
Submitted:
06 March 2024
Posted:
07 March 2024
You are already at the latest version
Abstract
Background: This study explored maternal gut, breast milk and infant gut microbiomes as possi-ble mediators of the observed effects of a relaxation intervention which reduced maternal stress and promoted infant weight gain. Methods: A randomized controlled trial was conducted in healthy Chinese primiparous mother-infant pairs. Mothers were randomly assigned to interven-tion group (IG, listening to relaxation meditation) or control group (CG). Outcomes were differ-ences in microbiome composition and diversity in maternal gut, breast milk and infant gut at 1- and 8-weeks between IG and CG, assessed by 16S rRNA gene amplicon sequencing of fecal and breastmilk samples. Results: 38 mother-infant pairs were included in this analysis (IG=19, CG=19). Overall microbiome community structure in the maternal gut was significantly different between IG and CG at 1-week and the difference was more significant at 8 weeks (Bray-Curtis distance R2=0.04 vs. R2=0.13). Post-intervention, the α-diversity was significantly lower in IG breast milk (observed features: CG=295 vs. IG=255, p=0.032); the Bifidobacterium genera presented higher relative abundance. In parallel, significantly higher α-diversity was observed in IG infant gut (observed features: CG=73 vs. IG=113, p
Keywords:
breastfeeding
; gut microbiome
; maternal stress
; infant weight
; mother-infant signalling
1. Introduction
Stress has an influence on the structure of the microbiota community in the gastrointestinal (GI) tract [1,2], potentially through the ‘gut-brain axis’ [3,4]. During lactation, the GI tract microbiome of mothers affected by stress could influence the milk microbiome via entero-mammary trafficking, which refers to the movement of microbiota from the maternal gut to the mammary gland [5,6,7]. Moreover, human breast milk contains bacteria including Lactic acid bacteria and Bifidobacteria [8] and several studies suggest that breast milk microbes influence the infant gut microbiome [9,10,11]. An emerging paradigm indicates that maternal psychological status could be associated with alterations in infant gut microbiome diversity [12] (eFigure 1, supplemental file).
The gut of the newborn infant is rapidly colonized by numerous microbes. The early life microbiome is known to influence infant development, and gut microbiome imbalances have been linked with an increased risk of certain autoimmune diseases [13]. The development of the neonatal gut microbiome can be influenced by multiple factors, including delivery mode (hence differential exposure to the maternal microbiome), feeding method, antibiotic intake in early life, stress and genetic factors [14,15]. Furthermore, the microbe and human milk oligosaccharide (HMO) content of human breast milk have a profound influence on early gut colonization of the infant [15,16,17].
We have shown in three randomized controlled trials (RCT) that a simple relaxation intervention significantly reduces psychosocial stress in breastfeeding women, whilst improving infant weight gain [18,19,20]. The present study analyzed data from one of these RCTs and investigated the role of the microbiome as a potential mediator of the observed intervention effects on weight gain in late preterm and early term infants (born at 34+0 to 37+6 weeks of gestation), a group that is relatively neglected compared to infants born at term or before 34 weeks. We used 16S rRNA gene sequencing to analyze the microbiome in the maternal gut (fecal), breast milk and infant gut (fecal) samples.
2. Materials and Methods
Details of the trial design and main outcomes have been published [18,21]. All mothers completed the 8-week data collection before COVID-19 lockdown in Beijing.
2.1. Study Design and Participants
Healthy primiparous mothers with a singleton infant born between 34+0-37+6 weeks gestation who aimed to exclusively breastfeed (EBF) for at least two months were recruited 3-5 days after birth. Recruitment was conducted in community clinics affiliated to Beijing Children’s Hospital in Beijing, China. Data collection was conducted through two home visits around 1-week and 8-weeks postpartum. To ensure consistency of procedures at each study center, all research assistants and nurses involved in the study attended training courses prior to the start of recruitment. Standard operating procedures for the study were printed and posted at each center. To control for the known effect of delivery mode on the infant microbiota, the present study only analyzed data from a subset of participants who delivered vaginally and provided maternal fecal, breast milk, and infant fecal samples at baseline (1-week) and 8 weeks home visits. The study was approved by the Research Ethics Committee of University College London (ID: 12681/002) and the Department of Child Health, Beijing Children’s Hospital (ID: 2018-167).
2.2. Randomization, Procedures, and Intervention
After obtaining written informed consent, participants were randomly assigned to either intervention group (IG, listening to relaxation meditation at least once a day with standard postpartum care from local clinics) or control group (CG, standard postpartum care from local clinics). The randomization sequence was computer generated by an independent researcher, and stratified by gestational age (34-35 versus 36-37 weeks), delivery method (vaginal versus caesarean) and recruitment location. Assignments were stored in sealed, opaque envelopes at Beijing Children’s Hospital. Participants were not told about the randomization until the end of the study; they were aware that the aim of the study is to investigate factors that could optimising breastfeeding outcomes. Nurses who collected the samples were aware of the groups; however, they had no particular interest or investment in the result of this study. The research technicians at Novogene who performed the 16s rRNA sequencing were blinded to the randomization status of the subjects.
2.3. Outcomes and Measures
Outcomes of the present study were differences in microbiome composition and diversity in maternal gut, breast milk and infant gut at 1- and 8-weeks between IG and CG, assessed by 16S rRNA gene amplicon sequencing of fecal and breastmilk samples.
Baseline characteristics of the mother-infant pairs were obtained using demographic questionnaires. The breast milk and maternal fecal samples were collected by mothers following the method in the published study protocol [18], mothers were asked to clean their areolar skin before collecting milk samples. Infant samples were collected by the nurse from infant diapers during home visits. All samples were collected into sterile tubes and stored at -80⁰C at the laboratory of Beijing Children’s Hospital. Samples for inclusion in the microbiome analysis were transported to the laboratory of Novogene Technology Inc. (Beijing, China) where the DNA extraction, library preparation, and the 16S rRNA gene amplicon sequencing was performed using standard procedures (eMethods in Supplemental file).
