
Submitted:
12 March 2024
Posted:
14 March 2024
You are already at the latest version
Abstract
Mitochondria are semiautonomous organelles, which are pivotal for the energy metabolism of eukaryotic cells. These organelles contain their own genetic system (mtDNA, mitogenome) that encodes to some structural RNAs and organellar proteins. The biogenesis and function of mitochondria is tightly regulated by nuclear-encoded protein cofactors. The mtDNAs of land plants are characteristically large, with a complex mode of gene organization and expression, in particularly at the post-transcriptional level. To become functional, the primary organellar transcripts undergo extensive maturation steps, which include endo- and exo-nucleolytic cleavage, RNA-base editing, and both ‘cis’- and ‘trans’-splicing events. These essential processing steps rely on the activities of a large set of nuclear-encoded factors. Among these, the RNA helicases serve as key players in organellar (mt-)RNA metabolism. This review summarizes the important roles played by these factors in regulating the highly dynamic processes of transcription, RNA processing, and translation within the mitochondria of land plants. We further discuss recent advancements in understanding how dysregulation of mitochondrial RNA helicases affects the organellar gene expression, respiratory functions, and hence the growth, development and physiology of land plants.
Keywords:
plant
; mitochondria
; RNA helicase
; RNA binding protein
; post-transcription
; RNA metabolism
; splicing
1. Introduction
1.1. The Regulation of Post-Transcriptional RNA Processing in Land Plant Mitochondria
Mitochondria serve as the powerhouse of eukaryotic cells. Accordingly, these organelles play a central role in cellular energy production through the oxidative phosphorylation (OXPHOS) pathway. As descendances of a free-living bacterium, mitochondria contain their own genetic material (mtDNA, or mitogenome), which encodes to tRNAs, rRNAs and some key proteins of the respiratory system. The vast majority of the proteins that reside within the mitochondria are encoded by nuclear gene-loci, and are post-translationally imported into the organelles. These include proteins of the OXPHOS system, as well as essential factors that regulate the mtDNA replication, organellar gene expression, and the assembly of organellar complexes [1,2,3]. The analyses of the genome sequences and transcriptome landscapes of mitochondria provided with key insights into the evolution of mitogenomes in animals, fungi and plants, in addition to the unique properties of mitochondrial RNA (mt-RNA) metabolism, transcriptional activities and protein synthesis in different organisms. Despite all mitochondria having a common bacterial ancestor, they have undergone divergent evolution in different organisms.
The mitochondria in animals typically exhibit a compact genome structure (harboring 37 genes, with 13 protein ORFs), which is mainly transcribed from two initiation sites, resulting in two long polycistronic pre-RNAs, denoted as the light (L) and heavy (H) strands. Post-transcriptional cleavage events generate ‘individual’ mRNA molecules, which are translated into the mitochondrial proteins. Additionally, tRNAs and rRNAs, that are essential for mitochondrial protein synthesis, are also processed from the precursor organellar molecules. The mitogenomes of land plants are notably larger and more complex in structure than their counterparts in Animalia, with a highly sophisticated mode of organellar gene expression in the post-transcriptional level [1]. To become functional RNAs (mature tRNAs, rRNAs and mRNAs), the precursor (pre-) RNAs that are transcribed within the mitochondria in plants must undergo a series of maturation steps. These include the processing of both 5’ and 3’ termini, numerous RNA ‘editing’ events (typically C-to-U deamination reactions in angiosperms’ mitochondria), and the removal of a large number of introns (mostly group II-type) which reside within the coding regions of many essential genes (e.g., respiratory proteins, cytochrome c maturation (CCM) factors, as well as ribosomal subunits). These RNA processing steps are crucial for plant organellar gene expression, and hence for the proper functioning of the respiratory apparatus, ATP synthesis, and for optimal plant growth and development.
Notably, the intricate interplay of different molecular machineries within the mitochondria is pivotal for the biogenesis of the respiratory machinery and, consequently, for maintaining cellular homeostasis and plant physiology. Among these components, RNA binding proteins (RBPs), which are encoded by nuclear gene loci and imported post-translationally into the organelle, stand out as key players in mitochondria gene expression [2,4,5]. Furthermore, the organellar-localized RBPs are also anticipated to serve as a key control mechanism for the regulation of mitochondrial biogenesis, the assembly of the OXPHOS complexes, and for optimal respiratory activities. Here, we focus on the functional and structural aspects of RNA helicases, an ancient group of enzymes, and discuss their significance in modulating mt-RNA splicing, and hence the fidelity of gene expression in land plant’s mitochondria.
2. RNA Helicases as Key Players in Organellar RNA Metabolism
RNA molecules, as their highly versatile proteinaceous counterparts, need to adopt a specific molecular conformation and to interact with protein cofactors in order to function correctly [6,7,8,9,10]. A cellular challenge is that unlike (most) proteins, RNAs can fall into different non-functional conformations that have similar energetically stable states, as resulting from their more chemical simplicity (i.e., combinations of ribonucleotides joined together by phosphodiester bonds) [11]. Specific changes in the RNA conformation can be facilitated by a variety of cellular factors, such as RNA-binding proteins (RBPs), ligands, and metal ions. Additionally, they can be influenced by variations in salt concentration, temperature, or pH [12]. This review focuses on RNA helicases as central regulators in the splicing and processing of group II-type introns in the genomes of plant mitochondria.
RNA helicases are a diverse family of enzymes that play key roles in determining RNA secondary (and tertiary) structures, in an ATP-dependent manner [13]. These also form one of the largest group of enzymes in eukaryotic RNA metabolism [14]. RNA helicases (and RNA helicase-like proteins) are present in the three kingdoms of life, as well as in the genomes of viruses, and are generally classified into six main groups [15]. The majority of the RNA helicases are grouped with the Super Family 2 (SF2) [16], which includes the DEAD, DEAH and DExH protein subfamilies. The core motif of the more common DEAD-box RNA helicases consists of two sequential RecA-like domains, which harbor several (i.e., ≥12) well-conserved amino acid subdomains termed Q, I, II, III, IV, V, and VI [16]. Figure 1 shows a characteristic structure of DEAD-box RNA helicase proteins.
The breaking of hydrogen bonds between ribonucleotides facilitates changes in RNA structures, thereby affecting the stability and function of various RNAs. By unwinding the secondary structures of organellar transcripts, proteinaceous factors, such as RNA helicases, regulate the processing of tRNAs, rRNAs, and mRNAs, control the stability of the RNA ligands, resolve RNA-RNA interactions, and modulate RNA-protein complexes (Figure 2). These activities are pivotal in numerous key processes, such as in transcription, post-transcriptional processing (such as in splicing), RNA decay, or translation (e.g., where the RNA fold may impede ribosome movement) [2,3,17]. Likewise, RNA helicases play key roles in the regulation of RNA metabolism in land plants, and may also link gene expression patterns with developmental or environmental signals [18]. Indeed, several RNA helicases were shown to respond to environmental signals, and were also associated with cellular response to abiotic stresses in prokaryotes [15]. Similarly, changes in temperature (or other environmental factors) can affect the expression and/or activity of the organellar RNA helicases, influencing the overall RNA metabolism within the mitochondria (or plastids) [4]. It is, therefore, not surprising that mutations in genes encoding RNA helicases can lead to notable organellar dysfunctions, highlighting the importance of these enzymes for cellular activities and (plant) physiology [2,3,19].
Accumulating data indicate that RNA helicases often act in multi-proteinaceous complexes, and affect various aspects of RNA metabolism and gene expression, e.g., translation initiation assemblies, or within the RNA splicing complexes [15]. Besides canonical helicase motifs, the RNA helicases' deduced protein sequences often feature additional N- and/or C-terminal domains, primarily comprising protein-protein interaction sites. However, these domains may also confer specific RNA ligand binding capabilities. We elaborate here on the established functions of RNA helicases in the regulation of group II introns splicing in plant mitochondria.
3. The Splicing of Plant Mitochondrial Group II Introns Relies on the Activities of Different Subset Nuclear-Encoded RNA Binding Cofactors
Splicing is a vital processing step in the maturation of some organellar transcripts in the mitochondria of fungi and in plant organelles. This intriguing RNA processing step has arisen as a fundamental process in the regulation of gene expression in the nuclear genomes of eukaryotic cells [20,21,22,23,24]. The splicing reaction entails the removal of intervening RNA sequences (i.e., introns) to allow the joining of the flanking ‘exons’, so as to generate functional RNA (mature, mRNA) transcripts. The introns in nature are divided into several distinct classes, termed as group I, group II, group III and nuclear-spliceosomal introns [25]. Each class represents a unique category of intronic sequences with distinct characteristics and functions. These classes serve as fundamental components of genetic information processing across various organisms, playing crucial roles in gene expression, regulation, and genome evolution. Canonical group I, group II, and group III introns are capable of self-splicing, generally under non-physiological (high temperature and salt) conditions. Group I introns are removed from the pre-RNA with a nucleophilic attack by a free nucleotide, usually a free guanine (G) residue. Group II sequences are spliced with a mechanism identical to the nuclear spliceosome, i.e., two transesterification reactions and the excision of the intron as a lariat loop. Following transcription, the group II RNAs can fold into a secondary and tertiary structure consisting of several domains. Group I RNAs fold into nine paired regions (i.e., P1-P9), which are organized into 2 main domains. Group II RNAs fold into a six-domain secondary structure (DI-DVI). Both group I and group II introns harbor an open reading frame (Intron Encoded Protein, IEP or maturase) which specifically binds to its own cognate intron and assist with its splicing under physiological conditions, in vivo. In group II introns, the maturases/IEPs are encoded within the fourth domain (DIV).
