
Submitted:
15 March 2024
Posted:
18 March 2024
You are already at the latest version
Abstract
Merkel cell carcinoma (MCC) is a rare, highly aggressive skin cancer characterized by two etiologies: the first one, observed in most cases, is attributed to Merkel cell polyomavirus (MCPyV), a DNA tumor virus, while the second one is due to ultraviolet (UV) radiation exposure. Currently, there are limited diagnostic, prognostic, and therapeutic biomarkers available for MCC. MicroRNAs (miRNAs) are small RNA molecules that do not code for proteins but regulate various cellular processes, including cell cycle progression, proliferation, differentiation, and apoptosis. The dysregulation of miRNAs has been associated with a wide range of cancers since they can act as either tumor suppressors or oncogenes. This paper aims to collect, summarize and re-organize the recent discoveries concerning miRNAs, whose dysregulation in the pathogenesis and progression of MCC is slowly emerging. Furthermore, the potential clinical applications of miRNAs as diagnostic biomarkers, possible differentiation between MCPyV-related and unrelated MCC, predictors of response to therapy and prognostics are also discussed. From this review, it emerges that miRNAs may have, in the near future, the potential to become clinically significant in improving the diagnostic accuracy, prognostic evaluation, and therapeutic options of MCCs. Finally, the new perspectives on the poorly explored field of artificial intelligence in MCC are provided.
Keywords:
merkel cell carcinoma
; microRNA
; miRNA
; exosomes
; circRNA
; artificial intelligence
; machine learning
; merkel cell polyomavirus
1. Introduction
1.1. Merkel Cell Carcinoma
Merkel cell carcinoma (MCC) is an uncommon and highly aggressive cancer characterized by both epithelial and endocrine features. It accounts for less than 1% of all skin malignancies [1]. The exact origin of MCC remains a matter of debate, with various theories proposing different cell types as potential precursors. These include Merkel cell precursors that may arise from epidermal stem cells or hair follicle stem cells, as well as pre-B cells, pro-B cells, or dermal fibroblast [2]. Risk factors for MCC development predominantly include exposure to ultraviolet (UV) light, advanced age, immune deficiencies, fair skin type, immunosenescence, the concurrent presence of other cancers, and chronic inflammation [1,3]. Two MCC types have been identified and classified based on specific molecular pathogenetic pathways related to mutational burden resulting from UV exposure and infection by the Merkel cell polyomavirus (MCPyV).
1.1.1. Polyomavirus-Related Merkel Cell Carcinoma
The oncogenic potential of MCPyV has been linked to two of its viral proteins: large T-antigen (LTAg) and small T-antigen (STAg), being detected in 80% of MCC samples [4]. In vitro studies have demonstrated their ability to transform cells, while animal studies have shown their capacity to induce tumorigenesis [5]. Other studies highlighted that MCPyV is normally present in dermal fibroblasts and that its viral particles are chronically eliminated, suggesting that direct physical contact could represent the possible mode of transmission [6]. The virus is detected in an episomal state in non-malignant cells, while it is integrated into a clonal model in all MCPyV-positive carcinomas; moreover, there is a nonsense mutation in the LTAg gene coding for a C-terminal truncated protein, whose mutation onset is not yet understood whether it occurs before or after integration or whether both possibilities can exist [7,8]. Truncated LTAg has lost its capability to support viral replication, nevertheless its oncogenic potential persists. Significantly, the early transforming genes encoded by MCPyV interfere with the retinoblastoma protein (RB) and p53 tumor suppressor pathways. Upon viral integration, a truncated form of the MCPyV LTAg is expressed, containing the LXCXE motif, which in turn enables the LTAg to bind to the RB protein and disable its tumor-suppressive functions, leading to tumor development [9]. The LTAg also seems to play a crucial role in tumor maintenance and cell growth. Additionally, MCPyV-positive MCC (MCCP) tumors express the MCPyV STAg. The binding of STAg to F-Box and WD Repeat Domain Containing 7 (Fbxw7), a critical tumor suppressor and one of the most deregulated proteins in the ubiquitin-proteasome system in human cancer, results in the accumulation of oncogenic proteins such as cyclin-E, c-Jun, mTOR, and truncated LTAg [10]. Moreover, the STAg is considered the primary driver gene for transformation and plays a significant role in the metastatic spread of MCC [11].
1.1.2. Ultraviolet-Related Merkel Cell Carcinoma
Conversely, UV-induced and MCPyV-negative MCC (MCCN) versus MCCP has been distinguished with peculiar molecular pathogenetic pathways. MCCN prevails in white patients living in areas with high UV exposure being characterized by a high tumor mutational burden-UV related, with RB and p53 as prevalent mutated genes. Nevertheless, both MCCN and MCCP are immunogenic with high intra-tumoral programmed-death ligand 1 (PD-L1) expression. Both UV-associated and viral-associated carcinogenesis lead to the development of highly immunogenic tumors due to the presence of multiple neoepitopes or viral proteins. These tumors are typically not clinically evident until they acquire immune escape mechanisms or when they are present in immunocompromised patients and cannot be effectively controlled by the immune system [3].
1.1.3. Merkel Cell Carcinoma Diagnosis and Treatment
MCC typically manifests as a firm, asymptomatic, non-tender flesh-colored or red nodule or plaque, rapidly increasing in size, with ulceration and bleeding occurring at an advanced stage. The sun-exposed areas of the head, neck and extremities represent the most frequent localizations [1]. Dermatoscopy can provide useful insights into MCC, revealing a prominent red color corresponding to numerous blood vessels or generalized erythema. A milky-red or pink structureless color is an additional characteristic, appearing as a pink background or smaller roundish areas known as milky-red areas, globules, or clods [12,13]. Various morphological types of vessels may be present, including dotted, glomerular, arborizing, and linear irregular vessels and polymorphous vascular patterns [12,13], with white areas also frequently observed [14]. However, the diagnosis is histological, characterized by the expression of cytokeratin 20 (CD20) and the absence of thyroid transcription factor 1 (TTF-1)[1].
Treatment options for MCC include surgery, radiotherapy, chemotherapy and immunotherapy. Surgery is the recommended approach for local and loco-regional primary MCC, aiming to remove the primary lesion with clear margins [15], while radiotherapy has historically been a primary treatment modality after surgical removal of the lesion. Chemotherapy using drugs such as platinum-based agents, etoposide, doxorubicin, taxanes, and anthracyclines, either alone or in combination, is utilized for tumor management, although with limited improvements in patient overall survival. The development of immune checkpoint therapy has been a breakthrough in MCC management, with inhibitors targeting PD-L1 and programmed cell death 1 (PD-1) reaching promising results in clinical trials. MCC employs a mechanism called "immune evasion" to evade the immune system, through the increased expression of PD-1 on tumor-infiltrating or peripheral antigen-specific T cells, as well as upregulated PD-L1 expression on tumor cells, intra-tumoral macrophages, and peritumoral immune cells. Therapies targeting both PD-1 and PD-L1 are expected to restore T-cell-mediated antitumoral reactions. Both MCCN, characterized by a high burden of UV-induced somatic mutations that generate neo-antigens, and MCCP, which expresses viral oncogenes, can be recognized by the immune system. However, high PD-L1 expression and a vigorous lymphocyte response are more commonly observed in MCCP compared to MCCN, suggesting that MCPyV antigens can enhance tumor PD-L1 expression, thus leading to increased immunogenicity in virus-positive MCC. Consequently, MCCP exhibits a better response to PD-1/PD-L1 blockade therapies compared to MCCN [1,5]. Figure 1 represents the clinical and dermoscopic features of a case of MCC.
1.2. MicroRNAs
1.2.1. microRNAs Biogenesis
MicroRNAs (miRNAs) are tiny RNA molecules, consisting of 20-25 nucleotides, that exert their regulatory effects by binding to messenger RNA (mRNA) molecules and modulating their stability and translation into proteins. One of the remarkable features of miRNAs is to target multiple genes simultaneously, allowing for the fine-tuning of complex biological networks [16]. By binding to the 3' untranslated region (UTR) of target mRNA molecules, miRNAs can induce translational repression or mRNA degradation, thereby downregulating the expression of specific genes. This post-transcriptional regulation mediated by miRNAs is involved in several biological processes, including cell proliferation, apoptosis, and immune response. MiRNAs are transcribed from specific genomic loci as long primary transcripts called pri-miRNAs. These pri-miRNAs undergo a series of enzymatic processing steps to generate mature miRNAs [17]. The initial processing event takes place in the nucleus, where the pri-miRNA is cleaved by the endonuclease Drosha, resulting in the formation of a hairpin-shaped precursor miRNA called pre-miRNA [18]. The pre-miRNA is then transported to the cytoplasm by nuclear export factor exportin-5 (XPO5), where it undergoes additional processing facilitated by the endonuclease Dicer. This processing leads to the generation of a mature miRNA duplex. Within this duplex, one of the miRNA strands, known as the “guide strand”, is selectively incorporated into the RNA-induced silencing complex (RISC), while the other strand, known as the “passenger strand”, is usually degraded. This incorporation of the guide strand into RISC allows for the precise targeting of mRNAs, resulting in their translational repression or degradation [19]. Recent data suggest that alongside the biogenesis pathway just described, known as the “canonical pathway”, there are non-canonical pathways for miRNA production defined as “Dicer independent” and “Droshoa independent” pathways [20,21]. The canonical pathway of miRNAs biogenesis is represented in Figure 2.
1.2.2. Functions of microRNAs in Cancers
Once loaded onto the RISC, the mature miRNA guides the complex to its target mRNA molecules through base pairing between the miRNA and the target mRNA. The regulation system is guided by the grade of complementarity between the miRNA and its target. The perfect or almost-perfect complementarity leads to mRNA degradation, while the partial complementarity results in translational repression. The discovery of miRNAs has revolutionized the understanding of gene regulation and provided novel insights into the complexity of cellular processes. Their dysregulation has been implicated in several diseases, such as cardiovascular disorders, neurodegenerative diseases, viral infections, and, above all, cancers. The up-to-date knowledge highlights that miRNAs, playing a crucial role in gene expression regulation, can both stimulate or attenuate metabolic activities, which, in turn, can lead to cancer development. Alterations of these tiny molecules’ production have been correlated to carcinogenesis through several and complex mechanisms, including chromosomal DNA mutations, epigenetic modifications, and transcriptional dysregulations [22,23]. On this topic, a more comprehensive understanding of the intricate link between miRNAs regulation and tumorigenesis is mandatory for advancing cancer prevention and treatment. Scientific research is increasingly focusing on the goal of exploiting the potential of miRNAs as diagnostic, prognostic and predictive markers of response to therapy, which appears to be promising for the development of innovative approaches in personalized medicine. In this context, a possible future role of artificial intelligence (AI) can be evaluated which, through big data analysis, can ease early diagnosis and guide physicians’ therapeutical choices for patient-tailored strategies.
This review aims to reorganize the knowledge known so far on the role of miRNAs in MCC pathogenesis and progression, also trying to offer a starting point on a new scientific field concerning the possible diagnostic perspectives and therapeutic approaches of these molecules. Also, new perspectives on the poorly explored field of AI in MCC are provided.
