
Submitted:
17 March 2024
Posted:
18 March 2024
You are already at the latest version
Abstract
DNA damage causes the mutations that are the principal source of genetic variation. DNA damage detection and repair mechanisms therefore play a determining role in generating the genetic diversity on which natural selection acts. Speciation, it is commonly assumed, occurs at a rate set by the level of standing allelic diversity in a population. The process of speciation is driven by a combination of two evolutionary forces: genetic drift and ecological selection. Genetic drift takes place under conditions of relaxed selection, and results in a balance between rates of mutation and rates of genetic substitution. These two processes, drift and selection, are necessarily mediated by a variety of mechanisms guaranteeing genome stability in any given species. One of the outstanding questions in evolutionary biology concerns the origin of the widely varying phylogenetic distribution of biodiversity across the tree of life, and how the forces of drift and selection contribute to shaping that distribution. The following examines some of the molecular mechanisms underlying genome stability and the adaptive radiations that are associated with biodiversity and the widely varying species richness and evenness in the different eukaryotic lineages.
Keywords:
Non-adaptive radiation
; mutation rates
; species richness
; species evenness
; karyotype diversity
; DNA repair systems
1. Introduction: Karyotype Diversity and Species Richness
The origins of the biodiversity comprising the Tree of Life involve longstanding and ongoing debates in evolutionary biology. Darwin characterized the astonishing species diversity among angiosperms (some 4,000 species) as the “abominable mystery”: “the rapid development … of all higher order plants within recent geological times” [1]. The generation of biodiversity involves two fundamental but biologically independent variables: mutation and selection. Mutation events, which alter genotypes, have been assumed to occur randomly at the molecular and genetic levels, while natural selection is expected to act non-randomly on the correspondingly altered phenotypes at the individual and population levels.
A major question of interest concerns how these two variables interact to establish an equilibrium, or balance, between the forces of mutation and selection during the process of speciation. Most mutations are deleterious to the organism and undergo negative, or purifying, selection while other mutations are beneficial and undergo positive selection, or adaptation. A third class of mutation is neither beneficial nor harmful but instead “neutral” or “nearly neutral”, meaning they are expected to have either negligible or no effect on an organism’s differential fitness. Neutral mutations become fixed, or substituted, in a population through random genetic drift rather than by Darwinian natural selection.
By analogy with symbiosis, deleterious mutations can be seen as acting as parasites that harm the organism; beneficial mutations can be seen as acting as mutualists that provide an advantage to both host and symbiont (the mutation itself); while neutral mutations can be seen as acting as commensals that benefit the symbiont without significantly harming the host. The analogy is not far-fetched: eukaryotes emerged as a result of a symbiotic and mutagenic event that eventually resulted in the invasion of non-coding symbiotic DNA into the host genome. The characteristic of being neutral has, along with other mutational events such as whole genome amplifications (polyploidy), resulted in the astonishing range of genome sizes and architectures across the eukaryote Tree of Life, in striking contrast to the relatively streamlined genome size range found in prokaryotes [2].
Eukaryote genomes are organized in individual units (chromosomes) of differing numbers and sizes (karyotypes). Genomes themselves vary enormously in size across both plants and animals. Animal genome sizes, for example, range from 0.04 picograms (pg; C-value = IC) in Trichoplax adhaerens to 133 pg in Protopterus aethiopicus, or about 3,300 fold. In plants, genome sizes range over 2,400 fold from 0.06 pg to about 152 pg, the largest known eukaryote genome [3,4]. In contrast the genomes of bacteria and Archaea range in size from 0.1 to 16 Mbp, or about 160 fold [5].
Changes in karyotypes and genome sizes are closely associated with corresponding differences in species richness within different taxonomic groups such as mammals and salamanders. Karyotype diversity (KD), moreover, is highly correlated with species richness (number of species in a clade) across closely and distantly related clades. The evenness in karyotype diversity also closely correlates with species evenness in the mammalian phylogenetic tree, evenness reflecting the balance/imbalance in the distribution of either KD or species richness (SR). In each case, the respective size distributions are significantly skewed in parallel, with lower KD aligning closely with lower SR and vice-verse (Figure 1).
A still unanswered question arises, at least concerning the Mammalian lineage: are the correlations between KD and SR and the correlations of evenness in the SR and KD distributions a trivial consequence of Neo-Darwinian natural selection acting on random genome modifications and rearrangements? Or do the correlations reflect processes of non-adaptive radiation that result from a balance between mutational inputs and neutral substitutions in a population? In other words, is speciation initially an adaptively neutral phenomenon resulting from relaxed selection, which generates the genetic diversity on which Darwinian natural selection ultimately acts? And if so, to what extent can relaxed selection and genetic drift explain or account for the widely varying distributions of SR observed in most, if not all, animal and plant lineages?
The question is pertinent given that most of the eurkayotic genome is comprised of non-coding, apparently neutral DNA that derives principally from a variety of different transposable elements (TE). This “neutral DNA” is largely responsible for the 64,000 fold range in genome sizes found in eurkaryotes [6], yet the number of genes in any given lineage varies little. The average number of genes in vertebrates, for example, is about 20,000, while the average number in invertebrates is about [7]. Rats and humans have approximately the same number of genes: 22,000.) [8]. Although neutral, the location of TE’s in the genome is not random but compartmentalized in the form of late replicating gene-poor heterochromatin.
It would appear then that species in a lineage differ from each other more in the amounts of non-coding DNA than in the amount of DNA on which natural selection is expected to act, namely genes, regulatory elements and the organization of genes in the genome (synteny). Synteny in mammals, for example, has been remarkably conserved for more than 300 My [9], yet the mammalian genome has a highly variable karyotype (2n = 6-7 in the Indian muntjac to over 100 in some rodents) and a significantly large range in C-value of about 358 fold [10,11].
Although genome size often correlates negatively with rates of evolution in plants and animals, genome stability and rates of change in C-value appear to underlie rates of macroevolution [12]. Salamanders, for example, have large and highly conserved genomes that are very slowly evolving (C-value: > 10 pg); frog’s have smaller genomes that have much faster rates of evolution (C-value: < 10 pg), while mammals have rates of genome evolution 20X faster than anurans and a much more restricted range of C-value compared to the Amphibia. Importantly, synteny is also highly conserved in frogs and salamanders [13] (Figure 2).
