluation of the Antioxidant Activity of Cell Extracts from Microalgae. Mar Drugs 2013, 11, 1256–1270. [Google Scholar] [CrossRef] [PubMed]
- Mohamed Hatha, A.A.; Sumayya, N.S. Antioxidants from Marine Cyanobacteria. In Marine Antioxidants; Elsevier, 2023; pp. 119–131. [Google Scholar]
- Lauritano, C.; Helland, K.; Riccio, G.; Andersen, J.H.; Ianora, A.; Hansen, E.H. Lysophosphatidylcholines and Chlorophyll-Derived Molecules from the Diatom Cylindrotheca Closterium with Anti-Inflammatory Activity. Mar Drugs 2020, 18, 166. [Google Scholar] [CrossRef] [PubMed]
- Riccio, G.; Lauritano, C. Microalgae with Immunomodulatory Activities. Mar Drugs 2019, 18, 2. [Google Scholar] [CrossRef]
- Patipong, T.; Hibino, T.; Waditee-Sirisattha, R.; Kageyama, H. Induction of Antioxidative Activity and Antioxidant Molecules in the Halotolerant Cyanobacterium Halothece Sp. PCC7418 by Temperature Shift. Nat Prod Commun 2019, 14, 1934578X1986568. [Google Scholar] [CrossRef]
- Chang, Z.; Sitachitta, N.; Rossi, J. V.; Roberts, M.A.; Flatt, P.M.; Jia, J.; Sherman, D.H.; Gerwick, W.H. Biosynthetic Pathway and Gene Cluster Analysis of Curacin A, an Antitubulin Natural Product from the Tropical Marine Cyanobacterium Lyngbya m Ajuscula. J Nat Prod 2004, 67, 1356–1367. [Google Scholar] [CrossRef]
- Costa, M.; Garcia, M.; Costa-Rodrigues, J.; Costa, M.; Ribeiro, M.; Fernandes, M.; Barros, P.; Barreiro, A.; Vasconcelos, V.; Martins, R. Exploring Bioactive Properties of Marine Cyanobacteria Isolated from the Portuguese Coast: High Potential as a Source of Anticancer Compounds. Mar Drugs 2013, 12, 98–114. [Google Scholar] [CrossRef]
- Nagarajan, M.; Maruthanayagam, V.; Sundararaman, M. A Review of Pharmacological and Toxicological Potentials of Marine Cyanobacterial Metabolites. Journal of Applied Toxicology 2012, 32, 153–185. [Google Scholar] [CrossRef]
- Muller-Feuga, A.; Robert, R.; Cahu, C.; Robin, J.; Divanach, P. Uses of Microalgae in Aquaculture. In Live Feeds in Marine Aquaculture; Blackwell Science Ltd: Oxford, UK; pp. 253–299.
- Nagappan, S.; Das, P.; AbdulQuadir, M.; Thaher, M.; Khan, S.; Mahata, C.; Al-Jabri, H.; Vatland, A.K.; Kumar, G. Potential of Microalgae as a Sustainable Feed Ingredient for Aquaculture. J Biotechnol 2021, 341, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Latif, H.M.R.; El-Ashram, S.; Yilmaz, S.; Naiel, M.A.E.; Abdul Kari, Z.; Hamid, N.K.A.; Dawood, M.A.O.; Nowosad, J.; Kucharczyk, D. The Effectiveness of Arthrospira Platensis and Microalgae in Relieving Stressful Conditions Affecting Finfish and Shellfish Species: An Overview. Aquac Rep 2022, 24, 101135. [Google Scholar] [CrossRef]
- Bahi, A.; Ramos-Vega, A.; Angulo, C.; Monreal-Escalante, E.; Guardiola, F.A. Microalgae with Immunomodulatory Effects on Fish. Rev Aquac 2023. [Google Scholar] [CrossRef]
- Kim, C.H.; Leong, J.-A. FISH VIRUSES. In Encyclopedia of Virology; Elsevier, 1999; pp. 558–568. [Google Scholar]
- Padhi, A.; Verghese, B. Detecting Positively Selected Codons in the Glycoprotein of Spring Viraemia of Carp Virus (SVCV) Isolates from the USA and China. J Fish Dis 2008, 31, 785–791. [Google Scholar] [CrossRef] [PubMed]
- OIE Spring Viraemia of Carp. In Manual of Diagnostic Tests for Aquatic Animals; 2021; pp. 1–18.
- Ashraf, U.; Lu, Y.; Lin, L.; Yuan, J.; Wang, M.; Liu, X. Spring Viraemia of Carp Virus: Recent Advances. Journal of General Virology 2016, 97, 1037–1051. [Google Scholar] [CrossRef]
- Fouad, A.M.; Elkamel, A.A.; Ibrahim, S.; El-Matbouli, M.; Soliman, H.; Abdallah, E.S.H. Control of Spring Viremia of Carp in Common Carp Using RNA Interference. Aquaculture 2022, 559, 738417. [Google Scholar] [CrossRef]
- de Souza Valente, C.; Wan, A.H.L. Vibrio and Major Commercially Important Vibriosis Diseases in Decapod Crustaceans. J Invertebr Pathol 2021, 181, 107527. [Google Scholar] [CrossRef] [PubMed]
- Irshath, A.A.; Rajan, A.P.; Vimal, S.; Prabhakaran, V.-S.; Ganesan, R. Bacterial Pathogenesis in Various Fish Diseases: Recent Advances and Specific Challenges in Vaccine Development. Vaccines (Basel) 2023, 11, 470. [Google Scholar] [CrossRef] [PubMed]
- Sanches-Fernandes, G.M.M.; Sá-Correia, I.; Costa, R. Vibriosis Outbreaks in Aquaculture: Addressing Environmental and Public Health Concerns and Preventive Therapies Using Gilthead Seabream Farming as a Model System. Front Microbiol 2022, 13. [Google Scholar] [CrossRef] [PubMed]
- Semwal, A.; Kumar, A.; Kumar, N. A Review on Pathogenicity of Aeromonas Hydrophila and Their Mitigation through Medicinal Herbs in Aquaculture. Heliyon 2023, 9, e14088. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Gai, C.; Ye, G.; An, J.; Liu, K.; Xu, L.; Cao, H. Aeromonas Hydrophila, an Emerging Causative Agent of Freshwater-Farmed Whiteleg Shrimp Litopenaeus Vannamei. Microorganisms 2019, 7, 450. [Google Scholar] [CrossRef]
- Pękala, A.; Paździor, E.; Antychowicz, J.; Bernad, A.; Głowacka, H.; Więcek, B.; Niemczuk, W. Kocuria Rhizophila and Micrococcus Luteus as Emerging Opportunist Pathogens in Brown Trout ( Salmo Trutta Linnaeus, 1758) and Rainbow Trout ( Oncorhynchus Mykiss Walbaum, 1792). Aquaculture 2018, 486, 285–289. [Google Scholar] [CrossRef]
- El Sayed, K.A. Natural Products as Antiviral Agents. In; 2000; pp. 473–572.
