
Submitted:
08 March 2024
Posted:
12 March 2024
You are already at the latest version
Abstract
Non-coding RNAs have been described as crucial regulators of gene expression and guards for cellular homeostasis. Some recent papers are focused on vault RNAs, one of the classes of non-coding RNA, and their role in cell proliferation, tumorigenesis, apoptosis, cancer response to therapy, and autophagy, which made them potential therapy targets in oncology. In the human genome, four vault RNA paralogues can be distinguished. They are associated with vault complexes, considered the largest ribonucleoprotein complex. The protein part of these complexes consists of a major vault protein (MVP) and two minor vault proteins (vPARP and TEP1). The name of the complex, as well as vault RNA, comes from the hollow barrel-shaped structure that reminds a vault. Their sequence and structure are highly evolutionary conserved and show many similarities in comparison with different species, but vault RNAs show various roles. Vaults were discovered in 1986, and their functions remained unclear for many years. Although not much is known about their contribution to cell metabolism, it has become clear that vault RNAs are involved in various processes and pathways associated with cancer progression and modulating cell functioning in normal and pathological stages. In this review, we discuss known functions of the human vault RNAs in the context of cellular metabolism, emphasizing processes related to cancer and cancer therapy efficacy.
Keywords:
vault RNA
; drug resistance
; apoptosis
; autophagy
; cancer
; aging
; cellular metabolism
; non-coding RNAs
; cancer therapy
1. Introduction
The non-coding RNA pool consists of a wide spectrum of RNA species with a broad size range, from the very small microRNA (17-25 nt) to large, long non-coding RNAs (more than 200 nt) and a broad range of cellular functions. Different biogenesis pathways characterize the subclasses of non-coding RNAs. They are transcribed by different polymerases (rRNAs are Pol I transcripts, miRNAs and lncRNAs are typically transcribed by RNA Pol II, while vtRNAs are Pol III transcripts), post-transcriptional processing, and their ability to interact with proteins [1]. Vault RNAs (vtRNAs) have been described in the mid-1980s as a class of small non-coding RNAs, and together with three proteins, i.e., TEP1 (telomerase associated protein 1), MVP (major vault protein), vPARP (vault-associated poly(ADP-ribose) polymerase), are components of the largest identified so far, eukaryotic ribonucleoprotein particles (RNPs), termed vaults [2]. They are massive barrel-shaped particles with a size of 40 x 40 x 70 nm and with a molecular weight ~ 13 MDa [3,4,5]. These assemblies comprise 78 copies of major vault protein (MVP) and additional proteins, TEP1 and vPARP [6]. Vaults are mainly located in the cytoplasm, but some reports confirmed their localization also in the nuclear membrane, within or near the nuclear pore complex (NPC) [7,8,9]. The vault RNP structure is characterized by rapid redistribution in response to external stimuli or stresses and is hypothesized to facilitate protein exchange between the cytoplasm and nucleus. Furthermore, the vault complex and its components are considered mediators of the nuclear-cytoplasmic translocation in normal and cancer cells because the shuttle of these molecules between cellular compartments in response to external factors involved in tumorigenesis may be crucial for the cellular vault function [6,10,11,12]. Interestingly, confocal laser microscopy and cryo-immunoelectron microscopy assessment in U373 cells revealed nuclear localization of part of the major vault protein [13]. The role of TEP1, MVP, and vPARP is reported in various cellular processes and diseases, such as multidrug resistance, apoptosis, cancer progression, and epilepsy[14,15,16,17,18,19,20,21,22,23,24]. The protein components of vaults are also considered risk and/or prognostic factors in cancer [17,25,26,27]. However, since the association status of the vault RNAs with the vault complex affects and modifies their action, the exact role of vault RNA still needs to be discovered. vtRNAs were initially thought to be solely located in vault particles, but literature data demonstrated that less than 5% are bound to vault complex. The majority of vtRNAs (95%) are not associated with vault complex and are located evenly in the cytoplasm as free molecules [28]. Moreover, published data strongly support the hypothesis suggesting that vaults bind the RNA molecules with different affinities depending on the cellular metabolism status, and other factors. The ratio in which the vault RNAs are associated with vaults might be of functional importance [8]. Thus, the mentioned differences in the level of RNA molecules in vault complexes and outside them may be significant in using vtRNAs as targets and/or markers in human diseases. Regarding the role of vtRNA in various cellular processes and their involvement in multiple mechanisms (Table 1), the first evidence came from the studies of EBV (Epstein-Barr virus) infection in B-cells. The studies showed that upregulation of vtRNA transcripts during infection reduced infected cells' apoptosis (inhibition of the extrinsic and intrinsic apoptotic pathways). The first report showed that the effect is mediated not via MVP but via specific action of vtRNA, namely vtRNA1-1. Interestingly, the central domain of vtRNA that distinguishes vtRNA1-1 from its paralogues is essential for its function, i.e., apoptosis resistance [29,30,31]. The members of non-coding vault RNAs are found in many eukaryotes. In human genome four vtRNA paralogs (vtRNA1-1, vtRNA1-2, vtRNA1-3, and vtRNA2-1) are found. RNA polymerase III transcribes them, and they are ca. 100 nt long (vtRNA1-1 98 nt, vtRNA1-2 88 nt, vtRNA1-3 88 nt, vtRNA2-1 100 nt) [32]. To date, among the four mentioned vtRNA paralogues in humans, vtRNA1-1 is the most well-known (Figure 1) [5,33,34,35,36,37,38].
Furthermore, the vtRNA1-1 gene shows the highest expression level among all vtRNA coding genes, potentially resulting from a unique secondary B-box (B2) element [8]. When it comes to the structure of vtRNAs coding genes, it is worth noting that a polymerase III type 2 promoter controls all vault RNA genes and contains box A and box B motives commonly present in tRNA coding genes [39]. Interestingly, the box A and box B sequences in VTRNA-1 and VTRNA-2 are crucial and bind transcription factors TFIIIC and TFIIIB, which facilitate polymerase III binding to the transcription start site. Thus, these elements play a critical role in tight control of polymerase III activity and, consequently, control of cellular homeostasis [40]. However, functional vtRNA genes also have many upstream elements that may distinguish active vtRNA genes from untranscribed ones. vtRNAs are rich in secondary structure elements (they form imperfect hairpin structures by base pairing; 2D structures of four human vault RNA paralogues are presented in the graphical abstract), but their role remains unclear [32,41]. Some single-stranded regions may pair with other transcripts, possibly with proteins and small molecules. Interestingly, vault RNAs are similar to other enigmatic but better-described family of ncRNAs called Y-RNAs, another class of Pol III transcripts [32,42]. Y-RNAs are part of Ro60 ribonucleoprotein particles that function in immune cell communication, immunopathology, antiviral immunity, and HIV-1 infection pathway [43]. Y-RNAs as other RNA Pol III transcripts due to their secondary structure mimicking viral structure may act as innate immune guardians during cell infection with viruses. Y-RNAs are cellular RIG-I ligands mobilized upon HIV-1 infections, thus they have an ability to trigger RIG-I-dependent immune response [43]. These molecules may be critical in maintaining cellular functions, mainly because their high abundance in human plasma [44] makes them good biomarker candidates, e.g., in breast cancer (BC). Notably, BC patients were characterized by increased levels of Y-RNA [44,45]. Although vtRNAs may play similar functions, they are less conserved, and it is suggested that they emerged recently in the evolution [32]. Consequently, the number of vtRNA genes varies between vertebrates and is absent in bacteria, fungi, nematodes, and plants. Both classes of RNAs have a similar size and are controlled by similar promoters, appear in a single or a small number of functional copies, and are typically transcribed from a small gene cluster. Interestingly, both families (vtRNA and Y-RNA) are stably related to their adjacent protein-coding genes for unknown reasons, and it is hypothesized that this association may significantly impact their function [32,42]. The only significant distinction is the composition of the Y-RNA cluster in tetrapods (four or five ancient paralogues). In contrast, clustered vtRNA paralogues are evolutionarily younger and have arisen independently multiple times. Because the molecular function of vtRNAs remains unclear, the speculation about the reason for lineage-specific evolutionary patterns seems pointless [32]. However, literature data revealed that the genes encoding vtRNAs and Y-RNA are associated with adjacent protein-coding genes. The entire, larger genome fragment was copied during evolution in various organisms [32,42,46]. In humans, four vault RNAs are encoded on chromosome 5q31 in two loci. The VTRNA-1 locus contains the genetic information for three vault RNAs (vtRNA1-1, vtRNA1-2, and vtRNA1-3) and is located between the zinc-finger matrin-type 2 gene and proto-cadherin cluster. Recent studies published by Alagia et al. revealed that Dicer-dependent small vault RNA1-2 (svtRNA1-2) is associated with Argonaute 2 (Ago2) and, in contrast to endogenous vtRNA1-2, is localized predominantly in the nucleus. It is postulated that svtRNA1-2 can modulate the expression of genes coding for cell membrane proteins, such as proteins involved in signaling and glycoproteins. Furthermore, the knock-down of vtRNA1-2 results in impaired cellular proliferation and upregulation of genes involved in cellular signaling associated with proliferation, cell adhesion, and migration.
Interestingly, protein-coding genes in the vtRNA1-2 neighborhood, zinc-finger matrin-type 2 and proto-cadherin, play similar roles. Thus, it suggests an association at the expression modulation level between genes coding for vtRNAs and adjacent proteins. However, the molecular basis of this functional link requires further research [32,47].
2. Non-Coding RNAs and Their Regulatory Role in Cancer
In general, non-coding RNAs are critical players in transcription regulation, and they can control the post-transcriptional regulation of mRNA under physiological conditions. The role and mode of action of microRNAs or lncRNAs, to date, have been widely examined under physiological and pathological conditions, and various reports suggest their role in splicing, RNA editing or mRNA degradation, protecting genomes from external nucleic acids, involvement in DNA synthesis or genome rearrangement [48]. Moreover, altered expression of non-coding RNAs can also contribute to several pathologies, including cancer [10], genetic disorders including DiGeorge syndrome, phenylketonuria, and neurodevelopmental defects (comprehensively reviewed by Mattick et al. [49]). Notably, the unknown mechanism of vtRNAs makes them the most mysterious among the various non-coding RNAs. Interestingly, as mentioned above, although vault RNAs were first identified as a component of vaults, data from sedimentation experiments clearly showed that only a fraction of vtRNAs is incorporated into vaults. These results suggest their role in various interactions and processes unrelated to ribonucleoparticles that they can be part of [32,50,51]. In 2000 and 2011, Hanahan and Weinberg proposed six hallmarks of cancer, constituting a logical outline for better understanding the diversity and complexity of neoplastic diseases. Among these characteristics, we can distinguish the ability to avoid apoptosis, insensitivity to anti-growth signals, self-sufficiency in growth signaling, limitless replicative potential, sustained angiogenesis, and increased invasion and metastasis. It has been underlined that the process normal cells must undergo to achieve a neoplastic state is multistep, and a succession of these hallmarks is required. These changes include signaling aberrations during cell proliferation, the ability to evade growth suppressors, and constant replication. Moreover, cancer cells are characterized by resistance to death signaling and active, increased angiogenesis. These changes result in invasive and metastatic phenotypes [52,53]. To date, the new, broad set of functional capabilities acquired by human cells for making their way from normal to neoplastic was proposed by Hanahan [54]. Accordingly, cancer cells are characterized by genome instability and mutations, deregulated cellular metabolism, the ability to evade growth suppressors, avoiding immune destruction, and sustaining proliferative signaling. Additionally, tumorigenesis is tightly associated with tumor-promoting inflammation and replicative immortality [54]. Like other non-coding RNAs, Vault RNAs are key players in various cancer-related processes, i.e., gene expression, regulation of cell proliferation, apoptosis, autophagy, and drug resistance, which is the leading cause of chemotherapy failure [31,35,36,55,56].
