
Submitted:
08 March 2024
Posted:
11 March 2024
You are already at the latest version
Abstract
The Australian tropical montane cloud forest (TMCF) is home to approximately 18 percent of the nation’s total vascular plant species. Over the past century, human activity and industrial development have caused global climate changes, posing a severe and irreversible danger to the entire land-based ecosystem, and WTWHA is no exception. The current average annual temperature of WTWHA in FNQ is 24 °C. However, in the coming years (by 2030), the average annual temperature increase is estimated to be between 0.5 and 1.4 °C compared to the climate observed between 1986 and 2005. Looking further ahead to 2070, the anticipated temperature rise is projected to be between 1.0 and 3.2 °C, with the exact range depending on future emissions. We identified 84 plant species endemic to the WTWHA in FNQ, which are already experiencing climate change threats. Few of these plants are used in herbal medicines. This study comprehensively reviewed metabolomics studies conducted on these 84 plant species until now toward understanding their physiological and metabolomics responses to global climate change. This review also discusses: i) recent developments in plant metabolomics studies that can be applied to study and understand the interactions of wet tropical plants with climatic stress, ii) medicinal plants and isolated phytochemicals with structural diversity, and iii) reported biological activities of crude extracts and isolated compounds.
Keywords:
wet tropics
; climate change
; metabolomics
; tropical montane cloud forest
; Wet Tropics World Heritage Area
; plant secondary metabolites
1. Introduction
Human activity and industrial development have led to significant and irreversible threats to the entire land-based ecosystem in the last century, primarily due to global climate changes. As the average global annual temperature continues to rise by 1℃ compared to the average temperature during the preindustrial era, and experts have predicted that the temperature will further rise by 3–5 ℃ by the end of this century, owing to the increasing concentration of greenhouse gases (GHGs), such as CO2 and methane in the atmosphere [1,2]. Looking further ahead to 2070, the anticipated temperature rise ranges from 1.0 to 3.2 °C, based on the intensity of future greenhouse gas emissions [3]. Climate change indirectly impacts species by diminishing the quantity and accessibility of habitat and eliminating species crucial for the survival of the species in question [4]. This impact is significantly felt in the Tropical montane (TM) regions within the evergreen forests that are enveloped in persistent and frequent low-level clouds form unique ecosystems called TM cloud forest (TMCF) [5]. Indeed, recent studies including climate model studies [6,7,8], predicted higher rates of temperature increase at higher elevations of TMCF regions compared to lower elevations. Furthermore, alterations in reliability and quantity of precipitation in TMCF are anticipated due to reductions in cloud cover [5,9,10,11]. These factors would bring significant challenges to species in TMCF, leading to shifts in altitudinal ranges, reshuffling of species compositions, and increased risk of extinction [5].
The Australian TMCF situated at or above 900 meters in elevation (Figure 1) across Wet Tropics World Heritage Area (WTWHA) of northeast Queensland, Australia, is no exception. Indeed, the impacts of climate change on Australian TMCF are anticipated to manifest within this century [7,12] as the plant species are particularly vulnerable to climatic stress compared to the lowland species [7,13]. The WTWHA, which is considered the sixth most important protected area globally for conserving biodiversity [14], is home to over 3,300 plant species, of which 700+ are endemic to the region, accounting for 18 percent of the nation’s total vascular plant species [15,16]. The ongoing project on climate-affected TMCF plants, which is led by the Australian Tropical Herbarium at James Cook University, has identified 84 plant species that are vulnerable to the impact of climate change in the WTWHA. Some of these plants are used in traditional medicines, including Aboriginal bush medicines.
These plants that are affected by climate change or biotic stressors exhibit both morphological and physiological plasticity in order to adapt, survive, and thrive [17]. Their adaptability relies on complex genetic or metabolic detection and communication systems that we are just starting to comprehend. Numerous research investigations have been carried out on various plants, for instance, Arabidopsis, aiming to unravel the intricate molecular mechanisms that plants exhibit in response to constantly changing environments [18,19,20,21,22]. More recently, technological advancements have enabled the acquisition of molecular data, including phenomics, epigenomics, transcriptomics, proteomics, and metabolomics data. These multidisciplinary approaches have enabled a better understanding of plants' responses to various environmental changes linked to climate change, including drought and cold [23,24]. The plant produces various biomolecules, which serve different biological roles throughout plant life cycles. These biomolecules can be categorized under two major groups: primary metabolites (PMs) and secondary metabolites (SMs). Primary metabolites, such as proteins, sugars, and organic acids, are widely recognized for their role in essential plant physiological processes, including photosynthesis, photorespiration, and the tricarboxylic acid cycle [25,26]. Secondary metabolites, mainly phenolics, terpenoids, and alkaloids, are produced in response to competitive environmental factors for survival and fulfilling various physiological functions [27]. These SMs do not play a direct role in plants' typical growth and survival; instead, they contribute to plant development and enhance resilience to stress [28]. For example, flavonoids are photo-protectants, shielding plants from damage caused by ultraviolet-B (UV-B) radiation [29,30]. Similarly, some terpenoids or alkaloids might act as antioxidants or osmotic regulators in response to abiotic stresses in plants [31,32]. In contrast, glucosinolates, limited to specific taxonomic groups, are considered essential antitoxins that play a crucial role in enabling plants to resist insect attacks [33].
Understanding the response patterns of these secondary metabolites to potential global climate changes is crucial due to their significance in plant growth, resistance, human health, and conservation efforts. Recently developed powerful omics techniques [34,35,36] and bioimaging [37] and biosensor tools [38,39] have been widely applied for understanding plant physiology, analyzing the plant metabolome, and discovering novel metabolomic pathways in response to the changing environment [40]. However, there is no comprehensive review that examined recent metabolomic and phytochemical studies on climate-affected plants of Australian TMCF.
This review comprehensively analysed the literature on the available metabolomics studies conducted on the 84 TMCF plant species and discusses i) recent developments in plant metabolomics studies that can be applied to study and understand the interactions of wet tropical plants with climatic stress, ii) medicinal plants and isolated phytochemicals with structural diversity, and iii) reported biological activities of crude extracts and isolated compounds. In doing so, we compiled and listed the plant species largely restricted to TMCF based on Hoyle et al. (2023) [5] and additional information from several sources: i) records in the Atlas of Living Australia [41], ii) the ‘Rainforest Key’ [42], and 3) expert knowledge [43]. We searched for studies of metabolomic profiles, medicinal plants use, phytochemical contents, and biological activities in Google Scholar, MEDLINE Ovid, Scopus, PubMed, and journal websites using the following keywords: “wet tropics climate-affected plants,” “secondary metabolites in plants,” “metabolomics studies of plants,” “phytochemical analysis of plants affected by climatic impact,” “biological activities,” and accepted plant name and their synonyms. The information we collected was analyzed and presented in tables and figures. Additionally, we utilized ChewDraw Professional software to create chemical structures, ensuring the accuracy of each structure by cross-referencing them with databases such as PubChem, ChemSpider, and HMDB databases.
2. Climate-Affected Australian Tropical Montane Cloud Forest Plants and Their Medicinal Uses
Using information from Hoyle et al. (2023) [5], expert opinion (botanists from the Australian Tropical Herbarium (ATH) at James Cook University in Cairns), and other literature, we found 84 climate-affected plant species largely restricted to TMCF. The plant names were cross-checked using the Australian Plant Census [44], WFO Plant List [45], and Australian Tropical Rainforest Plants information system [42]. Based on these 84 plant species lists, we generated information on their botanical names, taxonomy, distribution, life form, and medicinal uses of each plant (Table 1).
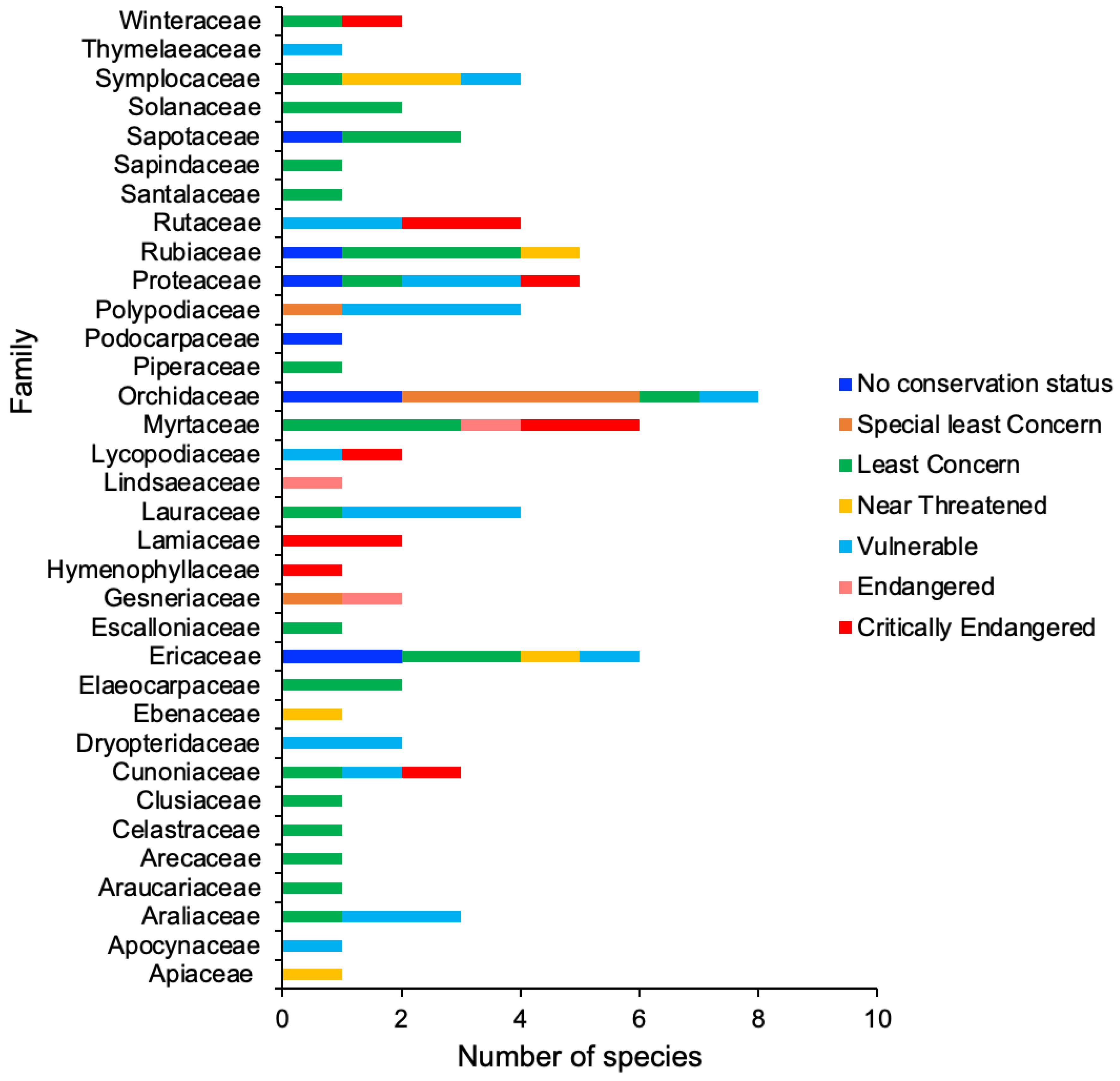
Of the 84 plant species, 54 were native to the WTWHA, 26 were endemic, and four were non-native (Table 1 and Figure 2A). Four non-native species were Parapolystichum grayi (D.J.Jones) J.J.S. Gardner & Nagalingum, Phlegmariurus creber (Alderw.) A.R.Field & Bostock, Oreogrammitis reinwardtii Blume, and Mischocarpus montanus C.T.White. Of 84 plant species, 29 were trees, followed by shrubs (28 species) and ferns (8 species) (Table 1 and Figure 2B). These 84 plant species belonged to 34 families, and the Orchidaceae family had the maximum number of species (8 species), trailed by the Ericaceae and Myrtaceae (6 species each) and Proteaceae and Rubiaceae (5 species each) (Figure 2C). Most of the families (15 families) had one species. When checked for their plant uses in traditional medicines, we found that most sTWHA plants were not used medicinally. This could be because most plants are endemic to WTWHA, and these endemic species, although used in Aboriginal bush medicines, their medicinal uses are not publicly available. Of 84 species, 43 belong to 29 medicinally important genera (Figure 2D). Of the 43 species, species of Planchonella, Tasmania, and Litsea were particularly indicated as traditionally used by Australian Aboriginal communities to treat various ailments, such as skin sores, scabies, and sore throat, as an antiseptic for boils, malaria, diarrhea, and cough (Table 1). Most of the genera were found to be used for medicinal purposes in traditional medicine systems of Asian countries, such as China, India, Indonesia, Malaysia, Japan, Taiwan, and Korea.
A report from 2010 by the Commonwealth Scientific and Industrial Research Organization (CSIRO) [46] indicates that numerous tree species are at risk of experiencing mean temperatures that exceed their typical tolerance levels. To illustrate, the recent increase in temperature may already be placing stress on a lowland tree species that has adapted to thrive within a mean annual temperature range of 23.0 to 24.0 (measured at 200 masl). To survive in a comparable temperature environment, they must relocate more than 1000 m upward by 2080. However, habitat fragmentation will severely constrain their capacity to respond in this way [16]. Figure 4 illustrates the conservation status of 84 Australian TMCF plant species affected by climate change. Of these, nearly half (41 species) are of conservation significance under Queensland State legislation: 21 species are listed as Vulnerable (V, sky blue bar), six are Near Threatened (NT, yellow bar), 3 are Endangered (E, orange bar), and 11 are Critically Endangered (CR, light red bar) (Table 1 and Figure 3). The remainder (43 species) are not currently listed as threatened, i.e., are categorized as Least Concern (LC), Special Least Concern (SL), or no conservation status indicated (No).
Through modelling analysis, Costion et al. [7] projected significant declines in suitable habitat for 19 of the 84 TMCF plant species listed in Table 1, with estimates ranging from 17% to 100% by 2040 and at least 46% by 2080. Roeble [47] further refined these predictions, modeling 37 plant species (including 8 from Costion’s study) and predicting a mean habitat loss of 63% by 2085. The study predicted that five out of 37 modelled species (Acrotriche baileyana, Gynochthodes constipata, Hymenophyllum whitei, Syzygium fratris, Tasmania sp. Mt. Bellenden Ker) will experience a total loss of their suitable habitat by 2035 and another two species (Cinnamomum propinquum and Leucopogon malayanus) by 2085 [48]. For only Bubbia whiteana was a substantial increase in the suitable habitat through 2085 predicted. Overall, both studies [7,48] suggest that a significant portion of Australian montane cloud forest species is either threatened or vulnerable to climatic stress. Hence, these plants must acclimatize and react swiftly to overcome environmental stresses or face extinction. Therefore, it is crucial to comprehend how plants react and adjust to shifts in their environment, striving to enhance their ability to withstand the challenges posed by climate change.