- Geetha Bai, R.; Tuvikene, R. Potential Antiviral Properties of Industrially Important Marine Algal Polysaccharides and Their Significance in Fighting a Future Viral Pandemic. Viruses 2021, 13, 1817. [Google Scholar] [CrossRef]
- Reis, J.G.; Cadamuro, R.D.; Cabral, A.C.; Thaís da Silva, I.; Rodríguez-Lázaro, D.; Fongaro, G. Broad Spectrum Algae Compounds Against Viruses. Front Microbiol 2022, 12. [Google Scholar] [CrossRef]
- Falaise, C.; François, C.; Travers, M.-A.; Morga, B.; Haure, J.; Tremblay, R.; Turcotte, F.; Pasetto, P.; Gastineau, R.; Hardivillier, Y.; et al. Antimicrobial Compounds from Eukaryotic Microalgae against Human Pathogens and Diseases in Aquaculture. Mar Drugs 2016, 14, 159. [Google Scholar] [CrossRef]
- Pradhan, J.; Das, B.K.; Sahu, S.; Marhual, N.P.; Swain, A.K.; Mishra, B.K.; Eknath, A.E. Traditional Antibacterial Activity of Freshwater Microalga Spirulina Platensis to Aquatic Pathogens. Aquac Res 2012, 43, 1287–1295. [Google Scholar] [CrossRef]
- Rojas, V.; Rivas, L.; Cárdenas, C.; Guzmán, F. Cyanobacteria and Eukaryotic Microalgae as Emerging Sources of Antibacterial Peptides. Molecules 2020, 25, 5804. [Google Scholar] [CrossRef]
- Das, B.; Pradhan, J. Antibacterial Properties of Selected Freshwater Microalgae against Pathogenic Bacteria. Indian J Fisheries 2010, 57, 61–66. [Google Scholar]
- Ma, K.; Bao, Q.; Wu, Y.; Chen, S.; Zhao, S.; Wu, H.; Fan, J. Evaluation of Microalgae as Immunostimulants and Recombinant Vaccines for Diseases Prevention and Control in Aquaculture. Front Bioeng Biotechnol 2020, 8. [Google Scholar] [CrossRef] [PubMed]
- Velichkova, K.; Sirakov, I.; Denev, S. In Vitro Antibacterial Effect of Lemna Minuta, Chlorella Vulgaris and Spirulina Sp. Extracts against Fish Pathogen Aeromonas Hydrophila. Aquac Aquar Conserv Legis 2019, 12, 936–940. [Google Scholar]
- Chénais, B. Algae and Microalgae and Their Bioactive Molecules for Human Health. Molecules 2021, 26, 1185. [Google Scholar] [CrossRef]
- Monteiro, M.; Santos, R.A.; Iglesias, P.; Couto, A.; Serra, C.R.; Gouvinhas, I.; Barros, A.; Oliva-Teles, A.; Enes, P.; Díaz-Rosales, P. Effect of Extraction Method and Solvent System on the Phenolic Content and Antioxidant Activity of Selected Macro- and Microalgae Extracts. J Appl Phycol 2020, 32, 349–362. [Google Scholar] [CrossRef]
- Montero-Lobato, Z.; Vázquez, M.; Navarro, F.; Fuentes, J.; Bermejo, E.; Garbayo, I.; Vílchez, C.; Cuaresma, M. Chemically-Induced Production of Anti-Inflammatory Molecules in Microalgae. Mar Drugs 2018, 16, 478. [Google Scholar] [CrossRef]
- Bennett, J.M.; Reeves, G.; Billman, G.E.; Sturmberg, J.P. Inflammation–Nature’s Way to Efficiently Respond to All Types of Challenges: Implications for Understanding and Managing “the Epidemic” of Chronic Diseases. Front Med (Lausanne) 2018, 5. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory Responses and Inflammation-Associated Diseases in Organs. Oncotarget 2018, 9, 7204–7218. [Google Scholar] [CrossRef]
- Sullivan, C.; Soos, B.-L.; Millard, P.J.; Kim, C.H.; King, B.L. Modeling Virus-Induced Inflammation in Zebrafish: A Balance Between Infection Control and Excessive Inflammation. Front Immunol 2021, 12. [Google Scholar] [CrossRef]
- Elagoz, A.M.; Ambrosino, L.; Lauritano, C. De Novo Transcriptome of the Diatom Cylindrotheca Closterium Identifies Genes Involved in the Metabolism of Anti-Inflammatory Compounds. Sci Rep 2020, 10, 4138. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, J.; Kubota, T. Bioactive Metabolites from Marine Dinoflagellates. In Comprehensive Natural Products II; Elsevier, 2010; pp. 263–325. [Google Scholar]
- Shah, M.R.; Samarakoon, K.W.; Ko, J.Y.; Lakmal, H.H.C.; Lee, J.H.; An, S.J.; Jeon, Y.J.; Lee, J.B. Potentiality of Benthic Dinoflagellate Cultures and Screening of Their Bioactivities in Jeju Island, Korea. Afr J Biotechnol 2014, 13, 792–805. [Google Scholar] [CrossRef]
- Jusidin, M.R.; Othman, R.; Shaleh, S.R.M.; Ching, F.F.; Senoo, S.; Oslan, S.N.H. In Vitro Antibacterial Activity of Marine Microalgae Extract against Vibrio Harveyi. Applied Sciences 2022, 12, 1148. [Google Scholar] [CrossRef]
- Yun, H.-S.; Kim, Y.-S.; Yoon, H.-S. Effect of Different Cultivation Modes (Photoautotrophic, Mixotrophic, and Heterotrophic) on the Growth of Chlorella Sp. and Biocompositions. Front Bioeng Biotechnol 2021, 9. [Google Scholar] [CrossRef] [PubMed]