3. The Role of vtRNAs in Cancer Cells Proliferation and Tumorigenesis
Cancer is a complex genetic disease arising from elaborate genome changes. These changes include a cumulative collection of gain-of-function mutations that induce oncogenes, loss-of-function mutations that inactivate tumor suppressor genes, and mutations that inactivate stability genes involved in proliferative cell division, all of which help facilitate the transformation of a normal cell to a malignant phenotype [57]. Many findings are compatible with the hypothesis that altered vault RNA expression is associated with tumorigenesis [5,28,55]. The novel pro-survival function of vtRNA1-1 has been described, and these findings support the hypothesis that this molecule plays a crucial role in tumor cell proliferation, tumorigenesis, and chemoresistance (Figure 1). Cell proliferation analyses revealed that the knockout of the vtRNA1-1 but not vtRNA1-3 in HeLa cells led to decreased cell proliferation [31]. A similar negative impact of vtRNA1-1 knockout on cell proliferation was observed in the Huh-7 cell line (hepatocellular adenocarcinoma cells) [55]. In the context of the vault RNAs role in cell proliferation and its regulation, another human paralogue was also analyzed [58]. Studies performed in prostate cancer indicate vtRNA2-1 as an oncogene and tumor suppressor gene. This small RNA molecule is repressed via epigenetic pathways in neoplastic cells, accompanied by increased cell proliferation and invasion. Consequently, it is speculated that vtRNA2-1 (or some derivatives, i.e., snc886-3p - small non-coding RNA derived from nc886) might be considered a valuable diagnostic/prognostic biomarker as it has been repeatedly detected in body fluids, but further research is required especially in the context of its methylation [59,60]. Fort et al. 2020 performed a re-analysis of PAR-CLIP and knockout of microRNA biogenesis enzymes and revealed the association of snc886-3p and snc886-5p products of DICER, independent of DROSHA with Argonaute 2 [59]. Furthermore, the overexpression of the vtRNA2-1 derivative led to the downregulation of mRNAs bearing sequences complementary to its "seed" in 3'-UTRs. Moreover, functional analyses performed in several prostate cancer cell lines revealed that this derivative (snc886-3p) could reduce cell cycle progression and increase apoptosis. Interestingly, the mode of action was similar to the one caused by its precursor vtRNA2-1 [59]. Some recent reports [61,62,63] revealed that vtRNA2-1 did not localize in the ribonucleoprotein complex, in contrast to canonical vault RNA (vtRNA1-1), and did not produce significant amounts of mature miRNAs. There are some indications that vault RNAs are known to produce microRNAs through a Dicer-dependent mechanism, and these microRNAs then operate similarly to regular miRNA [64]. There are some suggestions that this non-coding RNA should be classified as neither miRNA nor vault RNA but rather a 102-nucleotide cytoplasmic small RNA that functionally inhibits PKR (RNA-dependent protein kinase), an interferon-induced kinase involved in innate immune pathways [65]. Vault RNAs can directly interact with other non-coding RNAs and affect cancer cell proliferation. Recently published studies revealed the crucial role of direct interaction of premature vtRNA2-1 with a novel lncRNA, i.e., OXCT1-AS1, in osteosarcoma progression by promoting tumor growth. Interestingly, this interaction is crucial for vtRNA2-1 maturation suppression in this type of cancer. As reported, it plays an inhibitory role in cell proliferation and was demonstrated to contribute to cancer initiation, progression, and, in consequence, metastasis [66,67]. It was also postulated to be associated with tumor evolution and cancer therapy response. Although the new signaling axis OXCT1-AS1/ vtRNA2-1 was identified, the mechanism and detailed functions on the molecular level of the downstream target genes of this axis remain unclear. This newly reported relationship requires further studies, not only in osteosarcoma but also in other cancer types. Especially since the obtained results strongly suggest that overexpression of OXCT1-AS1 and a ratio of mature/premature vtRNA2-1 might be useful prognostic parameters in osteosarcoma patients [66,67]. Moreover, OXCT1-AS1 is known as an indicator of poor prognosis in glioblastoma (involved in the regulation of the miR-195/CDC25A axis [68] as well as a metastasis promoter in non-small-cell lung cancer (by stabilizing LEF1/lymphoid enhancer factor 1) [69].
4. vaultRNAs as Powerful Diagnostic and Cancer Therapy Monitoring Markers
Interestingly, literature data clarified the involvement of vtRNAs in acquiring an aggressive profile of TNBC (triple negative breast cancer). It was shown that TNBC-associated vtRNA1-1 was upregulated in estrogen receptor (ER) and progesterone receptor (PR) negative tumors, whereas vtRNA1-2 seems to be associated only with ER-negative type of breast cancer. Furthermore, it is suggested that vtRNA1-1 may discriminate cancer from normal tissues, inhibit apoptosis, and affect chemoresistance. These findings support the hypothesis suggesting prognostic and therapeutic potential of vtRNAs in breast cancer, especially vtRNA1-1 [70]. Consequently, it was shown as an excellent therapeutic biomarker for endocrine tumors (Bornstein et al. [71]). Interestingly, the correlation between vtRNA1-1 level and prognosis and/or response to treatment in endocrine tumors may be independent of tumor type and applied therapy but dependent on specific responsiveness. It was demonstrated that intracellular upregulation of vault RNAs and their specific exosomal release into the tumor microenvironment (TME) was directly correlated with successful tumor tissue damage after ASA 404 (Vadimezan), EDPM (etoposide, doxorubicin, cisplatin, mitotane) or LEDPM (etoposide, liposomal doxorubicin, liposomal cisplatin, and mitotane) administration [71]. Recently published data from Kato et al. revealed that in patients with blood diseases, the serum vtRNA1-1 levels might show some clinical significance [61]. Serum vtRNA1-1 levels correlated with leukocyte counts and significantly increased in leukemia and lymphoma patients. During intensive chemotherapy treatment, the level of vtRNA1-1 in serum decreased within these groups of patients. Notably, the serum vtRNA1-1 levels varied significantly in patients with hematological malignancies. Thus, the level of vtRNA1-1 might be potentially used as a biomarker of normal and malignant hematological disorders [58]. Furthermore, these molecule may be considered also as therapy efficacy index.
5. Epigenetic Regulation of vtRNAs Expression in Cancer
Genome methylation, one of the most critical mechanisms that control gene expression, also refers to genes coding for vtRNAs. As shown in acute myeloid leukemia (AML), vault RNA2-1 could play a potential role in the biology, progression, and prognosis of AML. It was found that vtRNA2-1 was monoallelically methylated in 75% of the control population, and the remaining 25% showed biallelic hypomethylation. It is worth noticing that leukemia patients with unmethylated vtRNA2-1 showed better overall survival compared to monoallelic or biallelic methylation carriers. Moreover, vtRNA2-1 induction (after demethylation provoked by 5-azanucleosides) was associated with a decreased level of phosphorylated PKR. This kinase activity has been shown to be modulated by vault RNA2-1 which suggested that vtRNA2-1 could be considered a tumor suppressor gene (TSG) in AML since pPKR is known to act as a leukemic cell survival promoter. Thus, the methylation status of vault RNA2-1 is a potential predictor of the therapy outcome in AML patients [72].
A variety of evidence revealed that vtRNA transcription is under the control of promoter gene methylation. Analysis of methylation status (and consequently expression profiling prediction) of the non-coding RNA promoters might be crucial in the context of the prognosis for cancer patients. In the case of hepatocellular carcinoma (HCC), a significant correlation between the methylation status of the vtRNA2-1 promoter and various features of the tumor that affects the prognosis for the patient, such as the presence of large tumors, pathological vascular invasion, more advanced tumor stage or tumor recurrence, was identified. Moreover, the methylation of the vtRNA2-1 promoter increased in stage III of HCC when compared with less advanced stages. The methylation status of the vtRNA2-1 promoter is also very beneficial for patient outcomes after liver resection. Patients with differentially hypermethylated promoters in tumor tissues compared to their adjacent normal tissues should receive more intensive care or adjuvant therapy after surgery [73].
Dugué and collaborators revealed that the methylation status of vtRNA2-1 region is heritable, modifiable, and highly important, associated with increased cancer risk and a worse prognosis [74]. Interestingly, the results of the present cohort study showed no link between epigenetic factors and the individual’s lifestyle, i.e., age, smoking, diet, and blood cells DNA methylation at vtRNA2-1. These results might prove the leading role of genetics and adult lifestyle in explaining methylation variability at the heritable vtRNA2-1 cluster. Moreover, the investigated genetic and non-genetic factors are minimal in explaining variation in blood cells DNA methylation at vtRNA2-1. Thus, their potential role in the observed association between vtRNA2-1 methylation status and breast cancer risk seems insufficient. Furthermore, the mechanism of DNA methylation inheritance in this region needs further investigation [74]. vtRNA2-1 was also perceived as a tumor suppressor gene (due to its ability to inhibit PKR) in prostate [75,76], cholangiocarcinoma [77], skin [78], and gastric cancer cells [79]. These reports remain concordant with the data concerning lower expression levels of vault RNA2-1 in cancer cell lines compared to normal tissues. Furthermore, the methylation of the vtRNA2-1 promoter increases from normal to tumor samples, whereas in the case of vtRNA1-2, the trend was the opposite. Interestingly, low DNA methylation at the promoter region of vtRNA1-1, vtRNA1-2, and vtRNA2-1 was associated with lower patients’ survival rate [80]. Furthermore, Kunkaew and collaborators showed the importance of vtRNA2-1 suppression in cholangiocarcionoma [77]. They found that some cancer patients were characterized by elevated PKR activity in cholangiocyte cells [77]. This finding was surprising because PKR is mainly known for its role as a pro-apoptotic factor through its ability to phosphorylate eIF2α in the host defense mechanism during viral infection. Researchers observed that vtRNA2-1 was capable of suppressing PKR via direct physical interaction. The starting point for further research suggests a potential role of the vtRNA2-1 suppression and then PKR activation during pre-malignant cells elimination in tumorigenesis. It is concluded that PKR plays a dual role during tumorigenesis. On the one hand, when the level of vtRNA2-1 is low (or repressed), cells are directed toward canonically programmed cell death pathways. Interestingly, PKR plays a dual role in tumorigenesis. On the one hand, active PKR in cholangiocarcinoma cells does not induce apoptosis or an eIF2α-dependent pathway. On the other hand, it promotes the pro-survival NF-κB pathway [77].
According to the data published by Hu et al.[81] higher level of vtRNA2-1 was expressed in the late phase of human endometrial cancer tissue, less than in the early phase but still higher than in normal human endometrial tissue. Significantly, after vtRNA2-1 silencing, the expression and protein levels of pPKR (phosphorylated PKR) and caspase-3 were increased, whereas NF-κB and VEGF were decreased. Furthermore, the apoptosis rate in the vtRNA2-1 -silenced group was increased, and the cell proliferation rate was slower compared to the control [81]. Interestingly, the opposite results indicating the oncogenic and anti-apoptotic mode of action of vtRNA2-1 was proposed in various cancer tissues, i.e. renal [82], endometrial [81], as well as ovarian [62].
In the male population, prostate cancer shows the highest incidence and mortality among all cancer types and is characterized by high heterogeneity. Researchers found that vtRNA2-1 dysregulation might be an essential and valuable prognostic marker in this type of cancer. It was found that upregulation of this ncRNA is not only associated with poor survival rate in patients but also significantly promoted proliferation and invasion as this molecule was a tumor promoter. Moreover, the knockdown of vtRNA2-1 related to inhibiting prostate cancer cell proliferation and their invasiveness. Therefore, it could be considered a novel therapeutic target [83]. The role of vtRNAs was also reported in cervical cancer (CSCC, cervical squamous cell carcinoma), the second-most common cancer in women worldwide[84]. Specifically, oncogenic activity and contribution to cervical cancer progression of vtRNA2-1-5p were reported. This non-coding RNA is AGO-associated small RNA derived from vtRNA2-1 and it was upregulated in human cervical squamous cell carcinoma compared to normal tissue [85]. Interestingly, published data demonstrate that vtRNA2-1-5p has a specific role only in cervical cancers. Moreover, vtRNA2-1-5p can directly target p53 expression and act as an oncomir. As observed, its inhibition was associated with decreased proliferation and tumorigenicity and increased apoptosis of CSCC. Thus, vtRNA2-1-5p was suggested to be a direct regulator of p53 expression, playing an essential role in the apoptosis and proliferation of tumor cells. Furthermore, the inhibition of vtRNA2-1-5p sensitized cells to cisplatin treatment and directed them to the apoptotic pathway [85].
The literature data point that the expression of vtRNA2-1-3p can be specifically reduced, e.g., by ARE (arecoline, a nicotinic acid-based mild parasympathomimetic stimulant alkaloid found in the areca nut) that was reported in oral squamous cell carcinoma. What is more, the mentioned suppression can play a critical role in the proliferation and migration of cancer cells and might affect the invasiveness of OSCC cells. These findings may be crucial in identifying new targets and treatment strategies in patients with this type of cancer [86].
Another interesting case showing the involvement of vault RNAs’ in tumorigenesis and patients’ prognosis was low-risk myelodysplastic syndrome (MDS). MDS are heterogeneous clonal diseases characterized by ineffective hematopoiesis associated with increased apoptosis and differentiation block of early progenitors. It was shown that vault RNAs might also act as tumor suppressors in a specific manner based on epigenetic modifications, mainly DNA methylation. The hypothesis put forward by Helbo, and colleagues suggests that vtRNA1-3 is silenced by DNA methylation in the acute promyelocytic leukemia cells (HL60) but was unmethylated in normal hematopoietic cells. Thus, the downregulation mentioned above seems to be cancer- specific. Furthermore, cohort studies revealed that vtRNA1-3 promoter hypermethylation was frequent in lower-risk MDS patients and was associated with a decreased overall survival. However, analysis of the relative impact of VTRNA1-3 promoter methylation on 5-year overall survival strongly suggests that it should not be considered an independent prognostic marker [87].