2.4. Statistical Analysis
Statistical analyses were conducted using R (version 4.12), and SPSS (version 26.0). We compared baseline characteristics, maternal stress and infant weight gain data between the 38 selected mother-infant pairs and the remainder of the 96 mother-infant pairs not involved in this analysis. We compared the diversity differences in maternal fecal, breast milk, and infant fecal samples between IG and CG at 1-and 8-weeks. The differences in α-diversity between IG and CG were examined by Wilcoxon rank-sum test using observed features. Differences in β-diversity between IG and CG were examined using Bray-Curtis dissimilarity metrics presented on a principal coordinates analysis plot (PCoA) and with differences between groups determined by ANOSIM. The relative abundance of the top 15 most abundant genera in all samples were examined and statistical differences between IG and CG samples were compared using the Wilcoxon rank-sum test and FDR adjusted p-value.
To determine the abundance of specific bacteria and potential associations with maternal stress and infant weight gain, Spearman-rank correlation was used, and results were presented in heatmaps. A standard p-value of < 0.05 was considered statistically significant for all analyses.
3. Results
3.1. Study Population and Baseline Characteristics
The original trial was conducted from October 2018 to October 2020. Of the 96 mother-infant pairs enrolled in the original trial, 38 eligible pairs were included in this secondary analysis (Figure 1, IG=19, CG=19); those 38 eligible pairs were recruited from February 2019 to January 2020. All mothers followed the traditional Chinese postpartum confinement practice during the data collection. There were no significant differences in baseline characteristics between those who were included in the microbiome analysis versus those that were not (eTable 1 in supplemental file). All 228 samples (38 maternal fecal, breast milk, and infant fecal samples at 1-and 8-weeks) were analyzed (Rarefaction curves are shown in eFigure 2). Distinct bacterial communities were observed in maternal gut, breast milk, and infant gut (eFigure 3 in supplemental file). Demographic characteristics of the participants are outlined in Table 1; there were no significant differences in participant characteristics between IG and CG at baseline.
3.2. Microbiome Composition and Diversity in the Maternal Gut
The number of observed bacterial taxa (observed features) in the maternal gut was not significantly different between IG and CG at both 1 and 8 weeks (Figure 2A & eTable 2).; moreover, the number of observed features did not significantly change between 1 and 8 weeks in either IG or CG. The overall composition of maternal gut microbiome (β-diversity) was significantly different between IG and CG at 1-week (Figure 2B, Bray-Curtis distance, R2=0.04, p=0.026) as determined by Bray-Curtis dissimilarity metrics. However, the separation between IG and CG was stronger at 8 weeks (Figure 2C, Bray-Curtis distance, R2=0.13, p=0.001).
3.3. Microbiome Composition and Diversity in Breast Milk
The number of observed bacterial features in breast milk was not significantly different between IG and CG at 1-week, but it was significantly lower in IG relative to CG at 8 weeks (Figure 2D & eTable 2, observed features 295 vs. 255, p=0.032). However, the difference lost significance after adjusting for 1-week baseline value (coefficient B= 37.5, 95%CI -42, 117, p=0.3). The overall bacterial composition was not significantly different between IG and CG at 1- or 8 weeks as determined by Bray-Curtis dissimilarity metrics (Figure 2E and 2F).
3.4. Microbiome Composition and Diversity in Infant Gut
Whilst no significant group differences were observed in infant gut microbiome at 1-week, the IG infant presented significantly higher evenness (eTable 2 & eFigure 4, Shannon index 1.94 vs. 2.27, p=0.015) and significantly higher number of observed bacterial features at 8 weeks (Figure 2G, observed features 73 vs. 113, p<0.001); interestingly, the difference was still significant after adjusting for 1-week baseline value (coefficient B=40.8, 95%CI 15.7, 65.9, p=0.002). The overall composition as determined by Bray-Curtis dissimilarity metrics was not significantly different between IG and CG at 1 (Figure 2H) or 8 weeks (Figure 2I).
3.5. Differences in Microbiome Composition in Maternal Gut, Breast Milk and Infant Gut between Groups
The top 15 most abundant bacteria in maternal gut, breast milk, and infant gut at 8 weeks after intervention were assessed, with group differences between IG and CG presented in Figure 3 and baseline group comparison presented in eFigure 5 (supplemental file). The relative abundance of some genera showed significant group differences at 8-weeks with no group difference at baseline, such as lower Veillonella and higher Faecalibacterium in the IG maternal gut at 8 weeks. In breast milk, on the other hand, Veillonella was significantly higher in IG at both baseline and 8 weeks. Veillonella was one of the common gut-associated obligate anaerobic genera shared between maternal gut, breast milk and infant gut, whilst Faecalibacterium was commonly shared between maternal gut and breast milk. Two other gut-associated anaerobic genera, Bifidobacterium and Blautia, both had higher relative abundance in the IG infant gut at 8 weeks, but this was only significant for Blautia. Moreover, although not significant, the relative abundance of Bifidobacterium in breast milk was lower in IG at baseline (eFigure 5 and eFigure 6 in supplemental file) but higher at 8 weeks after intervention (Figure 3). The relative abundance of two putative pathogens, Enterococcus and Acinetobacter, was also significantly lower in IG at 8 weeks.
3.6. Correlation between the Top 15 Microbial and Maternal Stress/Infant Weight
We examined whether the infant gut microbiome 8 weeks was correlated with infant weight at 8 weeks and weight gain from 1 to 8 weeks, as well as with maternal stress at 8 weeks; the top 15 bacteria were included in the analyses. Higher abundance of Ralstonia in infant gut was significantly correlated with infant weight gain from 1 to 8 weeks (r=0.38, p=0.017) and higher absolute infant weight at 8 weeks (r= 0.33 p=0.04). No significant correlation was identified between any of the 15 bacteria and maternal stress (Figure 4).
3.7. Unintended Effects
No unintended effects were reported by mothers included in this study.
4. Discussion
Using an experimental approach, we found that a simple relaxation intervention which reduced maternal stress and increased infant weight gain led to changes in the microbiome composition and diversity in the maternal gut, breast milk and infant gut. This is consistent with the hypothesis that the microbiome might be a mediator of the observed relaxation intervention effects via gut-brain and entero-mammary pathways. Key support for this finding comes from the differences in the microbial communities when comparing IG and CG of maternal gut, breast milk, and infant gut microbiomes.