Remarkably, natural selection has not purged the eurkaryote genome of this ostensibly useless DNA suggesting that it might play a role in adaptation and speciation, for example, in consolidating reproductive isolation [14]. While non-coding DNA itself might be biologically inert, the heterochromatin that it forms plays a number of vital roles in transcription, DNA repair, DNA replication timing, and differentiation and development. The following will look at the potential biological functions of non-coding DNA and heterochromatin in relationship to those factors contributing to adaptation and speciation.
2. Non-Adaptive Radiation: Ecological Selection vs. Genetic Drift
Motoo Kimura proposed a hypothesis of non-adaptive radiation (NAR) based on genetic drift, or the random fixation of an allele or genotype in a population [15]. The neo-Darwinian hypothesis, in contrast, holds that natural selection acting on an advantageous variant phenotype is the primary and principle driver behind fixing a genotype variant in a population [16,17]. The NAR hypothesis rests on the assumption that substitution rates equal mutation rates (mutation/substitution balance): mutation rates determine rates of substitution and, consequently, rates of speciation.
Although genetic drift might drive a mutation to substitution and fixation in a group with a small effective population size (Ne), Kimura’s NAR does not assume that Darwinian natural selection plays a minor or insignificant role in establishing reproductive isolation, for example, through the effects of speciation genes, genomic modifications resulting in incompatible karyotypes and other pre-and post zygotic barriers to gene flow [18]. It remains unclear, however, how these two evolutionary forces, drift and selection, interact during the processes of speciation and adaptive radiation. [19]
Theories of non-adaptive radiation have been proposed ever since Darwin. Non-adaptive radiation corresponds to lineage diversification in the absence of environmental shifts or evident niche divergence [20,21,22]. In contrast to ecological based theories of non-adaptive radiations, Kimura’s theory focuses on niche neutral genotype radiations at the molecular genetic level. The theory rests on four fundamental stages defining the speciation process:
- 1)
- Relaxation of a selective constraint (a weakened negative, or purifying, selection) resulting in a burst in the number of new gene and genotype variants;
- 2)
- Differential fixation of variants in a population, or subpopulations, under the force of genetic drift;
- 3)
- Rapid habitat-driven diversification into new niches and environments (ecological selection);
- 4)
- Competitive exclusion between related groups leading to extensive adaptive evolution and radically different taxa following successful adaptation to new ecological niches.
A substantial amount of evidence has accumulated in support of the NAR hypothesis since it was first formulated in 1991. The role of relaxed selection in influencing evolutionary rates is well established in plants and animals (Stage 1) [23,24,25,26]. Relaxed purifying selection is associated with changes in genome size (both expansions and reductions) and altered genome architecture and karyotypes [27,28]. The role of genetic drift in modulating genome sizes, however, remains unclear (Stage 2), but is expected to contribute significantly in the ancestral population during early stages of adaptive radiation [29,30,31]. The expected increase in mutational loads under relaxed selection in populations with small Ne enhances the levels of standing genetic variation under conditions of balancing selection (Stage 3) [32,33,34,35]. Balancing selection acts to maintain diversity in a population over long periods of time [36,37]. The corresponding elevated levels of genetic diversity (GD) in turn promote speciation when variants invade new niches and habitats (Stage 4). Population differentiation, for example, is related to speciation rates over evolutionary time [38].
Implicit in the NAR hypothesis is a time lag between stage 1 (stochastic divergence between isolated populations) and stage 4 (ecological selection and adaptation) [39]: the four stages take place in succession, or nearly in succession, over millions of years rather than simultaneously or in parallel [40]. On a microevolutionary scale, diversification without morphological change has been observed in plants, lizards and salamanders: rates of species diversification are not coincident with ecological and phenotypic evolution, while ecological and phenotypic evolution co-occur in time as expected according to ecological speciation [41]. These findings are more consistent with a primarily niche neutral diversification model than with models of simple density-dependent diversification [42]. Hence, the speciation process corresponds to a repeated cycle of niche neutral diversification followed by a period of density dependent ecological adaptation.
Other examples of neutral genotype diversification relate to genotype-phenotype maps and the neutral sets or networks they form [43]. More than one genotype can code for a single phenotype. The size distribution of neutral sets varies substantially, with any given phenotype mapping to multiple genotypes [44]. Since ecological selection acts on the individual phenotype, neutral sets of genotypes indicate a widely varying amount of degeneracy that is perhaps a signature of genetic drift [45].
Genotype-phenotype degeneracy can then be seen as analogous to the degeneracy in the genetic code [46,47], which provided an initial insight into the neutral theory of evolution. Neutral divergence of the genotype is therefore operating within the selective constraints that fix a phenotype in a population [48]. Phenotype plasticity and “epigenomic drift”, or the accumulation of stochastic epigenetic modifications, can also generate other forms of neutral and non-neutral genomic and genetic diversity [49,50,51,52]. Another example of protein evolution via the force of genetic drift concerns rapidly evolving intrinsically disordered proteins, which increase in number with organism complexity [53].
3. Genome Stability and Rates of Speciation: Karyotype Diversity Versus Gene Diversity in Determining Species Richness
As early as the 1970′s a clear distinction had been established between karyotype diversity and genetic diversity and their respective relationships to species richness [54,55]. Taken together, the observations suggested that “evolution at the organismal level is correlated more highly with karyotype evolution than with structural gene evolution.” [56]. Moreover, it was found that rates of karyotype evolution varied significantly among different taxonomic groups whereas rates of change in structural genes were about the same.
Another study found a negative correlation between levels of gene heterozygosity and rates of chromosomal speciation, suggesting that rates of speciation increase in populations with small Ne (low heterozygosity) [57]. The only feasible way, however, of estimating Ne is to rely on measures of within-population nucleotide diversity at neutral genomic sites, such as silent sites in codons (dS) [58]. While dependence of heterozygosity on Ne is necessarily true for isolated populations of the same species (same mutation rate per individual), it is not entirely clear whether the use of such measures can be applied to whole taxonomic groups for comparative studies [59]. Absolute rates of silent site divergence, for example, are 7X faster in angiosperms compared to gymnosperms [60], which might (or might not) affect biological conclusions based strictly on Ne.
Consistent with the earlier studies in the 1970′s, other biological features such as genome stability also seem to be highly associated with evolutionary rates. Rates of genome evolution appear to be closely correlated with levels of species richness. In mammals, a strong correlation between species richness and karyotype diversity was first reported in 1980. The author proposed that: “properties of stable or unstable karyotype may indicate that the cytological factors of importance are all of a submicroscopic nature.” [61]. Indeed, the phylogenetic trees of mammals, frogs and urodeles show significant differences in species richness among the three different taxa when accounting for the fossil record (Figure 1).