The organellar introns in plants, fungi and some protists, belong mainly to group I and group II introns [26,27,28,29,30,31,32]. Group II introns are predominantly prevalent in the mtDNAs of vascular plants [33]. One hypothesis suggests that the spliceosome, along with the intron-exon structure of genes, originated from ancient catalytic group II-type ribozymes. According to this hypothesis, the group II intron ribozymes, along with their maturase (intron-encoded) factors, have invaded the eukaryotic genomes during the endosymbiosis of an α-proteobacterium, which ultimately led to the establishment of mitochondria [20,21,22,23,24]. The catalytic and mobile intron-RNAs then ‘infected’ and spread through the eukaryotic genome, gradually degenerated, such as they lost their self-splicing activities and instead acquired protein cofactors to assist with their splicing in vivo. The nuclear introns and their cognate splicing cofactors later became part of the general splicing machinery, known as the spliceosome. Yet why splicing has expanded in plant mitochondria, while it has been lost in the organelles of mammals (and most other animals), remains an intriguing subject in evolutionary biology [4,21,22,26,34].
While the splicing of nuclear introns is a well-studied process, the cellular mechanisms and factors that enable the excision of the plant mitochondrial group II introns are much less understood [2,21,23,33]. Genetic and biochemical analysis led to the characterization of many proteinaceous cofactors that function in the processing of mitochondrial group II introns in fungi and plants [3,19,35,36]. The majority of the organellar splicing factors are encoded in the nucleus, thus acting as mediators of communication between the nucleus and the organelles [21]. These include a few proteins are related to intron-encoded maturases (MATs, Pfam-01348; [37]) that have translocated from the mitochondria into the nucleus [21,38], while the majority of the organellar splicing factors include proteins which harbor RNA binding modules, such as the pentatricopeptide motif (PPR, Pfam-13812; [39]), the mitochondrial transcription termination factor domain, (mTERFs, Pfam-02536; [40,41,42]), the plant organelle RNA recognition motif (PORR, Pfam-11955), as well as the ubiquitous DExD/H box RNA helicases (Pfam-00270; [3,19,35]) (Figure 1) that are highly conserved in nature and participate in nearly all aspects of RNA metabolism [14]. Table 1 summarizes a list of various DEAD-box RNA helicases found in the organelles of yeast and Arabidopsis, some of which were shown to act in the splicing of group II introns found in the mitogenomes of fungi and land plants.
4. Mitochondrial RNA Helicases Factors in Saccharomyces cerevisiae (Yeast)
RNA helicases play pivotal roles in mtRNA turnover, splicing and translation, as well as in the maintenance of the mitochondrial genome in humans, fungi, and plants. These were extensively studied in the context of group II intron splicing, especially in yeast mitochondria. A few mitochondrially localized RNA helicases have been described in S. cerevisiae, IRC3, MRH4, MSS116 and SUV3. Mutants in genes encoding these four mitochondrial helicases all show mtDNA defects and exhibit a petite mutant phenotype (i.e., formation of small anerobic like colonies) [59]. The petite phenotype is tightly correlated with altered respiratory functions, while mtDNA maintenance can be directly rely on the activities of RNA helicases, or may indirectly influenced by variations in mitochondria gene expression. The mtDNA copy numbers were shown be regulated by both internal and external stimuli. Proteins that control mtDNA replication can indirectly also affect organellar gene expression, by increasing the number of gene-loci that are being actively expressed. Correspondingly, altered mtDNA expression (in the transcriptional, post-transcriptional or translational levels) may also affects the maintenance or stability of the mtDNA [60,61], a phenomenon that may relate to deleterious effects of partially assembled respiratory complexes, e.g., as indicated in the case of ATP synthase enzyme assembly intermediates [62,63]. Hence, the loss, or a partial reduction, in mtDNA copy numbers may allow the cells to survive under conditions that disrupt the assembly of enzymes of the OXPHOS pathway [64].
The IRC3 enzyme contains a DEAH tetrapeptide within its RNA helicase motif II (Figure 2), but the deduced protein sequence seems to be closely related to the DEAD subfamily [65]. IRC3 was identified in a genetic screen for genes required for respiratory growth, mtDNA maintenance and mitochondrial protein synthesis in Saccharomyces cerevisiae [66], and was mainly associated with mitogenome maintenance or replication [64]. The specific roles of IRC3 in mtDNA maintenance, RNA metabolism and or organellar functions are still investigated. Genetic analyses indicate that the loss of IRC3 results in mtDNA fragmentation [43]. However, as indicated above, genetic analyses showed that the mtDNA defect phenotypes might be indirectly related to altered mitochondrial gene expression or RNA metabolism defects in the yeast cells [60].
Another member of the mitochondrial DEAD-box helicases of S. cerevisiae is MRH4, which its loss also results with a petite mutant phenotype and altered mitogenome stability [67]. The MRH4 gene was identified in a genetic screen of nuclear genes that assist splicing of mitochondrial localized group II introns in yeast [67]. The MRH4 was originally defined as a suppressor of a splicing defect linked to a point mutation in the ai5γ intron [67]. However, the main function(s) of the protein seem to be rather associated with mitochondrial translation and/or ribosome biogenesis [64]. Accordingly, null mrh4 mutants display defects in the assembly of the mitochondrial large ribosomal subunit (LSU), and the MRH4 is suggested to play key roles in the late stages of the mitoribosome assembly by ‘promoting remodeling of the 21S rRNA-protein interactions’ [44].
The SUV3 gene-locus (Table 1) encodes an extensively studied RNA helicase that belongs to the DExH subfamily, and harbors an atypical D-E-I-Q sequence within the helicase motif II (Figure 2) [46,47]. Genetic and biochemical studies indicate that SUV3 is a component of the mitochondrial degradosome, and acts as a 3’ to 5’ exonuclease complex together with the DSS1 ribonuclease [47,68,69,70,71,72]. These data also suggested that SUV3 functions in splicing and the degradation of excised introns (i.e., null mutations in the degradosome components result with overaccumulation of intronic sequences and a reduction in mRNA levels), although the functions of the protein seem to be primarily associated with global mtRNA turnover and surveillance [73,74]. Defects in the organellar degradosomes led to defects in the processing of structural RNAs (e.g., rRNA and tRNAs), which result in altered organellar translation and mitogenome instability. It is hypothesized that the degradation of the excised organellar introns may enables the recycling of lowly-expressed splicing cofactors, which are typically tightly associated with their intron RNA ligands.
MSS116 is a model DEAD box helicase, which contains the canonical tetrapeptide motif. The functions of MSS116 protein are mainly linked to organellar splicing (Table 1). The MSS116 gene was initially identified in a genetic screen for factors that regulate the splicing of organellar introns in yeast [75]. Experimental data further indicated that MSS116 assists in the splicing of virtually all the group I and II introns in yeast mitochondria [45]. MSS116 was also found to affect organellar translation, as indicated in mutant lines of yeast strains lacking mitochondrial introns, where the most notable reductions in protein levels were apparent in the cases of Cox1 and Cox3 [75]. Notably, the analysis of MSS116 sheds important light on the functions of RNA helicases in intron RNA splicing. In the group I ai5β intron, MSS116 was shown to facilitates a conformational change in the intron structure, following the first transesterification step (i.e., the release of the 5’ exon) that is necessary for the exon’s ligation reaction [76]. Also, MSS116 was found to assist in the folding of the group II ai2 intron, together with the ai2 maturase factor, into its catalytically active form [45]. These activities could relate to the roles of RNA helicases [77] as either RNA chaperons by unwinding undesirable structure intermediates (kinetic traps) of short duplexes [78], or alternatively may assist in the folding of the RNA by binding and stabilizing specific secondary and/or tertiary RNA base interactions (‘on-pathway intermediates’), in an ATP-independent manner [79]. Analysis of the 3D structure and biochemical assays indicated that the MSS116’s helicase core cooperates with its C-terminal domain to facilitate the intron splicing [80,81]. These analyses suggested that the C-terminal region of MSS116 has two distinct roles, by assisting in RNA binding as well as n stabilizing the helicase core. Remarkably, the functions of MSS116 in ai5γ intron splicing can be complemented by some other RNA helicases, such as the mitochondrial CYT-19 of Neurospora crassa, or the cytoplasmic DED1 helicase in yeast that its functions are related to translation initiation [45,82].
5. Mitochondrial RNA Helicases Factors in Arabidopsis thaliana Plants
The nuclear genomes of land plants seem to encode a high number of DExH/D RNA helicases, as compared with other organisms [83]. More than 100 SF2 DNA or RNA helicase genes were previously described in the nuclear genome of the model angiosperms Arabidopsis thaliana and rice (Oryza sativa) [83,84]. The majority of the genes seem to be constitutively expressed in the plant, and are likely to encode housekeeping proteins which are required for the maintenance of basic cellular functions. Although these are expected to have important roles in DNA or RNA metabolism, the specific functions of the majority of these enzymes are currently unknown.