Compared to baseline, the overall microbial community structures in maternal gut showed greater separation between IG and CG at 8 weeks, suggesting that the microbiome community structure was altered after the intervention. Nevertheless, the number of observed taxa in the maternal gut did not significantly differ between IG and CG and did not change significantly from 1-to 8 weeks, underlining that the microbiome richness in the adult gut may be relatively stable. Comparatively, the observed features and evenness of the microbiome in infant gut were significantly higher in IG than CG at 8 weeks. This may be regarded as beneficial, since studies have suggested that higher infant gut α-diversity reflects a more mature, adult-like community [23,24]. A study conducted in Mexican school-aged children showed a significant difference in microbiome composition among children in normal-weight, undernourished and obese groups (p < 0.01), with the normal-weight group showing greater α-diversity than undernourished and obese groups [25]. However, it should be mentioned that the interpretation of α-diversity is complex and can depend on various factors and research context. These finding merits further investigation with larger sample size.
As previously hypothesized, the changes in infant gut microbiome could be related to the maternal gut microbiome though breast feeding. However, whilst the α-diversity was significantly higher in IG infant feces, it was significantly lower in IG breast milk. On the one hand, a lower microbial diversity in breast milk could potentially be more consistent in its effects on infant health [26], since in a lower-diversity community, there might be stronger microbial competition. This competition could potentially inhibit the growth of harmful or pathogenic microorganisms, contributing to a healthier microbial balance in the infant’s gut [27]. On the other hand, whilst breastfeeding plays a key role in infant intestinal colonization [28], the infant gut microbiome does not share the composition and community structure seen in breast milk. Breast milk promotes a balanced microbiota development for the newborn, owing to its high content of unique oligosaccharides. These HMOs are the third most abundant solid component in breast milk after lactose and lipids, and can promote intestinal colonization in the infant gut [29,30]. Therefore, the content of HMOs in breast milk is an important factor in determining the microbiota diversity and composition in the infant gut. We did not measure the HMO content of breast milk in our trial. However, it is possible that HMOs were more abundant in the breast milk of IG mothers following the intervention, since previous evidence reported more abundant HMOs in mothers with good mental health compared to those who were distressed [31,32].
Bifidobacterium are predominant in the gut microbiota of infants, and they are considered to be important for infant health and development [8,33,34]. Bifidobacterium in breast milk has been reported to activate immunoglobulin A (IgA)-producing plasma cells in the neonatal gut [35] and could control inflammation through mucosal host-microbe crosstalk [36]. The present study showed a lower relative abundance of Bifidobacterium in IG breast milk at baseline but higher abundance after the relaxation intervention, suggesting the intervention may have increased the relative abundance of Bifidobacterium in IG breast milk. However, the increase in Bifidobacterium in infant gut following the intervention was not as obvious as that in breast milk. This finding is in agreement with studies showing that Bifidobacterium colonizes the infant gut rapidly within the first few months [11,34], although the CG infants showed a significantly lower baseline abundance than IG, it increased in both groups and no significant group difference was observed at 8 weeks. Overall, we suggest that the relaxation intervention contributed to an increase of Bifidobacterium in breast milk but had less impact on its colonization in infant gut.
Modulation of the Blautia genera is worth noting. Although its role in infant gut homeostasis remains less known, studies in adult patients undergoing allogeneic hematopoietic stem cell transplantation for leukemia highlight a positive association of Blautia with reduced rate of gut Graft versus Host Disease (GvHD) post-transplantation [37]. Future studies investigating potential crosstalk between beneficial bacteria that colonize the infant gut including Bifidobacterium, Lactobacillus and Blautia genera are warranted.
The published main outcomes from our trial included greater infant weight gain (mean difference in z-score=0.51, 95%CI: 0.2, 0.9), and lower maternal stress (mean difference in Perceived Stress Scale=2.7, 95%CI: 0.8, 4.5) in the IG at 8 weeks [18]. The correlation analysis in the present study further identified a significant association between higher abundance of infant gut Ralstonia and greater infant weight at 8 weeks. Ralstonia has commonly been observed in breast milk, whilst in the human gut, Ralstonia has mostly been reported as an opportunistic pathogen causing nosocomial infections in immunocompromised patients. In the present study, Ralstonia was observed rarely in the CG yet a significantly higher relative abundance of Ralstonia was observed in IG maternal and infant gut at both baseline and 8 weeks, with a positive correlation between Ralstonia and infant weight gain and the absolute weight value at 8 weeks. Again, these finding merits further investigation, since Ralstonia can be a common contaminant of DNA extraction kits or PCR reagents, which may lead to its erroneous appearance in microbiota or metagenomic datasets [38]. However, it is less plausible that only IG gut samples would show contamination, since all samples were coded before being sent for analysis and the research assistants were unable to distinguish between the groups.
Our study has several limitations. For reasons of privacy, most samples were collected by mothers without supervision, and although clear instructions were provided in advance, this might have led to contamination of the collected samples. Additionally, we did not collect samples from the mouth of mother or infant, maternal areolar skin or vagina, all of which could contribute additional bacteria to the infant gut microbiome. We also did not collect maternal dietary data which could have influenced microbiome composition. However, all mothers were randomly assigned into relaxation or control groups with no difference in baseline characteristics between groups; moreover, mothers were following the traditional postpartum confinement practice with similar diet and lifestyle, thus reducing concerns of bias. The analyses are somewhat limited by the relatively small sample size. However, the study sample was characterized by a high degree of homogeneity, as all mothers were primiparous Chinese women following vaginal delivery at 34-37 weeks, which increased the power to detect significant difference between IG and CG, and reduced potential bias. Furthermore, it should be noted that although we randomly assigned mothers with no significant differences observed in baseline demographics, the baseline β-diversity was significantly different between IG and CG mothers’ gut. Compared to baseline status, our results showed a stronger difference in gut microbiome diversity after the intervention, which may imply potential effects of the intervention, However, this should be treated with caution and needs to be further confirmed in larger trials.
This is the first study to test the hypothesis that the microbiome could act as a signal between mother and infant during breastfeeding using an experimental approach. By altering maternal psychological status using the relaxation intervention, we could evaluate the causal impact of maternal stress on maternal gut, breastmilk microbiome and subsequent consequences on infant gut microbiome and health. Consistent with our hypothesis, we found significant differences in microbiome composition and diversity between groups. Together with observed differences in the enrichment of specific genera and correlations between biomarkers and clinical outcomes, which are best considered as hypothesis-generating, these findings can inform the design of future studies, including larger trials in different populations, ideally with maternal dietary data and the collection of the additional biological samples mentioned above; and the application of metagenomic sequencing and metabolomics.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, CONSORT checklist; eFigure 1; eTable 1; eFigure 2; eFigure 3; eTable 2; eFigure 4; eFigure 5; eFigure 6.
