
Submitted:
06 March 2024
Posted:
07 March 2024
You are already at the latest version
Abstract
Candidate vaccines against African swine fever virus (ASFV) based on naturally attenuated or genetically modified viruses have the potential to generate protective immune responses, although there is no consensus on what defines a protective immune response against ASFV. Studies, espe-cially in sensible host species, focused on unravelling protective mechanisms will contribute to the development of safer and more effective vaccines. The present study provides a detailed analysis of phenotypic and functional data on cellular responses induced by intradermal immunization and subsequent boosting of domestic pigs with the naturally attenuated field strain Lv17/WB/Rie1, as well as the mechanisms underlying protection against intramuscular challenge with the virulent genotype II Armenia/07 strain. The transient mild to moderate increase of IL-8 and IL-10 in serum observed after immunization might be directly correlated with survival. Protection was also associated with a robust ASFV-specific polyfunctional memory T cell response, where CD4CD8 and CD8 T cells were identified as the main cellular sources of virus-specific IFNγ and TNFα. In parallel to cytokine response, these T-cell subsets also showed specific cytotoxic activity as evidenced by the increased expression of the CD107a degranulation marker. Along with virus-specific multifunctional CD4CD8 and CD8 T responses, the increased levels of antigen experienced cytotoxic CD4 T cells observed after challenge in immunized pigs might also contribute to control virulent infection by killing mechanisms targeting infected antigen-presenting cells. Future studies should elucidate whether the memory T-cell responses evidenced in the present study persist and provide long-term protection against further ASFV infections.
Keywords:
African swine fever
; live-attenuated virus
; vaccine
; protective cellular response
; T-cell
; cytokines
; domestic pigs
1. Introduction
African swine fever (ASF) is currently the main problem facing the pig industry worldwide [1]. ASF is a devastating haemorrhagic infectious disease caused by African swine fever virus (ASFV), a large and complex enveloped double-stranded DNA virus of the genus Asfirvirus (family Asfarviridae). The disease was endemic in most sub-Saharan African countries and in the island of Sardinia (Italy) until 2007, when highly virulent isolates belonging to genotype II appeared in Eastern Europe. Since then, ASF has become endemic in many European and Asian countries spreading without control into Eastern Europe, China (since 2018) and most Southeast Asian countries, causing a huge economic impact. Outbreaks of uncertain origin have also occurred since 2021 in some Caribbean countries (Dominican Republic and Haiti), posing a threat to the nearby North American pork industry [2]. The disease, which affects domestic and wild suids of all breeds and ages, presents a variable lethality depending on the virulence of the isolate and the immune status of the infected animals [3,4,5]. Thus, highly virulent isolates often have a lethality close to 100% in naïve animals, which usually die within two weeks of infection [6,7].
Vaccines are essential to control viral diseases. The lack of vaccines against ASFV can largely be attributed to gaps in knowledge of the strategies used by the virus to evade host innate and adaptive immunity and the functions of virus proteins responsible for inducing protective immune responses [8]. Evidence suggests the importance of both arms of adaptive immunity for protection. However, the immunological correlates of protection are not yet understood. On the one hand, antibodies have been suggested to be an essential component of protective immunity against virulent ASFV, although alone they are not sufficient to induce protection [9]. Conversely, evidence suggests that cellular immune response, even in the absence of specific antibodies against ASFV, may be necessary for effective protection, highlighting the role of CD8+ T lymphocytes [10,11,12,13,14,15,16].
Vaccines against ASFV based on naturally attenuated live virus (LAV) induce robust immune protection, stimulating both innate and adaptive (cellular and humoral) immunity. LAV immunizations, although characterized by an absence or mild presence of both clinical signs and viremia levels, have traditionally raised safety concerns, making their commercialization unlikely [8,17,18]. Selective deletion of genes involved in virus attenuation and/or induction of protection has also been used as a strategy to produce safe and effective live attenuated vaccines and to differentiate between vaccinated and infected animals (DIVA vaccines). In both cases, viruses attenuated naturally or by gene deletion usually render recovered animals protected from subsequent infections with related viruses, although this does not usually guarantee protection against divergent viruses [18,19,20,21]. Previous studies have demonstrated that immunization of domestic pigs (DP) and wild boar with a naturally attenuated non-haemadsorbing (non-HAD) genotype II ASFV isolate, obtained from a wild boar hunted in Latvia in 2017 (Lv17/WB/Rie1 strain), conferred high levels of protection against challenges with a virulent ASFV genotype II isolate (Armenia/07) [22,23,24]. It is known that immunizations with this strain induce some mild clinical signs and transient viremia. To improve its safety, the deletion of virulence-associated genes has recently been carried out. Although some deletion mutants generated showed a slight reduction in pathogenicity and lethality during the "in vivo" evaluations in DP, did not show a significant reduction in side effects with respect to the parental virus [25,26]. These results highlight the potential use of Lv17/WB/Rie1 strain not just as a vaccine prototype, but also as an excellent platform to elucidate protective mechanisms in immunized animals with especial attention to the role of the cellular immune response. With the objective of characterizing the immunological mechanisms of protection against ASFV, and with special attention to the role played by different subsets of T lymphocytes, we have systematically analyzed these mechanisms in DP immunized with the genotype II Lv17/WB/Rie1 strain and that were protected against virulent challenge with Armenia/07 (Arm07). In addition, several immunomodulatory cytokines were studied to complete the understanding of the protective mechanisms elicited by this vaccine prototype.
2. Materials and Methods
2.1. Viruses and Cells
The naturally attenuated, non-haemadsorbing (non-HAD) genotype II ASFV strain Lv17/WB/Rie1 was used for the immunization of domestic pigs. The haemadsorbing (HAD) ASFV strain Armenia/07 (Arm07), a virulent genotype II strain [27], was used to challenge the animals. Lv17/WB/Rie1, isolated for the first time from the serum of a wild boar hunted in Latvia in 2017, was previously described and tested in experimental trials in domestic pigs [24] and wild boar [22]. For the “in vivo” immunization/challenge studies, Lv17/WB/Rie1 was propagated in porcine alveolar macrophages (PAM) cultures, while Arm07 was propagated and titrated in porcine blood leukocytes (PBL) as described previously [28]. Titers of virus were defined as the amount of virus causing HAD (for the HAD strain) or infection, assessed by immunoperoxidase (IP) staining (for non-HAD strain), in 50% of infected cell cultures (HAD50/mL or TCID50/mL, respectively). For the “ex vivo" stimulation assays, Lv17/WB/Rie1 and Arm07 were propagated in PAM, stocks were titrated in 48-well plates using the African green monkey fibroblast-like cell line (Cos-7 cells cells), and viral titers (TCID50/mL) were estimated by IP staining. Mock virus supernatant was prepared from uninfected PAM and stock was titrated in the same way as virus stocks.
Cos-7 cells culture was originally obtained from the American Type Culture Collection (ATCC CRL-1651) and it was grown in Dulbecco’s modified Eagle Medium (Corning, Manassas, USA) supplemented with 2mM L-glutamine, 100U of gentamicin per ml (Sigma-Aldrich, Saint Louis, MO, USA), 1% Na Pyruvate (Thermo Scientific, Waltham, Massachusetts, USA) and 1% non-essential amino acids (Thermo Scientific, Waltham, Massachusetts, USA). Cells were cultured at 37°C in medium supplemented with heat inactivated (HI) fetal bovine serum (FBS) (Sigma-Aldrich, Saint Louis, MO, USA).
The MS-stable monkey kidney cell line (ECACC, 91070510) was used for the preparation of the ASFV MS-adapted E70 isolate (E70 MS 48); coated 96-well plates. These plates were then used as the antigen in the indirect IP test (IPT) [29].
2.2. Animals and Experimental Design
The “in vivo” experiments were conducted under biosafety level 3 (BSL3) conditions at the animal facilities of Centro de Investigación en Sanidad Animal (CISA-INIA-CSIC), in accordance with EC Directive 2010/63/UE and approved by the Spanish Ethical and Animal Welfare Committee (Ref nº PROEX/101/8.21). Nine ASFV-free and ASFV antibody-free 8-week-old European hybrid female were randomly allocated to two groups. Animals in one group (n=5; pig 6, 7, 8, 9 and 10) were immunized on day 0 (prime) by intradermal (ID) inoculation using the IDAL 3G+ needle-less vaccinator (IDAL®, MSD Animal Health) with 1 mL containing 102 TCID50 of the attenuated ASFV Lv17/WB/Rie1 strain. These animals received identical immunization on day 21 (boost). Animals in the other group (n=4) remained as non-immunized controls. 35 days after the first immunization, pigs were challenged by intramuscular (IM) route with 1 mL containing 10 HAD50 of the virulent Arm07 strain. In the control group, non-immunized animals were challenged in parallel using the same route and dose.
2.3. Sampling, Clinical and Post-Mortem Analysis
For this study, blood samples were taken from all pigs on a weekly basis. Serum was collected in BD SST vacutainer tubes (Fisher Scientific). Anticoagulated blood was collected in heparin tubes for peripheral blood mononuclear cells (PBMC) isolation and in EDTA for virological studies (BD vacutainer tubes, Fisher Scientific). Clinical signs were recorded daily post-immunization (dpi) and post-challenge (dpc) and expressed with a quantitative clinical score by summing the values of eight clinical signs as previously described [30,31]. The parameters of fever, anorexia, recumbence, skin hemorrhage or cyanosis, joint swelling, respiratory distress, ocular discharge, and digestive findings were assigned points on a severity scale of 0 to 3, with one being mild, two being moderate, and three the most serious. The sum of the points was recorded as the clinical score (CS), which was also used to define humane endpoints. Pre-determined humane endpoints included pigs displaying severe signs of temperature, anorexia, recumbence, respiratory distress and digestive findings for more than two consecutive days, or a total CS in a pig of > 18. Pigs in the immunized group were euthanized on day 65 after prime immunization (30 days after challenge) for tissue collection. Non-immunized pigs were culled from 7 dpc after reaching a specified humane endpoint. Macroscopic lesions were evaluated during necropsies, using scores based on previous standardized protocols [32]. Twenty-one different types of tissues and organs were obtained from each necropsied animal and frozen at -80°C for ASFV genome detection.
2.4. Samples Processing
Serum tubes were spun at 1300xg for 10 minutes at room temperature, aliquoted and stored at -80°C. PBMC were isolated by density gradient centrifugation as described previously [33]. Briefly, heparinized blood was diluted in phosphate-buffered saline (PBS) and poured into Histopaque®-1077 pre-filled LeucosepTM tubes. After centrifugation, PBMC were aspirated from the interphase above the separation medium, washed with PBS before red blood cells (RBC) lysis and washed again with PBS before resuspending in complete RPMI 1640 medium (cRPMI) supplemented with 10% HI FBS and antibiotics. PBMC were used immediately at the cell density required for the different immunological assays or cryopreserved in liquid nitrogen in 10% dimethyl sulphoxide (DMSO) in HI FBS for subsequent analysis. Organ and tissue samples were homogenized using stablished protocols [4,34]. Briefly, 10% (w/v) clarified homogenized tissue suspensions were prepared in PBS. Supernatants were filter with MINISART filters 0.45µm and then treated with 0.1 % of gentamicin sulphate 50mg/ml (BioWhittaker) during 1h at 4±3ºC prior to use for virus detection.
2.5. Assessment of the Presence of Virus in Blood and Tissues
DNA was isolated from EDTA-blood and homogenized tissue samples using the High Pure PCR Template Preparation kit (Roche Diagnostics GmbH, Roche Applied Science, Mannheim, Germany) and the Universal Probe Library (UPL) real-time PCR (PCR) [34,35] was carried out for each sample. ASFV-positives samples were those with a Ct < 40. Virus isolation (VI) was performed on PCR positive blood and tissues samples using PBL in 96-well plates as described in the WOAH Manual [34]. The plates were examined over a period of seven days to evidence the presence of HAD or cytopathic effect (CPE). In absence of HAD, the samples were blind passage three times and subjected to the UPL PCR [34] to verify the replication of the non-HAD Lv17/WB/Rie1 ASFV strain. Titers were estimated by end-point dilution as described previously in section 2.1.
2.6. Assessment of ASFV-Specific Antibodies and Cytokines in Serum
Sera were assayed for ASFV-specific antibodies (Ab) using a commercial ELISA kit (Ingenasa-Ingezim PPA Compac K3; Gold Standard Diagnostic, Madrid, Spain). Antibody titers were determined by end-point dilution using the IPT [29,34] and expressed as the reciprocal of the highest dilution showing a positive result. Commercial ELISA kits were used to analyze serum concentrations of different cytokines with important immunoregulatory functions according to the manufacturer´s instructions: TNFα, IFNγ, IL-8, IL-10, IL-4, IL-12 and IL-6 (R&D Systems, Abingdon, UK) and IFNα (Millipore-Sigma, Saint Louis, MO, USA). For cytokine analysis, serum samples collected at the same time points during the course of the study from immunized pigs and non-immunized control pigs were used to compare the different cytokine profiles. Only serum samples collected before challenge from two of the four pigs included in the control group were analyzed in the cytokine ELISA assays, while serum samples collected after challenge from all animals in this group were included in the assay.
2.7. Assessment of ASFV-Specific T Cell Cytokine Responses in Blood
2.7.1. IFNγ ELISpot Assay
A broad assessment of virus specific T cell responses was studied by longitudinal measurement of IFN-γ secretion using a porcine ELISpot assay [33,36,37]. IFN-γ responses to antigenic stimulation were assessed after “in vitro” stimulation of PBMC at 0, 7, 21, 28, 35, 42, 49 and 56 dpi either with Lv17/WB/Rie1 or Arm07 at a multiplicity of infection (MOI) of 0.1. Mock virus-infected PAM cell supernatant used in triplicated wells and 10 µg/mL concanavalin A (ConA) (Sigma-Aldrich) in one well were used as negative and positive controls, respectively. Briefly, multiscreen 96-well ELISpot plates (Millipore, Watford, UK) were coated with 1µg/mL anti-porcine IFN-γ capture antibody (BD Biosciences, Oxford, UK) diluted in PBS. After blocking the plates with cRPMI, 2.5 × 105 cells were tested in triplicated wells with an MOI of 0.1 for each virus/mock and plates were incubated overnight at 37°C in 5% CO2 humidified atmosphere. After 18 hours, cells were lysed with cold water and wells were washed with PBS/0.05% tween20 to remove cell debris. Plates were incubated with 0.17 µg/mL biotinylated anti-porcine IFN-γ antibody (BD Biosciences) and to visualize the reaction, streptavidin-alkaline phosphatase enzyme conjugate (R&D Systems, Abingdon, UK) and the BCIP/NBT substrate (R&D Systems) were used. Finally, reaction was stopped with distilled water, plates were washed extensively on both sides and allowed to dry protected from light. Spots were visualized and counted using an automated ELISpot reader (AID AutoImmun Diagnostika, Straβberg, Germany). Results were expressed as the mock-corrected number of IFN-γ secreting cells or spots forming units (SFU) per million PBMC.