Table 1.
List of climate-affected Australian montane cloud forest plants: their distribution, life form, conservation status and medicinal uses.
Table 1.
List of climate-affected Australian montane cloud forest plants: their distribution, life form, conservation status and medicinal uses.
| Botanical name, family, and synonyms | Distribution | Life Form | Medicinal uses | Metabolomics profile studied | Conservation status (QLD) |
|---|---|---|---|---|---|
| Pteridophyta | |||||
| Dryopteridaceae | |||||
|
Parapolystichum grayi (D.J.Jones) J.J.S. Gardner & Nagalingum Syn. Lastreopsis grayi D.L.Jones |
Africa, the Neotropics, north-eastern Australia, Madagascar, Pacific Island, and southern Asia | Fern | NU | No |
V |
|
Parapolystichum tinarooense (Tindale) Labiak, Sundue & R.C.Moran Syn. Lastreopsis tinarooensis Tindale |
Wet Tropics region (Australia) | Fern | NU | No | V |
| Hymenophyllaceae | |||||
| Hymenophyllum whitei Goy | Wet Tropics region (Australia) | Fern | NU | No | CR |
| Lindsaeaceae | |||||
| Lindsaea terrae-reginae K.U.Kramer | Wet Tropics region (Australia) | Fern | NU | No | E |
| Lycopodiaceae | |||||
|
Phlegmariurus creber (Alderw.) A.R.Field & Bostock Syn. Huperzia crebra (Alderw.) Holub |
Wet Tropics region (Australia), PNG, Hawaii | Epiphyte | Phlegmariurus/Huperzia species are traditionally used as vermifuge, purgative, and laxative [49]. | No | CR |
|
Phlegmariurus delbrueckii (Herter) A.R.Field & Bostock Syn. Huperzia delbrueckii (Herter) Holub |
Wet Tropics region (Australia) | Epiphyte | No | V | |
| Polypodiaceae | |||||
|
Oreogrammitis albosetosa (F.M.Bailey) Parris Syn. Polypodium albosetosum F. M.Bailey |
Wet Tropics region (Australia) | Fern | NU | No | V |
|
Oreogrammitis leonardii (Parris) Parris Syn. Grammitis leonardii Parris |
Wet Tropics region (Australia) | Fern | NU | No | V |
| Oreogrammitis reinwardtii Blume | Wet Tropics region (Australia), Sri Lanka, Philippines, Papua New Guinea, Solomon Islands, Malaysia |
Fern | NU | No | V |
|
Oreogrammitis wurunuran (Parris) Parris Syn. Grammitis wurunuran Parris |
Wet Tropics region (Australia) | Fern | NU | No | SL |
|
Magnoliophyta |
|||||
| Apiaceae | |||||
| Trachymene geraniifolia F.M.Bailey | Wet Tropics region (Australia) | Herb | NU | No | NT |
| Apocynaceae | |||||
| Parsonsia bartlensis J.B.Williams | Wet Tropics region (Australia) | Climber | NU | No | V |
| Araliaceae | |||||
| Hydrocotyle miranda A.R.Bean & Henwood | Wet Tropics region (Australia) | Herb | Hydrocotyle species are used as anti-inflammatory herbs in Taiwanese folk medicines [50]. | No | V |
| Polyscias bellendenkerensis (F.M.Bailey) Philipson | Wet Tropics region (Australia) | Shrub | Polyscias species are traditionally used to treat ailments, such as malaria, obesity, and mental disorders [51]. | No | V |
| Polyscias willmottii (F.Muell.) Philipson | Wet Tropics region (Australia) | Tree | No | LC | |
| Araucariaceae | |||||
| Agathis atropurpurea B.Hyland | Australia | Tree | Agathis species are traditionally used to treat myalgia and headaches [52]. | Yes | LC |
| Arecaceae | |||||
| Linospadix apetiolatus Dowe & A.K.Irivine | Wet Tropics region (Australia) | Tree | NU | No | LC |
| Celastraceae | |||||
| Hypsophila halleyana F.Muell. | Wet Tropics region (Australia) | Shrub | NU | No | LC |
| Clusiaceae | |||||
| Garcinia brassii C.T.White | Wet Tropics region (Australia) | Tree | Infusions prepared from fruits of Garcinia species are traditionally used to treat dysentery, ulcers, and wounds [53]. | No | LC |
| Cunoniaceae | |||||
| Ceratopetalum corymbosum C.T.White | Wet Tropics region (Australia) | Tree | NU | No | V |
| Ceratopetalum hylandii Rozefelds & R.W.Barnes | Wet Tropics region (Australia) | Tree | NU | No | LC |
| Eucryphia wilkiei B.Hyland | Wet Tropics region (Australia) | Shrub | NU | Yes | CR |
| Ebenaceae | |||||
| Diospyros granitica Jessup | Wet Tropics region (Australia) | Tree | Diospyros species are used traditionally used as sedative, astringent, carminative, febrifuge, anti-hypertensive, vermifuge, antidiuretic, and to relieve constipation [54]. | No | NT |
| Elaeocarpaceae | |||||
| Elaeocarpus linsmithii Guymer | Wet Tropics region (Australia) | Tree | Elaeocarpus species are the source of popular spiritual beads (known as Rudraksha in Asia), which are used to treat various ailments, including mental/neurological disorders (stress, depression, anxiety, hypertension, epilepsy, migraine, and neuralgia), asthma, and also used as analgesic [55]. | No | LC |
| Elaeocarpus hylobroma Y.Baba & Crayn | Wet Tropics region (Australia) | Tree | No | LC | |
| Ericaceae | |||||
| Acrotriche baileyana (Domin) J.M.Powell | Wet Tropics region (Australia) | Shrub | NU | No | NT |
| Dracophyllum sayeri F.Muell | Wet Tropics region (Australia) | Tree | NU | No | V |
|
Leucopogon malayanus subsp. novoguineensis (Sleumer) Pedley Syn. Styphelia malayana subsp. novoguineensis (Sleumer) Hislop, Crayn & Puente-Lel. |
Wet Tropics region (Australia) | Shrub | NU | No | No |
|
Rhododendron lochiae F.Muell. Syn. Rhododendron notiale, Craven |
Wet Tropics region (Australia) | Shrub | Rhododendron species are used to prevent and treat many ailments, including respiratory disorders like asthma and bronchitis, dysentery, diarrhea, constipation, fever, cardiac disorders, and inflammation [56]. | No | No |
| Rhododendron viriosum Craven | Wet Tropics region (Australia) | Tree | No | LC | |
| Trochocarpa bellendenkerensis Domin | Wet Tropics region (Australia) | Tree | NU | No | LC |
| Escalloniaceae | |||||
| Polyosma reducta F.Muell. | Wet Tropics region (Australia) | Tree | NU | No | LC |
| Gesneriaceae | |||||
| Boea kinneari (F.Muell.) B.L.Burtt | Wet Tropics region (Australia) | Herb | NU | No | E |
| Lenbrassia australiana (C.T.White) G.W.Gillett | Wet Tropics region (Australia) | Shrub | NU | No | SL |
| Lamiaceae | |||||
| Prostanthera albohirta C.T.White | Mount Emerald, Wet Tropics region (Australia) | Shrub | Some Prostanthera species are used for topical applications to treat skin sores and infections [57,58]. | No | CR |
| Prostanthera athertoniana B.J.Conn & T.C.Wilson | Wet Tropics region (Australia) | Shrub | No | CR | |
| Lauraceae | |||||
| Cinnamomum propinquum F.M.Bailey | Wet Tropics region (Australia) | Tree | Cinnamomum species are most commonly used in traditional Chinese medicines to treat multiple disorders, including indigestion, microbial infections, and cough and cold [59]. | Yes | V |
| Cryptocarya bellendenkerana B.Hyland | Wet Tropics region (Australia) | Tree | NU | Yes | LC |
| Endiandra jonesii B.Hyland | Wet Tropics region (Australia) | Tree | Endiandra species are traditionally used to treat rheumatism, headache, dysentery, pulmonary disorders, and uterine tumors [60]. | No | V |
| Litsea granitica B.Hyland | Wet Tropics region (Australia) | Tree | Litsea species are used traditionally by Aboriginal communities to treat skin infections such as sores and scabies, and also used an antiseptic [61]. | No | V |
| Myrtaceae | |||||
| Leptospermum wooroonooran F.M.Bailey | Wet Tropics region (Australia) | Tree | Leptospermum species are traditionally used in Malaysia to relieve menstrual and stomach disorders [62,63]. | Yes | LC |
| Micromyrtus delicata A.R.Bean | Wet Tropics region (Australia) | Shrub | NU | No | E |
| Pilidiostigma sessile N.Snow | Wet Tropics region (Australia) | Shrub | NU | No | LC |
| Rhodamnia longisepala N.Snow & A.J.Ford | Wet Tropics region (Australia) | Shrub | Rhodamnia species are used traditionally in Indonesia to treat scars, toothache, and cough [64]. | No | CR |
| Syzygium fratris Craven | Wet Tropic region (Australia) | Shrub | NU | No | CR |
| Uromyrtus metrosideros (F.M.Bailey) A.J.Scott | Wet Tropics region (Australia) | Shrub | NU | Yes | LC |
| Orchidaceae | |||||
| Bulbophyllum lilianiae Rendle | Wet Tropics region (Australia) | Epiphyte | Bulbophyllum species are traditionally used to treat skin diseases, cardiovascular diseases, and rheumatism [65]. | No | LC |
|
Bulbophyllum wadsworthii Dockrill Syn. Oxysepala wadsworthii (Dockrill) D.L.Jones & M.A.Clem. |
Australia | Epiphyte | No | SL | |
|
Bulbophyllum windsorense B.Gray & D.L.Jones Syn. Oxysepala windsorensis (B.Gray & D.L.Jones) D.L.Jones & M.A.Clem. |
Wet Tropics region (Australia) |
Epiphyte |
No | V | |
|
Dendrobium brevicaudum D.L.Jones & M.A.Clem. Syn. Dockrillia brevicauda (D.L.Jones & M.A.Clem.) M.A.Clem. & D.L.Jones |
Wet Tropics region (Australia) | Herb, Epiphyte | Dendrobium species are used in traditional Chinese and Indian medicine systems as a source of tonic for longevity and also as an antipyretic, analgesic, astringent, and anti-inflammatory agent [66]. | No | No |
|
Dendrobium carrii Rupp & C.T.White Syn. Australorchis carrii (Rupp & C.T.White) D.L.Jones & M.A.Clem. |
Wet Tropics region (Australia) | Herb, Epiphyte | No | SL | |
|
Dendrobium finniganense D.L.Jones Syn. Thelychiton finniganensis (D.L.Jones) M.A.Clem. & D.L.Jones |
Wet Tropics region (Australia) | Herb, Epiphyte | No | SL | |
| Liparis fleckeri Nicholls | Wet Tropics region (Australia) | Lithophyte | Liparis species are traditionally used in Chinese medicine to treat inflammatory diseases, including hemoptysis, metrorrhagia, traumatic hemorrhage, and pneumonia; it is also used to stop bleeding from wounds and to detoxify snakebite [67]. | No | No |
|
Octarrhena pusilla (F.M.Bailey) M.A.Clem. & D.L.Jones Syn. Octarrhena pusilla (F.M.Bailey) Dockrill |
Wet Tropics region (Australia) | Epiphyte | NU | No | SL |
| Piperaceae | |||||
| Peperomia hunteriana P.I.Forst. | Wet Tropics region (Australia) | Herb | Peperomia species are traditionally used for treating pain and inflammation, gastric ulcers, asthma, and bacterial infections [68,69]. | No | LC |
| Podocarpaceae | |||||
|
Prumnopitys ladei (F.M.Bailey) de Laub Syn. Stachycarpus ladei (Bailey) Gaussen, Podocarpus ladei F.M.Bailey |
Endemic to Wet Tropics Australia | Tree | Fruits and bark of Prunmnopitys species are considered medicinal [70]. | Yes | No |
| Proteaceae | |||||
| Austromuellera valida B.Hyland | Endemic to Wet Tropics region | Tree | NU | No | V |
| Helicia lewisensis Foreman | Endemic to Wet Tropics region | Tree | Helicia species are used for treating mouth and skin sores and also kidney and gastric problems [71,72,73,74]. | No | V |
| Helicia recurva Foreman | Endemic to Wet Tropics region | Tree | No | No | |
|
Hollandaea porphyrocarpa A.J.Ford & P.H.Weston Syn. Hollandaea sp. Pinnacle Rock Track (P.I.Forster PIF10714) |
Endemic to Wet Tropics region | Shrub | NU | No | CR |
|
Nothorites megacarpus (A.S.George & B.Hyland) P.H.Weston & A.R.Mast Syn. Orites megacarpa A.S.George & B.Hyland |
Endemic to Wet Tropics region | Tree | NU | No | LC |
| Rubiaceae | |||||
| Aidia gyropetala A.J.Ford and Halford | Endemic to Wet Tropics region | Tree | Aidia species are used for treating body/muscle pains and pains due to gastric disorders [75]. | No | LC |
|
Gynochthodes constipata (Halford & A.J.Ford) Razafim. & B.Bremer Syn. Morinda constipata Halford & A.J.Ford |
Endemic to Wet Tropics region | Climber | Gynochthodes/Morinda species are traditionally used for treating diabetes, inflammation, cancer, psychiatric disorders, and microbial infections [76]. | No | LC |
|
Gynochthodes podistra (Halford & A.J.Ford) Razafim. & B.Bremer Syn. Morinda podistra Halford & A.J.Ford |
Endemic to Wet Tropics region | Climber | No | LC | |
|
Ixora orophila C.T.White Syn. Psydrax montigena S.T.Reynolds & R.J.F.Hend. |
Endemic to Wet Tropics region | Shrub | Ixora species are used in Ayurvedic medicine against leucorrhoea, dys, hypertension, menstrual irregularities, sprains, bronchitis fever, sores, chronic ulcers, scabies, and skin diseases [77]. | No | No |
| Wendlandia connata C.T.White | Endemic to Wet Tropics region | Shrub | Wendlandia species are traditionally used for treating fever, dysentery, cough, hypertension, diabetes, constipation, inflammations, and hyperlipidemia [78]. | No | NT |
| Rutaceae | |||||
| Flindersia oppositifolia (F.Muell.) T.G.Hartley & Jessup | Wet Tropics region (Australia) | Tree | NU | Yes | V |
| Leionema ellipticum Paul G. Wilson | Endemic to Wet Tropics region | Shrub | NU | Yes | V |
| Zieria alata Duretto & P.I.Forst. | Endemic to Wet Tropics region | Shrub | NU | No | CR |
| Zieria madida Duretto & P.I.Forst. | Endemic to Wet Tropics region | Shrub | NU | No | CR |
| Santalaceae | |||||
| Korthalsella grayi Barlow | Endemic to Wet Tropics region | Herb | No | LC | |
| Sapindaceae | |||||
|
Mischocarpus montanus C.T.White Syn. Mischocarpus pyriformis subsp. retusus (Radlk.) R.W.Ham, Mischocarpus retusus Radlk. |
Wet Tropics region (Australia), New Guinea | Tree | NU | No | LC |
| Sapotaceae | |||||
|
Pleioluma singuliflora (C.T.White & W.D.Francis) Swenson Syn. Planchonella singuliflora (C.T.White & W.D.Francis) P.Royen, Pouteria singuliflora (C.T.White & W.D.Francis) Baehni |
Endemic to Wet Tropic region | Shrub | NU | No | LC |
|
Sersalisia sessiliflora (C.T.White) Aubrév. Syn. Pouteria sylvatica Baehni, Lucuma sessiliflora C.T.White |
Endemic to Wet Tropics region | Tree | NU | No | LC |
| Planchonella sp. Mt. Lewis (B.Hyland 14048) Qld Herbarium | Endemic to Wet Tropics region | Tree | Planchonella species have been used by Aboriginal medicine system to treat sores/sore throat and as an antiseptic for boils [61]. | No | No |
| Solanaceae | |||||
| Solanum dimorphispinum C.T.White | Endemic to Wet Tropics region | Shrub | Solanum species have been traditionally used against infectious diseases and also as anti-microbial agents and insecticidal against mosquitoes [79]. | No | LC |
| Solanum eminens A.R.Bean | Endemic to Wet Tropics region | Climber | No | LC | |
| Symplocaceae | |||||
|
Symplocos bullata Jessup Syn. Symplocos sp. North Mary (B. Gray 2543) |
Endemic to Wet Tropics region | Shrub | Symplocos species are traditionally known for treating diseases such as malaria, ulcers, leprosy, leucorrhea, menorrhagia, and gynecological disorders [80]. | No | LC |
| Symplocos graniticola Jessup | Endemic to Wet Tropics region | Shrub | No | V | |
|
Symplocos oresbia Jessup Syn. Symplocos sp. Mt Finnigan (L.J. Brass 20129) |
Endemic to Wet Tropics region | Shrub | No | NT | |
|
Symplocos wooroonooran Jessup Syn. Symplocos stawellii var. montana C.T.White, Symplocos cochinchinensis var. montana (C.T.White) Noot |
Endemic to Wet Tropics region | Shrub | No | NT | |
| Thymelaeaceae | |||||
|
Phaleria biflora (C.T.White) Herber Syn. Oreodendron biflorum C.T.White |
Endemic to Wet Tropics region | Tree | Phaleria species are used for treating stomachache, general pain, diarrhea, lowering glucose/cholesterol levels in blood, and also known for anti-cancer properties [81]. | No | V |
| Winteraceae | |||||
|
Bubbia whiteana A.C.Sm. Syn. Zygogynum semecarpoides var. whiteanum Vink, Bubbia semecarpoides var. whiteana Vink |
Endemic to Wet Tropics region | Shrub | NU | No | CR |
| Tasmannia sp. Mt Bellenden Ker (J.R.Clarkson 6571) | Wet Tropics region (Australia) | Shrub | Tasmania species are traditionally used for treating malaria, diarrhea, and cough [82]. | No | LC |
The scientific names and plant families follow the Australian Plant Census. Where taxonomy differs in "Plants of the World Online" [83], the synonym is given; Distribution and plant life form were sourced from: Atlas of Living Australia [41], Australian Tropical Rainforest Plants system [42], and Australian Tropical Rainforest Orchids [84]. Conservation status is per the Queensland Nature Conservation Act 1992 [85]: Abbreviations - SL: Special Least Concern; LC: Least Concern; NT: Near Threatened; V: Vulnerable; E: Endangered; CR: Critically Endangered; No: Species for which no conservation status is indicated; NU: Not used medicinally.
