
Submitted:
15 March 2024
Posted:
15 March 2024
You are already at the latest version
Abstract
Rheumatoid arthritis (RA) is a chronic inflammatory disease. Despite new methods of diagnostics and treatment as well as extensive biological and immunosuppressive treatment, the etiology of RA is not fully understood. Moreover, the problem of diagnosis and treatment of RA patients is still current and affects a large group of patients. It is suggested that these ER-related features may impair adaptation to chronic stress, inferring the risk of rheumatoid arthritis. The main goal in this study was evaluation of changes in mRNA translation to determine chronic ER stress conditions in rheumatoid arthritis patients. The study group consist of 86 individuals including total 56 rheumatoid arthritis patients and 30 healthy controls. The expression level of mRNA form blood samples of RA patients as well as controls of the PERK-UPR associated genes: p-eIF2, BCL-2, PERK, ATF4, BAX were investigated by Real-Time qPCR. GAPDH expression was used as a standard control. Considering the median, the expression of PERK, BCL-2, p-eIF2, ATF4, BAX was found to be significantly increased in the blood of RA patients compared with the control group. The p-value for the PERK gene was 0.0000000036, the p-value for the BCL-2 gene was 0.000000014, the p-value for the p-eIF2 gene was 0.006948, the p-value for the ATF4 gene was 0.0000056 and the p-value for the BAX gene was 0.00019, respectively. Thus, it can be concluded that targeting of the components of the PERK-dependent UPR signaling pathway via small-molecule PERK inhibitors, may contribute to the development of novel, innovative treatment strategies against rheumatoid arthritis.
Keywords:
endoplasmic reticulum stress
; unfolded protein response
; ER-adaptosome
; PERK inhibitor
; rheumatoid arthritis.
1. Introduction
Rheumatoid arthritis (RA) is a systemic condition affecting the joints and their surrounding tissues. RA presents a significant medical concern, and, to date, constitutes a major therapeutic challenge. Approximately, 1% of the global population suffer from RA, making it the most prevalent chronic autoimmune disorder [1]. Typically, the age of initial symptoms is estimated between 35 to 60. It is characterized by severe joint pain, stiffness, and swelling, and if not treated effectively, it can lead to disability, chronic pain, and distress. Proper treatment for RA alleviates pain, enhances functionality, and improves the overall quality of life. However, a substantial number of RA patients do not achieve remission [2]. The exact cause of RA remains unknown, with the primary risk factors encompassing genetics, environmental factors, lifestyle, and hormonal influences. Recent scientific reports indicate that Endoplasmic Reticulum (ER) stress constitutes a significant etiological factor in various human diseases, including conditions associated with the development of inflammation and oxidative stress [3]. Specifically examining the PERK (protein kinase RNA-like ER kinase) cellular signaling pathway, there is promising potential for understanding the pathological mechanisms related to rheumatoid arthritis. Specifically examining the PERK cellular signaling pathway, there is promising potential for understanding the pathological mechanisms related to the mentioned health condition.
It is known that rheumatoid arthritis is a chronic inflammatory disease characterized by abnormal proliferation of synoviocytes, leukocyte infiltration, and angiogenesis. Despite new methods of diagnostics and treatment as well as extensive biological and immunosuppressive treatment, which significantly slow down the course of the disease, the problem of diagnosis and treatment of RA patients is still current and affects a large group of patients. The etiology of RA is not fully understood. It has been suggested that its development is influenced by autoimmune, environmental, and genetic factors.
The endoplasmic reticulum (ER) is the site of biosynthesis for all secreted and membrane proteins. The accumulation of unfolded proteins leads to a condition of ER stress. Eukaryotic cells respond to the accumulation of unfolded proteins in the endoplasmic reticulum by activating the unfolded protein response (UPR) [4]. One of the transducers of the mammalian UPR is PERK kinase, which upon ER stress causes the global attenuation of protein synthesis mediated by the phosphorylation of eIF2α. However, protein synthesis largely recovers while stress ensues, indicating an adpatation process. A group of genes, termed the ER-adaptosome, was induced transcriptionally and escaped translation repression under chronic ER stress conditions [5]. Eukaryotic cells with developed ER, express different ER-translated mRNAs than normal cells [6]. It is suggested that these ER-related features may impair adaptation to chronic stress, inferring the risk of rheumatoid arthritis (RA). Thus, we are going to applied Real-Time qPCR expression of mRNA (the genes of the PERK-UPR) to determine global changes in mRNA translation specific for chronic ER stress conditions in rheumatoid arthritis patients.
2. Results
2.1. Expression of Endoplasmic Reticulum Stress Genes
Expression analysis included 56 blood samples from the rheumatoid arthritis (RA) group and 30 blood samples from the control group. Gene expression analysis was performed for six genes, including the endogenous control gene GAPDH and five test genes PERK, BCL-2, p-eIF2, ATF4, BAX. Considering the median, the expression of ER stress genes (PERK, BCL-2, p-eIF2, ATF4, BAX) was found to be significantly increased in the blood of RA patients compared with the control group, the results were presented in Figure 1. The p-value for the PERK gene was 0.0000000036, the p-value for the BCL-2 gene was 0.000000014, the p-value for the p-eIF2 gene was 0.006948, the p-value for the ATF4 gene was 0.0000056 and the p-value for the BAX gene was 0.00019, respectively.
Additionally, expression was analyzed in blood samples from groups of RA patients and control groups divided by patient gender. In the female group, the expression of ER stress genes (PERK, BCL-2, p-eIF2, ATF4, BAX) was found to be significantly increased in the blood of RA patients compared with the control group. The results were shown in Figure 2. The p-value for the PERK gene was 0.000001026, the p-value for the BCL-2 gene was 0.000004234, the p-value for the p-eIF2 gene was 0.02843, the p-value for the ATF4 gene was 0.00002514 and the p-value for the BAX gene was 0.002822, respectively.