Author Contributions
JY contributed to manuscript generating and statistical analysis; YZ contributed to microbiome analysis and preparation of the supplemental documents; MF contributed to revision of the manuscript, JY and ZW conducted the original trial, J.C.W, M.B.E, and D.S.N provided valuable comments and contributed to refining the manuscript. All authors read and approved the final manuscript.
Funding
The original research was conducted as part of a PhD and expenses were covered from the research funds held by Professor Fewtrell in The Childhood Nutrition Research Group. All research at Great Ormond Street Hospital NHS Foundation Trust and UCL Great Ormond Street Institute of Child Health is made possible by the NIHR Great Ormond Street Hospital Biomedical Research Centre. The views expressed are those of the author(s) and not necessarily those of the NHS, the NIHR or the Department of Health.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki, and approved by the Research Ethics Committee of University College London (ID: 12681/002) and the Department of Child Health, Beijing Children’s Hospital (ID: 2018-167). .
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
All raw sequencing data associated with this study have been uploaded to the Sequence Read Archive (SRA) under citation accession PRJNA1000236. Data described in the manuscript, code book, and analytic code will be made available upon request.
Acknowledgments
We would like to thank Novogene Technology Inc. (Beijing, China) for conducting the 16S rRNA sequencing. We also thank Xueling, Li, Yufeng An, Wenming, Cui, Changeshuan Yang and all research nurses from Beijing Children’s Hospital for assistance during data collection.
Conflicts of Interest
Professor Mary Fewtrell receives an unrestricted donation for research on infant nutrition from Philips (Amsterdam, NL). The remaining authors declare no other conflicts.
References
- Bailey, M.T.; Dowd, S.E.; Galley, J.D.; Hufnagle, A.R.; Allen, R.G.; Lyte, M. Exposure to a social stressor alters the structure of the intestinal microbiota: implications for stressor-induced immunomodulation. Brain Behav Immun 2011, 25, 397–407. [Google Scholar] [CrossRef]
- O’Mahony, S.M.; Marchesi, J.R.; Scully, P.; Codling, C.; Ceolho, A.M.; Quigley, E.M.; Cryan, J.F.; Dinan, T.G. Early life stress alters behavior, immunity, and microbiota in rats: implications for irritable bowel syndrome and psychiatric illnesses. Biol Psychiatry 2009, 65, 263–267. [Google Scholar] [CrossRef]
- Clarke, G.; Grenham, S.; Scully, P.; Fitzgerald, P.; Moloney, R.D.; Shanahan, F.; Dinan, T.G.; Cryan, J.F. The microbiome-gut-brain axis during early life regulates the hippocampal serotonergic system in a sex-dependent manner. Mol Psychiatry 2013, 18, 666–673. [Google Scholar] [CrossRef]
- Grenham, S.; Clarke, G.; Cryan, J.F.; Dinan, T.G. Brain-gut-microbe communication in health and disease. Front Physiol 2011, 2, 94. [Google Scholar] [CrossRef]
- Donnet-Hughes, A.; Perez, P.F.; Dore, J.; Leclerc, M.; Levenez, F.; Benyacoub, J.; Serrant, P.; Segura-Roggero, I.; Schiffrin, E.J. Potential role of the intestinal microbiota of the mother in neonatal immune education. Proc Nutr Soc 2010, 69, 407–415. [Google Scholar] [CrossRef]
- Fernandez, L.; Langa, S.; Martin, V.; Maldonado, A.; Jimenez, E.; Martin, R.; Rodriguez, J.M. The human milk microbiota: origin and potential roles in health and disease. Pharmacol Res 2013, 69, 1–10. [Google Scholar] [CrossRef]
- Perez, P.F.; Dore, J.; Leclerc, M.; Levenez, F.; Benyacoub, J.; Serrant, P.; Segura-Roggero, I.; Schiffrin, E.J.; Donnet-Hughes, A. Bacterial imprinting of the neonatal immune system: lessons from maternal cells? Pediatrics 2007, 119, e724–732. [Google Scholar] [CrossRef] [PubMed]
- Kapourchali, F.R.; Cresci, G.A. Early-Life gut microbiome—the importance of maternal and infant factors in its establishment. Nutrition in Clinical Practice 2020, 35, 386–405. [Google Scholar] [CrossRef] [PubMed]
- Pannaraj, P.S.; Li, F.; Cerini, C.; Bender, J.M.; Yang, S.; Rollie, A.; Adisetiyo, H.; Zabih, S.; Lincez, P.J.; Bittinger, K. Association between breast milk bacterial communities and establishment and development of the infant gut microbiome. JAMA pediatrics 2017, 171, 647–654. [Google Scholar] [CrossRef] [PubMed]
- Jost, T.; Lacroix, C.; Braegger, C.P.; Rochat, F.; Chassard, C. Vertical mother–neonate transfer of maternal gut bacteria via breastfeeding. Environmental microbiology 2014, 16, 2891–2904. [Google Scholar] [CrossRef]
- Nagpal, R.; Kurakawa, T.; Tsuji, H.; Takahashi, T.; Kawashima, K.; Nagata, S.; Nomoto, K.; Yamashiro, Y. Evolution of gut Bifidobacterium population in healthy Japanese infants over the first three years of life: a quantitative assessment. Scientific reports 2017, 7, 10097. [Google Scholar] [CrossRef] [PubMed]
- Galley, J.D.; Mashburn-Warren, L.; Blalock, L.C.; Lauber, C.L.; Carroll, J.E.; Ross, K.M.; Hobel, C.; Coussons-Read, M.; Schetter, C.D.; Gur, T.L. Maternal anxiety, depression and stress affects offspring gut microbiome diversity and bifidobacterial abundances. Brain, Behavior, and Immunity 2023, 107, 253–264. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Xu, J.; Ren, E.; Su, Y.; Zhu, W. Co-occurrence of early gut colonization in neonatal piglets with microbiota in the maternal and surrounding delivery environments. Anaerobe 2018, 49, 30–40. [Google Scholar] [CrossRef] [PubMed]
- Marques, T.M.; Wall, R.; Ross, R.P.; Fitzgerald, G.F.; Ryan, C.A.; Stanton, C. Programming infant gut microbiota: influence of dietary and environmental factors. Current opinion in biotechnology 2010, 21, 149–156. [Google Scholar] [CrossRef]
- Cong, X.; Xu, W.; Romisher, R.; Poveda, S.; Forte, S.; Starkweather, A.; Henderson, W.A. Focus: Microbiome: Gut microbiome and infant health: Brain-gut-microbiota axis and host genetic factors. The Yale journal of biology and medicine 2016, 89, 299. [Google Scholar]
- Tanaka, M.; Nakayama, J. Development of the gut microbiota in infancy and its impact on health in later life. Allergology International 2017, 66, 515–522. [Google Scholar] [CrossRef]