Figure 3.
Conservation status of climate-affected Australian montane cloud forest plants in the Wet Tropics World Heritage Area, Queensland. Conservation status are as per the Queensland Nature Conservation Act 1992 [85].
Figure 3.
Conservation status of climate-affected Australian montane cloud forest plants in the Wet Tropics World Heritage Area, Queensland. Conservation status are as per the Queensland Nature Conservation Act 1992 [85].
3. Metabolomic Profile of Climate-Affected Plants in WTWHA
The anticipated impact of global climate changes on plant secondary metabolism is significant, but a comprehensive understanding of these effects is currently absent. Changes in the metabolome (defined as the complete set of metabolites found in a biological sample) can occur rapidly in seconds or minutes due to living organisms' responses, acclimation, and adaptation to environmental conditions [86,87,88]. Investigations into climate effects on plants have shown that plants growing under various climatic stresses in their natural habitat produce various SMs that could potentially have a role in adaptation to the changing environment [89,90,91]. Studies have also revealed that abiotic stress factors, such as increased temperature and ultraviolet (UV) radiation, stimulate plants to reprogram their genetic codes for metabolic pathways, leading to the accumulation of new and unique secondary metabolites [92].
For example, it was demonstrated that elevated temperatures can lead to increased production of terpenoids, phenolic acids, and flavonoids in plants [93,94]. These compounds act as protective pigments when trees are exposed to UV-B radiation (wavelengths between 280-315 nm) [95]. Likewise, higher ozone (O3) concentrations have been linked to heightened production of antioxidant compounds such as glutathione, gamma-aminobutyric acid (GABA), terpenoids, and volatile organic compounds (VOCs) [96]. For example, the production of phenolics in plants plays an integral part in protecting mesophyll tissue from UV radiation and water stress [97,98]. It was discovered that drought conditions enhance plant productivity, leading to increased production of SMs, terpenes, complex phenols, and alkaloids [99,100,101]. Moreover, secondary metabolites with antioxidant properties, such as phenolic compounds and tocopherols, were known to scavenge the ROS (Reactive Oxygen Species) generated, thus adapting to a new environment [102].
In addition to metabolites, plants store proteins like hydrolases, enzymes for detoxifying reactive oxygen species (ROS), and enzymes for modifying cell walls. These proteins act as regulatory agents, governing plant growth and development [103]. Similarly, salinity also impacts the plant’s growth and development. It leads to an abnormal ion composition, causing toxicity, osmotic stress, producing reactive oxygen species (ROS), cellular harm, and degrading membrane lipids, proteins, and nucleic acids [102]. In response to saline soil stress, plants undergo a biochemical process that produces ions that can act against ion toxicity and abnormal osmotic pressure developed from salinity [104]. In addition, when a huge amount of sodium (Na+) ions prevails, plants respond to an abundance of Na+ ions by activating a sophisticated defense system, which enables them to regulate cellular and ion balance effectively [105]
These studies enable us to understand the pattern of metabolite/micronutrient changes, including compositions, variations, and biosynthetic pathways resulting from plants’ responses to biotic and abiotic stressors is known as the Plant metabolomics [106,107]. It can provide insights into plant phenotypic relations to their physiological and resistance development and biodiversity [108]. More than 200,000 secondary metabolites (SMs) have been identified [109] from over 391,000 plant species known worldwide [110] through metabolomics studies. The projected number for the plant kingdom is expected to surpass 200,000.[111,112]. Hence, plant metabolomics poses a significant hurdle for researchers in plant science. The comprehensive research workflow in plant metabolomics encompasses experimental planning, sample gathering, sample handling, sample preparation, detection and examination, data handling, as well as the analysis of metabolic pathways and networks [113].
From our literature review on 84 plant species affected by climate change in WTWHA of FNQ, only nine species were studied for their metabolomic profiles/phytochemical contents (Table 2). A total of 279 metabolites (251 identified and 18 isolated) were identified/isolated from parts of nine plant species. The identified metabolites were mostly flavonoids, terpenoids, alkaloids, and glucosides (Table 2). However, none of these metabolomics studies included in Table 2 were conducted to investigate their response to climatic stress conditions under in situ or ex-situ conditions. There is a need for this type of study, but the challenge would be to control various factors influencing plant responses, which is why we see most of the studies conducted under controlled conditions in glass houses. Currently, our group is conducting a first-of-its-kind study on selected WTWHA plants, in which we are comparing the metabolome profile and chemical variation between the wild and the domesticated plant population.
4. Phytochemicals Isolated from Climate-Affected Plants in WTWHA
Out of 84 plant species included in this review, phytochemicals were isolated from only three plant species, namely, Uromyrtus metrosideros, Flindersia oppositifolia, and Leionema ellipticum (Table 2). A total of 19 compounds/secondary metabolites were isolated from these three plant species (Table 2), and these compounds belong to four different chemical groups (alkaloids, flavonoids, benzopyrans, and glucosides). For example, two new galloyl glucosides (galloyl-lawsoniaside A and uromyrtoside) and four known compounds were isolated from Uromyrtus metrosideros [114]. These six compounds were characterised using low and high-resolution mass spectrometry (L/HRMS) and Nuclear Magnetic Resonance (NMR) spectroscopy. All three studies involving three plants were conducted to identify pharmacological drug leads (U. metrosideros and F. oppositifolia) and solve the taxonomic discrepancies (L. ellipticum). They did not suggest their role in response to climatic stress factors.
Table 2.
List of climate-affected Australian montane cloud forest plants studied for their phytochemical contents and bioactivity.
Table 2.
List of climate-affected Australian montane cloud forest plants studied for their phytochemical contents and bioactivity.
| Botanical name | Medicinal uses | Number and major metabolites identified | Isolated Compounds | Chemical class | Biological Activities of compounds |
|---|---|---|---|---|---|
| Agathis atropurpurea | Agathis species are traditionally used to treat myalgia and headaches [52]. | 27 metabolites; major metabolites are ⍺-pinene, ⍺-copaene, bicyclogermacrene, 𝛿-cadinene, phyllocladane, and 16-kaurene [115] | NA | Terpenoid | Antimicrobial, antibacterial, antiviral, anti-cancer activity (⍺-pinene) [116,117,118], antioxidant activity (⍺-copaene) [119] |
| Eucryphia wilkiei | NU | 2 unknown metabolites [120] | NA | Flavonoid | NA |
| Cinnamomum propinquum | Cinnamomum species are most commonly used in traditional Chinese medicines to treat multiple disorders, including indigestion, microbial infections, and cough and cold [59]. | 40 metabolites; Major metabolites are p-cymene, ⍺-pinene, and β-eudesmol [121] |
NA | Terpenoid | Anti-cancer activity (p-cymene) [122], anti-allergic and anti-angiogenic effect (β-eudesmol) [123,124] |
| Cryptocarya bellendenkerana | NU | 39 metabolites; major metabolites are ⍺-pinene, limonene, β-phellandrene, p-cymene, viridiflorene, E-β-farnesene, ⍺-copaene, β-and ⍺-selinene, 𝛿-cadinene, bicyclogermacrene, calamenene, and cubeban-11-ol [125]. | NA | Antioxidant, antidiabetic, anticancer, anti-inflammatory (limonene) [126,127], anti-fungal (β-phellandrene) [128], antioxidant and antitumour properties (viridiflorene) [129,130], insect repellent (E-β-farnesene) [131] antioxidant activity (copaene) [119]. | |
| Leptospermum wooroonooran | Leptospermum species are traditionally used in Malaysia to relieve menstrual and stomach disorders [62,63]. | 45 metabolites; major metabolitesare ⍺-pinene, β-pinene, sabinene, ⍺-terpinene, 𝛾-terpinene, terpinen-4-ol and ⍺-terpineol [132] | NA | Reduce skeletal muscle atrophy (sabinene) [133], antibacterial and antibiofilm activities (terpinene-4-ol) [134] | |
| Uromyrtus metrosideros | NU | 27 metabolites; major metabolites are ⍺-pinene, β-pinene, spathulenol and aromadendrene [135] | norbergenin, bergenin, (6S,9R)-roseoside, (4S)-𝜶-terpineol 8-O-β-D-(6-O-galloyl) glucopyranoside, galloyl-lawsoniaside A, and uromyrtoside [114] |
Benzopyran, Glucoside, |
Anti-inflammatory (galloyl-lawsoniaside A) [114]; reduced hypertension and allergic reaction (roseoside) [136,137] |
| Prumnopitys ladei | Fruits and bark of Prunmnopitys species are considered medicinal [70]. | 44-metabolites; major compounds are ⍺-pinene, limonene, verbenone, and p-cymene. β-caryophyllene, caryophyllene oxide, spathulenol, and ⍺-humulene [138] |
NA | Antimicrobial, anticarcinogenic, anti-inflammatory, antioxidant, and local anesthetic effects (β-caryophyllene) [139,140,141] | |
|
Flindersia oppositifolia |
NU | 37 metabolites; major compounds are β-caryophyllene and bicyclogermacrene [142]; Identified 8 alkaloids from leaf [143]. |
pimentelamine A, pimentelamine B, pimentelamine C, 2-isoprenyl-N-N-dimethyltryptamine, 4-methylborreverine, borreverine, dimethylisoborreverine, quercitrin, and carpachromene [142]; harmalan, pimentelamine B, isoborreverine, skimmianine, kokusaginine, maculosidine, flindersiamine, 8-methoxy-N-methylflindersine [143]. | Terpene, Alkaloid | Antiplasmodial (pimentelamine C) [144,145] |
| Leionema ellipticum | NU | 3,4ʹ,5-trimethoxyflavone-7-O-⍺-rhamnoside, boropinol-B, and osthol [146] |
Flavonoid | Neuroprotective (boropinol-B) [147,148]; anti-inflammatory (osthol) [149,150] | |
Chemical class for isolated chemicals were referred from human metabolome database (https://hmdb.ca); Abbreviations - NA: Not available; NU: Not used medicinally.