Among the three taxa, Urodela have the fewest number of species (816 newts and salamanders; time of emergence: 230 Mya [62]. Anurans are substantially more speciose than salamanders (7,682 frogs and toads; emerging 180 Mya; [63], and Mammalia have a similar number of species (6,495 of which about one third, or 2,276, belong to Rodentia). Mammals first evolved 225 Mya, but experienced a rapid adaptive radiation 65,8 Mya among placentals, much later than the anuran radiation [64]. Hence, salamanders are evolving more slowly than frogs, which are evolving more slowly than mammals [65,66]
The question emerges from these and other observations: what are the submicroscopic factors that might explain the correlations between SR and KD and the manifest differences in SR and species evenness in the respective phylogenetic trees—assuming that those cellular and presumably nuclear factors and mechanisms are genuinely associated with the correlations and their respective differences? If that assumption holds true, to what extent then would those yet unidentified factors contribute to—or contrast with—the prevailing view that most if not all speciation and adaptive radiations are attributable to ecological speciation alone instead of to NARs resulting from DNA damage, mutation and diversification? [16,67]. What these submicroscopic factors might be remains unknown.
4. DNA Damage Detection and Repair Systems (DDR) and Chromatin Structure.
Genomes with larger amounts of functional DNA (number of genes, regulatory sequences, etc.) are expected to have lower mutation rates; yet larger genomes are more prone to DNA damage and mutation. The apparent paradox can be resolved by noting that the eukaryote genome is compartmentalized into two broad and varying forms of chromatin: euchromatin (EC) and heterochromatin (HC) [68]. Euchromatin is characterized by large DNA loops that are more accessible to regulatory enzymes and are more rich in genes. Heterochromatin, facultative or constitutive [69], is more compact, has a much lower gene density and is more refractory to enzymes involved in DNA metabolism (replication, transcription and repair).
This spatial compartmentalization also imposes temporal compartmentalization according to a replication timing (RT) program [70]: EC replicates early during the S-phase of the cell cycle and HC replicates late. Late replicating DNA protects the genome and cell against mitotic catastrophe, or premature entry into the mitotic phase, which would damage unreplicated gene dense EC and cause apoptosis. Late replicating DNA also serves as a substrate for the ATR/ATM checkpoint system that mediates DNA repair by inhibiting the activation of late and/or dormant DNA replication origins until the cell is ready to recover at mid S-phase from DNA damage. The S-phase and G2/M-phase checkpoint proteins Chk1 and Chk2 govern these functions and organize a multi-factorial cell cycle replication timing program.
Importantly, this temporal compartmentalization corresponds to the differential employment of the two main eukaryote DNA repair systems: error free homologous recombination (HR), which operates more efficiently in the open euchromatin that replicates early in S-phase, and error prone non-homologous end-joining (NHEJ), which operates throughout S-phase but dominates in M and G1 phases [71,72,73]. The ratio between HC and NHEJ decreases with genome size across eukaryotes: species with larger genomes rely more heavily on NHEJ than do species with smaller genomes [74]. Consequently, they tend to have much larger introns and higher intron density [75,76].
Thus, individual mutation events, although assumed to be randomly occurring, are not randomly distributed across the genome [77]. Several studies have established that rates of mutation depend highly on the replication timing (RT) of subregions of the eukaryote genome. The RT program therefore serves to limit mutation rates in gene rich EC: mutation rates are significantly higher in late replicating HC [78,79,80,81,82,83,84], suggesting that mutation rates in early and late replicating DNA are anti-correlated to a degree directly proportional to the quantity of late replicating HC relative to early replicating EC. DNA damage prone polymerases, the Y-family of translesion polymerases, also might account for the higher mutation rate in late S-phase. The elevated mutation rates in late replicating DNA in yeast, for example, are suppressed when DNA translesion polymerases are rendered inactive [79].
Not surprisingly, the functional identity of genes—gene ontology—is also unequally distributed across the genome. Essential house keeping genes, required for the survival of all cells, are universally early replicating while adaptive genes, such as the olfactory complex, are generally late replicating and located in or near heterochromatic domains [85,86]. Speciation genes tend to be non-essential in contrast to house-keeping genes [87].
The late replicating status of non-essential mutation prone speciation/adaptive genes remains to be firmly established, but some studies strongly suggest that the epigenome biases mutation [88,89], which might promote the adaptive functions exhibited by immune system genes, which are late replicating, and other ecologically responsive, or sensitive, genes. The generation of genetic diversity by promoting DNA damage in late replicating genes can thus be viewed as analogous to the programmed DNA damaging processes involved in the generation of antibody diversity in the immune system [90].
5. Limb Regeneration, Cell Differentiation, Development and Aging.
Species with large C-values have longer introns and correspondingly slower rates of transcription, a phenomenon known as “intron delay” [91,92]. Consequently, they have much slower cell and life cycles. Other features associated with species with either large or more stable genomes are long maximum lifespans (MLS), slow rates of development and in some cases the ability to regenerate ablated tissue [93]. Salamanders, for example, can regenerate not only limbs but also internal organs including the brain [94]. Tissue regeneration depends on a strong DNA damage response system that converges on the cell cycle checkpoint regulators Chk1 and Chk2: inhibition Chk1 and Chk2 impairs regeneration [95].
Given the role of the heterochromatin-DDR complex in RT, DNA repair, cell cycle regulation, limb regeneration and aging, it is not surprising that an embryonic state of chromatin also facilitates experimental cloning of animals [96]. This might suggest that the limb regeneration and slow aging phenotypes in salamanders are associated with the substantially larger amounts of heterochromatin in their genomes compared to other species with smaller genomes. Obligate neotenes consistently have genomes much larger than metamorphic or direct developing salamanders [97,98]. Larger amounts of heterochromatin might therefore facilitate DNA repair, slow the rate of aging, enhance MLS and retard developmental rate.
DNA and histone methylation are features of heterochromatin, and are associated with developmental genes, gene regulatory regions and the polycomb repressive complex 2, a histone methyl transferase associated with repressed transcriptionally silent facultative herterochromatin and X-chromosme inactivation [14]. Histone methylation also participates indirectly in the DDR [99]. Additionally, epigenetic drift involves the erosion of CpG methylation, and is closely associated with aging: higher densities of CpG methylation buffer against epigenetic drift and extend MLS [51]. Other important chromatin modulators such as Sirt6 are also involved in the HC-DDR complex and influence mutation rates and aging [100]. The link between heterochromatin, genome stability and aging perhaps can be extended to rates of speciation [101,102,103].
