
Submitted:
11 March 2024
Posted:
11 March 2024
You are already at the latest version
Abstract
Use of probiotic lactobacilli has been proposed as a strategy to mitigate damage associated to exposure to toxic metals. Their protective effect against cationic metal ions, such as those of mercury or lead, is believed to stem from their chelating and accumulating potential. However, their retention of anionic toxic metalloids, such as inorganic arsenic, is generally lower. Through the construction of mutants in phosphate transporter genes (pst) in Lactiplantibacillus plantarum and Lacticaseibacillus paracasei strains, coupled with arsenate [As(V)] uptake and toxicity assays, we determined that the incorporation of As(V), which structurally resembles phosphate, is likely facilitated by phosphate transporters. Surprisingly, inactivation in Lc. paracasei of PhoP, the tran-scriptional regulator of the two-component system PhoPR, a signal transducer involved in phosphate sensing, led to an increased resistance to arsenite [As(III)]. In comparison to the wild type, the phoP strain exhibited no differences in the ability to retain As(III), and there were no observed changes in the oxidation of As(III) to the less toxic As(V). These results reinforce the idea that specific transport, and not unspecific cell retention plays a role in As(V) biosorption by lactobacilli, while they reveal an unexpected phenotype for the lack of the pleiotropic regulator PhoP.
Keywords:
Lactiplantibacillus plantarum
; Lacticaseibacillus paracasei
; arsenate
; arsenite
; phosphate transport
; two-component system
; Pho
1. Introduction
Inorganic arsenic [iAs: arsenite, As(III) and arsenate, As(V)] represents a serious health threat and it is estimated that a population of more than 100 million people are exposed to elevated amounts of this metalloid throughout the world, mainly via drinking water [1]. Owing to the severe adverse effects of iAs, international agencies recommend reducing dietary exposure [2]. In situations where achieving this goal is not feasible (for example, populations with no access to uncontaminated water sources), employing alternative approaches to alleviate the effects of iAs exposure, such as strategies focused on diminishing its entry into the systemic circulation or reducing its toxicity, may contribute to minimize potential damage.
Intestinal damage caused by heavy metals can be responsible in some instances for other systemic pathologies or contribute to increased entry of the toxicant in the organism. Strains of lactic acid bacteria have been postulated as agents (probiotics) aimed to prevent this adverse effect [3,4,5]. Selected strains of lactobacilli have been shown to alleviate inflammation, oxidative stress, increased intestinal permeability and metal organ accumulation upon oral exposure through food or drink in animal models [6,7,8]. However, assays in human populations are still very scarce [9,10]. Chelation or sequestration of cationic toxic metals by negatively-charged molecules of the cellular envelope has been hypothesized as a mechanism responsible for the observed positive effects [11], although activation of anti-inflammatory and antioxidant pathways of the host possibly also play a role [6]. In the case of iAs, which is an anionic pollutant, protective effects of probiotics on animal models have also been determined [12,13,14]. Strains of lactobacilli have been reported to complex iAs and to remove it from aqueous solutions with different efficacies [15]. However, owing to its anionic nature, the chelation/surface binding of iAs is supposed to play a less important role compared to metallic cations, and other processes in addition to iAS sequestration are probably at play.
The sequestration or uptake of chemical contaminants by microorganisms can, in some cases, reside in microbial internalization mediated by specific transport systems [4]. As(V), which structurally resembles phosphate, can possibly enter the cells by the same natural mechanisms as phosphate [16]. The most studied phosphate uptake system in bacteria is the widely distributed Pst ABC-type high-affinity phosphate transporter [17]. This system is composed by a surface bound phosphate-binding protein (PstS), transmembrane transport subunits (PstCA) and cytoplasmic ATPase subunits (PstB) which energize transport. In many instances an auxiliary gene, phoU, encode a small regulatory protein which in the model organism E. coli has been shown to link phosphate uptake by the ABC system to activity of the two-component system (TCS) PhoPR [18]. Under low-phosphate conditions, the conformational changes derived via transport through PstCA and ATP-hydrolysis by PstB are sensed by PhoU, that interacts with the sensor kinase PhoR, triggering its phosphorylation function on PhoP, the response regulator of the TCS [19]. PhoP controls the expression of phosphate-regulated genes, including the pst genes, activating their transcription under conditions of low phosphate availability [18].
A PhoPR TCS has been studied in Lacticaseibacillus paracasei BL23, showing that mutating the response regulator phoP resulted in slow growth and acid sensitivity in MRS medium [20]. This TCS was adjacent to a pstSCAB1B2-phoU gene cluster, but this operon has not been studied in any member of the Lactobacillaceae family. In this study, our aim was to elucidate the mechanisms responsible for the binding of iAs in lactobacilli. Through the construction of mutants, we demonstrated that As(V) is likely taken up by the Pst ABC transport system in Lactiplantibacillus plantarum. Additionally, we made an unprecedented finding, reporting a link between the activity of the PhoP response regulator and As(III) sensitivity in Lacticaseibacillus paracasei.
2. Results
2.1. As(V) and As(III) Incorporation by Lactobacilli Strains
The low ability of some lactobacilli strains to take up or retain the anionic forms of inorganic metalloid As has been previously described [21]. The percentages of retention for a collection of lactobacilli after incubation of high-density cell suspensions in phosphate buffered saline (PBS) with As(V) or As(III) or with the organic arsenic form dimethylarsinic acid (DMA) were below 10% in all cases (Table 1). For As(V), this last circumstance could be attributed to the fact that phosphate structurally resembles this As species and may interfere with its incorporation [21]. Furthermore, previous studies reported the need of live cells for As(V) incorporation, while this was not a requisite for As(III) or for other metals such as Hg [21,22]
Therefore, we hypothesized that incorporation of As(V) in lactobacilli could take place through specific phosphate transporter(s). In bacteria, a high affinity-low capacity phosphate transporter of the ABC type family is widely distributed (pstSCAB). In many instances, the encoding genes are clustered with the genes encoding the TCS PhoPR, putatively involved in sensing the extracellular phosphate concentration and controlling expression of phosphate-regulated genes. Inspection of available genome sequences within the Lactobacillaceae family, showed this typical organization (supplementary Figure S1; [25]), in which a phoU gene also follows a pstSCAB cluster. This operon generally carried two tandem copies of the ATPase-encoding gene pstB (pstSCAB1B2) and phoPR genes were found upstream. In order to assess the contribution of this system to As(V) incorporation, we decided to construct mutants in the transmembrane phosphate permease PstC in Lp. plantarum Lpp+ and in the reference Lp. plantarum strain WCFS1 [23]. An insertional mutant in pstC was also constructed in the model lactic acid bacterium Lacticaseibacillus paracasei BL23 [26]. In addition, the phoP transcriptional regulator and phoU were also targeted with integrative plasmids in these strains. While pstC, phoP and phoU mutants were successfully obtained for WCFS1 and BL23 strains, for unknown reasons and despite several attempts, only pstC disruptants of the Lpp+ strain could be achieved.
2.2. Mutation of Pst Genes Coding for ABC Phosphate Transporters Impacts As(V) Uptake and Toxicity
We performed time-course As(V) incorporation assays in the Lp. plantarum strains defective in pstC, phoP or phoU genes (Figure 1). In these experiments the absence of phosphate allowed As(V) uptake by the cells, including WCFS1 strain, which had showed low As(V) uptake when phosphate was present (Table 1). As(V) incorporation by WCFS1 strain was lower compared to Lpp+ strain. However, mutation of the PstC component of the phosphate ABC transporter in Lpp+ resulted in reduced As(V) uptake, while it was almost abolished in the WCFS1 pstC derivative (Figure 2). The capacity of the WCFS1 phoU strains to incorporate As(V) was comparable to that of the pstC strain, whereas a phoP mutant had an intermediate behavior. The capacity to incorporate As(V) by Lc. paracasei BL23 under these conditions was very low and did not show differences between the wild type or the pstC defective strain (data not shown).
We next determined the degree of toxicity of As(V) by culturing the different strains in growth media with high or low-phosphate concentration (MEI and LP-MEI medium, respectively) containing different concentrations of As(V). The toxicity and the effects of the mutations varied between both Lp. plantarum strains but, lower As(V) toxicity in high-phosphate conditions (Supplementary Figure S2) than under low-phosphate conditions (Figure 2) was observed in the WCFS1 strain. Furthermore, inactivation of either pstC or phoP enhanced As(V) resistance when growth experiments were performed under low phosphate in WCFS1. Inactivation of phoU in strain WCFS1 also led to a slight increase in resistance against As(V) (Figure 2). The effects of the mutations were much less evident when the strains were grown in MEI medium, supporting that the presence of phosphate had a protective effect on As(V) toxicity, probably through competition with a common incorporating system. Furthermore, a pstC mutation displayed no effects on As(V) toxicity in the Lp. pantarum Lpp+, irrespective of the phosphate concentration in the medium (supplementary Figure S3). Similar to WCFS1, increased resistance was found in the BL23 pstC strain, only under low phosphate conditions (supplementary Figure S3).
2.3. Mutations in the PhoPR TCS Result in Increased As(III) Resistance in Lc. paracasei
While conducting the As toxicity tests in the mutant lactobacilli strains we made an unexpected observation. Disruption of phoP in the Lc. paracasei BL23 strain resulted in increased resistance to As(III) (data not shown). The disruption of phoP in BL23 had been obtained by plasmid integration, which can result in strong polar effects on the expression of downstream phoR or pst genes. In order to avoid this, we constructed an in-frame internal deletion of phoP in BL23 which resulted in expression of a mutant PhoP protein devoid of 189 amino acids (PhoP(13-201), which included the phosphorylatable Asp-52 residue.