2. Role of miRNAs in Merkel Cell Carcinoma
2.1. miR-375 and Merkel Cell Carcinoma
Several miRNAs have been described as dysregulated in MCC, for both MCCN and MCCP subtypes. One of the most deeply studied miRNAs in MCC is hsa-miR-375 [23,24,25,26,27,28,29,30,31], which seems to play a dichotomous function based on specific tumor type [32], since it can act as an oncogenic miRNA or, conversely, as an anti-tumoral miRNA [33,34,35,36]. By conducting deep sequencing of RNA from different sample groups, including normal skin, basal cell carcinoma (BCC) and MCC, it was discovered that miR-375 is specifically associated with MCC, with its concentration being 60 times higher compared to the non-MCC group (consisting of normal skin and BCC samples). Moreover, the levels of miR-375 were independent of the viral state, thus suggesting that the presence of the virus does not influence the expression of miR-375[29]. Confirming this hypothesis, Fan et al. screened a set of sera from a group of MCC patients for circulating hsa-miR-375 and highlighted that hsa-miR-375 levels strongly correlated to the tumor volume, hypotizing the possible involvement of this miRNA in MCC pathogenesis [28]. Other studies highlighted that silencing hsa-miR-375 in MCC cell lines did not affect cell proliferation, morphology, apoptosis, or oncogenic signaling pathways, so it was proposed to play a more likely role in intercellular communication in MCC as an “exosome-shuttle miRNA”[37]. It was found that when transferred horizontally, hsa-miR-375 induced polarization of fibroblasts towards cancer-associated fibroblasts (CAFs) by downregulating genes involved in fibroblast polarization, such as recombination signal binding protein for immunoglobulin kappa J region (RBPJ) and TP53. These findings suggest that miR-375's function in MCC cells is to induce a pro-tumorigenic microenvironment through fibroblast polarization, rather than directly affecting MCC cells themselves [38]. However, other studies have shown different conclusions, reporting both an intracellular role for hsa-miR-375 in MCC and a dual nature of this miRNA in different pathophysiological conditions, particularly concerning MCPyV positivity [30,39].
2.1.1. miR-375 Related to the Viral State
Differently than what was stated before, just as the dual role of hsa-miR-375 in promoting and inhibiting carcinogenesis has been reported, a similar dual role has been described in MCC depending on MCPyV status, since an overexpression of hsa-miR-375 in MCCP tissues and cells compared to MCCN has been observed [31,39]. The inhibition of hsa-miR-375 expression can reduce cell growth and increase apoptosis in MCCP cells, while its ectopic expression can decrease proliferation and migration in MCCN cells, resulting in proapoptotic effects and cell cycle arrest. The functional expression of hsa-miR-375 seems to be mediated by MCPyV truncated LTAg through the expression of atonal bHLH transcription factor 1 (ATOH1), a key regulator of Merkel cell development, which directly activates hsa-miR-375 expression during MCPyV-driven carcinogenesis [40]. Demonstrating the upstream role played by ATOH1, some authors showed that hsa-miR-375 expression was enhanced following the forced overexpression of ATOH1 in MCCN cells [40]. However, the ATOH1 role is not still completely understood as it is downregulated during MCC progression but overexpressed in MCCP carcinogenesis [41,42].
Another non-unique aspect concerns the different expression of hsa-miR-375 based on the histological differentiation of MCCs. Hsa-miR-375 has been observed to exhibit high expression levels in well-differentiated MCC cells, while it is silenced in undifferentiated and aggressive MCC cells. When artificially induced in these aggressive MCC cells, hsa-miR-375 promotes neuroendocrine differentiation and reduces cell viability, migration, invasion, and survival, which suggests a potential tumor-suppressive role for this miRNA. The inhibition of two target genes, NOTCH2 and RBPJ, belonging to the Notch signaling pathway by hsa-miR-375, seems to contribute to the neuroendocrine differentiation process, suggesting that hsa-miR-375-driven MCC may arise from a cutaneous subpopulation of neuroendocrine cells, including both Merkel cells or their precursors. Concerning MCPyV positivity, the suppression of hsa-miR-375 did not impact NOTCH2 expression and cell proliferation in MCC cells, but it led to increased RBPJ expression, thus highlighting that MCCP cells may rely on MCPyV LTAg/STAg for sustained growth and survival, unlike MCCN cells, whose proliferation is likely driven by hsa-miR-375 [30].
In MCCP cells, functional inhibition of hsa-miR-375 expression can lead to reduced cell growth and increased apoptosis while, on the other hand, ectopic expression of hsa-miR-375 in MCCN cells can decrease proliferation and migration, resulting in proapoptotic effects and cell cycle arrest. Simultaneously, a negative correlation has been observed between the expression of lactate dehydrogenase B (LDHB), a target of hsa-miR-375 known to play important roles in cancer cell growth and progression, and the expression of hsa-miR-375 in MCCP cells. Specifically, silencing LDHB counteracted the growth effects caused by the inhibition of hsa-miR-375 in MCCP cells, while LDHB inhibition reduced cell growth in MCCN cells [39]. Further experiments on MCCP cells revealed that the expression of hsa-miR-375 is also promoted by two autophagy-related genes, autophagy-related 7 (ATG7) and ubiquitin-binding protein p62 (p62). This suggests that MCPyV oncoproteins might suppress autophagy in MCCP cells by targeting autophagy-related genes, thereby protecting the cells from autophagy-induced cell death [27].
In conclusion, Hsa-miR-375 might function as a tumor suppressor in MCCN, while acting as an oncomiR in MCCP. Figure 3 highlights the dual role of has-miR-375 in MCCP and MCCN.
2.2. Other miRNAs Dysregulated in Merkel Cell Carcinoma
The biological role of other several miRNAs in MCC is yet to be fully explored, despite their reported dual role as tumor-promoting and tumor-inhibiting molecules. In an initial study, using next-generation sequencing, Ning et al. analyzed the microRNAome of three MCCs, with unknown MCPyV status, one melanoma, one squamous cell carcinoma (SCC), one BCC, and one normal skin sample, aiming to distinguish miRNAs that are specifically associated with MCC. Their findings revealed that eight miRNAs were upregulated in MCC (miR-502-3p, miR-9, miR-7, miR-340, miR-182, miR-190b, miR-873, and miR-183), whereas three miRNAs were downregulated when compared to non-MCC cutaneous tumors and normal skin (miR-3170, miR-125b, and miR-374c)[43].
Among these microRNAs, hsa-miR-182, hsa-miR-190b, hsa-miR-183, and hsa-miR-340 were also found to be overexpressed in a single MCCP cell line compared to various non-MCC cells. However, the functional roles of these microRNAs in MCC and their relationship with MCPyV require further investigation, since the expression patterns of several miRNAs have been found to differ between MCCP and MCCN tumors [43]. Specifically, hsa-miR-30a, hsa-miR-30a-3p, hsa-miR-30a-5p, hsa-miR-34a, hsa-miR-769-5p, hsa-miR-142-3p, and hsa-miR-1539 were overexpressed in MCCP tumors, while hsa-miR-203 was downregulated [44]. Conversely, miRNAs that exhibit higher expression levels in MCCN, as compared to MCCP, include miR-125b, miR-374c, and miR-3170[43]. Among these, miR-125b seems to play a dual role in suppressing and promoting cancer progression, while also down-regulating the activation and cytotoxicity activity of γδ T cells as well as cells involved in anti-tumor surveillance [45,46]. On the other hand, miR-374c has been described as a new miRNA in cervical cancer, while miR-3170 was first detected in the miRNAome of melanoma and subsequently observed in breast cancer as well [47,48].
A comparative analysis of the intracellular miRNA expression profiles in 10 MCCN and 16 MCCP by a miRNA microarray-based method revealed 36 over-expressed and 20 under-expressed miRNAs in MCCP compared to MCCN [31]. Among these, miR-30a-3p, miR-30a-5p, miR-34a, miR-375, and miR-769-5p showed a significant overexpression, while miR-203 exhibited a significant underexpression, as confirmed by qRT-PCR. Regarding hsa-miR-30a-3p and hsa-miR-30a-5p, functional studies conducted in MCCP cells have shown that the expression of these miRNAs can be upregulated by MCPyV oncoproteins LTAg/STAg. These miRNAs were found to target Beclin 1 (BECN1), a protein involved in autophagy and cell death [27]. The upregulation of hsa-miR-30a-3p and hsa-miR-30a-5p by MCPyV oncoproteins suggests a role in suppressing autophagy to protect cancer cells from cell death, highlighting an oncomiR function for these miRNAs in MCCP.
The potential role of miR-203 in the development of MCC was investigated by overexpressing miR-203 into three MCCN cell lines [31]. This led to a decrease in cell growth, an increase in the number of cells in the G1 phase of the cell cycle, and a decrease in the number of cells in the G2 phase, but with no noticeable impact on apoptosis compared to cells transfected with a miRNA mimic control. The expression of the anti-apoptotic gene BIRC5 (survivin) was negatively regulated by the over-expression of miR-203, but not inducing apoptosis. On the other hand, when miR-203 was over-expressed in the MCCP WaGa cell line, it did not significantly affect cell proliferation, cell cycle progression, or survivin expression levels, thus suggesting that miR-203 regulates the expression of survivin in MCCN cell lines, but not in MCCP cell lines. These evidences indicate a tumor-suppressive function for hsa-miR-203 in MCC, particularly in cases MCCN [49].
Another miRNA investigated for its established strong association with cancer is miR-20a-5p [50]. This is usually due to dysregulation (overexpression or downregulation) of the upstream processing machinery components, resulting in aberrant biosynthesis of miR-20a-5p [50]. On this topic, a recent in vitro study investigating the involvement of the histone methyltransferase PR/SET domain 8 (PRDM8) in MCCN onset identified hsa-miR-20a-5p as a negative upstream regulator of this gene. Hsa-miR-20a-5p was found to be downregulated in MCCN cells, thus leading to high levels of PRDM8 protein and the repressive histone mark H3 lysine 9 trimethylation, which is involved in silencing tumor suppressor genes [51]. These findings highlight a novel epigenetic mechanism involving the regulation of PRDM8 by hsa-miR-20a-5p in MCCN development, suggesting hsa-miR-20a-5p as a potential tumor suppressor miRNA candidate in MCCN treatment. Gravemeyer et al. demonstrated that MCCP exhibited lower expression of genes associated with EMT, a process involved in the acquisition of migratory and invasive properties by cells. Concurrently, they found higher expression levels of miRNA clusters, namely miR-200c-141 and miR-183-96-182, which are known regulators of EMT [52].
Finally, the differential expression of miRNAs in primary and metastatic MCC has also been investigated, thus highlighting that 92 miRNAs were over-expressed in metastatic MCC as compared to primary tumors [31]. Among the miRNAs analyzed, miR-150 was identified as the only miRNA significantly upregulated after qRT-PCR validation, despite miR-142-3p, miR-483-5p and miR-630 being initially identified as among the most upregulated [31]. Table 1 summarizes miRNAs dysregulation in MCC. Figure 4 summarizes the main dysregulated miRNAs in MCCP.
2.3. Exosomal miRNAs in Merkel Cell Carcinoma
Exosomes, one of the three main types of extracellular vesicles (EVs)[53] are nanoscale vesicles, having a round or oval shape [54] and ranging in diameter from 30 to 130 nm, which are actively released through the fusion of multivesicular endosomes with the plasma membrane and persist in various bodily fluids such as blood and lymph [55]. It is well-established that exosomes act as vehicles for transporting various molecules, including DNA, miRNAs, mRNAs, proteins, lipids and circular RNAs (circRNAs)[55]. Among these, miRNAs are highly abundant in exosomes due to their small size of 19-24 nucleotides. Some miRNAs are specifically enriched in exosomes and are referred to as “exosome-shuttle miRNAs”, including the already mentioned has-miR-375, playing a significant role in the communication between tumor cells and stromal cells, and influencing various aspects of tumor biology, including proliferation, angiogenesis, metastasis, immune responses, as well as the activation of CAFs, which are present in the tumor microenvironment [55]. Exosome-shuttle miRNAs contribute to the complex network of cellular interactions that shape tumor behavior and can have profound effects on tumor progression and metastatic process, being recognized as crucial mediators of molecular communication and material transfer between primary tumor sites and distant metastasis sites [55]. Moving from these premises, exosomal miRNAs have gained attention as promising cancer biomarkers for various cancers, including melanoma, breast, colon, prostate, renal, and gastric cancer [56,57,58,59,60,61,62], due to their easy retrieval from body fluids, eliminating the need for tumor biopsy [63,64,65]. However, limited research has been conducted on exosomal-miRNAs specifically in MCC, and they have not yet been utilized as biomarkers. Referring to the most studied miRNA, hsa-miR-375 has been demonstrated to induce fibroblast polarization towards CAFs by regulating genes involved in fibroblast polarization, including RBPJ and TP53[37]. This suggests that miR-375 in MCCP cells may induce a pro-tumorigenic microenvironment, rather than directly affecting the MCCP cells themselves. Moreover, silencing hsa-miR-375 in MCC cell lines had no significant effects on cell proliferation, morphology, apoptosis, or oncogenic signaling pathways, thus ruling out an intracellular role for miR-375 in MCCP [38].