- Sanniyasi, E.; Venkatasubramanian, G.; Anbalagan, M.M.; Raj, P.P.; Gopal, R.K. In Vitro Anti-HIV-1 Activity of the Bioactive Compound Extracted and Purified from Two Different Marine Macroalgae (Seaweeds) (Dictyota Bartayesiana J.V.Lamouroux and Turbinaria Decurrens Bory). Sci Rep 2019, 9, 12185. [Google Scholar] [CrossRef]
- Kuda, T.; Nishizawa, M.; Toshima, D.; Matsushima, K.; Yoshida, S.; Takahashi, H.; Kimura, B.; Yamagishi, T. Antioxidant and Anti-Norovirus Properties of Aqueous Acetic Acid Macromolecular Extracts of Edible Brown Macroalgae. LWT 2021, 141, 110942. [Google Scholar] [CrossRef]
- Kristjánsson, J.; Rolfsson, Ó. Virucidal Activity of a Proprietary Blend of Plant-Based Oils (Viruxal) against SARS-CoV-2 and Influenza Viruses—An in Vitro Study. bioRxiv 2021, 1–11. [Google Scholar]
- Zakaryan, H.; Arabyan, E.; Oo, A.; Zandi, K. Flavonoids: Promising Natural Compounds against Viral Infections. Arch Virol 2017, 162, 2539–2551. [Google Scholar] [CrossRef]
- Monteiro, M.; Lavrador, A.S.; Santos, R.; Rangel, F.; Iglesias, P.; Tárraga, M.; Couto, A.; Serra, C.R.; Tafalla, C.; Da Costa, E.; et al. Evaluation of the Potential of Marine Algae Extracts as a Source of Functional Ingredients Using Zebrafish as Animal Model for Aquaculture. Marine Biotechnology 2021, 23, 529–545. [Google Scholar] [CrossRef] [PubMed]
- Lassudrie, M.; Soudant, P.; Nicolas, J.-L.; Fabioux, C.; Lambert, C.; Miner, P.; Le Grand, J.; Petton, B.; Hégaret, H. Interaction between Toxic Dinoflagellate Alexandrium Catenella Exposure and Disease Associated with Herpesvirus OsHV-1 ΜVar in Pacific Oyster Spat Crassostrea Gigas. Harmful Algae 2015, 45, 53–61. [Google Scholar] [CrossRef]
- Fabra, M.J.; Falcó, I.; Randazzo, W.; Sánchez, G.; López-Rubio, A. Antiviral and Antioxidant Properties of Active Alginate Edible Films Containing Phenolic Extracts. Food Hydrocoll 2018, 81, 96–103. [Google Scholar] [CrossRef]
- Yim, J.H.; Kim, S.J.; Ahn, S.H.; Lee, H.K. Characterization of a Novel Bioflocculant, p-KG03, from a Marine Dinoflagellate, Gyrodinium Impudicum KG03. Bioresour Technol 2007, 98, 361–367. [Google Scholar] [CrossRef]
- Azizan, A.; Ahamad Bustamam, M.; Maulidiani, M.; Shaari, K.; Ismail, I.; Nagao, N.; Abas, F. Metabolite Profiling of the Microalgal Diatom Chaetoceros Calcitrans and Correlation with Antioxidant and Nitric Oxide Inhibitory Activities via 1H NMR-Based Metabolomics. Mar Drugs 2018, 16, 154. [Google Scholar] [CrossRef]
- Abidizadegan, M.; Peltomaa, E.; Blomster, J. The Potential of Cryptophyte Algae in Biomedical and Pharmaceutical Applications. Front Pharmacol 2021, 11. [Google Scholar] [CrossRef]
- Parra-Riofrio, G.; Moreno, P.; García-Rosado, E.; Alonso, M.C.; Uribe-Tapia, E.; Abdala-Diaz, R.T.; Bejar, J. Tetraselmis Suecica and Porphyridium Cruentum Exopolysaccharides Show Anti-VHSV Activity on RTG-2 Cells. Aquaculture International 2023. [Google Scholar] [CrossRef]
- Raposo, M.; De Morais, R.; Bernardo de Morais, A. Bioactivity and Applications of Sulphated Polysaccharides from Marine Microalgae. Mar Drugs 2013, 11, 233–252. [Google Scholar] [CrossRef]
- Desbois, A.P.; Smith, V.J. Antibacterial Free Fatty Acids: Activities, Mechanisms of Action and Biotechnological Potential. Appl Microbiol Biotechnol 2010, 85, 1629–1642. [Google Scholar] [CrossRef]
- Agoramoorthy, G.; Chandrasekaran, M.; Venkatesalu, V.; Hsu, M.J. Antibacterial and Antifungal Activities of Fatty Acid Methyl Esters of the Blind-Your-Eye Mangrove from India. Brazilian Journal of Microbiology 2007, 38, 739–742. [Google Scholar] [CrossRef]
- Ramos, D.F.; Bartolomeu Halicki, P.C.; da Silva Canielles Caprara, C.; Borges, P.; da R., M. D’Oca, C.; de Fátima C. Santos, M.; D’Oca, M.G.M.; Roselet, F.; Almeida da Silva, P.E.; Abreu, P.C. Chemical Profile and Antimicrobial Activity of the Marine Diatom Chaetoceros Muelleri. Chem Biodivers 2022, 19. [Google Scholar] [CrossRef]
- Viso, A.C.; Pesando, D.; Baby, C. Antibacterial and Antifungal Properties of Some Marine Diatoms in Culture. botm 1987, 30, 41–46. [Google Scholar] [CrossRef]
- Asha shalini, A.; Syed Ali, M.; Anuradha, V.; Yogananth, N.; Bhuvana, P. GCMS Analysis and Invitro Antibacterial and Anti-Inflammatory Study on Methanolic Extract of Thalassiosira Weissflogii. Biocatal Agric Biotechnol 2019, 19, 101148. [Google Scholar] [CrossRef]