6. Vault RNAs as a Regulator during the Autophagy
Autophagy is a fundamental eukaryotic pathway that helps maintain metabolic homeostasis. This process is observed under physiological conditions, i.e., autophagy promotes embryonic organogenesis such as neural tube formation, neuronal differentiation, reticulocyte development, myocardial growth, chondrocyte survival, and differentiation [88]. Autophagy is also observed in pathological conditions, i.e., inflammation, neurodegeneration, and cardiovascular diseases or carcinogenesis [89]. Autophagy in cancer is highly correlated with the innate and adaptive immune system and is mediated by cytokine signaling [90]. One of the critical autophagy markers, p62 protein, also called sequestosome 1 (SQSTM1), acts as a critical receptor that identifies and binds ubiquitinated proteins, delivering them via membrane-bound LC3 to the phagophore for subsequent degradation. p62 is also an RNA-binding protein that can engage small non-coding vault RNA1-1 as a major interacting RNA. Vault RNA1-1 regulates p62-dependent autophagy via interference with p62 multimerization-critical step for biological activity of p62 protein. Vault RNA1-1 directly modulates the p62 oligomerization that is critical for autophagy control. Specifically, vtRNA1-1 acts as a riboregulator of key cellular processes via direct interaction/direct binding to the autophagic receptor p62, thus changing the protein function [91]. Interesting results in the context of the regulatory role of vtRNA1-1 in lysosome metabolism were published by Ferro and collaborators. Specifically, it was revealed that lack of vtRNA1-1, both in vitro and in vivo mouse model, reduced drug lysosomotropism and inhibited tumor cell proliferation in hepatocellular carcinoma cells [55]. Moreover, vtRNA1-1 knockout in human hepatocellular carcinoma (HCC) cells leads to lysosomal compartment dysfunction via TFEB nuclear translocation inhibition, resulting in a downregulation of the CLEAR network genes. Thus, TFEB affects the expression of genes involved in lysosomal biogenesis, lysosome-to-nucleus signaling and lipid catabolism. vtRNA1-1 depletion resulted in an increased activation of the MAPK (mitogen-activated protein kinase) cascade, MAPK1/ERK2-MAPK3/ERK1, responsible for TFEB inactivation and cytoplasmic retention [55]. Kinases involved in signaling axes ERK1/2 MAPK and PI3K/Akt are described as autophagy regulators [92], cell growth, and proliferation modulators, and are involved in tumorigenesis (reviewed in [93]).
Horos et al. showed that overexpression of human vtRNA1-1 inhibits, while its antisense LNA-mediated knockdown enhances p62-dependent autophagy [34]. Interestingly, starvation significantly reduces the steady state, and p62-bound levels of vault RNA1-1 that lead to autophagy induction [34]. These findings are also significant in tumorigenesis as, to date, p62/SQSTM1 is the best studied mammalian substrate that showed autophagy specificity. Moreover, the modulation of this multidomain protein during autophagy is an inestimable factor in tumorigenesis. Data from extensive research employing knock-out, transgenic, and knock-in mice strongly suggest its critical role in several cellular functions, including bone remodeling, obesity, and cancer development and progression (reviewed in [94]). To date, the mechanism through which vtRNA1-1 can inhibit p62 oligomerization and the specificity of their interaction is lacking. Recently, Büscher et al. showed crucial components for binding between those two molecules [35]. It was found that the adjacent linker region of p62, PB1 domain, and central flexible loop within vault RNA1-1 are necessary for specific binding with vtRNA1-1. Moreover, they revealed lysine 7 and arginine 21 as the two critical regulatory points of p62 [35].
7. Vault RNAs Act as Apoptosis Modulators
Apoptosis is a tightly regulated and highly conserved type of cell death. This cascade of reactions and processes plays crucial roles in normal physiological conditions and pathological states, e.g., cancer. It involves the activation of caspases[95] but can also be associated with the major vault protein that was postulated as an apoptosis inhibitor in senescent cells [24]. However, very little is known about the role of vtRNA in apoptosis. In 2015, the first data concerning the general apoptotic resistance upon vtRNA1-1 expression in malignant B cells was published. Amort and collaborators identified latent membrane protein 1 (LMP1) expressed in most EBV-associated lymphoproliferative diseases and malignancies [96] as a trigger for NF-κB-dependent vtRNA1-1 expression. Ectopic expression of vtRNA1-1 in a B-cell line that usually lacks this ncRNA led to increased susceptibility to EBV infection due to attenuated apoptosis. Furthermore, knockdown experiments of the MVP clearly showed that the anti-apoptotic effect is driven by vtRNA1-1 and is not associated with the vault complex. Moreover, vtRNA1-1 modulates both the intrinsic and extrinsic apoptosis pathway [56]. By next-generation deep sequencing of the mRNome, Bracher and collaborators identified the PI3K/Akt pathway and the ERK1/2 MAPK cascade, which were dysregulated in the absence of vtRNA1-1 during starvation-mediated cell death. They showed that the expression of 24 nucleotides of the vtRNA1-1 central domain seems crucial for maintaining apoptosis resistance. Interestingly, similar resistance to apoptosis in the lack of vtRNA1-1 was also observed in two other cell lines (A549 and HEK293) [31]. These results strongly suggest that apoptosis is dependent on vtRNA1-1 level.
Kunkaew and collaborators discovered a cytotoxic mechanism based on the role of cellular non-coding RNA vtRNA2-1 and PKR that acts as a proapoptotic protein. In normal cells, vtRNA2-1, right after its transcription, can bind to PKR and prevent PKR from aberrant activation. In cancer it was observed that vtRNA2-1 transcription was turned off after DOX treatment due to Pol III detachment from DNA. When the available pool of vtRNA2-1 runs out, PKR does not have a binding partner, and cells are directed to apoptosis.
8. vtRNAs and Drug Resistance
Identifying efficient cancer treatment remains an important goal for both researchers and clinicians. Currently, chemotherapy is seen as one of the most promising cancer treatments to reduce the burden of cancer. However, chemotherapy fails in nearly 90% of cases as cancer cells develop resistance to the anticancer agents, resulting in increased cancer invasion, progression, and metastasis [97]. Participation of vault particles in development of drug resistance in cancer cells has already been demonstrated. Moreover, MVP (also known as LRP, lung resistance-related protein) is generally overexpressed in human cancer cells and is involved in resistance phenotype formation [19,21,98]. According to The Human Protein Atlas data, MVP is mainly accumulated in colorectal, renal, pancreatic, lung and thyroid cancer (access: 2024/02/27). The role of vaults in therapy resistance may be associated with their ability to transport drugs away from their intracellular targets, e.g., from the nucleus or mediate the function of efflux pumps or exocytotic vesicles. Moreover, based on the characteristics of the minor vault proteins, vaults or vault components are likely involved in the maintenance of genomic stability (reviewed in [19]). Specifically, vault RNAs, especially vtRNA1-1 and vtRNA1-2, were reported to bind mitoxantrone, one of the chemotherapeutic agents commonly used in malignancies, leading to the sequestration of this drug and, consequently, decreased treatment efficacy. Further investigation revealed overexpression of vault RNAs in cell lines derived from human glioblastoma, leukemia, and osteocarcinoma. Moreover, these cells showed increased resistance to mitoxantrone, and vtRNA downregulation led to significant decrease in cells’ tolerance to this chemotherapeutic agent. These data strongly suggest an essential role of vault RNAs in susceptibility to mitoxantrone in tested malignant cells [33].
Furthermore, van Zon and collaborators showed that the elevated level of vtRNA1-3 may be mediated by the change in the affinity of the vault complex to the vault RNA species in response to specific functional cues. Moreover, multidrug-resistant (MDR) cells were characterized by higher levels of vault RNA1-3 associated with vault complexes. Consequently, the ratio of the vault-associated RNA species was suggested to influence the drug response [8].
Vault RNAs can mediate drug resistance in an MVP/vault complex-independent manner. By vtRNA1-1 or MVP knockdown in MCF-7 cells, Chen et al. revealed that chemoresistance mediated by vtRNA did not depend on MVP and, what is more, confirmed the role of vtRNA1-1 in regulating PSF (polypyrimidine tract binding protein associated splicing factor) transcriptional activity independent with the expression of MVP [36]. It has been found that vtRNA1-1 could bind to the RBD on PSF protein, and when the PSF expression was compromised, the cancer cells became sensitive to doxorubicin. On the other hand, the binding of vtRNA1-1 to PSF released its target gene, GAGE6, and thus affected its expression level, and cells became more sensitive to the treatment. Thus, vtRNA1-1 shows a very important role in cancer cell drug resistance [36]. Moreover, it is suggested that a crucial function in drug resistance and cancer progression is played by lysosomes (reviewed in [99]). Ferro et al. showed that vtRNA1-1 knockout in human hepatocellular carcinoma cells (HCC) led to lysosomal compartment dysfunction. This abnormality resulted from inhibited TFEB (transcription factor EB) nuclear translocation and downregulation of the CLEAR (coordinated lysosomal expression and regulation) network genes. Moreover, the depletion of vtRNA1-1 was associated with increased activation of the MAPK cascade [55].
vtRNA2-1 was identified as a molecular signal to sense DOX, and this finding explains why drugs that can cause DNA damage might also be cytotoxic for cells with the competent vtRNA2-1/PKR pathway [61]. One of the leading hypotheses on how vtRNAs and vault particles could be associated with drug resistance is based on PKR activity. Interestingly, it was shown that one of the vault RNA derivatives (svRNAb) is associated with Argonaute proteins and can drive specific cleavage, thus acting and regulating gene expression similarly to miRNAs. svRNAb can downregulate one of the most essential enzymes in drug metabolism, CYP3A4. This finding might be beneficial in the association of vault RNA and drug resistance. It can help explain the broad spectrum of potential mechanisms involved in these processes [38].
9. Vault RNAs in Aging
Undoubtedly, the occurrence and development of many diseases, including cancer, are related to aging. In recent years, an increasing number of researchers have concluded that immunological factors play an increasingly important role in physical degeneration and pathological changes, which may be essential targets for assessing and treating older individuals with cancer. Aging is a complex process that profoundly affects the immune system. The impairment of the immune system with age is manifested by increased susceptibility to infectious diseases, poorer response to vaccinations, autoimmune and other chronic diseases, and increased cancer incidence [100]. To date, recent reports indicate that overexpression of vtRNA1-1 may attenuate apoptosis in a cellular model of Epstein-Barr virus infection [29,56] and accelerate apoptosis in influenza virus via PKR pathway (the major component of the cellular antiviral system) inactivation [101,102]. Furthermore, influenza A virus induced robust expression of vtRNAs in host through NS1 protein. In turn, the increased expression of vtRNAs was essential for NS1-mediated inhibition of PKR activation. These findings strongly support hypothesis that viruses, including influenza A virus, can overcome PKR-mediated innate immune response through NS1-dependent upregulation of vtRNAs [102]. Since vtRNAs play an essential role in the epigenetic regulation of target gene expression, they are suggested to contribute to cancer or other disease development and aging. Kim and collaborators were interested in determining whether chemical compounds found in green tea that can control vtRNA2-1 expression also have a role in the senescence of fibroblasts. Specifically, they tested AbsoluTea Concentrate 2.0 (ATC, absolute extract from green tea leaves obtained from Camelina sinensis var. sinensis cv. Jangwon No.3). They revealed that tested extracts led to increased vtRNA2-1 expression that resulted in a suppression of cellular senescence that was manifested by a decrease in accumulation of cell senescence biomarkers, such as SA--gal activity and p16INK4A, and p21Waf1/Cip1 expression. Thus, this non-coding RNA might be a good candidate for an anti-aging target [103].
Similar observations were made by Kwang-Soo Lee and colleagues, who suggested the role of vtRNA2-1 in controlling photoaging and inflammation in skin cells [78] (Figure 2). This study investigated how binding between vtRNA2-1 and PKR and regulation of PKR activity may affect skin aging. They showed that right after UVB radiation, the expression of vtRNA2-1 decreases, whereas the phosphorylation status of PKR via MAPK kinases was increased. Consequently, together with reduced expression of vtRNA2-1, uncontrolled PKR activity, and increased expression of inflammatory cytokines (such as IL-1α, IL-6, and IL-8), cyclooxygenase (COX-2), collagenase type IV, MMP-9 (matrix metalloproteinase-9) (Figure 2), the inflammatory response and aging processes in the skin cells were significantly accelerated [78].