The Arabidopsis Information Resource (TAIR) and the UniProt databases suggest the presence of numerous genes encoding DNA or RNA helicases, with 92 of them being postulated to function as RNA helicases (Supplementary Table S1, Figure 3).
Similar to previous reports [83,84], our phylogenetic analysis also indicates that the plant RNA helicases are clustered into 3 main subgroups, which include DEAH, DExH and DEAD -box RNA helicase family proteins (Figure 3). At least nine of the RNA helicases genes encode protein cofactors, which are predicted or shown to reside within the mitochondria in Arabidopsis (Table 1, Figure 3). These include a homolog of the yeast SUV3 protein that is also identified in rice, however, the specific roles of this factor in plant organellar RNA (or DNA) metabolism await further characterization (Table1, and [53,54,55]). Notably, three specific RNA helicases (i.e., ABO6, PMH2 and RH33), which were also subjected to detailed analysis, were all shown to be involved in mitochondrial group II introns splicing.
PUTATIVE MITOCHONDRIAL RNA HELICASE 1 (PMH1) and PMH2 that are situated very close to one another within chromosome 3 (Table1), and also share a high sequence similarity, are likely representing a recent gene duplication event [85]. The PMH1 and PMH2 proteins harbor a DEAD-box helicase subdomain, and a N-terminal region that is related to plant mitochondria localization signals. Accordingly, both PMH1 and PMH2 are identified in high molecular weight ribonucleoprotein (RNP) complexes in Arabidopsis mitochondria [50]. The PMH1 gene is expressed at low levels in a tissue-specific manner (mainly in the flowers) and seem to be induced by cold stress, while PMH2 likely encodes a housekeeping factor that is expressed in all tissues throughout the plant growth and development. PMH2 was found to function as a general splicing factor, influencing the processing of numerous group II introns in Arabidopsis mitochondria [35,51]. Notably, the plant maturase factor (nMAT2; [86]) and PMH2 function in the splicing of a similar subset of group II introns [35,51]. It is, therefore, tempting to speculate that similarly to the roles of MSS116 in yeast, PMH2 may also function as an RNA chaperone that assists in the folding of maturase-bound pre-RNA transcripts in Arabidopsis mitochondria. Yet, much less is known about the roles of the PMH1 paralog in organellar RNA metabolism and gene expression. Based on its high sequence similarity with PMH2, and the lack of molecular phenotypes of PMH1 knockout mutant lines, it is plausible that PMH1 may possess some functional redundancies with those of PMH2, especially in the flower tissues or under specific growth conditions, where the expression of the gene is upregulated [50].
ABA OVERLY SENSITIVE 6 (ABO6) encodes a member of the DExH box helicase subfamily, which was identified in a genetic screen for ABA mediated inhibition of primary root growth [48]. A point mutation in the sixth exon of ABO6 gene-locus leads to reduced splicing efficiency of several mitochondrial pre-RNAs encoding the respiratory complex I Nad1, Nad2, Nad4 and Nad5 subunits. The point mutation (Gly-334 to Glu-334) in abo6-1 mutant is located within the DExH motif of ABO6, in which the Gly-334 is identified as a well-preserved amino acid in ABO6 orthologs in different plant species. The functions of ABO6 seems to be essential during embryogenesis and early plant development, as heterozygous T-DNA insertional lines found within the coding region of AT5G04895 locus are unable to produce homozygous progeny. The growth and developmental defect phenotypes, and reduced levels of various organellar nad mRNA transcripts in abo6 mutants, are likely associated with altered respiratory complex I (CI) biogenesis and function [19,87,88,89]. The accumulation of holo-CI (or partially-assembled CI intermediates) in abo6 mutant-lines has not been analyzed yet, and no data currently exists regarding the respiratory functions of the mutants versus the wild type plants. However, the mutants are anticipated to exhibit altered OXPHOS functions, as the abo6 mutants show an induction of the alternative respiratory pathway (i.e., alternative oxidases and rotenone-insensitive NADH reductases) and increased ROS production [48].
The DEA(D/H)-box RNA helicase family protein, RH33 (encoded by the At2g07750 gene locus) (Table 1 and Table S1), is another mitochondrial helicase, which based on own preliminary data, is associated with mitochondrial group II introns splicing in Arabidopsis plants (Supplementary Figures S1 and S2). Based on its N-terminal region, the RH33 protein product is predicted to be localized to the mitochondria (Table 1 and Table S1; [57]). To establish the putative roles of RH33 in mitochondria RNA (mtRNA) metabolism, we analyzed the growth phenotypes and RNA profiles of three individual T-DNA insertional lines within the AT2G07750 gene locus, i.e., Sail_604_A01 (rh33-1) (found within the 5’-UTR, 16 nucleotides upstream to the AUG site), Salk_119034 (rh33-2) (located 221 nts upstream to the AUG site within the 5’-UTR), and Salk_119725 (rh33-3; found in the coding region, inside the exon 7 region) (Figure S1A). The rh33-1 mutants exhibited normal growth phenotypes when grown on MS-agar plats, under optimal growth conditions (i.e., at 22oC), but showed retarded growth phenotypes, and more notably altered root morphology, when grown at 28oC (Figure S1B). An association between mtRNA processing defects and altered root development was recently indicated for a few temperature-sensitive mutants encoding mitochondrial RBPs, i.e., the RRD1 a ribonuclease (altered mRNA deadenylation), and two PPR editing factors, RRD2 and RID4 (Otsuka et al. 2021), as well as the rpd1 mutants [90,91,92], which are affected in the expression of the PORR-family splicing cofactor, ROOT PRIMORDIUM DEFECTIVE 1 [92]. Yet, the phenotypes of the homozygous rh33-2 mutant were similar to those of the wild type (Col-0) plants, while no homozygous plants could be established for the rh33-3 line that contains a T-DNA insertion within the 7th exon of RH33 gene (Figure S1A), suggesting that the functions of RH33 are essential during embryogenesis and for normal seed development. A primary analysis of the RNA profiles of rh33 plants indicated to reduced mRNA accumulation of various nad1 and nad7 transcripts in rh33-1 plants, which their coding regions are both interrupted by several group II introns (Figure S1C). However, the RNA profiles of rh33-2 plants (containing a T-DNA insertion in the 5’ UTR) were similar to those of the wild-type plants, possibly because the insertion within the 5’ UTR of rh33-2 has not affected the expression of the gene [93]. We further assumed that the reduced mRNA levels we see in nad1 exons 1-2, 2-3, 3-4, and 4-5, and nad7 exons 2-3 in and rh33-1, may correspond to defects in the excision of the introns that reside within the three mitochondrial nad genes. Indeed, RT-qPCRs further indicated to splicing defects (i.e., accumulation of pre-RNAs that are correlated with reduced levels of their corresponding mRNAs) in the rh33-1 mutant (Figure S2). These data support a role for RH33 in the splicing of various group II-type introns in Arabidopsis mitochondria. The intracellular location(s) and the effects of altered RH33 expression on the respiratory functions, OXPHOS complexes assembly and organellar biogenesis are currently analyzed.
Analysis of a mitochondria-localized RNA helicase in corn (Zea mays), termed as RH48 (Table 1 and Table S1), indicated that the protein facilitates the processing of several organellar group II introns, including the second intron in nad2 (i.e., nad2 i2), nad5 i1, nad7 introns 1, 2 and 3, and the single intron within the ccmFc pre-RNA transcript [52]. The Zm-rh48 mutants displayed seed and embryo developmental defect phenotypes, which are postulated to be associated with altered complex I and cytochrome C biogenesis [52]. Two-hybrid screens indicated that RH48 interacts with several known splicing factors, including PPR-SMR1 and SPR2 [52]. ZmRH48 may also play a role in the C-to-U deamination of (i.e., RNA editing) of C-515 of the atp6 transcript [52]. however, its function in the editing atp6-515 is less pronounced, which may indicate a pleotropic effect due to the altered mtRNA metabolism [32,94,95,96,97]. Arabidopsis contains a homolog of ZmRH48 (AtRH48, At1G63250), but its functions in organellar gene expression and function remain to be addressed.
Another DEAD RNA helicase that is localized to the mitochondria is the INCREASED SIZE EXCLUSION LIMIT 1 (ISE1) protein (Table 1; [49]). Similar to the functions of several other mtRNA processing enzymes, [2,3,19,87], the functions of ISE1 are essential for normal embryogenesis. The ise1 mutants further show altered plasmodesmata (PD) structure and increased PD mediated transport of fluorescent tracers, which may relate to altered cellular metabolism [49]. Similarly, reduced expression of ISE1 in tobacco (Nicotiana benthamiana) cells also leads to increased intercellular movement of a GFP-TMV-MP fusion protein through the PD [49].
In addition to these factors, Arabidopsis plants encode a homologue (i.e., AT4G14760, AtSUV3) of the OsSUV3 protein, which were found to possess RNA unwinding activity and to provide salt tolerance in transgenic over-expressed lines [53,55]. Due to their high homology with the yeast SUV3 protein [46,47], it has been proposed that the plant orthologs also play a role in the mitochondrial degradosomes, but such hypotheses need to be experimentally supported. No data currently exist for the DExH18 protein, which is encoded by the AT5G39840 gene-locus in Arabidopsis and predicted to be localized to the mitochondria (Table 1). The phylogenetic data indicates that AT5G39840 is clustered together with various DExH helicase proteins (Figure 3).