- Nur Fadillah, S.; Natsir, H.; Ahmad, A.; Karim, A.; Taba, P.; Nur fadillah, S. Extraction and Fractionation of Active Protein from Microalgae Nitzschia Sp. as Antimicrobial Agent. Egypt J Chem 2022, 0, 0–0. [Google Scholar] [CrossRef]
- Long, M.; Krock, B.; Castrec, J.; Tillmann, U. Unknown Extracellular and Bioactive Metabolites of the Genus Alexandrium: A Review of Overlooked Toxins. Toxins (Basel) 2021, 13, 905. [Google Scholar] [CrossRef]
- Nagai, H.; Satake, M.; Yasumoto, T. Antimicrobial Activities of Polyether Compounds of Dinoflagellate Origins. J Appl Phycol 1990, 2, 305–308. [Google Scholar] [CrossRef]
- Mussai, P.; Larsen, J.; Alrefaei, A.F.; Jeewon, R. Ribosomal DNA Sequence-Based Taxonomy and Antimicrobial Activity of Prorocentrum Spp. (Dinophyceae) from Mauritius Coastal Waters, South-West Indian Ocean. Mar Drugs 2023, 21, 216. [Google Scholar] [CrossRef]
- Wencheng, L.; Cho, K.; Yamasaki, Y.; Takeshita, S.; Hwang, K.; Kim, D.; Oda, T. Photo-Induced Antibacterial Activity of a Porphyrin Derivative Isolated from the Harmful Dinoflagellate Heterocapsa Circularisquama. Aquatic Toxicology 2018, 201, 119–128. [Google Scholar] [CrossRef]
- Zhu, P.; You, Y.; Chu, J.; Jin, H.; Yan, X. [Screening and Characterization of Antibacterial and Cytotoxic Marine Bacteria Associated with Karlodinium Micrum]. Wei Sheng Wu Xue Bao 2010, 50, 1044–1050. [Google Scholar]
- Quijano-Scheggia, S. The Inhibitory Effect of a Non-Yessotoxin-Producing Dinoflagellate, Lingulodinium Polyedrum (Stein) Dodge, towards Vibrio Vulnificus and Staphylococcus Aureus. Rev Biol Trop 2016, 64, 805. [Google Scholar] [CrossRef]
- Aveiro, S.S.; Melo, T.; Figueiredo, A.; Domingues, P.; Pereira, H.; Maia, I.B.; Silva, J.; Domingues, M.R.; Nunes, C.; Moreira, A.S.P. The Polar Lipidome of Cultured Emiliania Huxleyi: A Source of Bioactive Lipids with Relevance for Biotechnological Applications. Biomolecules 2020, 10, 1434. [Google Scholar] [CrossRef]
- Guzmán; Wong; Román; Cárdenas; Alvárez; Schmitt; Albericio; Rojas Identification of Antimicrobial Peptides from the Microalgae Tetraselmis Suecica (Kylin) Butcher and Bactericidal Activity Improvement. Mar Drugs 2019, 17, 453. [CrossRef]
- Cordero-Maldonado, M.L.; Siverio-Mota, D.; Vicet-Muro, L.; Wilches-Arizábala, I.M.; Esguerra, C. V.; de Witte, P.A.M.; Crawford, A.D. Optimization and Pharmacological Validation of a Leukocyte Migration Assay in Zebrafish Larvae for the Rapid In Vivo Bioactivity Analysis of Anti-Inflammatory Secondary Metabolites. PLoS One 2013, 8, e75404. [Google Scholar] [CrossRef]
- Kim, J.; Choi, Y.K.; Lee, J.-H.; Kim, S.-Y.; Kim, H.-S. “Anti-Inflammatory Activity on LPS-Stimulated in Vitro RAW 264.7 Cells and in Vivo Zebrafish of Heterosigma Akshiwo.”. Journal of Chitin and Chitosan 2017, 22, 185–193. [Google Scholar] [CrossRef]
- Mathias, J.R.; Perrin, B.J.; Liu, T.-X.; Kanki, J.; Look, A.T.; Huttenlocher, A. Resolution of Inflammation by Retrograde Chemotaxis of Neutrophils in Transgenic Zebrafish. J Leukoc Biol 2006, 80, 1281–1288. [Google Scholar] [CrossRef]
- Renshaw, S.A.; Loynes, C.A.; Trushell, D.M.I.; Elworthy, S.; Ingham, P.W.; Whyte, M.K.B. A Transgenic Zebrafish Model of Neutrophilic Inflammation. Blood 2006, 108, 3976–3978. [Google Scholar] [CrossRef]
- Xie, Y.; Meijer, A.H.; Schaaf, M.J.M. Modeling Inflammation in Zebrafish for the Development of Anti-Inflammatory Drugs. Front Cell Dev Biol 2021, 8. [Google Scholar] [CrossRef]
- Gugulothu, P. “Bioactive Compound from Micro Algae and Their Anti-Cancer Properties.”. Biomed J Sci Tech Res 2022, 42. [Google Scholar] [CrossRef]
- Hussein, H.A.; Abdullah, M.A. Anticancer Compounds Derived from Marine Diatoms. Mar Drugs 2020, 18, 356. [Google Scholar] [CrossRef]
- Saxena, A.; Raj, A.; Tiwari, A. Exploring the Anti-Cancer Potential of Microalgae. In Progress in Microalgae Research - A Path for Shaping Sustainable Futures; IntechOpen, 2022. [Google Scholar]
- Skjånes, K.; Aesoy, R.; Herfindal, L.; Skomedal, H. Bioactive Peptides from Microalgae: Focus on Anti-cancer and Immunomodulating Activity. Physiol Plant 2021, 173, 612–623. [Google Scholar] [CrossRef]
- Boyd, M.R.; Paull, K.D. Some Practical Considerations and Applications of the National Cancer Institute in Vitro Anticancer Drug Discovery Screen. Drug Dev Res 1995, 34, 91–109. [Google Scholar] [CrossRef]
- McCauley, J.; Zivanovic, A.; Skropeta, D. Bioassays for Anticancer Activities. In; 2013; pp. 191–205.