Figure 3 depicts the diameters of the inhibitory halos resulting from spotting filter papers impregnated with As(III) on lawns of bacteria in medium with high or low phosphate contents. While clear growth inhibition was observed for the wild type, no such inhibition haloes were evidenced for the phoP strain under low-phosphate conditions, while they were strongly reduced under high phosphate conditions. Compared to the wild type, this reduction in the diameter of haloes caused by As(III) could also be observed in the pstC and phoU strains only under low phosphate conditions. However, the effect of these mutations was weaker compared to phoP.
Growth assays confirmed the reduced sensitivity to As(III) of phoP strain (Figure 4). Furthermore, complementation of the phoP strain by expressing wild-type PhoP from a plasmid restored As(III) sensitivity. This excluded the possibility that polar effects on adjacent genes due to mutation (insertion or deletion) were responsible for the observed phenotype in BL23 phoP strains. The As(III) resistance phenotype upon phoP mutation was exclusive for Lc. paracasei, as in the equivalent Lp. plantarum mutant strains no apparent changes in As(III) sensitivity were observed (supplementary Figure S4).
Functionality of TCS usually requires the concerted action of a sensor histidine kinase and a response regulator [27]. Therefore, we next explored whether eliminating PhoR in BL23 strain resulted in a similar As(III) resistance phenotype as that detected in phoP cells. To this end, a new strain where phoR was completely eliminated by double recombination was obtained. The toxicity of As(III) was reduced compared to the wild-type strain, reaching levels similar to that of a phoP strain. Although transforming this strain with a plasmid bearing phoR did not fully complement the inactivation of phoR (Figure 4), these results suggested that resistance to As(III) was possibly linked to genes under control of PhoRP.
2.4. No Differences in As(III) Oxidation Are Observed in Lc. paracasei phoP or phoR Mutants
The fact that eliminating PhoP triggered resistance to As(III) in Lc. paracasei was puzzling, as this inorganic arsenic species does not resemble phosphate and its links with phosphate transport/metabolism are not obvious. We hypothesized that being a pleiotropic transcriptional regulator, the lack of PhoP could lead to changes in the bacterial cell surface capacity/characteristics to retain As(III), leading to decreased toxicity of the metalloid. When Lc paracasei strains were incubated with 5 mg/L As(III) at an OD595 of 10, they showed low capacity to bind As(III) and most of As(III) added to the cell suspensions was recovered by a first washing step in both the wild type and phoP strains (98.4 ± 0.5 % and 96.4 ± 3.0 %, respectively). In these experiments, As(III) retained by bacteria after two washing steps was somewhat higher for the phoP strain (56.3 ± 40.2 ng As per ml of cells at an OD595 of 10) compared to the wild-type (4.9 ± 2.3 ng As per ml of cells at an OD595 of 10). This excluded the possibility that reduced surface interaction or binding resulted in reduced toxicity in the phoP strains. A second possibility was that in phoP or phoR strains a likely oxidation of As(III) to the less toxic As(V) species was enhanced or taking place, affecting thus the toxicity of iAs. We tested this possibility by performing experiments in which As(III) was present during growth in MEI medium in the wild type, phoP and phoR strains of BL23 and determining the speciation of different iAs forms present in the cells. Oxidation of As(III) to As(V) did not take place in these assays, as the proportion of As(V) was always very low and similar for all strains (between 0.9 to 2% of total As; Figure 5). This indicated that increased As(III) resistance upon phoP or phoR deletion probably resides in mechanisms different to As(III) oxidation. In these experiments, the capacity of mutant bacteria to retain As(III) was higher compared to the wild type. However, it has to be considered that in these experiments, owing to the intrinsic As(III) toxicity, the wild type strain displayed lower growth compared to phoP and phoR strains.
3. Discussion
Strains of lactobacilli have demonstrated the ability to incorporate inorganic arsenic (iAs), predominantly in the As(V) form [15,28]. They have been proposed as potential tools for protecting against metal toxicity, and even as a means of eliminating it from water or other beverages by using these food-grade microorganisms, akin to similar applications suggested for addressing other contaminating or toxic substances like mycotoxins [15,29]. However, the presence of phosphate, which is typically widespread, may impose a limitation on the capacity for As(V) accumulation, hindering its effective utilization. This situation mirrors that observed with other toxic metals, such as mercury, which in food matrices generally forms complexes with thiolated compounds [11]. These compounds alter the affinity of probiotic strains for mercury, thereby interfering with the chelation process.
We have demonstrated that the As(V) uptake capability of Lp. plantarum WCFS1 may be associated with the Pst ABC-type phosphate transporter. The presence of pstSCAB clusters in lactobacilli (see supplementary Figure S1) suggests that this transporter could be a major phosphate uptake system in this bacterial group, although no prior studies on it have been reported for these microorganisms. While Pst transporters have been extensively characterized in bacteria and thoroughly studied in model organisms such as E. coli, other phosphate transporters like the Pit permeases have also been identified for phosphate uptake [17,30]. The existence of alternative and additional phosphate transporter(s) might explain the varied effects of pstC inactivation on As(V) incorporation in the two employed strains of Lp. plantarum . In Lc. paracasei BL23, a strain which does not display any remarkable As(V) incorporation capacity, mutating pstC also resulted in increased As(V) resistance when cells were cultured in a medium with low phosphate. The effects of phoP and phoU mutations in WCFS1 and BL23 strains have, however, no straightforward interpretation and may indicate that in the absence of these putative regulators, possible changes in PstSCAB expression may impact As(V) incorporation.
In E. coli, strains harboring a phoU mutation exhibit deregulation of phosphate transport, enabling the incorporation of higher amounts [31]. In this bacterium, PhoU serves a regulatory function, linking the transport activity through Pst components to the auto-phosphorylation of the sensor kinase PhoR. This mechanism provides a means of sensing external phosphate concentrations and subsequently regulating PhoP activity [17,18]. Presently, the exact role of PhoU in lactobacilli remains unknown. While PhoU is present in some bacteria possessing PstSCAB transporters and PhoPR orthologues, it is absent from others, including some lactobacilli. It is also remarkable that in some members of the Lactobacillaceae (e.g.: species of Lacticaseibacillus, Lactiplantibacillus, Latilactobacillus, Loigolactobacillus, Levilactobacillus, Lentilactobacillus, Liquorilactobacillus, Agrilactobacillus or Secundilactobacillus), a gene encoding a putative protein with eight transmembrane segments and a C-terminal cytoplasmic PDZ-domain (LCABL_10470 in Lc. paracasei BL23) is always located upstream of phoP, forming a likely operon structure with phoPR (supplementary Figure S1). Therefore, the possible function of this gene in the Pho-Pst regulatory network deserves further investigation.
In addition to the observed acid sensitivity and reduced growth rate in the Lc. paracasei BL23 phoP strain in MRS medium [20], we have uncovered an unexpected As(III) resistance phenotype. Our investigation ruled out the possibility that the BL23 phoP strain was incorporating less As(III) or promoting its oxidation to As(V). In fact, Lc. paracasei phoP and phoR mutants accumulated more As(III) compared to the wild-type strain, although As(III) uptake in Lc. paracasei occurs at very low levels. The reasons for this effect are unknown. In E. coli, the pho regulon is well characterized. This regulon includes genes dedicated to phosphate uptake and phosphate scavenging under conditions of limited phosphate supply, such as genes encoding phosphatases and the pst genes [18]. However, owing to the metabolic relevance of phosphate, a defect in PhoP, and hence in cell phosphate supply may have important pleiotropic effects. How a defect in phoP in the BL23 strain increases As(III) resistance is still not known. A defect in the sensor kinase encoding gene phoR in Lc. casei BL23 also resulted in a resistance phenotype compared to phoP. This suggests that the resistance in a phoR mutant may be attributed to the absence of PhoP activation through phosphorylation due to the lack of its cognate kinase. However, the possibility that deletion of phoR resulted in altered expression of phoP cannot be excluded, as we could not confirm complementation when we transformed the phoR strain with a plasmid bearing phoR.
Mechanisms to counteract the intrinsic toxicity of iAs have been described in a number of bacteria. Arsenic resistance generally relies in a mechanism by which As(V) molecules that enter the cell (probably by mimicking phosphate) are transformed to As(III) by arsenate reductase enzymes (Ars). Subsequently, this As species, which is even more toxic than As(V), can be expelled from the cells by specific As(III) detoxifying pumps [32]. Although these mechanisms are not common or widespread in lactobacilli, they have been genetically identified in some strains [33,34]. However, no specific As(III) pump, nor arsenate reductase encoding genes, can be identified in the Lc. paracasei BL23 genome that could be eventually overexpressed in the phoP mutant. Links between As(III) and PhoPR have been described in other bacteria, revealing other metabolic connections between As and phosphate metabolism. In Halomonas sp., the TCS PhoBR (equivalent to PhoPR) regulates the expression of the aioBA genes coding for As(III) oxidases which promote As(III) to As(V) conversion depending on phosphate availability [35]. In Agrobacterium tumefaciens an antimonite [Sb(III)] detoxifying mechanism that promotes Sb(III) oxidation to antimonate [Sb(V)] has also been described, mediated by the Sb(III) oxidase AnoA, which shows cross reactivity with As(III), and whose expression is also controlled by phosphate through PhoB (PhoP) [36]. However, mutations in phoB in these bacteria lead to reduced expression of these oxidases, which decreases As(III) conversion and Sb(III) detoxification, and aioBA, or anoA homologous genes are not present in the BL23 genome.