- Robertson, R.C.; Manges, A.R.; Finlay, B.B.; Prendergast, A.J. The human microbiome and child growth–first 1000 days and beyond. Trends in microbiology 2019, 27, 131–147. [Google Scholar] [CrossRef]
- Yu, J.; Wei, Z.; Wells, J.C.; Fewtrell, M. Effects of relaxation therapy on maternal psychological status and infant growth following late preterm and early term delivery: a randomized controlled trial. The American Journal of Clinical Nutrition 2022. [Google Scholar] [CrossRef]
- Dib, S.; Wells, J.C.; Eaton, S.; Fewtrell, M. A Breastfeeding Relaxation Intervention Promotes Growth in Late Preterm and Early Term Infants: Results from a Randomized Controlled Trial. Nutrients 2022, 14, 5041. [Google Scholar] [CrossRef]
- Mohd Shukri, N.H.; Wells, J.; Eaton, S.; Mukhtar, F.; Petelin, A.; Jenko-Pražnikar, Z.; Fewtrell, M. Randomized controlled trial investigating the effects of a breastfeeding relaxation intervention on maternal psychological state, breast milk outcomes, and infant behavior and growth. The American journal of clinical nutrition 2019, 110, 121–130. [Google Scholar] [CrossRef]
- Yu, J.; Wells, J.; Wei, Z.; Fewtrell, M. Effects of relaxation therapy on maternal psychological state, infant growth and gut microbiome: protocol for a randomised controlled trial investigating mother-infant signalling during lactation following late preterm and early term delivery. Int Breastfeed J 2019, 14, 50. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Cai, Y.; Dai, W.; Jiang, W.; Tang, W. The difference of gut microbiome in different biliary diseases in infant before operation and the changes after operation. BMC pediatrics 2022, 22, 502. [Google Scholar] [CrossRef] [PubMed]
- Bokulich, N.A.; Kaehler, B.D.; Rideout, J.R.; Dillon, M.; Bolyen, E.; Knight, R.; Huttley, G.A.; Gregory Caporaso, J. Optimizing taxonomic classification of marker-gene amplicon sequences with QIIME 2′s q2-feature-classifier plugin. Microbiome 2018, 6, 1–17. [Google Scholar] [CrossRef]
- Bäckhed, F.; Roswall, J.; Peng, Y.; Feng, Q.; Jia, H.; Kovatcheva-Datchary, P.; Li, Y.; Xia, Y.; Xie, H.; Zhong, H. Dynamics and stabilization of the human gut microbiome during the first year of life. Cell host & microbe 2015, 17, 690–703. [Google Scholar]
- Méndez-Salazar, E.O.; Ortiz-López, M.G.; Granados-Silvestre, M.d.l.Á.; Palacios-González, B.; Menjivar, M. Altered gut microbiota and compositional changes in Firmicutes and Proteobacteria in Mexican undernourished and obese children. Frontiers in microbiology 2018, 2494. [Google Scholar]
- Rajilić-Stojanović, M.; De Vos, W.M. The first 1000 cultured species of the human gastrointestinal microbiota. FEMS microbiology reviews 2014, 38, 996–1047. [Google Scholar] [CrossRef]
- LaTuga, M.S.; Stuebe, A.; Seed, P.C. A review of the source and function of microbiota in breast milk. In Proceedings of Seminars in reproductive medicine; pp. 068–073.
- Moloney, R.D.; Desbonnet, L.; Clarke, G.; Dinan, T.G.; Cryan, J.F. The microbiome: stress, health and disease. Mammalian Genome 2014, 25, 49–74. [Google Scholar] [CrossRef]
- Marcobal, A.; Barboza, M.; Froehlich, J.W.; Block, D.E.; German, J.B.; Lebrilla, C.B.; Mills, D.A. Consumption of human milk oligosaccharides by gut-related microbes. J Agric Food Chem 2010, 58, 5334–5340. [Google Scholar] [CrossRef]
- Vazquez, E.; Barranco, A.; Ramirez, M.; Gruart, A.; Delgado-Garcia, J.M.; Martinez-Lara, E.; Blanco, S.; Martin, M.J.; Castanys, E.; Buck, R. , et al. Effects of a human milk oligosaccharide, 2’-fucosyllactose, on hippocampal long-term potentiation and learning capabilities in rodents. J Nutr Biochem 2015, 26, 455–465. [Google Scholar] [CrossRef]
- Smilowitz, J.T.; O’sullivan, A.; Barile, D.; German, J.B.; Lönnerdal, B.; Slupsky, C.M. The human milk metabolome reveals diverse oligosaccharide profiles. The Journal of nutrition 2013, 143, 1709–1718. [Google Scholar] [CrossRef]
- Kortesniemi, M.; Slupsky, C.M.; Aatsinki, A.-K.; Sinkkonen, J.; Karlsson, L.; Linderborg, K.M.; Yang, B.; Karlsson, H.; Kailanto, H.-M. Human milk metabolome is associated with symptoms of maternal psychological distress and milk cortisol. Food Chemistry 2021, 356, 129628. [Google Scholar] [CrossRef]
- Mikami, K.; Kimura, M.; Takahashi, H. Influence of maternal bifidobacteria on the development of gut bifidobacteria in infants. Pharmaceuticals 2012, 5, 629–642. [Google Scholar] [CrossRef]
- Makino, H.; Kushiro, A.; Ishikawa, E.; Kubota, H.; Gawad, A.; Sakai, T.; Oishi, K.; Martin, R.; Ben-Amor, K.; Knol, J. Mother-to-infant transmission of intestinal bifidobacterial strains has an impact on the early development of vaginally delivered infant’s microbiota. PloS one 2013, 8, e78331. [Google Scholar] [CrossRef]
- Fernández, L.; Ruiz, L.; Jara, J.; Orgaz, B.; Rodríguez, J.M. Strategies for the preservation, restoration and modulation of the human milk microbiota. Implications for human milk banks and neonatal intensive care units. Frontiers in Microbiology 2018, 9, 2676. [Google Scholar] [CrossRef]
- LeBouder, E.; Rey-Nores, J.E.; Raby, A.-C.; Affolter, M.; Vidal, K.; Thornton, C.A.; Labéta, M.O. Modulation of neonatal microbial recognition: TLR-mediated innate immune responses are specifically and differentially modulated by human milk. The Journal of Immunology 2006, 176, 3742–3752. [Google Scholar] [CrossRef]
- Benítez-Páez, A.; Gómez del Pugar, E.M.; López-Almela, I.; Moya-Pérez, Á.; Codoñer-Franch, P.; Sanz, Y. Depletion of Blautia species in the microbiota of obese children relates to intestinal inflammation and metabolic phenotype worsening. Msystems 2020, 5. [Google Scholar] [CrossRef]
- Salter, S.J.; Cox, M.J.; Turek, E.M.; Calus, S.T.; Cookson, W.O.; Moffatt, M.F.; Turner, P.; Parkhill, J.; Loman, N.J.; Walker, A.W. Reagent and laboratory contamination can critically impact sequence-based microbiome analyses. BMC biology 2014, 12, 1–12. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
CONSORT Flowchart of the Randomized Controlled Trial. Notes: GA=gestational age, EBF=exclusive breastfeeding.