- Emtyazjoo, Mo.; Moghadasi, Z.; Rabbani, M.; Emtyazjoo, Ma.; Samadi, S.; Mossaffa, N. Anticancer Effect of Dunaliella Salina under Stress and Normal Conditions against Skin Carcinoma Cell Line A431 in Vitro. Iran J Fish Sci 2012, 11, 283–293. [Google Scholar]
- Kobayashi, J. Amphidinolides and Its Related Macrolides from Marine Dinoflagellates. J Antibiot (Tokyo) 2008, 61, 271–284. [Google Scholar] [CrossRef]
- Ebrahimi Nigjeh, S.; Yusoff, F.M.; Mohamed Alitheen, N.B.; Rasoli, M.; Keong, Y.S.; Omar, A.R. bin Cytotoxic Effect of Ethanol Extract of Microalga, Chaetoceros Calcitrans, and Its Mechanisms in Inducing Apoptosis in Human Breast Cancer Cell Line. Biomed Res Int 2013, 2013, 1–8. [Google Scholar] [CrossRef]
- Umemura, K.; Yanase, K.; Suzuki, M.; Okutani, K.; Yamori, T.; Andoh, T. Inhibition of DNA Topoisomerases I and II, and Growth Inhibition of Human Cancer Cell Lines by a Marine Microalgal Polysaccharide. Biochem Pharmacol 2003, 66, 481–487. [Google Scholar] [CrossRef]
- Sanniyasi, E.; Patrick, A.P.R.; Rajagopalan, K.; Gopal, R.K.; Damodharan, R. Characterization and in Vitro Anticancer Potential of Exopolysaccharide Extracted from a Freshwater Diatom Nitzschia Palea (Kütz.) W.Sm. 1856. Sci Rep 2022, 12, 22114. [Google Scholar] [CrossRef]
- Cutignano, A.; Conte, M.; Tirino, V.; Del Vecchio, V.; De Angelis, R.; Nebbioso, A.; Altucci, L.; Romano, G. Cytotoxic Potential of the Marine Diatom Thalassiosira Rotula: Insights into Bioactivity of 24-Methylene Cholesterol. Mar Drugs 2022, 20, 595. [Google Scholar] [CrossRef]
- Edwards, D.J.; Marquez, B.L.; Nogle, L.M.; McPhail, K.; Goeger, D.E.; Roberts, M.A.; Gerwick, W.H. Structure and Biosynthesis of the Jamaicamides, New Mixed Polyketide-Peptide Neurotoxins from the Marine Cyanobacterium Lyngbya Majuscula. Chem Biol 2004, 11, 817–833. [Google Scholar] [CrossRef]
- Reed, L.J.; Muench, H. A Simple Method of Estimating Fifty Percent Endpoints. Am J Hyg 1938, 27, 493–497. [Google Scholar]
- Hall, C.; Flores, M.V.; Storm, T.; Crosier, K.; Crosier, P. The Zebrafish Lysozyme C Promoter Drives Myeloid-Specific Expression in Transgenic Fish. BMC Dev Biol 2007, 7, 42. [Google Scholar] [CrossRef]
Figure 1.
Antiviral activity of the extracts against SVCV. The different algal groups selected are indicated. The viral titer was determined at day 6 post-infection as the viral dilution that causes an infection of 50% of the cell line (TCID50). Results were expressed as the mean and SD of four titrations. A T-test was used to determine significant differences at p-value < 0.05 (*) between control (SVCV infected cells) and cells treated with the extracts at day 6 post-infection. Using the calculated percentage of inhibition (PI), the antiviral activity was scored as strong (PI>90%), moderate (PI between 50 and 90%) and weak (PI<50%) according to Monteiro et al. [55].
Figure 1.
Antiviral activity of the extracts against SVCV. The different algal groups selected are indicated. The viral titer was determined at day 6 post-infection as the viral dilution that causes an infection of 50% of the cell line (TCID50). Results were expressed as the mean and SD of four titrations. A T-test was used to determine significant differences at p-value < 0.05 (*) between control (SVCV infected cells) and cells treated with the extracts at day 6 post-infection. Using the calculated percentage of inhibition (PI), the antiviral activity was scored as strong (PI>90%), moderate (PI between 50 and 90%) and weak (PI<50%) according to Monteiro et al. [55].
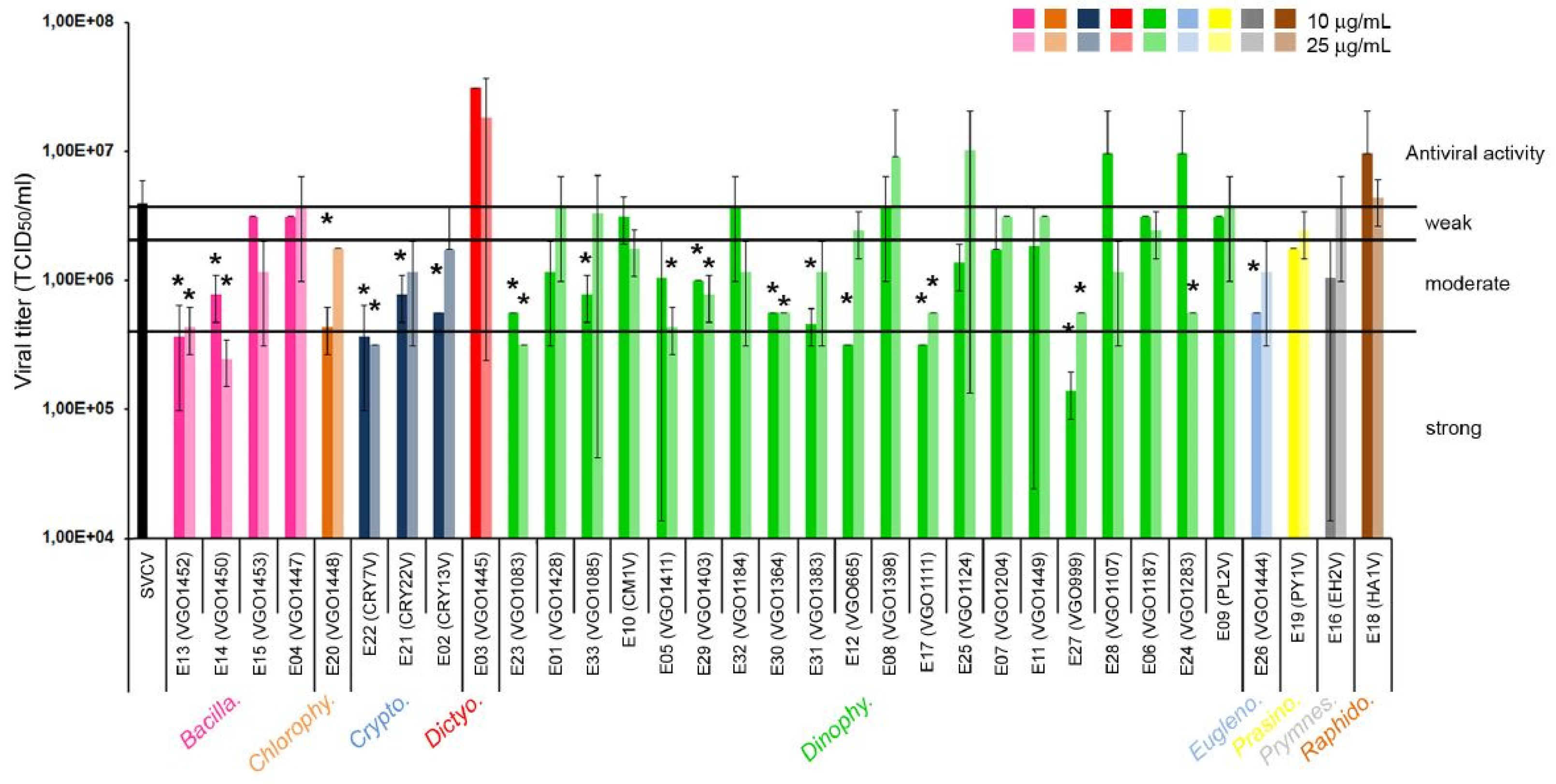
Figure 2.