In this work we determined the relationship between As(V) and phosphate transport in Lp. plantarum. Our results highlight the importance of the PstSCAB system in phosphate metabolism in lactobacilli and point to the existence of alternative phosphate transporters. At the same time, they show that specific uptake, and not unspecific cell retention plays a role in As(V) bioaccumulation by lactobacilli. Notwithstanding, the potential beneficial effects of lactobacilli on iAs toxicity may depend on both bioaccumulation of iAs, leading to reduced bioaccessibility at the gastrointestinal tract, but they can extend beyond mere iAs capture and may involve additional mechanisms, such as anti-inflammatory and antioxidant properties unique to certain strains. As an example, recent experiments in animal models showed that strains like Lc. paracasei BL23, which does not possess As(III) [nor As(V)] binding abilities reduce most of the intestinal toxic effects of As(III) exposure through drinking water in a murine model [12].
We have also demonstrated that Lc. paracasei lacking phoP or phoR does not exhibit apparent As(III) detoxification (oxidation) but displays enhanced resistance to it. Characterizing the pho regulon in Lb. paracasei and investigating transcriptomic/proteomic changes resulting from phoP elimination will contribute to understand the alterations caused by the absence of this regulator, which triggers As(III) resistance. In particular, elucidating the cross-talk mechanisms associated with phosphate sensing via PhoPR, which in other microorganisms involve carbon, nitrogen, iron, potassium, sodium, and sulfur metabolism, as well as resistance to general stresses [37], will enhance our comprehension of this regulatory network.
4. Materials and Methods
4.1. Bacterial Culture Conditions
The lactobacilli strains used in this study (Table 1 and Table 2) were routinely grown in de Man, Rogosa and Sharpe (MRS) medium (BD Difco) at 30 or 37 °C, under static conditions. For iAs toxicity assays, MEI medium was employed [38] containing (w/v): 0.5% yeast extract, 0.5% tryptone, 0.4% K2HPO4, 0.5% KH2PO4, 0.02%, MgSO4·7H2O, 0.005% MnSO4, 0.05% cysteine, 0.5% glucose and 1 ml of Tween 80 per liter. When low phosphate conditions were employed, no K2HPO4 and KH2PO4 were added to the MEI medium (LP-MEI medium). E. coli DH10B [F− endA1 recA1 galE15 galK16 nupG rpsL ΔlacX74 Φ80lacZΔM15 araD139 Δ(ara,leu)7697 mcrA Δ(mrr-hsdRMS-mcrBC) λ−] was employed for cloning purposes and it was grown in LB medium at 37 °C under strong agitation (200 rpm). For E. coli clone selection, ampicillin was used at 100 μg/mL. Erythromycin at 5 g/mL was used for selection of recombinant clones in lactobacilli. Solid media were made by adding 1.8% (w/v) agar.
4.2. Construction of Strains Mutated in Pst and Pho Genes
The oligonucleotides used in this work are listed in the supplementary Table S1. Chromosomal DNA from Lp. plantarum and Lc. paracasei strains was isolated with the DNA Isolation Kit for Cells and Tissues (Roche Applied Science). Internal fragments ranging from 300 to 500 pb from pstC, phoP and phoU were amplified by PCR from chromosomal DNA with NZYTaq II DNA polymerase (NZYtech). The obtained fragments were purified with the GFX PCR DNA and Gel Band Purification Kit (Cytiva), digested with appropriated restriction enzymes and cloned into the integrative plasmid pRV300 [39] digested with the same enzymes. The ligation mixtures were transformed into E. coli DH10B and recombinant clones were selected in LB agar plates with ampicillin, 40 g/mL X-gal and 0.1 mM IPTG. The obtained plasmids were purified from E. coli with the NucleoSpin Plasmid Kit (Machery-Nagel) and used to transform Lp. plantarum WCFS1, Lp. plantarum Lpp+ and Lc. paracasei BL23 to obtain disruption mutants by single cross-over integration. Plasmid integration at the correct locus was checked by PCR with one oligonucleotide which hybridized in the targeted gene outside the cloned fragment and an oligonucleotide hybridizing in the pRV300 plasmid.
To obtain phoP and phoR derivatives from BL23 strain, fragments of 1 kb upstream and downstream of the desired deletion were synthesized by PCR with Phusion High-Fidelity DNA Polymerase (Thermo Scientific). These fragments overlapped by 20 bp and they were fused by a second PCR reaction using them as template. The obtained 2 kb fragments were digested with appropriated restriction enzymes and cloned into pRV300. The integrative plasmids thus obtained were used to transform Lc. paracasei BL23 and strains with a first integration of the plasmid in the chromosome were isolated on MRS plates containing erythromycin. One transformant was selected from each construction and they were grown for approximately 200 generations in the absence of antibiotic. Strains in which a second recombination event took place, leading to plasmid excision, were selected by erythromycin sensibility by replica-plating. Among the erythromycin-sensible clones obtained from each construction, the presence of the desired deletion was checked by PCR and confirmed by sequencing.
Lp. plantarum and Lc. paracasei were transformed by electroporation with a Gene Pulser apparatus (Bio-Rad). Lp. plantarum electrocompetent bacteria were prepared in polyethylene glycol (PEG) 1500 as described [40] with some modifications. The bacteria were cultured in 50 mL of MRS supplemented with 1% (w/v) glycine to an OD595 of 0.4-0.6. After washing with 1 vol of cold 1 mM MgCl2, the cells were washed with half vol of cold 30% PEG 1500 and resuspended in 500 μL of 30% PEG 1500. Cells were electroporated in 0.2 cm cuvettes at 1.5 kV, 25 μF and 400 Ω, with 0.5-2 g of purified plasmids and resuspended in 1 mL of MRS. After incubation at 30 °C for 2 h the transformed bacteria were plated on MRS plates containing 5 g/mL of erythromycin and incubated at 30 °C for 48 h. Lc. paracasei BL23 was transformed by electroporation as previously described [41] and the transformants were isolated on MRS plates with 5 g/L erythromycin at 37 °C
4.3. Strains Complementation
The phoP and phoR genes from Lc. paracasei BL23 were amplified by PCR with Phusion High-Fidelity DNA Polymerase (Thermo Scientific) and appropriated oligonucleotides (supplementary Table S1). The purified PCR fragments were ligated to BglII/SpeI-digested pT1NX [42] plasmid with the GeneArt™ Gibson Assembly EX kit (Invitrogen), leading to phoP and phoR genes in which their expression was under the control of the lactococcal P1 constitutive promoter, respectively. The products of the Gibson reaction were used to transform Lactococcus lactis MG1363 electrocompetent cells [43] and transformants were selected on M17 (Oxoid) agar plates containing 0.5 % (w/v) glucose (GM17) plus 5 g/mL erythromycin after incubation at 30 °C. Colonies were checked by PCR and positive clones bearing inserts were grown in 5 mL of GM17 for plasmid isolation with the NucleoSpin Plasmid Kit (Macherey-Nagel) with modifications. Cells were incubated in STE buffer (20% sucrose, 10 mM Tris-HCl [pH 8.0], 10 mM EDTA, 50 mM NaCl) supplemented with lysozyme (1 mg/mL) for 30 min at 37 °C prior cell lysis. Subsequent steps were carried out as indicated by the manufacturer. pT1NX derivatives carrying phoP and phoR were sequenced and these expression plasmids were used to transform Lc. paracasei phoP and phoR mutant strains, respectively, by electroporation.
4.4. As Toxicity, Incorporation and Speciation Assays
As(V) (stock solution of 1000 mg/L, As2O5) was purchased from Merck. The As(III) solution (1000 mg/L) was prepared by dissolving 1.320 g of As2O3 (Riedel-de Haën) in 25 mL of KOH 20% w/v. After neutralization with 20% H2SO4 v/v, this solution was made up to a final volume of 1 L with H2SO4 1% v/v. DMA(V) [dimethylarsinic acid; (CH3)2AsNaO2·3H2O (Fluka)] stock solution was prepared in water.
The lactobacilli strains were cultured in MEI or LP-MEI media with different As(III) or As(V) concentrations in 96-well plates (200 L per well) at 30 °C (Lp. pantarum) or 37 °C (Lc. paracasei) in a Spectrostar Nano plate reader (BMG-Labtech). OD readings at 595nm were recorded every 30 min. As(III) inhibition assays on plates were carried out by placing Whatman 3MM filter paper discs (5 mm diameter) impregnated with 3 L of As(III) 1000 mg/L onto 90 mm MRS agar plates with an overlay of 5 ml of MRS with 0.8 % agar containing 106 CFU of different Lc. paracasei strains. After incubation for 24 h, the diameter of inhibition haloes was measured.
As(V), As(III) and DMA retention capacity in different lactobacilli was tested in cells from overnight cultures (5 mL) resuspended in phosphate-buffered saline (PBS) to an OD at 595 nm of 10. Cell suspensions were supplemented with 1 mg/L of As(V), As(III) or DMA, respectively, and incubated for 1h at 37 °C. Cells were centrifuged at 5000 x g for 10 min and washed with 5 ml of PBS. The As retained in the bacterial pellets was determined as indicated below.