When DNA damage occurs, cells face three possible outcomes depending on the amount of damage: 1) checkpoint mediated cell cycle arrest and DNA repair (DDR activation), 2) cellular senescence (aging) and 3) apoptosis (programmed cell death). A fourth fate involves cellular differentiation [104,105]. Apoptosis is an integral feature of both the DDR and the cellular differentiation that drives embryogenesis and development [106]. Chk1 is activated, for example, at the midblastula transition during embryogenesis when the cellular transcription program is switched on [107]; and it acts to extend the cell cycle and initiate cellularization in the developing embryo [108].
It has been claimed that cellular differentiation, a feature driving the evolution of metazoans, emerged as a defense against lethal DNA damage and oncogenesis [105]. The idea is that cellular differentiation is an evolutionary adaptation to DNA damage and a prophylaxis against oncogenesis in metazoans. This raises an interesting question: are rates of evolution constrained by rates of development? Rates of development in salamanders, for example, are constrained by a nucleotypic effect relating to genome size [109]. If so, could speciation rates scale with the timing of the program of differentiation and development in the individual organism?
Additionally, the lower levels of DNA damage and the stronger DDRs associated with higher levels of heterochromatin might contribute to the slower rates of evolution observed in Urodela compared to Anura and Mammalia. Such a relationship is also apparent within the Urodela lineage: species richness at the family level taxonomic clade is negatively correlated with C-value. Although the latter observation remains to be rigorously established, slow aging and longer developmental programs, which result in longer MLS, provide the organism with more time to repair DNA damage, thus promoting the efficiency of DNA repair and enhancing genome stability by reducing mutation/substitution rates.
If mutation rates set rates of speciation as Kimura’s NAR hypothesis proposes and if mutation rates vary substantially across animal and plant lineages, the DDR and HC must play important roles in determining speciation rates across the Tree of Life (Figure 3). Speciation rates might indeed be related to developmental rates [110], a question that has long intrigued evolutionary and developmental biologists. This would suggest that, in more than just a metaphorical sense (though not exactly in a literal sense), “ontogeny recapitulates phylogeny.” [111,112]. It would be interesting, nonetheless, to investigate how rates of speciation and phylogenesis scale with rates of development and ontogenesis should it turn out that the DDR and heterochromatin are in fact limiting for cell cycle progression and mutation [113,114,115,116].
Discussion
This review has attempted to adumbrate some of the various mechanisms in which heterochromatin and DNA repair might play a role in maintaining genome integrity and stability, biological features that are increasingly associated with rates of speciation and adaptive radiations [117]. The central question addressed here concerns to what extent molecular mechanisms mediating genome dynamics determine rates of evolution in parallel to, or even in concert with, gene specific mutation rates.
Mutation rates in vertebrates, for example, are very similar to rates of TE transposition [118,119,120], which is regulated by heterochromatin and ecological variables that shape phenotype plasticity. A role for TE activity in punctuated equilibria has also been suggested [121]. Heterochromatin, however, might not be in and of itself a determining factor of SR and KD, but instead might operate more indirectly through the multiple pathways, both molecular and ecological, that affect and influence evolutionary outcomes. It has now become clear, however, that heterochromatin plays vital regulatory roles in RT, DNA repair, transcription and development. Its role in speciation merits further investigation.
NAR, in its molecular formulation, might imply a biphasic mode of evolution: 1) a lag period of drift involving chromosomal and genome rearrangements in a neutral niche occupied by an ancestor population (stem group), followed by 2) niche diversification and neo-Darwinian positive selection on adaptive genes resulting in ecological speciation (crown group) [122,123] (Figure 4). The fact that synteny is highly conserved in salamanders, frogs and mammals while rates of structural change in genes are fairly constant supports the proposal that karyotypes evolve neutrally whereas the transcriptome and corresponding phenotypes evolve according to positive (and purifying) selection. It is also notable that synteny is correlated with MLS in mammals (unpublished).
Both features, karyotype diversification and genetic diversification, might contribute successively or in tandem (and in concert) to the processes of reproductive isolation and adaptation. Might there then be two distinct molecular clocks determining the mode and tempo of evolution: a gene based molecular clock that sets a constant rate of genetic evolution across lineages, and a genome/junk based molecular clock that sets a given rate of speciation that varies from lineage to lineage? [124,125].
The central tenet of Kimura’s NAR hypothesis relies on the assumption that mutation rates directly influence substitution rates (mutation-substitution balance) and therefore speciation rates. Ecological speciation, in contrast, rests on the assumption that environmental shifts acting on functional DNA alone (or predominantly) determine speciation rates. It has been repeatedly found in every organism examined so far (including salamanders) that substitutions at non-silent sites in gene codons (amino acid substitutions) are correlated with substitutions at silent sites, suggesting that selection acts not only on genes but also on gene locations and regions in the genome (eg. early vs. late replicating DNA, heterochromatin vs. euchromatin) [88].
This raises an interesting, perhaps provocative question: To what extent do mutation rates and DNA repair efficiencies influence, or set, substitution rates—and hence speciation rates—independently of ecological selection? It has been pointed out that “locational selection would have to be realized through the influence of the local mutation rate on the amino acid changing mutation rate” [88]. If this hypothesis is correct—selection based on gene location—and if it is a reflection of the non-random distribution of DNA damage events, it would not be unreasonable to expect that such a relationship/correlation between gene location, DNA damage and DNA repair efficiency would apply not only within genomes, but also across taxa (salamanders vs. frogs vs. mammals) in a manner that sets variations in speciation rates within lineages and explains, at least in part, the striking differences in species richness and evenness observed in the Tree of Life.
Conclusion
What role, if any, non-coding DNA and heterochromatin play in the process of speciation and adaptive radiations remains largely unknown and awaits further investigation into the finer molecular details of genome architecture and DNA metabolism. The growing body of genome wide data from different taxa and the emergence of new bioinformatic tools have opened up the field of the genomics of speciation by allowing for detailed analyses at the levels of the gene, the genome and the transcriptome [126,127,128,129].