6. Conclusions and Perspectives
RNA helicases are key components in the complex cellular RNA metabolism activities. These factors also play pivotal roles in maintaining mtDNA maintenance, and in organellar gene expression. Their unwinding activity is fundamental for the proper maturation and functioning of numerous RNA molecules, thereby contributing to the regulation of gene expression (transcription, post-transcriptional RNA processing and translation) and overall cellular homeostasis. Ongoing research continues to uncover the specific roles and mechanisms of different RNA helicases in cellular, nuclear and organellar RNA metabolism. As indicated above, canonical group I and II introns are catalytic RNAs that are able to catalyze their own excision in vitro (usually under non-physiological conditions) [26,32,33,34,98,99]. However, for their efficient splicing in vivo, all group I or II introns rely on the activities of various proteinaceous cofactors. In yeast (and plant) mitochondria these include maturases, as well as various nuclear-encoded RNA biding proteins, which function on specific introns or may generally affect the splicing of several or many pre-RNAs [21]. RNA helicases were shown to be required for efficient splicing of all the mitochondrial group I and II introns, including those that do not require the assistance of an intron-encoded maturase
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1: The RNA helicase RH33 affects the processing of various protein-encoding transcripts in Arabidopsis mitochondria; Figure S2: Analysis of the splicing efficiencies of mitochondrial group II-type introns in rh33-1 mutant; Table S1: RNA helicases encoded in Arabidopsis thaliana genome.
Author Contributions
methodology, R.M. and O.O.B.; formal analysis, R.M.; writing—original draft preparation R.M. and O.O.B.; writing—review and editing, R.M. and O.O.B.; supervision, O.O.B.; funding acquisition, O.O.B. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by grants to O.O.B from the ‘Israeli Science Foundation’ ISF grants no. 1834/20 and 3254-2020.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We thank Dr. Hagit Zer for useful comments and discussions about the manuscript data and writing.
Conflicts of Interest
The authors declare no conflict of interest. The funders played no role in the writing of the manuscript or the decision to publish the results.
References
- Gualberto, J. M.; Newton, K. J. , Plant mitochondrial genomes: Dynamics and mechanisms of mutation. Annu Rev Plant Biol 2017, 68, 225–252. [Google Scholar] [CrossRef]
- Zmudjak, M.; Ostersetzer-Biran, O., RNA Metabolism and Transcript Regulation. In Annu Plant Rev online, 2018; pp 143-184.
- Small, I.; Melonek, J.; Bohne, A.-V.; Nickelsen, J.; Schmitz-Linneweber, C. , Plant organellar RNA maturation. Plant Cell 2023, 35(6), 1727–1751. [Google Scholar] [CrossRef]
- Best, C.; Mizrahi, R.; Ostersetzer-Biran, O. , Why so complex? The intricacy of genome structure and gene expression, associated with angiosperm mitochondria, may relate to the regulation of embryo quiescence or dormancy - intrinsic blocks to early plant life. Plants 2020, 9(5), 598. [Google Scholar] [CrossRef]
- Fuchs, P.; Rugen, N.; Carrie, C.; Elsasser, M.; Finkemeier, I.; Giese, J.; Hildebrandt, T. M.; Kuhn, K.; Maurino, V. G.; Ruberti, C.; Schallenberg-Rudinger, M.; Steinbeck, J.; Braun, H. P.; Eubel, H.; Meyer, E. H.; Muller-Schussele, S. J.; Schwarzlander, M. , Single organelle function and organization as estimated from Arabidopsis mitochondrial proteomics. Plant J 2020, 101(2), 420–441. [Google Scholar] [CrossRef]
- Zeng, C.; Jian, Y.; Vosoughi, S.; Zeng, C.; Zhao, Y. , Evaluating native-like structures of RNA-protein complexes through the deep learning method. Nat Comm 2023, 14(1), 1060. [Google Scholar] [CrossRef]
- Chung, C. S.; Tseng, C. K.; Lai, Y. H.; Wang, H. F.; Newman, A. J.; Cheng, S. C. , Dynamic protein-RNA interactions in mediating splicing catalysis. Nucleic Acids Res 2019, 47(2), 899–910. [Google Scholar] [CrossRef] [PubMed]
- Glisovic, T.; Bachorik, J. L.; Yong, J.; Dreyfuss, G. , RNA-binding proteins and post-transcriptional gene regulation. FEBS Lett 2008, 582(14), 1977–86. [Google Scholar] [CrossRef] [PubMed]
- Licatalosi, D. D.; Darnell, R. B. , RNA processing and its regulation: global insights into biological networks. Nat Rev Genet 2010, 11(1), 75–87. [Google Scholar] [CrossRef] [PubMed]
- Spitale, R. C.; Incarnato, D. , Probing the dynamic RNA structurome and its functions. Nat Rev Genet 2023, 24(3), 178–196. [Google Scholar] [CrossRef] [PubMed]
- Assmann, S. M.; Chou, H.-L.; Bevilacqua, P. C. , Rock, scissors, paper: How RNA structure informs function. Plant Cell 2023, 35(6), 1671–1707. [Google Scholar] [CrossRef] [PubMed]
- Cruz, J. A.; Westhof, E. , The Dynamic Landscapes of RNA Architecture. Cell 2009, 136(4), 604–609. [Google Scholar] [CrossRef]
- Bourgeois, C. F.; Mortreux, F.; Auboeuf, D. , The multiple functions of RNA helicases as drivers and regulators of gene expression. Nat Rev Mol Cell Biol 2016, 17(7), 426–438. [Google Scholar] [CrossRef]
- Jankowsky, E. , RNA helicases at work: binding and rearranging. Trends Biochem Scie 2011, 36(1), 19–29. [Google Scholar] [CrossRef]
- Owttrim, G. W. , RNA helicases: diverse roles in prokaryotic response to abiotic stress. RNA biol 2013, 10(1), 96–110. [Google Scholar] [CrossRef] [PubMed]
- Fairman-Williams, M. E.; Guenther, U. P.; Jankowsky, E. , SF1 and SF2 helicases: family matters. Curr Opin Struct Biol 2010, 20(3), 313–24. [Google Scholar] [CrossRef] [PubMed]
- Hammani, K.; Giege, P. , RNA metabolism in plant mitochondria. Trends Plant Sci 2014, 19(6), 380–9. [Google Scholar] [CrossRef] [PubMed]
- Nawaz, G.; Kang, H. , Chloroplast- or Mitochondria-Targeted DEAD-Box RNA Helicases Play Essential Roles in Organellar RNA Metabolism and Abiotic Stress Responses. Front Plant Sci 2017, 8, 871. [Google Scholar] [CrossRef] [PubMed]
- Colas des Francs-Small, C.; Small, I. , Surrogate mutants for studying mitochondrially encoded functions. Biochimie 2014, 100(0), 234–242. [Google Scholar] [CrossRef] [PubMed]
- Martin, W.; Koonin, E. V. , Introns and the origin of nucleus-cytosol compartmentalization. Nature 2006, 440(7080), 41–45. [Google Scholar] [CrossRef]
- Schmitz-Linneweber, C.; Lampe, M.-K.; Sultan, L. D.; Ostersetzer-Biran, O., Organellar maturases: A window into the evolution of the spliceosome. BBA - Bioenergetics 2015, 1847, (9), 798-808. [CrossRef]
- Cech, T. R. , The RNA worlds in context. Cold Spring Harb Perspect Biol 2012, 4(7), a006742. [Google Scholar] [CrossRef]
- Zimmerly, S.; Semper, C. , Evolution of group II introns. Mobile DNA 2015, 6(1), 1–19. [Google Scholar] [CrossRef]
- Lambowitz, A. M.; Belfort, M. , Mobile bacterial group II introns at the crux of eukaryotic evolution. Microbiol spect 2015, 3(1), 1–26. [Google Scholar]
- Catania, F.; Lynch, M. , Where do introns come from? PLoS Biol 2008, 6(11), e283. [Google Scholar] [CrossRef]
- Cech, T. R. , The generality of self-splicing RNA: Relationship to nuclear messenger-RNA splicing. Cell 1986, 44(2), 207–210. [Google Scholar] [CrossRef]
- ambowitz, A. M.; Caprara, M. G.; Zimmerly, S.; Perlman, P., Group I and Group II Ribozymes as RNPs: Clues to the Past and Guides to the Future. In The RNA World, R. Gesteland, T. Cech, and J. Atkins, eds (Cold Spring Harbor: Cold Spring Harbor Laboratory Press). 1999, pp. 451-485.
- Pyle, A. M.; Lambowitz, A. M., Group II Introns: Ribozymes that splice RNA and invade DNA. In: The RNA World, R. Gesteland, TR. Cech, and J. Atkins (eds), 3rd eddition. Cold Spring Harbor:. 1999.