For time-course As(V) incorporation assays, strains were cultured overnight in 50 mL of MRS, pelleted by centrifugation (5000 x g, 10 min) and washed with 1 vol of 0.9% NaCl. Washed bacteria were resuspended in 0.9% NaCl and OD at 595nm was adjusted to 10. Aliquots of the cell suspensions (1 mL) were incubated at 30 °C for 5 min in a water bath before As(V) was added to a final concentration of 5 mg/L. Samples of 300 L were withdrawn at different time intervals and quickly filtered under vacuum using 0.45 m nitrocellulose filters (Millipore). The filters were washed twice with 5 ml of 0.9% NaCl and air dried before As quantification.
The As(III) retention capacity of Lc. paracasei strains was estimated by measuring the amount of As(III) retained by 1 mL of cells at an OD at 595nm of 10. Cell suspensions were incubated with 5 mg/L As(III) for 1 h at 37 °C in 0.9 % NaCl. After centrifugation at 10000 x g 5 min, bacterial pellets were washed twice with 1 mL of 0.9 % NaCl and As(III) in washing supernatants and the bacterial pellet were determined.
For total As determination in samples (bacterial washing supernatants, cell pellets and filters), after a dry ashing step, As quantification was carried out by flow injection-hydride generation-atomic absorption spectrometry (FI-HG-AAS), following the procedure described by Clemente et al. [44].
To determine As(III) oxidation to As(V) by Lc. paracasei cells. 50 ml of MEI medium containing 5 mg/L As(III) were inoculated with Lc. paracasei BL23 and their derived phoP and phoR mutants at an initial OD at 595nm of 0.01. After 16h of incubation at 37 °C, cells were washed at 4 °C with cold 0.9% NaCl (4000 x g 10 min) and the bacterial pellets were kept at -20 °C until analysis. iAs was extracted from bacterial pellets with 5 mL of 0.28 M HNO3 at 95 °C for 1.5 h [45] and the As(III) and As(V) contents were determined by HPLC with anion exchange column (Hamilton PRP X100, 150 × 4.1 mm, particle size 5 μm) coupled to a PerkinElmer Nex-Ion™300X ICP-MS as described [46].
4.5. Statistical Analysis
One-way ANOVA with Tukey's multiple comparison test and Student's t test were carried out with GraphPad Prism 5. Differences were considered statistically significant at p < 0.05.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: Genomic context of pstSCAB genes in members of the Lactobacillaceae; Figure S2: Growth of lactobacilli strains in MEI medium (high phosphate) with different concentrations of As(V); Figure S3: Growth of Lp. plantarum Lpp+ and Lc. paracasei BL23 strains and their derived pstC mutants in MEI and LP-MEI media with different concentrations of As(V); Figure S4: Grow curves of Lp. plantarum WCFS1 and its derived phoP mutant in MEI medium with different amounts of As(III); Table S1: Oligonucleotides used in this study;.
Author Contributions
Conceptualization, Dinoraz Vélez, Vicenta Devesa, Vicente Monedero and Manuel Zúñiga; Funding acquisition, Dinoraz Vélez, Vicenta Devesa, Vicente Monedero and Manuel Zúñiga; Investigation, Daniela Corrales, Cristina Alcántara, María Jesús Clemente, Dinoraz Vélez, Vicenta Devesa, Vicente Monedero and Manuel Zúñiga; Methodology, Dinoraz Vélez, Vicenta Devesa, Vicente Monedero and Manuel Zúñiga; Project administration, Vicenta Devesa and Manuel Zúñiga; Supervision, Vicente Monedero and Manuel Zúñiga; Writing – original draft, Vicente Monedero and Manuel Zúñiga; Writing – review & editing, Daniela Corrales, Cristina Alcántara, Dinoraz Vélez, Vicenta Devesa, Vicente Monedero and Manuel Zúñiga. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Ministerio de Ciencia, Innovación y Universidades/Agencia Estatal de Investigación (MCIU/AEI/10.13039/501100011033) from Spain, grant number RTI2018-098071-B-I00 and it was co-funded by the European Commission through the European Regional Development Fund (Multiregional operative program for Spain 2014–2020) and by Generalitat Valenciana, grant number AICO/2019/008. The Accreditation as Center of Excellence Severo Ochoa CEX2021-001189-S funded by MCIU/AEI is also fully acknowledged.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors on request.
Acknowledgments
D. Corrales is grateful for her doctoral fellowship from the Ministerio de Ciencia, Tecnología e Innovación of Colombia (Convocatoria 906 de 2021, Doctorados en el Exterior). R. Domínguez (Grupo de Elementos Traza, Espectroscopía y Especiación, Universidad de Santiago de Compostela, Spain) is acknowledged for the As speciation analysis. We thank M. Bovea and E. Cebriá for their help during strain construction.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- WHO, Arsenic fact sheet. 2012, No. 372, Geneva.
- Mayne, S. T. , The FDA's action plan to reduce dietary exposure to arsenic, lead, cadmium, and mercury for infants and young children. Am J Clin Nutr 2023, 117, 647–648. [Google Scholar] [CrossRef] [PubMed]
- Monachese, M.; Burton, J. P.; Reid, G. , Bioremediation and tolerance of humans to heavy metals through microbial processes: a potential role for probiotics? Appl Environ Microbiol 2012, 78, 6397–404. [Google Scholar] [CrossRef] [PubMed]
- Chiocchetti, G.M.; Jadán-Piedra, C.; Monedero, V.; Zúñiga, M.; Vélez, D.; Devesa, V. Use of lactic acid bacteria and yeasts to reduce exposure to chemical food contaminants and toxicity. Crit. Rev. Food Sci. Nutr. 2019, 59, 1534–1545. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Han, J.; Ren, Q.; Liu, Z.; Zhang, X.; Wu, Z. The Involvement of Lactic Acid Bacteria and Their Exopolysaccharides in the Biosorption and Detoxication of Heavy Metals in the Gut. Biol. Trace Element Res. 2024, 202, 671–684. [Google Scholar] [CrossRef]
- Rodríguez-Viso, P.; Domene, A.; Vélez, D.; Devesa, V.; Zúñiga, M.; Monedero, V. Protective effects of oral administration of lactic acid bacteria strains against methylmercury-induced intestinal toxicity in a murine model. Food Chem. Toxicol. 2024, 114461. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Gu, S.; Liu, D.; Zhao, L.; Xia, S.; He, X.; Chen, H.; Ge, J. , Lactobacillus brevis 23017 relieves mercury toxicity in the colon by modulation of oxidative stress and inflammation through the interplay of MAPK and NF-kappaB signaling cascades. Front Microbiol 2018, 9, 2425. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Yu, L.; Shen, X.; Tian, F.; Zhao, J.; Zhang, H.; Chen, W.; Zhai, Q. Protective Effects of Lactobacillus plantarum CCFM8610 against Acute Toxicity Caused by Different Food-Derived Forms of Cadmium in Mice. Int. J. Mol. Sci. 2021, 22, 11045. [Google Scholar] [CrossRef]
- Bisanz, J.E.; Enos, M.K.; Mwanga, J.R.; Changalucha, J.P.; Burton, J.; Gloor, G.B.; Reid, G. Randomized Open-Label Pilot Study of the Influence of Probiotics and the Gut Microbiome on Toxic Metal Levels in Tanzanian Pregnant Women and School Children. mBio 2014, 5, e01580–14. [Google Scholar] [CrossRef]
- Astolfi, M.L.; Protano, C.; Schiavi, E.; Marconi, E.; Capobianco, D.; Massimi, L.; Ristorini, M.; Baldassarre, M.E.; Laforgia, N.; Vitali, M.; et al. A prophylactic multi-strain probiotic treatment to reduce the absorption of toxic elements: In-vitro study and biomonitoring of breast milk and infant stools. Environ. Int. 2019, 130, 104818. [Google Scholar] [CrossRef]
- Jadán-Piedra, C.; Alcántara, C.; Monedero, V.; Zúñiga, M.; Vélez, D.; Devesa, V. The use of lactic acid bacteria to reduce mercury bioaccessibility. Food Chem. 2017, 228, 158–166. [Google Scholar] [CrossRef]
- Domene, A.; Orozco, H.; Rodríguez-Viso, P.; Monedero, V.; Zúñiga, M.; Vélez, D.; Devesa, V. Lactobacillus strains reduce the toxic effects of a subchronic exposure to arsenite through drinking water. Environ. Res. 2024, 245, 117989. [Google Scholar] [CrossRef]
- Jain, A.; Jain, R.; Jain, S. K. , Assessment of Lactobacillus rhamnosus mediated protection against arsenic-induced toxicity in zebrafish: a qPCR-based analysis of Firmicutes and Bacteroidetes groups and embryonic development. Arch Microbiol 2023, 205, 316. [Google Scholar] [CrossRef]