5. Pharmacological Activities of Isolated Phytochemicals of Climate-affected Plants in WTWHA
Studies have suggested that SMs, which function as plant defense mechanisms, possess intriguing pharmacological properties, including antioxidant and anti-inflammatory properties [151]. For instance, a novel galloyl-lawsoniaside A isolated from U. metrosideros leaf significantly suppressed pro-inflammatory cytokines, such as interferon-gamma and interleukins-17 (IL-17) and IL-18, and thus were identified as new anti-inflammatory drug-lead molecule [114]. Osthol isolated from Leionema ellipticum also showed anti-inflammatory activity [149,150]. A study by Yeshi et al. [151] analysed crude extracts from leaves of seven plant species endemic to WTWHA of FNQ. Five of seven plant species showed potent antioxidant and anti-inflammatory activities in in-vitro human peripheral blood cells (PBMCs) assay [151]. About 30 plant species growing in the WTWHA were reported as medicinal plants used for many years by indigenous communities to treat various diseases and ailments, including inflammation-related diseases [152]. Many metabolites identified through metabolomic studies (Table 2) were also studied for numerous biological properties (Table 2). Some major and bioactive metabolites were ⍺-pinene, p-cymene, β-endemol, limonene, viridiflorene, E-β-farnesene, copaene, and β-caryophyllene (Table 2). Figure 6 shows some interesting structures of these isolated compounds. They showed a wide array of pharmacological activities, from anti-microbial to anti-cancer and anti-plasmodial properties. Of nine plant species listed in Table 2, four were tested for anti-inflammatory and anti-cancer properties, three each were tested for anti-microbial and antioxidant activities, two were tested for anti-allergic reactions, and the rest were tested for anti-diabetic, anti-malarial, and neuro-protective properties (one plant species each). For instance, galloyl-lawsoniaside A isolated from U. metrosideros leaf showed promising anti-inflammatory activity through significant suppression of pro-inflammatory cytokines, interferon-gamma (IFN-γ), and interleukin-17A (IL-17A) by phorbol myristate acetate/ionomycin (P/I)-activated cells [114]. Moreover, it also significantly suppressed the release of IL-8 by the anti-CD3/anti-CD28-activated cells [114]. There are increasing studies on identifying anti-inflammatory molecules by targeting the 5-lipoxygenase (5-LOX) pathway, as 5-LOX drives inflammation by producing inflammatory mediators, such as leukotrienes [153,154]. Osthol isolated from aerial parts of Leionema ellipticum showed selective inhibition of the 5-LOX pathway [146]. The anti-cancer/anti-tumor activity was mainly tested with crude extracts or essential oils by studying their inhibitory effect on tumor growth using tumor cell lines such as sarcoma 180 ascites tumor cells [129]. The anti-cancer activity was attributed to the major metabolite constituents, such as limonene, p-cymene, α-pinene, and viridiflorene (Table 2), and was not tested against the single compound. A few isolated compounds also exhibited anti-plasmodial activity. For example, pimentelamine C, isolated from the leaf of Flindersia pimentaliana showed moderate anti-plasmodial activity against Plasmodium falciparum with IC50 values of 3.6 ± 0.7 (against chloroquine-sensitive strain) and 2.7 ± 0.3 (against chloroquine-resistant strains) [144]. Figure 6 shows some interesting structures of these isolated compounds.
6. Metabolomics Approaches, Tools, and Techniques Used in Plant Metabolomics
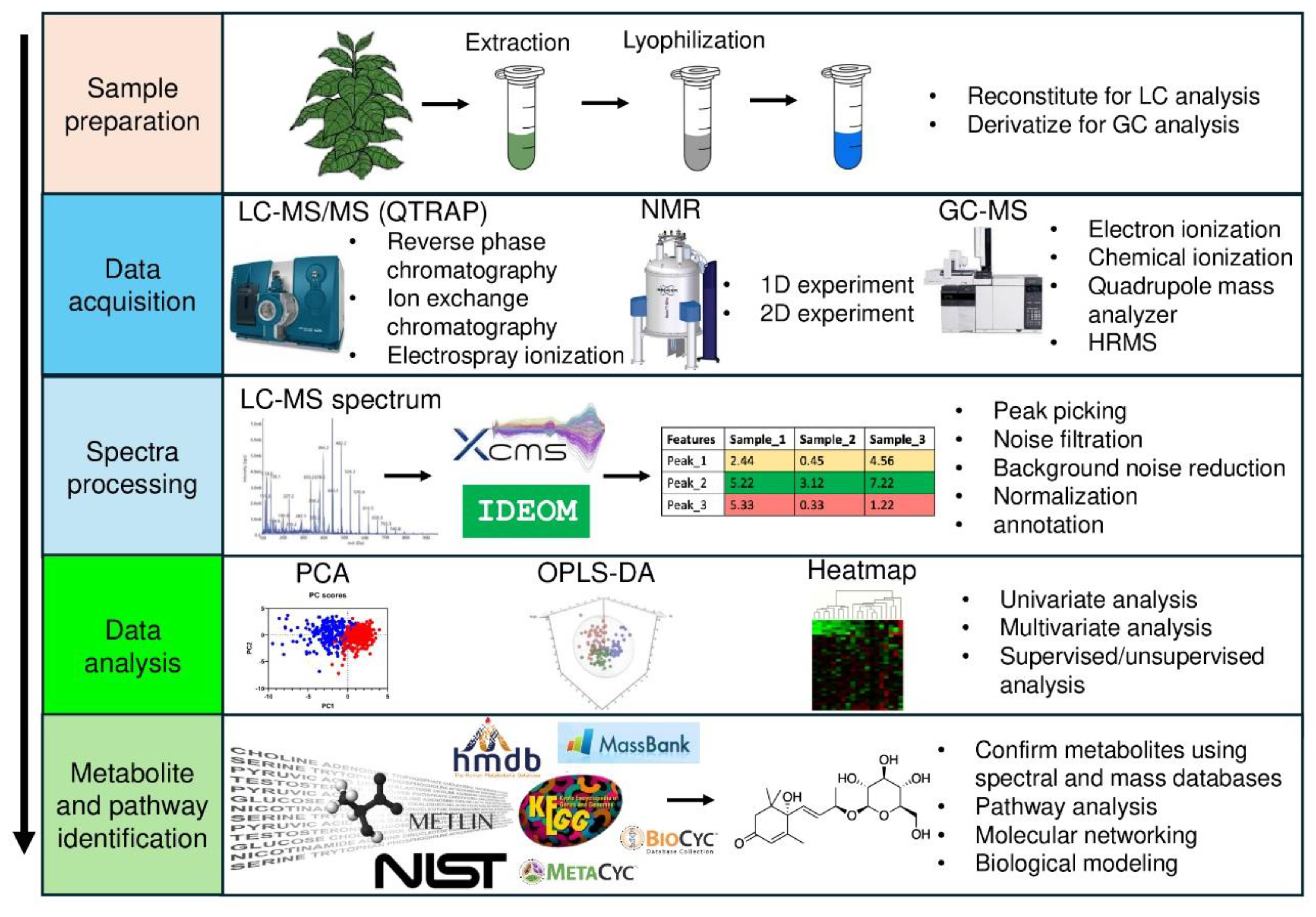
Understanding plants’ physiological and metabolomic responses to global change is key to identifying potential traits, including their genetic mutations and changes in their metabolomic pathways. Additionally, it is possible to predict potential changes in the composition of plant communities by assessing the ability of various plant species to adapt to environmental shifts [155,156]. Metabolites from living matters can be identified using i) isolation techniques and ii) metabolomics platforms. Metabolomics platforms, in general, rely on mass spectrometry (MS)-based techniques, namely capillary electrophoresis mass spectrometry (CE-MS), liquid chromatography-mass spectrometry (LC-MS), and gas chromatography-mass spectrometry (GC-MS) [157,158]. Two innovative technological methods that do not require the use of metabolite chromatography include Nuclear Magnetic Resonance (NMR) analysis of unrefined extracts and the direct inspection of unrefined extracts using Mass Spectrometry (MS), specifically either quadrupole (Q) TOF-MS or ultra-high-resolution Fourier transform ion cyclotron MS (FT-MS) [108], which are discussed in-depth in later sections. Compared to conventional methods in the postgenomic era, metabolomics analysis offers numerous advantages and potential applications [158]. It has various steps, as shown in Figure 5, including sample preparation, spectra processing, data analysis, and metabolite identification.
The NMR-based metabolomic analysis offers a potent, non-invasive method, delivering precise structural details about metabolites [159]. While metabolomic analysis using mass spectrometry is inherently destructive, it is highly sensitive and can detect traces of metabolites, and thus, it has gained more popularity [160]. Mass spectrometry (MS) methods are frequently integrated with chromatographic separation methods, including gas chromatography (GC) and liquid chromatography (LC) [160]. Since the metabolome is a complex mixture of many small molecules, chromatographic separation is necessary prior to ion detection, particularly to distinguish isobaric compounds with a similar mass. Alternatively, the direct-infusion mass spectrometry (DIMS) approach is applied to measure metabolites directly without a prior chromatographic separation [161]. Most of the plant-based metabolomics published studies so far use the Orbitrap or TOF (time-of-flight) equipment [162]. One of the main reasons for using TOF equipment could be due to its mass resolution values (i.e., 30,000–40,000) [163,164], and the resolution power is unaffected by chromatography acquisition rates [165,166,167]. On the contrary, Orbitrap mass spectrometers can rapidly acquire tandem MS spectra up to 240,000 mass resolution, and thus, they are mainly applied in the shotgun metabolomics [165,168,169].
The DIMS methodology has recently been expanded to swift, high-throughput fingerprinting techniques employing advanced mass spectrometers with high resolution, such as Fourier Transform Ion Cyclotron Resonance (FT-ICR) mass spectrometers (FT-ICR-MS) [165], its extensive usage in plant metabolomics greatly aids in understanding plant development, responses to both biotic and abiotic stresses, and the exploration of novel natural nutraceutical compounds [162,170]. Fourier Transform Ion Cyclotron Resonance mass spectrometers have a higher resolution power (105 to >106), mass accuracy (typically 0.1–2 ppm), and sensitivity [171,172]. For instance, FT-ICR-MS can analyse and evaluate approximately 50,000 molecular formulas in complex samples, such as plant-derived crude oils [173,174]. Another reason for widely using FT-ICR-MS is that this application has a wide range of ionisation sources, including electron spray ionisation (ESI), atmospheric pressure chemical ionization, and photoionization, thus, enabling analyses of different sample types [175]. For example, Shahbazya et al. [176] used FT-ICR-MS to study the response of thyme plants (Thymus vulgaris) to drought stress. The study identified galactose metabolism as the most significant for drought adaptive response in thyme. Other studies, including metabolomics changes in poplar species in response to salinity stress [177] and UV-B radiation, also applied the same technique [178].
Nevertheless, because metabolites exhibit various chemical properties and are found abundantly in various cells, no single analytical platform can encompass the entire metabolome. Therefore, multi-omics technology has enabled the exploration of genes and metabolites in response to various climatic stress factors, particularly by combining transcriptomics and metabolomics approaches. For instance, Wang et al., [24], in their study about Poa crymophila, applied transcriptomics and metabolomics and identified the phenylpropanoid pathway as the main mechanism that facilitates this plant to survive against the unfavourable environment of Qinghai-Tibet Plateau. Liu et al., [23] also applied a combination of transcriptomics with physiological analyses to understand the chilling response in pumpkins and found that α-linolenic acid biosynthesis was one of the key pathways in the response.
These instruments employ three types of approaches to characterize metabolites, namely, (i) targeted analysis, (ii) untargeted analysis, and (iii) metabolic fingerprinting [179]. Unlike the targeted approach (identify a set of targeted metabolites with reference to available standards), the untargeted approach generates a large volume and complex data requiring specialised computational methods, such as artificial intelligence (AI) and machine learning (ML) algorithms, to process and interpret data [179]. In contrast, metabolic fingerprinting or exometabolomics involves characterising extracellular metabolites (i.e., metabolic by-products of organisms produced in response to environmental factors in which they survive) [180,181]. For plant metabolomic profiling, ‘ecometabolomics’ is a commonly applied technique. The term ‘’ecometabolomics’’ first appeared in the scientific literature in 2009 [34,182]. This study investigates how living organisms respond, acclimate, and adapt to environmental conditions by a nontargeted approach [86,87,88]. Metabolite identification can be achieved at four different metabolite standard initiative (MSI) levels. Metabolite standard initiative level-1 (MSI-1) is considered the highest level of identification as it identifies metabolites after comparing with their chemical standards [183,184]. Level 2 (MSI-2) and level-3 identifications are only putative, as metabolites (MSI-2) or metabolite class (MSI-3) are not compared to their chemical standards, whereas MSI level-4 (MSI-4) putatively annotates unknown metabolites [183,184].
Figure 5.
Common metabolomic workflow applied in plant metabolomics studies. The figure was adapted from Xu and Fu [185], and all databases’ logos used in this figure were obtained from their respective websites. Abbreviations: - LC-MS: Liquid chromatography-mass spectrometry; NMR: Nuclear magnetic resonance; 1D: one dimensional; 2D: two dimensional; GC-MS: Gas chromatography-mass spectrometry; QTRAP: The Quadrupole Ion Trap; XCMS: eXtensible Computational Mass Spectrometry; HMDB: Human metabolome database; PCA: principal component analysis; OPLS-DA: Orthogonal Partial Least Squares Discriminant Analysis; NIST: National Institute of Standards and Technology; METLIN: Metabolite and chemical entity database. .
Figure 5.
Common metabolomic workflow applied in plant metabolomics studies. The figure was adapted from Xu and Fu [185], and all databases’ logos used in this figure were obtained from their respective websites. Abbreviations: - LC-MS: Liquid chromatography-mass spectrometry; NMR: Nuclear magnetic resonance; 1D: one dimensional; 2D: two dimensional; GC-MS: Gas chromatography-mass spectrometry; QTRAP: The Quadrupole Ion Trap; XCMS: eXtensible Computational Mass Spectrometry; HMDB: Human metabolome database; PCA: principal component analysis; OPLS-DA: Orthogonal Partial Least Squares Discriminant Analysis; NIST: National Institute of Standards and Technology; METLIN: Metabolite and chemical entity database. .
7. Conclusions and Future Directions
The Australian Tropical Montane Cloud Forest (TMCF), which lies in the Wet Tropics World Heritage Area in FNQ, has rich and unique biodiversity, with over 700 endemic plant species. The current study identified that 84 plant species were affected by climate change, with some species already being endangered in their natural habitat. Recent studies of 37 of these species predicted total loss of suitable habitat for five species by 2035 and seven species by 2085 if greenhouse gas emission (e.g., CO2) into the atmosphere continues at the current speed. Recently, many powerful technologies have been developed, including omics techniques, bioimaging, and biosensor tools, which have been widely applied to understanding plants' physiology and metabolome and discovering novel metabolomic pathways in response to global climate change. However, our literature review revealed that these 84 Australian TMCF plants were scarcely studied for their biomolecules, and we understand little about their medicinal uses, chemical profiles, and biological functions. Of 84 species, 43 belong to 29 medicinally important genera with various medical properties, and only seven species were studied for their metabolite compositions. There is an urgent need for enhanced metabolomics studies of these least studied plants, given that they are at risk of significant habitat loss because of climate change. Additionally, it is urgent to understand and identify potential traits in these least studied plants, including possible genetic mutations that may have led to the change in the pattern of secondary metabolite accumulation and their metabolomic pathways in the adaptive response to climatic stress factors. Such studies will produce more data to holistically understand the interactive effect of climate change on the growth and fitness of these plants. This, in turn, would enable us to predict the adaptive response of plants specific to future climatic conditions and, thus, design the appropriate conservation measures to rescue those already identified as endangered and nearly threatened plant species.