Moreover, Eunsun Jung Lab goes further with studies regarding the role of vtRNA2-1 in photoaging and consistently obtained results suggesting that ultraviolet irradiation can accelerate the methylation of vtRNA2-1 gene [104]. Therefore, its expression is reduced, the PKR pathway is activated, and various pro-inflammatory factors are secreted. However, when vtRNA2-1 was overexpressed, expression and production of those pro-inflammatory factors were inhibited. Moreover, the extract from Laminaria japonica was protecting vault RNA2-1 against UVB, and the biosynthesis of UV-induced inflammatory factors mediated by the PKR pathway was inhibited (Figure 2) [104].
Moreover, overexpression of vtRNA2-1 reduced the production of MMP-9 and inflammatory cytokines. In turn, UVB led to the inhibition of vtRNA2-1 expression by increasing its methylation status. Simultaneously, UVB irradiation accelerated the phosphorylation of PKR, p38, JNK, c-Jun, and ATF-2. These results revealed that increasing vtRNA2-1 expression, which is inhibited by UVB exposure, can inhibit the MMP-9 and inflammatory cytokines. The extract from Laminaria japonica suppressed PKR phosphorylation. Furthermore, treatment with Laminaria japonica extract inhibited the reduction of vtRNA2-1 caused by UVB irradiation and blocked signal transduction via the PKR pathway to decrease the production of MMP-9 and inflammatory cytokines [104].
10. Vault RNAs and Immune Response
Literature data strongly suggest that vtRNA2-1 suppresses the IFN- signaling and controls the inflammation processes. In response to the treatment with various pathogens, vtRNA2-1 not only suppresses the activation of IRF3 (Interferon Regulatory Factor 3) but also, via inhibition of PKR, can inhibit NF-B (Nuclear Factor kappa-light-chain-enhancer of activated B cells) and AP-1 (Activator Protein 1). These findings prove that vtRNA2-1 is tightly connected with the regulation expression of IFN- and the factors needed to activate its promoter. Thus, vtRNA is responsible for decreased expression of IFN- and genes induced by interferon [105].
Interferons, innate immune response elements, mediate this reaction. Among various interferons, the first line of response to the pathogen activity is occupied by IFN-. The impaired control of IFN- might cause autoimmune diseases. Thus, the induction of this molecule and its secretion should be tightly controlled on various levels. vtRNA2-1 might be a direct molecular link between the dependency of innate immunity on epigenetics and environmental factors such as UVB (Figure 2).
The development and proper functioning of the human immune system are closely related to and dependent on commensal microbiota. The microbiota plays emerging roles in the host’s innate and adaptive immune systems. The symbiosis between host and microbes is crucial for avoiding many of immune-related disorders, such as inflammatory bowel disease (IBS), cardiometabolic disease, and cancer (reviewed in [106]). A recently published paper also showed the role of vtRNA2-1 in disrupting the gut epithelial barrier via interaction with HuR, one of the RNA-binding proteins. Increased level of this vtRNA was observed in intestinal mucosal tissue samples from patients with IBS and samples from mice with colitis or sepsis. Higher ectopic expression of vtRNA2-1 was associated with a decreased level of intracellular junction proteins, i.e., claudin 1, occluding, and e-cadherin, which resulted in intestinal epithelial barrier dysfunction in vitro. In contrast, vtRNA2-1 silencing supports the functioning of the intestinal barrier. The results observed in mice agreed with those from in vitro experiments. Mechanistically, vtRNA2-1 interaction with HuR prevents HuR binding to the claudin 1 and occludin mRNA and decreases their translation [107].
11. vtRNAs Role in Neural System Development and Pathology
Vault RNA1-1 functions as a riboregulator of synaptogenesis via MAPK signaling pathway modulation in neurons. In an in vitro synapse formation model, it was shown that murine vtRNA promotes synapse formation. In overview, murine vtRNA is firstly transported to the distal part of neuronal cells as a component of the vault complex and, secondly, released from the complex in the neurite by a mitotic kinase Aurora-A-dependent phosphorylation of MVP. Interestingly, the role in binding and activating MEK1 (mitogen-activated protein kinase) and enhancing MEK1-mediated ERK (extracellular signal-regulated kinases) activation in neurites by vtRNA per se, not as a part of ribonucleoprotein complex, suggests the existence of a novel molecular mechanism for synaptogenesis with vtRNA as a riboregulator in this process [108,109]. Further investigation revealed that only one human vtRNA paralogue could affect synaptogenesis. Expression of vtRNA1-1 increases phosphorylation of ERK as well as the number of PSD95 (postsynaptic density protein 95) and Synapsin I – crucial proteins for synapse maturation and biological function [110]. Interestingly, there is an emerging hypothesis associating vtRNA with an increased prevalence of ASD (autism spectrum disorder). The work of Geoffray and colleagues supports these claims. They found a link between dysregulation of the RAS MAPK signaling pathway and ASD. As MAPK is regulated by vtRNA1-1, it is postulated that vtRNA may also be associated with RASopathies - a group of inherited disorders caused by pathogenic variants of genes encoding regulatory proteins within the RAS-MAPK signaling pathway [111]. These findings need further investigation to find a direct link between vtRNA expression and ASD. Moreover, vtRNA expression in ASD could support the possible clinical translation of ASD-related MEK inhibitor drug therapy [112]. To date, it has been shown that ASD is not the only neurological pathology with the potential role of vtRNA in etiology as well as a potential therapy target. Miñones-Moyano and collaborators suggest that vtRNA2-1 and its deregulation could contribute to pathogenesis in the initial stages of Parkinson’s disease (PD). It was observed that the expression level of this vtRNA was increased in the advanced stages of PD. However, the upregulation level was not strictly correlated with the severity of the neuropathological lesions. Researchers focused their studies on the amygdala in patients without motor symptoms in the early stages without pharmacological treatment. Thus, it might be excluded that drug treatment in PD patients could somehow affect the vtRNAs and other genes [113].
12. Conclusions and Future Perspectives
Vault RNAs play a role in cellular metabolism, and it is crucial to gain a better understanding of how these molecules are involved in normal cells metabolism, processes involved in tumorigenesis and tumor response to treatment. Although vault complexes and vault RNAs are known since over 30 years, still there is an important gap regarding mechanism of their action and molecular basis of their regulatory role. Vault RNAs and whole ribonucleoprotein complexes which they are part of, affect such important processes for cell functioning as autophagy, apoptosis and proliferation. Based on literature data we can assume that vault RNAs may play the role in cell signaling control (e.g. PKR). Vault RNAs are also involved in neural cells development, synaptogenesis, metabolism and pathologies in neural system, neurodegenerative diseases, i.e.; Parkinson’s Disease or Alzheimer’s Disease. This might be extremely important in the context of aging societies and the need to search for new mechanisms aimed at preventing, delaying or even repairing neuronal damage. Moreover, vault RNAs can contribute to multidrug response, especially vault RNA1-1 and vault RNA1-2, via their ability to bind chemical compounds, including chemotherapy agents. Furthermore, since they can bind chemotherapeutic drugs, there is a hypothesis that they can also bind other chemicals including xenobiotics. That fact might be crucial, due to growing environmental pollution level and the need to counteract and prevent their effects on cellular metabolism. Vault RNAs and their regulatory role in variety pathways indicating that they may be considered as potential prognostic and diagnostic markers in cancer. As a subclass of pivotal regulators, vault RNAs, similarly to other non-coding RNAs, may be also crucial for cells development, differentiation and together with their interaction network may be involved in aging and aging-related disorders. Many non-coding RNA have been reported as key regulators and as they can be involved in the regulation of cancer-related genes expression. It is extremely encouraging that also in case of vault RNAs it is possible to combine their activity and expression level with traditional therapy approach curation for cancer patients better prognosis and outcome.
List of abbreviations
| Ago2 | Argonaute 2 |
| AML | acute myeloid leukemia |
| AP1 | activator protein 1 |
| ARE | arecoline |
| ASD | autism spectrum disorder |
| ATF-2 | activating transcription factor 2 |
| ATG5 | autophagy related 5 protein |
| BC | breast cancer |
| CDC25A | cell division cycle 25 A |
| c-Jun | transcription factor Jun |
| CLEAR | coordinated lysosomal expression and regulation network |
| COX-2 | cyclooxygenase |
| CSCC | cervical squamous cel carcinoma |
| CYP3A4 | cytochrome P450 3A4 |
| DICER | endoribonuclease Dicer |
| DOX | doxorubicin |
| DROSHA | class 2 ribonuclease III enzyme |
| EBV | Epstein-Barr virus |
| eiF2α | eukaryotic initiation factor 2 |
| ERK1/ERK2 | extracellular signal-regulated kinases |
| ERKs | extracellular signal-regulated kinases |
| GAGE6 | G Antigen 6 |
| HCC | hepatocellular carcinoma cells |
| IBS | inflammatory bowel disease |
| IFN-β | interferon β |
| IRF3 | interferon regulatory factor 3 |
| JNK | c-Jun N-terminal kinases |
| LAMP1 | lysosomal associated membrane protein 1 |
| LC3II | autophagosome-associated protein LC3-II |
| LEF1 | lymphoid enhancer factor 1 |
| lncRNA | long non-coding RNA |
| MAPK | mitogen activated protein kinase |
| MDR | multidrug resistance |
| MDS | myelodysplastic syndrome |
| miRNA | micro RNA |
| MMP-9 | matrix metaloproteinase-9 |
| MVP | major vault protein |
| NF-κB | nuclear factor kappa-light-chain-enhancer of activated B cells |