2.7.2. Flow Cytometric Analysis
In order to characterize the phenotype and functions of virus-specific T cells a flow cytometry analysis was performed [33,36,37,38]. Cryopreserved PBMC from selected days (0, 7, 21, 28, 35, 42, 49 and 56 dpi) were recovered from liquid nitrogen storage and resuscitated in pre-warmed cRPMI. After washing, PBMC were resuspended in cRPMI at the appropriated cell density and 5 × 105 cells were seeded per well in a 96-well round bottom plate (Nunc, Thermo Scientific). Cells were stimulated in triplicate with 0.1 MOI of virus using either Lv17/WB/Rie1 or Arm07. Medium only or mock virus supernatant were used as negative controls. In the positive control wells, a cell activation cocktail containing phorbol 12-myristate-13-acetate and ionomycin (PMA/Ionomycin) (BioLegend, San Diego, CA, USA) was used. After 14-16 hours at 37°C, Brefeldin A (GolgiPlug, BD Biosciences) was added to every well and cells were incubated for another 6 hours at 37°C. In defined experiments, CD107a-Alexa Fluor 647 monoclonal Ab (mAb) (Bio-Rad, Kidlington, UK), in conjunction with Brefeldin A (GolgiPlug) and Monensin (GolgiStop) (BD Biosciences), were used to study cytotoxic degranulation. After 6 hours incubation, prior to intracellular cytokine staining (ICS), cells were surface stained with Zombie NIR fixable viability dye (Biolegend) to distinguish live from dead cells and with the mAbs CD3ε-FITC, CD4α-PerCP-Cy5.5 and CD8α-PE (BD Bioscience). At selected time points and experiments, mouse anti-pig swine leukocyte antigen (SLA) class II DR mAb (Bio-Rad) was added to study cell activation. Following surface staining, cells were fixed (Fixation buffer, Biolegend) and permeabilized (Permeabilization wash buffer, Biolegend), and subsequently labelled with IFN-γ- Alexa Fluor 647 (Bio-Rad) and TNFα-Brilliant Violet 421 (Biolegend) to detect both intracellular cytokines. Flow cytometric data were analyzed using a BD FACSCelesta Cell Analyzer (BD Biosciences). This analysis was carried out by selection of lymphocytes, followed by selection of singlet and live events. Then CD3+CD4+CD8int/low (CD4+CD8+ T cells), CD3+CD4negCD8high (CD8+ T cells) and CD3+CD4+CD8neg (CD4+ T cells) cell subpopulations were gated and their frequencies relative to the live cells analyzed. Moreover, frequencies of cells positives to IFNγ, CD107a or SLA class II (SLA-II), as well as co-expression of TNFα, were analyzed for all those subpopulation. Mean frequencies of the different cell populations in the mock-stimulated wells were subtracted from each experimental value of each animal. Likewise, mean % of IFNγ+ or IFNγ+TNFα+ secreting cells, mean % of CD107a+ or CD107a+TNFα+ cells, as well as mean % of SLA-II+ or SLA-II+ TNFα+ cells in the mock-stimulated wells were subtracted from each experimental value of each animal in order to analyze the virus-specific cellular responses. The numbers of singlet live lymphocytes acquired for analysis ranged from 30,000 to 50,000 events.
2.8. Statistical Data Analysis
For graphical and statistical analysis of data, the software GraphPad Prism v8.0.1 (GraphPad Software, San Diego, CA, USA) was used. Flow cytometry data were analyzed using FlowJo v10 (BD). In order to compare virus-specific cellular responses from 0 dpi onwards in the immunized group and between groups and/or virus stimulus at different time points, a two-way analysis of variance (ANOVA) or a mixed-effects model were used. Antibody titer data were log transformed before analysis. P-values < 0.05 were considered statistically significant.
3. Results
3.1. Evaluation of Immunization Efficacy
3.1.1. Clinical Signs and Viraemia
All the pigs in the immunized group were successfully immunized with Lv17/WB/Rie1. Except for one pig (pig 7), the animals showed none or mild clinical signs, and remained healthy across the entire observation period. After the prime immunization, the pig 7 developed moderated clinical signs, showing high temperature (41-41.6oC) from 10 to 14 dpi, which coincided with moderate levels of viraemia (Ct values from 26.3 to 25.6). Pig 7 was euthanized at 14 dpi because it reached a mean CS of 12, and consequently, it was excluded from further immunological evaluations. The remaining immunized animals exhibited mild clinical signs (mean CS 2.4 ± 0.9) with low and sporadic viraemia (Ct values above 34) between 7 and 21 dpi (Figure 1A). Excluding pig 7, virus was solely isolated from blood samples obtained from pig 10 at 10 and 14 dpi, with an average viral titer of 9.8 x 102 TCID50. Animals were protected against virulent challenge with Arm07, without apparent clinical signs and with barely detectable virus levels in the blood (Ct values above 37.3) at 42 dpi (7 dpc). No infectious virus was recovered by virus isolation from blood of any of the immunized pigs after the challenge. Non-immunized control pigs became severely ill and died or were humanely culled from 7 dpc, showing clinical signs and viraemia levels characteristic of acute ASF.
3.1.2. Pathological Finding and Presence of Virus Genome in Tissues
Non-immunized control pigs displayed characteristic ASF lesions after challenge. In contrast, no such lesions were observed in immunized animals during necropsies carried out at the end of the study (65 dpi/30 dpc). The detection of the ASFV genome by PCR in tissue samples taken during necropsies of immunized pigs (30 dpc) is shown in Table 1. The ASFV genome was found in 6/21 tissues (28.6%) in the pig 6, 1/21 (4.8%) in the pig 8, 5/21 (23.8%) in the pig 9 and 14/21 (66,6%) in the pig 10. Lv17/WB/Rie1 was isolated from some tissues (lymph nodes and articular cartilage) obtained from two immunized pigs (pig 6 and pig 10), whereas Arm07 was isolated from the palatine tonsil in one of the immunized pigs (pig 10) (Table 1).
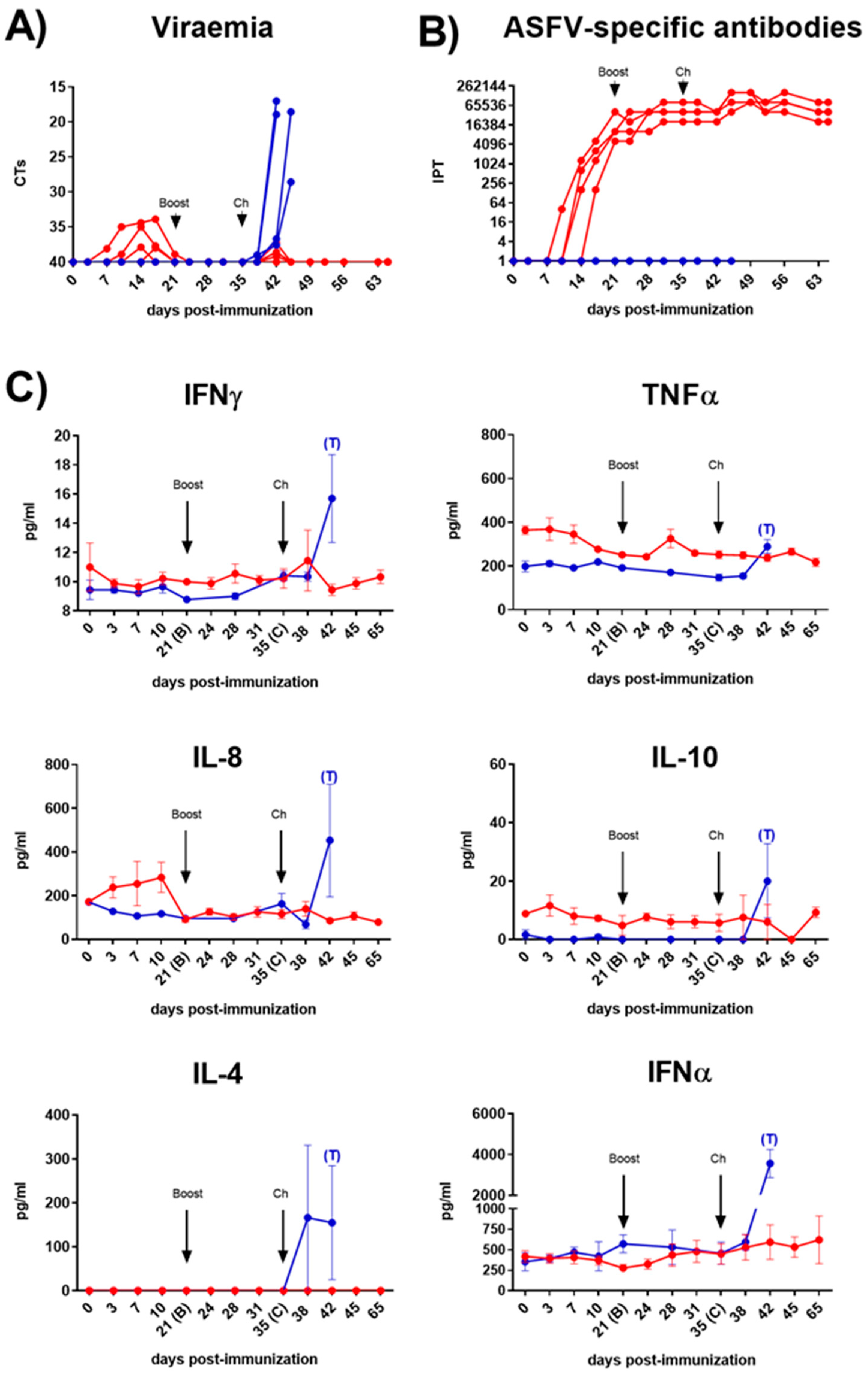
Figure 1.
Viraemia (A), ASFV-specific antibody response (B) and cytokines in serum samples (C) following immunization of pigs on day 0 (prime) and 21 (boost) dpi with Lv17/WB/Rie1 and challenge at 35 dpi with Armenia07. Immunized pigs (red line) and non-immunized pigs (blue line). Values at each time point for each individual pig are shown (A and B). Mean data ± SEM are shown for each group (C).
Figure 1.
Viraemia (A), ASFV-specific antibody response (B) and cytokines in serum samples (C) following immunization of pigs on day 0 (prime) and 21 (boost) dpi with Lv17/WB/Rie1 and challenge at 35 dpi with Armenia07. Immunized pigs (red line) and non-immunized pigs (blue line). Values at each time point for each individual pig are shown (A and B). Mean data ± SEM are shown for each group (C).
3.2. Evaluation of ASFV-Specific Antibodies and Cytokine Profiles in Serum
All pigs seroconverted from 12±1.6 dpi, showing a robust response from 21 dpi, which was maintained until the end of the study (Figure 1B). Serum levels of key immunoregulatory cytokines were monitored throughout the study (Figure 1C). Immunization with Lv17/WB/Rie1 did not result in a significant increase in serum IFNγ levels. However, although barely perceptible, we observed small increases of this cytokine in some of the immunized animals, displaying a first peak 7 days after booster immunization (28 dpi) and a second and more marked increase 3 days after the challenge (38 dpi). TNFα levels exhibited a gradual decrease following primary immunization, achieving significant differences regarding pre-immunization values between 10 and 24 dpi (p<0.01). At 28 dpi, seven days after the booster immunization, a non-significant but punctual increase of TNFα was detected. Other important pro-inflammatory cytokine, such as IFNα, did not show significant changes at any time in the immunized group. However, between 10 and 24 dpi, in parallel with the TNFα, a slight decrease of IFNα was also observed. Conversely, a remarkable, albeit not statistically significant, increase in serum levels of IL-8 (also known as CXCL-8) was observed between 3 and 10 dpi. IL-10 levels remained generally low throughout the study, with a slight, non-significant peak at 3 dpi, while IL-4 was undetectable at any time point. Finally, other important cytokines, such as IL-12 or IL-6 (data not shown), did not exhibit appreciable changes in the immunized animals. By contrast, following the challenge, serum levels of all studied cytokines increased in the non-immunized control pigs, displaying some significant differences with the immunized animals, particularly notable for IFNγ and IFNα (p<0.001), and slightly lower for IL-8 (p<0.05).
3.3. Evaluation of ASFV-Induced Protective Cellular Responses after Immunization with the Lv17/WB/Rie1 Strain and Homologous Virulent Challenge with Armenia/07
3.3.1. Analysis of Virus-Specific IFNγ Responses by IFNγ T Cell ELISpot
Virus-specific T cell response was quantified throughout the experiment using an IFNγ ELISpot assay after “ex vivo” stimulation of PBMC using either Lv17/WB/Rie1 or Arm07 (Figure 2A). The immunization of pigs with Lv17/WB/Rie1 induced a strong virus-specific IFNγ T cell response, which was observed following the prime immunization, with significant increases (p<0.001) of the responding cells against both viruses (Lv17/WB/Rie1 and Arm07) at 21 dpi. Both responses were even greater after the boost, displaying the maximum peaks after one week, at 28 dpi (p<0.0001), showing the highest response after stimulating with Lv17/WB/Rie1. A significant drop in the responses was observed just before the challenge (35 dpi), although both responses increased again after the challenge (from 42 dpi onward), showing a similar trend between both viral stimuli. Increases in the number of IFNγ responding cells were significant from 42 dpi [6] and highly significant (p<0.0001) from 49 dpi. On the contrary, no virus specific T cells responses were observed in the non-immunized control pigs at any time during the study.