- Michel, F.; Umesono, K.; Ozeki, H. , Comparative and functional anatomy of group II catalytic introns — a review. Gene 1989, 82(1), 5–30. [Google Scholar] [CrossRef]
- Ferat, J. L.; Michel, F. , Group-II Self-Splicing Introns in Bacteria. Nature 1993, 364(6435), 358–361. [Google Scholar] [CrossRef]
- Bonen, L.; Vogel, J. , The ins and outs of group II introns. Trends Genet 2001, 17(6), 322–331. [Google Scholar] [CrossRef]
- Bonen, L. , Cis- and trans-splicing of group II introns in plant mitochondria. Mitochondrion 2008, 8(1), 26–34. [Google Scholar] [CrossRef]
- Brown, G. G.; Colas des Francs-Small, C.; Ostersetzer-Biran, O. , Group II intron splicing factors in plant mitochondria. Front Plant Sci 2014, 5(35), 1–13. [Google Scholar] [CrossRef] [PubMed]
- Sharp, P. A. , On the origin of RNA splicing and introns. Cell 1985, 42(2), 397–400. [Google Scholar] [CrossRef]
- Zmudjak, M.; Shevtsov, S.; Sultan, L. D.; Keren, I.; Ostersetzer-Biran, O. , Analysis of the roles of the Arabidopsis nMAT2 and PMH2 proteins provided with new insights into the regulation of group II intron splicing in land-plant mitochondria. Int J Mol Sci 2017, 18(11), 1–25. [Google Scholar] [CrossRef] [PubMed]
- Møller, I. M.; Rasmusson, A. G.; Van Aken, O. , Plant mitochondria – past, present and future. Plant J 2021, 108(4), 912–959. [Google Scholar] [CrossRef] [PubMed]
- Mizrahi, R.; Shevtsov-Tal, S.; Ostersetzer-Biran, O. , Group II intron-encoded proteins (IEPs/maturases) as key regulators of Nad1 expression and Complex I biogenesis in land plant mitochondria. Genes 2022, 13(7), 1–16. [Google Scholar] [CrossRef] [PubMed]
- Mohr, G.; Lambowitz, A. M. , Putative proteins related to group II intron reverse transcriptase/maturases are encoded by nuclear genes in higher plants. Nucleic Acids Res 2003, 31(2), 647–652. [Google Scholar] [CrossRef] [PubMed]
- Small, I. D.; Peeters, N. , The PPR motif - a TPR-related motif prevalent in plant organellar proteins. Trends Biochem Sci 2000, 25(2), 46–7. [Google Scholar] [CrossRef]
- Fernandez-Silva, P.; Martinez-Azorin, F.; Micol, V.; Attardi, G. , The human mitochondrial transcription termination factor (mTERF) is a multizipper protein but binds to DNA as a monomer, with evidence pointing to intramolecular leucine zipper interactions. EMBO J 1997, 16(5), 1066–79. [Google Scholar] [CrossRef]
- Kleine, T. , Arabidopsis thaliana mTERF proteins: evolution and functional classification. Front Plant Sci 2012, 3, 233. [Google Scholar] [CrossRef]
- Robles, P.; Micol, J. L.; Quesada, V. , Unveiling Plant mTERF Functions. Mol Plant 2012, 5(2), 294–296. [Google Scholar] [CrossRef]
- Sedman, T.; Gaidutšik, I.; Villemson, K.; Hou, Y.; Sedman, J. , Double-stranded DNA-dependent ATPase Irc3p is directly involved in mitochondrial genome maintenance. Nucleic Acids Res 2014, 42(21), 13214–13227. [Google Scholar] [CrossRef]
- De Silva, D.; Fontanesi, F.; Barrientos, A. , The DEAD box protein Mrh4 functions in the assembly of the mitochondrial large ribosomal subunit. Cell Metabol 2013, 18(5), 712–25. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.-R.; Rowe, C. E.; Mohr, S.; Jiang, Y.; Lambowitz, A. M.; Perlman, P. S. , The splicing of yeast mitochondrial group I and group II introns requires a DEAD-box protein with RNA chaperone function. Proc Natl Acad Sci U S A 2005, 102(1), 163–168. [Google Scholar] [CrossRef] [PubMed]
- Conrad-Webb, H.; Perlman, P. S.; Zhu, H.; Butow, R. A. , The nuclear SUV3-1 mutation affects a variety of post-transcriptional processes in yeast mitochondria. Nucleic Acids Res 1990, 18, 1369–1376. [Google Scholar] [CrossRef] [PubMed]
- Stepien, P. P.; Margossian, S. P.; Landsman, D.; Butow, R. A. , The yeast nuclear gene suv3 affecting mitochondrial post-transcriptional processes encodes a putative ATP-dependent RNA helicase. Proc Natl Acad Sci U S A 1992, 89(15), 6813–6817. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Duan, Y.; Hua, D.; Fan, G.; Wang, L.; Liu, Y.; Chen, Z.; Han, L.; Qu, L.-J.; Gong, Z. , DEXH box RNA helicase–mediated mitochondrial reactive oxygen species production in Arabidopsis mediates crosstalk between abscisic acid and auxin signaling. Plant Cell 2012, 24(5), 1815–1833. [Google Scholar] [CrossRef]
- Stonebloom, S.; Burch-Smith, T.; Kim, I.; Meinke, D.; Mindrinos, M.; Zambryski, P. , Loss of the plant DEAD-box protein ISE1 leads to defective mitochondria and increased cell-to-cell transport via plasmodesmata. Proc Natl Acad Sci U S A 2009, 106(40), 17229–34. [Google Scholar] [CrossRef] [PubMed]
- Matthes, A.; Schmidt-Gattung, S.; Köhler, D.; Forner, J.; Wildum, S.; Raabe, M.; Urlaub, H.; Binder, S. , Two DEAD-box proteins may be part of RNA-dependent high-molecular-mass protein complexes in Arabidopsis mitochondria. Plant Physiol 2007, 145(4), 1637–1646. [Google Scholar] [CrossRef] [PubMed]
- Kohler, D.; Schmidt-Gattung, S.; Binder, S., The DEAD-box protein PMH2 is required for efficient group II intron splicing in mitochondria of Arabidopsis thaliana. Plant Mol Biol 2010, 72, (4-5), 459-67. [CrossRef]
- Yang, Y. Z.; Ding, S.; Liu, X. Y.; Xu, C.; Sun, F.; Tan, B. C. , The DEAD-box RNA helicase ZmRH48 is required for the splicing of multiple mitochondrial introns, mitochondrial complex biosynthesis, and seed development in maize. J Integr Plant Biol 2023, 65(11), 2456–2468. [Google Scholar] [CrossRef] [PubMed]
- Gagliardi, D.; Kuhn, J.; Spadinger, U.; Brennicke, A.; Leaver, C. J.; Binder, S. , An RNA helicase (AtSUV3) is present in Arabidopsis thaliana mitochondria. FEBS Letters 1999, 458(3), 337–342. [Google Scholar] [CrossRef]
- Tuteja, N.; Tarique, M.; Tuteja, R. , Rice SUV3 is a bidirectional helicase that binds both DNA and RNA. BMC Plant Biol 2014, 14, 283. [Google Scholar] [CrossRef]
- Sahoo, R. K.; Ansari, M. W.; Tuteja, R.; Tuteja, N., OsSUV3 transgenic rice maintains higher endogenous levels of plant hormones that mitigates adverse effects of salinity and sustains crop productivity. Rice (N Y) 2014, 7, (1), 17. [CrossRef]
- Wallet, C.; Le Ret, M.; Bergdoll, M.; Bichara, M.; Dietrich, A.; Gualberto, J. M. , The RECG1 DNA Translocase Is a Key Factor in Recombination Surveillance, Repair, and Segregation of the Mitochondrial DNA in Arabidopsis. Plant Cell 2015, 27(10), 2907–2925. [Google Scholar] [CrossRef]
- Hooper, C. M.; Castleden, I. R.; Tanz, S. K.; Aryamanesh, N.; Millar, A. H. , SUBA4: the interactive data analysis centre for Arabidopsis subcellular protein locations. Nucleic Acids Res 2017, 45(D1), D1064–D1074. [Google Scholar] [CrossRef]
- Almagro Armenteros, J. J.; Salvatore, M.; Emanuelsson, O.; Winther, O.; von Heijne, G.; Elofsson, A.; Nielsen, H., Detecting sequence signals in targeting peptides using deep learning. Life Sci Alliance 2019, 2, (5). [CrossRef]
- Bernardi, G. , The petite mutation in yeast. Trends Biochem Sci 1979, 4(9), 197–201. [Google Scholar] [CrossRef]
- Myers, A. M.; Pape, L. K.; Tzagoloff, A. , Mitochondrial protein synthesis is required for maintenance of intact mitochondrial genomes in Saccharomyces cerevisiae. EMBO J 1985, 4(8), 2087–2092. [Google Scholar] [CrossRef]
- Lipinski, K. A.; Kaniak-Golik, A.; Golik, P., Maintenance and expression of the S. cerevisiae mitochondrial genome—from genetics to evolution and systems biology. BBA-Bioenergetics 2010, 1797, (6-7), 1086-1098.
- Duvezin-Caubet, S.; Rak, M.; Lefebvre-Legendre, L.; Tetaud, E.; Bonnefoy, N.; di Rago, J.-P. , A "petite obligate" mutant of Saccharomyces cerevisiae: functional mtDNA is lethal in cells lacking the delta subunit of mitochondrial F1-ATPase. J Biol Chem 2006, 281(24), 16305–16313. [Google Scholar] [CrossRef]
- Mueller, D. M. , Partial Assembly of the Yeast Mitochondrial ATP Synthase1. J Bioenerg Biomem 2000, 32(4), 391–400. [Google Scholar] [CrossRef] [PubMed]
- Szczesny, R. J.; Wojcik, M. A.; Borowski, L. S.; Szewczyk, M. J.; Skrok, M. M.; Golik, P.; Stepien, P. P., Yeast and human mitochondrial helicases. Biochimica et Biophysica Acta (BBA) - Gene Reg Mechan 2013, 1829, (8), 842-853.