- Bora, S.; Lakshman, M.; Madhuri, D.; Kalakumar, B.; Udayakumar, M. Protective Effect of Lactobacillus sporogenes against Arsenic-Induced Hematological Alterations in Male Albino Wistar Rats. Biol. Trace Element Res. 2022, 200, 4744–4749. [Google Scholar] [CrossRef]
- Elsanhoty, R.M.; Al-Turki, I.A.; Ramadan, M.F. Application of lactic acid bacteria in removing heavy metals and aflatoxin B1 from contaminated water. Water Sci. Technol. 2016, 74, 625–638. [Google Scholar] [CrossRef]
- Villa-Bellosta, R.; Sorribas, V. Role of rat sodium/phosphate cotransporters in the cell membrane transport of arsenate. Toxicol. Appl. Pharmacol. 2008, 232, 125–134. [Google Scholar] [CrossRef]
- Hsieh, Y.-J.; Wanner, B.L. Global regulation by the seven-component Pi signaling system. Curr. Opin. Microbiol. 2010, 13, 198–203. [Google Scholar] [CrossRef]
- Gardner, S. G.; McCleary, W. R. , Control of the phoBR regulon in Escherichia coli. EcoSal Plus 2019, 8. [Google Scholar] [CrossRef] [PubMed]
- Gardner, S.G.; Johns, K.D.; Tanner, R.; McCleary, W.R. The PhoU Protein from Escherichia coli Interacts with PhoR, PstB, and Metals To Form a Phosphate-Signaling Complex at the Membrane. J. Bacteriol. 2014, 196, 1741–1752. [Google Scholar] [CrossRef] [PubMed]
- Alcántara, C.; Revilla-Guarinos, A.; Zúñiga, M. Influence of Two-Component Signal Transduction Systems of Lactobacillus casei BL23 on Tolerance to Stress Conditions. Appl. Environ. Microbiol. 2011, 77, 1516–1519. [Google Scholar] [CrossRef] [PubMed]
- Clemente, M.J.; Vivó, M.d.L. .; Puig, S.; Zúñiga, M.; Monedero, V.; Devesa, V.; Vélez, D. In vitro evaluation of the efficacy of lactobacilli and yeasts in reducing bioavailability of inorganic arsenic. LWT 2020, 126, 109272. [Google Scholar] [CrossRef]
- Alcántara, C.; Jadán-Piedra, C.; Vélez, D.; Devesa, V.; Zúñiga, M.; Monedero, V. Characterization of the binding capacity of mercurial species in Lactobacillus strains. J. Sci. Food Agric. 2017, 97, 5107–5113. [Google Scholar] [CrossRef] [PubMed]
- Kleerebezem, M.; Boekhorst, J.; Van Kranenburg, R.; Molenaar, D.; Kuipers, O.P.; Leer, R.; Tarchini, R.; Peters, S.A.; Sandbrink, H.M.; Fiers, M.W.E.J.; et al. Complete genome sequence of Lactobacillus plantarum WCFS1. Proc. Natl. Acad. Sci. USA 2003, 100, 1990–1995. [Google Scholar] [CrossRef] [PubMed]
- Cesena, C.; Morelli, L.; Alander, M.; Siljander, T.; Tuomola, E.; Salminen, S.; Mattila-Sandholm, T.; Vilpponen-Salmela, T.; von Wright, A. Lactobacillus crispatus and its Nonaggregating Mutant in Human Colonization Trials. J. Dairy Sci. 2001, 84, 1001–1010. [Google Scholar] [CrossRef] [PubMed]
- Botas, J.; del Río. R.; Giner-Lamia, J.; Huerta-Cepas, J. GeCoViz: genomic context visualisation of prokaryotic genes from a functional and evolutionary perspective. Nucleic Acids Res. 2022, 50, W352–W357. [Google Scholar] [CrossRef] [PubMed]
- Mazé, A.; Boël, G.; Zúñiga, M.; Bourand, A.; Loux, V.; Yebra, M.J.; Monedero, V.; Correia, K.; Jacques, N.; Beaufils, S.; et al. Complete Genome Sequence of the Probiotic Lactobacillus casei Strain BL23. J. Bacteriol. 2010, 192, 2647–2648. [Google Scholar] [CrossRef]
- Buschiazzo, A.; Trajtenberg, F. , Two-component sensing and regulation: how do histidine kinases talk with response regulators at the molecular level? Annual Review of Microbiology 2019, 73, 507–528. [Google Scholar] [CrossRef] [PubMed]
- Halttunen, T.; Finell, M.; Salminen, S. Arsenic removal by native and chemically modified lactic acid bacteria. Int. J. Food Microbiol. 2007, 120, 173–178. [Google Scholar] [CrossRef]
- Zoghi, A.; Khosravi-Darani, K.; Sohrabvandi, S.; Attar, H. Patulin removal from synbiotic apple juice using Lactobacillus plantarum ATCC 8014. J. Appl. Microbiol. 2018, 126, 1149–1160. [Google Scholar] [CrossRef]
- Atalla, A.; Schumann, W. , The pst operon of Bacillus subtilis is specifically induced by alkali stress. Journal of Bacteriology 2003, 185, 5019–5022. [Google Scholar] [CrossRef]
- Morohoshi, T.; Maruo, T.; Shirai, Y.; Kato, J.; Ikeda, T.; Takiguchi, N.; Ohtake, H.; Kuroda, A. , Accumulation of inorganic polyphosphate in phoU mutants of Escherichia coli and Synechocystis sp. strain PCC6803. Appl Environ Microbiol 2002, 68, 4107–10. [Google Scholar] [CrossRef]
- Rosen, B.P. Biochemistry of arsenic detoxification. FEBS Lett. 2002, 529, 86–92. [Google Scholar] [CrossRef]
- Isokpehi, R.D.; Udensi, U.K.; Simmons, S.S.; Hollman, A.L.; Cain, A.E.; Olofinsae, S.A.; Hassan, O.A.; Kashim, Z.A.; Enejoh, O.A.; Fasesan, D.E.; et al. Evaluative Profiling of Arsenic Sensing and Regulatory Systems in the Human Microbiome Project Genomes. Microbiol. Insights 2014, 7, MBI–S18076. [Google Scholar] [CrossRef] [PubMed]
- Gustaw, K.; Koper, P.; Polak-Berecka, M.; Rachwał, K.; Skrzypczak, K.; Waśko, A. Genome and Pangenome Analysis of Lactobacillus hilgardii FLUB—A New Strain Isolated from Mead. Int. J. Mol. Sci. 2021, 22, 3780. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Cao, Y.; Wei, S.; Li, Y.; Li, X.; Wang, Q.; Wang, G. Regulation of arsenite oxidation by the phosphate two-component system PhoBR in Halomonas sp. HAL1. Front. Microbiol. 2015, 6, 923. [Google Scholar] [CrossRef]
- Li, J.; Qiao, Z.; Shi, M.; Zhang, Y.; Wang, G. Regulation of antimonite oxidation and resistance by the phosphate regulator PhoB in Agrobacterium tumefaciens GW4. Microbiol. Res. 2019, 226, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Santos-Beneit, F. The Pho regulon: a huge regulatory network in bacteria. Front. Microbiol. 2015, 6, 402. [Google Scholar] [CrossRef]
- Alcántara, C.; Coll-Marqués, J.M.; Jadán-Piedra, C.; Vélez, D.; Devesa, V.; Zúñiga, M.; Monedero, V. Polyphosphate in Lactobacillus and Its Link to Stress Tolerance and Probiotic Properties. Front. Microbiol. 2018, 9, 1944. [Google Scholar] [CrossRef]
- Leloup, L.; Ehrlich, S.D.; Zagorec, M.; Morel-Deville, F. Single-crossover integration in the Lactobacillus sake chromosome and insertional inactivation of the ptsI and lacL genes. Appl. Environ. Microbiol. 1997, 63, 2117–2123. [Google Scholar] [CrossRef]
- Aukrust, T.; Blom, H. Transformation of Lactobacillus strains used in meat and vegetable fermentations. Food Res. Int. 1992, 25, 253–261. [Google Scholar] [CrossRef]
- Posno, M.; Leer, R.J.; van Luijk, N.; van Giezen, M.J.F.; Heuvelmans, P.T.H.M.; Lokman, B.C.; Pouwels, P.H. Incompatibility of Lactobacillus Vectors with Replicons Derived from Small Cryptic Lactobacillus Plasmids and Segregational Instability of the Introduced Vectors. Appl. Environ. Microbiol. 1991, 57, 1822–1828. [Google Scholar] [CrossRef]
- Schotte, L.; Steidler, L.; Vandekerckhove, J.; Remaut, E. Secretion of biologically active murine interleukin-10 by Lactococcus lactis. Enzym. Microb. Technol. 2000, 27, 761–765. [Google Scholar] [CrossRef] [PubMed]
- Holo, H.; Nes, I. F. , High-frequency transformation, by electroporation, of Lactococcus lactis subsp. cremoris grown with glycine in osmotically stabilized media. Appl Environ Microbiol 1989, 55, 3119–23. [Google Scholar] [PubMed]
- Clemente, M.J.; Devesa, V.; Vélez, D. In Vitro Reduction of Arsenic Bioavailability Using Dietary Strategies. J. Agric. Food Chem. 2017, 65, 3956–3964. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.-H.; Ilgen, G.; Fecher, P. Quantitative chemical extraction for arsenic speciation in rice grains. J. Anal. At. Spectrom. 2010, 25, 800–802. [Google Scholar] [CrossRef]
- Domínguez-González, M.R.; Barciela-Alonso, M.C.; Calvo-Millán, V.G.; Herbello-Hermelo, P.; Bermejo-Barrera, P. The bioavailability of arsenic species in rice. Anal. Bioanal. Chem. 2020, 412, 3253–3259. [Google Scholar] [CrossRef]
Figure 1.
Incorporation of As(V) in Lp. plantarum strains and their derivative mutants affected in pst and pho genes (a) L. plantarum Lpp+; (b) Lp. plantarum WCFS1. Columns represent average amounts of As retained by 300 L of bacterial cells at an OD595 of 10 when exposed to 5 mg/L As(V) (n=3). Error bars represent standard deviations. * (p<0.05); ** (p<0.01), statistical differences with respect to wild-type strain for each time point.