Many metabolites reported from those plants that have already been studied have shown numerous pharmacological activities. Studies have also reported that plants produce defensive/protective secondary metabolites in response to climate change. Most of these defensive secondary metabolites are antioxidative/anti-inflammatory. Therefore, Australian TMCF plants also present an exciting avenue to discover novel pharmaceutical leads.
Author Contributions
Conceptualization, P.W., and K.Y.; writing—original draft preparation, N.G., and K.Y; writing—review and editing, P.W. and D.C.; supervision, P.W., and D.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
Not Applicable.
Acknowledgments
We acknowledge funding support from the FNQH grant and the NHMRC Ideas grant (APP1183323) awarded to P.W and the Ian Potter Foundation grant awarded to D.C. Authors sincerely acknowledge the office of Wet Tropics Management Authority office, state of Queensland for generating and allowing us to use a map of the Wet Tropics bioregion depicting WTWHA in this study.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Arnell, N.W.; Lowe, J.A.; Challinor, A.J.; Osborn, T.J. Global and regional impacts of climate change at different levels of global temperature increase. Climatic Change 2019, 155, 377-391. [CrossRef]
- Cohen, I.; Zandalinas, S.I.; Huck, C.; Fritschi, F.B.; Mittler, R. Meta-analysis of drought and heat stress combination impact on crop yield and yield components. Physiologia Plantarum 2021, 171, 66-76. [CrossRef]
- QLD. Climate change in the Far North Queensland region. 2019.
- Morris, R.J. Anthropogenic impacts on tropical forest biodiversity: a network structure and ecosystem functioning perspective. Philosophical Transactions of the Royal Society B: Biological Sciences 2010, 365, 3709-3718. [CrossRef]
- Hoyle, G.L.; Sommerville, K.D.; Liyanage, G.S.; Worboys, S.; Guja, L.K.; Stevens, A.V.; Crayn, D.M. Seed banking is more applicable to the preservation of tropical montane flora than previously assumed: A review and cloud forest case study. Global Ecology and Conservation 2023, 47. [CrossRef]
- Helmer, E.H.; Gerson, E.A.; Baggett, L.S.; Bird, B.J.; Ruzycki, T.S.; Voggesser, S.M. Neotropical cloud forests and páramo to contract and dry from declines in cloud immersion and frost. PLOS ONE 2019, 14, e0213155. [CrossRef]
- Costion, C.M.; Simpson, L.; Pert, P.L.; Carlsen, M.M.; John Kress, W.; Crayn, D. Will tropical mountaintop plant species survive climate change? Identifying key knowledge gaps using species distribution modelling in Australia. Biological Conservation 2015, 191, 322-330. [CrossRef]
- Karger, D.N.; Kessler, M.; Lehnert, M.; Jetz, W. Limited protection and ongoing loss of tropical cloud forest biodiversity and ecosystems worldwide. Nature Ecology & Evolution 2021, 5, 854-862. [CrossRef]
- Still, C.J.; Foster, P.N.; Schneider, S.H. Simulating the effects of climate change on tropical montane cloud forests. Nature 1999, 398, 608-610. [CrossRef]
- Foster, P. The potential negative impacts of global climate change on tropical montane cloud forests. Earth-Science Reviews 2001, 55, 73-106. [CrossRef]
- Hu, J.; Riveros-Iregui, D.A. Life in the clouds: are tropical montane cloud forests responding to changes in climate? Oecologia 2016, 180, 1061-1073. [CrossRef]
- Williams, S.E.; Bolitho, E.E.; Fox, S. Climate change in Australian tropical rainforests: an impending environmental catastrophe. Proc. R. Soc. B Biol. Sci. 2003, 270, 1887-1892. [CrossRef]
- Williams, S.E.; Bolitho, E.E.; Fox, S. Climate change in Australian tropical rainforests: an impending environmental catastrophe. Proc Biol Sci 2003, 270, 1887-1892. [CrossRef]
- Le Saout, S.; Hoffmann, M.; Shi, Y.; Hughes, A.; Bernard, C.; Brooks, T.M.; Bertzky, B.; Butchart, S.H.; Stuart, S.N.; Badman, T.; et al. Conservation. Protected areas and effective biodiversity conservation. Science 2013, 342, 803-805. [CrossRef]
- UNESCO World Heritage Convention. Wet Tropics of Queensland. Available online: https://whc.unesco.org/en/list/486/ (accessed on August 6 2023).
- Weber, E.T.; Catterall, C.P.; Locke, J.; Ota, L.S.; Prideaux, B.; Shirreffs, L.; Talbot, L.; Gordon, I.J. Managing a World Heritage Site in the Face of Climate Change: A Case Study of the Wet Tropics in Northern Queensland. Earth 2021, 2, 248-271. [CrossRef]
- Grossmann, G.; Krebs, M.; Maizel, A.; Stahl, Y.; Vermeer, J.E.M.; Ott, T. Green light for quantitative live-cell imaging in plants. Journal of Cell Science 2018, 131. [CrossRef]
- Awlia, M.; Alshareef, N.; Saber, N.; Korte, A.; Oakey, H.; Panzarová, K.; Trtílek, M.; Negrão, S.; Tester, M.; Julkowska, M.M. Genetic mapping of the early responses to salt stress in Arabidopsis thaliana. The Plant Journal 2021, 107, 544-563. [CrossRef]
- Berg, C.S.; Brown, J.L.; Weber, J.J. An examination of climate-driven flowering-time shifts at large spatial scales over 153 years in a common weedy annual. American journal of botany 2019, 106, 1435-1443. [CrossRef]
- Cortijo, S.; Charoensawan, V.; Brestovitsky, A.; Buning, R.; Ravarani, C.; Rhodes, D.; van Noort, J.; Jaeger, K.E.; Wigge, P.A. Transcriptional regulation of the ambient temperature response by H2A. Z nucleosomes and HSF1 transcription factors in Arabidopsis. Molecular plant 2017, 10, 1258-1273. [CrossRef]
- Sriden, N.; Charoensawan, V. Large-scale comparative transcriptomic analysis of temperature-responsive genes in Arabidopsis thaliana. Plant Molecular Biology 2022, 110, 425-443. [CrossRef]
- Zhao, Y.; Antoniou-Kourounioti, R.L.; Calder, G.; Dean, C.; Howard, M. Temperature-dependent growth contributes to long-term cold sensing. Nature 2020, 583, 825-829. [CrossRef]
- Liu, W.; Zhang, R.; Xiang, C.; Zhang, R.; Wang, Q.; Wang, T.; Li, X.; Lu, X.; Gao, S.; Liu, Z.; et al. Transcriptomic and Physiological Analysis Reveal That alpha-Linolenic Acid Biosynthesis Responds to Early Chilling Tolerance in Pumpkin Rootstock Varieties. Front Plant Sci 2021, 12, 669565. [CrossRef]
- Wang, Y.; Li, X.Y.; Li, C.X.; He, Y.; Hou, X.Y.; Ma, X.R. The Regulation of Adaptation to Cold and Drought Stresses in Poa crymophila Keng Revealed by Integrative Transcriptomics and Metabolomics Analysis. Front Plant Sci 2021, 12, 631117. [CrossRef]
- Sun, Y.; Alseekh, S.; Fernie, A.R. Plant secondary metabolic responses to global climate change: A meta-analysis in medicinal and aromatic plants. Global Change Biology 2023, 29, 477-504. [CrossRef]
- Hodges, M.; Dellero, Y.; Keech, O.; Betti, M.; Raghavendra, A.S.; Sage, R.; Zhu, X.-G.; Allen, D.K.; Weber, A.P. Perspectives for a better understanding of the metabolic integration of photorespiration within a complex plant primary metabolism network. Journal of Experimental Botany 2016, 67, 3015-3026. [CrossRef]
- Ncube, B.; Van Staden, J. Tilting plant metabolism for improved metabolite biosynthesis and enhanced human benefit. Molecules 2015, 20, 12698-12731.
- Yang, L.; Wen, K.-S.; Ruan, X.; Zhao, Y.-X.; Wei, F.; Wang, Q. Response of plant secondary metabolites to environmental factors. Molecules 2018, 23, 762. [CrossRef]
- Peng, M.; Shahzad, R.; Gul, A.; Subthain, H.; Shen, S.; Lei, L.; Zheng, Z.; Zhou, J.; Lu, D.; Wang, S. Differentially evolved glucosyltransferases determine natural variation of rice flavone accumulation and UV-tolerance. Nature communications 2017, 8, 1975. [CrossRef]
- Tohge, T.; Fernie, A.R. Leveraging natural variance towards enhanced understanding of phytochemical sunscreens. Trends in plant science 2017, 22, 308-315. [CrossRef]
- Boncan, D.A.T.; Tsang, S.S.; Li, C.; Lee, I.H.; Lam, H.-M.; Chan, T.-F.; Hui, J.H. Terpenes and terpenoids in plants: Interactions with environment and insects. Int. J. Mol. Sci. 2020, 21, 7382. [CrossRef]
- Matsuura, H.N.; Rau, M.R.; Fett-Neto, A.G. Oxidative stress and production of bioactive monoterpene indole alkaloids: biotechnological implications. Biotechnology letters 2014, 36, 191-200. [CrossRef]
- Bakhtiari, M.; Rasmann, S. Variation in below-to aboveground systemic induction of glucosinolates mediates plant fitness consequences under herbivore attack. Journal of chemical ecology 2020, 46, 317-329. [CrossRef]
- Sardans, J.; Gargallo-Garriga, A.; Urban, O.; Klem, K.; Walker, T.W.N.; Holub, P.; Janssens, I.A.; Peñuelas, J. Ecometabolomics for a Better Understanding of Plant Responses and Acclimation to Abiotic Factors Linked to Global Change. Metabolites 2020, 10. [CrossRef]
- Ma, A.; Qi, X. Mining plant metabolomes: Methods, applications, and perspectives. Plant Commun 2021, 2, 100238. [CrossRef]
- Oh, S.-W.; Imran, M.; Kim, E.-H.; Park, S.-Y.; Lee, S.-G.; Park, H.-M.; Jung, J.-W.; Ryu, T.-H. Approach strategies and application of metabolomics to biotechnology in plants. Frontiers in Plant Science 2023, 14. [CrossRef]
- Colin, L.; Martin-Arevalillo, R.; Bovio, S.; Bauer, A.; Vernoux, T.; Caillaud, M.-C.; Landrein, B.; Jaillais, Y. Imaging the living plant cell: From probes to quantification. The Plant Cell 2021, 34, 247-272. [CrossRef]
- Hsiao, A.-S.; Huang, J.-Y. Bioimaging tools move plant physiology studies forward. Frontiers in Plant Science 2022, 13. [CrossRef]
- Uslu, V.V.; Grossmann, G. The biosensor toolbox for plant developmental biology. Current Opinion in Plant Biology 2016, 29, 138-147. [CrossRef]
- Gamalero, E.; Bona, E.; Glick, B.R. Current Techniques to Study Beneficial Plant-Microbe Interactions. Microorganisms 2022, 10. [CrossRef]
- Belbin, L.; Wallis, E.; Hobern, D.; Zerger, A. The Atlas of Living Australia: History, current state and future directions. Biodiversity Data Journal 9: e65023. Available online: (accessed on September 19).
- Zich, F.A.; Hyland, B.P.M.; Whiffin, T.; Kerrigan, R.A. Australian Tropical Rainforest Plants, Edition 8. 2020.
- Crayn, D.; Worboys, S. Personal communication. 2023.
- APC. Australian Plant Census IBIS database, Centre for Australian National Biodiversity Research, Council of Heads of Australasian Herbaria. 2024.
- WFO. World Flora Online. Version 2023.03. 2023. [CrossRef]
- CSIRO. CSIRO Annual Report 2010-11; ACT: Australia, 2011; pp. 1-176.
- E., R. Modelling the vulnerability of endemic montane flora to climate change in the Australian Wet Tropics. MSc thesis. Imperial College London, UK, 2018.
- Roeble, E. Modelling the vulnerability of endemic montane flora to climate change in the Australian Wet Tropics. Imperial College London, UK, 2018.
- Armijos, C.; Gilardoni, G.; Amay, L.; Lozano, A.; Bracco, F.; Ramirez, J.; Bec, N.; Larroque, C.; Finzi, P.V.; Vidari, G. Phytochemical and ethnomedicinal study of Huperzia species used in the traditional medicine of Saraguros in Southern Ecuador; AChE and MAO inhibitory activity. Journal of Ethnopharmacology 2016, 193, 546-554. [CrossRef]
- Hamdy, S.A.; El Hefnawy, H.M.; Azzam, S.M.; Aboutabl, E.A. Botanical and genetic characterization of Hydrocotyle umbellata L. cultivated in Egypt. Bulletin of Faculty of Pharmacy, Cairo University 2018, 56, 46-53. [CrossRef]
- Śliwińska, A.; Figat, R.; Zgadzaj, A.; Wileńska, B.; Misicka, A.; Nałęcz-Jawecki, G.; Pietrosiuk, A.; Sykłowska-Baranek, K. Polyscias filicifolia (Araliaceae) Hairy Roots with Antigenotoxic and Anti-Photogenotoxic Activity. Molecules 2021, 27. [CrossRef]
- Ho, Y.T.; Liu, I.H.; Chang, S.T.; Wang, S.Y.; Chang, H.T. In Vitro and In Vivo Antimelanogenesis Effects of Leaf Essential Oil from Agathis dammara. Pharmaceutics 2023, 15. [CrossRef]
- Espirito Santo, B.; Santana, L.F.; Kato Junior, W.H.; de Araújo, F.O.; Bogo, D.; Freitas, K.C.; Guimarães, R.C.A.; Hiane, P.A.; Pott, A.; Filiú, W.F.O.; et al. Medicinal Potential of Garcinia Species and Their Compounds. Molecules 2020, 25. [CrossRef]
- Rauf, A.; Uddin, G.; Patel, S.; Khan, A.; Halim, S.A.; Bawazeer, S.; Ahmad, K.; Muhammad, N.; Mubarak, M.S. Diospyros, an under-utilized, multi-purpose plant genus: A review. Biomedicine & Pharmacotherapy 2017, 91, 714-730. [CrossRef]
- Sudradjat, S.E.; Timotius, K.H. Pharmacological properties and phytochemical components of Elaeocarpus: A comparative study. Phytomedicine Plus 2022, 2, 100365. [CrossRef]
- Nisar, M.; Ali, S.; Qaisar, M.; Gilani, S.N.; Shah, M.R.; Khan, I.; Ali, G. Antifungal activity of bioactive constituents and bark extracts of Rhododendron arboreum. ||| Bangladesh Journal of Pharmacology 2013, 8, 218-222. [CrossRef]
- Sadgrove, N.J.; Padilla-González, G.F.; Telford, I.R.H.; Greatrex, B.W.; Jones, G.L.; Andrew, R.; Bruhl, J.J.; Langat, M.K.; Melnikovova, I.; Fernandez-Cusimamani, E. Prostanthera (Lamiaceae) as a 'Cradle of Incense': Chemophenetics of Rare Essential Oils from Both New and Forgotten Australian 'Mint Bush' Species. Plants (Basel) 2020, 9. [CrossRef]
- Lassak, E.V.; McCarthy, T. Australian medicinal plants. (No Title) 1983.