Figure 1.
CONSORT Flowchart of the Randomized Controlled Trial. Notes: GA=gestational age, EBF=exclusive breastfeeding.
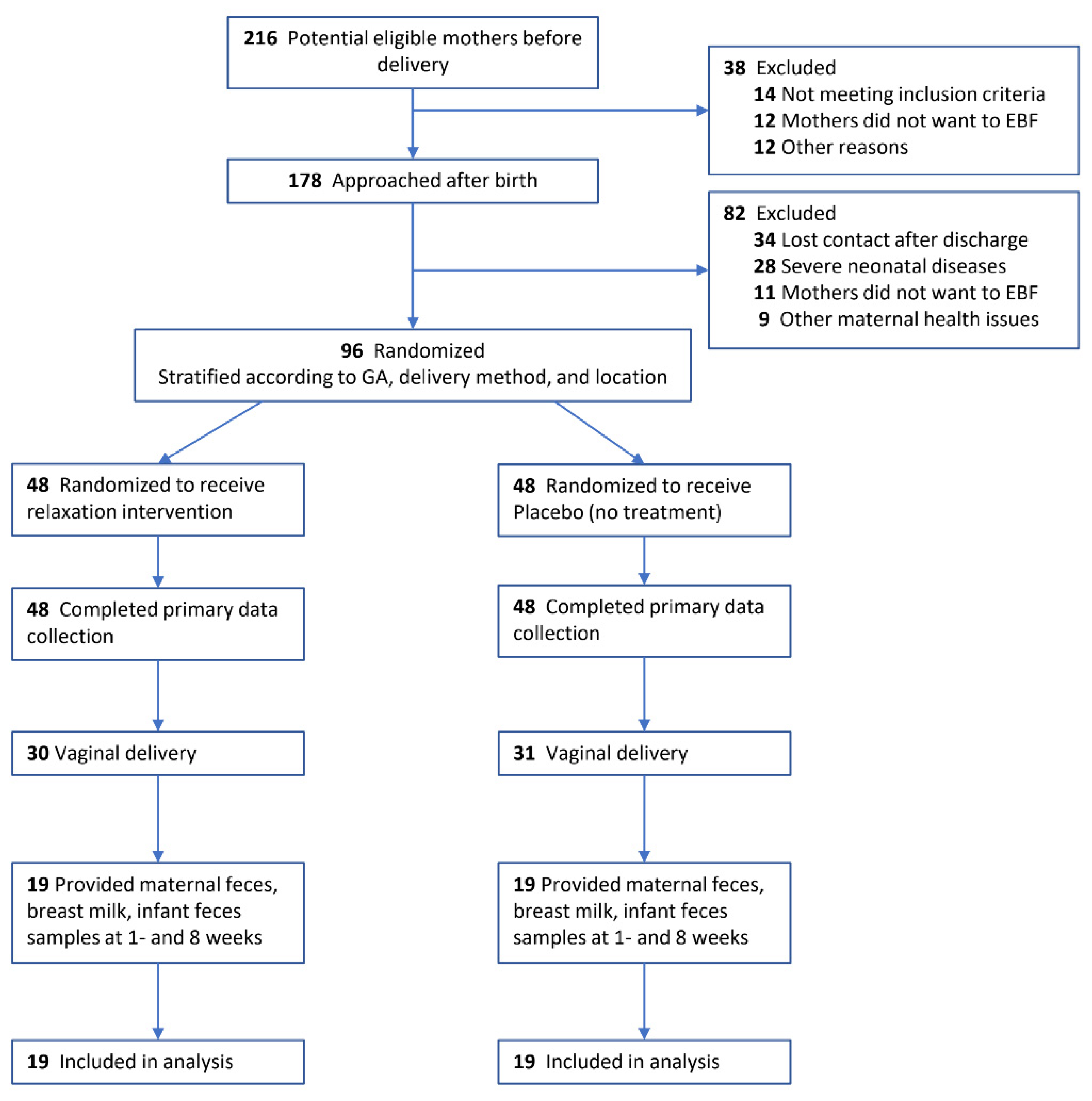
Figure 2.
Microbiome Diversity in Maternal Gut, Breast Milk, and Infant Gut. Notes: Gut microbiome diversity was assessed by analyzing microbiome composition in fecal samples and breast milk. Differences in maternal gut between intervention and control group at 1 and 8 weeks were shown in A (α-diversity, within sample diversity), B and C (β-diversity, between sample diversity); differences in breast milk were shown in, D(α-diversity), E and F (β-diversity); differences in infant gut were shown in G (α-diversity), H and I (β-diversity). Difference in α-diversity was assessed by Wilcoxon rank-sum test based on observed features. Differences in β-diversity was presented using principal coordinates analysis plot (PCoA) based on Bray-Curtis distance matrix.
Figure 2.