Other lines of investigation will further integrate cell cycle regulation and DNA metabolism into the existing and future theoretical models, clarifying the role of DNA metabolism in the process of speciation that is, as of now, incompletely understood. In short, the molecular basis of NAR suggests that the evolutionary outcomes of totipotent cells forming different lineages within a single metazoan organism during development, and the evolutionary outcomes of an individual organism/population forming a new species within a genus during speciation, might be as much a question of chance and genetic drift as it is a question of necessity and ecological adaptation.
References
- Soltis P.S., Folk R.A., Soltis D.E. Darwin review: angiosperm phylogeny and evolutionary radiations. Proc. R. Soc. B.2862019009920190099. [CrossRef]
- Lynch M. Streamlining and simplification of microbial genome architecture. Annu Rev Microbiol. 2006;60:327-49. 10.1146/annurev.micro.60.080805.142300.
- Pellicer J, Hidalgo O, Dodsworth S, Leitch IJ. Genome Size Diversity and Its Impact on the Evolution of Land Plants. Genes (Basel). 2018 Feb 14;9(2):88. [CrossRef]
- Pellicer, J., Fay, M.F., Leitch, I.J. The largest eukaryotic genome of them all?, Botanical Journal of the Linnean Society, Volume 164, Issue 1, September 2010, Pages 10–15, . [CrossRef]
- 5. Alejandro Rodríguez-Gijón, Moritz Buck, Anders F Andersson, Dandan Izabel-Shen, Francisco J A Nascimento, Sarahi L Garcia, Linking prokaryotic genome size variation to metabolic potential and environment, ISME Communications, Volume 3, Issue 1, December 2023, 25, . [CrossRef]
- Hidalgo, O., Pellicer, J., Christenhusz, M., Schneider, A., R. Leitch, A.R., Leitch, I.J. Is There an Upper Limit to Genome Size? Trends in Plant Science, Volume 22, Issue 7, 2017, Pages 567-573.
- Prachumwat A, Li WH. Gene number expansion and contraction in vertebrate genomes with respect to invertebrate genomes. Genome Res. 2008 Feb;18(2):221-32. [CrossRef]
- Demuth JP, Bie TD, Stajich JE, Cristianini N, Hahn MW (2006) The Evolution of Mammalian Gene Families. PLoS ONE 1(1): e85. [CrossRef]
- Damas J, Corbo M, Kim J, Turner-Maier J, Farré M, Larkin DM, Ryder OA, Steiner C, Houck ML, Hall S, Shiue L, Thomas S, Swale T, Daly M, Korlach J, Uliano-Silva M, Mazzoni CJ, Birren BW, Genereux DP, Johnson J, Lindblad-Toh K, Karlsson EK, Nweeia MT, Johnson RN; Zoonomia Consortium; Lewin HA. Evolution of the ancestral mammalian karyotype and syntenic regions. Proc Natl Acad Sci U S A. 2022 Oct 4;119(40):e2209139119. [CrossRef]
- Graphodatsky, A.S., Trifonov, V.A. & Stanyon, R. The genome diversity and karyotype evolution of mammals. Mol Cytogenet 4, 22 (2011). [CrossRef]
- T. RYAN GREGORY, The C-value Enigma in Plants and Animals: A Review of Parallels and an Appeal for Partnership, Annals of Botany, Volume 95, Issue 1, January 2005, Pages 133–146, . [CrossRef]
- Puttick M. N., Clark, J., Donoghue, P. C. J. 2015 Size is not everything: rates of genome size evolution, not C-value, correlate with speciation in angiosperms. Proc. R. Soc. B.2822015228920152289. [CrossRef]
- Voss SR, Kump DK, Putta S, Pauly N, Reynolds A, Henry RJ, Basa S, Walker JA, Smith JJ. Origin of amphibian and avian chromosomes by fission, fusion, and retention of ancestral chromosomes. Genome Res. 2011 Aug;21(8):1306-12. [CrossRef]
- Liu, J., M., Zhou, Q.. Establishment and evolution of heterochromatin. First Published: 04 February 2020, . [CrossRef]
- Kimura M. The neutral theory of molecular evolution: a review of recent evidence. Jpn J Genet. 1991 Aug;66(4):367-86. [CrossRef]
- Schluter D, Conte GL. Genetics and ecological speciation. Proc Natl Acad Sci U S A. 2009 Jun 16;106 Suppl 1(Suppl 1):9955-62. [CrossRef]
- M. Ravinet, R. Faria, R. K. Butlin, J. Galindo, N. Bierne, M. Rafajlović, M. A. F. Noor, B. Mehlig, A. M. Westram, Interpreting the genomic landscape of speciation: a road map for finding barriers to gene flow, Journal of Evolutionary Biology, Volume 30, Issue 8, 1 August 2017, Pages 1450–1477, . [CrossRef]
- The Paradox Behind the Pattern of Rapid Adaptive Radiation: How Can the Speciation Process Sustain Itself Through an Early Burst? Christopher H. Martin and Emilie J. Richards. Annual Review of Ecology, Evolution, and Systematics 2019 50:1, 569-593. [CrossRef]
- 19. Rosemary G Gillespie, Gordon M Bennett, Luc De Meester, Jeffrey L Feder, Robert C Fleischer, Luke J Harmon, Andrew P Hendry, Matthew L Knope, James Mallet, Christopher Martin, Christine E Parent, Austin H Patton, Karin S Pfennig, Daniel Rubinoff, Dolph Schluter, Ole Seehausen, Kerry L Shaw, Elizabeth Stacy, Martin Stervander, James T Stroud, Catherine Wagner, Guinevere O U Wogan, Comparing Adaptive Radiations Across Space, Time, and Taxa, Journal of Heredity, Volume 111, Issue 1, January 2020, Pages 1–20, . [CrossRef]
- Czekanski-Moir JE, Rundell RJ. The Ecology of Nonecological Speciation and Nonadaptive Radiations. Trends Ecol Evol. 2019 May;34(5):400-415. [CrossRef]
- The Next Generation of Adaptive Radiation Studies in Plants John J. Schenk. International Journal of Plant Sciences 2021 182:4, 245-262.