Figure 1.
Incorporation of As(V) in Lp. plantarum strains and their derivative mutants affected in pst and pho genes (a) L. plantarum Lpp+; (b) Lp. plantarum WCFS1. Columns represent average amounts of As retained by 300 L of bacterial cells at an OD595 of 10 when exposed to 5 mg/L As(V) (n=3). Error bars represent standard deviations. * (p<0.05); ** (p<0.01), statistical differences with respect to wild-type strain for each time point.
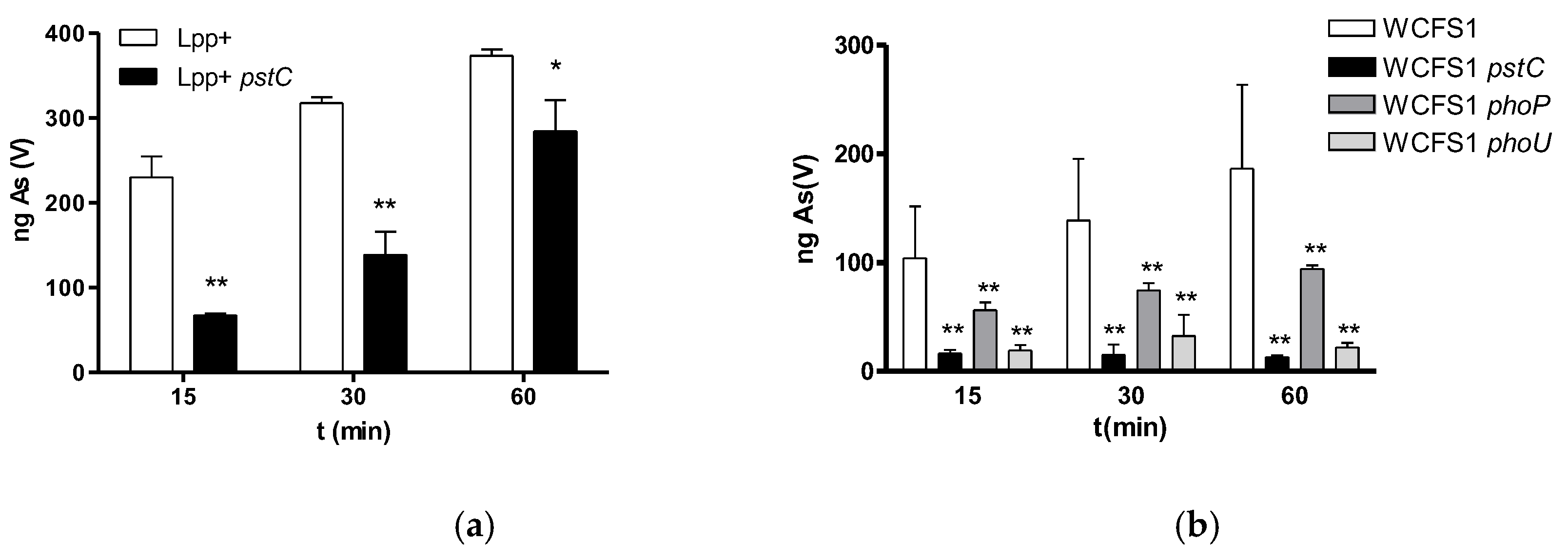
Figure 2.
Growth curves of Lp. plantarum WCFS1 and different pst and pho derived mutants in LP-MEI medium with different amounts of As(V) indicated in mg/L. (a) wild type; (b) pstC mutant; (c) phoP mutant; (d) phoU mutant.
Figure 2.
Growth curves of Lp. plantarum WCFS1 and different pst and pho derived mutants in LP-MEI medium with different amounts of As(V) indicated in mg/L. (a) wild type; (b) pstC mutant; (c) phoP mutant; (d) phoU mutant.
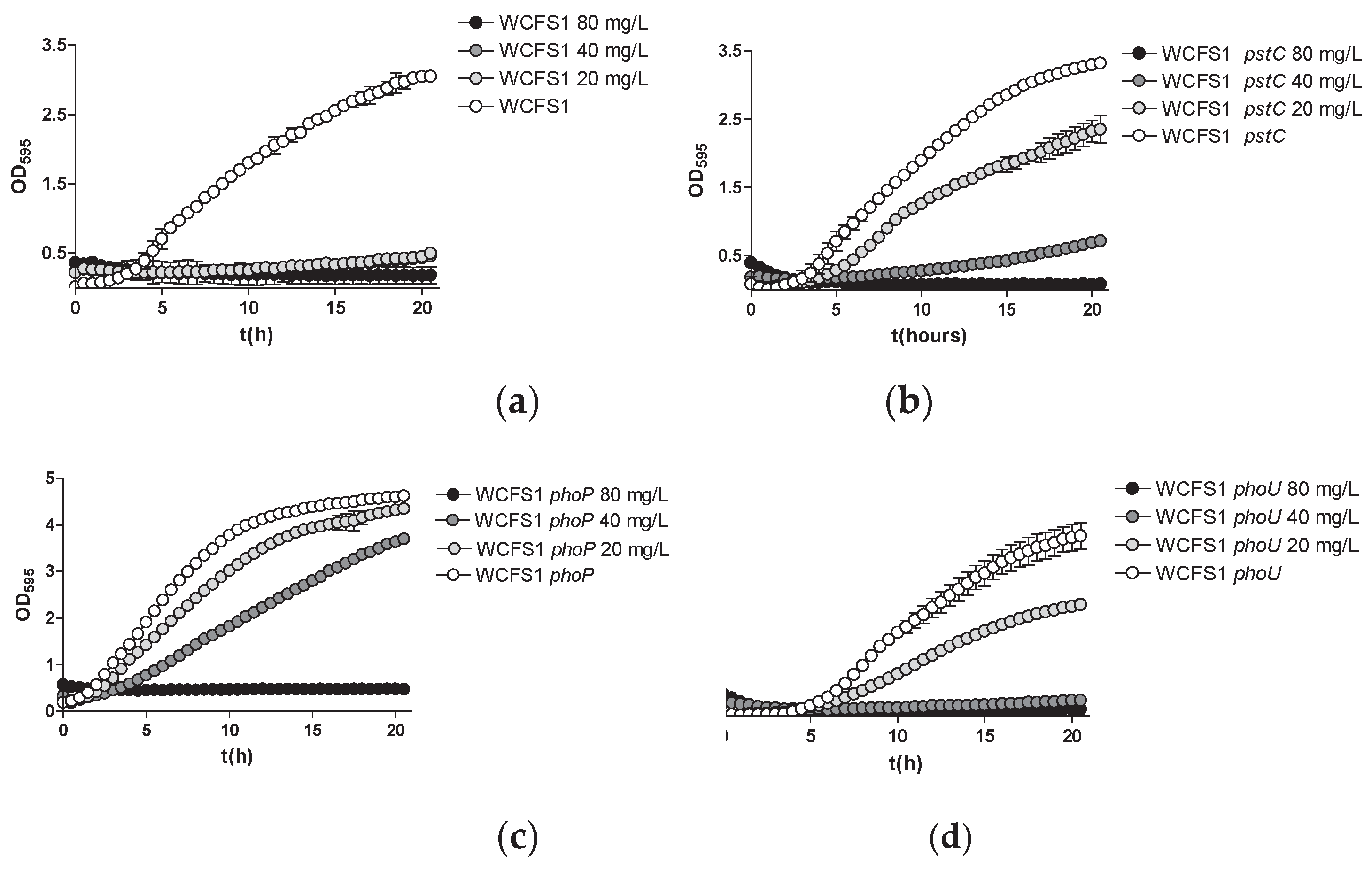
Figure 3.
Inhibition haloes of As(III) on MEI (high phosphate) and LP-MEI (low phosphate) plates seeded with different strains of Lc. paracasei BL23 wild type and mutants in pst and pho genes. Columns represent average diameters (n=3). Error bars represent standard deviations. Asterisks indicate statistical differences compared to wild-type for each growth condition (p<0.01).
Figure 3.
Inhibition haloes of As(III) on MEI (high phosphate) and LP-MEI (low phosphate) plates seeded with different strains of Lc. paracasei BL23 wild type and mutants in pst and pho genes. Columns represent average diameters (n=3). Error bars represent standard deviations. Asterisks indicate statistical differences compared to wild-type for each growth condition (p<0.01).