The antibacterial activity of extracts (10 μg mL-1) against Gram (-) and Gram (+) bacteria. The different algal groups selected are indicated. The bacterial growth was evaluated by measuring the OD600 nm for 24h: (A) The kinetics obtained in samples treated with the extract 9 was selected as representative result; (B and C) T-test was used to determine significant differences at p-value < 0.05 (*). Tables show the sampling points where the differences in OD 600 nm were statistically significant at p<0.05 against A. hydrophyla (B) and M. luteus (C). The percentage of bacterial growth reduction is specified in each sampling point.
Figure 2.
The antibacterial activity of extracts (10 μg mL-1) against Gram (-) and Gram (+) bacteria. The different algal groups selected are indicated. The bacterial growth was evaluated by measuring the OD600 nm for 24h: (A) The kinetics obtained in samples treated with the extract 9 was selected as representative result; (B and C) T-test was used to determine significant differences at p-value < 0.05 (*). Tables show the sampling points where the differences in OD 600 nm were statistically significant at p<0.05 against A. hydrophyla (B) and M. luteus (C). The percentage of bacterial growth reduction is specified in each sampling point.
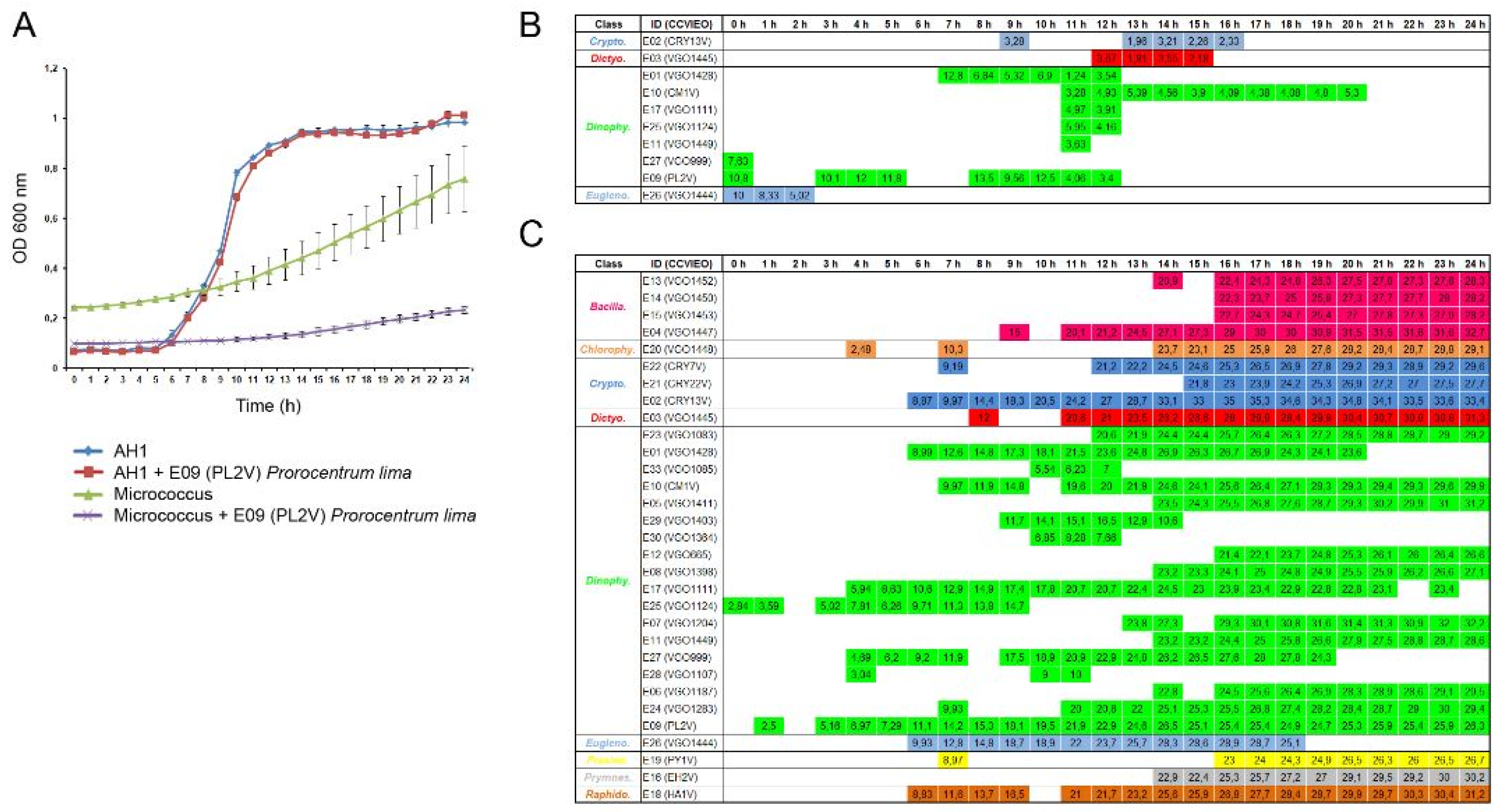
Figure 3.
In vivo evaluation of the anti-inflammatory activity of the extracts (at 25 μg mL-1) using zebrafish larvae. The different algal groups selected are indicated. (A) The transgenic zebrafish larvae Tg(lyz:DsRed2) was used. The lysozyme-expressing cells (neutrophils) are marked in red and can be analyzed in live animals; (B) Control animals were injured in the tail and the number of neutrophils was measured at 2h, 24h and 48h. Scale bar = 100 μm; (C) Evolution in the number of neutrophils at the injury in control fish. The graph was created using information obtained from 50 animals; (D). Effect of the extracts on the number of neutrophils at the injured fin. Results represent the mean and SD. (*) asterisks represent significant differences (p<0.05) compared to controls.