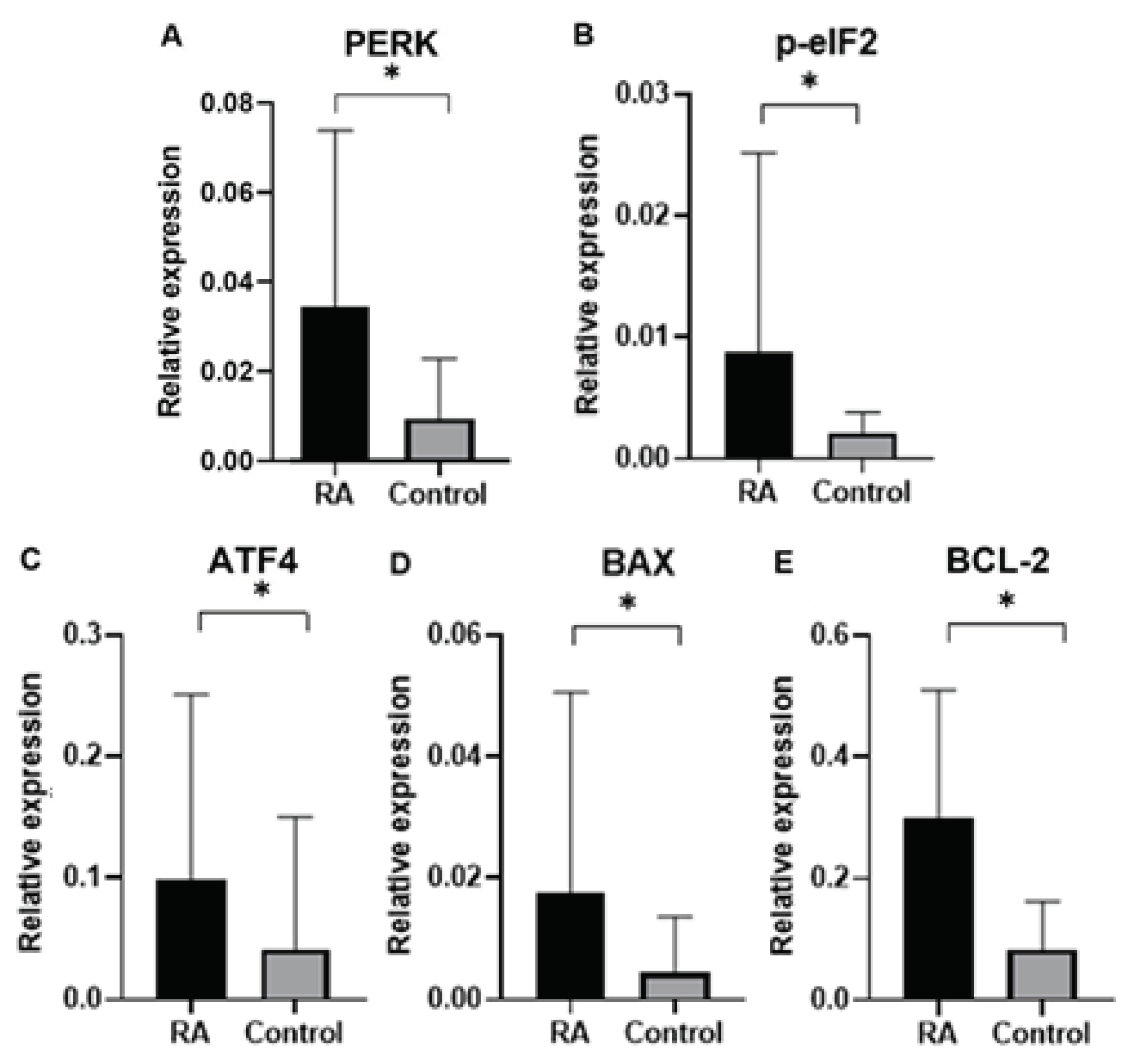
In the male group, the expression of ER stress genes (PERK, BCL-2, ATF4, BAX) in the blood of RA patients was significantly increased compared with the control group. The p-value for the PERK gene is 0.0004593. The p-value for the BCL-2 gene is 0.00009458. The p-value for the ATF4 gene is 0.02951. The p-value for the BAX gene is 0.01845. However, no significant statistical differences were observed for the p-eIF2 gene (p value 0.05535). The results were shown in Figure 3.
Analysis of PERK, BCL-2, p-eIF2, ATF4, and BAX gene expression between the female and male groups with RA showed no statistically significant differences. The p-value for the PERK gene was 0.5182.The p-value for the BCL-2 gene was 0.6165, the p-value for the ATF4 gene was 0.2705, the p-value for the BAX gene was 0.3483 and the p-value for the p-eIF2 gene was 0.5662, respectively.
3. Discussion
Rheumatoid arthritis (RA) is a chronic autoimmune inflammatory disease of the joints of unknown etiopathogenesis, manifested by an inflammatory process mainly in the small metacarpophalangeal and metatarsophalangeal joints, as well as in the wrists, leading to deformation and permanent damage to the joints, as well as to simultaneous multiorgan changes. Despite new methods of diagnostics and treatment as well as extensive biological and immunosuppressive treatment, which significantly slow down the course of the disease, the problem of diagnosis and treatment of RA patients is still current and affects a large group of patients [7].
RA is the most common inflammatory joint disease, affecting approximately 0.3-1.5% of the population. The incidence is 0.1-0.5% in adults. The peak incidence occurs in the 4th and 5th decade of life. Women get sick 3 times more often than men, and the age of onset for women is usually over 50. Burdensome family history increases the risk of developing the disease many times over. The attempt to classify RA is made using the ACR / EULAR 2010 criteria. The quality of life in RA patients is assessed using the AIMS-2 scale [8].
Despite numerous studies, the etiology of RA is not fully understood. It has been suggested that its development is influenced by genetic, autoimmune, and environmental factors. The heredity of rheumatoid arthritis is estimated to be 66%. There is an increased risk of developing rheumatoid arthritis in first-degree relatives. The presence of the disease in a parent increases the risk of its development in a child 2-5 times [7]. Genome sequencing revealed the association of many genes with an increased risk of disease development [8].
Rheumatoid arthritis, due to the course of the disease and its scope, as well as treatment costs, is a social disease that requires constant introduction of new therapies tailored to the patient’s needs. Due to the large group of disabled and invalids, there is a need to assess the accuracy of diagnostics and personalized treatment with the use of new targeted molecular drugs. To better understand the etiopathogenesis of the disease and act on the initiating factor of the avalanche of the progressive RA process, it is necessary to continue research at the molecular level so that the applied therapy protects the patient against the advanced effects of the disease [1-3abc].