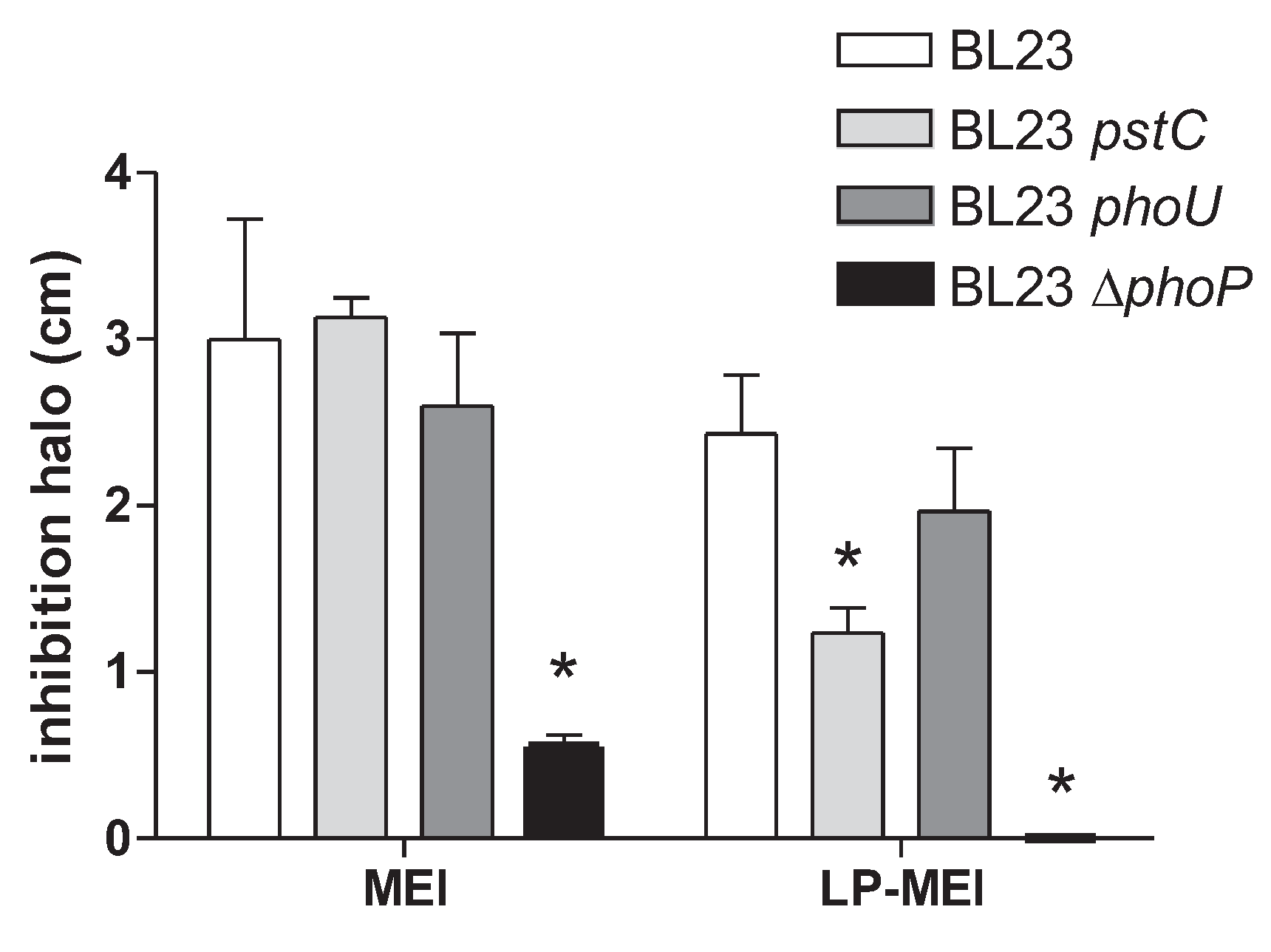
Figure 4.
Growth curves of Lc. paracasei BL23 and phoP or phoR mutants and their complemented strains in MEI medium with different amounts of As(III). (a) BL23 (wild type); (b) phoP; (c) phoP [pT1phoP], complemented strain; (d) phoR; (e) phoR[pT1phoR], complemented strain.
Figure 4.
Growth curves of Lc. paracasei BL23 and phoP or phoR mutants and their complemented strains in MEI medium with different amounts of As(III). (a) BL23 (wild type); (b) phoP; (c) phoP [pT1phoP], complemented strain; (d) phoR; (e) phoR[pT1phoR], complemented strain.
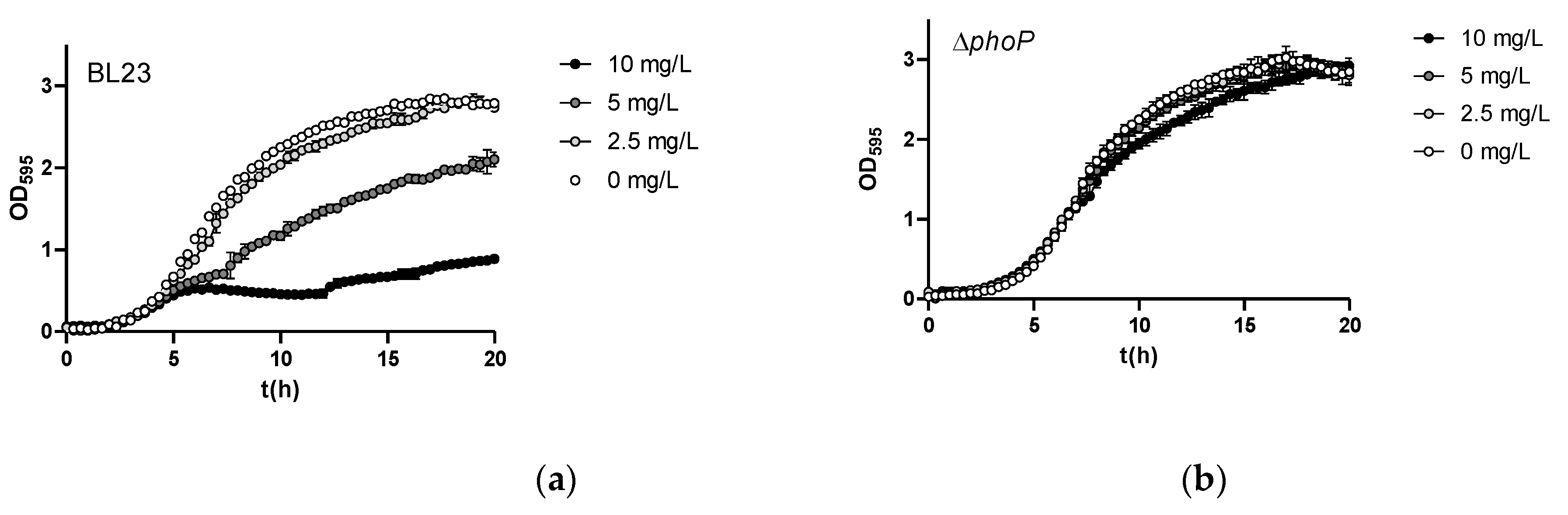
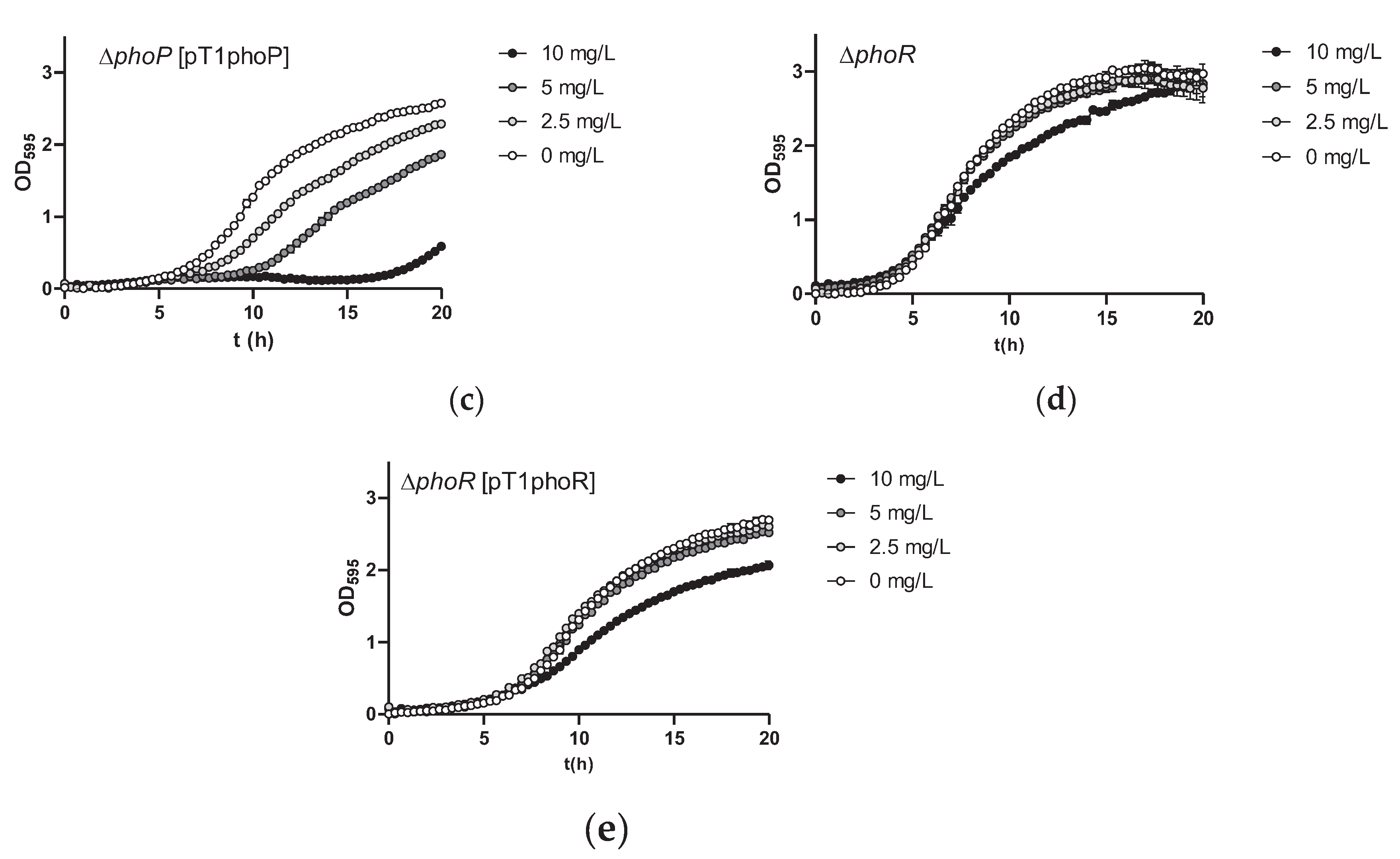
Figure 5.