3.3.2. Analysis of Primary Cellular Sources of ASFV-Specific IFNγ and TNFα by Flow Cytometry
In parallel, we used flow cytometry to identify the cellular source of virus specific IFNγ responses (Figure 2B-E). The flow cytometry gating strategy defining the phenotype of the different cell populations analysed is described in the Supplementary Figure S1. CD8 T cells (Figure 2B and D, Figure 5A and D) and CD4CD8 T cells (Figure 2C and E, Figure 5B and E), defined by the CD3+CD4negCD8high cytotoxic phenotype and by the CD3+CD4+CD8low activated/memory phenotype respectively, were identified as the main source of ASFV-specific IFNγ+ and IFNγ+TNFα+ after immunization with the Lv17/WB/Rie1 strain. In the immunized pigs, significant ASFV-specific cytokine responses (IFNγTNFα) from T cells CD8 (Figure 2B, Figure 5D) and CD4CD8 (Figure 2C, Figure 5E) were observed following “ex vivo” stimulation of PBMC with both viruses. Both responses were evident from 21 dpi; although they were not significant (compared with 0 dpi) until day 28. Generally, the CD4CD8 specific response observed was greater (in terms of percentages of cytokine-secreting cells) than the CD8 specific response. Similarly to what was observed with the IFNγ ELISpot, the highest cytokine responses were observed at 28 dpi, seven days after the boost, displaying both responses (CD8 and CD4CD8), highly significant increases (p<0.0001) against both viral stimuli, Lv17/WB/Rie1 and Arm07. Nevertheless, just before the challenge at 35 dpi, both responses dropped significantly, although these increased again following the challenge with Arm07. These increases, although without statistical significance, were gradual and maintained against both virus stimuli until the end of study, being particularly evident in the case of the CD4CD8 T cell response at 56 dpi. With regard to CD8 responses, these increased after challenge peaking at 49 dpi (14 dpc) against both stimuli, although only achieved statistical significance (p<0.01) upon stimulation with Lv17/WB/Rie1. Virus-specific responses for the CD3+CD4+CD8neg helper T cell phenotype (CD4 T cells) were absent (Figure 5 C and F).
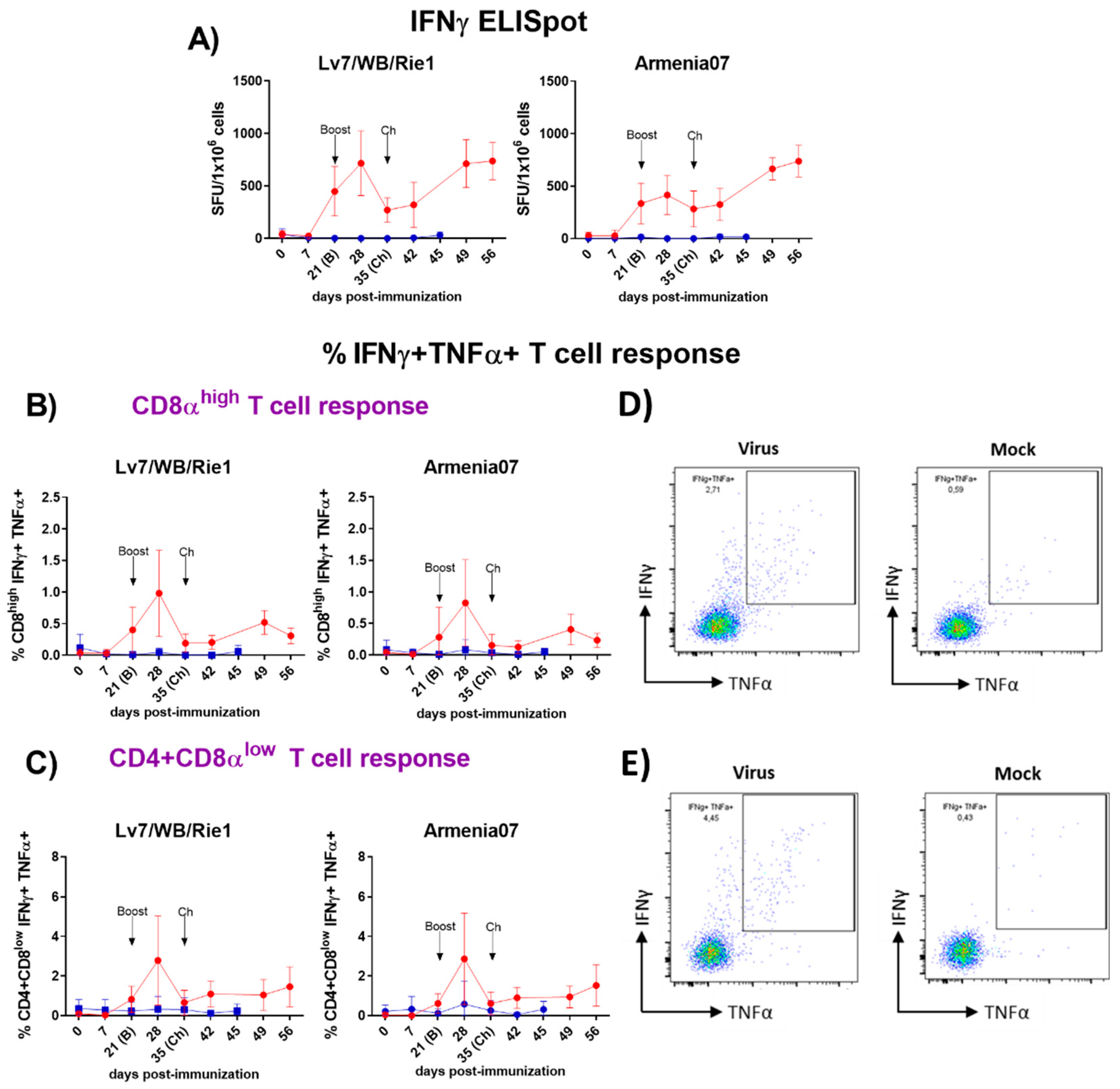
Figure 2.
(A) Evaluation by IFNγ ELISpot of virus-specific IFNγ T cell responses following immunization with Lv7/WB/Rie1 on 0 (prime) and 21 (boost) dpi and challenge with Armenia07 at 35 dpi. Responses of PBMC to stimulation with Lv7/WB/Rie1 and Arm07 are presented as the mock-corrected number of IFNγ spot forming units (SFU) per million cells. B-E) Phenotyping of virus-specific IFNγ responder cells by intracellular cytokine staining (ICS) using flow cytometry following immunization with Lv7/WB/Rie1 on 0 (prime) and 21 (boost) dpi and challenge with Armenia07 at 35 dpi. T cell populations were defined as CD3+CD4negCD8high (cytotoxic CD8+) (B) and CD3+CD4+CD8low (memory CD4+CD8+) (C). Intracytoplasmic co-expression of IFNγ and TNFα was assessed in each population. Longitudinal responses of resucited PBMC to stimulation with Lv7/WB/Rie1 and Armenia07 are presented as the mock-corrected %IFNγ+TNFα+CD8αhigh (B) and %IFNγ+TNFα+CD4+CD8αlow T cells (C). Immunized group (red line) and non-immunized control group (blue line). Mean data ± SD from four pigs/time point are shown for each group. D) Representative dot plots showing gates defining double expression of IFNγ+TNFα+ in singlet, live cytotoxic CD8+ T lymphocytes after stimulation with virus or mock. E) Representative dot plots showing gates defining double expression of IFNγ+TNFα+ in singlet, live memory CD4+CD8+ T lymphocytes after stimulation with virus or mock. Representative dot plots from one pig of the immunized group at 28 dpi are displayed. Final gates define percentages of double expression %IFNγ+TNFα+. .
Figure 2.
(A) Evaluation by IFNγ ELISpot of virus-specific IFNγ T cell responses following immunization with Lv7/WB/Rie1 on 0 (prime) and 21 (boost) dpi and challenge with Armenia07 at 35 dpi. Responses of PBMC to stimulation with Lv7/WB/Rie1 and Arm07 are presented as the mock-corrected number of IFNγ spot forming units (SFU) per million cells. B-E) Phenotyping of virus-specific IFNγ responder cells by intracellular cytokine staining (ICS) using flow cytometry following immunization with Lv7/WB/Rie1 on 0 (prime) and 21 (boost) dpi and challenge with Armenia07 at 35 dpi. T cell populations were defined as CD3+CD4negCD8high (cytotoxic CD8+) (B) and CD3+CD4+CD8low (memory CD4+CD8+) (C). Intracytoplasmic co-expression of IFNγ and TNFα was assessed in each population. Longitudinal responses of resucited PBMC to stimulation with Lv7/WB/Rie1 and Armenia07 are presented as the mock-corrected %IFNγ+TNFα+CD8αhigh (B) and %IFNγ+TNFα+CD4+CD8αlow T cells (C). Immunized group (red line) and non-immunized control group (blue line). Mean data ± SD from four pigs/time point are shown for each group. D) Representative dot plots showing gates defining double expression of IFNγ+TNFα+ in singlet, live cytotoxic CD8+ T lymphocytes after stimulation with virus or mock. E) Representative dot plots showing gates defining double expression of IFNγ+TNFα+ in singlet, live memory CD4+CD8+ T lymphocytes after stimulation with virus or mock. Representative dot plots from one pig of the immunized group at 28 dpi are displayed. Final gates define percentages of double expression %IFNγ+TNFα+. .
3.3.3. Comprehensive Functional Characterization of ASFV-Induced T-Cell Responses by Flow Cytometry in the Immunized/Challenged Animals
A more detailed flow cytometry study combining phenotypic analysis with functional assays was performed at selected time points (0, 7, 21, 28, 35, 42 dpi) with PBMC from three representative animals in the immunized/challenge group. Following the “ex vivo” stimulation of PBMC with virus, mock or medium, frequencies of CD3 T cells and the CD3+ T subpopulations defined as CD3+CD4+CD8neg (helper CD4), CD3+CD4negCD8high (cytotoxic CD8) and CD3+CD4+CD8low (activated/memory CD4CD8), in combination with the simultaneous detection of intracellular IFNγ and TNFα, the cytotoxic degranulation marker CD107a and the detection of the major histocompatibility complex (MHC)-class II (SLA-class II in pigs) were studied.
(a) Changes in the frequencies of CD3 and CD3+T cell subsets.
The analyses were performed with PBMC from immunized/challenged pigs after priming immune responses with medium or virus (Figure 3). Basal responses to medium in the immunized animals (Figure 3A) evidenced a moderate and transient, yet not significant, increase in frequencies of CD4CD8 cells 7 days after the prime immunization. On the contrary, CD8 and CD4 frequencies experienced a clear decrease at 7 dpi. All the CD3+ subsets returned to pre-immunization values at 21 dpi, except for the helper CD4 cells, which remained at lower levels from 7 dpi and during all the study. Although not statistically significant, additional changes were noticed after the boost (28 dpi). Thus, second inoculation of Lv17/WB/Rie1 induced a new drop of CD8 cells and a small and transitory decrease of CD4CD8 and CD4 cells. CD3 cells raised from 35 dpi onwards, remaining high and above pre-immunization levels until the end of study. However, none of the three CD3 subsets studied exhibited a similar trend, except for CD4CD8 and to a lesser extent CD8 at 42 dpi, which increased again after the challenge. Nevertheless, these changes were not statistically significant compared to previous values. On the other hand, the frequency of specific CD3 T cells in response to recall antigen increased most markedly between 7 and 28 dpi (Figure 3B), showing less individual variability and reaching the highest numbers, on average, between 21 and 28 dpi against both virus stimuli. At 21 dpi, a moderate, albeit not statistically significant, increase in the frequency of virus-specific CD8 T cells was observed. No noticeable changes in the virus recall response were observed among the other populations studied.
(b) Assessment of the expression of IFNγ, degranulation marker CD107a, MHC-class II and the co-expression of TNFα.
Simultaneous analyses of phenotype and function were performed with PBMC from immunized/challenged pigs after priming immune responses with medium (Figure 4) or virus (Figure 5). Spontaneous/basal responses to medium in the immunized pigs (Figure 4) evidenced some changes in cytotoxic CD8 (Figure 4A and D), activated/memory CD4CD8 (Figure 4B and E) and helper CD4 (Figure 4C and F) T cells after the prime immunization in the immunized/challenge animals. Immunization induced a moderate, yet not significant, increase on baseline levels of CD4CD8 T cells expressing IFNγ alone or in combination with TNFα, mainly after the prime (7 dpi) and the boost (28 dpi). This CD4CD8 response was reduced to almost absent levels from 35 dpi until the end of the study (Figure 4B and E). Baseline IFNγ and IFNγTNFα CD8 responses to medium were very poor and barely suffered any changes during the study (Figure 4A and D). Meanwhile, spontaneous cytokine response from CD4 T cells was absent during all study (Figure 4C and F).
To further elucidate this response, the ability of these cells to degranulate and secrete both cytokines was investigated. All subsets of CD3 T cells studied (CD8, CD4CD8 and CD4) showed evidences of non-specific cytotoxicity after immunization, as indicated by the increase in CD107a expression from 21 dpi onward. Before this increase took place, all subsets experienced a clear loss of CD107a expression at 7 dpi. CD107a+CD8 showed the highest increase at 28 dpi, although this increase did not reach statistical significance at any time point during the study (Figure 4A). CD107a+CD8 and CD107a+CD4CD8 cytotoxic T-lymphocytes (CTL) secreting simultaneously TNFα also showed a similar trend (Figure 4D-E), displaying the CD4CD8 CTL the highest frequencies and maximal increases, on average, at days 21 and 28 (Figure 4E). Although differences observed at these time points were not statistically significant due to variability between individuals, they were more pronounced for CD4CD8 CTL (Figure 4E). Interestingly, basal frequencies of cells expressing CD107a were significantly high for CD4CD8 (p<0.0001) (Figure 4B) and CD4 T cells (p<0.001) (Figure 4C) before challenge (35 dpi), and for CD4 T cells (p<0.05) also after challenge (42 dpi) (Figure 4C). This indicated that both T cell subpopulations exhibited high levels of cytotoxic activity even before challenge, which may have contributed to the defense against viral challenge.