MiR-30a, miR-34, and miR-375 have been also reported in MCC [29]. Although miR-30 and miR-375 were found in exosomes derived from MCC cell lines, it remains unknown whether these miRNAs are present in non-transformed Merkel cells. Moreover, miR-30a and miR-34 have shown increased levels in MCCP compared to MCCN cases, suggesting their potential application in distinguishing between MCCP and MCCN [31,44]. Among the miRNAs previously identified in intracellular MCC samples from MCC cell lines, the initial findings confirmed the presence of miR-30a, miR-125b, miR-183, miR-190b, and miR-375 in the exosomes [23]. Moving to clinical applications, to improve diagnosis, real-time PCR methods targeting exosomal miRNAs specific to MCCP could replace conventional PCR using DNA extracted from tumor samples, with the advantage of accessibility of body fluids like blood or urine, which are easier to obtain for patients compared to biopsies, thus saving time and the need for invasive tests for patients. Figure 5 highlights exosomal miRNAs dysregulated in MCC.
2.4. CircRNAs in Merkel Cell Carcinoma
CircRNAs are a class of non-coding RNA molecules that are characterized by their covalently closed loop structure, lacking 5' caps and 3' polyadenylated tails and generated through a process called “back-splicing”, where a downstream splice acceptor site is joined to an upstream splice donor site, resulting in a circularized RNA molecule. CircRNAs have been implicated in various biological processes, including gene expression regulation, RNA splicing modulation, and protein interaction modulation [66]. Their presence and functional relevance have been discovered across different species, suggesting their evolutionary conservation and potential biological significance [67,68]. CircRNAs exhibit cell-, tissue-, or disease-specific activities, leading to contrasting effects in different contexts, acting as oncogenic drivers or tumor suppressors [69]. However, studies have reported abnormal expression of circRNAs in cancer tissues, implicating their role in tumor initiation and progression. The imbalances in circRNA levels in neoplastic diseases can be related to different mechanisms that alter circRNA circularization during cancer development [70]. In fact, the expression patterns of circRNAs have been extensively investigated in solid cancers such as epithelial ovarian cancer, breast cancer, and esophageal squamous cell carcinoma [70]. Recent research has shed light on the role of circRNAs in MCC pathogenesis [71], with specific circRNAs being identified as dysregulated compared to healthy tissues [71], which seem to contribute to tumor progression through various mechanisms. For instance, some circRNAs may act as sponges for miRNAs, sequestering them and preventing their interaction with target mRNAs, thereby indirectly modulating gene expression. Others circRNAs can interact with RNA-binding proteins and influence their activities, modulating cellular processes and signaling pathways involved in MCC development and progression [65]. Moreover, certain circRNAs have been found to participate in the regulation of key cancer-related pathways, such as cell proliferation, apoptosis, migration, and invasion. By affecting these pathways, circRNAs can influence the behavior of MCC cells, promoting tumor growth, metastasis, or immune evasion [65]. On this topic, a recent study conducted by Abere et al. has revealed the presence of a circRNA called circMCV-T in the genome of MCPyV. These circRNAs could counteract the effects of MCPyV-miR-M1, a miRNA encoded by the virus. Specifically, circMCV-T is a circular RNA molecule that is 762 nucleotides long and is generated through the process of back-splicing of the MCPyV transcript from the viral early region, which is expressed simultaneously with the viral LTAg and inhibits MCPyV-miR-M1, thereby promoting LTAg expression during viral replication. This intricate interplay between viral circRNAs, miRNAs, and mRNA molecules modulates the replication of MCPyV DNA. However, the biological significance of these RNAs in the context of MCCP is yet to be fully understood [71]. Yang et al. conducted a study that identified protein-coding circALTO RNAs derived from both MCPyV and Trichodysplasia spinulosa polyomavirus (TSPyV)[72]. In the case of MCPyV, two circRNAs (circALTO1 and circALTO2) were found in the early region surrounding the ALTO gene in MCCP cell lines and patient tumors, suggesting that MCPyV circALTO can enhance the transcriptional activity of specific promoters.
We suggest that understanding the specific circRNAs involved in MCC and elucidating their functional roles can provide insights into the underlying mechanisms of the disease and potentially reveal novel therapeutic targets.
3. Possible Uses and Limitations of miRNAs in Merkel Cell Carcinoma
3.1. miRNAs possible Clinical Applications
MiRNAs are gaining clinical significance as early disease and prognostic biomarkers for their ability to either enable early disease identification or predict patient outcomes, particularly using liquid biopsies which are noninvasive and easily repeatable, allowing for efficient and repeated quantification of miRNAs [73]. Despite encouraging data, only a few miRNAs have been attributed with potential diagnostic and prognostic roles in MCC. Hsa-miR-375 has been extensively studied in MCC tissues and cells [28,29,30,31], while limited data are available for other miRNAs, such as hsa-miR-30a and hsa-mir-34a, which have been identified in MCC tissues and cells just in two independent studies [31,44]. The prognostic importance of miRNAs in MCC has been demonstrated for hsa-miR-150, which is associated with metastasis and poor prognosis [31], as well as circulating hsa-miR-375, which has been suggested as a patient monitoring tool [28]. In fact, elevated levels of circulating hsa-miR-375 have been observed in MCC patients compared to tumor-free patients, with high diagnostic accuracy, indicating its potential as a reliable liquid biopsy-based biomarker for tumor burden in MCC patients without restriction to MCPyV positivity. This data suggests the possible use of hsa-miR-375 for therapy monitoring and the follow-up of MCC patients [28]. As already discussed, the first-line treatment for mMCC includes immunotherapy, which is showing promising results in mMCC treatment by targeting the PD1-PDL-1 pathway using anti-PD1 antibodies (pembrolizumab, nivolumab) or anti-PDL-1 antibodies (avelumab)[74]. These therapies have demonstrated favorable responses, with a six-month progression-free survival rate ranging from 40% to 85% and even complete tumor resolution in some patients [75,76,77,78,79,80]. However, not all patients respond positively to these therapies, highlighting the need for the development of additional treatment approaches. Drugs targeting MCC-specific miRNAs or their target transcripts could be used in combination with the already approved immunotherapy treatments, but unfortunately clinical trials involving miRNAs have been initiated for various pathological conditions, although not specifically for MCC [81,82]. To establish useful miRNA biomarkers, multicentric studies analyzing the microRNAome on a significant number of MCC samples or biological fluids obtained from patients are mandatory to correlate miRNA data with clinical features, tumor stage, viral status, expression of specific markers, patient age, and gender.
3.2. miRNAs Limitations
Despite the presence of promising data, currently available studies on miRNA dysregulation in MCC do not provide sufficient evidence to establish definitive conclusions in clinical applications, with the main limitation caused by incomplete and often conflicting results [83,84]. One of the challenges in studying miRNAs in MCC is the lack of knowledge about the specific miRNA network of normal Merkel cells, partly due to the rarity of Merkel cells and their unknown origin. Additionally, there are too few Merkel cells in the skin to account for the high copies of the MCPyV genome detected on healthy human skin [85]. Further studies, particularly using single-cell transcriptomic/proteomic approaches, are needed to better characterize Merkel cells and identify the cells of origin for MCC. Moreover, it remains unclear whether the miRNAs derived from MCC are specifically expressed in MCC cells or also present in non-malignant Merkel cells [23]. Standard tumor sampling may not effectively separate the tumor from histologically benign tissue, resulting in contamination by nonmalignant cells such as epithelial, endothelial, white and blood cells, and fibroblasts [86,87], which can impact the reliability of miRNA-based studies. Primary cell cultures established from fresh MCC specimens can potentially reduce contamination bias during RNA isolation by isolating RNA from tumor cells separately [86,87]. Finally, obtaining large sets of MCC tissues is challenging due to the rarity of this cutaneous cancer, resulting in some studies being conducted with a relatively small number of samples, thus leading to insufficient statistical power. To overcome these challenges, multicenter studies should be designed to collect MCC tissues/liquid biopsies from different centers, including both MCCP and MCCN samples.
4. Application of Artificial Intelligence in MCC
In recent years, advancements in the field of AI have shown promise in enhancing the diagnosis, management, and assessment of skin cancers, sometimes performing better than dermatologists [88]. Machine learning (ML) models for skin cancer recognition can analyze medical images, including dermoscopic images or photographs of skin lesions, identifying specific patterns and features of MCC, thereby helping early detection and diagnosis, and potentially allowing for earlier intervention and improved patient outcomes. However, some issues are emerging: when trained ML models are deployed into real-life experience, the detection of rare but aggressive skin cancers, including MCC, represents an important challenge, since these cancers are not well-covered in training datasets. Aiming to overcome these limitations, a data set of MCC, amelanotic melanoma, and common benign lesions was obtained by Steele et al. Model1 (direct to consumer) returns a high/low-risk binary output, while Model2 (search) provides a list of the top five diagnoses with prediction confidence. However, in the direct-to-consumer model, approximately 20% of MCCs were misdiagnosed as low-risk lesions, and the false-positive rate for common benign lesions was greater than 60%. This limitation arises from the fact that these models are trained on the most frequent tumors and lesions, excluding the rarest ones such as MCC, which limits their application in real-life clinical practice. Indeed, models trained without MCC may erroneously learn that the red color is strongly suggestive only for benign vascular lesions, obtaining excellent results during ML setup, but not diagnosing MCC in clinical practice [89]. From this emerges the need to train these ML models also with specific images and features specific to rare tumors.
AI models can also integrate clinical and genomic data to assess the prognosis, risk of recurrence and metastasis in MCC patients. A study conducted by Cheng et al. identified and characterized the landscape of cellular states and ecotypes in MCC based on a ML framework, highlighting that the genes highly expressed in cellular states were significantly enriched in immune- and cancer hallmark-related pathways. Transcriptional regulation analysis revealed the critical transcription factors (i.e. E2F1, E2F3 and E2F7), which play important roles in regulating the TME of MCC, providing an initial knowledge to understand the intrinsic subtypes of MCCs and the pathways involved in distinct subtype oncogenesis [90]. Moreover, liquid-biopsies-based detection of miRNA biomarkers in AI-based models for early-stage cancer subtyping and prognosis is emerging, which could be useful in machine-based predictive modeling of cancer staging and progression [91]. In dermatological fields, this line of research has been already developed in cutaneous melanoma, with miRNAs being analyzed in a non-linear classification model to predict melanoma metastasis and recurrence [92]. Finally, in the future, AI algorithms could accelerate the identification and development of novel therapeutic targets for MCC through the analysis of large datasets of genomic and molecular information or, conversely, could predict the efficacy of existing drugs or repurposed compounds. AI systems, by analyzing complex datasets, can serve as decision support tools for oncologists and dermatologists in the management of patients with MCC, providing evidence-based recommendations and assisting physicians in treatment decisions. However, it is mandatory to validate AI models rigorously and integrate them effectively into clinical practice to realize their full potential in dermo-oncology. Finally, the “explainability”, the ability to understand the decisions or predictions made by an ML model, particularly deep learning models, which are often seen as "black boxes" due to their complex architectures and high amount of data, results crucial to understand, validate, and potentially correct the decisions made by AI systems, especially in the oncological field.