Inorganic As speciation in bacterial cell pellets of different Lc. paracasei strains. Cells were grown in MEI medium containing 5 mg/L As(III) and the contents of As(III) and As(V) in the bacteria were quantified and referred to g As per gram of wet mass (n=3). Error bars are standard deviations. Asterisks indicate statistical differences compared to wild-type (p<0.01).
Figure 5.
Inorganic As speciation in bacterial cell pellets of different Lc. paracasei strains. Cells were grown in MEI medium containing 5 mg/L As(III) and the contents of As(III) and As(V) in the bacteria were quantified and referred to g As per gram of wet mass (n=3). Error bars are standard deviations. Asterisks indicate statistical differences compared to wild-type (p<0.01).
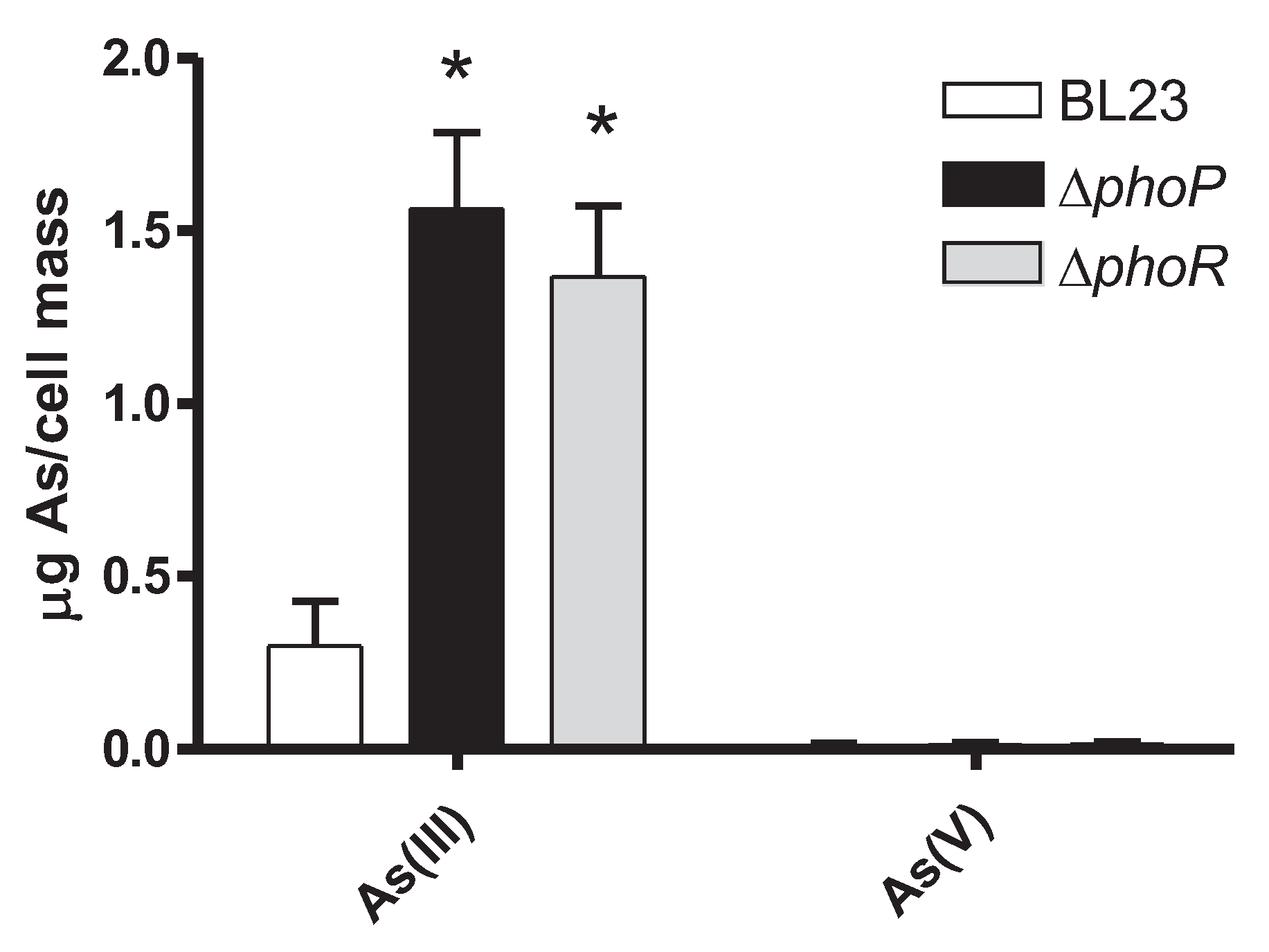

Table 1.
Percentages of retention of As(III), As(V) or DMA by different lactobacilli strains.
| Strain designation and collection code | % As(III) retention | % As(V) retention | % DMA retention |
|---|---|---|---|
| BL7 Levilactobacillus brevis DSMZa 1268 | 0.48 ± 0.08 | 0.06 ± 0.01 | 0.09 ± 0.02 |
| BL10 Lactobacillus acidophilus ATCCb 9224 | 3.28 ± 1.40 | 0.80 ± 0.18 | 0.74 ± 0.13 |
| BL17 Lactobacillus acidophilus ATCCb 4356 | 3.99 ± 0.95 | 0.41 ± 0.27 | 0.88 ± 0.27 |
| BL23 Lacticaseibacillus paracasei CECTc 5275 | 0.08 ± 0.07 | 0.09 ± 0.03 | 0.03 ± 0.02 |
| BL36 Levilactobacillus brevis ATCCb 14869 | 3.01 ± 0.19 | 2.92 ± 0.05 | 0.35 ± 0.01 |
| BL73 Lactobacillus acidophilus CNRZd 55 | 2.60 ± 0.32 | 0.77 ± 0.42 | 0.83 ± 0.35 |
| BL75 Lactobacillus acidophilus CNRZd 21 | 3.27 ± 0.06 | 0.49 ± 0.12 | 0.51 ± 0.35 |
| BL166 Lactiplantibacillus plantarum WCFS1e | 1.26 ± 0.12 | 0.01 ± 0.03 | 0.47 ± 0.05 |
| BL221 Lactobacillus crispatus M247f | 1.20 ± 0.10 | 0.36 ± 0.14 | 0.24 ± 0.08 |
| BL278 Lactobacillus crispatus DSMZa 20584 | 1.38 ± 0.04 | 0.27 ±0.01 | 0.37 ± 0.04 |
| BL279 Lactobacillus acidophilus CECTc 4529 | 1.25 ± 0.12 | 0.20 ± 0.09 | 0.41 ± 0.10 |
| BL280 Lactobacillus acidophilus CECTc 4179 | 2.41 ± 0.15 | 0.41 ± 0.13 | 0.76 ± 0.41 |
| Lpp+ Lactiplantibacillus plantarumg | 2.55 ± 0.16 | 5.70 ± 0.16 | 1.12 ± 0.09 |
The strains were incubated at 37°C for 1 h with different As species (1 mg/L) at an adjusted OD595nm of 10 in PBS. Cells were washed with one volume of PBS and As retained by the cells was quantified. DMA, dimethylarsinic acid. aDeutsche Sammlung von Mikroorganismen und Zellkulturen; bAmerican Type Strain Culture Collection; cColección Española de Cultivos Tipo (CECT); dCentre National de Recherches Zootechniques (CNRZ); e[23]; f[24]; gCollection of the Lactic Bacteria and Probiotics Laboratory [Instituto de Agroquímica y Tecnología de Alimentos (IATA)].
Table 2.
Lp. plantarum and Lc. paracasei strains and their derived mutants used in this study.
| Strain | genotype | reference |
|---|---|---|
| Lp. plantarum WCFS1 | wild type | [23] |
| Lp. plantarum DC421 | WCFS1 pstC::pRV300; eryRa | This work |
| Lp. plantarum DC423 | WCFS1 phoP::pRV300; eryR | This work |
| Lp. plantarum DC425 | WCFS1 phoU::pRV300; eryR | This work |
| Lp. plantarum Lpp+ | wild type | [38] |
| Lp. plantarum DC424 | Lpp+ pstC::pRV300; eryR | This work |
| Lc. paracasei BL23 | wild type | [26] |
| Lc. paracasei DC399 | BL23 pstC::pRV300; eryR | This work |
| Lc. paracasei TC04 | BL23 phoP::pRV300; eryR | [20] |
| Lc. paracasei DC398 | BL23 phoU::pRV300; eryR | This work |
| Lc. paracasei DC487 | BL23 phoP | This work |
| Lc. paracasei DC488 | BL23 phoP [pT1phoP]; eryR | This work |
| Lc. paracasei DC489 | BL23 phoR | This work |
| Lc. paracasei DC490 | BL23 phoR [pT1phoR]; eryR | This work |
a erythromycin resistance.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