Microbiome Diversity in Maternal Gut, Breast Milk, and Infant Gut. Notes: Gut microbiome diversity was assessed by analyzing microbiome composition in fecal samples and breast milk. Differences in maternal gut between intervention and control group at 1 and 8 weeks were shown in A (α-diversity, within sample diversity), B and C (β-diversity, between sample diversity); differences in breast milk were shown in, D(α-diversity), E and F (β-diversity); differences in infant gut were shown in G (α-diversity), H and I (β-diversity). Difference in α-diversity was assessed by Wilcoxon rank-sum test based on observed features. Differences in β-diversity was presented using principal coordinates analysis plot (PCoA) based on Bray-Curtis distance matrix.
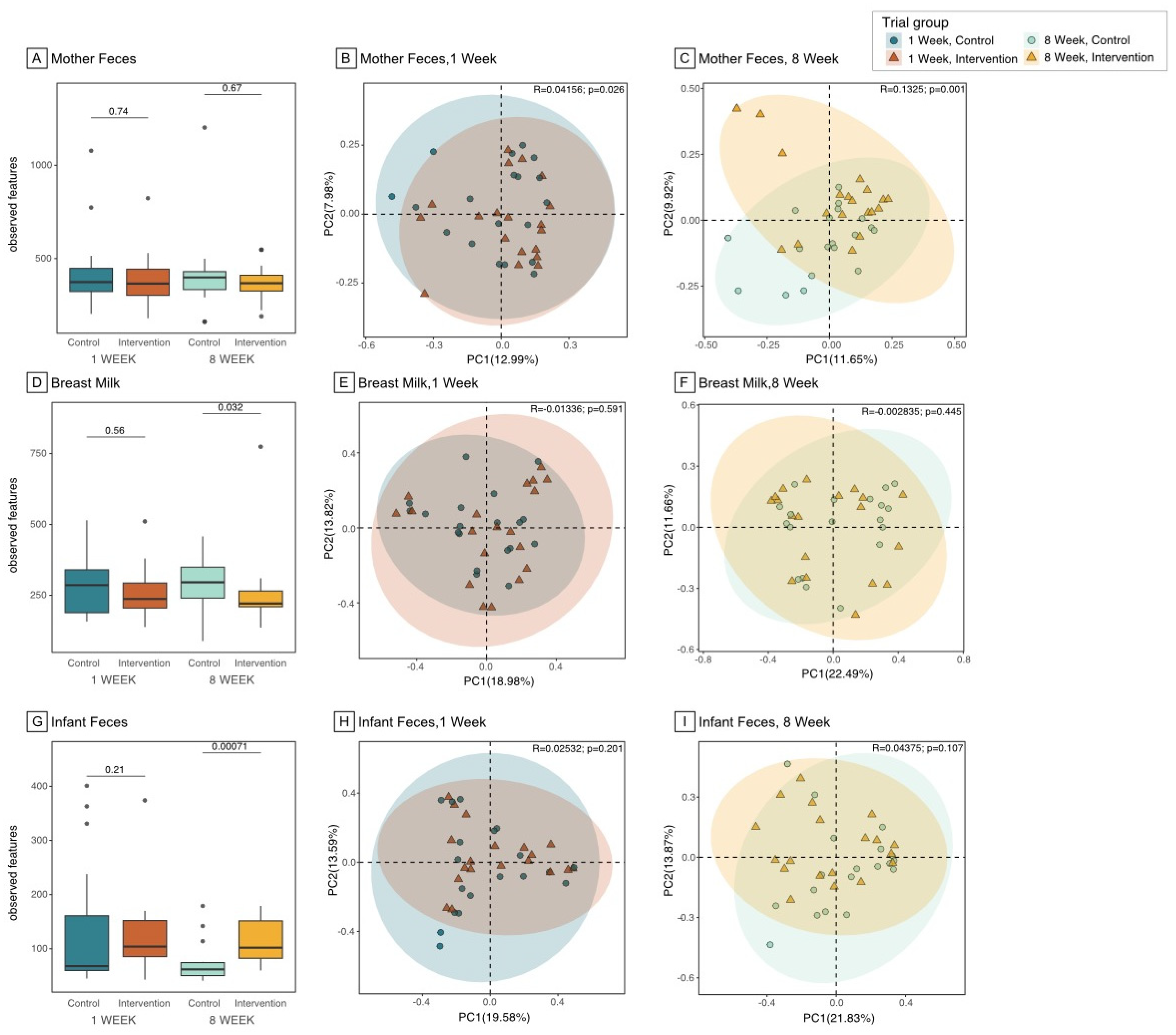
Figure 3.
Relative abundance of the Top 15 bacteria among Maternal Feces, Breast Milk, and Infant Feces Samples. Notes: Relative abundance of the top 15 most abundant genera in maternal feces, breast milk, and infant feces were examined and statistical differences between intervention and control group were compared using the Wilcoxon rank-sum test and FDR adjusted p-value.
Figure 3.
Relative abundance of the Top 15 bacteria among Maternal Feces, Breast Milk, and Infant Feces Samples. Notes: Relative abundance of the top 15 most abundant genera in maternal feces, breast milk, and infant feces were examined and statistical differences between intervention and control group were compared using the Wilcoxon rank-sum test and FDR adjusted p-value.