Most secretory and transmembrane proteins fold and mature in the endoplasmic reticulum (ER). The flux of proteins entering the ER is dynamic and regulated. In demanding states, the protein load in the ER is increased and must be met by the organelle folding capacity. Adaptation to the load requires quality control mechanisms which monitor the levels of unfolded proteins and prevent their accumulation for risks of aggregation. Once these quality control mechanisms are compromised, or when cells undergo an insult that changes the physiology of the secretory pathway, such as viral infection or in response to hypoxia [9,10], the balance between folded and unfolded proteins in the ER is tipped, resulting in the accumulation of misfolded proteins, a state referred to as ER stress.
Eukaryotic cells respond to ER stress by activating a signaling pathway coined the Unfolded Protein Response (UPR). The UPR is a collection of signaling pathways which can resolve ER stress by integrating mRNA translation control with the regulation of gene transcription. If ER stress persists despite the activation of these feedback, the UPR will initiate apoptosis [4]. In mammalian cells the UPR is comprised of three major branches: inositol requiring enzyme 1 (IRE1); double-stranded RNA-activated protein kinase (PKR)- like ER kinase (PERK) and activating transcription factor 6 (ATF6), each termed after the ER-transmembrane sensors which gauge the levels of misfolded proteins in the ER lumen and consequently activate their respective downstream signaling cascades. Briefly, upon ER stress, the ATF6 transmembrane sensor travels from the ER to the Golgi, where it is cleaved in a manner that liberates the N-terminus domain of ATF6 (ATF6(N)). ATF6(N) translocates to the nucleus and functions as a transcription factor. PERK is activated by oligomerization, and once activated, phosphorylates the translation initiation factor eIF2α. This reduces translation initiation, leading to a global decrease in protein synthesis. Paradoxically, eIF2α phosphorylation increases synthesis of select transcripts some of which contain short overlapping open reading frames in their 5′UTR, such as ATF4, a transcription factor that coordinates transcription of genes that determine cell fate following ER stress [5]. The third UPR sensor, IRE1, is kinase and endonuclease. Once activated it splices the mRNA of the transcription factor XBP1, excising a 26-nucleotide intron. This non-canonical splicing causes a shift in the reading frame, yielding the spliced form of XBP1 (XBP1s) [11,12]. XBP1s is a highly potent transcription factor that promotes the level of a large variety of ER chaperones and induces expansion of the ER [13].
In addition to its roles in development and cell function, the UPR modulates prominent diseases, such as diabetes, liver steatosis, inflammatory bowel disease, cancers and more [14,15,16,17]. A role for the ER stress in rheumatoid arthritis was initially suggested [18,19]. However, the involvement of the UPR in RA patients has turned out to be much more general with important contributions to disease initiation, progression, and response to therapy [20,21].
The cellular program controlled by PERK in mouse embryonic fibroblasts (MEFs), represent in normal cells, is widespread and includes many downstream genes regulated by multiple mechanisms [22]. However, PERK affects its targets in a cell type and physiological context-dependent manner [23]. For instance, PERK has been found to be essential to the progression of BRAF-mutated melanoma, less of non- BRAF mutated tumors [24]. PERK regulates cellular redox by directly phosphorylating and activating NRF2 [25,26]. PERK interphases with the circadian oscillations by induction of miRNA that represses major circadian genes in a manner that affects Burkitt lymphoma progression [27]. Hence, it is not surprising that multiple pharmaceutical companies developed high affinity inhibitors of PERK. Glaxo Smith Kline developed GSK2606414 (GSK414); Amgen developed AMG PERK 44; Eli Lilly developed Ly4. In our laboratory we developed specific inhibitors for PERK treatment in neurodegenerative disorders including glaucoma (termed PERKi) [28,29,30,31]. The intricacies of the transcription and translation program controlled by PERK invites research in different tissue types and in combinations with other drugs to assess its role as an efficacious therapeutic target.
Translation repression by PERK in response to chronic ER stress is reversed by adaptation. The phosphorylation of eIF2α by PERK and by additional kinases attenuates translation initiation by the sequestration of the multi- subunit GEF eIF2B. This results in reduction of the ternary complex of translation initiation and leads to a global repression in protein synthesis [32]. Since cells cannot survive under prolonged translation repression, homeostatic mechanisms are engaged to gradually restore protein synthesis during chronic ER stress. One such mechanism, described by Hatzoglou et al, is a conversion from the classical and efficient CAP-dependent mRNA translation, driven by eIF4E, to a less favorable mechanism that relies on recruitment of the ribosome by eIF3 [33]. This adaptation process is dependent on the constant repression of eIF2B activity [34]. Analysis of the stress specific transcriptome and translatome in MEFs subjected to a prolonged ER stress identified a set of 567 genes that were induced at the level of transcription and were translated under the chronic conditions [6]. Remarkably, 35 of these genes encode proteins that function in the ER in protein folding, glycosylation, trafficking and degradation. Genes were indentified under ER stress conditions using thapsigargin (Tg:16h vs. Tg:1h). Because the ER protein processing pathway includes in total approximately 141 genes (according to KEGG, PATHWAY: ko04141 in www.genome.jp), it is of note that a relatively large subset of genes specifically in this category was upregulated during adaptation to ER chronic stress. When PERKi was applied post-establishment of the transcriptional and translational reprogramming, this led to a 50% larger shift in fold changes for ER-translated mRNAs, which was determined by the comparison of polysome-associated to total mRNAs. Consistently, these mRNAs were regulated via both, changes in translation efficiency (61 genes) and mRNA abundance (105 genes). We use the term ER- adaptosome to describe the group of genes congruently induced during chronic ER stress.
The Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analysis revealed that the group of genes congruently up-regulated during chronic ER stress is enriched in those that encode proteins involved in ER functions, including genes listed in reference to PERK-dependent UPR [6]. ER-adaptosome genes (including 35 genes of the ER protein processing pathway) are known targets of UPR-induced transcription factors, including ATF6, which exhibits protective functions during chronic ER stress and is negatively regulated by PERKi [35]. One of these factors, the ER transmembrane glycoprotein wolframin, is a regulator of ER calcium levels, which plays a crucial role in ER homeostasis [36]. Because ATF4 induction requires PERK and is necessary for maximal induction of ATF6 [35], it is suggested that sustained PERK activity during chronic ER stress maintains ER proteostasis in concert with congruently up-regulated genes. This, in combination with the effects on the ER-localized translation, suggests that chronic ER stress is tailored to maintain ER function by coordinating protein load and processing capacity of the ER. Transcripts that are predominantly translated at the ER [37] showed congruent decreases in polysome-association and cytosolic mRNA levels during chronic ER stress and in large part reversed by PERKi. Thus, ER-associated mRNA translation is modulated during chronic ER stress in a PERK-dependent manner.