- Wang, J.; Su, B.; Jiang, H.; Cui, N.; Yu, Z.; Yang, Y.; Sun, Y. Traditional uses, phytochemistry and pharmacological activities of the genus Cinnamomum (Lauraceae): A review. Fitoterapia 2020, 146, 104675. [CrossRef]
- Salleh, W.M.N.H.W.; Farediah, A.; Khong, H.Y.; Zulkifli, R. A Review of Endiandric Acid Analogues. International Journal of Pharmacognosy and Phytochemical Research 2015, 7, 844-856.
- Cock, I.E. Medicinal and aromatic plants—Australia. In Ethnopharmacology Section, Biological, Physiological and Health Sciences, Encyclopedia of Life Support Systems (EOLSS), Developed under the Auspices of the UNESCO; EOLSS Publishers: Oxford, UK, 2011. Available online: http://www.eolss.net (accessed on 30 April 2022).
- Caputo, L.; Smeriglio, A.; Trombetta, D.; Cornara, L.; Trevena, G.; Valussi, M.; Fratianni, F.; De Feo, V.; Nazzaro, F. Chemical Composition and Biological Activities of the Essential Oils of Leptospermum petersonii and Eucalyptus gunnii. Front Microbiol 2020, 11, 409. [CrossRef]
- Riley, M. Māori healing and herbal: New Zealand ethnobotanical sourcebook; Viking Sevenseas NZ: 1994.
- Oktavia, D.; Pratiwi, S.D.; Munawaroh, S.; Hikmat, A.; Hilwan, I. The potential of medicinal plants from heath forest: Local knowledge from Kelubi Village, Belitung Island, Indonesia. Biodiversitas Journal of Biological Diversity 2022, 23. [CrossRef]
- Sharifi-Rad, J.; Quispe, C.; Bouyahya, A.; El Menyiy, N.; El Omari, N.; Shahinozzaman, M.; Ara Haque Ovey, M.; Koirala, N.; Panthi, M.; Ertani, A.; et al. Ethnobotany, Phytochemistry, Biological Activities, and Health-Promoting Effects of the Genus Bulbophyllum. Evid Based Complement Alternat Med 2022, 2022, 6727609. [CrossRef]
- Cakova, V.; Bonte, F.; Lobstein, A. Dendrobium: Sources of Active Ingredients to Treat Age-Related Pathologies. Aging Dis 2017, 8, 827-849. [CrossRef]
- Liang, W.; Guo, X.; Nagle, D.G.; Zhang, W.-D.; Tian, X.-H. Genus Liparis: A review of its traditional uses in China, phytochemistry and pharmacology. Journal of Ethnopharmacology 2019, 234, 154-171. [CrossRef]
- Ware, I.; Franke, K.; Hussain, H.; Morgan, I.; Rennert, R.; Wessjohann, L.A. Bioactive Phenolic Compounds from Peperomia obtusifolia. Molecules 2022, 27. [CrossRef]
- Al-Madhagi, W.M.; Mohd Hashim, N.; Awad Ali, N.A.; Alhadi, A.A.; Abdul Halim, S.N.; Othman, R. -Chemical profiling and biological activity of Peperomia blanda (Jacq.) Kunth. PeerJ 2018, 6, e4839. [CrossRef]
- Inostroza-Blancheteau, C.; Sandoval, Y.; Reyes-Díaz, M.; Tighe-Neira, R.; González-Villagra, J. Phytochemical characterization and antioxidant properties of Prumnopitys andina fruits in different ripening stages in southern Chile. Chilean journal of agricultural research 2022, 82, 285-293. [CrossRef]
- Tlau, L.; Lalawmpuii, L. Commonly used medicinal plants in N. Mualcheng, Mizoram, India. Science Vision 2020, 20, 156-161. [CrossRef]
- Ray, S.; Saini, M.K. Impending threats to the plants with medicinal value in the Eastern Himalayas Region: An analysis on the alternatives to its non-availability. Phytomedicine Plus 2022, 2, 100151. [CrossRef]
- Cock, I.E. Medicinal and aromatic plants–Australia. Ethnopharmacology, Encyclopedia of Life Support Systems (EOLSS) 2011. [CrossRef]
- Palombo, E.A.; Semple, S.J. Antibacterial activity of traditional Australian medicinal plants. Journal of ethnopharmacology 2001, 77, 151-157. [CrossRef]
- Awang-Jamil, Z.; Basri, A.; Ahmad, N.; Taha, H. Phytochemical analysis, antimicrobial and antioxidant activities of Aidia borneensis leaf extracts. Journal of Applied Biology & Biotechnology 2019, 7. [CrossRef]
- Singh, B.; Sharma, R.A. Indian Morinda species: A review. Phytotherapy Research 2020, 34, 924-1007. [CrossRef]
- Baliga, M.S.; Kurian, P.J. Ixora coccinea Linn.: Traditional uses, phytochemistry and pharmacology. Chinese Journal of Integrative Medicine 2012, 18, 72-79. [CrossRef]
- Hossain, M.J.; Maliha, F.; Hawlader, M.B.; Farzana, M.; Rashid, M.A. Ethnomedicinal uses, phytochemistry, pharmacology and toxicological aspects of genus Wendlandia: an overview. Journal of Bangladesh Academy of Sciences 2023. [CrossRef]
- Chidambaram, K.; Alqahtani, T.; Alghazwani, Y.; Aldahish, A.; Annadurai, S.; Venkatesan, K.; Dhandapani, K.; Thilagam, E.; Venkatesan, K.; Paulsamy, P.; et al. Medicinal Plants of Solanum Species: The Promising Sources of Phyto-Insecticidal Compounds. J Trop Med 2022, 2022, 4952221. [CrossRef]
- Badoni, R.; Semwal, D.K.; Kothiyal, S.K.; Rawat, U. Chemical constituents and biological applications of the genus Symplocos. J Asian Nat Prod Res 2010, 12, 1069-1080. [CrossRef]
- Ahmad, R.; Khairul Nizam Mazlan, M.; Firdaus Abdul Aziz, A.; Mohd Gazzali, A.; Amir Rawa, M.S.; Wahab, H.A. Phaleria macrocarpa (Scheff.) Boerl.: An updated review of pharmacological effects, toxicity studies, and separation techniques. Saudi pharmaceutical journal : SPJ : the official publication of the Saudi Pharmaceutical Society 2023, 31, 874-888. [CrossRef]
- Mohanty, S. Bioactive properties of Australian native fruits, Tasmannia lanceolata and Terminalia ferdinandiana: The Characterization of their Active Compounds. Griffith University Australia 2016.
- POWO. Plants of the World Online. Facilitated by the Royal Botanic Gardens, Kew. Published on the Internet; http://www.plantsoftheworldonline.org/. 2023.
- Jones, D.L.; Hopley, T.; Duffy, S.M. Australian Tropical Rainforest Orchids. 2010.
- QLD. Nature Conservation Act 1992. 2017.
- Rivas-Ubach, A.; Pérez-Trujillo, M.; Sardans, J.; Gargallo-Garriga, A.; Parella, T.; Peñuelas, J. Ecometabolomics: optimized NMR-based method. Methods Ecol. Evol. 4, 464–473. 2013. [CrossRef]
- Rivas-Ubach, A.; Peñuelas, J.; Hódar, J.A.; Oravec, M.; Paša-Tolić, L.; Urban, O.; Sardans, J. We are what we eat: A stoichiometric and ecometabolomic study of caterpillars feeding on two pine subspecies of Pinus sylvestris. Int. J. Mol. Sci. 2018, 20, 59. [CrossRef]
- Allevato, D.M.; Kiyota, E.; Mazzafera, P.; Nixon, K.C. Ecometabolomic analysis of wild populations of Pilocarpus pennatifolius (Rutaceae) using unimodal analyses. Frontiers in plant science 2019, 10, 258. [CrossRef]
- Berini, J.L.; Brockman, S.A.; Hegeman, A.D.; Reich, P.B.; Muthukrishnan, R.; Montgomery, R.A.; Forester, J.D. Combinations of abiotic factors differentially alter production of plant secondary metabolites in five woody plant species in the boreal-temperate transition zone. Frontiers in plant science 2018, 9, 1257. [CrossRef]
- Steinbauer, M.J.; Grytnes, J.-A.; Jurasinski, G.; Kulonen, A.; Lenoir, J.; Pauli, H.; Rixen, C.; Winkler, M.; Bardy-Durchhalter, M.; Barni, E. Accelerated increase in plant species richness on mountain summits is linked to warming. Nature 2018, 556, 231-234. [CrossRef]
- Lavola, A.; Julkunen-Tiitto, R.; Aphalo, P.; de la Rosa, T.; Lehto, T. The effect of UV-B radiation on UV-absorbing secondary metabolites in birch seedlings grown under simulated forest soil conditions. The New Phytologist 1997, 137, 617-621.
- Salam, U.; Ullah, S.; Tang, Z.H.; Elateeq, A.A.; Khan, Y.; Khan, J.; Khan, A.; Ali, S. Plant Metabolomics: An Overview of the Role of Primary and Secondary Metabolites against Different Environmental Stress Factors. Life (Basel) 2023, 13. [CrossRef]
- Sallas, L.; Luomala, E.-M.; Utriainen, J.; Kainulainen, P.; Holopainen, J.K. Contrasting effects of elevated carbon dioxide concentration and temperature on Rubisco activity, chlorophyll fluorescence, needle ultrastructure and secondary metabolites in conifer seedlings. Tree Physiology 2003, 23, 97-108. [CrossRef]
- Večeřová, K.; Klem, K.; Veselá, B.; Holub, P.; Grace, J.; Urban, O. Combined Effect of Altitude, Season and Light on the Accumulation of Extractable Terpenes in Norway Spruce Needles. Forests 2021, 12. [CrossRef]
- Yeshi, K.; Crayn, D.; Ritmejeryte, E.; Wangchuk, P. Plant Secondary Metabolites Produced in Response to Abiotic Stresses Has Potential Application in Pharmaceutical Product Development. Molecules 2022, 27. [CrossRef]
- Pinto, D.M.; Blande, J.D.; Souza, S.R.; Nerg, A.M.; Holopainen, J.K. Plant volatile organic compounds (VOCs) in ozone (O3) polluted atmospheres: the ecological effects. J Chem Ecol 2010, 36, 22-34. [CrossRef]
- Schneider, G.F.; Coley, P.D.; Younkin, G.C.; Forrister, D.L.; Mills, A.G.; Kursar, T.A. Phenolics lie at the centre of functional versatility in the responses of two phytochemically diverse tropical trees to canopy thinning. J Exp Bot 2019, 70, 5853-5864. [CrossRef]
- Pinasseau, L.; Vallverdu-Queralt, A.; Verbaere, A.; Roques, M.; Meudec, E.; Le Cunff, L.; Peros, J.P.; Ageorges, A.; Sommerer, N.; Boulet, J.C.; et al. Cultivar Diversity of Grape Skin Polyphenol Composition and Changes in Response to Drought Investigated by LC-MS Based Metabolomics. Front Plant Sci 2017, 8, 1826. [CrossRef]
- Sampaio, B.L.; Edrada-Ebel, R.; Da Costa, F.B. Effect of the environment on the secondary metabolic profile of Tithonia diversifolia: a model for environmental metabolomics of plants. Sci 2016, 6, 29265. [CrossRef]
- Niinemets, Ü. Uncovering the hidden facets of drought stress: secondary metabolites make the difference. Tree Physiology 2016, 36, 129-132. [CrossRef]
- Afzal, S.F.; Yar, A.K.; Ullah, R.H.; Ali, B.G.; Ali, J.S.; Ahmad, J.S.; Fu, S. Impact of drought stress on active secondary metabolite production in Cichorium intybus roots. J Appl Environ Biol Sci 2017, 7, 39-43.
- Punia, H.; Tokas, J.; Malik, A.; Bajguz, A.; El-Sheikh, M.A.; Ahmad, P. Ascorbate-Glutathione Oxidant Scavengers, Metabolome Analysis and Adaptation Mechanisms of Ion Exclusion in Sorghum under Salt Stress. Int J Mol Sci 2021, 22. [CrossRef]
- Singiri, J.R.; Swetha, B.; Sikron-persi, N.; Grafi, G. Differential response to single and combined salt and heat stresses: Impact on accumulation of proteins and metabolites in dead pericarps of Brassica juncea. Int. J. Mol. Sci. 2021, 22. [CrossRef]
- Munns, R.; Gilliham, M. Salinity tolerance of crops–what is the cost? New phytologist 2015, 208, 668-673.
- Goche, T.; Shargie, N.G.; Cummins, I.; Brown, A.P.; Chivasa, S.; Ngara, R. Comparative physiological and root proteome analyses of two sorghum varieties responding to water limitation. Sci. Rep. 2020, 10, 11835. [CrossRef]
- Xiao, Q.; Mu, X.; Liu, J.; Li, B.; Liu, H.; Zhang, B.; Xiao, P. Plant metabolomics: a new strategy and tool for quality evaluation of Chinese medicinal materials. Chinese Medicine 2022, 17, 45. [CrossRef]
- Guy, C.; Kopka, J.; Moritz, T. Plant metabolomics coming of age. Physiol Plant 2008, 132, 113-116. [CrossRef]
- Hall, R.; Beale, M.; Fiehn, O.; Hardy, N.; Sumner, L.; Bino, R. Plant metabolomics: the missing link in functional genomics strategies. Plant Cell 2002, 14, 1437-1440. [CrossRef]
- Neilson, E.H.; Goodger, J.Q.; Woodrow, I.E.; Møller, B.L. Plant chemical defense: at what cost? Trends in plant science 2013, 18, 250-258.
- Kesselmeier, J.; Staudt, M. Biogenic volatile organic compounds (VOC): an overview on emission, physiology and ecology. Journal of atmospheric chemistry 1999, 33, 23-88.
- Pichersky, E.; Gang, D.R. Genetics and biochemistry of secondary metabolites in plants: an evolutionary perspective. Trends Plant Sci 2000, 5, 439-445. [CrossRef]
- Fiehn, O. Combining genomics, metabolome analysis, and biochemical modelling to understand metabolic networks. Comp Funct Genomics 2001, 2, 155-168. [CrossRef]
- Kim, H.K.; Verpoorte, R. Sample preparation for plant metabolomics. Phytochem Anal 2010, 21, 4-13. [CrossRef]
- Ritmejeryte, E.; Ryan, R.Y.M.; Byatt, B.J.; Peck, Y.; Yeshi, K.; Daly, N.L.; Zhao, G.; Crayn, D.; Loukas, A.; Pyne, S.G.; et al. Anti-inflammatory properties of novel galloyl glucosides isolated from the Australian tropical plant Uromyrtus metrosideros. Chem Biol Interact 2022, 368, 110124. [CrossRef]
- Garrison, M.S.; Irvine, A.K.; Setzer, W.N. Chemical composition of the resin essential oil from Agathis atropurpurea from North Queensland, Australia. Am. J. Essent. Oils Nat. Prod 2016, 4, 4-5.