5. Conclusions and Future Perspectives
Our review highlights the increasing significance of miRNAs in understanding MCC pathogenesis and progression, encompassing both MCCP and MCCN subtypes. Several miRNAs are being studied, and although some of them are showing promise as potential candidates for playing a role in MCC pathogenesis and progression, their precise roles remain not fully elucidated. Specific miRNA levels were correlated with MCCP, while others were correlated with MCCN. Some miRNAs identified in the MCC have been found to play a dual function, acting as both oncogenes and tumor suppressors in other cellular systems. Consequently, targeting the expression of these miRNAs may present a double-edged sword, which could affect both pro-tumor and anti-tumor pathways. Therefore, it is crucial to elucidate the precise roles of miRNAs in MCC to develop effective and specific therapeutic future strategies. By understanding the specific contributions of miRNAs in the MCC, it becomes possible to design interventions that selectively target oncogenic pathways while sparing tumor suppressive mechanisms. Therefore, the need to conduct further functional investigations to unravel the mechanistic involvement of miRNAs in MCC is arising. Monitoring MCC patients based on miRNA profiles could offer valuable insights during therapy. Furthermore, miRNAs have demonstrated potential as prognostic indicators for therapy resistance and are emerging as viable targets for tumor treatment, but their implication in drug resistance and therapeutic applications in MCC has not been explored. Nevertheless, miRNAs present several limitations. These molecules are present in very low levels in biological samples, particularly in liquid biopsies, and they are often not detected by PCR. The lack of understanding regarding the specific miRNA network in normal Merkel cells is another limitation that could be addressed in the future through single-cell transcriptomic and proteomic studies. Contamination from non-malignant cells in MCC tissues could be mitigated by establishing primary cell cultures. Lastly, MCC is a very rare cutaneous cancer, therefore the issue of small sample sizes in MCC studies has emerged. From this, we suggest the need for multicenter investigations to increase sample size and statistical power. Finally, a prospective role of AI in the analysis of miRNAs, especially based on ML systems, is emerging in the dermato-oncology field, proving to be a possible future support for clinicians in the early diagnosis, staging, and prognosis of MCC patients.
Author Contributions
Conceptualization, F.L.P. and F.B.; methodology, F.L.P. and F.B.; validation, F.L.P, G.B. and S.B.; formal analysis, F.L.P; investigation, F.L.P.; resources, F.L.P.; data curation, F.L.P., L.I. and M.V.; writing—original draft preparation, F.L.P; writing—review and editing, F.L.P. and L.I; visualization, F.L.P.; supervision, S.G., G.B., L.I., A.R. and F.B.; project administration, G.B., L.I. and A.R. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Gauci, M.-L.; Aristei, C.; Becker, J.C.; Blom, A.; Bataille, V.; Dreno, B.; Del Marmol, V.; Forsea, A.M.; Fargnoli, M.C.; Grob, J.-J.; et al. Diagnosis and Treatment of Merkel Cell Carcinoma: European Consensus-Based Interdisciplinary Guideline – Update 2022. Eur J Cancer 2022, 171, 203–231. [CrossRef]
- Samimi, M.; Kervarrec, T.; Touze, A. Immunobiology of Merkel Cell Carcinoma. Curr Opin Oncol 2020, 32, 114–121. [CrossRef]
- Papa, V.; Li Pomi, F.; Borgia, F.; Vaccaro, M.; Pioggia, G.; Gangemi, S. Immunosenescence and Skin: A State of Art of Its Etiopathogenetic Role and Crucial Watershed for Systemic Implications. Int J Mol Sci 2023, 24, 7956. [CrossRef]
- Liu, W.; MacDonald, M.; You, J. Merkel Cell Polyomavirus Infection and Merkel Cell Carcinoma. Curr Opin Virol 2016, 20, 20–27. [CrossRef]
- Zaggana, E.; Konstantinou, M.P.; Krasagakis, G.H.; de Bree, E.; Kalpakis, K.; Mavroudis, D.; Krasagakis, K. Merkel Cell Carcinoma—Update on Diagnosis, Management and Future Perspectives. Cancers (Basel) 2022, 15, 103. [CrossRef]
- Schowalter, R.M.; Pastrana, D. V.; Pumphrey, K.A.; Moyer, A.L.; Buck, C.B. Merkel Cell Polyomavirus and Two Previously Unknown Polyomaviruses Are Chronically Shed from Human Skin. Cell Host Microbe 2010, 7, 509–515. [CrossRef]
- Stakaitytė, G.; Wood, J.; Knight, L.; Abdul-Sada, H.; Adzahar, N.; Nwogu, N.; Macdonald, A.; Whitehouse, A. Merkel Cell Polyomavirus: Molecular Insights into the Most Recently Discovered Human Tumour Virus. Cancers (Basel) 2014, 6, 1267–1297. [CrossRef]
- Arora, R.; Chang, Y.; Moore, P.S. MCV and Merkel Cell Carcinoma: A Molecular Success Story. Curr Opin Virol 2012, 2, 489–498. [CrossRef]
- DeCaprio, J.A. Merkel Cell Polyomavirus and Merkel Cell Carcinoma. Philosophical Transactions of the Royal Society B: Biological Sciences 2017, 372, 20160276. [CrossRef]
- Verhaegen, M.E.; Mangelberger, D.; Harms, P.W.; Vozheiko, T.D.; Weick, J.W.; Wilbert, D.M.; Saunders, T.L.; Ermilov, A.N.; Bichakjian, C.K.; Johnson, T.M.; et al. Merkel Cell Polyomavirus Small T Antigen Is Oncogenic in Transgenic Mice. Journal of Investigative Dermatology 2015, 135, 1415–1424. [CrossRef]
- Pietropaolo, V.; Prezioso, C.; Moens, U. Merkel Cell Polyomavirus and Merkel Cell Carcinoma. Cancers (Basel) 2020, 12, 1774. [CrossRef]
- Dalle, S.; Parmentier, L.; Moscarella, E.; Phan, A.; Argenziano, G.; Thomas, L. Dermoscopy of Merkel Cell Carcinoma. Dermatology 2012, 224, 140–144. [CrossRef]
- Lallas, A.; Moscarella, E.; Argenziano, G.; Longo, C.; Apalla, Z.; Ferrara, G.; Piana, S.; Rosato, S.; Zalaudek, I. Dermoscopy of Uncommon Skin Tumours. Australasian Journal of Dermatology 2014, 55, 53–62. [CrossRef]
- Jalilian, C.; Chamberlain, A.J.; Haskett, M.; Rosendahl, C.; Goh, M.; Beck, H.; Keir, J.; Varghese, P.; Mar, A.; Hosking, S.; et al. Clinical and Dermoscopic Characteristics of Merkel Cell Carcinoma. British Journal of Dermatology 2013, 169, 294–297. [CrossRef]
- Kachare, S.D.; Wong, J.H.; Vohra, N.A.; Zervos, E.E.; Fitzgerald, T.L. Sentinel Lymph Node Biopsy Is Associated with Improved Survival in Merkel Cell Carcinoma. Ann Surg Oncol 2014, 21, 1624–1630. [CrossRef]
- Oliveto, S.; Mancino, M.; Manfrini, N.; Biffo, S. Role of MicroRNAs in Translation Regulation and Cancer. World J Biol Chem 2017, 8, 45. [CrossRef]
- Rani, V.; Sengar, R.S. Biogenesis and Mechanisms of MicroRNA-mediated Gene Regulation. Biotechnol Bioeng 2022, 119, 685–692. [CrossRef]
- Dexheimer, P.J.; Cochella, L. MicroRNAs: From Mechanism to Organism. Front Cell Dev Biol 2020, 8. [CrossRef]
- Lin, Y.-C.; Chen, T.-H.; Huang, Y.-M.; Wei, P.-L.; Lin, J.-C. Involvement of MicroRNA in Solid Cancer: Role and Regulatory Mechanisms. Biomedicines 2021, 9, 343. [CrossRef]
- Rizk, N.I.; Midan, H.M.; Helal, G.K.; Abulsoud, A.I.; Elshaer, S.S.; El-Husseiny, A.A.; Fathi, D.; Abdelmaksoud, N.M.; Abdel Mageed, S.S.; Elballal, M.S.; et al. The Emerging Role of MiRNAs in Merkel Cell Carcinoma Pathogenesis: Signaling Pathway Crosstalk. Pathol Res Pract 2023, 249, 154771. [CrossRef]
- Miyoshi, K.; Miyoshi, T.; Siomi, H. Many Ways to Generate MicroRNA-like Small RNAs: Non-Canonical Pathways for MicroRNA Production. Molecular Genetics and Genomics 2010, 284, 95–103. [CrossRef]
- Mazziotta, C.; Cervellera, C.F.; Lanzillotti, C.; Touzé, A.; Gaboriaud, P.; Tognon, M.; Martini, F.; Rotondo, J.C. MicroRNA Dysregulations in Merkel Cell Carcinoma: Molecular Mechanisms and Clinical Applications. J Med Virol 2023, 95. [CrossRef]
- Konstantinell, A.; Coucheron, D.; Sveinbjørnsson, B.; Moens, U. MicroRNAs as Potential Biomarkers in Merkel Cell Carcinoma. Int J Mol Sci 2018, 19, 1873. [CrossRef]
- Rotondo, J.C.; Mazziotta, C.; Lanzillotti, C.; Tognon, M.; Martini, F. Epigenetic Dysregulations in Merkel Cell Polyomavirus-Driven Merkel Cell Carcinoma. Int J Mol Sci 2021, 22, 11464. [CrossRef]
- Neagu, M.; Constantin, C.; Cretoiu, S.M.; Zurac, S. MiRNAs in the Diagnosis and Prognosis of Skin Cancer. Front Cell Dev Biol 2020, 8. [CrossRef]