| NPC | nuclear pore complex |
| OSCC | oral squamous cel carcinoma |
| OXCT1-AS1 | OXCT1 antisense RNA 1 |
| p16INK4A | protein bound to cyclin dependent kinase 4 and 6 |
| p21Waf1/Cip1 | |
| p38 | p38 mitogen-activated protein kinases |
| p62 | nuclear pore glycoprotein p62 |
| PB1 | Phox and Bem1 domain |
| PI3K/Akt | phosphatidyl inositol 3-kinase/ v-akt murine thymoma viral oncogene homolog 1 |
| PKR | RNA-dependent protein kinase |
| pPKR | phosphorylated PKR |
| PSD95 | postsynaptic density protein 95 |
| PSF | polypyrimidine tract binding protein associated factor |
| RBD | RNA-binding domain |
| RNP | ribonucleoprotein |
| rRNA | ribosomal RNA |
| SA-β-gal | senescence-associated β-galactosidase |
| SQSTM1 | sequestosome 1 |
| TEP1 | telomerase associated protein 1 |
| TFEB | transcription factor EB |
| TME | tumor microenvironment |
| TNBC | triple-negative breast cancer |
| TSG | tumor suppressor gene |
| VEGF | vascular endothelial growth factor |
| vPARP | vault-associated poly (ADP-ribose) polymerase |
Author Contributions
M.T. conceived the work, wrote, reviewed, and edited the manuscript. N.L. wrote, reviewed, and edited the manuscript. E.T. reviewed the manuscript and prepared the figures. B.R. reviewed and edited the manuscript, supervised the work. All authors have read and agreed to the published version of the manuscript.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Zhou, S.; Van Bortle, K. The Pol III Transcriptome: Basic Features, Recurrent Patterns, and Emerging Roles in Cancer. WIREs RNA 2023, e1782. [Google Scholar] [CrossRef] [PubMed]
- Kedersha, N.L.; Rome, L.H. Isolation and Characterization of a Novel Ribonucleoprotein Particle: Large Structures Contain a Single Species of Small RNA. J. Cell Biol. 1986, 103, 699–709. [Google Scholar] [CrossRef] [PubMed]
- Abbondanza, C.; Rossi, V.; Roscigno, A.; Gallo, L.; Belsito, A.; Piluso, G.; Medici, N.; Nigro, V.; Molinari, A.M.; Moncharmont, B.; et al. Interaction of Vault Particles with Estrogen Receptor in the MCF-7 Breast Cancer Cell. J. Cell Biol. 1998, 141, 1301–1310. [Google Scholar] [CrossRef] [PubMed]
- Kedersha, N.L.; Heuser, J.E.; Chugani, D.C.; Rome, L.H. Vaults. III. Vault Ribonucleoprotein Particles Open into Flower-like Structures with Octagonal Symmetry. J. Cell Biol. 1991, 112, 225–235. [Google Scholar] [CrossRef] [PubMed]
- Gopinath, S.C.B.; Wadhwa, R.; Kumar, P.K.R. Expression of Noncoding Vault RNA in Human Malignant Cells and Its Importance in Mitoxantrone Resistance. Mol. Cancer Res. 2010, 8, 1536–1546. [Google Scholar] [CrossRef] [PubMed]
- Frascotti, G.; Galbiati, E.; Mazzucchelli, M.; Pozzi, M.; Salvioni, L.; Vertemara, J.; Tortora, P. The Vault Nanoparticle: A Gigantic Ribonucleoprotein Assembly Involved in Diverse Physiological and Pathological Phenomena and an Ideal Nanovector for Drug Delivery and Therapy. Cancers 2021, 13, 707. [Google Scholar] [CrossRef]
- Chugani, D.C.; Rome, L.H.; Kedersha, N.L. Evidence That Vault Ribonucleoprotein Particles Localize to the Nuclear Pore Complex. J. Cell Sci. 1993, 106, 23–29. [Google Scholar] [CrossRef]
- van Zon, A.; Mossink, M.H.; Schoester, M.; Scheffer, G.L.; Scheper, R.J.; Sonneveld, P.; Wiemer, E.A.C. Multiple Human Vault RNAs. J. Biol. Chem. 2001, 276, 37715–37721. [Google Scholar] [CrossRef]
- Van Zon, A.; Mossink, M.H.; Scheper, R.J.; Sonneveld, P.; Wiemer, E.A.C. The Vault Complex. Cell. Mol. Life Sci. CMLS 2003, 60, 1828–1837. [Google Scholar] [CrossRef]
- Hahne, J.C.; Lampis, A.; Valeri, N. Vault RNAs: Hidden Gems in RNA and Protein Regulation. Cell. Mol. Life Sci. 2021, 78, 1487–1499. [Google Scholar] [CrossRef]
- Kim, E.; Lee, S.; Mian, M.F.; Yun, S.U.; Song, M.; Yi, K.; Ryu, S.H.; Suh, P. Crosstalk between Src and Major Vault Protein in Epidermal Growth Factor-dependent Cell Signalling. FEBS J. 2006, 273, 793–804. [Google Scholar] [CrossRef] [PubMed]
- Chung, J.-H.; Ginn-Pease, M.E.; Eng, C. Phosphatase and Tensin Homologue Deleted on Chromosome 10 (PTEN) Has Nuclear Localization Signal–Like Sequences for Nuclear Import Mediated by Major Vault Protein. Cancer Res. 2005, 65, 4108–4116. [Google Scholar] [CrossRef]
- Slesina, M.; Inman, E.M.; Rome, L.H.; Volknandt, W. Nuclear Localization of the Major Vault Protein in U373 Cells. Cell Tissue Res. 2005, 321, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Pietras, P.; Leśniczak-Staszak, M.; Kasprzak, A.; Andrzejewska, M.; Jopek, K.; Sowiński, M.; Rucinski, M.; Lyons, S.M.; Ivanov, P.; Szaflarski, W. MVP Expression Facilitates Tumor Cell Proliferation and Migration Supporting the Metastasis of Colorectal Cancer Cells. Int. J. Mol. Sci. 2021, 22, 12121. [Google Scholar] [CrossRef]
- Szaflarski, W.; Sujka-Kordowska, P.; Pula, B.; Jaszczyńska-Nowinka, K.; Andrzejewska, M.; Zawierucha, P.; Dziegiel, P.; Nowicki, M.; Ivanov, P.; Zabel, M. Expression Profiles of Vault Components MVP, TEP1 and vPARP and Their Correlation to Other Multidrug Resistance Proteins in Ovarian Cancer. Int. J. Oncol. 2013, 43, 513–520. [Google Scholar] [CrossRef] [PubMed]
- Lara, P.C.; Pruschy, M.; Zimmermann, M.; Henríquez-Hernández, L.A. MVP and Vaults: A Role in the Radiation Response. Radiat. Oncol. 2011, 6, 148. [Google Scholar] [CrossRef]
- Lee, H.M.; Joh, J.W.; Seo, S.-R.; Kim, W.-T.; Kim, M.K.; Choi, H.S.; Kim, S.Y.; Jang, Y.-J.; Sinn, D.H.; Choi, G.S.; et al. Cell-Surface Major Vault Protein Promotes Cancer Progression through Harboring Mesenchymal and Intermediate Circulating Tumor Cells in Hepatocellular Carcinomas. Sci. Rep. 2017, 7, 13201. [Google Scholar] [CrossRef]
- Balan, S.; Lekshmi, S.; Radha, K.; Sathyan, S.; Vijai, J.; Banerjee, M.; Radhakrishnan, K. Major Vault Protein (MVP) Gene Polymorphisms and Drug Resistance in Mesial Temporal Lobe Epilepsy with Hippocampal Sclerosis. Gene 2013, 526, 449–453. [Google Scholar] [CrossRef]
- Mossink, M.H.; van Zon, A.; Scheper, R.J.; Sonneveld, P.; Wiemer, E.A. Vaults: A Ribonucleoprotein Particle Involved in Drug Resistance? Oncogene 2003, 22, 7458–7467. [Google Scholar] [CrossRef]
- Han, M.; Lv, Q.; Tang, X.-J.; Hu, Y.-L.; Xu, D.-H.; Li, F.-Z.; Liang, W.-Q.; Gao, J.-Q. Overcoming Drug Resistance of MCF-7/ADR Cells by Altering Intracellular Distribution of Doxorubicin via MVP Knockdown with a Novel siRNA Polyamidoamine-Hyaluronic Acid Complex. J. Controlled Release 2012, 163, 136–144. [Google Scholar] [CrossRef]
- Wojtowicz, K.; Januchowski, R.; Nowicki, M.; Zabel, M. vPARP Adjusts MVP Expression in Drug-Resistant Cell Lines in Conjunction with MDR Proteins. Anticancer Res. 2017, 37. [Google Scholar] [CrossRef]
- Jin, D.-H.; Kim, S.; Kim, D.-H.; Park, J. Two Genetic Variants in Telomerase-Associated Protein 1 Are Associated with Stomach Cancer Risk. J. Hum. Genet. 2016, 61, 885–889. [Google Scholar] [CrossRef] [PubMed]
- Kickhoefer, V.A.; Liu, Y.; Kong, L.B.; Snow, B.E.; Stewart, P.L.; Harrington, L.; Rome, L.H. The Telomerase/Vault-Associated Protein Tep1 Is Required for Vault RNA Stability and Its Association with the Vault Particle. J. Cell Biol. 2001, 152, 157–164. [Google Scholar] [CrossRef] [PubMed]
- Ryu, S.J.; An, H.J.; Oh, Y.S.; Choi, H.R.; Ha, M.K.; Park, S.C. On the Role of Major Vault Protein in the Resistance of Senescent Human Diploid Fibroblasts to Apoptosis. Cell Death Differ. 2008, 15, 1673–1680. [Google Scholar] [CrossRef]
- Dong, X.; Akuetteh, P.D.P.; Song, J.; Ni, C.; Jin, C.; Li, H.; Jiang, W.; Si, Y.; Zhang, X.; Zhang, Q.; et al. Major Vault Protein (MVP) Associated With BRAFV600E Mutation Is an Immune Microenvironment-Related Biomarker Promoting the Progression of Papillary Thyroid Cancer via MAPK/ERK and PI3K/AKT Pathways. Front. Cell Dev. Biol. 2022, 9, 688370. [Google Scholar] [CrossRef]
- Silva, P.; West, C.M.; Slevin, N.; Valentine, H.; Ryder, W.D.J.; Hampson, L.; Bibi, R.; Sloan, P.; Thakker, N.; Homer, J.; et al. Tumor Epression of Major Vault Protein Is an Adverse Prognostic Factor for Radiotherapy Outcome in Oropharyngeal Carcinoma. Int. J. Radiat. Oncol. 2007, 69, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Ramberg, H.; Richardsen, E.; De Souza, G.A.; Rakaee, M.; Stensland, M.E.; Braadland, P.R.; Nygård, S.; Ögren, O.; Guldvik, I.J.; Berge, V.; et al. Proteomic Analyses Identify Major Vault Protein as a Prognostic Biomarker for Fatal Prostate Cancer. Carcinogenesis 2021, 42, 685–693. [Google Scholar] [CrossRef]
- Gallo, S.; Kong, E.; Ferro, I.; Polacek, N. Small but Powerful: The Human Vault RNAs as Multifaceted Modulators of Pro-Survival Characteristics and Tumorigenesis. Cancers 2022, 14, 2787. [Google Scholar] [CrossRef]
- Nandy, C.; Mrázek, J.; Stoiber, H.; Grässer, F.A.; Hüttenhofer, A.; Polacek, N. Epstein–Barr Virus-Induced Expression of a Novel Human Vault RNA. J. Mol. Biol. 2009, 388, 776–784. [Google Scholar] [CrossRef]
- Mrazek, J.; Kreutmayer, S.B.; Grasser, F.A.; Polacek, N.; Huttenhofer, A. Subtractive Hybridization Identifies Novel Differentially Expressed ncRNA Species in EBV-Infected Human B Cells. Nucleic Acids Res. 2007, 35, e73–e73. [Google Scholar] [CrossRef]
- Bracher, L.; Ferro, I.; Pulido-Quetglas, C.; Ruepp, M.-D.; Johnson, R.; Polacek, N. Human vtRNA1-1 Levels Modulate Signaling Pathways and Regulate Apoptosis in Human Cancer Cells. Biomolecules 2020, 10, 614. [Google Scholar] [CrossRef] [PubMed]
- Stadler, P.F.; Chen, J.J.-L.; Hackermuller, J.; Hoffmann, S.; Horn, F.; Khaitovich, P.; Kretzschmar, A.K.; Mosig, A.; Prohaska, S.J.; Qi, X.; et al. Evolution of Vault RNAs. Mol. Biol. Evol. 2009, 26, 1975–1991. [Google Scholar] [CrossRef] [PubMed]
- Gopinath, S.C.B.; Matsugami, A.; Katahira, M.; Kumar, P.K.R. Human Vault-Associated Non-Coding RNAs Bind to Mitoxantrone, a Chemotherapeutic Compound. Nucleic Acids Res. 2005, 33, 4874–4881. [Google Scholar] [CrossRef] [PubMed]
- Horos, R.; Büscher, M.; Kleinendorst, R.; Alleaume, A.-M.; Tarafder, A.K.; Schwarzl, T.; Dziuba, D.; Tischer, C.; Zielonka, E.M.; Adak, A.; et al. The Small Non-Coding Vault RNA1-1 Acts as a Riboregulator of Autophagy. Cell 2019, 176, 1054–1067e12. [Google Scholar] [CrossRef] [PubMed]
- Büscher, M.; Horos, R.; Huppertz, I.; Haubrich, K.; Dobrev, N.; Baudin, F.; Hennig, J.; Hentze, M.W. Vault RNA1–1 Riboregulates the Autophagic Function of P62 by Binding to Lysine 7 and Arginine 21, Both of Which Are Critical for P62 Oligomerization. RNA 2022, 28, 742–755. [Google Scholar] [CrossRef]