Finally, in our study we analyzed gene expression of ER proteins processing pathway in blood samples of rheumatoid arthritis (RA) patients in comparison to control group subjects. Gene expression analysis was performed for genes of PERK, BCL-2, p-eIF2, ATF4, BAX as well as endogenous control gene GAPDH. An expression of ER stress genes of PERK, BCL-2, p-eIF2, ATF4, BAX were found to be significantly higher in RA patients than in the control group. Interestingly, ER stress genes of PERK, BCL-2, p-eIF2, ATF4, BAX in patients divided by gender was found higher according to controls in both male and female group, which suggested that UPR-unfolded proteins response is a global process involved in RA pathogenesis. In agreement with the PERK-mediated maintenance of the adaptive state during chronic stress conditions, inclusion of PERKi resulted in the formation of large membrane distensions, associated with perturbation of the ER-adaptosome. These intracellular distensions gradually ballooned, and the foamy cells eventually died in a manner that coincided with the rupture of the vacuoles [27]. However, in agreement with the idea that the intracellular ballooning is a sign of ER dysfunction and not a programmed cell death, was finding that the vacuoles were reversed back to normal ER structures and perfectly functioning cells if the stress was removed shortly after the appearance of ER distension. It is named ER-dysfunction mediated cell death mechanism, Ballooning Endoplasmic Reticulum cell Death (BERD). Thus, how the adaptation to chronic ER stress is modulated in rheumatoid arthritis cells and whether a failure to adapt can be used for BERD-mediated treatment of patients with rheumatoid arthritis should be the subject of future studies, to determine the unfolded protein response pathway as a modulator of rheumatoid arthritis initiation, progression, and therapy.
4. Materials and Methods
4.1. Patients and Study Specimens
The material was collected from a total 56 rheumatoid arthritis patients, including 39 women and 17 men. The average age among women was 73 years, while among men it was 63 years. Study participants were patients with newly diagnosed rheumatoid arthritis according to ACR/EULAR 2010 guidelines. Positive family history of inflammatory joint diseases was present in 21 women and 10 men. The mean erythrocyte sedimentation rate (ESR) before treatment initiation in the female group was 47 mm/h, whereas in the male group it was 64 mm/h. The mean C-reactive protein (CRP) level before treatment initiation in the female group was 31 mg/l, while among men it was 38 mg/l. The Disease Activity Score (DAS28) was also analyzed, which is commonly used in clinical practice to assess disease activity and joint damage. The score is calculated using a calculator that considers: 1) The number of swollen joints; 2) The number of tender joints (28 joints are considered: wrists, metacarpophalangeal, proximal interphalangeal, elbows, shoulders, and knees); 3) ESR or CRP; 4) The patient’s overall assessment of disease activity on a visual analogue scale (VAS, 0–100).
The possible values on this scale range from 0 to 9.4. This scale allows us to assess disease activity as follows: <2.6 indicates remission; ≤3.2 indicates low activity; 3.2 and ≤5.1 indicate moderate activity; 5.1 is interpreted as high activity. This scale also facilitates an objective assessment of treatment response, where: a good response is defined by a change in activity of ≥1.2 and low activity; a moderate response is defined by a change of >0.6 and <1.2 and low or moderate activity, or in the case of a change of ≥1.2 and high or moderate activity; no response is observed when the change is <0.6 or <1.2 and the activity is high.
The mean DAS28 score before treatment initiation in the female group was 5.76, while in the male group it was 6.23. All patients were treated according to current ACR/EULAR recommendations. Methotrexate was the first-choice drug among disease-modifying antirheumatic drugs (DMARDs) at an initial dose of 10 mg/week. Subsequently, the dose was gradually increased by 5 mg every 2 or 4 weeks to a maximum dose of 25-30 mg/week or to a dose tolerated by the patient. In cases where methotrexate was contraindicated or early intolerance occurred (within 6 weeks of starting treatment), leflunomide or sulfasalazine was used as the first choice DMARD (5 women and 1 man). Short-term oral glucocorticoid (methylprednisolone) therapy was administered to 27 women and 16 men during the initiation or modification of DMARD therapy at various doses, but they were tapered as quickly as the clinical condition of the patients allowed. The mean DAS28 score after 3 months of treatment initiation was 2.69 among women and 2.58 among men, indicating achievement of remission. After treatment initiation, a significant decrease in inflammatory parameters was also observed. In the female group, the mean ESR was 19 mm/h, while in the male group it was 12 mm/h. The mean CRP level among women was 6 mg/l, while among men it was 4 mg/l.

Table 1.
Clinical characteristic of Rheumatoid Arthritis patients.
| Category | Mean value | |
| Number of patients (n) | 56 | |
| Number of patients treated with methotrexate (MTX) (n) | 50 | |
| Positive family history (n) | 31 | |
| Number of patients bridging with GCS (n) | 43 | |
| Mean ESR before treatment initiation (mm/h) | 55,5 | |
| Mean ESR after treatment initiation (mm/h) | 15,5 | |
| Mean CRP before treatment initiation (mg/l) | 34,5 | |
| Mean CRP after treatment initiation (mg/l) | 5 | |
| Mean DAS-28 before treatment | 5,99 | |
| Mean DAS-28 after treatment | 2,63 |
Overall, the research encompassed a group of 86 individuals. The study group comprised 56 patients with diagnosed Rheumatoid Arthritis selected from patients of the the Vadimed Medical Center in Krakow, Poland. Concurrently, the control group included 30 volunteers selected from healthy subjects admitted to the Department for other reasons not associated with chronic inflammatory, cancer nor neurodegenerative disorders. The control group was matched to the study group regarding sex and age. Blood samples were collected from both groups for gene expression assessment. The study received approval from the Institutional Bioethics Committee (protocol no. 7/KBL/OIL/2022.). All participants provided written informed consent to participate in the study. Prior to commencing the experiments, all participants underwent comprehensive medical examinations.