Analysis of basal expression of the porcine MHC-class II (SLA-II) in the immunized/challenged group following the prime immunization with Lv17/WB/Rie1 revealed some significant changes. Thus, immunization led to an evident increase in the frequency of CD8, CD4CD8 and CD4 T cells that expressed this marker. This increase was observed after priming (7 dpi) and after boosting (28 dpi) in the three CD3 T subsets. In all cases, the increase in SLA-II expression was most notable after the prime, particularly evident at 7 dpi, although statistical significance was observed only for CD4 at this time point (p<0.01) and after the boost at 28 dpi (p<0.05) (Figure 4C). SLA-II expression levels remained elevated for CD8 (Figure 4A) and for CD4CD8 (Figure 4B) after boost (from 21 dpi onwards), although it is worth pointing out that the challenge with Arm07 triggered a further rise in the percentage of SLA-II+CD4CD8 cells (42 dpi) but not in the others. The SLA-II+CD4 cells frequencies decreased markedly from 35 dpi, falling below pre-immunization levels and remaining low until the end of the study, when levels show a statistically significant decrease (p<0.05) (Figure 4C). After immunization, the double positively stained (SLA-II+TNFα+) CD8 (Figure 4D) and CD4CD8 (Figure 4E) cell subsets increased from 7 to 28 dpi, peaking between day 21 and 28; however, significant differences to pre-immunization levels could be observed only for the SLA-II+TNFα+CD8 by day 21 (p<0.05) (Figure 4D). At the time of challenge (35 dpi), frequencies of CD8 and CD4CD8 that were expressing simultaneously SLA-II and TNFα dropped to almost pre-immunization levels that were maintained until the end of the study (Figure 4D-E). Percentages of SLA-II+TNFα+CD4CD8 at 35 and 42 dpi were even lower than the ones observed at day 0 (Figure 4E). Finally, baseline SLA-II+TNFα+CD4 responses to medium were barely detected and did not suffer any changes during the study (Figure 4F).
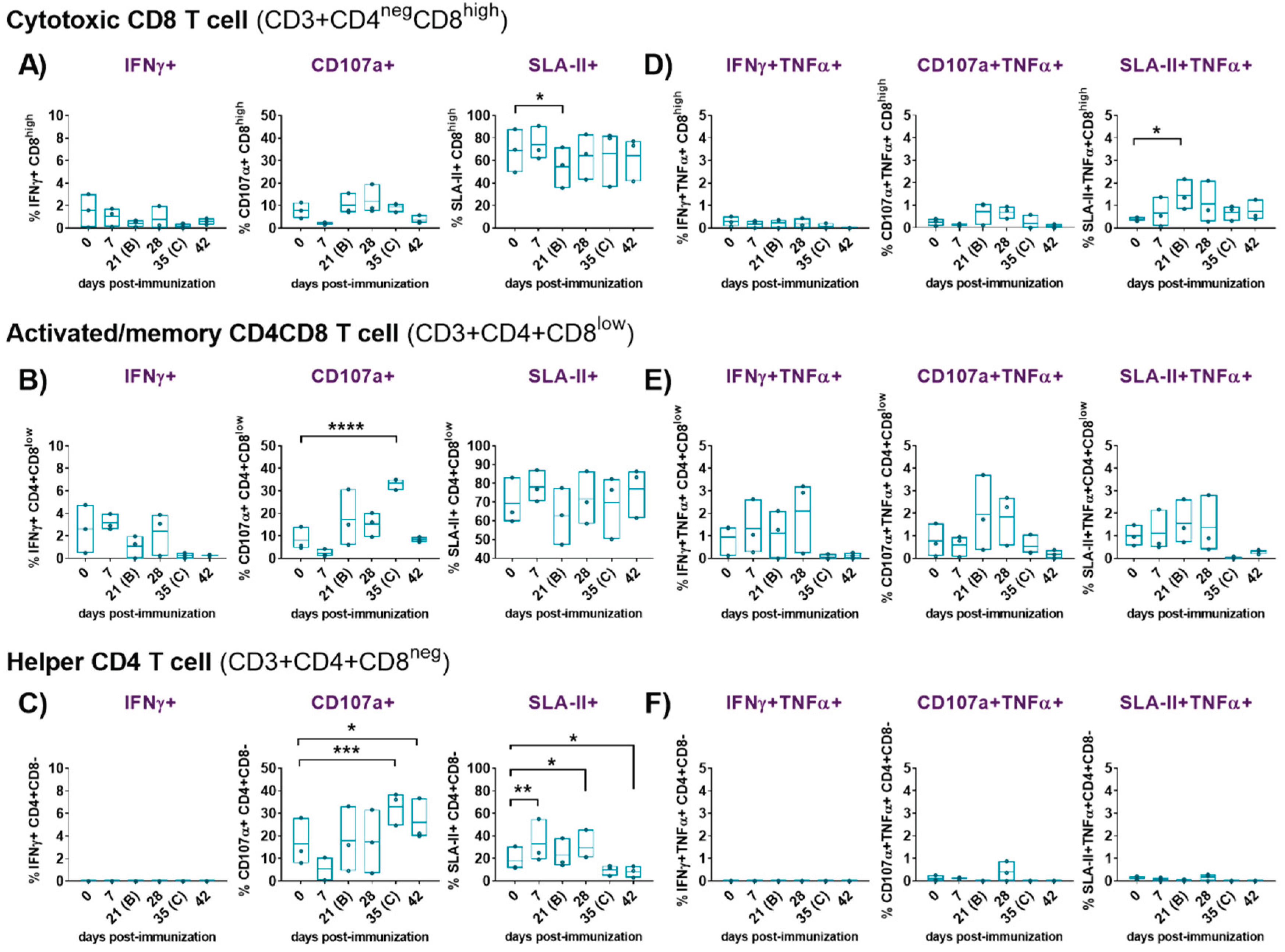
Figure 4.
Evaluation of IFNγ response, cytotoxicity (CD107a degranulation marker ) and expression of porcine class II major histocompatibility complex (SLA-II) of cytotoxic CD8, activated/memory CD4CD8 and helper CD4 T cells following immunization with Lv7/WB/Rie1 on 0 (prime) and 21 (boost) dpi and after challenge with Armenia07 (35 dpi). Responses of resucited PBMC to stimulation with medium were assessed by flow cytometry at selected days post-immunization (dpi) (0, 7, 21, 28, 35 and 42 dpi). T cell populations were defined as CD3+CD4negCD8high (cytotoxic CD8), CD3+CD4+CD8low (memory CD4CD8) and CD3+CD4+CD8neg (helper CD4). Single expression of intracytoplasmic IFNγ, CD107a and SLA-II was assessed for CD8 (A), CD4CD8 (B) and CD4 (C) T cells. Representative data for each cellular population on live, singlet lymphocytes after medium stimulation are presented as % IFNγ+, %CD107a+ and % SLA-II+. Double expression of intracytoplasmic IFNγ and TNFα, CD107a and TNFα, and SLA-II and TNFα, was assessed for CD8 (D), CD4CD8 (E) and CD4 (F) T cells. Representative data for each cellular population on live, singlet lymphocytes after medium stimulation are presented as % IFNγ+TNFα+, % CD107a+TNFα+ and % SLA-II+TNFα+. Spontaneous/basal responses to medium in the immunized pigs. Floating bars of individual data and line at mean from three pigs are shown for each time point. Significance is indicated by: *(p<0.05), **(p<0.01), ***(p<0.001) and ****(p<0.0001).
Figure 4.
Evaluation of IFNγ response, cytotoxicity (CD107a degranulation marker ) and expression of porcine class II major histocompatibility complex (SLA-II) of cytotoxic CD8, activated/memory CD4CD8 and helper CD4 T cells following immunization with Lv7/WB/Rie1 on 0 (prime) and 21 (boost) dpi and after challenge with Armenia07 (35 dpi). Responses of resucited PBMC to stimulation with medium were assessed by flow cytometry at selected days post-immunization (dpi) (0, 7, 21, 28, 35 and 42 dpi). T cell populations were defined as CD3+CD4negCD8high (cytotoxic CD8), CD3+CD4+CD8low (memory CD4CD8) and CD3+CD4+CD8neg (helper CD4). Single expression of intracytoplasmic IFNγ, CD107a and SLA-II was assessed for CD8 (A), CD4CD8 (B) and CD4 (C) T cells. Representative data for each cellular population on live, singlet lymphocytes after medium stimulation are presented as % IFNγ+, %CD107a+ and % SLA-II+. Double expression of intracytoplasmic IFNγ and TNFα, CD107a and TNFα, and SLA-II and TNFα, was assessed for CD8 (D), CD4CD8 (E) and CD4 (F) T cells. Representative data for each cellular population on live, singlet lymphocytes after medium stimulation are presented as % IFNγ+TNFα+, % CD107a+TNFα+ and % SLA-II+TNFα+. Spontaneous/basal responses to medium in the immunized pigs. Floating bars of individual data and line at mean from three pigs are shown for each time point. Significance is indicated by: *(p<0.05), **(p<0.01), ***(p<0.001) and ****(p<0.0001).
The detailed functional analysis of ASFV-specific responses after priming PBMC with both viruses revealed significant findings (Figure 5). In addition to the virus-specific IFNγ+TNFα+CD8 and IFNγ+TNFα+CD8CD4 T cell responses observed in the immunized animals after the prime immunization (described in section 3.3.2), a high proportion of the responding cells also expressed CD107a, indicating cytotoxic activity. The ASFV-specific CD107a+CD8 CTL (Figure 5A) and CD107a+CD8CD4 CTL (Figure 5B) responses peaked between 21 and 28 dpi, with the CD8CD4 response achieving statistical significance at 28 dpi (p<0.05). On day 42, a virus-specific CD8CD4 cytotoxic response was mounted against both virus stimuli, although only reached significant difference (p<0.05) compared to day 0 after priming with Arm07 (Figure 5B). A proportion of the responding CD8 CTL and CD8CD4 CTL also co-expressed TNF-α (Figure 5D-E), and such responses were more pronounced from 21 to 28 dpi. However, only the CD107a+TNFα+CD8CD4 response achieved a significant difference (p<0.01) at 28 dpi (Figure 5E). These findings, illustrated in Figure 5, indicated robust cytotoxic and cytokine-producing responses mounted by T cells following priming with ASFV, particularly observed in the CD8CD4 subset.
With regards to CD4 helper T cells, notable cytotoxic responses were observed at 42 dpi (7 days post-challenge), when a highly significant CD4 CTL response was mounted against both Lv17/WB/Rie1 (p<0.01) and Arm07 (p<0.001) (Figure 5C). However, neither CD107a+ nor CD107a+TNFα+ ASFV-specific responses were clearly observed in the CD4 T cell subset before this point, indicating minimal or absent cytotoxic activity of these cells until day 42 (Figure 5C and F).
For SLA-II, a virus-specific SLA-II+CD8 response triggered by stimulation with both viruses was observed from day 7 onwards (Figure 5A). These responses, although not statistically significant, peaked between 21 and 28 dpi. Beyond this point (28 dpi), SLA-II+CD8 response decreased, although a new increase following the challenge was detected. Similar trend was observed for the SLA-II+CD4CD8 cells (Figure 5B). On the other hand, SLA-II+CD4 response was of lower intensity and appeared earlier, showing a small peak at day 7 (Figure 5C). In line with the induction of virus-specific IFNγ+TNFα+ and CD107a+ TNFα+ expression displayed by both CD8 and CD8CD4 T cells subsets, a highly significant virus-specific SLA-II+TNFα+ response was mounted against Lv17/WB/Rie1 and Arm07 from both subsets at 21 and 28 dpi (Figure 5D-E). In particular, the specific SLA-II+TNFα+CD8 response (Figure 5D) against Lv17/WB/Rie1 achieved significant differences at day 21 (p<0.01), displaying even higher differences at day 28 (p<0.001). While the differences at day 21 and 28 were also significant against Arm07, these achieved lower significance (p<0.05). Likewise, the SLA-II+TNFα+CD4CD8 response (Figure 5E) against both virus stimuli was statistically significant (p<0.01) at both days (21 and 28 dpi). No SLA-II+TNFα+ ASFV-specific responses were observed for the CD4 subset (Figure 5F).
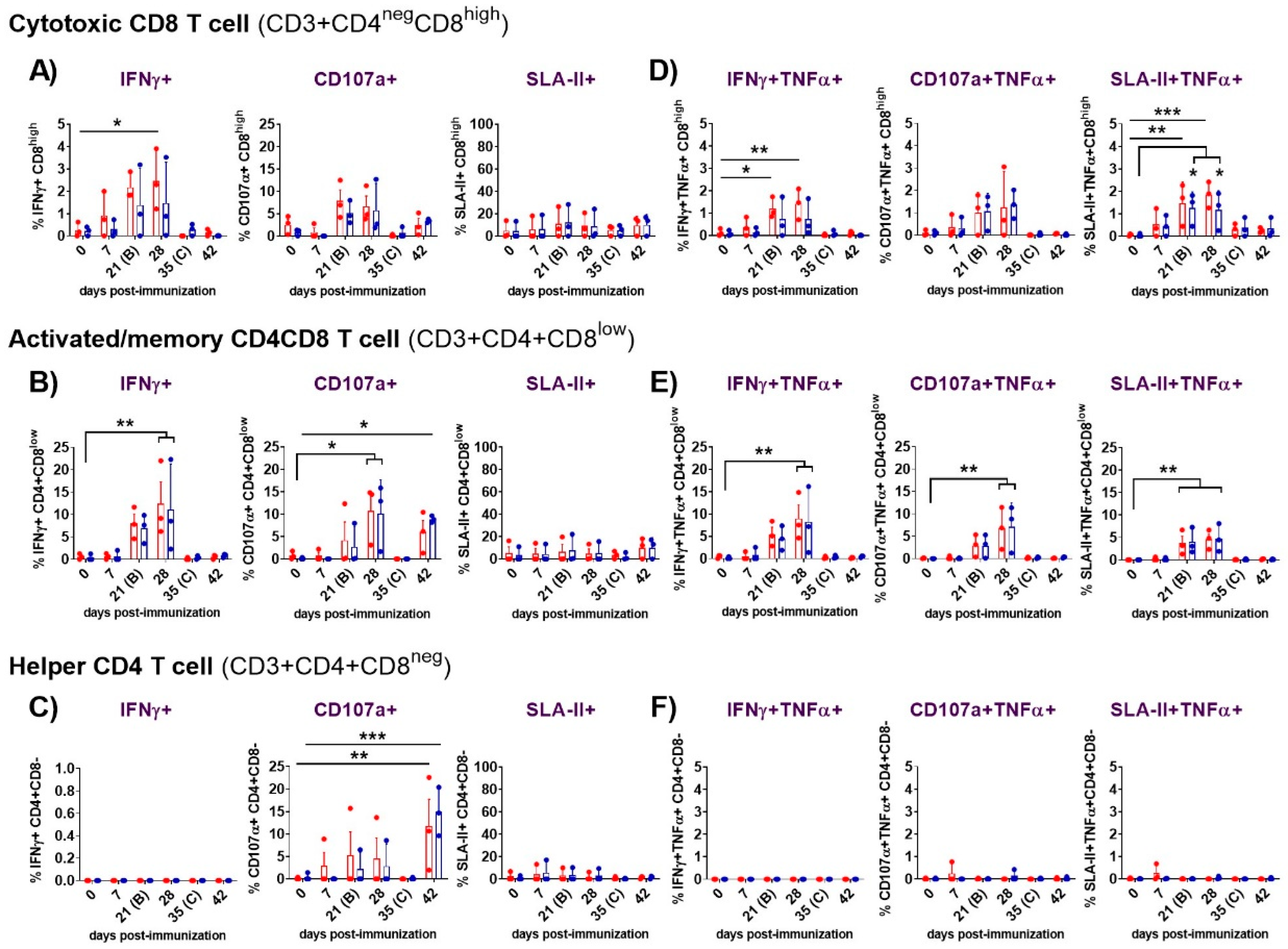
Figure 5.