- Chen, J.; OuYang, H.; An, X.; Liu, S. Vault RNAs Partially Induces Drug Resistance of Human Tumor Cells MCF-7 by Binding to the RNA/DNA-Binding Protein PSF and Inducing Oncogene GAGE6. PLOS ONE 2018, 13, e0191325. [Google Scholar] [CrossRef] [PubMed]
- Hussain, S.; Sajini, A.A.; Blanco, S.; Dietmann, S.; Lombard, P.; Sugimoto, Y.; Paramor, M.; Gleeson, J.G.; Odom, D.T.; Ule, J.; et al. NSun2-Mediated Cytosine-5 Methylation of Vault Noncoding RNA Determines Its Processing into Regulatory Small RNAs. Cell Rep. 2013, 4, 255–261. [Google Scholar] [CrossRef]
- Persson, H.; Kvist, A.; Vallon-Christersson, J.; Medstrand, P.; Borg, Å.; Rovira, C. The Non-Coding RNA of the Multidrug Resistance-Linked Vault Particle Encodes Multiple Regulatory Small RNAs. Nat. Cell Biol. 2009, 11, 1268–1271. [Google Scholar] [CrossRef]
- Kickhoefer, V.A.; Searles, R.P.; Kedersha, N.L.; Garber, M.E.; Johnson, D.L.; Rome, L.H. Vault Ribonucleoprotein Particles from Rat and Bullfrog Contain a Related Small RNA That Is Transcribed by RNA Polymerase III. J. Biol. Chem. 1993, 268, 7868–7873. [Google Scholar] [CrossRef]
- Schramm, L.; Hernandez, N. Recruitment of RNA Polymerase III to Its Target Promoters. Genes Dev. 2002, 16, 2593–2620. [Google Scholar] [CrossRef]
- Bergeron, D.; Faucher-Giguère, L.; Emmerichs, A.-K.; Choquet, K.; Song, K.S.; Deschamps-Francoeur, G.; Fafard-Couture, É.; Rivera, A.; Couture, S.; Churchman, L.S.; et al. Intronic Small Nucleolar RNAs Regulate Host Gene Splicing through Base Pairing with Their Adjacent Intronic Sequences. Genome Biol. 2023, 24, 160. [Google Scholar] [CrossRef]
- Perreault, J.; Perreault, J.-P.; Boire, G. Ro-Associated Y RNAs in Metazoans: Evolution and Diversification. Mol. Biol. Evol. 2007, 24, 1678–1689. [Google Scholar] [CrossRef] [PubMed]
- Vabret, N.; Najburg, V.; Solovyov, A.; Gopal, R.; McClain, C.; Šulc, P.; Balan, S.; Rahou, Y.; Beauclair, G.; Chazal, M.; et al. Y RNAs Are Conserved Endogenous RIG-I Ligands across RNA Virus Infection and Are Targeted by HIV-1. iScience 2022, 25, 104599. [Google Scholar] [CrossRef] [PubMed]
- Driedonks, T.A.P.; Nolte-’t Hoen, E.N.M. Circulating Y-RNAs in Extracellular Vesicles and Ribonucleoprotein Complexes; Implications for the Immune System. Front. Immunol. 2019, 9, 3164. [Google Scholar] [CrossRef] [PubMed]
- Dhahbi, J.M.; Spindler, S.R.; Atamna, H.; Boffelli, D.; Martin, D.I.K. Deep Sequencing of Serum Small RNAs Identifies Patterns of 5′ tRNA Half and YRNA Fragment Expression Associated with Breast Cancer. Biomark. Cancer 2014, 6, BICS20764. [Google Scholar] [CrossRef] [PubMed]
- Mosig, A.; Guofeng, M.; Stadler, B.M.R.; Stadler, P.F. Evolution of the Vertebrate Y RNA Cluster. Theory Biosci. 2007, 126, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Alagia, A.; Tereňová, J.; Ketley, R.F.; Di Fazio, A.; Chelysheva, I.; Gullerova, M. Small Vault RNA1-2 Modulates Expression of Cell Membrane Proteins through Nascent RNA Silencing. Life Sci. Alliance 2023, 6, e202302054. [Google Scholar] [CrossRef]
- Mattick, J.S.; Makunin, I.V. Non-Coding RNA. Hum. Mol. Genet. 2006, 15, R17–R29. [Google Scholar] [CrossRef]
- Mattick, J.S.; Amaral, P.P.; Carninci, P.; Carpenter, S.; Chang, H.Y.; Chen, L.-L.; Chen, R.; Dean, C.; Dinger, M.E.; Fitzgerald, K.A.; et al. Long Non-Coding RNAs: Definitions, Functions, Challenges and Recommendations. Nat. Rev. Mol. Cell Biol. 2023, 24, 430–447. [Google Scholar] [CrossRef]
- Kickhoefer, V.A.; Poderycki, M.J.; Chan, E.K.L.; Rome, L.H. The La RNA-Binding Protein Interacts with the Vault RNA and Is a Vault-Associated Protein. J. Biol. Chem. 2002, 277, 41282–41286. [Google Scholar] [CrossRef]
- Kickhoefer, V.A.; Rajavel, K.S.; Scheffer, G.L.; Dalton, W.S.; Scheper, R.J.; Rome, L.H. Vaults Are Up-Regulated in Multidrug-Resistant Cancer Cell Lines. J. Biol. Chem. 1998, 273, 8971–8974. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. The Hallmarks of Cancer. Cell 2000, 100, 57–70. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. Hallmarks of Cancer: The Next Generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D. Hallmarks of Cancer: New Dimensions. Cancer Discov. 2022, 12, 31–46. [Google Scholar] [CrossRef] [PubMed]
- Ferro, I.; Gavini, J.; Gallo, S.; Bracher, L.; Landolfo, M.; Candinas, D.; Stroka, D.M.; Polacek, N. The Human Vault RNA Enhances Tumorigenesis and Chemoresistance through the Lysosome in Hepatocellular Carcinoma. Autophagy 2022, 18, 191–203. [Google Scholar] [CrossRef] [PubMed]
- Amort, M.; Nachbauer, B.; Tuzlak, S.; Kieser, A.; Schepers, A.; Villunger, A.; Polacek, N. Expression of the Vault RNA Protects Cells from Undergoing Apoptosis. Nat. Commun. 2015, 6, 7030. [Google Scholar] [CrossRef] [PubMed]
- Schiliro, C.; Firestein, B.L. Mechanisms of Metabolic Reprogramming in Cancer Cells Supporting Enhanced Growth and Proliferation. Cells 2021, 10, 1056. [Google Scholar] [CrossRef]
- Kato, H.; Hashimoto, Y.; Hatayama, Y.; Shimohiro, H.; Motokura, T. Serum Levels of Vault RNA Significantly Varied in Patients with Haematological Malignancies. Mol. Med. Rep. 2023, 28, 190. [Google Scholar] [CrossRef]
- Fort, R.S.; Garat, B.; Sotelo-Silveira, J.R.; Duhagon, M.A. vtRNA2-1/Nc886 Produces a Small RNA That Contributes to Its Tumor Suppression Action through the microRNA Pathway in Prostate Cancer. Non-Coding RNA 2020, 6, 7. [Google Scholar] [CrossRef]
- McDonald, A.C.; Vira, M.; Walter, V.; Shen, J.; Raman, J.D.; Sanda, M.G.; Patil, D.; Taioli, E. Circulating microRNAs in Plasma among Men with Low-grade and High-grade Prostate Cancer at Prostate Biopsy. The Prostate 2019, 79, 961–968. [Google Scholar] [CrossRef]
- Kunkeaw, N.; Lee, Y.-S.; Im, W.R.; Jang, J.J.; Song, M.-J.; Yang, B.; Park, J.-L.; Kim, S.-Y.; Ku, Y.; Kim, Y.; et al. Mechanism Mediated by a Noncoding RNA, Nc886, in the Cytotoxicity of a DNA-Reactive Compound. Proc. Natl. Acad. Sci. 2019, 116, 8289–8294. [Google Scholar] [CrossRef] [PubMed]
- Ahn, J.-H.; Lee, H.-S.; Lee, J.-S.; Lee, Y.-S.; Park, J.-L.; Kim, S.-Y.; Hwang, J.-A.; Kunkeaw, N.; Jung, S.Y.; Kim, T.J.; et al. Nc886 Is Induced by TGF-β and Suppresses the microRNA Pathway in Ovarian Cancer. Nat. Commun. 2018, 9, 1166. [Google Scholar] [CrossRef]
- Lee, K.; Kunkeaw, N.; Jeon, S.H.; Lee, I.; Johnson, B.H.; Kang, G.-Y.; Bang, J.Y.; Park, H.S.; Leelayuwat, C.; Lee, Y.S. Precursor miR-886, a Novel Noncoding RNA Repressed in Cancer, Associates with PKR and Modulates Its Activity. RNA 2011, 17, 1076–1089. [Google Scholar] [CrossRef]
- Izquierdo, M.A.; Scheffer, G.L.; Schroeijers, A.B.; De Jong, M.C.; Scheper, R.J. Vault-Related Resistance to Anticancer Drugs Determined by the Expression of the Major Vault Protein LRP. Cytotechnology 1998, 27, 137–148. [Google Scholar] [CrossRef]
- Jeon, S.H.; Lee, K.; Lee, K.S.; Kunkeaw, N.; Johnson, B.H.; Holthauzen, L.M.F.; Gong, B.; Leelayuwat, C.; Lee, Y.S. Characterization of the Direct Physical Interaction of Nc886, a Cellular Non-Coding RNA, and PKR. FEBS Lett. 2012, 586, 3477–3484. [Google Scholar] [CrossRef] [PubMed]
- Dai, W.; Liu, H. MicroRNA-886 Suppresses Osteosarcoma Cell Proliferation and Its Maturation Is Suppressed by Long Non-Coding RNA OXCT1-AS1. Bioengineered 2022, 13, 5769–5778. [Google Scholar] [CrossRef] [PubMed]
- Weinberg, R.A. How Cancer Arises. Sci. Am. 1996, 275, 62–70. [Google Scholar] [CrossRef]
- Zhong, C.; Yu, Q.; Peng, Y.; Zhou, S.; Liu, Z.; Deng, Y.; Guo, L.; Zhao, S.; Chen, G. Novel LncRNA OXCT1-AS1 Indicates Poor Prognosis and Contributes to Tumorigenesis by Regulating miR-195/CDC25A Axis in Glioblastoma. J. Exp. Clin. Cancer Res. 2021, 40, 123. [Google Scholar] [CrossRef]
- Li, B.; Zhu, L.; Li, L.; Ma, R. lncRNA OXCT1-AS1 Promotes Metastasis in Non-Small-Cell Lung Cancer by Stabilizing LEF1, In Vitro and In Vivo. BioMed Res. Int. 2021, 2021, 1–15. [Google Scholar] [CrossRef]
- Kärkkäinen, E.; Heikkinen, S.; Tengström, M.; Kosma, V.-M.; Mannermaa, A.; Hartikainen, J.M. Expression Profiles of Small Non-Coding RNAs in Breast Cancer Tumors Characterize Clinicopathological Features and Show Prognostic and Predictive Potential. Sci. Rep. 2022, 12, 22614. [Google Scholar] [CrossRef]
- Bornstein, S.; Shapiro, I.; Mazumdar, A.; Zitzmann, K.; Nölting, S.; Luca, E.; Beuschlein, F.; Sharma, A.; Hantel, C. The Vault Complex Is Significantly Involved in Therapeutic Responsiveness of Endocrine Tumors and Linked to Autophagy under Chemotherapeutic Conditions. Cancers 2023, 15, 1783. [Google Scholar] [CrossRef] [PubMed]
- Treppendahl, M.B.; Qiu, X.; Søgaard, A.; Yang, X.; Nandrup-Bus, C.; Hother, C.; Andersen, M.K.; Kjeldsen, L.; Möllgaard, L.; Hellström-Lindberg, E.; et al. Allelic Methylation Levels of the Noncoding VTRNA2-1 Located on Chromosome 5q31. 1 Predict Outcome in AML. Blood 2012, 119, 206–216. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.-C.; Lee, C.-W.; Lin, C.-H.; Wu, C.-H.; Lee, Y.-S.; Tsai, C.-L.; Tsai, C.-N. Differential Hypermethylation of the VTRNA2-1 Promoter in Hepatocellular Carcinoma as a Prognostic Factor: Tumor Marker Prognostic Study. Int. J. Surg. 2020, 79, 282–289. [Google Scholar] [CrossRef] [PubMed]
- Dugué, P.-A.; Yu, C.; McKay, T.; Wong, E.M.; Joo, J.E.; Tsimiklis, H.; Hammet, F.; Mahmoodi, M.; Theys, D. ; kConFab; et al. VTRNA2-1: Genetic Variation, Heritable Methylation and Disease Association. Int. J. Mol. Sci. 2021, 22, 2535. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Wang, M.; Zhou, Y.; Yang, J.; Wang, L.; Yang, R.; Wen, M.; Kong, L. Noncoding RNA 886 Alleviates Tumor Cellular Immunological Rejection in Host C57BL/C Mice. Cancer Med. 2020, 9, 5258–5271. [Google Scholar] [CrossRef]
- Fort, R.S.; Mathó, C.; Geraldo, M.V.; Ottati, M.C.; Yamashita, A.S.; Saito, K.C.; Leite, K.R.M.; Méndez, M.; Maedo, N.; Méndez, L.; et al. Nc886 Is Epigenetically Repressed in Prostate Cancer and Acts as a Tumor Suppressor through the Inhibition of Cell Growth. BMC Cancer 2018, 18, 127. [Google Scholar] [CrossRef] [PubMed]
- Kunkeaw, N.; Jeon, S.H.; Lee, K.; Johnson, B.H.; Tanasanvimon, S.; Javle, M.; Pairojkul, C.; Chamgramol, Y.; Wongfieng, W.; Gong, B.; et al. Cell Death/Proliferation Roles for Nc886, a Non-Coding RNA, in the Protein Kinase R Pathway in Cholangiocarcinoma. Oncogene 2013, 32, 3722–3731. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.-S.; Shin, S.; Cho, E.; Im, W.K.; Jeon, S.H.; Kim, Y.; Park, D.; Fréchet, M.; Chajra, H.; Jung, E. Nc886, a Non-Coding RNA, Inhibits UVB-Induced MMP-9 and COX-2 Expression via the PKR Pathway in Human Keratinocytes. Biochem. Biophys. Res. Commun. 2019, 512, 647–652. [Google Scholar] [CrossRef]