- Fan, K.; Gravemeyer, J.; Ritter, C.; Rasheed, K.; Gambichler, T.; Moens, U.; Shuda, M.; Schrama, D.; Becker, J.C. MCPyV Large T Antigen-Induced Atonal Homolog 1 Is a Lineage-Dependency Oncogene in Merkel Cell Carcinoma. Journal of Investigative Dermatology 2020, 140, 56-65.e3. [CrossRef]
- Kumar, S.; Xie, H.; Shi, H.; Gao, J.; Juhlin, C.C.; Björnhagen, V.; Höög, A.; Lee, L.; Larsson, C.; Lui, W. Merkel Cell Polyomavirus Oncoproteins Induce MicroRNAs That Suppress Multiple Autophagy Genes. Int J Cancer 2020, 146, 1652–1666. [CrossRef]
- Fan, K.; Ritter, C.; Nghiem, P.; Blom, A.; Verhaegen, M.E.; Dlugosz, A.; Ødum, N.; Woetmann, A.; Tothill, R.W.; Hicks, R.J.; et al. Circulating Cell-Free MiR-375 as Surrogate Marker of Tumor Burden in Merkel Cell Carcinoma. Clinical Cancer Research 2018, 24, 5873–5882. [CrossRef]
- Renwick, N.; Cekan, P.; Masry, P.A.; McGeary, S.E.; Miller, J.B.; Hafner, M.; Li, Z.; Mihailovic, A.; Morozov, P.; Brown, M.; et al. Multicolor MicroRNA FISH Effectively Differentiates Tumor Types. Journal of Clinical Investigation 2013, 123, 2694–2702. [CrossRef]
- Abraham, K.J.; Zhang, X.; Vidal, R.; Paré, G.C.; Feilotter, H.E.; Tron, V.A. Roles for MiR-375 in Neuroendocrine Differentiation and Tumor Suppression via Notch Pathway Suppression in Merkel Cell Carcinoma. Am J Pathol 2016, 186, 1025–1035. [CrossRef]
- Xie, H.; Lee, L.; Caramuta, S.; Höög, A.; Browaldh, N.; Björnhagen, V.; Larsson, C.; Lui, W.-O. MicroRNA Expression Patterns Related to Merkel Cell Polyomavirus Infection in Human Merkel Cell Carcinoma. Journal of Investigative Dermatology 2014, 134, 507–517. [CrossRef]
- Costa-Pinheiro, P.; Ramalho-Carvalho, J.; Vieira, F.Q.; Torres-Ferreira, J.; Oliveira, J.; Gonçalves, C.S.; Costa, B.M.; Henrique, R.; Jerónimo, C. MicroRNA-375 Plays a Dual Role in Prostate Carcinogenesis. Clin Epigenetics 2015, 7, 42. [CrossRef]
- Shi, Z.; Chu, X.; Wu, Y.; Wu, J.; Lu, C.; Lü, R.; Ding, M.; Mao, N. MicroRNA-375 Functions as a Tumor Suppressor in Osteosarcoma by Targeting PIK3CA. Tumor Biology 2015, 36, 8579–8584. [CrossRef]
- CUI, F.; WANG, S.; LAO, I.; ZHOU, C.; KONG, H.; BAYAXI, N.; LI, J.; CHEN, Q.; ZHU, T.; ZHU, H. MiR-375 Inhibits the Invasion and Metastasis of Colorectal Cancer via Targeting SP1 and Regulating EMT-Associated Genes. Oncol Rep 2016, 36, 487–493. [CrossRef]
- Osako, Y.; Seki, N.; Kita, Y.; Yonemori, K.; Koshizuka, K.; Kurozumi, A.; Omoto, I.; Sasaki, K.; Uchikado, Y.; Kurahara, H.; et al. Regulation of MMP13 by Antitumor MicroRNA-375 Markedly Inhibits Cancer Cell Migration and Invasion in Esophageal Squamous Cell Carcinoma. Int J Oncol 2016, 49, 2255–2264. [CrossRef]
- Chang, Y.; Lin, J.; Tsung, A. Manipulation of Autophagy by MIR375 Generates Antitumor Effects in Liver Cancer. Autophagy 2012, 8, 1833–1834. [CrossRef]
- Fan, K.; Zebisch, A.; Horny, K.; Schrama, D.; Becker, J.C. Highly Expressed MiR-375 Is Not an Intracellular Oncogene in Merkel Cell Polyomavirus-Associated Merkel Cell Carcinoma. Cancers (Basel) 2020, 12, 529. [CrossRef]
- Fan, K.; Spassova, I.; Gravemeyer, J.; Ritter, C.; Horny, K.; Lange, A.; Gambichler, T.; Ødum, N.; Schrama, D.; Schadendorf, D.; et al. Merkel Cell Carcinoma-Derived Exosome-Shuttle MiR-375 Induces Fibroblast Polarization by Inhibition of RBPJ and P53. Oncogene 2021, 40, 980–996. [CrossRef]
- Kumar, S.; Xie, H.; Scicluna, P.; Lee, L.; Björnhagen, V.; Höög, A.; Larsson, C.; Lui, W.-O. MiR-375 Regulation of LDHB Plays Distinct Roles in Polyomavirus-Positive and -Negative Merkel Cell Carcinoma. Cancers (Basel) 2018, 10, 443. [CrossRef]
- Fan, K.; Gravemeyer, J.; Ritter, C.; Rasheed, K.; Gambichler, T.; Moens, U.; Shuda, M.; Schrama, D.; Becker, J.C. MCPyV Large T Antigen-Induced Atonal Homolog 1 Is a Lineage-Dependency Oncogene in Merkel Cell Carcinoma. Journal of Investigative Dermatology 2020, 140, 56-65.e3. [CrossRef]
- Gambichler, T.; Mohtezebsade, S.; Wieland, U.; Silling, S.; Höh, A.-K.; Dreißigacker, M.; Schaller, J.; Schulze, H.-J.; Oellig, F.; Kreuter, A.; et al. Prognostic Relevance of High Atonal Homolog-1 Expression in Merkel Cell Carcinoma. J Cancer Res Clin Oncol 2017, 143, 43–49. [CrossRef]
- Ostrowski, S.M.; Wright, M.C.; Bolock, A.M.; Geng, X.; Maricich, S.M. Ectopic Atoh1 Expression Drives Merkel Cell Production in Embryonic, Postnatal and Adult Epidermis. Development 2015. [CrossRef]
- Ning, M.S.; Kim, A.S.; Prasad, N.; Levy, S.E.; Zhang, H.; Andl, T. Characterization of the Merkel Cell Carcinoma MiRNome. J Skin Cancer 2014, 2014, 1–9. [CrossRef]
- Veija, T.; Sahi, H.; Koljonen, V.; Bohling, T.; Knuutila, S.; Mosakhani, N. MiRNA-34a Underexpressed in Merkel Cell Polyomavirus-Negative Merkel Cell Carcinoma. Virchows Archiv 2015, 466, 289–295. [CrossRef]
- Yin, H.; Sun, Y.; Wang, X.; Park, J.; Zhang, Y.; Li, M.; Yin, J.; Liu, Q.; Wei, M. Progress on the Relationship between MiR-125 Family and Tumorigenesis. Exp Cell Res 2015, 339, 252–260. [CrossRef]
- Zou, C.; Zhao, P.; Xiao, Z.; Han, X.; Fu, F.; Fu, L. Γδ T Cells in Cancer Immunotherapy. Oncotarget 2017, 8, 8900–8909. [CrossRef]
- Stark, M.S.; Tyagi, S.; Nancarrow, D.J.; Boyle, G.M.; Cook, A.L.; Whiteman, D.C.; Parsons, P.G.; Schmidt, C.; Sturm, R.A.; Hayward, N.K. Characterization of the Melanoma MiRNAome by Deep Sequencing. PLoS One 2010, 5, e9685. [CrossRef]
- Persson, H.; Kvist, A.; Rego, N.; Staaf, J.; Vallon-Christersson, J.; Luts, L.; Loman, N.; Jonsson, G.; Naya, H.; Hoglund, M.; et al. Identification of New MicroRNAs in Paired Normal and Tumor Breast Tissue Suggests a Dual Role for the ERBB2/Her2 Gene. Cancer Res 2011, 71, 78–86. [CrossRef]
- Arora, R.; Shuda, M.; Guastafierro, A.; Feng, H.; Toptan, T.; Tolstov, Y.; Normolle, D.; Vollmer, L.L.; Vogt, A.; Dömling, A.; et al. Survivin Is a Therapeutic Target in Merkel Cell Carcinoma. Sci Transl Med 2012, 4. [CrossRef]
- Huang, W.; Wu, X.; Xiang, S.; Qiao, M.; Cen, X.; Pan, X.; Huang, X.; Zhao, Z. Regulatory Mechanism of MiR-20a-5p Expression in Cancer. Cell Death Discov 2022, 8, 262. [CrossRef]
- Orouji, E.; Peitsch, W.K.; Orouji, A.; Houben, R.; Utikal, J. Unique Role of Histone Methyltransferase PRDM8 in the Tumorigenesis of Virus-Negative Merkel Cell Carcinoma. Cancers (Basel) 2020, 12, 1057. [CrossRef]
- Gravemeyer, J.; Lange, A.; Ritter, C.; Spassova, I.; Song, L.; Picard, D.; Remke, M.; Horny, K.; Sriram, A.; Gambichler, T.; et al. Classical and Variant Merkel Cell Carcinoma Cell Lines Display Different Degrees of Neuroendocrine Differentiation and Epithelial-Mesenchymal Transition. Journal of Investigative Dermatology 2021, 141, 1675-1686.e4. [CrossRef]
- Raposo, G.; Stoorvogel, W. Extracellular Vesicles: Exosomes, Microvesicles, and Friends. Journal of Cell Biology 2013, 200, 373–383. [CrossRef]
- Han, Z.; Chen, H.; Guo, Z.; Shen, J.; Luo, W.; Xie, F.; Wan, Y.; Wang, S.; Li, J.; He, J. Circular RNAs and Their Role in Exosomes. Front Oncol 2022, 12. [CrossRef]
- Mashouri, L.; Yousefi, H.; Aref, A.R.; Ahadi, A. mohammad; Molaei, F.; Alahari, S.K. Exosomes: Composition, Biogenesis, and Mechanisms in Cancer Metastasis and Drug Resistance. Mol Cancer 2019, 18, 75. [CrossRef]
- HUANG, K.; TANG, Y.; HE, L.; DAI, Y. MicroRNA-340 Inhibits Prostate Cancer Cell Proliferation and Metastasis by Targeting the MDM2-P53 Pathway. Oncol Rep 2016, 35, 887–895. [CrossRef]
- Pfeffer, S.; Grossmann, K.; Cassidy, P.; Yang, C.; Fan, M.; Kopelovich, L.; Leachman, S.; Pfeffer, L. Detection of Exosomal MiRNAs in the Plasma of Melanoma Patients. J Clin Med 2015, 4, 2012–2027. [CrossRef]
- Imamura, T.; Komatsu, S.; Ichikawa, D.; Miyamae, M.; Okajima, W.; Ohashi, T.; Kiuchi, J.; Nishibeppu, K.; Kosuga, T.; Konishi, H.; et al. Low Plasma Levels of MiR-101 Are Associated with Tumor Progression in Gastric Cancer. Oncotarget 2017, 8, 106538–106550. [CrossRef]
- Zhao, Q.; Deng, S.; Wang, G.; Liu, C.; Meng, L.; Qiao, S.; Shen, L.; Zhang, Y.; Lü, J.; Li, W.; et al. A Direct Quantification Method for Measuring Plasma MicroRNAs Identified Potential Biomarkers for Detecting Metastatic Breast Cancer. Oncotarget 2016, 7, 21865–21874. [CrossRef]
- Foj, L.; Ferrer, F.; Serra, M.; Arévalo, A.; Gavagnach, M.; Giménez, N.; Filella, X. Exosomal and Non-Exosomal Urinary MiRNAs in Prostate Cancer Detection and Prognosis. Prostate 2017, 77, 573–583. [CrossRef]