- Kozak K. H., Weisrock D. W., Larson A. 2006Rapid lineage accumulation in a non-adaptive radiation: phylogenetic analysis of diversification rates in eastern North American woodland salamanders (Plethodontidae: Plethodon) Proc. R. Soc. B. 273, 539–546. [CrossRef]
- Joel O. Wertheim, Ben Murrell, Martin D. Smith, Sergei L. Kosakovsky Pond, Konrad Scheffler, RELAX: Detecting Relaxed Selection in a Phylogenetic Framework, Molecular Biology and Evolution, Volume 32, Issue 3, March 2015, Pages 820–832, . [CrossRef]
- Hunt BG, Ometto L, Wurm Y, Shoemaker D, Yi SV, Keller L, Goodisman MA. Relaxed selection is a precursor to the evolution of phenotypic plasticity. Proc Natl Acad Sci U S A. 2011 Sep 20;108(38):15936-41. [CrossRef]
- Persi, E., Wolf, Y., Koonin, E. Positive and strongly relaxed purifying selection drive the evolution of repeats in proteins. Nat Commun 7, 13570 (2016). [CrossRef]
- Lynch M, Conery JS. The origins of genome complexity. Science. 2003 Nov 21;302(5649):1401-4. [CrossRef]
- Fuselli S, Greco S, Biello R, Palmitessa S, Lago M, Meneghetti C, McDougall C, Trucchi E, Rota Stabelli O, Biscotti AM, Schmidt DJ, Roberts DT, Espinoza T, Hughes JM, Ometto L, Gerdol M, Bertorelle G. Relaxation of Natural Selection in the Evolution of the Giant Lungfish Genomes. Mol Biol Evol. 2023 Sep 1;40(9):msad193. [CrossRef]
- Mohlhenrich ER, Mueller RL. Genetic drift and mutational hazard in the evolution of salamander genomic gigantism. Evolution. 2016 Dec;70(12):2865-2878. [CrossRef]
- Ai B, Wang ZS, Ge S. Genome size is not correlated with effective population size in the Oryza species. Evolution. 2012 Oct;66(10):3302-10. [CrossRef]
- Whitney KD, Baack EJ, Hamrick JL, Godt MJ, Barringer BC, Bennett MD, Eckert CG, Goodwillie C, Kalisz S, Leitch IJ, Ross-Ibarra J. A role for nonadaptive processes in plant genome size evolution? Evolution. 2010 Jul;64(7):2097-109. [CrossRef]
- Blommaert J. Genome size evolution: towards new model systems for old questions. Proc Biol Sci. 2020 Aug 26;287(1933):20201441. [CrossRef]
- Bourgeois, Yann, and Stéphane Boissinot. 2019. “On the Population Dynamics of Junk: A Review on the Population Genomics of Transposable Elements” Genes 10, no. 6: 419. [CrossRef]
- Llaurens V, Whibley A, Joron M. Genetic architecture and balancing selection: the life and death of differentiated variants. Mol Ecol. 2017 May;26(9):2430-2448. [CrossRef]
- Ding, G., Hasselmann, M., Huang, J. et al. Global allele polymorphism indicates a high rate of allele genesis at a locus under balancing selection. Heredity 126, 163–177 (2021). [CrossRef]
- Avise JC. Genic heterozygosity and rate of speciation. Paleobiology. 1977; 3(4):422-432. [CrossRef]
- Charlesworth D. Balancing selection and its effects on sequences in nearby genome regions. PLoS Genet. 2006 Apr;2(4):e64. [CrossRef]
- 37. Bárbara D Bitarello, Débora Y C Brandt, Diogo Meyer, Aida M Andrés, Inferring Balancing Selection From Genome-Scale Data, Genome Biology and Evolution, Volume 15, Issue 3, March 2023, evad032, . [CrossRef]
- Harvey MG, Seeholzer GF, Smith BT, Rabosky DL, Cuervo AM, Brumfield RT. Positive association between population genetic differentiation and speciation rates in New World birds. Proc Natl Acad Sci U S A. 2017 Jun 13;114(24):6328-6333. [CrossRef]
- Mani GS, Clarke BC. Mutational order: a major stochastic process in evolution. Proc R Soc Lond B Biol Sci. 1990 May 22;240(1297):29-37. [CrossRef]
- Uyeda JC, Hansen TF, Arnold SJ, Pienaar J. The million-year wait for macroevolutionary bursts. Proc Natl Acad Sci U S A. 2011 Sep 20;108(38):15908-13. [CrossRef]
- Folk RA, Stubbs RL, Mort ME, Cellinese N, Allen JM, Soltis PS, Soltis DE, Guralnick RP. Rates of niche and phenotype evolution lag behind diversification in a temperate radiation. Proc Natl Acad Sci U S A. 2019 May 28; 116(22):10874-10882. [CrossRef]
- Aguilée R, Gascuel F, Lambert A, Ferriere R. Clade diversification dynamics and the biotic and abiotic controls of speciation and extinction rates. Nat Commun. 2018 Aug 1;9(1):3013. [CrossRef]
- Rezazadegan, R., Reidys, C. Degeneracy and genetic assimilation in RNA evolution. BMC Bioinformatics 19, 543 (2018). [CrossRef]
- Mohanty V, Greenbury SF, Sarkany T, Narayanan S, Dingle K, Ahnert SE, Louis AA. Maximum mutational robustness in genotype-phenotype maps follows a self-similar blancmange-like curve. J R Soc Interface. 2023 Jul;20(204):20230169. [CrossRef]
- Manrubia S, Cuesta JA, Aguirre J, Ahnert SE, Altenberg L, Cano AV, Catalán P, Diaz-Uriarte R, Elena SF, García-Martín JA, Hogeweg P, Khatri BS, Krug J, Louis AA, Martin NS, Payne JL, Tarnowski MJ, Weiß M. From genotypes to organisms: State-of-the-art and perspectives of a cornerstone in evolutionary dynamics. Phys Life Rev. 2021 Sep;38:55-106. [CrossRef]
- Whitacre JM, Atamas SP. Degeneracy allows for both apparent homogeneity and diversification in populations. Biosystems. 2012 Oct;110(1):34-42. [CrossRef]
- Paaby AB, Rockman MV. Cryptic genetic variation: evolution’s hidden substrate. Nat Rev Genet. 2014 Apr;15(4):247-58. [CrossRef]
- Wideman JG, Novick A, Muñoz-Gómez SA, Doolittle WF. Neutral evolution of cellular phenotypes. Curr Opin Genet Dev. 2019 Oct;58-59:87-94. [CrossRef]
- Ryszard Maleszka, Paul H. Mason, Andrew B. Barron, Epigenomics and the concept of degeneracy in biological systems, Briefings in Functional Genomics, Volume 13, Issue 3, May 2014, Pages 191-202, . [CrossRef]
- Milosavljevic A. Emerging patterns of epigenomic variation. Trends Genet. 2011 Jun;27(6):242-50. [CrossRef]
- Bertucci-Richter, E.M., Parrott, B.B. The rate of epigenetic drift scales with maximum lifespan across mammals. Nat Commun 14, 7731 (2023). [CrossRef]
- Charlesworth D, Barton NH, Charlesworth B. The sources of adaptive variation. Proc Biol Sci. 2017 May 31;284(1855):20162864. [CrossRef]
- Afanasyeva A, Bockwoldt M, Cooney CR, Heiland I, Gossmann TI. Human long intrinsically disordered protein regions are frequent targets of positive selection. Genome Res. 2018 Jul;28(7):975-982. [CrossRef]
- Wilson AC, Sarich VM, Maxson LR. The importance of gene rearrangement in evolution: evidence from studies on rates of chromosomal, protein, and anatomical evolution. Proc Natl Acad Sci U S A. 1974 Aug; 71(8):3028-30. [CrossRef]
- Bush GL, Case SM, Wilson AC, Patton JL. Rapid speciation and chromosomal evolution in mammals. Proc Natl Acad Sci U S A. 1977 Sep;74(9):3942-6. [CrossRef]
- Maxson LER, Wilson AC. RATES OF MOLECULAR AND CHROMOSOMAL EVOLUTION IN SALAMANDERS. Evolution. 1979 Jun; 33(2):734-740. [CrossRef]
- Coyne, J.A. Correlation between Heterozygosity and Rate of Chromosome Evolution in Animals. The American Naturalist 1984 123:5, 725-729.