Figure 3.
In vivo evaluation of the anti-inflammatory activity of the extracts (at 25 μg mL-1) using zebrafish larvae. The different algal groups selected are indicated. (A) The transgenic zebrafish larvae Tg(lyz:DsRed2) was used. The lysozyme-expressing cells (neutrophils) are marked in red and can be analyzed in live animals; (B) Control animals were injured in the tail and the number of neutrophils was measured at 2h, 24h and 48h. Scale bar = 100 μm; (C) Evolution in the number of neutrophils at the injury in control fish. The graph was created using information obtained from 50 animals; (D). Effect of the extracts on the number of neutrophils at the injured fin. Results represent the mean and SD. (*) asterisks represent significant differences (p<0.05) compared to controls.
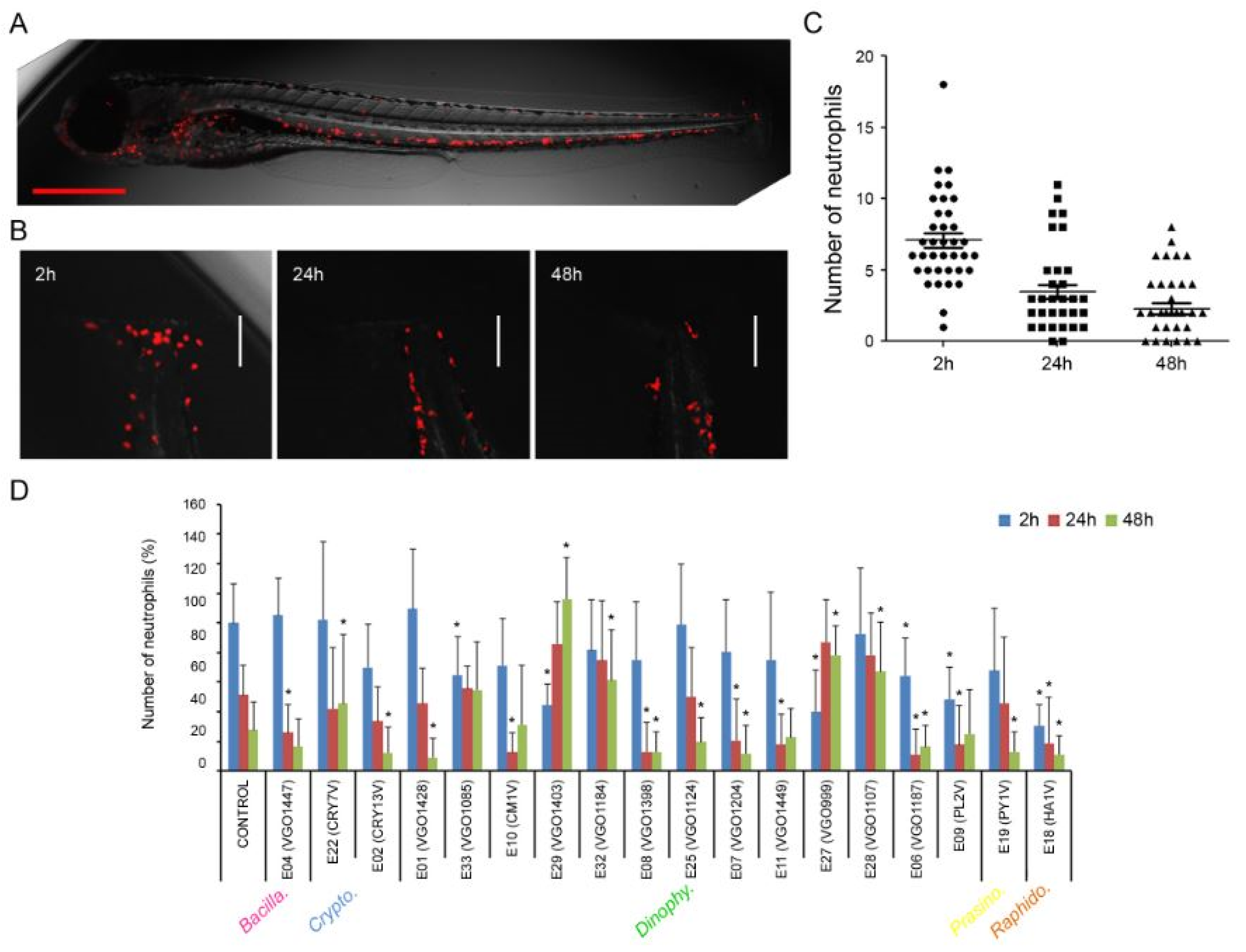
Figure 4.
Anti-cancer activity of the extracts against HCT 116, HepG2 and MG-63. Results represent the mean and SD of three independent experiments. STP (staurosporine).
Figure 4.
Anti-cancer activity of the extracts against HCT 116, HepG2 and MG-63. Results represent the mean and SD of three independent experiments. STP (staurosporine).