Evaluation of IFNγ response, cytotoxicity (CD107a degranulation marker ) and expression of porcine class II major histocompatibility complex (SLA-II) of cytotoxic CD8, activated/memory CD4CD8 and helper CD4 T cells following immunization with Lv7/WB/Rie1 on 0 (prime) and 21 (boost) dpi and after challenge with Armenia07 (35 dpi). Responses of resucited PBMC to stimulation either with Lv7/WB/Rie1 (red dots and box), Armenia07 (blue dots and box) or mock virus supernantant were assessed by flow cytometry at selected days post-immunization (dpi) (0, 7, 21, 28, 35 and 42 dpi). T cell populations were defined as CD3+CD4negCD8high (cytotoxic CD8), CD3+CD4+CD8low (memory CD4CD8) and CD3+CD4+CD8neg (helper CD4). Single expression of intracytoplasmic IFNγ, CD107a and SLA-II was assessed for CD8+ (A), CD4+CD8+ (B) and CD4+ (C) T cells. Representative data for each cellular population on live, singlet lymphocytes after virus stimulation are presented as the mock-corrected % IFNγ+, %CD107a+ and % SLA-II+. Double expression of intracytoplasmic IFNγ and TNFα, CD107a and TNFα, and SLA-II and TNFα, was assessed for CD8 (D), CD4CD8 (E) and CD4 (F) T cells. Representative data for each cellular population on live, singlet lymphocytes after virus stimulation are presented as the mock-corrected % IFNγ+TNFα+, % CD107a+TNFα+ and % SLA-II+TNFα+. Individual and mean data ± SD from three pigs are shown for each time point. Significance is indicated by: *(p<0.05), **(p<0.01), ***(p<0.001) and ****(p<0.0001).
Figure 5.
Evaluation of IFNγ response, cytotoxicity (CD107a degranulation marker ) and expression of porcine class II major histocompatibility complex (SLA-II) of cytotoxic CD8, activated/memory CD4CD8 and helper CD4 T cells following immunization with Lv7/WB/Rie1 on 0 (prime) and 21 (boost) dpi and after challenge with Armenia07 (35 dpi). Responses of resucited PBMC to stimulation either with Lv7/WB/Rie1 (red dots and box), Armenia07 (blue dots and box) or mock virus supernantant were assessed by flow cytometry at selected days post-immunization (dpi) (0, 7, 21, 28, 35 and 42 dpi). T cell populations were defined as CD3+CD4negCD8high (cytotoxic CD8), CD3+CD4+CD8low (memory CD4CD8) and CD3+CD4+CD8neg (helper CD4). Single expression of intracytoplasmic IFNγ, CD107a and SLA-II was assessed for CD8+ (A), CD4+CD8+ (B) and CD4+ (C) T cells. Representative data for each cellular population on live, singlet lymphocytes after virus stimulation are presented as the mock-corrected % IFNγ+, %CD107a+ and % SLA-II+. Double expression of intracytoplasmic IFNγ and TNFα, CD107a and TNFα, and SLA-II and TNFα, was assessed for CD8 (D), CD4CD8 (E) and CD4 (F) T cells. Representative data for each cellular population on live, singlet lymphocytes after virus stimulation are presented as the mock-corrected % IFNγ+TNFα+, % CD107a+TNFα+ and % SLA-II+TNFα+. Individual and mean data ± SD from three pigs are shown for each time point. Significance is indicated by: *(p<0.05), **(p<0.01), ***(p<0.001) and ****(p<0.0001).
4. Discussion
In this study, intradermal immunization of domestic pigs, using a prime/boost regime with the attenuated Lv17/WB/Rie1 strain conferred high levels of protection against virulent challenge with Arm07. Unlike intramuscular inoculation, which may lead to adverse reactions, the intradermal approach not only conferred robust protection but also reduced adverse reactions typically associated with live vaccine candidates in domestic pigs [24,25]. Although one of the five immunized pigs developed viraemia and moderate clinical signs after the first intradermal inoculation, leading to its euthanasia prior to the boost, the remaining four animals were fully protected. These four pigs developed robust immunity that was evident after the prime immunization, characterized by the induction of ASFV-specific antibodies and virus-specific IFNγ-T cell responses. This pattern of immune response aligns with previous findings, suggesting that the combination of antibody induction with a potent virus-specific cellular response improves protection against subsequent infections [39,40,41]. Hence, this experimental model provides an excellent opportunity to study in detail the mechanisms defining the protective immune response induced by attenuated strains against virulent ASFV challenge in natural hosts.
The mild and transient increase in body temperature observed after prime immunization with the attenuated strain, coupled with low and transient viremia, is consistent with previous observations in both domestic pigs [4,24,25] and wild boar [22,23]. Unlike the findings in wild boar immunized orally with repeated doses of the Lv17/WB/Rie1 strain [23], the intradermal route did not induce notable viremia or clinical signs after the booster dose. These differences could be attributed to the different routes of administration used (intradermal vs. oral). Previous studies have proven that intradermal vaccination is apparently more effective than other routes of vaccination, such as intramuscular or subcutaneous, even if the latter are administered in repeated doses [42,43,44]. Although vaccination by these routes may be equally immunogenic, the dose may be reduced when the vaccine is administered intradermally [45]. Dermis is rich in resident dendritic cells (DC), especially Langerhans cells and dermal DC, and although plasmacytoid DC (pDC) are rare in skin, they quickly infiltrate this organ during inflammation. It is known that pDC become stimulated by the virus to produce type I IFN during acute ASFV infections (reviewed in [46]), cytokine that seems to be crucial in innate protection against some attenuated ASFV strains [21,47]. Therefore, as has been suggested in other immunization studies carried out against haemorrhagic viruses such as Ebola [48], intradermal immunization would trigger enhanced adaptive immune responses by recruiting more dermal DC subsets to the inoculation site, including pDC. It would increase the chances of success against subsequent infections even using reduced or single doses during immunizations, hypothesis that requires further studies with ASFV in domestic pigs and wild boar.
An increase in circulating levels of all cytokines studied was observed after challenge in the non-immunized control pigs. This event, also known as "cytokine storm," is directly associated with severe disease caused by virulent strains when animals lack protection [49,50]. Unlike controls after challenge, where IL-8 levels increased significantly in parallel with the development of clinical signs and viremia, IL-8 levels in the immunized animals remained relatively stable after the initial peak following the first immunization (between 3-10 dpi), with minimal fluctuations observed after booster and challenge. Although studies on IL-8 are contradictory, and often fail to observe a significant modulation of this chemokine following infection with virulent or attenuated ASFV isolates, our findings align with previous studies indicating an increase in circulating levels of IL-8 following infection with the virulent Arm07 strain [51]. The early and controlled increase of IL-8 in immunized pigs suggests an "in vivo" modulation of circulating IL-8 after immunization with the attenuated Lv17/WB/Rie1 strain, which appears to be correlated with protection against ASFV.
Similarly, IL-10 levels remained minimally affected in the immunized/challenged group, except for some animals that exhibited a mild increase shortly after the first immunization (3 dpi). In contrast, in the control group, a substantial increase in IL-10 was observed consistent with the rise in IL-8 after challenge, indicating, in line with other studies (reviewed in [52]), a direct correlation between elevated levels of both cytokines and an exacerbated and/or uncontrolled inflammatory immune response after Arm07 challenge.
The controlled and early increases of IL-10, cytokine with a potent and broad anti-inflammatory activity, have also been described in wild boar experimentally immunized with Lv17/WB/Rie1 [53], as well as in domestic pigs experimentally inoculated with some attenuated ASFV vaccine strains [19,21,54]. Thus, our results are consistent with the premise that a controlled and early increase in IL-10 may contribute to controlling viral replication and dampening the exacerbated inflammatory response that often leads to fatal outcomes during ASFV infections. The simultaneous decrease in circulating levels of TNFα and IFNα, two cytokines with important pro-inflammatory functions, which are normally associated with tissue damage and the appearance of clinical signs [49,52,55], showed evidence of a restrained inflammatory response in the immunized animals. In summary, the transient mild to moderate increase in IL-8 and IL-10 observed in pigs immunized with Lv17/WB/Rie1 seems to be directly correlated with survival, by contributing to the control of virus spread and the inflammatory response. Thus, moderate and controlled serum levels of these two cytokines, but especially IL-8, could be good markers of protection as well as of favorable evolution of ASFV infection.
IFNγ plays a pivotal role in inducing and modulating immune responses but it remains unclear whether protection against ASFV is linked to IFNγ production. A study conducted in domestic pigs reported a significant rise in circulating IFNγ, as well as TNFα, seven days after immunization with the attenuated vaccine candidate HLJ/18-7GD [16]. Similarly, significant increases were observed at 28 dpi in wild boar orally inoculated with Lv17/WB/Rie1 [53]. However, other studies did not find remarkable changes in circulating levels of IFNγ following immunization of pigs with different attenuated ASFV isolates [21,56,57], including the Lv17/WB/Rie1/d110-11L and Lv17/WB/Rie1 ASFV strains inoculated intramuscularly [25]. In our study, intradermal immunization with Lv17/WB/Rie1 induced detectable, albeit not significant, increases of circulating IFNγ, as well as TNFα, 7 days after boost (28 dpi), indicating that the presence of these small peaks seems to be directly related to virus recognition and protection against reinfections with the same strain. A second IFNγ peak observed three days after challenge (38 dpi) in some animals would suggest that IFNγ production may also be linked with protection against virulent challenge with Arm07.
Regarding the phenotypic characterization of the T cell responses, the different subpopulations of PBMC in the immunized animals suffered changes in their frequencies throughout the experiment. Without viral stimulation of PBMC, analysis of CD3+ and CD3+ subset frequencies showed that CD3, together with the activated/memory CD4CD8 cells, experienced a moderate and transient increase 7 days after immunization with Lv17/WB/Rie1, while, at the same time, cytotoxic CD8 and helper CD4 T cells experienced a clear drop in their frequencies. Significant increases in CD4CD8 cells have been reported as early as 4 days after immunization with the attenuated genotype I vaccine candidates OURT88/3 and Benin ΔMGF [54]. Fluctuations in cytotoxic CD8 cells after immunization with both vaccine strains have also been reported [54]. However, other authors did not observe changes in pigs immunized with OURT88/3 prior to challenge [41]. In our study, these populations returned to pre-immunization values at 21 dpi, except for the helper CD4 cells, which remained low from 7 dpi and throughout the study. A progressive decrease of circulating CD4 helper T cells has also been described with the same genotype I vaccine strains listed above, in both protected and unprotected pigs [41,54]. All subset of CD3 T cells increased again after the challenge, mainly CD4CD8, moderately the CD8 cells and to a lesser extent the CD4 cells. Increases in CD3+ cells, mainly CD4CD8 and CD8, have also been reported following virulent challenge in pigs that became protected after immunization with OURT88/3 [41]. Frequency analysis of these populations in response to recall ASFV showed the induction of a CD3-specific proliferative response between 21 and 28 dpi, although among the CD3+ subsets studied, only CD8 showed a small increase in response to recall virus at 21 dpi in some of the immunized animals. The increase in these circulating T cell subsets provides evidence of activation of the adaptive cellular immune response following immunization capable of responding to challenge with Arm07.
A peculiarity of the porcine immune system is the high expression of MHC class II (SLA-II) DR in resting lymphocytes, although the use of this marker, in combination with CD8α, has demonstrated to be of great help identifying activation in porcine helper T cells [58]. Previously to immunizing the animals, circulating CD4 helper T cells lacked CD8 expression and had variable expression of SLA-II, phenotype that matched the normal description of resting helper T cells. CD4CD8 and CD8 T cells, however, expressed high levels of SLA-II, which is in line with the description carried out of these T cell subsets in blood of healthy pigs [59]. Immunization with Lv17/WB/Rie1 led to the transient upregulation of the SLA-II surface protein in circulating CD8, CD4CD8 and CD4 T cells after prime and boost; however, these increases resulted significant only for the CD4 T cells. The transitory upregulation of SLA-II protein expression induced after immunization with Lv17/WB/Rie1 would indicate an increase in antigen presentation phenomena and would confirm the existence of regulatory mechanisms that activate an adaptive immune response. Due to the fact that co-expression between SLA-II and CD8α appears strongly associated in TCR- αβ T lymphocytes [58,59], it was not surprising that SLA-II expression remained high in both CD8 positive T cells during all study.