- Lynch M, Ali F, Lin T, Wang Y, Ni J, Long H. The divergence of mutation rates and spectra across the Tree of Life. EMBO Rep. 2023 Oct 9; 24(10):e57561. [CrossRef]
- 59. Adam B Roddy, David Alvarez-Ponce, Scott W Roy, Mammals with Small Populations Do Not Exhibit Larger Genomes, Molecular Biology and Evolution, Volume 38, Issue 9, September 2021, Pages 3737–3741, . [CrossRef]
- Amanda R. De La Torre, Zhen Li, Yves Van de Peer, Pär K. Ingvarsson, Contrasting Rates of Molecular Evolution and Patterns of Selection among Gymnosperms and Flowering Plants, Molecular Biology and Evolution, Volume 34, Issue 6, June 2017, Pages 1363–1377, . [CrossRef]
- Bengtsson BO. Rates of karyotype evolution in placental mammals. Hereditas. 1980;92(1):37-47. [CrossRef]
- Schoch RR, Werneburg R, Voigt S. A Triassic stem-salamander from Kyrgyzstan and the origin of salamanders. Proc Natl Acad Sci U S A. 2020 May 26;117(21):11584-11588. [CrossRef]
- Portik DM, Streicher JW, Wiens JJ. Frog phylogeny: A time-calibrated, species-level tree based on hundreds of loci and 5,242 species. Mol Phylogenet Evol. 2023 Nov;188:107907. [CrossRef]
- Hunter P. The rise of the mammals: Fossil discoveries combined with dating advances give insight into the great mammal expansion. EMBO Rep. 2020 Nov 5;21(11):e51617. [CrossRef]
- Bredeson, J.V., Mudd, A.B., Medina-Ruiz, S. et al. Conserved chromatin and repetitive patterns reveal slow genome evolution in frogs. Nat Commun 15, 579 (2024). [CrossRef]
- Liedtke, H.C., Gower, D.J., Wilkinson, M. et al. Macroevolutionary shift in the size of amphibian genomes and the role of life history and climate. Nat Ecol Evol 2, 1792–1799 (2018). [CrossRef]
- Rundell RJ, Price TD. Adaptive radiation, nonadaptive radiation, ecological speciation and nonecological speciation. Trends Ecol Evol. 2009 Jul;24(7):394-9. [CrossRef]
- Janssen A, Colmenares SU, Karpen GH. Heterochromatin: Guardian of the Genome. Annu Rev Cell Dev Biol. 2018 Oct 6;34:265-288. [CrossRef]
- Zylicz, J.J. and Heard, E., Molecular Mechanisms of Facultative Heterochromatin Formation: An X-Chromosome Perspective. Annual Review of Biochemistry, 2020, 59: 255-282, . [CrossRef]
- Nakatani, T., Schauer, T., Altamirano-Pacheco, L. et al. Emergence of replication timing during early mammalian development. Nature 625, 401–409 (2024). [CrossRef]
- Mao Z, Bozzella M, Seluanov A, Gorbunova V. DNA repair by nonhomologous end joining and homologous recombination during cell cycle in human cells. Cell Cycle. 2008 Sep 15;7(18):2902-6. [CrossRef]
- Mao Z, Bozzella M, Seluanov A, Gorbunova V. Comparison of nonhomologous end joining and homologous recombination in human cells. DNA Repair (Amst). 2008 Oct 1;7(10):1765-71. [CrossRef]
- Chen Z, Tyler JK. The Chromatin Landscape Channels DNA Double-Strand Breaks to Distinct Repair Pathways. Front Cell Dev Biol. 2022 Jun 8;10:909696. [CrossRef]
- Sonoda E, Hochegger H, Saberi A, Taniguchi Y, Takeda S. Differential usage of non-homologous end-joining and homologous recombination in double strand break repair. DNA Repair (Amst). 2006 Sep 8;5(9-10):1021-9. [CrossRef]
- Vinogradov, A. Intron–Genome Size Relationship on a Large Evolutionary Scale. J Mol Evol 49, 376–384 (1999). [CrossRef]
- Farlow A, Meduri E, Schlötterer C. DNA double-strand break repair and the evolution of intron density. Trends Genet. 2011 Jan; 27(1):1-6. [CrossRef]
- Kin Chan and Dmitry A. Gordenin Clusters of Multiple Mutations: Incidence and Molecular Mechanisms. Annual Review of Genetics 2015 49:1, 243-267. [CrossRef]
- Stamatoyannopoulos, J., Adzhubei, I., Thurman, R. et al. Human mutation rate associated with DNA replication timing. Nat Genet 41, 393–395 (2009). [CrossRef]
- Lang GI, Murray AW. Mutation rates across budding yeast chromosome VI are correlated with replication timing. Genome Biol Evol. 2011; 3:799-811. [CrossRef]
- Catherine J. Pink, Laurence D. Hurst, Timing of Replication Is a Determinant of Neutral Substitution Rates but Does Not Explain Slow Y Chromosome Evolution in Rodents, Molecular Biology and Evolution, Volume 27, Issue 5, May 2010, Pages 1077–1086, . [CrossRef]
- J. Pink, Laurence D. Hurst, Late-Replicating Domains Have Higher Divergence and Diversity in Drosophila melanogaster, Molecular Biology and Evolution, Volume 29, Issue 2, February 2012, Pages 873–882, . [CrossRef]
- Agier, F., Fischer, G. The Mutational Profile of the Yeast Genome Is Shaped by Replication, Molecular Biology and Evolution, Volume 29, Issue 3, March 2012, Pages 905–913, . [CrossRef]