- Wang, J.; Yan, F.; Zhao, Q.; Zhan, F.; Wang, R.; Wang, L.; Zhang, Y.; Huang, X. Circulating Exosomal MiR-125a-3p as a Novel Biomarker for Early-Stage Colon Cancer. Sci Rep 2017, 7, 4150. [CrossRef]
- Butz, H.; Nofech-Mozes, R.; Ding, Q.; Khella, H.W.Z.; Szabó, P.M.; Jewett, M.; Finelli, A.; Lee, J.; Ordon, M.; Stewart, R.; et al. Exosomal MicroRNAs Are Diagnostic Biomarkers and Can Mediate Cell–Cell Communication in Renal Cell Carcinoma. Eur Urol Focus 2016, 2, 210–218. [CrossRef]
- Hayes, J.; Peruzzi, P.P.; Lawler, S. MicroRNAs in Cancer: Biomarkers, Functions and Therapy. Trends Mol Med 2014, 20, 460–469. [CrossRef]
- Lan, H.; Lu, H.; Wang, X.; Jin, H. MicroRNAs as Potential Biomarkers in Cancer: Opportunities and Challenges. Biomed Res Int 2015, 2015, 1–17. [CrossRef]
- Wang, H.; Peng, R.; Wang, J.; Qin, Z.; Xue, L. Circulating MicroRNAs as Potential Cancer Biomarkers: The Advantage and Disadvantage. Clin Epigenetics 2018, 10, 59. [CrossRef]
- Ma, S.; Kong, S.; Wang, F.; Ju, S. CircRNAs: Biogenesis, Functions, and Role in Drug-Resistant Tumours. Mol Cancer 2020, 19, 119. [CrossRef]
- Holdt, L.M.; Kohlmaier, A.; Teupser, D. Molecular Roles and Function of Circular RNAs in Eukaryotic Cells. Cellular and Molecular Life Sciences 2018, 75, 1071–1098. [CrossRef]
- Salzman, J.; Chen, R.E.; Olsen, M.N.; Wang, P.L.; Brown, P.O. Cell-Type Specific Features of Circular RNA Expression. PLoS Genet 2013, 9, e1003777. [CrossRef]
- Fontemaggi, G.; Turco, C.; Esposito, G.; Di Agostino, S. New Molecular Mechanisms and Clinical Impact of CircRNAs in Human Cancer. Cancers (Basel) 2021, 13, 3154. [CrossRef]
- Allegra, A.; Cicero, N.; Tonacci, A.; Musolino, C.; Gangemi, S. Circular RNA as a Novel Biomarker for Diagnosis and Prognosis and Potential Therapeutic Targets in Multiple Myeloma. Cancers (Basel) 2022, 14, 1700. [CrossRef]
- Abere, B.; Zhou, H.; Li, J.; Cao, S.; Toptan, T.; Grundhoff, A.; Fischer, N.; Moore, P.S.; Chang, Y. Merkel Cell Polyomavirus Encodes Circular RNAs (CircRNAs) Enabling a Dynamic CircRNA/MicroRNA/MRNA Regulatory Network. mBio 2020, 11. [CrossRef]
- Jin, Y.; Yang, H.; Ji, W.; Wu, W.; Chen, S.; Zhang, W.; Duan, G. Virology, Epidemiology, Pathogenesis, and Control of COVID-19. Viruses 2020, 12, 372. [CrossRef]
- Boyer, M.; Cayrefourcq, L.; Dereure, O.; Meunier, L.; Becquart, O.; Alix-Panabières, C. Clinical Relevance of Liquid Biopsy in Melanoma and Merkel Cell Carcinoma. Cancers (Basel) 2020, 12, 960. [CrossRef]
- Badalamenti, G.; Incorvaia, L.; Algeri, L.; Carreca, I.U.; Brando, C.; Madonia, G.; Peri, M.; Cucinella, A.; Perez, A.; Barraco, N.; et al. Immunometabolic Predictive Factors in Merkel Cell Carcinoma (MCC) Patients Treated with Avelumab. Journal of Clinical Oncology 2022, 40, e21525–e21525. [CrossRef]
- Mantripragada, K.; Birnbaum, A. Response to Anti-PD-1 Therapy in Metastatic Merkel Cell Carcinoma Metastatic to the Heart and Pancreas. Cureus 2015. [CrossRef]
- Nghiem, P.T.; Bhatia, S.; Lipson, E.J.; Kudchadkar, R.R.; Miller, N.J.; Annamalai, L.; Berry, S.; Chartash, E.K.; Daud, A.; Fling, S.P.; et al. PD-1 Blockade with Pembrolizumab in Advanced Merkel-Cell Carcinoma. New England Journal of Medicine 2016, 374, 2542–2552. [CrossRef]
- Kaufman, H.L.; Russell, J.; Hamid, O.; Bhatia, S.; Terheyden, P.; D’Angelo, S.P.; Shih, K.C.; Lebbé, C.; Linette, G.P.; Milella, M.; et al. Avelumab in Patients with Chemotherapy-Refractory Metastatic Merkel Cell Carcinoma: A Multicentre, Single-Group, Open-Label, Phase 2 Trial. Lancet Oncol 2016, 17, 1374–1385. [CrossRef]
- Walocko, F.M.; Scheier, B.Y.; Harms, P.W.; Fecher, L.A.; Lao, C.D. Metastatic Merkel Cell Carcinoma Response to Nivolumab. J Immunother Cancer 2016, 4, 79. [CrossRef]
- Thiem, A.; Kneitz, H.; Schummer, P.; Herz, S.; Schrama, D.; Houben, R.; Goebeler, M.; Schilling, B.; Gesierich, A. Coincident Metastatic Melanoma and Merkel Cell Carcinoma with Complete Remission on Treatment with Pembrolizumab. Acta Dermato Venereologica 2017, 97, 1252–1254. [CrossRef]
- Kaufman, H.L.; Russell, J.S.; Hamid, O.; Bhatia, S.; Terheyden, P.; D’Angelo, S.P.; Shih, K.C.; Lebbé, C.; Milella, M.; Brownell, I.; et al. Updated Efficacy of Avelumab in Patients with Previously Treated Metastatic Merkel Cell Carcinoma after ≥1 Year of Follow-up: JAVELIN Merkel 200, a Phase 2 Clinical Trial. J Immunother Cancer 2018, 6, 7. [CrossRef]
- Chakraborty, C.; Sharma, A.R.; Sharma, G.; Doss, C.G.P.; Lee, S.-S. Therapeutic MiRNA and SiRNA: Moving from Bench to Clinic as Next Generation Medicine. Mol Ther Nucleic Acids 2017, 8, 132–143. [CrossRef]
- Incorvaia, L.; Fanale, D.; Badalamenti, G.; Brando, C.; Bono, M.; De Luca, I.; Algeri, L.; Bonasera, A.; Corsini, L.R.; Scurria, S.; et al. A “Lymphocyte MicroRNA Signature” as Predictive Biomarker of Immunotherapy Response and Plasma PD-1/PD-L1 Expression Levels in Patients with Metastatic Renal Cell Carcinoma: Pointing towards Epigenetic Reprogramming. Cancers (Basel) 2020, 12, 3396. [CrossRef]
- Cabibi, D.; Caruso, S.; Bazan, V.; Castiglia, M.; Bronte, G.; Ingrao, S.; Fanale, D.; Cangemi, A.; Calò, V.; Listì, A.; et al. Analysis of Tissue and Circulating MicroRNA Expression during Metaplastic Transformation of the Esophagus. Oncotarget 2016, 7, 47821–47830. [CrossRef]
- Fanale, D.; Amodeo, V.; Bazan, V.; Insalaco, L.; Incorvaia, L.; Barraco, N.; Castiglia, M.; Rizzo, S.; Santini, D.; Giordano, A.; et al. Can the MicroRNA Expression Profile Help to Identify Novel Targets for Zoledronic Acid in Breast Cancer? Oncotarget 2016, 7, 29321–29332. [CrossRef]
- Schowalter, R.M.; Pastrana, D. V.; Pumphrey, K.A.; Moyer, A.L.; Buck, C.B. Merkel Cell Polyomavirus and Two Previously Unknown Polyomaviruses Are Chronically Shed from Human Skin. Cell Host Microbe 2010, 7, 509–515. [CrossRef]
- Elloumi, F.; Hu, Z.; Li, Y.; Parker, J.S.; Gulley, M.L.; Amos, K.D.; Troester, M.A. Systematic Bias in Genomic Classification Due to Contaminating Non-Neoplastic Tissue in Breast Tumor Samples. BMC Med Genomics 2011, 4, 54. [CrossRef]
- Litchfield, K.; Stanislaw, S.; Spain, L.; Gallegos, L.L.; Rowan, A.; Schnidrig, D.; Rosenbaum, H.; Harle, A.; Au, L.; Hill, S.M.; et al. Representative Sequencing: Unbiased Sampling of Solid Tumor Tissue. Cell Rep 2020, 31, 107550. [CrossRef]
- Jutzi, T.B.; Krieghoff-Henning, E.I.; Holland-Letz, T.; Utikal, J.S.; Hauschild, A.; Schadendorf, D.; Sondermann, W.; Fröhling, S.; Hekler, A.; Schmitt, M.; et al. Artificial Intelligence in Skin Cancer Diagnostics: The Patients’ Perspective. Front Med (Lausanne) 2020, 7. [CrossRef]
- Steele, L.; Velazquez-Pimentel, D.; Thomas, B.R. Do AI Models Recognise Rare, Aggressive Skin Cancers? An Assessment of a Direct-to-consumer Application in the Diagnosis of Merkel Cell Carcinoma and Amelanotic Melanoma. Journal of the European Academy of Dermatology and Venereology 2021, 35. [CrossRef]
- Cheng, S.; Li, S.; Yang, P.; Wang, R.; Zhou, P.; Li, J. Dissecting the Tumour Immune Microenvironment in Merkel Cell Carcinoma Based on a Machine Learning Framework. Artif Cells Nanomed Biotechnol 2023, 51, 397–407. [CrossRef]
- Muthamilselvan, S.; Ramasami Sundhar Baabu, P.; Palaniappan, A. Microfluidics for Profiling MiRNA Biomarker Panels in AI-Assisted Cancer Diagnosis and Prognosis. Technol Cancer Res Treat 2023, 22. [CrossRef]
- Korfiati, A.; Grafanaki, K.; Kyriakopoulos, G.C.; Skeparnias, I.; Georgiou, S.; Sakellaropoulos, G.; Stathopoulos, C. Revisiting MiRNA Association with Melanoma Recurrence and Metastasis from a Machine Learning Point of View. Int J Mol Sci 2022, 23, 1299. [CrossRef]
Figure 1.
clinical examination of a MCC in a 74-year-old man, characterized by a reddish nodule rapidly growing of the pre-auricular left region (a); dermoscopy (10x) highlights milky red areas with white streaks and irregular vessels.
Figure 1.
clinical examination of a MCC in a 74-year-old man, characterized by a reddish nodule rapidly growing of the pre-auricular left region (a); dermoscopy (10x) highlights milky red areas with white streaks and irregular vessels.
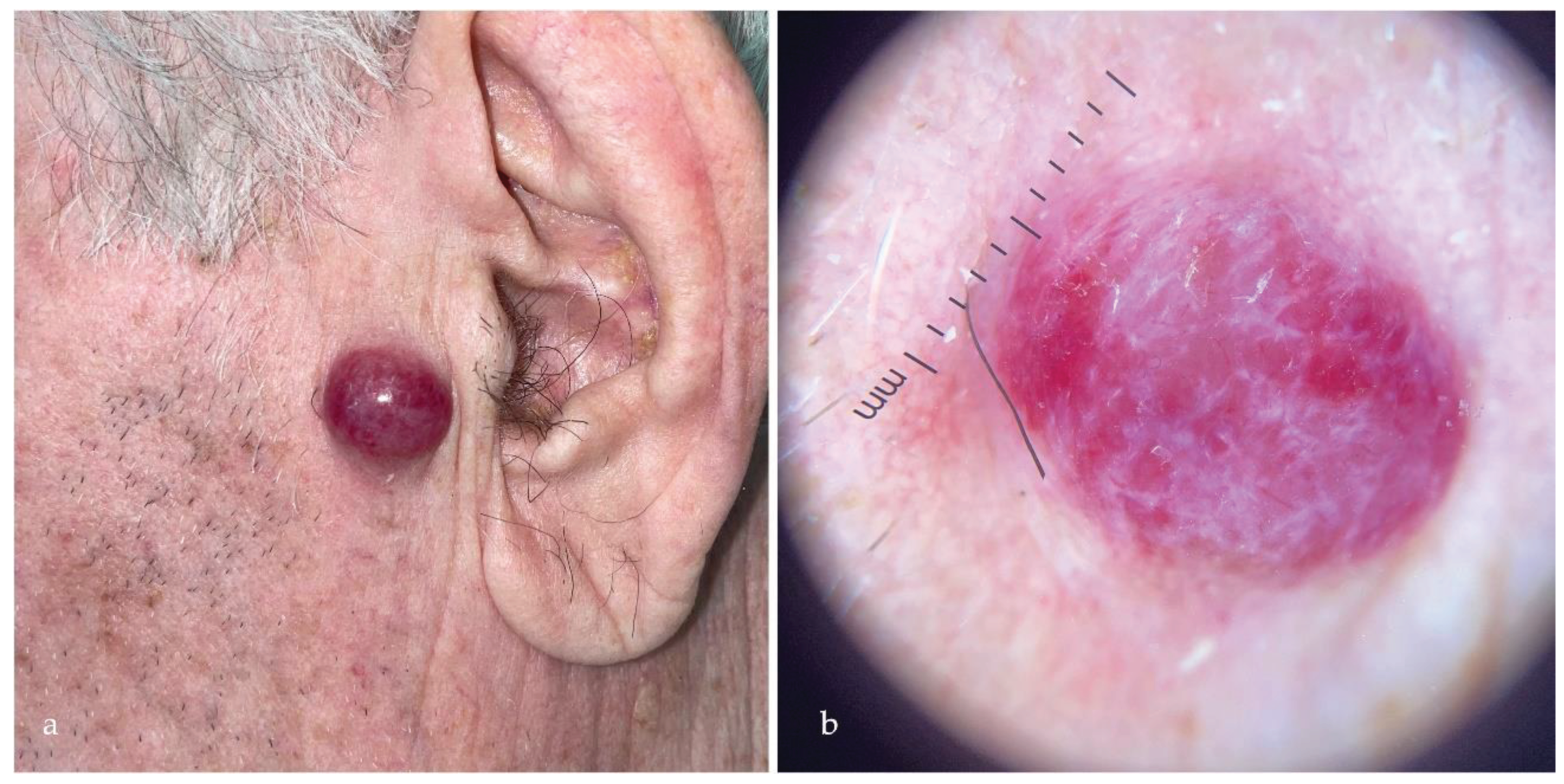
Figure 2.