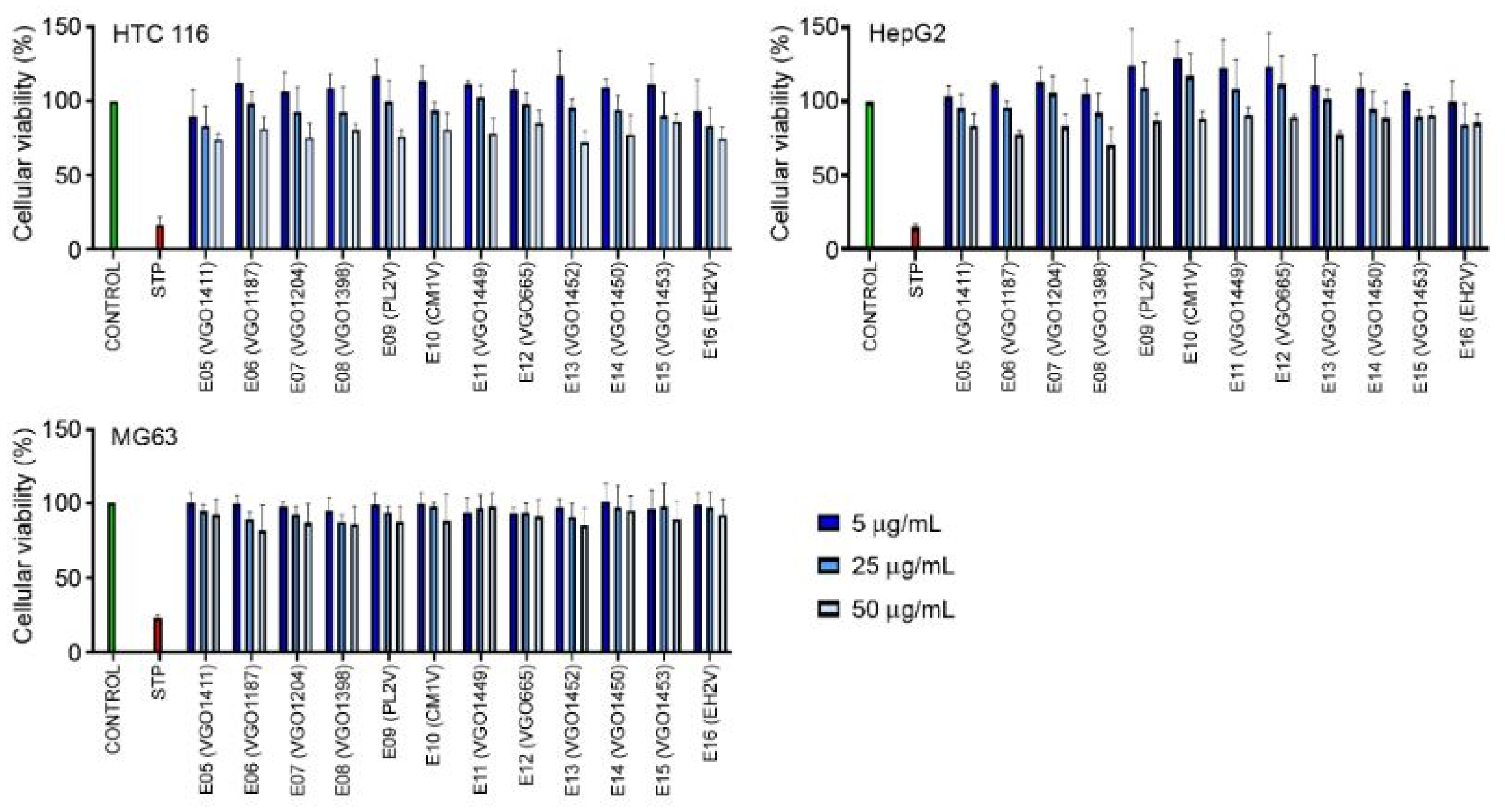

Table 1.
Microalgal strains selected in the present study and culture conditions. L1 codes for L1 medium without silicates.
Table 1.
Microalgal strains selected in the present study and culture conditions. L1 codes for L1 medium without silicates.
| Class | Species | ID (CCVIEO) | Temp (ºC) | Irradiance( E m2s-1) | Culture medium |
|---|---|---|---|---|---|
| Bacillariophyceae | Chaetoceros dichatoensis | E13 (VGO1452) | 16 | 150 | L1(+Si) |
| Nitzschia sp. | E14 (VGO1450) | 16 | 150 | L1(+Si) | |
| Pseudonitzschia australis | E15 (VGO1453) | 16 | 150 | L1(+Si) | |
| Thalassiosira delicatula | E04 (VGO1447) | 16 | 150 | L1(+Si) | |
| Chlorophyceae | Tetraselmis convolutae | E20 (VGO1448) | 16 | 150 | L1 |
| Cryptophyceae | Falcomonas sp. | E22 (CRY7V) | 16 | 150 | L1 |
| Guillardia theta | E21 (CRY22V) | 16 | 150 | L1 | |
| Teleaulax amphioxeia | E02 (CRY13V) | 16 | 150 | L1 | |
| Dictyochophyceae | Pseudopedinella elastica | E03 (VGO1445) | 19 | 150 | L1 |
| Dinophyceae | Alexandrium mediterraneum | E23 (VGO1083) | 19 | 150 | L1 |
| Alexandrium minutum | E01 (VGO1428) | 19 | 150 | L1 | |
| Alexandrium tamarense | E33 (VGO1085) | 19 | 150 | L1 | |
| Coolia monotis | E10 (CM1V) | 19 | 150 | L1 | |
| Dinophysis acuminata | E05 (VGO1411) | 19 | 150 | L1 (/20) | |
| Dinophysis caudata | E29 (VGO1403) | 19 | 150 | L1 (/20) | |
| Gambierdiscus australes | E32 (VGO1184) | 25 | 80 | K/2 | |
| Gambierdiscus caribaeus | E30 (VGO1364) | 25 | 80 | K/2 | |
| Gambierdiscus excentricus | E31 (VGO1383) | 25 | 80 | K/2 | |
| Gymnodinium impudicum | E12 (VGO665) | 19 | 150 | L1 | |
| Heterocapsa minima | E08 (VGO1398) | 19 | 150 | L1 | |
| Karlodinium veneficum | E17 (VGO1111) | 19 | 150 | L1 | |
| Kryptoperidinium foliaceum | E25 (VGO1124) | 19 | 150 | L1 | |
| Lingulodinium polyedrum | E07 (VGO1204) | 19 | 150 | L1 | |
| Matsuokaea loeblichii | E11 (VGO1449) | 19 | 150 | L1 | |
| Ostreopsis fattorussoi | E27 (VGO999) | 25 | 80 | L1 | |
| Ostreopsis cf. ovata | E28 (VGO1107) | 25 | 80 | L1 | |
| Ostreopsis siamensis | E06 (VGO1187) | 19 | 150 | L1 | |
| Prorocentrum hoffmannianum | E24 (VGO1283) | 25 | 80 | L1 | |
| Prorocentrum lima | E09 (PL2V) | 19 | 150 | L1 | |
| Euglenophyceae | Eutreptiella gymnastica | E26 (VGO1444) | 19 | 150 | L1 |
| Prasinophyceae | Pyramimonas sp. | E19 (PY1V) | 19 | 150 | L1 |
| Prymnesiophyceae | Emiliania huxleyi | E16 (EH2V) | 19 | 150 | L1 |
| Raphidophyceae | Heterosigma akashiwo | E18 (HA1V) | 19 | 150 | L1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