4.2. RNA Isolation
Total RNA was extracted from whole blood in sterile environment using a commercially available RiboPure™-Blood Kit (Invitrogen™, ThermoFisher Scientific, Waltham, MA, USA) in accordance with the manufacturer’s protocol. The concentration and quality of the extracted RNA were evaluated by spectrophotometric measurement of samples at 260 and 280 nm using a Multiskan SkyHigh Microplate Spectrophotometer (Thermo Scientific™, ThermoFisher Scientific, Waltham, MA, USA).
4.3. Generation of Single-Stranded cDNA
Obtained RNA was used for the quantitative conversion of 100 ng of total RNA into cDNA in single 10 μl reaction using the High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems™, ThermoFisher Scientific, Waltham, MA, USA) in accordance with the manufacturer’s instructions. To perform reverse transcription the thermal cycler VeritiPro (Applied Biosystems™, ThermoFisher Scientific, Waltham, MA, USA) was used using the conditions suggested by the manufacturer.
4.4. Quantitative Real-Time PCR (qPCR)
The expression of genes associated with selected PERK signaling pathway in the study and control groups was evaluated by TaqMan technique. For qPCR reactions, 10ng of generated cDNA was used for analysis on the CFX Connect Real-Time PCR Detection System (Bio-Rad, Hercules, CA, USA). Quantification of mRNA expression was performed using the TaqMan™ gene expression assays including TaqMan Universal PCR Master Mix (Applied Biosystems™, ThermoFisher Scientific, Waltham, MA, USA) with the predeveloped TaqMan assays for EIF2A (Assay ID HS00230684_m1), BCL-2 (Assay ID HS00708019_s1), PERK (Assay ID HS00984003_n1), p-eIF2 (Assay ID Hs00909569_g1), BAX (Assay ID Hs00180269_m1) and GAPDH (Assay ID HS02786624_g1). GAPDH expression was used as an endogenous standard. The targeted transcripts were run in triplicate. Real-Time PCR conditions were as universal cycling conditions in accordance with standard protocol. The quantities of selected genes in relation to the housekeeping gene were assessed using the comparative Ct method by Livak.
4.5. Statistical Analysis of Gene Expression
Statistical analyses were performed using Statistica 13.1 software (StatSoft, Tulsa, OK, USA). To evaluate the significance of expression differences between the study and control groups, a non-parametric test, U Mann-Whitney was used. The distribution of variables was examined using the Shapiro–Wilk test. Statistical significance was considered for p-values less than 0.05.
5. Conclusions
Rheumatoid arthritis is a chronic inflammatory disease. Despite new methods of diagnostics and treatment as well as extensive biological and immunosuppressive treatment, the etiology of RA is not fully understood. Moreover, the problem of diagnosis and treatment of RA patients is still current and affects a large group of patients. Thus, based on the latest literature data and results obtained from the present study, gene expression profiling to unfolded proteins response can be considered as a risk modulator of patients with rheumatoid arthritis. Finally, it can be concluded that targeting of the components of the PERK-dependent UPR signaling pathway via small-molecule PERK inhibitors, may contribute to the development of novel, innovative treatment strategies against rheumatoid arthritis.
Author Contributions
Conceptualization, I.M.; methodology, A.K-L., M.S., A.B. and I.M.; formal analysis, A.K-L., M.S. and I.M.; investigation, A.K-L., M.S. and A.B.; resources, A.K-L and I.M.; writing-original draft preparation, A.L-K., M.S. and I.M.; writing-review and editing, M.S. and I.M.; visualization, A.K-L and M.S; supervision, I.M.; project administration, I.M.; funding acquisition, A.K-L. and I.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by grant from Medical University of Lodz, Poland no. 503/1-156-07/503-11-001.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Radu, A.F.; Bungau, S.G. Management of Rheumatoid Arthritis: An Overview. Cells. 2021, 10, 2857. [Google Scholar] [CrossRef]
- Zhao, J.; Guo, S.; et al. Molecular and Cellular Heterogeneity in Rheumatoid Arthritis: Mechanisms and Clinical Implications. Front Immunol. 2021, 12, 790122. [Google Scholar] [CrossRef] [PubMed]
- Mollereau, B.; Manié, S.; Napoletano, F. Getting the better of ER stress. J Cell Commun Signal. 2014, 8, 311–21. [Google Scholar] [CrossRef] [PubMed]
- Ron, D.; Walter, P. Signal integration in the endoplasmic reticulum unfolded protein response. Nat Rev Mol Cell Biol. 2007, 8, 519–529. [Google Scholar] [CrossRef] [PubMed]
- Harding, H.P.; Novoa, I.; Zhang, Y.; Zeng, H.; Wek, R.; Schapira, M.; Ron, D. Regulated translation initiation controls stress-induced gene expression in mammalian cells. Molecular cell. 2000, 6, 1099–1108. [Google Scholar] [CrossRef] [PubMed]
- Guan, B.J.; van Hoef, V.; Jobava, R.; Elroy-Stein, O.; Valasek, L.S.; Cargnello, M.; Gao, X.H.; Krokowski, D.; Merrick, W.C.; Kimball, S.R.; et al. A Unique ISR Program Determines Cellular Responses to Chronic Stress. Mol Cell 2017, 68, 885–900. [Google Scholar] [CrossRef] [PubMed]
- Puszczewicz, M.; Grygiel-Górniak, B. Diagnostyka reumatoidalnego zapalenia stawów. Przegląd Reumatologiczny; 2018.