Simultaneous production of different cytokines or effector molecules on the single T-cell level has been proposed to be a hallmark of protective immune responses. For this purpose, we aimed to identify potential multifunctional virus-specific T cells. The correlation between virus-specific IFNγ-producing cells and protection has been described in some “in vivo” studies by using different techniques such as ELISA, ELISpot assay or flow cytometry [11,40,60,61]. In other studies, in which the authors carried out a phenotypic characterization by flow cytometry of IFNγ-producing cells in vaccinated pigs, they did not find a clear relationship between protection and induction of IFNγ-specific T cells. However, they could not rule out a possible protective role for these cells [39,41,54]. The combined expression of two cytokines, such as IFNγ and TNFα, is a good indicator of the quality of the responses. Our results showed a high proportion of CD8 and CD4CD8 cells that, along with IFNγ, also co-expressed TNFα, with CD4CD8 cells exhibiting the highest virus-specific IFNγ+TNFα+ response throughout the study. Although a specific CD8 T-cell response was detected in some animals as early as 7 dpi, the induction of both responses became clearer between 21 and 28 dpi, showing the highest level of significance at 28 dpi (7 days after the boost). The induction of elevated percentages of ASFV-specific polyfunctional memory T cells, i.e. IFNγ+TNFα+ CD4CD8 T cells in pigs immunized with the BA71ΔCD2 deleted mutant, a vaccine candidate that conferred protection against virulent challenge with genotype II Georgia2007/1 strain, has also been described recently [40]. In our study, the high levels of circulating IFNγ+TNFα+ CD4CD8 cells detected 7 days after boosting (28 dpi), in the absence of recall virus antigen, would also suggest that CD4CD8 T cells might be involved of the spontaneous increases of these cytokines detected in sera at this time point. Hence, intradermal immunization with Lv17/WB/Rie1 induced a robust ASFV-specific IFNγ T cell response, which was clearly detectable after the first immunization at 21 dpi, prior to booster, where CD4CD8 and CD8 T cells were identified as the main cellular sources of virus-specific IFNγ and TNFα. These results demonstrate the correlation between the induction of virus-specific CD4CD8 and CD8 T cells and protection against subsequent infections with both attenuated and virulent strains of ASFV genotype II.
One of our aims was to study the role of specific cytotoxic T-lymphocytes (CTL) in protection against subsequent ASFV infection. Although surface expression of CD8 has traditionally been attributed to cytotoxic functions, it is important to note that not all CD8 cells exhibit this capability [62]. Therefore, the inclusion of markers indicating cytotoxicity, such as perforin or CD107a, may be useful in defining CTL subpopulations. The CD107a assay has been used to study cytotoxic degranulation associated with loss of perforin in porcine T cells following antigenic stimulation in other porcine viral diseases such as classical swine fever [36,38], porcine respiratory and reproductive syndrome (PRRS) [37,63] or swine influenza A [64]. Although CD4+CD8+ and CD8 T cell cytotoxic activity, demonstrated by the detection of perforin expression, has been described during experimental ASFV infections [15,40,62,65], to our knowledge, this is the first study reporting the use of CD107a assay to identify CTL in ASFV-infected pigs. Immunization induced a variable, although progressive, increase of T cells with cytotoxic function in blood, as demonstrated by detection of the CD107a marker. CD8 CTL, CD4CD8 CTL as well as CD4 CTL increased from 21 dpi, and in the case of CD8 CTL they were maintained until 28 dpi. Mainly in the case of CD4CD8 CTL, this increased cytotoxic activity was accompanied by a marked and spontaneous secretion of cytokines and by increased expression of SLA-II. Only in the case of CD4 CTL, their progressive increase in blood was not accompanied by simultaneous cytokine production, although this increase in circulating CD4 CTL did appear to be associated, only after the boost (at 28 dpi), with an up-regulation of SLA-II expression on CD4 helper T cells. Similarly, upon virus stimulation, IFNγ and IFNγ+TNFα+ producing T cells were limited mainly to cytotoxic CD4CD8, and also to cytotoxic CD8. Taken together, these results confirm the important role of CD4CD8 T cells during the early stages of infection with attenuated ASFV strains in stopping viral replication, but also their key role mounting an effective adaptive immune response that induces protection against subsequent infections. The generation of an important subpopulation of antigen-experienced CD4CD8 T cells during the induction of the adaptive immune response was also confirmed. Furthermore, a second subset of memory CD8 T cells (CD4negCD8high) was also identified. These cells were able to proliferate quickly after antigen re-encounter, as indicated by the quality of these virus-specific multifunctional CD4CD8 and CD8 T responses elicited upon stimulation with both viruses at day 21 and especially at 28 dpi. Beyond 28 dpi, only CD4CD8 CTL and CD4 CTL remained very significantly elevated. However, after the challenge, while CD4CD8 CTL declined dramatically, CD4 CTL frequencies remained markedly elevated. The strong correlation between CD4 CTL appearance and control of infection with Arm07 suggested an important protective role of this subset in the control of early replication and infection with virulent virus in previously immunized pigs. It is noteworthy that, although the specific cytokine secretion by these T-cell subsets after challenge was weak, all of them (mainly CD4, followed by CD4CD8) were able to mount significant virus-specific CTL responses against both viral stimuli, indicating also the correlation between CD4 and CD4CD8 CTL subsets and protection against subsequent infection with a virulent genotype II isolate. The inclusion of additional markers, such as CD25, might help to better differentiate memory subpopulations among these CTL subsets.
Recent evidence highlights the potential role of CD4 CTL in controlling and protecting against viral diseases in pigs, particularly in the context of porcine respiratory and reproductive syndrome virus (PRRSV) infections [63,66]. These studies suggest that, in addition to the high levels of PRRSV-specific CD4CD8 CTL acquired by vaccination or previous infection, elevated levels of PRRSV-specific CD4 CTL are crucial in host defense against subsequent infections, even in the absence of neutralizing antibodies [63,66]. While CD4 CTL are unlikely to replace the function of CD8 CTL or CD8CD4 CTL, CD4 cytotoxic activity contributes to immune responses by targeting antigen-presenting cells (APC) via the MHC class II pathway. In mice and humans, CD4 CTL presence is associated with chronic viral infections, autoimmune diseases and cancer, attributing them important antiviral functions and suggesting their potential importance during adaptive cytotoxic immune responses [67]. The induction of CD4 CTL responses targeting APC may be particularly relevant in scenarios where CD8 CTL responses are insufficient or compromised due to sustained antigenic stimulation. It is possible that the immunization regimen used in our study, involving prime and boost, led to CD8 cytotoxic cell fatigue, potentially explaining the relatively weak specific responses observed just before challenge. Additionally, viruses can evade the host immune system by down-regulating MHC class I expression in infected cells, hindering T-cell recognition of viral antigens. Although this evasion mechanism has not been confirmed during "in vivo" infections with virulent ASFV isolates, it has been demonstrated “in vitro”, implicating the viral protein EP153R [68]. Thus, the increased frequency of CD4 CTL observed after challenge in our immunized/challenged pigs may play a crucial role in combating "de novo" infection with virulent ASFV, potentially through CD4 CTL-mediated killing of infected APC, such as monocyte/macrophages, which are primary target cells for ASFV. Further elucidation of the mechanisms underlying CD4 CTL differentiation could inform the development of more effective ASFV vaccines.
5. Conclusions
Intradermal immunization of domestic pigs using a prime/boost regime with Lv17/WB/Rie1 strain conferred high levels of protection against virulent challenge with Arm07. The transient mild to moderate increase of IL-8 and IL-10 in serum observed after immunization might be directly correlated with survival. Protection was also associated with a robust ASFV-specific polyfunctional memory T cell response, where CD4CD8 and CD8 T cells were identified as the main cellular sources of virus-specific IFNγ and TNFα. In parallel to cytokine response, these T-cell subsets also showed specific cytotoxic activity as evidenced by the increased expression of the CD107a degranulation marker. Along with virus-specific multifunctional CD4CD8 and CD8 T responses, the increased levels of antigen experienced cytotoxic CD4 T cells observed after challenge in immunized pigs might also contribute to control virulent infection by killing mechanisms targeting infected antigen-presenting cells. Future studies should elucidate whether the memory T-cell responses evidenced in the present study persist and provide long-term protection against further ASFV infections.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: Flow cytometry gating strategy used for phenotyping and to interrogate immune responses.
Author Contributions
Conceptualization, M.P., M.A. and C.G.; methodology, M.P., A.S., A.S., N.C., C.P., M.A.G-C and P.J.S-C; formal analysis, M.P. and C.G.; investigation, M.P., A.S., P.F-P., P.J.S-C., M.A. and C.G.; writing—original draft preparation, M.P.; writing—review and editing, M.P., P.J.S-C., M.A. and C.G.; project administration, M.P., M.A. and C.G.; funding acquisition, M.A. and C.G. All authors have read and agreed to the published version of the manuscript.
Funding
The research project “VACDIVA” was funded by the European Union’s Horizon 2020 Research and Innovation Program under the grant agreement no. 862874.
Institutional Review Board Statement
The animal study was in accordance with European regulations (EC Directive 2010/63/UE) and reviewed and approved by the Spanish Ethical and Animal Welfare Committee (ethical approval reference number: PROEX/101/8.21).
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors would like to thank the animal care staff, veterinary services and Veterinary Pathology Unit at the BSL3 facilities (CISA-INIA/CSIC) for the technical assistance with the animals and during necropsies. Authors would like to thank the support received from the European project VACDIVA (EU-H2020-862874).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Ward, M.P.; Tian, K.; Nowotny, N. African Swine Fever, the forgotten pandemic. Transbound Emerg Dis 2021, 68, 2637–2639. [Google Scholar] [CrossRef] [PubMed]
- African swine fever (ASF) - Situation Report 45 - WOAH - World Organisation for Animal Health. 2024.
- Sánchez-Cordón, P.J.; Nunez, A.; Neimanis, A.; Wikström-Lassa, E.; Montoya, M.; Crooke, H.; Gavier-Widén, D. African Swine Fever: Disease Dynamics in Wild Boar Experimentally Infected with ASFV Isolates Belonging to Genotype I and II. Viruses 2019, 11, 852. [Google Scholar] [CrossRef] [PubMed]