The canonical pathway of miRNAs biogenesis is represented.
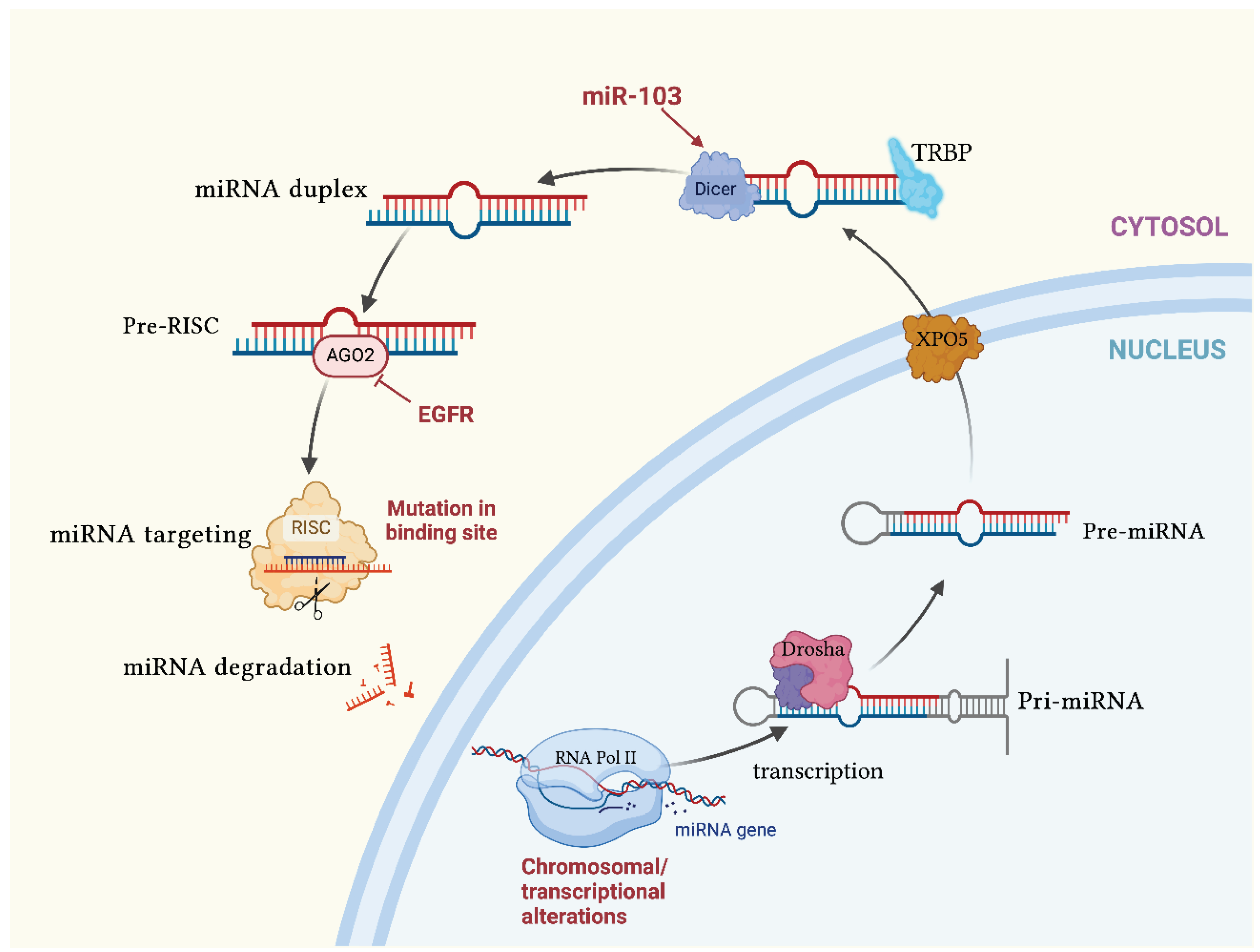
Figure 3.
the possible dual role of hsa-miR-375 in MCCP and MCCN. In MCCP emerges its role as an oncomiR, promoting cell proliferation, apoptosis inhibition, cell migration and invasion, while in MCCN it acts as a tumor-suppressor miRNA, promoting neuroendocrine differentiation and reducing cell viability, migration and survival. Created with BioRender.com.
Figure 3.
the possible dual role of hsa-miR-375 in MCCP and MCCN. In MCCP emerges its role as an oncomiR, promoting cell proliferation, apoptosis inhibition, cell migration and invasion, while in MCCN it acts as a tumor-suppressor miRNA, promoting neuroendocrine differentiation and reducing cell viability, migration and survival. Created with BioRender.com.
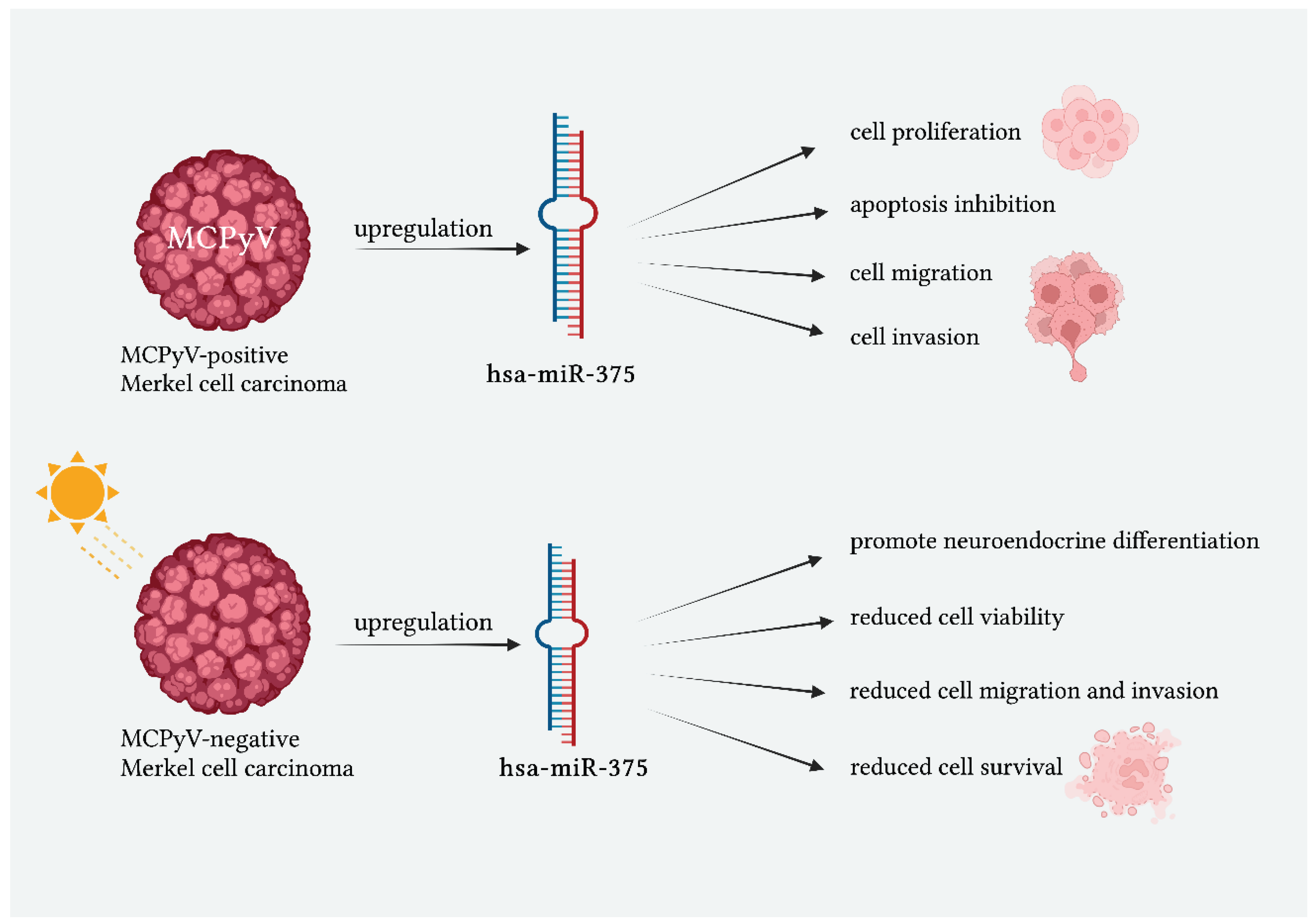
Figure 4.
during MCPyV-driven carcinogenesis, MCPyV truncated LTAg mediates the functional expression of hsa-miR-375 through ATOH1. Hsa-miR-375 promotes neuroendocrine differentiation and reduces cell viability, migration, invasion, and survival, indicating a potential tumor-suppressive role for this miRNA. The functional repression of NOTCH2 and RBPJ contributes to the neuroendocrine differentiation process. Hsa-miR-375 does not impact NOTCH2-related cell proliferation, while the inhibition of RBPJ expression, through has-miR-375, leads to a pro-tumoral microenvironment. Downregulation of TP53 causes cancer-associated fibroblast polarization. Silencing LDHB counteracts the growth effects caused by the inhibition of hsa-miR-375 in MCCP cells. Also, ATG7 and p62, two autophagy-related genes, are targeted by hsa-miR-375, thus suggesting that MCPyV oncoproteins might suppress autophagy in MCCP cells. Also, hsa-miR-30a-3p and hsa-miR-30a-5p can be upregulated by MCPyV oncoprotein LTAg/STAg. These miRNAs target, through inhibition, BECN1 which is involved in autophagy and cell death, suggesting a role in suppressing autophagy to protect cancer cells from cell death. Finally, the upregulation of miR-200c-141 and miR-183-96-182 causes a lower expression of EMT-related genes. .
Figure 4.
during MCPyV-driven carcinogenesis, MCPyV truncated LTAg mediates the functional expression of hsa-miR-375 through ATOH1. Hsa-miR-375 promotes neuroendocrine differentiation and reduces cell viability, migration, invasion, and survival, indicating a potential tumor-suppressive role for this miRNA. The functional repression of NOTCH2 and RBPJ contributes to the neuroendocrine differentiation process. Hsa-miR-375 does not impact NOTCH2-related cell proliferation, while the inhibition of RBPJ expression, through has-miR-375, leads to a pro-tumoral microenvironment. Downregulation of TP53 causes cancer-associated fibroblast polarization. Silencing LDHB counteracts the growth effects caused by the inhibition of hsa-miR-375 in MCCP cells. Also, ATG7 and p62, two autophagy-related genes, are targeted by hsa-miR-375, thus suggesting that MCPyV oncoproteins might suppress autophagy in MCCP cells. Also, hsa-miR-30a-3p and hsa-miR-30a-5p can be upregulated by MCPyV oncoprotein LTAg/STAg. These miRNAs target, through inhibition, BECN1 which is involved in autophagy and cell death, suggesting a role in suppressing autophagy to protect cancer cells from cell death. Finally, the upregulation of miR-200c-141 and miR-183-96-182 causes a lower expression of EMT-related genes. .