- Lee, K.-S.; Park, J.-L.; Lee, K.; Richardson, L.E.; Johnson, B.H.; Lee, H.-S.; Lee, J.-S.; Kim, S.-B.; Kwon, O.-H.; Song, K.S.; et al. Nc886, a Non-Coding RNA of Anti-Proliferative Role, Is Suppressed by CpG DNA Methylation in Human Gastric Cancer. Oncotarget 2014, 5, 3944–3955. [Google Scholar] [CrossRef]
- Fort, R.S.; Duhagon, M.A. Pan-Cancer Chromatin Analysis of the Human vtRNA Genes Uncovers Their Association with Cancer Biology. F1000Research 2021, 10, 182. [Google Scholar] [CrossRef]
- Hu, Z.; Zhang, H.; Tang, L.; Lou, M.; Geng, Y. Silencing Nc886, a Non-Coding RNA, Induces Apoptosis of Human Endometrial Cancer Cells-1A In Vitro. Med. Sci. Monit. 2017, 23, 1317–1324. [Google Scholar] [CrossRef]
- Lei, J.; Xiao, J.-H.; Zhang, S.-H.; Liu, Z.-Q.; Huang, K.; Luo, Z.-P.; Xiao, X.-L.; Hong, Z.-D. Non-Coding RNA 886 Promotes Renal Cell Carcinoma Growth and Metastasis through the Janus Kinase 2/Signal Transducer and Activator of Transcription 3 Signaling Pathway. Mol. Med. Rep. 2017, 16, 4273–4278. [Google Scholar] [CrossRef] [PubMed]
- Pan, B.; Yu, J.; Liu, X. Upregulation of miR-886 Indicates Poor Prognosis and Promotes Tumour Progression of Prostate Cancer. Andrologia 2022, 54. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA. Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Kong, L.; Hao, Q.; Wang, Y.; Zhou, P.; Zou, B.; Zhang, Y. Regulation of P53 Expression and Apoptosis by Vault RNA2-1-5p in Cervical Cancer Cells. Oncotarget 2015, 6, 28371–28388. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, X.; Han, S.; Wang, Y.; Liu, R.; Meng, F.; Su, Z.; Huo, F. Suppression of Mir-886-3P Mediated by Arecoline (Are) Contributes to the Progression of Oral Squamous Cell Carcinoma. J. Investig. Med. 2021, 69, 377–381. [Google Scholar] [CrossRef] [PubMed]
- Helbo, A.; Treppendahl, M.; Aslan, D.; Dimopoulos, K.; Nandrup-Bus, C.; Holm, M.; Andersen, M.; Liang, G.; Kristensen, L.; Grønbæk, K. Hypermethylation of the VTRNA1-3 Promoter Is Associated with Poor Outcome in Lower Risk Myelodysplastic Syndrome Patients. Genes 2015, 6, 977–990. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Ye, X.; Zhao, T. The Physiological Roles of Autophagy in the Mammalian Life Cycle: Autophagy in the Mammal Life Cycle. Biol. Rev. 2019, 94, 503–516. [Google Scholar] [CrossRef]
- Klionsky, D.J.; Petroni, G.; Amaravadi, R.K.; Baehrecke, E.H.; Ballabio, A.; Boya, P.; Bravo-San Pedro, J.M.; Cadwell, K.; Cecconi, F.; Choi, A.M.K.; et al. Autophagy in Major Human Diseases. EMBO J. 2021, 40. [Google Scholar] [CrossRef]
- Deretic, V.; Saitoh, T.; Akira, S. Autophagy in Infection, Inflammation and Immunity. Nat. Rev. Immunol. 2013, 13, 722–737. [Google Scholar] [CrossRef]
- Büscher, M.; Horos, R.; Hentze, M.W. ‘High Vault-Age’: Non-Coding RNA Control of Autophagy. Open Biol. 2020, 10, 190307. [Google Scholar] [CrossRef] [PubMed]
- Ferreira-Marques, M.; Carvalho, A.; Cavadas, C.; Aveleira, C.A. PI3K/AKT/MTOR and ERK1/2-MAPK Signaling Pathways Are Involved in Autophagy Stimulation Induced by Caloric Restriction or Caloric Restriction Mimetics in Cortical Neurons. Aging 2021, 13, 7872–7882. [Google Scholar] [CrossRef]
- Cao, Z.; Liao, Q.; Su, M.; Huang, K.; Jin, J.; Cao, D. AKT and ERK Dual Inhibitors: The Way Forward? Cancer Lett. 2019, 459, 30–40. [Google Scholar] [CrossRef] [PubMed]
- Moscat, J.; Diaz-Meco, M.T. P62 at the Crossroads of Autophagy, Apoptosis, and Cancer. Cell 2009, 137, 1001–1004. [Google Scholar] [CrossRef] [PubMed]
- Morana, O.; Wood, W.; Gregory, C.D. The Apoptosis Paradox in Cancer. Int. J. Mol. Sci. 2022, 23, 1328. [Google Scholar] [CrossRef] [PubMed]
- Kieser, A.; Sterz, K.R. The Latent Membrane Protein 1 (LMP1). In Epstein Barr Virus Volume 2; Münz, C., Ed.; Current Topics in Microbiology and Immunology; Springer International Publishing: Cham, 2015; ISBN 978-3-319-22833-4. [Google Scholar]
- Emran, T.B.; Shahriar, A.; Mahmud, A.R.; Rahman, T.; Abir, M.H. ; Siddiquee, Mohd. F.-R.; Ahmed, H.; Rahman, N.; Nainu, F.; Wahyudin, E.; et al. Multidrug Resistance in Cancer: Understanding Molecular Mechanisms, Immunoprevention and Therapeutic Approaches. Front. Oncol. 2022, 12, 891652. [Google Scholar] [CrossRef]
- Steiner, E.; Holzmann, K.; Elbling, L.; Micksche, M.; Berger, W. Cellular Functions of Vaults and Their Involvement in Multidrug Resistance. Curr. Drug Targets 2006, 7, 923–934. [Google Scholar] [CrossRef]
- Geisslinger, F.; Müller, M.; Vollmar, A.M.; Bartel, K. Targeting Lysosomes in Cancer as Promising Strategy to Overcome Chemoresistance—A Mini Review. Front. Oncol. 2020, 10, 1156. [Google Scholar] [CrossRef]
- Hong, H.; Wang, Q.; Li, J.; Liu, H.; Meng, X.; Zhang, H. Aging, Cancer and Immunity. J. Cancer 2019, 10, 3021–3027. [Google Scholar] [CrossRef]
- Li, S.; Min, J.-Y.; Krug, R.M.; Sen, G.C. Binding of the Influenza A Virus NS1 Protein to PKR Mediates the Inhibition of Its Activation by Either PACT or Double-Stranded RNA. Virology 2006, 349, 13–21. [Google Scholar] [CrossRef]
- Li, F.; Chen, Y.; Zhang, Z.; Ouyang, J.; Wang, Y.; Yan, R.; Huang, S.; Gao, G.F.; Guo, G.; Chen, J.-L. Robust Expression of Vault RNAs Induced by Influenza A Virus Plays a Critical Role in Suppression of PKR-Mediated Innate Immunity. Nucleic Acids Res. 2015, gkv1078. [Google Scholar] [CrossRef]
- Kim, Y.; Ji, H.; Cho, E.; Park, N.-H.; Hwang, K.; Park, W.; Lee, K.-S.; Park, D.; Jung, E. Nc886, a Non-Coding RNA, Is a New Biomarker and Epigenetic Mediator of Cellular Senescence in Fibroblasts. Int. J. Mol. Sci. 2021, 22, 13673. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.-S.; Cho, E.; Weon, J.B.; Park, D.; Fréchet, M.; Chajra, H.; Jung, E. Inhibition of UVB-Induced Inflammation by Laminaria Japonica Extract via Regulation of Nc886-PKR Pathway. Nutrients 2020, 12, 1958. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.-S.; Bao, X.; Lee, H.-H.; Jang, J.J.; Saruuldalai, E.; Park, G.; Im, W.R.; Park, J.-L.; Kim, S.-Y.; Shin, S.; et al. Nc886, a Novel Suppressor of the Type I Interferon Response Upon Pathogen Intrusion. Int. J. Mol. Sci. 2021, 22, 2003. [Google Scholar] [CrossRef] [PubMed]
- Zheng, D.; Liwinski, T.; Elinav, E. Interaction between Microbiota and Immunity in Health and Disease. Cell Res. 2020, 30, 492–506. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Xiao, L.; Wen, S.J.; Yu, T.; Sharma, S.; Chung, H.K.; Warner, B.; Mallard, C.G.; Rao, J.N.; Gorospe, M.; et al. Small Noncoding Vault RNA2 -1 Disrupts Gut Epithelial Barrier Function via Interaction with HUR. EMBO Rep. 2023, 24, e54925. [Google Scholar] [CrossRef] [PubMed]
- Wakatsuki, S.; Araki, T. Novel Molecular Basis for Synapse Formation: Small Non-Coding Vault RNA Functions as a Riboregulator of MEK1 to Modulate Synaptogenesis. Front. Mol. Neurosci. 2021, 14, 748721. [Google Scholar] [CrossRef]
- Wakatsuki, S.; Takahashi, Y.; Shibata, M.; Adachi, N.; Numakawa, T.; Kunugi, H.; Araki, T. Small Noncoding Vault RNA Modulates Synapse Formation by Amplifying MAPK Signaling. J. Cell Biol. 2021, 220, e201911078. [Google Scholar] [CrossRef]
- Wakatsuki, S.; Ohno, M.; Araki, T. Human Vault RNA1-1, but Not Vault RNA2-1, Modulates Synaptogenesis. Commun. Integr. Biol. 2021, 14, 61–65. [Google Scholar] [CrossRef]
- Geoffray, M.-M.; Falissard, B.; Green, J.; Kerr, B.; Evans, D.G.; Huson, S.; Burkitt-Wright, E.; Garg, S. Autism Spectrum Disorder Symptom Profile Across the RASopathies. Front. Psychiatry 2021, 11, 585700. [Google Scholar] [CrossRef]
- Borrie, S.C.; Plasschaert, E.; Callaerts-Vegh, Z.; Yoshimura, A.; D’Hooge, R.; Elgersma, Y.; Kushner, S.A.; Legius, E.; Brems, H. MEK Inhibition Ameliorates Social Behavior Phenotypes in a Spred1 Knockout Mouse Model for RASopathy Disorders. Mol. Autism 2021, 12, 53. [Google Scholar] [CrossRef]
- Miñones-Moyano, E.; Friedländer, M.R.; Pallares, J.; Kagerbauer, B.; Porta, S.; Escaramís, G.; Ferrer, I.; Estivill, X.; Martí, E. Upregulation of a Small Vault RNA (svtRNA2-1a) Is an Early Event in Parkinson Disease and Induces Neuronal Dysfunction. RNA Biol. 2013, 10, 1093–1106. [Google Scholar] [CrossRef]
Figure 1.
The biological potential of vault RNA1-1 in the modulation of cell metabolism related to tumorigenesis and drug resistance. vtRNA1-1 is the most well-known among four vtRNA paralogues in humans, and the vtRNA1-1 gene shows the highest expression level in the group of vtRNA coding genes. vtRNA1-1 is involved in the regulation of crucial cellular processes associated with homeostasis. It can regulate apoptosis and autophagy, processes necessary to maintain the correct number of properly functioning cells. Moreover, this vault RNA is tightly associated with tumorigenesis, cell proliferation, and tumor ability to metastasize. Vault RNA1-1 is also considered a potential target in cancer therapy since its role in cell proliferation and drug resistance was demonstrated. vtRNA1-1 showed a pro-survival mode of action, stimulating tumor cell proliferation. Overexpression of vtRNA1-1 led to autophagy, apoptosis inhibition, and drug resistance induction. Arrows indicate induction (↑) or inhibition (↓) of a specific mechanism/pathway. .
Figure 1.
The biological potential of vault RNA1-1 in the modulation of cell metabolism related to tumorigenesis and drug resistance. vtRNA1-1 is the most well-known among four vtRNA paralogues in humans, and the vtRNA1-1 gene shows the highest expression level in the group of vtRNA coding genes. vtRNA1-1 is involved in the regulation of crucial cellular processes associated with homeostasis. It can regulate apoptosis and autophagy, processes necessary to maintain the correct number of properly functioning cells. Moreover, this vault RNA is tightly associated with tumorigenesis, cell proliferation, and tumor ability to metastasize. Vault RNA1-1 is also considered a potential target in cancer therapy since its role in cell proliferation and drug resistance was demonstrated. vtRNA1-1 showed a pro-survival mode of action, stimulating tumor cell proliferation. Overexpression of vtRNA1-1 led to autophagy, apoptosis inhibition, and drug resistance induction. Arrows indicate induction (↑) or inhibition (↓) of a specific mechanism/pathway. .