- Gallardo, C.; Soler, A.; Nurmoja, I.; Cano-Gómez, C.; Cvetkova, S.; Frant, M.; Woźniakowski, G.; Simón, A.; Pérez, C.; Nieto, R.; Arias, M. Dynamics of African swine fever virus (ASFV) infection in domestic pigs infected with virulent, moderate virulent and attenuated genotype II ASFV European isolates. Transbound Emerg Dis 2021, 68, 2826–2841. [Google Scholar] [CrossRef] [PubMed]
- Radulovic, E.; Mehinagic, K.; Wüthrich, T.; Hilty, M.; Posthaus, H.; Summerfield, A.; Ruggli, N.; Benarafa, C. The baseline immunological and hygienic status of pigs impact disease severity of African swine fever. PLoS Pathog 2022, 18, e1010522. [Google Scholar] [CrossRef]
- Blome, S.; Gabriel, C.; Beer, M. Pathogenesis of African swine fever in domestic pigs and European wild boar. Virus Res 2013, 173, 122–30. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Cordón, P.J.; Montoya, M.; Reis, A.L.; Dixon, L.K. African swine fever: A re-emerging viral disease threatening the global pig industry. Vet J 2018, 233, 41–48. [Google Scholar] [CrossRef]
- Rock, D.L. Thoughts on African Swine Fever Vaccines. Viruses 2021, 13, 943. [Google Scholar] [CrossRef]
- Onisk, D.V.; Borca, M.V.; Kutish, G.; Kramer, E.; Irusta, P.; Rock, D.L. Passively transferred African swine fever virus antibodies protect swine against lethal infection. Virology 1994, 198, 350–4. [Google Scholar] [CrossRef]
- Oura, C.A.L.; Denyer, M.S.; Takamatsu, H.; Parkhouse, R.M.E. In vivo depletion of CD8+ T lymphocytes abrogates protective immunity to African swine fever virus. J Gen Virol 2005, 86, 2445–2450. [Google Scholar] [CrossRef]
- King, K.; Chapman, D.; Argilaguet, J.M.; Fishbourne, E.; Hutet, E.; Cariolet, R.; Hutchings, G.; Oura, C.A.; Netherton, C.L.; Moffat, K.; Taylor, G.; Le Potier, M.F.; Dixon, L.K.; Takamatsu, H.H. Protection of European domestic pigs from virulent African isolates of African swine fever virus by experimental immunisation. Vaccine 2011, 29, 4593–600. [Google Scholar] [CrossRef]
- Takamatsu, H.H.; Denyer, M.S.; Lacasta, A.; Stirling, C.M.; Argilaguet, J.M.; Netherton, C.L.; Oura, C.A.; Martins, C.; Rodríguez, F. Cellular immunity in ASFV responses. Virus Res 2013, 173, 110–121. [Google Scholar] [CrossRef]
- Argilaguet, J.M.; Pérez-Martín, E.; López, S.; Goethe, M.; Escribano, J.M.; Giesow, K.; Keil, G.M.; Rodríguez, F. BacMam immunization partially protects pigs against sublethal challenge with African swine fever virus. Antiviral Res 2013, 98, 61–5. [Google Scholar] [CrossRef]
- Lacasta, A.; Ballester, M.; Monteagudo, P.L.; Rodríguez, J.M.; Salas, M.L.; Accensi, F.; Pina-Pedrero, S.; Bensaid, A.; Argilaguet, J.; López-Soria, S.; Hutet, E.; Le Potier, M.F.; Rodríguez, F. Expression library immunization can confer protection against lethal challenge with African swine fever virus. J Virol 2014, 88, 13322–32. [Google Scholar] [CrossRef]
- Schäfer, A.; Franzoni, G.; Netherton, C.L.; Hartmann, L.; Blome, S.; Blohm, U. Adaptive Cellular Immunity against African Swine Fever Virus Infections. Pathogens 2022, 11, 274. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Chen, W.; Jiang, C.; Zhang, X.; Sun, Y.; Liu, R.; Wang, J.; Yang, D.; Zhao, D.; Bu, Z.; He, X. Host Responses to Live-Attenuated ASFV (HLJ/18-7GD). Viruses 2022, 14, 2003. [Google Scholar] [CrossRef] [PubMed]
- Arias, M.; Jurado, C.; Gallardo, C.; Fernández-Pinero, J.; Sánchez-Vizcaíno, J.M. Gaps in African swine fever: Analysis and priorities. Transbound Emerg Dis 2018, 65 Suppl 1, 235–247. [Google Scholar] [CrossRef]
- Gladue, D.P.; Borca, M.V. Recombinant ASF Live Attenuated Virus Strains as Experimental Vaccine Candidates. Viruses 2022, 14, 878. [Google Scholar] [CrossRef] [PubMed]
- Lacasta, A.; Monteagudo, P.L.; Jiménez-Marín, Á.; Accensi, F.; Ballester, M.; Argilaguet, J.; Galindo-Cardiel, I.; Segalés, J.; Salas, M.L.; Domínguez, J.; Moreno, Á.; Garrido, J.J.; Rodríguez, F. Live attenuated African swine fever viruses as ideal tools to dissect the mechanisms involved in viral pathogenesis and immune protection. Vet Res 2015, 46, 135. [Google Scholar] [CrossRef]
- Mulumba-Mfumu, L.K.; Goatley, L.C.; Saegerman, C.; Takamatsu, H.H.; Dixon, L.K. Immunization of African Indigenous Pigs with Attenuated Genotype I African Swine Fever Virus OURT88/3 Induces Protection Against Challenge with Virulent Strains of Genotype I. Transbound Emerg Dis 2016, 63, e323–7. [Google Scholar] [CrossRef]
- Sánchez-Cordón, P.J.; Jabbar, T.; Berrezaie, M.; Chapman, D.; Reis, A.; Sastre, P.; Rueda, P.; Goatley, L.; Dixon, L.K. Evaluation of protection induced by immunisation of domestic pigs with deletion mutant African swine fever virus BeninΔMGF by different doses and routes. Vaccine 2018, 36, 707–715. [Google Scholar] [CrossRef]
- Barasona, J.A.; Gallardo, C.; Cadenas-Fernández, E.; Jurado, C.; Rivera, B.; Rodríguez-Bertos, A.; Arias, M.; Sánchez-Vizcaíno, J.M. First Oral Vaccination of Eurasian Wild Boar Against African Swine Fever Virus Genotype II. Front Vet Science 2019, 6, 137. [Google Scholar] [CrossRef]
- Barasona, J.A.; Cadenas-Fernández, E.; Kosowska, A.; Barroso-Arévalo, S.; Rivera, B.; Sánchez, R.; Porras, N.; Gallardo, C.; Sánchez-Vizcaíno, J.M. Safety of African Swine Fever Vaccine Candidate Lv17/WB/Rie1 in Wild Boar: Overdose and Repeated Doses. Front Immunol 2021, 12, 761753. [Google Scholar] [CrossRef]
- Gallardo, C.; Soler, A.; Rodze, I.; Nieto, R.; Cano-Gómez, C.; Fernandez-Pinero, J.; Arias, M. Attenuated and non-haemadsorbing (non-HAD) genotype II African swine fever virus (ASFV) isolated in Europe, Latvia 2017. Transbound Emerg Dis 2019, 66, 1399–1404. [Google Scholar] [CrossRef]
- Franzoni, G.; Petrini, S.; Mészáros, I.; Dei Giudici, S.; Righi, C.; Olasz, F.; Zinellu, S.; Tamás, V.; Pela, M.; Gallardo, C.; Zádori, Z.; Oggiano, A.; Feliziani, F. Evaluation of Haematological and Immunological Parameters of the ASFV Lv17/WB/Rie1 Strain and Its Derived Mutant Lv17/WB/Rie1/d110-11L against ASFV Challenge Infection in Domestic Pigs. Vaccines 2023, 11, 1277. [Google Scholar] [CrossRef]
- Tamás, V.; Righi, C.; Mészáros, I.; D'Errico, F.; Olasz, F.; Casciari, C.; Zádori, Z.; Magyar, T.; Petrini, S.; Feliziani, F. Involvement of the MGF 110-11L Gene in the African Swine Fever Replication and Virulence. Vaccines 2023, 11, 846. [Google Scholar] [CrossRef]
- Pérez-Núñez, D.; Castillo-Rosa, E.; Vigara-Astillero, G.; García-Belmonte, R.; Gallardo, C.; Revilla, Y. Identification and Isolation of Two Different Subpopulations Within African Swine Fever Virus Arm/07 Stock. Vaccines 2020, 8, 625. [Google Scholar] [CrossRef]
- Carrascosa, A.L.; Bustos, M.J.; de Leon, P. Methods for growing and titrating African swine fever virus: field and laboratory samples. Curr Protoc Cell Biol 2011, Chapter 26, 14.1-26.14.25. [CrossRef]
- Carmina, G.; Nieto, R.; Arias, M. Indirect Immunoperoxidase Test (IPT) for Detection of Antibodies Against African Swine Fever Virus (ASFV) on African Green Monkey Cell Lines (Vero, MS). Methods Mol Biol 2022, 2503, 147–158. [Google Scholar] [CrossRef]
- Gallardo, C.; Soler, A.; Nieto, R.; Sánchez, M.A; Martins, C.; Pelayo, V.; Carrascosa, A.; Revilla, Y.; Simón, A. , Briones, V.; Sánchez-Vizcaíno, J.M.; Arias, M. Experimental Transmission of African Swine Fever (ASF) Low Virulent Isolate NH/P68 by Surviving Pigs. Transbound Emerg Dis 2015, 62, 612-22. [CrossRef]
- Gallardo, C.; Soler, A.; Nieto, R.; Cano, C.; Pelayo, V.; Sánchez, M.A.; Pridotkas, G.; Fernandez-Pinero, J.; Briones, V.; Arias, M. Experimental Infection of Domestic Pigs with African Swine Fever Virus Lithuania 2014 Genotype II Field Isolate. Transbound Emerg Dis 2017, 64, 300–304. [Google Scholar] [CrossRef]
- Sánchez-Cordón, P.J.; Floyd, T.; Hicks, D.; Crooke, H.R.; McCleary, S.; McCarthy, R.R.; Strong, R. , Dixon, L.K.; Neimanis, A.; Wikström-Lassa, E.; Gavier-Widén, D.; Núñez, A. Evaluation of Lesions and Viral Antigen Distribution in Domestic Pigs Inoculated Intranasally with African Swine Fever Virus Ken05/Tk1 (Genotype X). Pathogens 2021, 10, 768. [CrossRef]
- Pedrera, M.; Macchi, F.; McLean, R.K.; Franceschi, V.; Thakur, N.; Russo, L.; Medfai, L.; Todd, S.; Tchilian, E.Z.; Audonnet, J.C.; Chappell, K.; Isaacs, A. , Watterson, D.; Young, P.R.; Marsh, G.A.; Bailey, D.; Graham, S.P.; Donofrio, G. Bovine Herpesvirus-4-Vectored Delivery of Nipah Virus Glycoproteins Enhances T Cell Immunogenicity in Pigs. Vaccines 2020, 8, 115. [CrossRef]
- Terrestrial Manual Online Access - WOAH - World Organisation for Animal Health. 2023.
- Fernández-Pinero, J.; Gallardo, C.; Elizalde, M.; Robles, A.; Gómez, C.; Bishop, R.; Heath, L.; Couacy-Hymann, E.; Fasina, F.O.; Pelayo, V.; Soler, A.; Arias, M. Molecular diagnosis of African Swine Fever by a new real-time PCR using universal probe library. Transbound Emerg Dis 2013, 60, 48–58. [Google Scholar] [CrossRef]
- Franzoni, G.; Kurkure, N.V.; Essler, S.E.; Pedrera, M.; Everett, H.E; Bodman-Smith, K.B.; Crooke, H.R.; Graham, S.P. Proteome-wide screening reveals immunodominance in the CD8 T cell response against classical swine fever virus with antigen-specificity dependent on MHC class I haplotype expression. PloS one 2013, 8, e84246. [Google Scholar] [CrossRef]
- Mokhtar, H.; Pedrera, M. ; Frossard. J.P.; Biffar, L.; Hammer, S.E.; Kvisgaard, L.K.; Larsen, L.E.; Stewart, G.R.; Somavarapu, S.; Steinbach, F.; Graham, S.P. The Non-structural Protein 5 and Matrix Protein Are Antigenic Targets of T Cell Immunity to Genotype 1 Porcine Reproductive and Respiratory Syndrome Viruses. Front Immunol 2016, 7, 40. [Google Scholar] [CrossRef]
- Franzoni, G.; Kurkure, N.V.; Edgar, D.S.; Everett, H.E.; Gerner, W.; Bodman-Smith, K.B.; Crooke, H.R.; Graham, S.P. Assessment of the phenotype and functionality of porcine CD8 T cell responses following vaccination with live attenuated classical swine fever virus (CSFV) and virulent CSFV challenge. Clin Vaccine Immunol 2013, 20, 1604–16. [Google Scholar] [CrossRef] [PubMed]
- Attreed, S.E.; Silva, C. , Abbott, S.; Ramirez-Medina, E.; Espinoza, N.; Borca, M.V.; Gladue, D.P.; Diaz-San Segundo, F. A Highly Effective African Swine Fever Virus Vaccine Elicits a Memory T Cell Response in Vaccinated Swine. Pathogens 2022, 11, 1438. [CrossRef]
- Bosch-Camós, L.; Alonso, U.; Esteve-Codina, A.; Chang, C.Y.; Martín-Mur, B.; Accensi, F.; Muñoz, M.; Navas, M.J.; Dabad, M.; Vidal, E.; Pina-Pedrero, S.; Pleguezuelos, P.; Caratù, G.; Salas, M.L.; Liu, L.; Bataklieva, S.; Gavrilov, B.; Rodríguez, F.; Argilaguet, J. Cross-protection against African swine fever virus upon intranasal vaccination is associated with an adaptive-innate immune crosstalk. PLoS pathog 2022, 18, e1010931. [Google Scholar] [CrossRef] [PubMed]
- Goatley, L.C.; Nash, R.H.; Andrews, C.; Hargreaves, Z.; Tng, P.; Reis, A.L.; Graham, S.P.; Netherton, C.L. Cellular and Humoral Immune Responses after Immunisation with Low Virulent African Swine Fever Virus in the Large White Inbred Babraham Line and Outbred Domestic Pigs. Viruses 2022, 14, 1487. [Google Scholar] [CrossRef]
- Sugimura, T.; Ito, Y.; Tananari, Y.; Ozaki, Y.; Maeno, Y.; Yamaoka, T.; Kudo, Y. Improved antibody responses in infants less than 1 year old using intradermal influenza vaccination. Vaccine 2008, 26, 2700–5. [Google Scholar] [CrossRef]
- Soonawala, D.; Verdijk, P.; Wijmenga-Monsuur, A.J.; Boog, C.J.; Koedam, P.; Visser, L.G.; Rots, N.Y. Intradermal fractional booster dose of inactivated poliomyelitis vaccine with a jet injector in healthy adults. Vaccine 2013, 31, 3688–94. [Google Scholar] [CrossRef] [PubMed]