- Scherer, H.U.; Häupl, T.; Burmester, G.R. The etiology of rheumatoid arthritis. J Autoimmun 2020, 110, 102400. [Google Scholar] [CrossRef]
- He, B. Viruses, endoplasmic reticulum stress, and interferon responses. Cell death and differentiation. 2006, 13, 393–403. [Google Scholar] [CrossRef]
- Feldman, D.E.; Chauhan, V.; Koong, A.C. The unfolded protein response: a novel component of the hypoxic stress response in tumors. Molecular cancer research. MCR 2005, 3, 597–605. [Google Scholar] [CrossRef]
- Calfon, M.; Zeng, H.; Urano, F.; Till, J.H.; Hubbard, S.R.; Harding, H.P.; Clark, S.G.; Ron, D. IRE1 couples endoplasmic reticulum load to secretory capacity by processing the XBP-1 mRNA. Nature. 2002, 415, 92–96. [Google Scholar] [CrossRef]
- Yoshida, H.; Matsui, T.; Yamamoto, A.; Okada, T.; Mori, K. XBP1 mRNA is induced by ATF6 and spliced by IRE1 in response to ER stress to produce a highly active transcription factor. Cell. 2001, 107, 881–891. [Google Scholar] [CrossRef]
- Lee, A.H.; Iwakoshi, N.N.; Glimcher, L.H. XBP-1 regulates a subset of endoplasmic reticulum resident chaperone genes in the unfolded protein response. Molecular and cellular biology. 2003, 23, 7448–7459. [Google Scholar] [CrossRef]
- Grootjans, J.; Kaser, A.; Kaufman, R.J.; Blumberg, R.S. The unfolded protein response in immunity and inflammation. Nat Rev Immunol. 2016, 16, 469–484. [Google Scholar] [CrossRef]
- Adolph, T.E.; Niederreiter, L.; Blumberg, R.S.; Kaser, A. Endoplasmic reticulum stress and inflammation. Dig Dis. 2012, 30, 341–346. [Google Scholar] [CrossRef]
- Han, J.; Kaufman, R.J. The role of ER stress in lipid metabolism and lipotoxicity. J Lipid Res. 2016, 57, 1329–1338. [Google Scholar] [CrossRef]
- Avril, T.; Vauleon, E.; Chevet, E. Endoplasmic reticulum stress signaling and chemotherapy resistance in solid cancers. Oncogenesis. 2017, 6, e373. [Google Scholar] [CrossRef]
- Rahmati, M.; Moosavi, M.A.; McDermott, M.F. ER Stress: A Therapeutic Target in Rheumatoid Arthritis? Trends Pharmacol Sci. 2018, 39, 610–623. [Google Scholar] [CrossRef]
- Park, Y.J.; Yoo, S.A.; Kim, W.U.J. Role of endoplasmic reticulum stress in rheumatoid arthritis pathogenesis. Korean Med Sci. 2014, 29, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.; Wang, J.; Zhang, X.; Tian, R.; Qiao, L.; Ge, L.; Pan, J.; Wang, L. TXNDC5 protects synovial fibroblasts of rheumatoid arthritis from the detrimental effects of endoplasmic reticulum stress. Intractable Rare Dis Res. 2020, 9, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Hu, F.; Tang, Y.; Wang, P.; Bai, M.; Li, X.; Song, J.; Shi, L.; Sun, X.; Jin, M.; Li, Z.; Li, X. Endoplasmic reticulum stress perpetuated toll-like receptor signalling-mediated inflammation in rheumatoid arthritis via X- box-binding protein-1. Clin Exp Rheumatol. 2021, 39, 859–867. [Google Scholar] [CrossRef] [PubMed]
- Gonen, N.; Sabath, N.; Burge, C.B.; Shalgi, R. Widespread PERK-dependent repression of ER targets in response to ER stress. Sci Rep. 2019, 9, 4330. [Google Scholar] [CrossRef]
- Adamson, B.; Norman, T.M.; Jost, M.; Cho, M.Y.; Nunez, J.K.; Chen, Y.; Villalta, J.E.; Gilbert, L.A.; Horlbeck, M.A.; Hein, M.Y.; et al. A Multiplexed Single-Cell CRISPR Screening Platform Enables Systematic Dissection of the Unfolded Protein Response. Cell 2016, 167, 1867–1882. [Google Scholar] [CrossRef]
- Pytel, D.; Gao, Y.; Mackiewicz, K.; Katlinskaya, Y.V.; Staschke, K.A.; Paredes, M.C.; Yoshida, A.; Qie, S.; Zhang, G.; Chajewski, O.S.; Wu, L.; Majsterek, I.; Herlyn, M.; Fuchs, S.Y.; Diehl, J.A. PERK Is a Haploinsufficient Tumor Suppressor: Gene Dose Determines Tumor- Suppressive Versus Tumor Promoting Properties of PERK in Melanoma. PLoS Genet. 2016, 12, e1006518. [Google Scholar] [CrossRef] [PubMed]
- Cullinan, S.B.; Diehl, J.A. PERK-dependent activation of Nrf2 contributes to redox homeostasis and cell survival following endoplasmic reticulum stress. The Journal of biological chemistry. 2004, 279, 20108–20117. [Google Scholar] [CrossRef] [PubMed]
- Bobrovnikova-Marjon, E.; Grigoriadou, C.; Pytel, D.; Zhang, F.; Ye, J.; Koumenis, C.; Cavener, D.; Diehl, J.A. PERK promotes cancer cell proliferation and tumor growth by limiting oxidative DNA damage. Oncogene. 2010, 29, 3881–3895. [Google Scholar] [CrossRef]