- Risner, D.; Marco, M.L.; Pace, S.A.; Spang, E.S. The Potential Production of the Bioactive Compound Pinene Using Whey Permeate. Processes 2020, 8, 263. [CrossRef]
- Salehi, B.; Upadhyay, S.; Erdogan Orhan, I.; Kumar Jugran, A.; S, L.D.J.; D, A.D.; Sharopov, F.; Taheri, Y.; Martins, N.; Baghalpour, N.; et al. Therapeutic Potential of α- and β-Pinene: A Miracle Gift of Nature. Biomolecules 2019, 9. [CrossRef]
- Rivas da Silva, A.C.; Lopes, P.M.; Barros de Azevedo, M.M.; Costa, D.C.; Alviano, C.S.; Alviano, D.S. Biological activities of α-pinene and β-pinene enantiomers. Molecules 2012, 17, 6305-6316. [CrossRef]
- Türkez, H.; Celik, K.; Toğar, B. Effects of copaene, a tricyclic sesquiterpene, on human lymphocytes cells in vitro. Cytotechnology 2014, 66, 597-603. [CrossRef]
- Wollenweber, E.; Dörr, M.; Rozefelds, A.C.; Minchin, P.; Forster, P.I. Variation in flavonoid exudates in Eucryphia species from Australia and South America. Biochemical Systematics and Ecology 2000, 28, 111-118. [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Forster, P.I. The Leaf Oils of the Australian Species ofCinnamomum(Lauraceae). Journal of Essential Oil Research 2001, 13, 332-335. [CrossRef]
- Balahbib, A.; El Omari, N.; Hachlafi, N.E.L.; Lakhdar, F.; El Menyiy, N.; Salhi, N.; Mrabti, H.N.; Bakrim, S.; Zengin, G.; Bouyahya, A. Health beneficial and pharmacological properties of p-cymene. Food and Chemical Toxicology 2021, 153, 112259. [CrossRef]
- Han, N.R.; Moon, P.D.; Ryu, K.J.; Jang, J.B.; Kim, H.M.; Jeong, H.J. β-eudesmol suppresses allergic reactions via inhibiting mast cell degranulation. Clin Exp Pharmacol Physiol 2017, 44, 257-265. [CrossRef]
- Tshering, G.; Pimtong, W.; Plengsuriyakarn, T.; Na-Bangchang, K. Anti-angiogenic effects of beta-eudesmol and atractylodin in developing zebrafish embryos. Comp Biochem Physiol C Toxicol Pharmacol 2021, 243, 108980. [CrossRef]
- Brophy, J.J.; Forster, P.I.; Goldsack, R.J. Coconut Laurels: The Leaf Essential Oils from Four Endemic Australian Cryptocarya Species: C. bellendenkerana, C. cocosoides, C. cunninghamii and C. lividula (Lauraceae). Nat Prod Commun 2016, 11, 255-258.
- Anandakumar, P.; Kamaraj, S.; Vanitha, M.K. D-limonene: A multifunctional compound with potent therapeutic effects. J Food Biochem 2021, 45, e13566. [CrossRef]
- Vieira, A.J.; Beserra, F.P.; Souza, M.C.; Totti, B.M.; Rozza, A.L. Limonene: Aroma of innovation in health and disease. Chem Biol Interact 2018, 283, 97-106. [CrossRef]
- Thangaleela, S.; Sivamaruthi, B.S.; Kesika, P.; Tiyajamorn, T.; Bharathi, M.; Chaiyasut, C. A Narrative Review on the Bioactivity and Health Benefits of Alpha-Phellandrene. Scientia Pharmaceutica 2022, 90, 57. [CrossRef]
- Ferraz, R.P.; Cardoso, G.M.; da Silva, T.B.; Fontes, J.E.; Prata, A.P.; Carvalho, A.A.; Moraes, M.O.; Pessoa, C.; Costa, E.V.; Bezerra, D.P. Antitumour properties of the leaf essential oil of Xylopia frutescens Aubl. (Annonaceae). Food Chem 2013, 141, 196-200. [CrossRef]
- Minh, P.T.H.; Tuan, N.T.; Van, N.T.H.; Bich, H.T.; Lam, D.T. Chemical Composition and Biological Activities of Essential Oils of Four Asarum Species Growing in Vietnam. Molecules 2023, 28. [CrossRef]
- Qin, Y.; Zhang, J.; Song, D.; Duan, H.; Li, W.; Yang, X. Novel (E)-β-Farnesene Analogues Containing 2-Nitroiminohexahydro-1,3,5-triazine: Synthesis and Biological Activity Evaluation. Molecules 2016, 21, 825.
- brophy2000.pdf.
- Ryu, Y.; Lee, D.; Jung, S.H.; Lee, K.J.; Jin, H.; Kim, S.J.; Lee, H.M.; Kim, B.; Won, K.J. Sabinene Prevents Skeletal Muscle Atrophy by Inhibiting the MAPK-MuRF-1 Pathway in Rats. Int J Mol Sci 2019, 20. [CrossRef]
- Cordeiro, L.; Figueiredo, P.; Souza, H.; Sousa, A.; Andrade-Júnior, F.; Medeiros, D.; Nóbrega, J.; Silva, D.; Martins, E.; Barbosa-Filho, J.; et al. Terpinen-4-ol as an Antibacterial and Antibiofilm Agent against Staphylococcus aureus. Int J Mol Sci 2020, 21. [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Forster, P.I. The Essential Oils of the Australian Species of Uromyrtus (Myrtaceae). Flavour and Fragrance Journal 1996, 11, 133-138. [CrossRef]
- Hong, E.Y.; Kim, T.Y.; Hong, G.U.; Kang, H.; Lee, J.Y.; Park, J.Y.; Kim, S.C.; Kim, Y.H.; Chung, M.H.; Kwon, Y.I.; et al. Inhibitory Effects of Roseoside and Icariside E4 Isolated from a Natural Product Mixture (No-ap) on the Expression of Angiotensin II Receptor 1 and Oxidative Stress in Angiotensin II-Stimulated H9C2 Cells. Molecules 2019, 24. [CrossRef]
- Yajima, A.; Oono, Y.; Nakagawa, R.; Nukada, T.; Yabuta, G. A simple synthesis of four stereoisomers of roseoside and their inhibitory activity on leukotriene release from mice bone marrow-derived cultured mast cells. Bioorg Med Chem 2009, 17, 189-194. [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Forster, P.I. Chemistry of the Australian Gymnosperms Part VIII. The Leaf Oil ofPrumnopitys ladei(Podocarpaceae). Journal of Essential Oil Research 2006, 18, 212-214. [CrossRef]
- Dahham, S.S.; Tabana, Y.M.; Iqbal, M.A.; Ahamed, M.B.; Ezzat, M.O.; Majid, A.S.; Majid, A.M. The Anticancer, Antioxidant and Antimicrobial Properties of the Sesquiterpene β-Caryophyllene from the Essential Oil of Aquilaria crassna. Molecules 2015, 20, 11808-11829. [CrossRef]
- Francomano, F.; Caruso, A.; Barbarossa, A.; Fazio, A.; La Torre, C.; Ceramella, J.; Mallamaci, R.; Saturnino, C.; Iacopetta, D.; Sinicropi, M.S. β-Caryophyllene: A Sesquiterpene with Countless Biological Properties. Applied Sciences 2019, 9, 5420. [CrossRef]
- Fidyt, K.; Fiedorowicz, A.; Strządała, L.; Szumny, A. β-caryophyllene and β-caryophyllene oxide-natural compounds of anticancer and analgesic properties. Cancer Med 2016, 5, 3007-3017. [CrossRef]
- Brophy, J.J.; Goldsack, R.J.; Forster, P.I. The Leaf Oils of the Australian Species of Flindersia (Rutaceae). Journal of Essential Oil Research 2005, 17, 388-395. [CrossRef]
- Robertson, L.P.; Hall, C.R.; Forster, P.I.; Carroll, A.R. Alkaloid diversity in the leaves of Australian Flindersia (Rutaceae) species driven by adaptation to aridity. Phytochemistry 2018, 152, 71-81. [CrossRef]
- Robertson, L.P.; Duffy, S.; Wang, Y.; Wang, D.; Avery, V.M.; Carroll, A.R. Pimentelamines A-C, Indole Alkaloids Isolated from the Leaves of the Australian Tree Flindersia pimenteliana. J Nat Prod 2017, 80, 3211-3217. [CrossRef]
- Robertson, L.P.; Lucantoni, L.; Avery, V.M.; Carroll, A.R. Antiplasmodial Bis-Indole Alkaloids from the Bark of Flindersia pimenteliana. Planta Med 2020, 86, 19-25. [CrossRef]
- Resch, M.; Steigel, A.; Chen, Z.-l.; Bauer, R. 5-Lipoxygenase and Cyclooxygenase-1 Inhibitory Active Compounds from Atractylodes lancea. J. Nat. Prod. 1998, 61, 347-350. [CrossRef]
- Mu, K.; Zhang, J.; Feng, X.; Zhang, D.; Li, K.; Li, R.; Yang, P.; Mao, S. Sedative-hypnotic effects of Boropinol-B on mice via activation of GABAA receptors. J Pharm Pharmacol 2023, 75, 57-65. [CrossRef]
- Hu, Q.; Luo, L.; Yang, P.; Mu, K.; Yang, H.; Mao, S. Neuroprotection of boropinol-B in cerebral ischemia-reperfusion injury by inhibiting inflammation and apoptosis. Brain Research 2023, 1798, 148132. [CrossRef]
- Liu, J.H.; Zschocke, S.; Reininger, E.; Bauer, R. Inhibitory effects of Angelica pubescens f. biserrata on 5-lipoxygenase and cyclooxygenase. Planta Med 1998, 64, 525-529. [CrossRef]
- Resch, M.; Steigel, A.; Chen, Z.L.; Bauer, R. 5-Lipoxygenase and cyclooxygenase-1 inhibitory active compounds from Atractylodes lancea. J Nat Prod 1998, 61, 347-350. [CrossRef]
- Yeshi, K.; Ruscher, R.; Miles, K.; Crayn, D.; Liddell, M.; Wangchuk, P. Antioxidant and Anti-Inflammatory Activities of Endemic Plants of the Australian Wet Tropics. Plants (Basel) 2022, 11. [CrossRef]
- Yeshi, K.; Wangchuk, P. Bush Medicinal Plants of the Australian Wet Tropics and Their Biodiscovery Potential. In Bioprospecting of Tropical Medicinal Plants, Arunachalam, K., Yang, X., Puthanpura Sasidharan, S., Eds.; Springer Nature Switzerland: Cham, 2023; pp. 357-379.
- Giménez-Bastida, J.A.; González-Sarrías, A.; Laparra-Llopis, J.M.; Schneider, C.; Espín, J.C. Targeting Mammalian 5-Lipoxygenase by Dietary Phenolics as an Anti-Inflammatory Mechanism: A Systematic Review. Int J Mol Sci 2021, 22. [CrossRef]
- Rådmark, O.; Samuelsson, B. 5-Lipoxygenase: mechanisms of regulation1. J. Lipid Res. 2009, 50, S40-S45. [CrossRef]
- Barbier de Reuille, P.; Routier-Kierzkowska, A.-L.; Kierzkowski, D.; Bassel, G.W.; Schüpbach, T.; Tauriello, G.; Bajpai, N.; Strauss, S.; Weber, A.; Kiss, A.; et al. MorphoGraphX: A platform for quantifying morphogenesis in 4D. eLife 2015, 4, e05864. [CrossRef]
- Fernandez, R.; Das, P.; Mirabet, V.; Moscardi, E.; Traas, J.; Verdeil, J.-L.; Malandain, G.; Godin, C. Imaging plant growth in 4D: robust tissue reconstruction and lineaging at cell resolution. Nature Methods 2010, 7, 547-553. [CrossRef]
- Perez de Souza, L.; Alseekh, S.; Scossa, F.; Fernie, A.R. Ultra-high-performance liquid chromatography high-resolution mass spectrometry variants for metabolomics research. Nat Methods 2021, 18, 733-746. [CrossRef]
- Jamtsho, T.; Yeshi, K.; Perry, M.J.; Loukas, A.; Wangchuk, P. Approaches, Strategies and Procedures for Identifying Anti-Inflammatory Drug Lead Molecules from Natural Products. Pharmaceuticals 2024, 17, 283. [CrossRef]
- Markley, J.L.; Brüschweiler, R.; Edison, A.S.; Eghbalnia, H.R.; Powers, R.; Raftery, D.; Wishart, D.S. The future of NMR-based metabolomics. Curr Opin Biotechnol 2017, 43, 34-40. [CrossRef]
- Dunn, W.B.; Bailey, N.J.; Johnson, H.E. Measuring the metabolome: current analytical technologies. Analyst 2005, 130, 606-625. [CrossRef]
- Castrillo, J.I.; Hayes, A.; Mohammed, S.; Gaskell, S.J.; Oliver, S.G. An optimized protocol for metabolome analysis in yeast using direct infusion electrospray mass spectrometry. Phytochemistry 2003, 62, 929-937. [CrossRef]
- Maia, M.; Figueiredo, A.; Cordeiro, C.; Sousa Silva, M. FT-ICR-MS-based metabolomics: A deep dive into plant metabolism. Mass Spectrometry Reviews 2023, 42, 1535-1556. [CrossRef]
- Andrews, G.L.; Simons, B.L.; Young, J.B.; Hawkridge, A.M.; Muddiman, D.C. Performance characteristics of a new hybrid quadrupole time-of-flight tandem mass spectrometer (TripleTOF 5600). Analytical chemistry 2011, 83, 5442-5446. [CrossRef]
- Pelander, A.; Decker, P.; Baessmann, C.; Ojanperä, I. Evaluation of a high resolving power time-of-flight mass spectrometer for drug analysis in terms of resolving power and acquisition rate. Journal of the American Society for Mass Spectrometry 2011, 22, 379-385. [CrossRef]
- Ghaste, M.; Mistrik, R.; Shulaev, V. Applications of fourier transform ion cyclotron resonance (FT-ICR) and orbitrap based high resolution mass spectrometry in metabolomics and lipidomics. Int. J. Mol. Sci. 2016, 17, 816. [CrossRef]
- Glauser, G.; Veyrat, N.; Rochat, B.; Wolfender, J.-L.; Turlings, T.C. Ultra-high pressure liquid chromatography–mass spectrometry for plant metabolomics: A systematic comparison of high-resolution quadrupole-time-of-flight and single stage Orbitrap mass spectrometers. Journal of chromatography A 2013, 1292, 151-159. [CrossRef]
- Park, S.-G.; Mohr, J.P.; Anderson, G.A.; Bruce, J.E. Application of frequency multiple FT-ICR MS signal acquisition for improved proteome research. International journal of mass spectrometry 2021, 465, 116578. [CrossRef]
- Schuhmann, K.; Herzog, R.; Schwudke, D.; Metelmann-Strupat, W.; Bornstein, S.R.; Shevchenko, A. Bottom-up shotgun lipidomics by higher energy collisional dissociation on LTQ Orbitrap mass spectrometers. Analytical chemistry 2011, 83, 5480-5487. [CrossRef]
- Schuhmann, K.; Almeida, R.; Baumert, M.; Herzog, R.; Bornstein, S.R.; Shevchenko, A. Shotgun lipidomics on a LTQ Orbitrap mass spectrometer by successive switching between acquisition polarity modes. Journal of Mass Spectrometry 2012, 47, 96-104. [CrossRef]
- Allwood, J.W.; Parker, D.; Beckmann, M.; Draper, J.; Goodacre, R. Fourier Transform Ion Cyclotron Resonance mass spectrometry for plant metabolite profiling and metabolite identification. Methods Mol Biol 2012, 860, 157-176. [CrossRef]
- Barrow, M.P.; Burkitt, W.I.; Derrick, P.J. Principles of Fourier transform ion cyclotron resonance mass spectrometry and its application in structural biology. Analyst 2005, 130, 18-28. [CrossRef]
- Hiraoka, K. Fundamentals of mass spectrometry; Springer: 2013; Volume 8.