- de la Cruz, J.; Kressler, D.; Linder, P. , Unwinding RNA in Saccharomyces cerevisiae: DEAD-box proteins and related families. Trends Biochem Sci 1999, 24(5), 192–198. [Google Scholar] [CrossRef]
- Merz, S.; Westermann, B. , Genome-wide deletion mutant analysis reveals genes required for respiratory growth, mitochondrial genome maintenance and mitochondrial protein synthesis in Saccharomyces cerevisiae. Genome Biol 2009, 10(9), 1–18. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, U.; Lehmann, K.; Stahl, U. , A novel mitochondrial DEAD box protein (Mrh4) required for maintenance of mtDNA in Saccharomyces cerevisiae. FEMS Yeast Res 2002, 2(3), 267–276. [Google Scholar]
- Zhu, H.; Conrad-Webb, H.; Sheng Liao, X.; Perlman, P. S.; Butow, R. A. , Functional Expression of a Yeast Mitochondrial Intron-Encoded Protein Requires RNA Processing at a Conserved Dodecamer Sequence at the 3′ End of the Gene. Mol Cell Biol 1989, 9(4), 1507–1512. [Google Scholar]
- Conrad-Webb, H.; Perlman, P. S.; Zhu, H.; Butow, R. A. , The nuclear SUV3-1 mutation affects a variety of post-transcriptional processes in yeast mitochondria. Nucleic Acids Res 1990, 18(6), 1369–1376. [Google Scholar] [CrossRef]
- Margossian, S. P.; Li, H.; Zassenhaus, H. P.; Butow, R. A. , The DExH box protein Suv3p is a component of a yeast mitochondrial 3′-to-5′ exoribonuclease that suppresses group I intron toxicity. Cell 1996, 84(2), 199–209. [Google Scholar] [CrossRef]
- Stepien, P. P.; Kokot, L.; Leski, T.; Bartnik, E. , The suv3 nuclear gene product is required for the in vivo processing of the yeast mitochondrial 21 s rRNA transcripts containing the r1 intron. Curr Genet 1995, 27, 234–238. [Google Scholar] [CrossRef]
- Golik, P.; Szczepanek, T.; Bartnik, E.; Stepien, P. P.; Lazowska, J. , The S. cerevisiae nuclear gene SUV3 encoding a putative RNA helicase is necessary for the stability of mitochondrial transcripts containing multiple introns. Curr Genet 1995, 28, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Dziembowski, A.; Piwowarski, J.; Hoser, R.; Minczuk, M.; Dmochowska, A.; Siep, M.; van der Spek, H.; Grivell, L.; Stepien, P. P. , The yeast mitochondrial degradosome: its composition, interplay between RNA helicase and RNase activities and the role in mitochondrial RNA metabolism. J Biol Chem 2003, 287(3), 1603–1611. [Google Scholar] [CrossRef] [PubMed]
- Rogowska, A. T.; Puchta, O.; Czarnecka, A. M.; Kaniak, A.; Stepien, P. P.; Golik, P. , Balance between transcription and RNA degradation is vital for Saccharomyces cerevisiae mitochondria: reduced transcription rescues the phenotype of deficient RNA degradation. Mol Biol Cell 2006, 17(3), 1184–1193. [Google Scholar] [CrossRef]
- Séraphin, B.; Simon, M.; Boulet, A.; Faye, G. , Mitochondrial splicing requires a protein from a novel helicase family. Nature 1989, 337(6202), 84–87. [Google Scholar] [CrossRef] [PubMed]
- Bifano, A. L.; Caprara, M. G. , A DExH/D-box protein coordinates the two steps of splicing in a group I intron. J Mol Biol 2008, 383(3), 667–682. [Google Scholar] [CrossRef] [PubMed]
- Rajkowitsch, L.; Chen, D.; Stampfl, S.; Semrad, K.; Waldsich, C.; Mayer, O.; Jantsch, M. F.; Konrat, R.; Bläsi, U.; Schroeder, R. , RNA Chaperones, RNA Annealers and RNA Helicases. RNA Biol 2007, 4(3), 118–130. [Google Scholar] [CrossRef]
- Russell, R.; Jarmoskaite, I.; Lambowitz, A. M. , Toward a molecular understanding of RNA remodeling by DEAD-box proteins. RNA Biol 2013, 10(1), 44–55. [Google Scholar] [CrossRef]
- Solem, A.; Zingler, N.; Pyle, A. M. , A DEAD protein that activates intron self-splicing without unwinding RNA. Mol Cell 2006, 24(4), 611–617. [Google Scholar] [CrossRef]
- Mohr, G.; Del Campo, M.; Mohr, S.; Yang, Q.; Jia, H.; Jankowsky, E.; Lambowitz, A. M. , Function of the C-terminal domain of the DEAD-box protein Mss116p analyzed in vivo and in vitro. J Mol Biol 2008, 375(5), 1344–1364. [Google Scholar] [CrossRef]
- Del Campo, M.; Lambowitz, A. M. , Structure of the Yeast DEAD box protein Mss116p reveals two wedges that crimp RNA. Mol Cell 2009, 35(5), 598–609. [Google Scholar] [CrossRef] [PubMed]
- Halls, C.; Mohr, S.; Del Campo, M.; Yang, Q.; Jankowsky, E.; Lambowitz, A. M., Involvement of DEAD-box proteins in group I and group II intron splicing. Biochemical characterization of Mss116p, ATP hydrolysis-dependent and-independent mechanisms, and general RNA chaperone activity. J Mol Biol 2007, 365, (3), 835-855. [CrossRef]
- Linder, P.; Owttrim, G. W. , Plant RNA helicases: linking aberrant and silencing RNA. Trends Plant Sci 2009, 14(6), 344–352. [Google Scholar] [CrossRef] [PubMed]
- Umate, P.; Tuteja, R.; Tuteja, N. , Genome-wide analysis of helicase gene family from rice and Arabidopsis: a comparison with yeast and human. Plant Mol Biol 2010, 73(4), 449–465. [Google Scholar] [CrossRef] [PubMed]
- Lallemand, T.; Leduc, M.; Landès, C.; Rizzon, C.; Lerat, E., An Overview of Duplicated Gene Detection Methods: Why the Duplication Mechanism Has to Be Accounted for in Their Choice. Genes 2020, 11, (9). [CrossRef]
- Keren, I.; Bezawork-Geleta, A.; Kolton, M.; Maayan, I.; Belausov, E.; Levy, M.; Mett, A.; Gidoni, D.; Shaya, F.; Ostersetzer-Biran, O. , AtnMat2, a nuclear-encoded maturase required for splicing of group-II introns in Arabidopsis mitochondria. RNA 2009, 15(12), 2299–2311. [Google Scholar] [CrossRef] [PubMed]
- Ostersetzer-Biran, O. , Respiratory complex I and embryo development. J Exp Bot 2016, 67(5), 1205–7. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, K.; Obata, T.; Feher, K.; Bock, R.; Fernie, A. R.; Meyer, E. H. , Complete mitochondrial complex I deficiency induces an up-regulation of respiratory fluxes that is abolished by traces of functional complex I. Plant Physiol 2015, 168(4), 1537–49. [Google Scholar] [CrossRef] [PubMed]
- Braun, H.-P.; Binder, S.; Brennicke, A.; Eubel, H.; Fernie, A. R.; Finkemeier, I.; Klodmann, J.; König, A.-C.; Kühn, K.; Meyer, E.; Obata, T.; Schwarzländer, M.; Takenaka, M.; Zehrmann, A. , The life of plant mitochondrial complex I. Mitochondrion 2014, 19, 295–313. [Google Scholar] [CrossRef]
- Konishi, M.; Sugiyama, M. , Genetic analysis of adventitious root formation with a novel series of temperature-sensitive mutants of Arabidopsis thaliana. Development 2003, 130(23), 5637–47. [Google Scholar] [CrossRef]
- Konishi, M.; Sugiyama, M. , A novel plant-specific family gene, ROOT PRIMORDIUM DEFECTIVE 1, is required for the maintenance of active cell proliferation. Plant Physiol 2006, 140(2), 591–602. [Google Scholar] [CrossRef]
- Edris, R.; Sultan, L. D.; Best, C.; Mizrahi, R.; Weinstein, O.; Chen, S.; Kamennaya, N. A.; Keren, N.; Zer, H.; Zhu, H.; Ostersetzer-Biran, O. , Root Primordium Defective 1 Encodes an Essential PORR Protein Required for the Splicing of Mitochondria Encoded Group II Introns and for Respiratory Complex I Biogenesis. Plant Cell Physiol 2023. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y. H. , How effective is T-DNA insertional mutagenesis in Arabidopsis? Journal of Biochemical Technology 2008, 1(1), 11–20. [Google Scholar]
- Castandet, B.; Choury, D.; Bégu, D.; Jordana, X.; Araya, A. , Intron RNA editing is essential for splicing in plant mitochondria. Nucleic Acids Res 2010, 38(20), 7112–21. [Google Scholar] [CrossRef] [PubMed]
- Oldenkott, B.; Yamaguchi, K.; Tsuji-Tsukinoki, S.; Knie, N.; Knoop, V. , Chloroplast RNA editing going extreme: more than 3400 events of C-to-U editing in the chloroplast transcriptome of the lycophyte Selaginella uncinata. RNA 2014, 20(10), 1499–1506. [Google Scholar] [CrossRef] [PubMed]
- Small, I. D.; Schallenberg-Rudinger, M.; Takenaka, M.; Mireau, H.; Ostersetzer-Biran, O. , Plant organellar RNA editing: what 30 years of research has revealed. Plant J 2020, 101(5), 1040–1056. [Google Scholar] [CrossRef]
- Yap, A.; Kindgren, P.; Colas des Francs-Small, C.; Kazama, T.; Tanz, S. K.; Toriyama, K.; Small, I. , AEF1/MPR25 is implicated in RNA editing of plastid atpF and mitochondrial nad5, and also promotes atpF splicing in Arabidopsis and rice. Plant J 2015, 81(5), 661–669. [Google Scholar] [CrossRef]
- Lambowitz, A. M.; Zimmerly, S. , Mobile group II introns. Annu Rev Genet 2004, 38(1), 1–35. [Google Scholar] [CrossRef]
- Michel, F.; Lang, B. F. , Mitochondrial class II introns encode proteins related to the reverse transcriptases of retroviruses. Nature 1985, 306(6029), 641–3. [Google Scholar] [CrossRef]
Figure 1.