- Beals, C.R.; Railkar, R.A.; Schaeffer, A.K.; Levin, Y.; Kochba, E.; Meyer, B.K.; Evans, R.K.; Sheldon, E.A.; Lasseter, K.; Lang, N.; Weinberg, A.; Canniff, J.; Levin, M.J. Immune response and reactogenicity of intradermal administration versus subcutaneous administration of varicella-zoster virus vaccine: an exploratory, randomised, partly blinded trial. Lancet Infect Dis 2016, 16, 915–22. [Google Scholar] [CrossRef]
- Schnyder, J.L.; Garcia Garrido, H.M.; De Pijper, C.A.; Daams, J.G.; Stijnis, C.; Goorhuis, A.; Grobusch, M.P. Comparison of equivalent fractional vaccine doses delivered by intradermal and intramuscular or subcutaneous routes: A systematic review. Travel Med Infect Dis 2021, 41, 102007. [Google Scholar] [CrossRef]
- Franzoni, G.; Graham, S.P.; Dei Giudici, S.; Oggiano, A. Porcine Dendritic Cells and Viruses: An Update. Viruses 2019, 11, 445. [Google Scholar] [CrossRef]
- Leitão, A.; Cartaxeiro, C.; Coelho, R.; Cruz, B.; Parkhouse, R.M.E.; Portugal, F.C.; Vigário, J.D.; Martins, C.L.V. The non-haemadsorbing African swine fever virus isolate ASFV/NH/P68 provides a model for defining the protective anti-virus immune response. J Gen Virol 2001, 82, 513–523. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhao, Z.; Wang, Y.; Wu, S.; Wang, B.; Zhang, J.; Song, X.; Chen, Y.; Lv, P.; Hou, L. Comparative immunogenicity analysis of intradermal versus intramuscular immunization with a recombinant human adenovirus type 5 vaccine against Ebola virus. Front Immunol 2022, 13, 963049. [Google Scholar] [CrossRef] [PubMed]
- Salguero, F.J.; Ruiz-Villamor, E.; Bautista, M.J.; Sánchez-Cordón, P.J.; Carrasco, L.; Gómez-Villamandos, J.C. Changes in macrophages in spleen and lymph nodes during acute African swine fever: expression of cytokines. Vet Immunol Immunopathol 2002, 90, 11–22. [Google Scholar] [CrossRef]
- Wang, S.; Zhang, J.; Zhang, Y.; Yang, J.; Wang, L.; Qi, Y.; Han, X.; Zhou, X.; Miao, F.; Chen, T.; Wang, Y.; Zhang, F.; Zhang, S.; Hu, R. Cytokine Storm in Domestic Pigs Induced by Infection of Virulent African Swine Fever Virus. Front Vet Sci 2021, 7, 601641. [Google Scholar] [CrossRef]
- Zakaryan, H.; Cholakyans, V.; Simonyan, L.; Misakyan, A.; Karalova, E.; Chavushyan, A.; Karalyan, Z. A study of lymphoid organs and serum proinflammatory cytokines in pigs infected with African swine fever virus genotype II. Arch Virol 2015, 160, 1407–14. [Google Scholar] [CrossRef]
- Franzoni, G.; Pedrera, M.; Sánchez-Cordón, P.J. African Swine Fever Virus Infection and Cytokine Response In Vivo: An Update. Viruses 2023, 15, 233. [Google Scholar] [CrossRef]
- Barroso-Arévalo, S.; Barasona, J.A.; Cadenas-Fernández, E.; Sánchez-Vizcaíno, J.M. The Role of Interleukine-10 and Interferon-γ as Potential Markers of the Evolution of African Swine Fever Virus Infection in Wild Boar. Pathogens 2021, 10, 757. [Google Scholar] [CrossRef]
- Sánchez-Cordón, P.J.; Jabbar, T.; Chapman, D.; Dixon, L.K.; Montoya, M. Absence of Long-Term Protection in Domestic Pigs Immunized with Attenuated African Swine Fever Virus Isolate OURT88/3 or BeninΔMGF Correlates with Increased Levels of Regulatory T Cells and Interleukin-10. J Virol 2020, 94, e00350–20. [Google Scholar] [CrossRef] [PubMed]
- Gómez del Moral, M.; Ortuño, E.; Fernández-Zapatero, P.; Alonso, F.; Alonso, C.; Ezquerra, A.; Domínguez, J. African swine fever virus infection induces tumor necrosis factor alpha production: implications in pathogenesis. J Virol 1999, 73, 2173–80. [Google Scholar] [CrossRef] [PubMed]
- Carlson, J.; O'Donnell, V.; Alfano, M.; Velazquez Salinas, L.; Holinka, L.G.; Krug, P.W.; Gladue, D.P.; Higgs, S.; Borca, M.V. Association of the Host Immune Response with Protection Using a Live Attenuated African Swine Fever Virus Model. Viruses 2016, 8, 291. [Google Scholar] [CrossRef]
- Sánchez-Cordón, P.J.; Chapman, D.; Jabbar, T.; Reis, A.L.; Goatley, L.; Netherton, C.L.; Taylor, G.; Montoya, M.; Dixon, L. Different routes and doses influence protection in pigs immunised with the naturally attenuated African swine fever virus isolate OURT88/3. Antiviral Res 2017, 138, 1–8. [Google Scholar] [CrossRef]
- Saalmüller, A.; Werner, T.; Fachinger, V. T-helper cells from naive to committed. Vet Immunol Immunopathol 2002, 87, 137–45. [Google Scholar] [CrossRef]
- Gerner, W.; Käser, T.; Saalmüller, A. Porcine T lymphocytes and NK cells-an update. Dev Comp Immunol 2009, 33, 310–20. [Google Scholar] [CrossRef]
- Revilla, Y.; Pena, L.; Viñuela, E. Interferon-gamma production by African swine fever virus-specific lymphocytes. Scand J Immunol 1992, 35, 225–30. [Google Scholar] [CrossRef]
- Reis, A.L.; Goatley, L.C.; Jabbar, T.; Sanchez-Cordon, P.J.; Netherton, C.L.; Chapman, D.A.G.; Dixon, L.K. Deletion of the African Swine Fever Virus Gene DP148R Does Not Reduce Virus Replication in Culture but Reduces Virus Virulence in Pigs and Induces High Levels of Protection against Challenge. J Virol 2017, 91, e01428–17. [Google Scholar] [CrossRef] [PubMed]
- Denyer, M.S.; Wileman, T.E.; Stirling, C.M.; Zuber, B.; Takamatsu, H.H. Perforin expression can define CD8 positive lymphocyte subsets in pigs allowing phenotypic and functional analysis of natural killer, cytotoxic T, natural killer T and MHC un-restricted cytotoxic T-cells. Vet Immunol Immunopathol 2006, 110, 279–92. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Díaz, I.; Martín-Valls, G.; Beyersdorf, N. , Mateu, E. Systemic CD4 cytotoxic T cells improve protection against PRRSV-1 transplacental infection. Front Immunol 2023, 13, 1020227. [CrossRef]
- Talker, S.C.; Koinig, H.C.; Stadler, M.; Graage, R.; Klingler, E.; Ladinig, A.; Mair, K.H.; Hammer, S.E.; Weissenböck, H.; Dürrwald, R.; Ritzmann, M.; Saalmüller, A.; Gerner, W. Magnitude and kinetics of multifunctional CD4+ and CD8β+ T cells in pigs infected with swine influenza A virus. Vet Res 2015, 46, 52. [Google Scholar] [CrossRef] [PubMed]
- Schäfer, A.; Zani, L.; Pikalo, J.; Hühr, J.; Sehl, J.; Mettenleiter, T.C.; Breithaupt, A.; Blome, S.; Blohm, U. T-cell responses in domestic pigs and wild boar upon infection with the moderately virulent African swine fever virus strain 'Estonia2014'. Transbound Emerg Dis 2021, 68, 2733–2749. [Google Scholar] [CrossRef] [PubMed]
- Chung, C.J.; Cha, S.H.; Grimm, A.L.; Ajithdoss, D.; Rzepka, J.; Chung, G.; Yu, J.; Davis, W.C.; Ho, C.S. Pigs that recover from porcine reproduction and respiratory syndrome virus infection develop cytotoxic CD4+CD8+ and CD4+CD8- T-cells that kill virus infected cells. PloS one 2018, 13, e0203482. [Google Scholar] [CrossRef] [PubMed]
- Preglej, T.; Ellmeier, W. CD4+ Cytotoxic T cells - Phenotype, Function and Transcriptional Networks Controlling Their Differentiation Pathways. Immunol Lett 2022, 247, 27–42. [Google Scholar] [CrossRef] [PubMed]
- Hurtado, C.; Bustos, M.J.; Granja, A.G.; de León, P.; Sabina, P.; López-Viñas, E.; Gómez-Puertas, P.; Revilla, Y.; Carrascosa, A.L. The African swine fever virus lectin EP153R modulates the surface membrane expression of MHC class I antigens. Arch Virol 2011, 156, 219–34. [Google Scholar] [CrossRef]
Figure 3.
At selected days post-immunization (dpi) with Lv7/WB/Rie1 on 0 (prime) and 21 (boost) dpi and challenge with Armenia07 at 35 dpi (0, 7, 21, 28, 35 and 42 dpi), resuscitated PBMC were stimulated in vitro with medium or ASFV (Lv7/WB/Rie1 or Armenia07) or mock virus supernatant. Phenotype and frequencies (Freq. of live) were assessed by flow cytometry. T cell populations were defined as CD3+ (CD3 T cell), CD3+CD4negCD8high (cytotoxic CD8 T cell), CD3+CD4+CD8low (activated/memory CD4CD8 T cell) and CD3+CD4+CD8neg (helper CD4 T cell). (A) Data of cellular populations on live, singlet lymphocytes after medium stimulation. Spontaneous/basal responses to medium in the immunized pigs. Floating bars of individual data and line at mean from three pigs are shown for each time point. (B) Data of cellular populations on live, singlet lymphocytes after virus stimulation and corrected against mock. Responses to Lv7/WB/Rie1 (red dots and box) and Armenia07 (blue dots and box) in the immunized pigs. Individual and mean data ± SD from three pigs are shown for each time point. Significance is indicated by: *(p<0.05), **(p<0.01), ***(p<0.001) and ****(p<0.0001).
Figure 3.
At selected days post-immunization (dpi) with Lv7/WB/Rie1 on 0 (prime) and 21 (boost) dpi and challenge with Armenia07 at 35 dpi (0, 7, 21, 28, 35 and 42 dpi), resuscitated PBMC were stimulated in vitro with medium or ASFV (Lv7/WB/Rie1 or Armenia07) or mock virus supernatant. Phenotype and frequencies (Freq. of live) were assessed by flow cytometry. T cell populations were defined as CD3+ (CD3 T cell), CD3+CD4negCD8high (cytotoxic CD8 T cell), CD3+CD4+CD8low (activated/memory CD4CD8 T cell) and CD3+CD4+CD8neg (helper CD4 T cell). (A) Data of cellular populations on live, singlet lymphocytes after medium stimulation. Spontaneous/basal responses to medium in the immunized pigs. Floating bars of individual data and line at mean from three pigs are shown for each time point. (B) Data of cellular populations on live, singlet lymphocytes after virus stimulation and corrected against mock. Responses to Lv7/WB/Rie1 (red dots and box) and Armenia07 (blue dots and box) in the immunized pigs. Individual and mean data ± SD from three pigs are shown for each time point. Significance is indicated by: *(p<0.05), **(p<0.01), ***(p<0.001) and ****(p<0.0001).
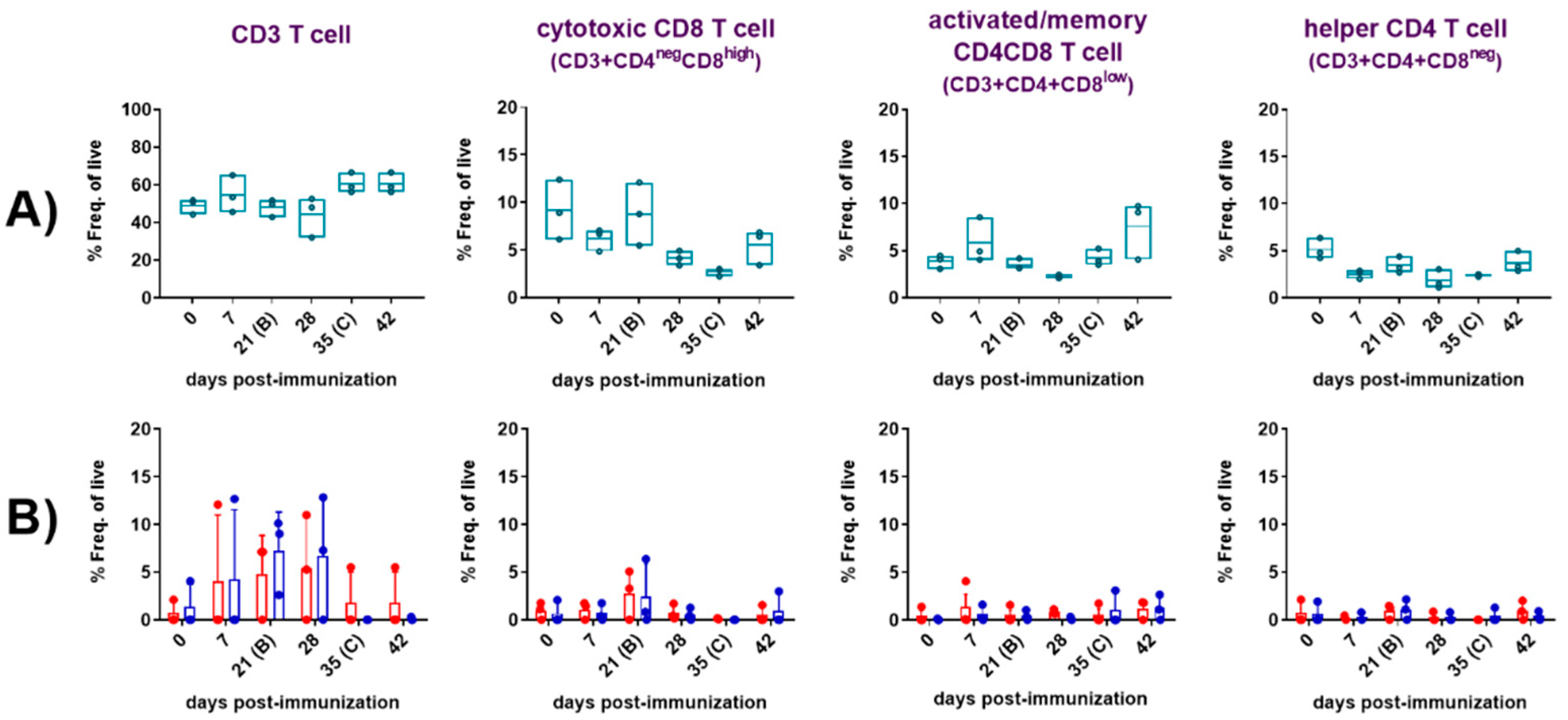

Table 1.
ASFV detection in tissues determined by real-time PCR in domestic pigs intradermally immunized with the Lv17/WB/Rie1. In grey it is indicated the positive virus isolation (VI) results after three passages in porcine blood leukocytes (PBL) in absence of haemadsorption (HAD). In red is indicated the positive HAD result indicating the presence of Arm07 ASFV.
Table 1.
ASFV detection in tissues determined by real-time PCR in domestic pigs intradermally immunized with the Lv17/WB/Rie1. In grey it is indicated the positive virus isolation (VI) results after three passages in porcine blood leukocytes (PBL) in absence of haemadsorption (HAD). In red is indicated the positive HAD result indicating the presence of Arm07 ASFV.
| ID domestic pig /Tissue | Pig 6 (D65/30) | Pig 8 (D65/30) |
Pig 9 (D64/29) |
Pig 10 (D64/29) |
|---|---|---|---|---|
| Liver | No Ct | No Ct | No Ct | No Ct |
| Lung | No Ct | No Ct | 33.7 | 35.7 |
| Kidney | 36.98 | No Ct | No Ct | No Ct |
| Heart | No Ct | No Ct | 37.5 | 36.8 |
| Spleen | No Ct | No Ct | No Ct | No Ct |
| Tonsil | No Ct | 37.8 | No Ct | 34 |
| Renal LN* | 39.05 | No Ct | No Ct | No Ct |
| Retropharyngeal LN | No Ct | No Ct | No Ct | No Ct |
| Gastro-hepatic LN | No Ct | No Ct | No Ct | 34.6 |
| Mesenteric LN | No Ct | No Ct | No Ct | 39.2 |
| Mediastinal LN | 33.93 | No Ct | No Ct | 37.8 |
| Inguinal LN | No Ct | No Ct | 38.4 | No Ct |
| Submandibular LN | No Ct | No Ct | No Ct | 36 |
| Splenic LN | No Ct | No Ct | No Ct | 35.3 |
| Popliteal LN | 39.65 | No Ct | 39.6 | 36.3 |
| Bone marrow | No Ct | No Ct | No Ct | No Ct |
| Diaphragm | 39.95 | No Ct | 37.8 | 32.3 |
| Front left IA** | 35.85 | No Ct | No Ct | 34.2 |
| Front right IA | No Ct | No Ct | No Ct | 29.8 |
| Back left IA | No Ct | No Ct | No Ct | 36 |
| Back right IA | No Ct | No Ct | No Ct | 37.3 |
| TOTAL PCR POS. | 6/21 (28.5%) |
1/21 (4,8%) |
5/21 (23.8%) |
14/21 (66.6%) |
| TOTAL VI POS. | 1/21 (4.8%) |
0/21 (0%) |
0/21 (0%) |
5/21 (23.8%) |
*Lymph node; ** articular cartilage.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