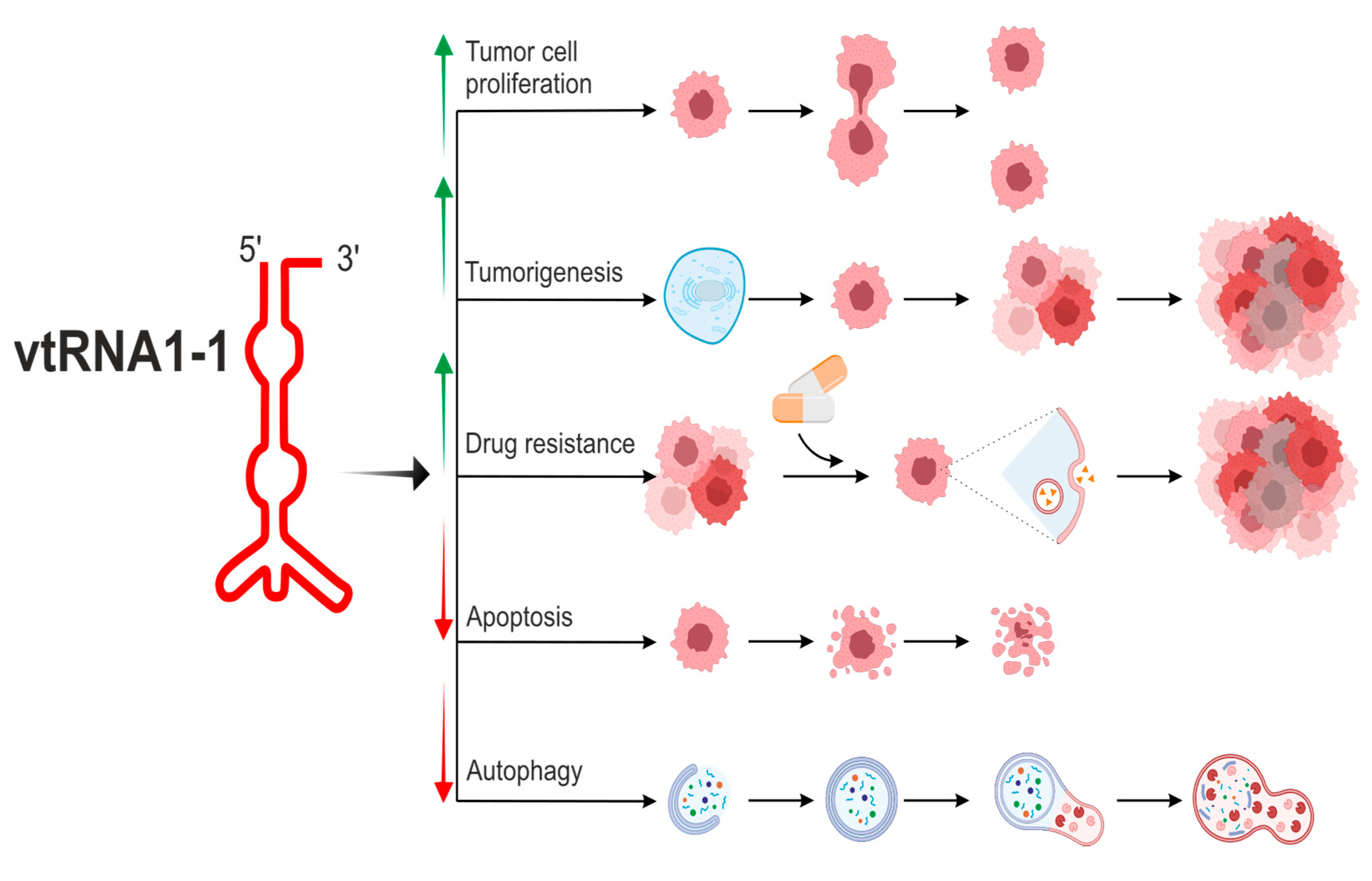
Figure 2.
The potential role of vtRNA2-1 in aging, senescence and immune response. vtRNA2-1 expression may be controlled via various genetic and epigenetic mechanisms. This tight control is crucial for the role of vtRNA2-1 in aging, and this molecule is a potential anti-aging target. Red arrows indicate up- (↑) or downregulation (↓) of target effect, respectively.
Figure 2.
The potential role of vtRNA2-1 in aging, senescence and immune response. vtRNA2-1 expression may be controlled via various genetic and epigenetic mechanisms. This tight control is crucial for the role of vtRNA2-1 in aging, and this molecule is a potential anti-aging target. Red arrows indicate up- (↑) or downregulation (↓) of target effect, respectively.
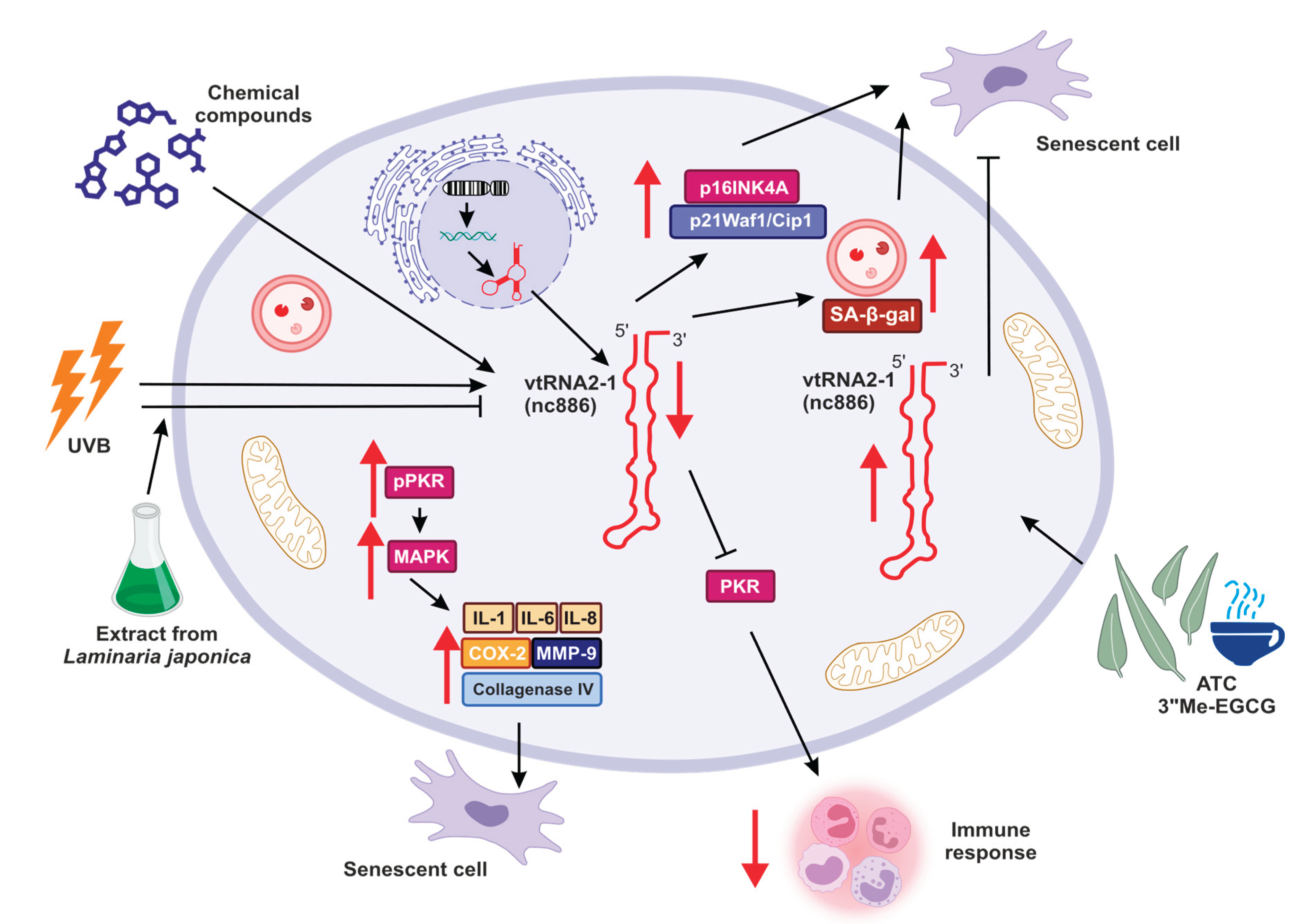

Table 1.
The role of human vault RNAs paralogues in cellular processes.
| vtRNA paralogue | structure | derivatives | role in cellular processes |
|---|---|---|---|
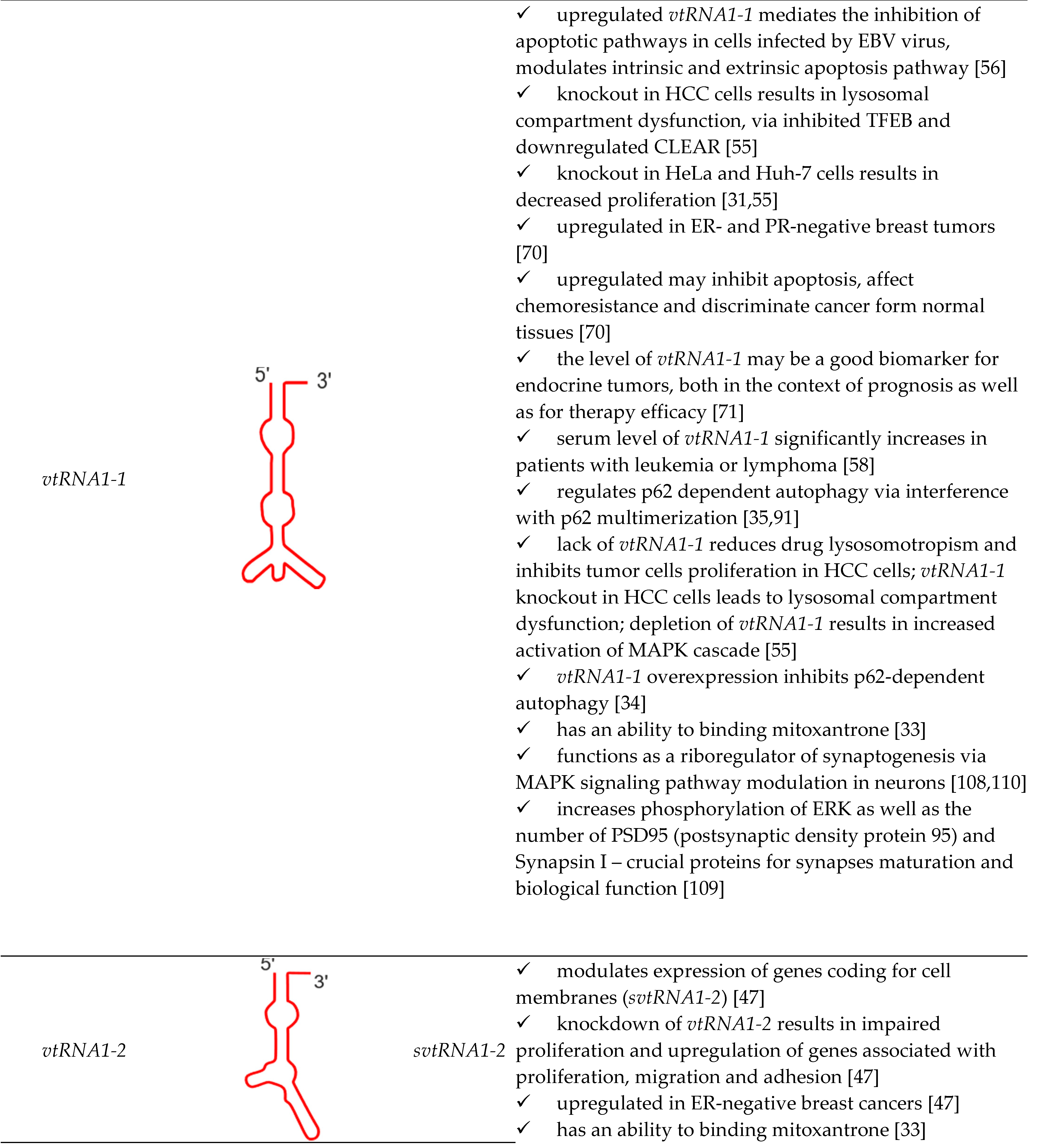 
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