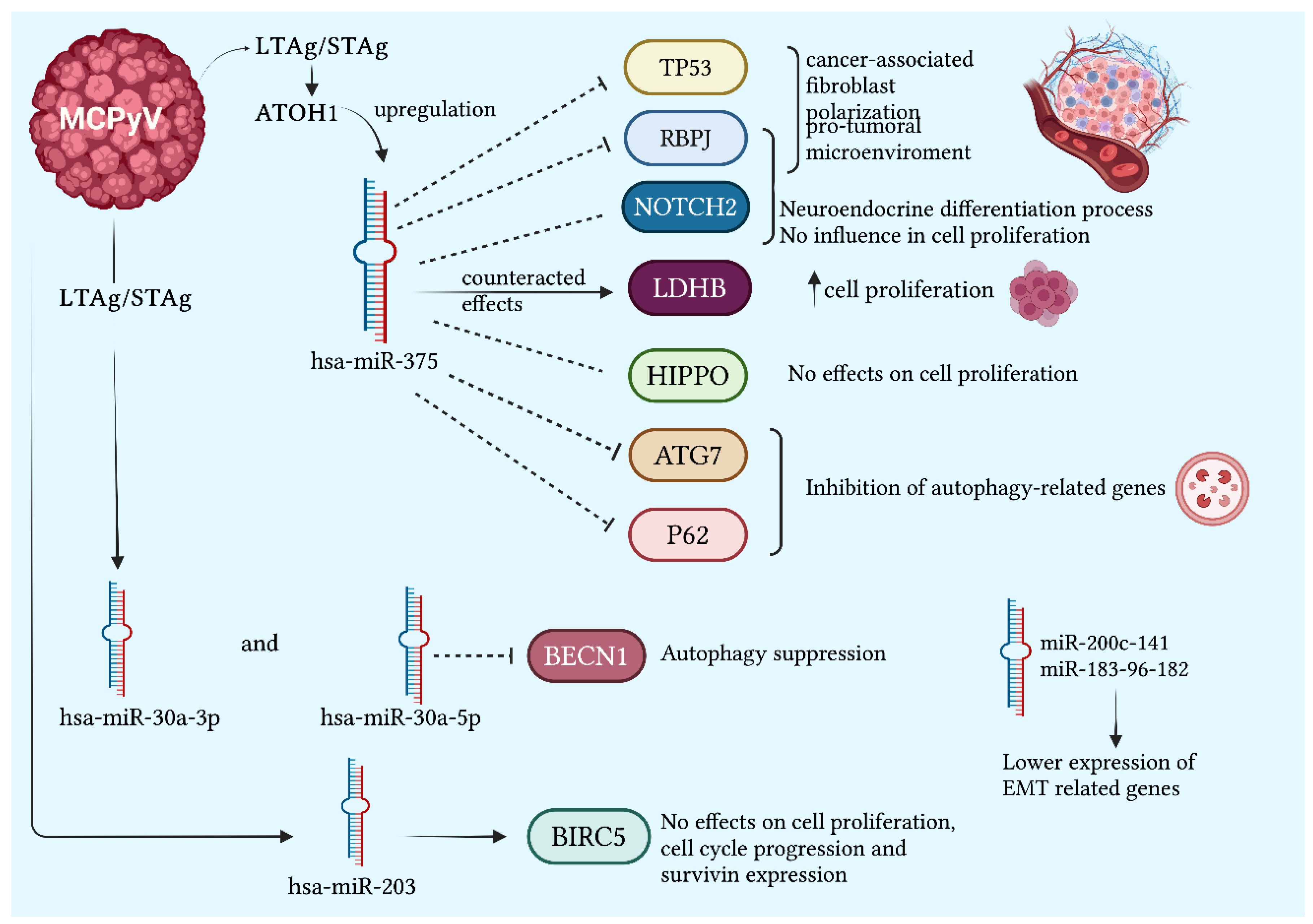
Figure 5.
exosomal miRNAs are extracted through liquid biopsies. miR-125b, miR-183 and miR-190b are overexpressed in MCC; miR-30a and miR-34 have been associated to MCCP specifically while has-miR375 induces fibroblast polarization towards CAFs, pro-tumoral microenvironment with no effect on cell proliferation.
Figure 5.
exosomal miRNAs are extracted through liquid biopsies. miR-125b, miR-183 and miR-190b are overexpressed in MCC; miR-30a and miR-34 have been associated to MCCP specifically while has-miR375 induces fibroblast polarization towards CAFs, pro-tumoral microenvironment with no effect on cell proliferation.
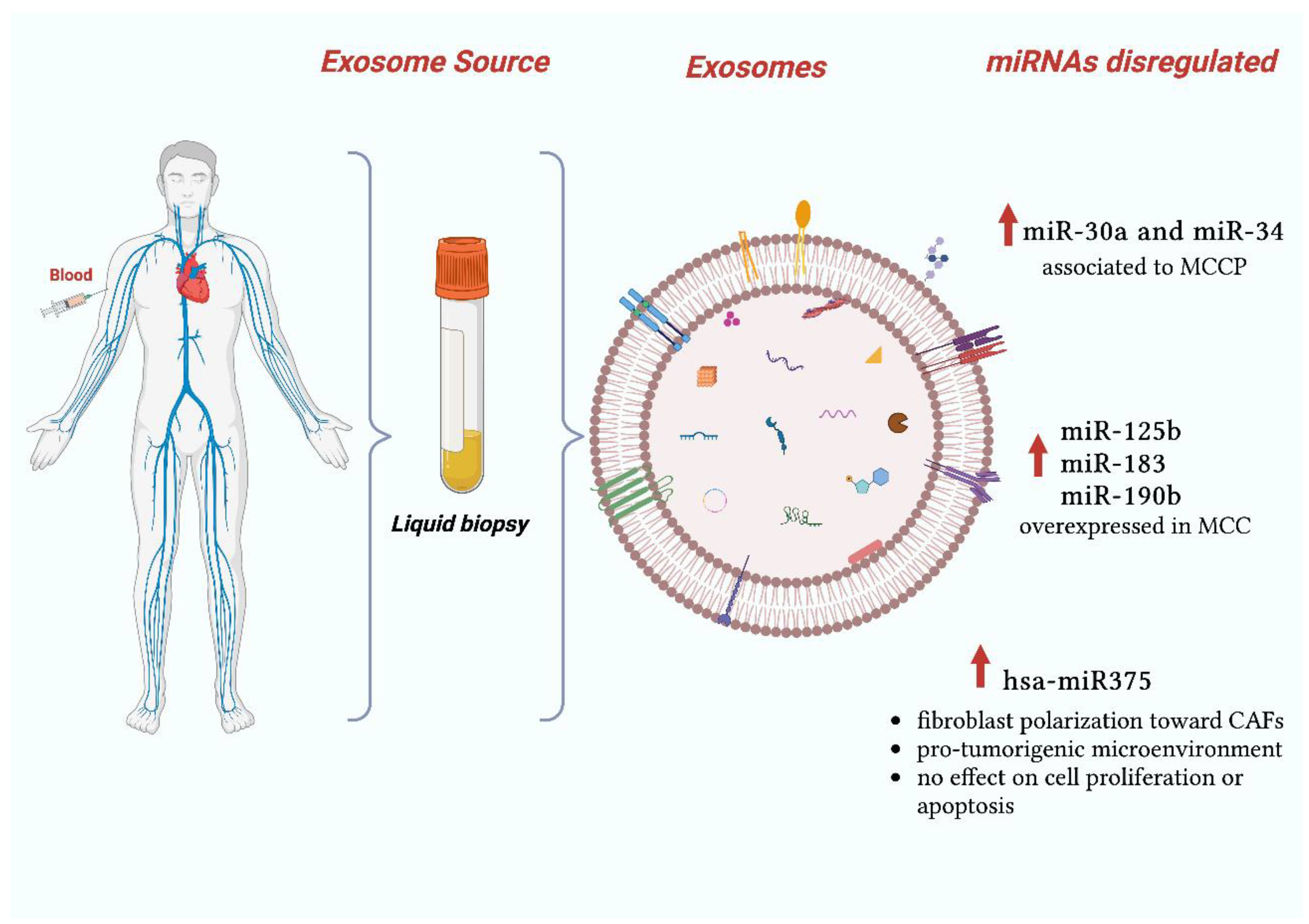

Table 1.
the main miRNAs dysregulation in MCC are summarized.
| Authors | miRNAs upregulated in MCC | Sample types | miRNAs’ role |
|---|---|---|---|
| Fan et al.[28] | miR-375 | MCCP versus MCCN tissues and cel lines | miR-375 serum level showed high diagnostic accuracy to discriminate tumor-bearing and tumor-free patients with MCC. |
| Ning et al.[43] | miRNA-9 miRNA-502-3p miRNA-7 miRNA-340 miRNA-182 miRNA-190b miRNA-873 miRNA-183 |
MCC tissues versus non-MCC skin tumor and normal skin | Enhancing tumor motility and promoting colony formation. Specifically miR-182 was localized within tumor cells and, as expected from the qRT-PCR data, expression in surrounding tissue and normal skin was low compared to that in MCC cells. |
| Renwick et al.[29] | miRNA-205 miRNA-375 |
MCC versus non-MCC tissue and cell lines | Downregulating the expression of gene targets through interaction with the three prime untranslated region (3′ UTR) of the respective genes. Potential diagnostic role by distinguish BCC from MCC |
| Xie et al.[31] | miR-150 | MCC metastases versus primary tumor | Association with tumor metastasis and disease-specific survival |
| Authors | miRNAs downregulated in MCC | Sample types | miRNAs’ role |
| Ning et al.[43] | miRNA-3170 miRNA-125b miRNA-374c |
MCC tissues versus non-MCC skin tumor and normal skin | Not reported. |
| Authors | miRNAs upregulatd in MCCP | Sample types | miRNAs’ role |
| Veija et al.[44] | miR-34a miR-1539 miR-30a miR-142-3p |
MCCP versus MCCN tissues | Possible role in the oncogenesis of MCCP. Slight underexpression of miR-181d was detectable in MCCP |
| Gravemeyer et al.[52] | miR-200c-141 miR-183-96-1832 miR-30a-5pmiR-375 |
MCCP versus MCCN cell lines | Establishment of a connection between the neuroendocrine features of MCC and the lack of EMT |
| Abraham et al. [30] | miR-375 | MCCP versus MCCN cell lines | Roles of MiR-375 in neuroendocrine differentiation |
| Xie et al.[31] |
miR-30a-3p miR-769-5p miR-34a miR-375 |
MCCP versus MCCN tissues | Specifically upregulated in MCCP compared to MCCN |
| Author | miRNAs disregulated in MCCN | Sample types | miRNAs’ role |
| Veija et al.[44] | miR-34a | MCCP versus MCCN tissues | Statistically significative reduction of miR-34a in MCCN |
| Xie et al.[31] | miR-203 | MCCP versus MCCN tissues | Specific activity only in MCCN |
| Kumar et al.[39] | miR-375 | MCCP versus MCCN tissues and cel lines | Silencing of LDHB reduced cell growth in MCPyV− cell lines, while its silencing in MCPyV+ cell lines rescued the cell growth effect mediated by miR-375 inhibition |
| Orouji et al. [51] | miR-20a-5p | miR-20a-5p downregulation in MCCN led to high levels of PRDM8 protein, which is involved in silencing tumor suppressor gene. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