Schematic illustration of functional helicase domains. The canonical core domains and the sub-helicase motifs fund within the model DEAD-box helicase DEAD in E. Coli, as well as two DExD/H-box helicases from Arabidopsis thaliana (i.e., The DEAD-box PMH2 and the DExH box ABO6) are illustrated. The different sub-motifs (i.e., Q, I, Ia, Ib, Ic, II, III, IV, IVa, V, Va, VI) composing the two RecA-like regions of the DExD/H box proteins are highlighted in light grey. Different colors represent the different biochemical functions of each sub-motif. Red indicates ATP binding sites; blue, RNA binding sites, whereas yellow color indicates linker regions between ATP- and RNA-binding sites). DExH and DEAH box proteins lack the Q sub-motif found in DEAD-box helicases.
Figure 1.
Schematic illustration of functional helicase domains. The canonical core domains and the sub-helicase motifs fund within the model DEAD-box helicase DEAD in E. Coli, as well as two DExD/H-box helicases from Arabidopsis thaliana (i.e., The DEAD-box PMH2 and the DExH box ABO6) are illustrated. The different sub-motifs (i.e., Q, I, Ia, Ib, Ic, II, III, IV, IVa, V, Va, VI) composing the two RecA-like regions of the DExD/H box proteins are highlighted in light grey. Different colors represent the different biochemical functions of each sub-motif. Red indicates ATP binding sites; blue, RNA binding sites, whereas yellow color indicates linker regions between ATP- and RNA-binding sites). DExH and DEAH box proteins lack the Q sub-motif found in DEAD-box helicases.
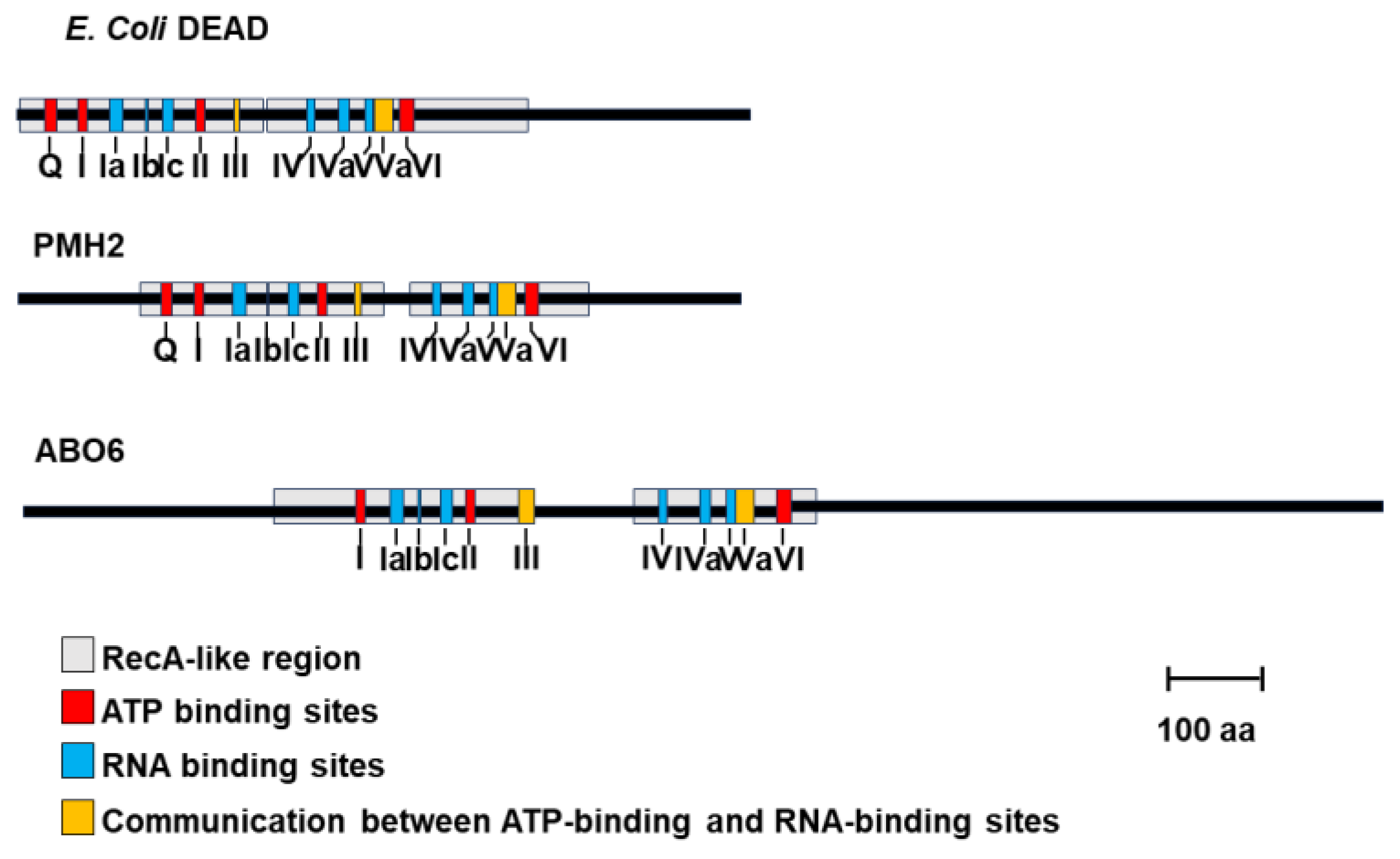
Figure 2.
RNA helicases as mediators of RNA structure remodeling. RNA helicases are primarily associated with rearranging RNA structures activities, mostly by unwinding double-stranded RNAs (dsRNAs) into single-stranded RNAs (ssRNAs) (i.e., hairpin winding/unwinding). The combined activities of annealing and unwinding of RNAs promote the exchange of RNA strands (strand switching). These complex processes rely on ATP hydrolysis, not solely for structural rearrangement, but also for facilitating the association of the helicase with the RNA substrate. Accordingly, the schematic figure of the DExD/H box helicases provides insights into the dynamic mechanisms by which RNA helicases remodel RNA structures, highlighting the dual functions of unwinding and annealing, as well as the vital process of ATP hydrolysis in helicase-RNA substrate interactions.
Figure 2.
RNA helicases as mediators of RNA structure remodeling. RNA helicases are primarily associated with rearranging RNA structures activities, mostly by unwinding double-stranded RNAs (dsRNAs) into single-stranded RNAs (ssRNAs) (i.e., hairpin winding/unwinding). The combined activities of annealing and unwinding of RNAs promote the exchange of RNA strands (strand switching). These complex processes rely on ATP hydrolysis, not solely for structural rearrangement, but also for facilitating the association of the helicase with the RNA substrate. Accordingly, the schematic figure of the DExD/H box helicases provides insights into the dynamic mechanisms by which RNA helicases remodel RNA structures, highlighting the dual functions of unwinding and annealing, as well as the vital process of ATP hydrolysis in helicase-RNA substrate interactions.