- Folli, G.S.; Souza, L.M.; Araújo, B.Q.; Romão, W.; Filgueiras, P.R. Estimating the intermediate precision in petroleum analysis by (±) electrospray ionization Fourier transform ion cyclotron resonance mass spectrometry. Rapid Communications in Mass Spectrometry 2020, 34, e8861. [CrossRef]
- Hughey, C.A.; Rodgers, R.P.; Marshall, A.G. Resolution of 11 000 compositionally distinct components in a single electrospray ionization Fourier transform ion cyclotron resonance mass spectrum of crude oil. Analytical Chemistry 2002, 74, 4145-4149. [CrossRef]
- Allwood, J.W.; De Vos, R.C.; Moing, A.; Deborde, C.; Erban, A.; Kopka, J.; Goodacre, R.; Hall, R.D. Plant metabolomics and its potential for systems biology research: Background concepts, technology, and methodology. Methods in enzymology 2011, 500, 299-336.
- Shahbazy, M.; Moradi, P.; Ertaylan, G.; Zahraei, A.; Kompany-Zareh, M. FTICR mass spectrometry-based multivariate analysis to explore distinctive metabolites and metabolic pathways: A comprehensive bioanalytical strategy toward time-course metabolic profiling of Thymus vulgaris plants responding to drought stress. Plant Science 2020, 290, 110257. [CrossRef]
- Janz, D.; Behnke, K.; Schnitzler, J.-P.; Kanawati, B.; Schmitt-Kopplin, P.; Polle, A. Pathway analysis of the transcriptome and metabolome of salt sensitive and tolerant poplar species reveals evolutionary adaption of stress tolerance mechanisms. BMC Plant Biology 2010, 10, 1-17. [CrossRef]
- Kaling, M.; Kanawati, B.; Ghirardo, A.; Albert, A.; Winkler, J.B.; Heller, W.; Barta, C.; Loreto, F.; SCHMITT-KOPPLIN, P.; SCHNITZLER, J.P. UV-B mediated metabolic rearrangements in poplar revealed by non-targeted metabolomics. Plant, cell & environment 2015, 38, 892-904.
- Fiehn, O. Metabolomics—The link between genotypes and phenotypes. Plant Mol Biol 2002, 48, 155–171.
- Silva, L.P.; Northen, T.R. Exometabolomics and MSI: deconstructing how cells interact to transform their small molecule environment. Curr Opin Biotechnol 2015, 34, 209-216. [CrossRef]
- Mapelli, V.; Olsson, L.; Nielsen, J. Metabolic footprinting in microbiology: methods and applications in functional genomics and biotechnology. Trends in Biotechnology 2008, 26, 490-497. [CrossRef]
- Kuzina, V.; Ekstrøm, C.T.; Andersen, S.B.; Nielsen, J.K.; Olsen, C.E.; Bak, S. Identification of defense compounds in Barbarea vulgaris against the herbivore Phyllotreta nemorum by an ecometabolomic approach. Plant physiology 2009, 151, 1977-1990. [CrossRef]
- Salek, R.M.; Steinbeck, C.; Viant, M.R.; Goodacre, R.; Dunn, W.B. The role of reporting standards for metabolite annotation and identification in metabolomic studies. GigaScience 2013, 2, 13. [CrossRef]
- Sumner, L.W.; Amberg, A.; Barrett, D.; Beale, M.H.; Beger, R.; Daykin, C.A.; Fan, T.W.; Fiehn, O.; Goodacre, R.; Griffin, J.L.; et al. Proposed minimum reporting standards for chemical analysis Chemical Analysis Working Group (CAWG) Metabolomics Standards Initiative (MSI). Metabolomics 2007, 3, 211-221. [CrossRef]
- Xu, Y.; Fu, X. Reprogramming of Plant Central Metabolism in Response to Abiotic Stresses: A Metabolomics View. Int. J. Mol. Sci. 2022, 23, 5716. [CrossRef]
Figure 1.
Map of Australia showing the State of Queensland and the Wet Tropics World Heritage Area (WTWHA) shaded in green. Tropical Montane Cloud Forest is restricted to the mountaintops of the WTWHA, typically areas above 900 m above sea level. A map depicting WTWHA was generated from the Wet Tropics Plan zoning map Edition 3.0 with the help of Wet Tropics Management Authority office, Queensland.
Figure 1.
Map of Australia showing the State of Queensland and the Wet Tropics World Heritage Area (WTWHA) shaded in green. Tropical Montane Cloud Forest is restricted to the mountaintops of the WTWHA, typically areas above 900 m above sea level. A map depicting WTWHA was generated from the Wet Tropics Plan zoning map Edition 3.0 with the help of Wet Tropics Management Authority office, Queensland.
Figure 2.
Climate-affected Australian montane cloud forest plants in the Wet Tropics World Heritage Area, Queensland: A) distribution, B) life form, C) family diversity, and D) medicinally important genus with species number.
Figure 2.
Climate-affected Australian montane cloud forest plants in the Wet Tropics World Heritage Area, Queensland: A) distribution, B) life form, C) family diversity, and D) medicinally important genus with species number.
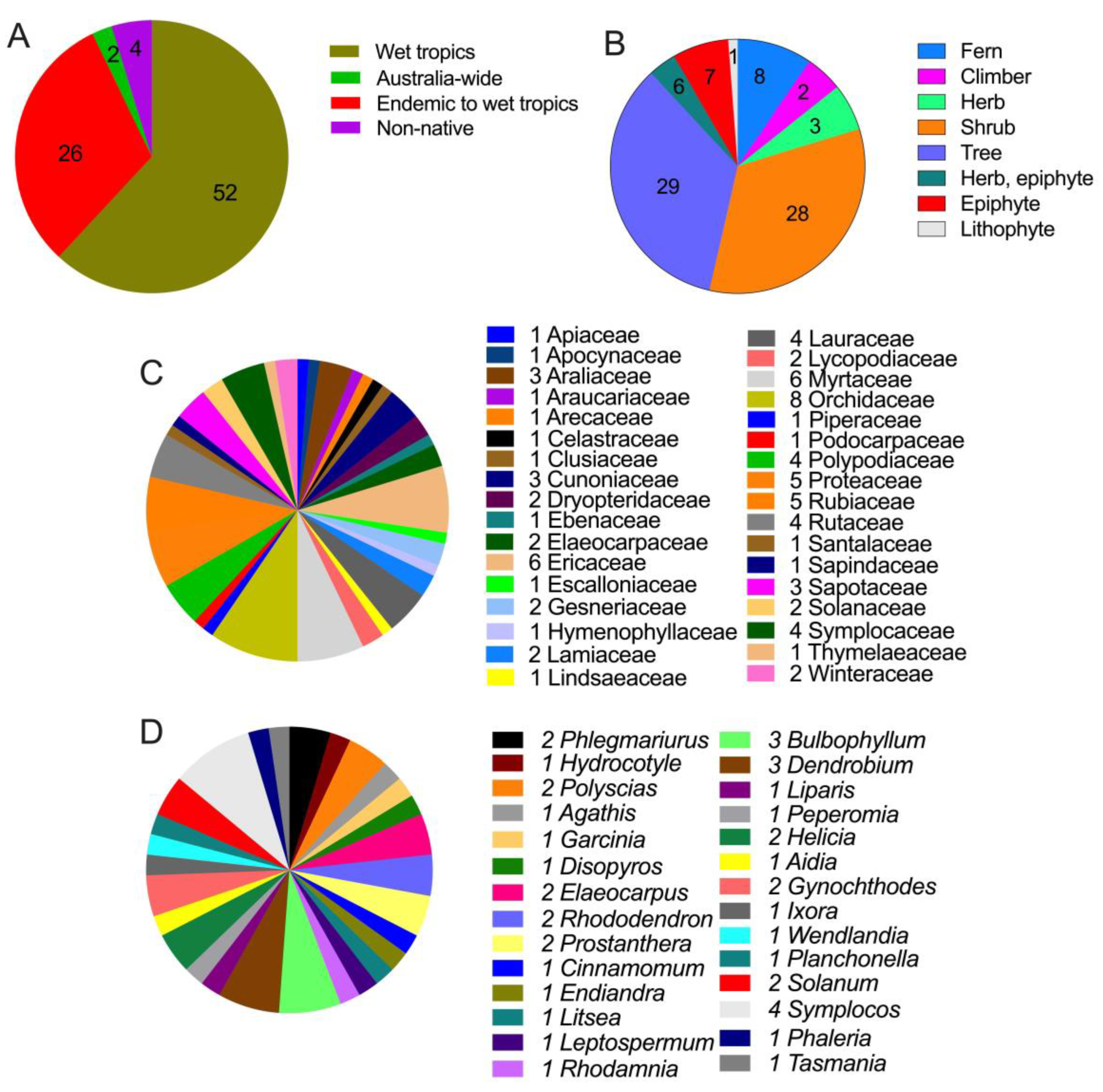
Figure 6.
Chemical structure of bioactive compounds isolated/identified from the climate-affected Australian montane cloud forest plants (also used medicinally) in the Wet Tropics World Heritage Area, Queensland.
Figure 6.
Chemical structure of bioactive compounds isolated/identified from the climate-affected Australian montane cloud forest plants (also used medicinally) in the Wet Tropics World Heritage Area, Queensland.
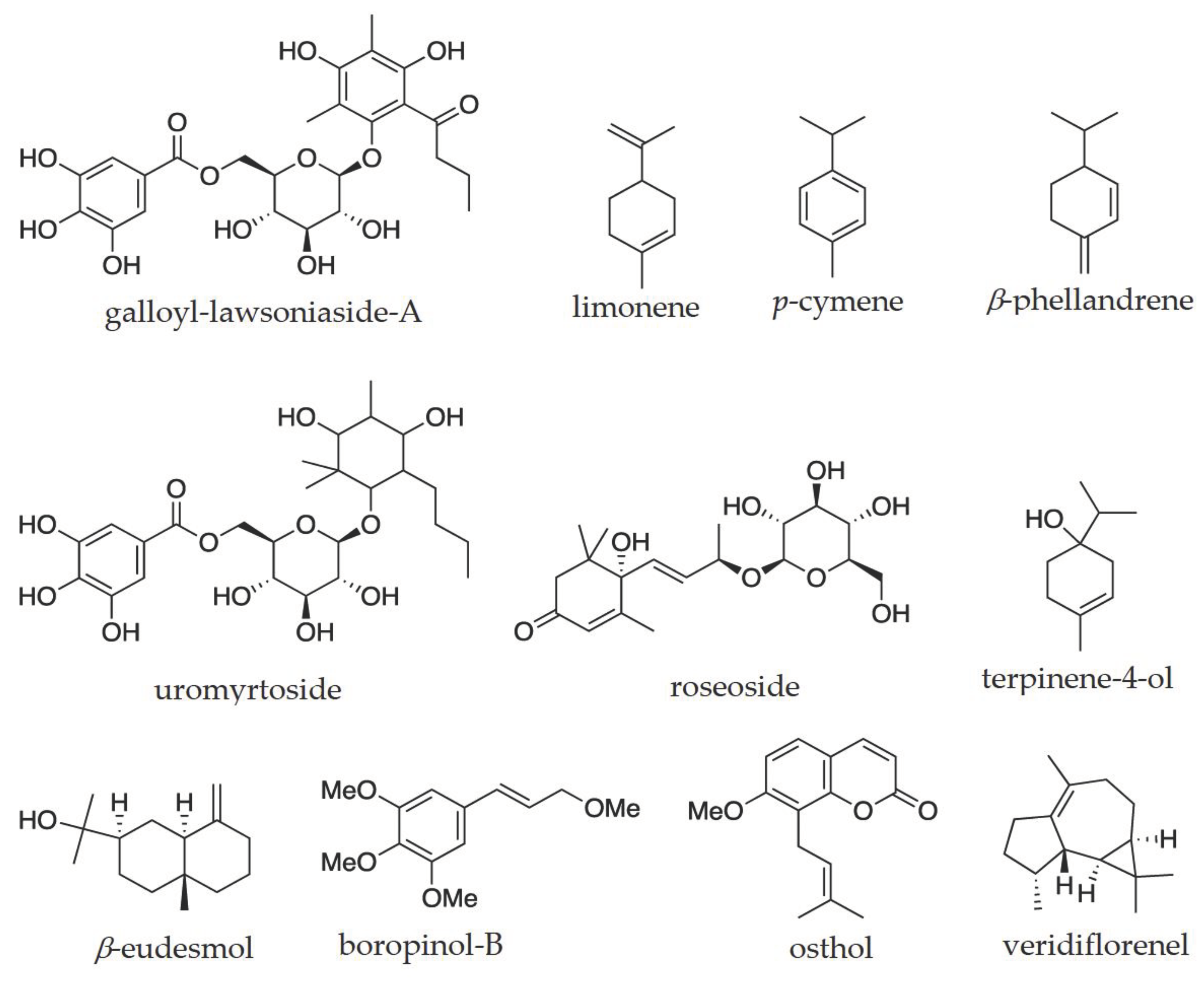
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