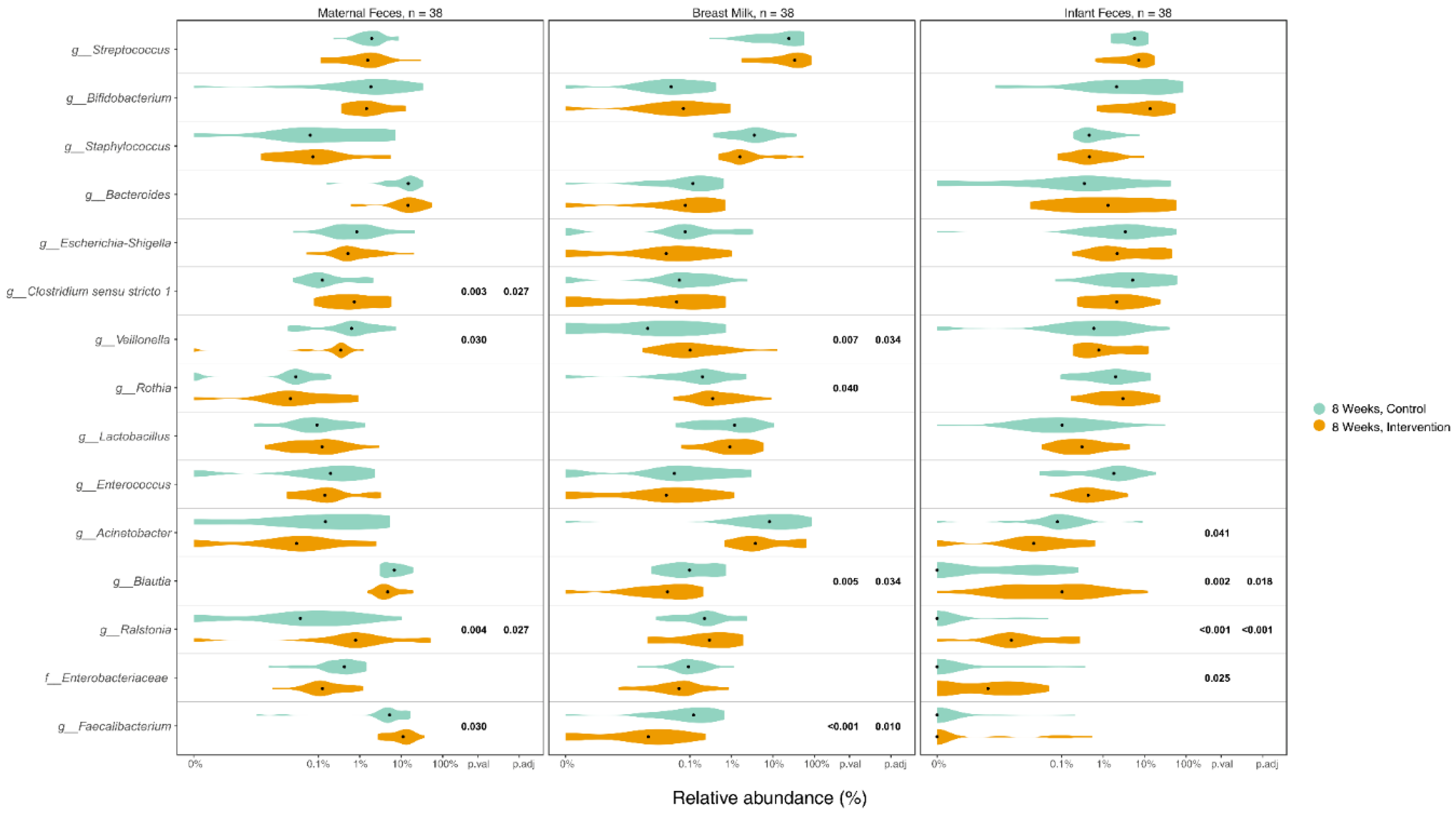
Figure 4.
Top 15 bacteria in infant feces samples and its correlation with infant weight at 8 weeks and weight gain from 1 to 8 weeks. Notes: wks= weeks, weight z-score was calculated based on 21st intergrowth study preterm newborn database.
Figure 4.
Top 15 bacteria in infant feces samples and its correlation with infant weight at 8 weeks and weight gain from 1 to 8 weeks. Notes: wks= weeks, weight z-score was calculated based on 21st intergrowth study preterm newborn database.


Table 1.
Baseline characteristics of the study participants.
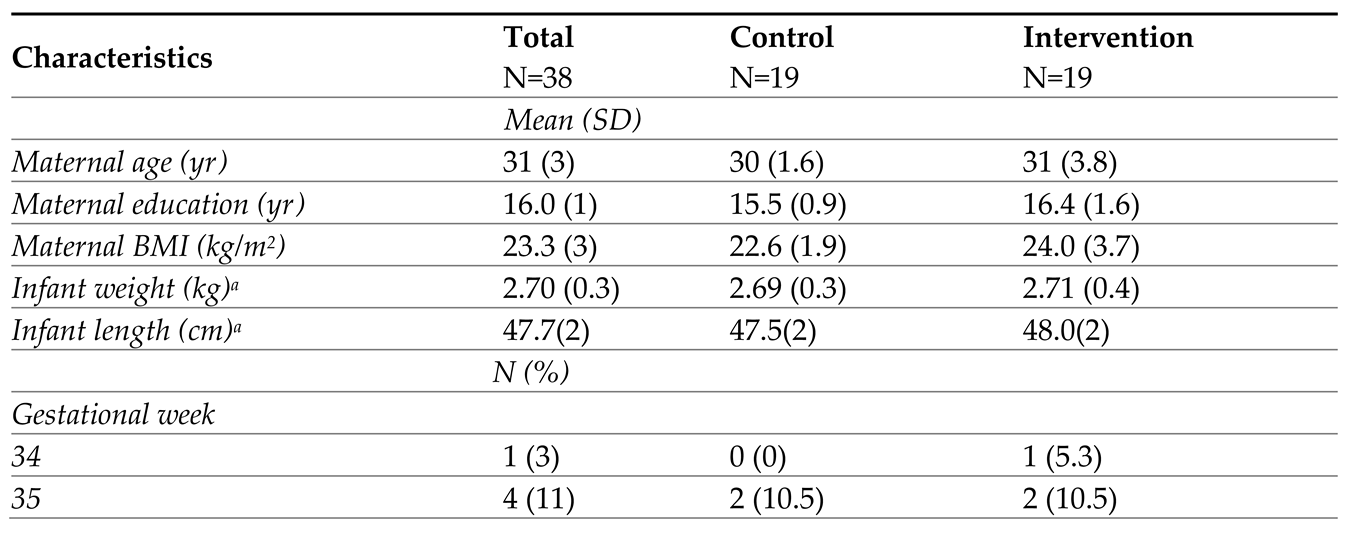 
|
Notes: SD=standard deviation. N=number. BMI=body mass index. EBF=exclusively breastfeed, self-reported by mothers with definition provided on the questionnaire. a Weight and length were measured using standard anthropometry assessment at 1-week home visit. b Infant could receive expressed breast milk or formula initially but had to be EBF at 1 week enrolment. C Apart from those infants who were exclusively breastfed at 8 weeks, the rest of infants were mostly breastfed (breast milk >70% of the feeding). d Three mothers took antibiotics during hospital stay due to vaginal incision. Two mothers in control group took Cephalosporins and Amoxicillin respectively; one mother in intervention group took Cephalosporins. All mothers reported no medicine intake during the study period.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