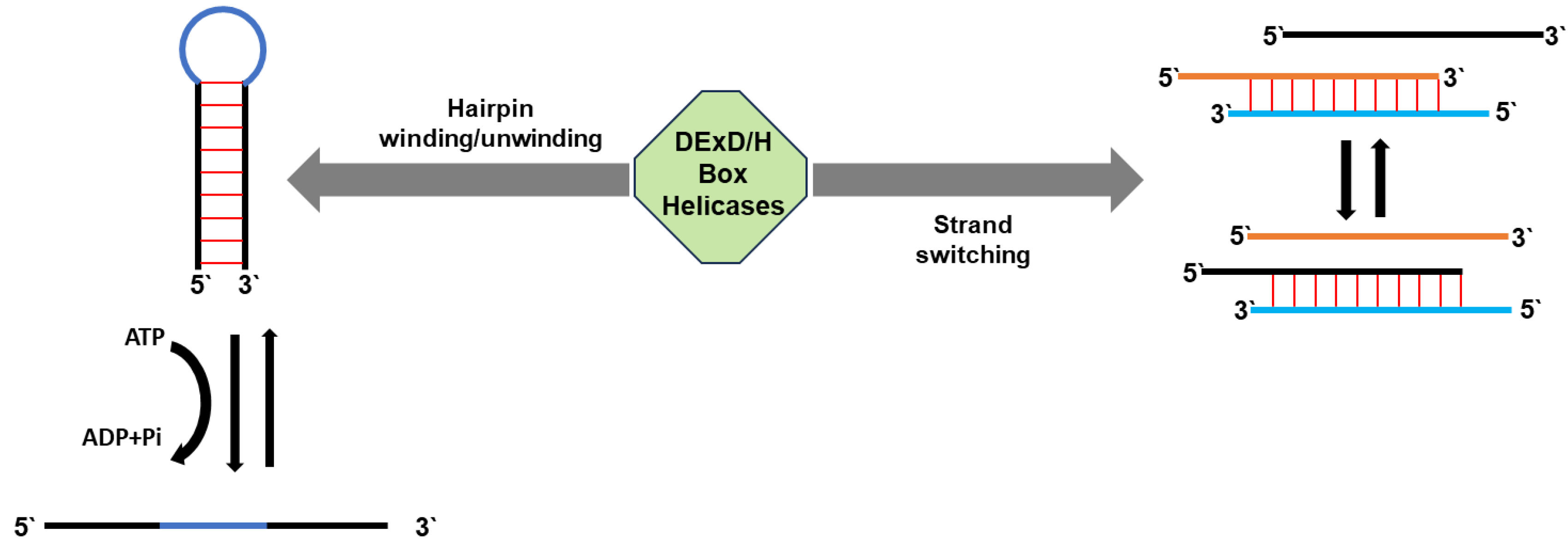
Figure 3.
A phylogenetic analysis of the DExD/H box RNA helicases in Arabidopsis thaliana. The evolutionary relationships among different DExD/box helicases are illustrates by a phylogenetic tree. The tree was constructed by using the ‘Multiple Sequence Comparison by Log-Expectation’ (MUSCLE), based on the deduced protein sequences of homologous genes in Arabidopsis thaliana. The ClustalW algorithm was used for the alignment of the proteins and the tree was built using a neighbor-joining algorithm based on this alignment. The three RNA helicase groups, DExH, DEAD and DEAH box, are indicated in the figure. The tree distances represent the degree of divergence in the amino acid sequence between each branch in the tree. The predicted cellular localization of the gene products, as assigned by the ‘subcellular localization database for Arabidopsis proteins’ (SUBA5) database [57], are indicated. Blue, for nuclear proteins, yellow for cytosolic, red for mitochondria, and green for plastid/chloroplast proteins.
Figure 3.
A phylogenetic analysis of the DExD/H box RNA helicases in Arabidopsis thaliana. The evolutionary relationships among different DExD/box helicases are illustrates by a phylogenetic tree. The tree was constructed by using the ‘Multiple Sequence Comparison by Log-Expectation’ (MUSCLE), based on the deduced protein sequences of homologous genes in Arabidopsis thaliana. The ClustalW algorithm was used for the alignment of the proteins and the tree was built using a neighbor-joining algorithm based on this alignment. The three RNA helicase groups, DExH, DEAD and DEAH box, are indicated in the figure. The tree distances represent the degree of divergence in the amino acid sequence between each branch in the tree. The predicted cellular localization of the gene products, as assigned by the ‘subcellular localization database for Arabidopsis proteins’ (SUBA5) database [57], are indicated. Blue, for nuclear proteins, yellow for cytosolic, red for mitochondria, and green for plastid/chloroplast proteins.
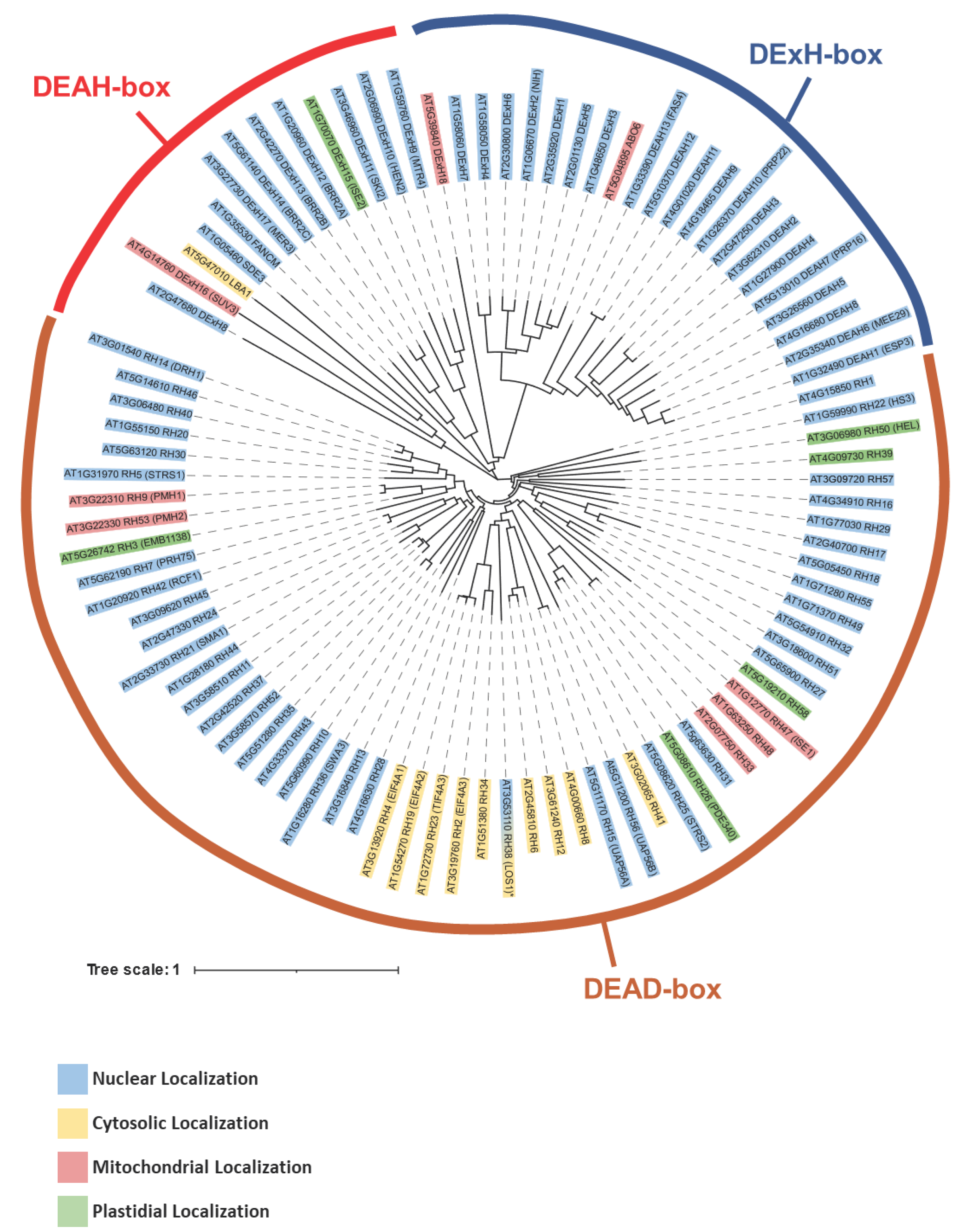

Table 1.
Mitochondrial RNA helicases in fungi and plants.
| RNA helicase | Gene I.D. | Organism*1 | Specific role(s) in mt-RNA metabolism | REFs |
|---|---|---|---|---|
| IRC3 | S000002740 | S.c. | mtDNA maintenance or stability | [43] |
| MRH4 | S000003032 | S.c. | Translation, ribosome biogenesis | [44] |
| MSS116 | S000002602 | S.c. | Splicing (group I and II introns) | [45] |
| SUV3 | S000005950 | S.c. | Nucleolysis (degredosomal factor) | [46,47] |
| ABO6 | At5g04895 | A.t. | Splicing (group II introns) | [48] |
| ISE1/EMB1586 | At1g12770 | A.t. | T.B.D*2., affects mitochondria biogenesis | [49] |
| DExH8 | At5g39640 | A.t. | T.B.D., predicted to the mitochondria*3 | --- |
| PMH1 | At3g22310 | A.t. | T.B.D., found in large RNP complexes*4 | [50] |
| PMH2 | At3g22330 | A.t. | Splicing (group II introns) | [35,50,51] |
| RH33 | At2g07750 | A.t. | Splicing (group II introns) | This study |
| RH48 | At1g63250 | A.t. | T.B.D., homolog of Zm-RH48 | --- |
| RH48 | GRMZM2G171801 | Z.m. | Splicing (group II introns) | [52] |
| SUV3 (At) | At4g14790 | A.t. | T.B.D., RNA metabolisms | [53] |
| SUV3 (Os) | GQ982584 | O.s. | T.B.D., DNA/RNA metabolisms | [54,55] |
| RECG1 | At2g01440 | A.t. | T.B.D., mtDNA and cpDNA maintenance | [56] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