- Bu, Y.; Yoshida, A.; Chitnis, N.; Altman, B.J.; Tameire, F.; Oran, A.; Gennaro, V.; Armeson, K.E.; McMahon, S.B.; Wertheim, G.B.; et al. A PERK-miR-211 axis suppresses circadian regulators and protein synthesis to promote cancer cell survival. Nat Cell Biol. 2018, 20, 104–115. [Google Scholar] [CrossRef] [PubMed]
- Rozpędek-Kamińska, W.; Galita, G.; Saramowicz, K.; Granek, Z.; Barczuk, J.; Siwecka, N.; Pytel, D.; Majsterek, I. Evaluation of the LDN-0060609 PERK Inhibitor as a Selective Treatment for Primary Open-Angle Glaucoma: An In Vitro Study on Human Retinal Astrocytes. Int J Mol Sci. 2014, 25, 728. [Google Scholar] [CrossRef] [PubMed]
- Rozpędek, W.; Pytel, D.; Popławski, T.; Walczak, A.; Gradzik, K.; Wawrzynkiewicz, A.; Wojtczak, R.; Mucha, B.; Diehl, J.A.; Majsterek, I. Inhibition of the PERK-Dependent Unfolded Protein Response Signaling Pathway Involved in the Pathogenesis of Alzheimer’s Disease. Curr Alzheimer Res. 2019, 16, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Rozpędek-Kamińska, W.; Galita, G.; Siwecka, N.; Carroll, S.L.; Diehl, J.A.; Kucharska, E.; Pytel, D.; Majsterek, I. The Potential Role of Small-Molecule PERK Inhibitor LDN-0060609 in Primary Open-Angle Glaucoma Treatment. Int J Mol Sci. 2021, 22, 4494. [Google Scholar] [CrossRef] [PubMed]
- Lusa, W.; Rozpędek-Kamińska, W.; Siwecka, N.; Galita, G.; Majsterek, I.; Kucharska, E. Small molecule PKR like endoplasmic reticulum kinase inhibitors as a novel targeted therapy for Parkinson’s disease. Mol Med Rep. 2023, 27, 102. [Google Scholar] [CrossRef]
- Krishnamoorthy, T.; Pavitt, G.D.; Zhang, F.; Dever, T.E.; Hinnebusch, A.G. Tight binding of the p3osphorylated alpha subunit of initiation factor 2 (eIF2alpha) to the regulatory subunits of guanine nucleotide exchange factor eIF2B is required for inhibition of translation initiation. Molecular and cellular biology. 2001, 21, 5018–5030. [Google Scholar] [CrossRef]
- Komar, A.A.; Hatzoglou, M. Exploring Internal Ribosome Entry Sites as Therapeutic Targets. Front Oncol. 2015, 5, 233. [Google Scholar] [CrossRef] [PubMed]
- Guan, B.J.; Krokowski, D.; Majumder, M.; Schmotzer, C.L.; Kimball, S.R.; Merrick, W.C.; Koromilas, A.E.; Hatzoglou, M. Translational control during endoplasmic reticulum stress beyond phosphorylation of the translation initiation factor eIF2α. J Biol Chem. 2014, 289, 12593–611. [Google Scholar] [CrossRef] [PubMed]
- Teske, B.F.; Wek, S.A.; Bunpo, P.; Cundiff, J.K.; McClintick, J.N.; Anthony, T.G.; Wek, R.C. The eIF2 kinase PERK and the integrated stress response facilitate activation of ATF6 during endoplasmic reticulum stress. Molecular biology of the cell. 2011, 22, 4390–4405. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, S.G.; Fukuma, M.; Lipson, K.L.; Nguyen, L.X.; Allen, J.R.; Oka, Y.; Urano, F. WFS1 is a novel component of the unfolded protein response and maintains homeostasis of the endoplasmic reticulum in pancreatic beta- cells. The Journal of biological chemistry. 2005, 280, 39609–39615. [Google Scholar] [CrossRef]
- Reid, D.W.; Chen, Q.; Tay, A.S.; Shenolikar, S.; Nicchitta, C.V. The unfolded protein response triggers selective mRNA release from the endoplasmic reticulum. Cell 2014, 158, 1362–1374. [Google Scholar] [CrossRef]
Figure 1.
Relative expression of ER stress genes in the blood of RA patients and controls: A. PERK gene expression in patients and healthy groups (*p<0.05); B. p-eIF2 gene expression in patients and healthy groups (*p<0.05); C. ATF4 gene expression in patients and healthy groups (*p<0.05); D. BAX gene expression in patients and healthy groups (*p<0.05); E. BCL-2 gene expression in patients and healthy groups (*p<0.05).
Figure 1.
Relative expression of ER stress genes in the blood of RA patients and controls: A. PERK gene expression in patients and healthy groups (*p<0.05); B. p-eIF2 gene expression in patients and healthy groups (*p<0.05); C. ATF4 gene expression in patients and healthy groups (*p<0.05); D. BAX gene expression in patients and healthy groups (*p<0.05); E. BCL-2 gene expression in patients and healthy groups (*p<0.05).
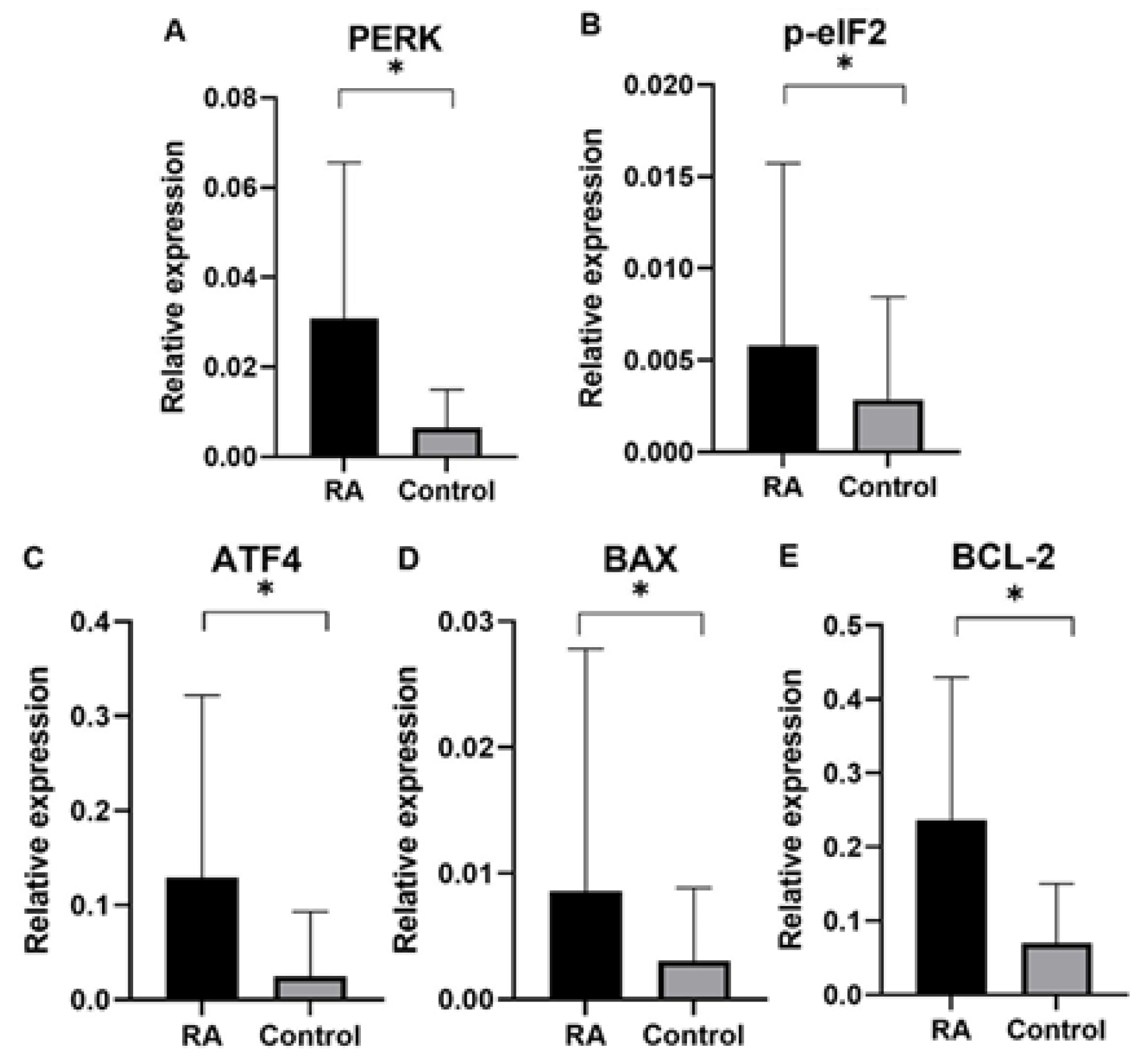
Figure 2.
Relative expression of ER stress genes in the blood of RA patients and controls in the female group: A. PERK gene expression in patients and healthy groups (*p<0.05); B. p-eIF2 gene expression in patients and healthy groups (*p<0.05); C. ATF4 gene expression in patients and healthy groups (*p<0.05); D. BAX gene expression in patients and healthy groups (*p<0.05); E. BCL-2 gene expression in patients and healthy groups (*p<0.05).
Figure 2.
Relative expression of ER stress genes in the blood of RA patients and controls in the female group: A. PERK gene expression in patients and healthy groups (*p<0.05); B. p-eIF2 gene expression in patients and healthy groups (*p<0.05); C. ATF4 gene expression in patients and healthy groups (*p<0.05); D. BAX gene expression in patients and healthy groups (*p<0.05); E. BCL-2 gene expression in patients and healthy groups (*p<0.05).
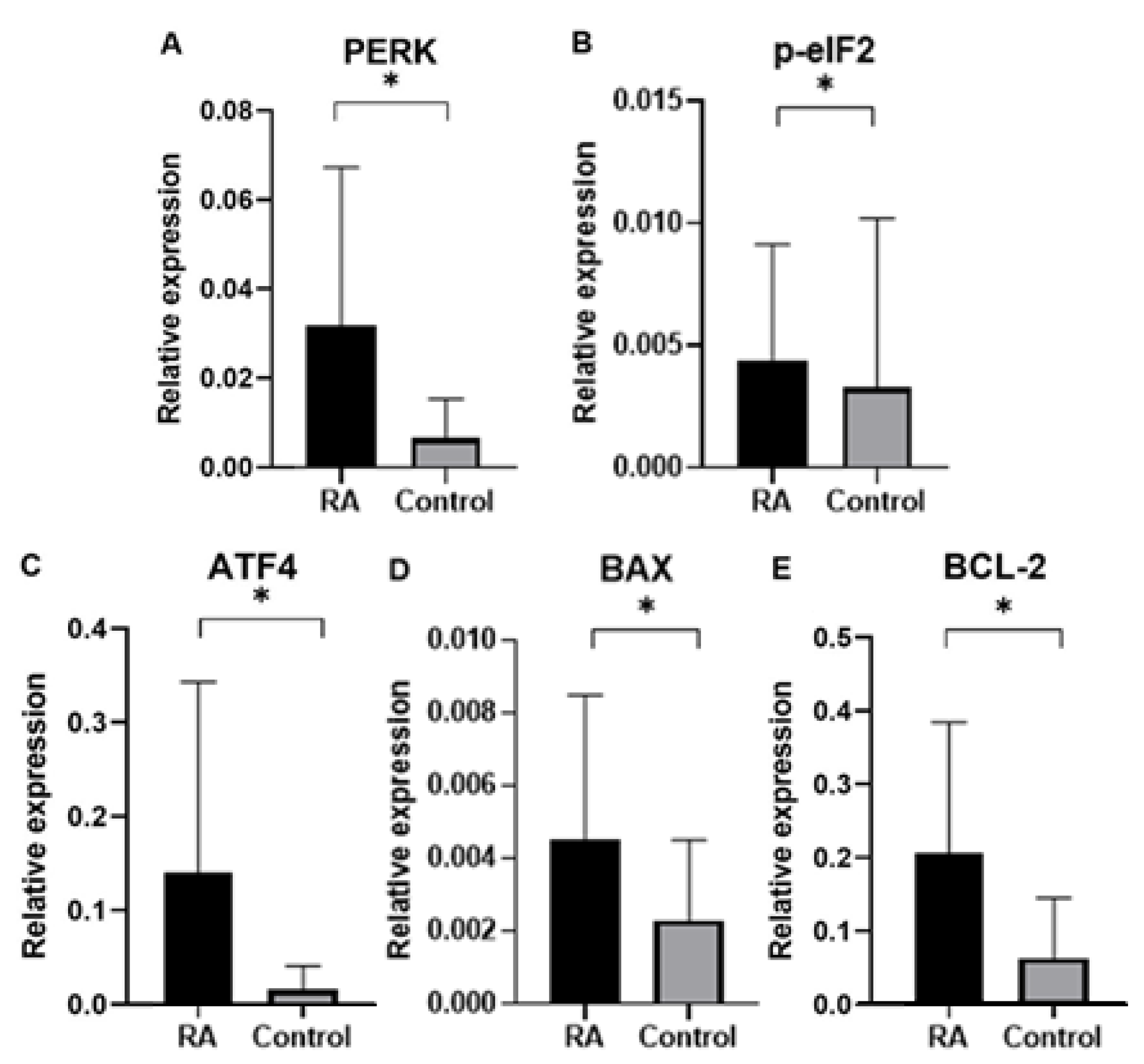
Figure 3.
Relative expression of ER stress genes in the blood of RA patients and controls in the male group: A. PERK gene expression in patients and healthy groups (*p<0.05); B. p-eIF2 gene expression in patients and healthy groups (*p<0.05); C. ATF4 gene expression in patients and healthy groups (*p<0.05); D. BAX gene expression in patients and healthy groups (*p<0.05); E. BCL-2 gene expression in patients and healthy groups (*p<0.05).
Figure 3.
Relative expression of ER stress genes in the blood of RA patients and controls in the male group: A. PERK gene expression in patients and healthy groups (*p<0.05); B. p-eIF2 gene expression in patients and healthy groups (*p<0.05); C. ATF4 gene expression in patients and healthy groups (*p<0.05); D. BAX gene expression in patients and healthy groups (*p<0.05); E. BCL-2 gene expression in patients and healthy groups (*p<0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