- Chen CL, Rappailles A, Duquenne L, Huvet M, Guilbaud G, Farinelli L, Audit B, d’Aubenton-Carafa Y, Arneodo A, Hyrien O, Thermes C. Impact of replication timing on non-CpG and CpG substitution rates in mammalian genomes. Genome Res. 2010 Apr;20(4):447-57. [CrossRef]
- Murat, P. et al. DNA replication initiation shapes the mutational landscape and expression of the human genome.Sci. Adv.8,eadd3686(2022). [CrossRef]
- Bomblies K, Peichel CL. Genetics of adaptation. Proc Natl Acad Sci U S A. 2022 Jul 26;119(30):e2122152119. [CrossRef]
- Bracci AN, Dallmann A, Ding Q, Hubisz MJ, Caballero M, Koren A. The evolution of the human DNA replication timing program. Proc Natl Acad Sci U S A. 2023 Mar 7;120(10):e2213896120. [CrossRef]
- Wu, CI., Ting, CT. Genes and speciation. Nat Rev Genet 5, 114–122 (2004). [CrossRef]
- Chuang JH, Li H. Functional bias and spatial organization of genes in mutational hot and cold regions in the human genome. PLoS Biol. 2004 Feb;2(2):E29. [CrossRef]
- Monroe, J.G., Srikant, T., Carbonell-Bejerano, P. et al. Mutation bias reflects natural selection in Arabidopsis thaliana. Nature 602, 101–105 (2022). [CrossRef]
- Cohn, M., Av Mitchison, N., Paul, W. et al. Reflections on the clonal-selection theory. Nat Rev Immunol 7, 823–830 (2007). [CrossRef]
- Heyn P, Kalinka AT, Tomancak P, Neugebauer KM. Introns and gene expression: cellular constraints, transcriptional regulation, and evolutionary consequences. Bioessays. 2015 Feb;37(2):148-54. [CrossRef]
- Abou Chakra M, Isserlin R, Tran TN, Bader GD. Control of tissue development and cell diversity by cell cycle-dependent transcriptional filtering. Elife. 2021 Jul 2;10:e64951. [CrossRef]
- Sessions SK, Wake DB. Forever young: Linking regeneration and genome size in salamanders. Dev Dyn. 2021 Jun;250(6):768-778. [CrossRef]
- Joven A, Elewa A, Simon A. Model systems for regeneration: salamanders. Development. 2019 Jul 22;146(14):dev167700. [CrossRef]
- Sousounis K, Bryant DM, Martinez Fernandez J, Eddy SS, Tsai SL, Gundberg GC, Han J, Courtemanche K, Levin M, Whited JL. Eya2 promotes cell cycle progression by regulating DNA damage response during vertebrate limb regeneration. Elife. 2020 Mar 6;9:e51217. [CrossRef]
- Lemaitre JM, Danis E, Pasero P, Vassetzky Y, Méchali M. Mitotic remodeling of the replicon and chromosome structure. Cell. 2005 Dec 2;123(5):787-801. [CrossRef]
- Gregory TR. Genome size and developmental complexity. Genetica. 2002 May;115(1):131-46. [CrossRef]
- 2023; 5, 98. R Lockridge Mueller, C E Cressler, R S Schwartz, R A Chong, M A Butler, Metamorphosis Imposes Variable Constraints on Genome Expansion through Effects on Development, Integrative Organismal Biology, Volume 5, Issue 1, 2023, obad015, . [CrossRef]
- Gong F, Miller KM. Histone methylation and the DNA damage response. Mutat Res Rev Mutat Res. 2019 Apr-Jun;780:37-47. [CrossRef]
- Tian X, Firsanov D, Zhang Z, Cheng Y, Luo L, Tombline G, Tan R, Simon M, Henderson S, Steffan J, Goldfarb A, Tam J, Zheng K, Cornwell A, Johnson A, Yang JN, Mao Z, Manta B, Dang W, Zhang Z, Vijg J, Wolfe A, Moody K, Kennedy BK, Bohmann D, Gladyshev VN, Seluanov A, Gorbunova V. SIRT6 Is Responsible for More Efficient DNA Double-Strand Break Repair in Long-Lived Species. Cell. 2019 Apr 18;177(3):622-638.e22. [CrossRef]
- Crofts, S.J.C., Latorre-Crespo, E. & Chandra, T. DNA methylation rates scale with maximum lifespan across mammals. Nat Aging 4, 27–32 (2024). [CrossRef]
- Pértille F, Da Silva VH, Johansson AM, Lindström T, Wright D, Coutinho LL, Jensen P, Guerrero-Bosagna C. Mutation dynamics of CpG dinucleotides during a recent event of vertebrate diversification. Epigenetics. 2019 Jul;14(7):685-707. [CrossRef]
- 103. Clare J Venney, Dafni Anastasiadi, Maren Wellenreuther, Louis Bernatchez, The Evolutionary Complexities of DNA Methylation in Animals: From Plasticity to Genetic Evolution, Genome Biology and Evolution, Volume 15, Issue 12, December 2023, evad216, . [CrossRef]
- Sjakste N, Riekstiņa U. DNA damage and repair in differentiation of stem cells and cells of connective cell lineages: A trigger or a complication? Eur J Histochem. 2021 May 3;65(2):3236. [CrossRef]
- Sherman MH, Bassing CH, Teitell MA. Regulation of cell differentiation by the DNA damage response. Trends Cell Biol. 2011 May;21(5):312-9. [CrossRef]
- Meier, P., Finch, A. & Evan, G. Apoptosis in development. Nature 407, 796–801 (2000). [CrossRef]
- Zhang M, Kothari P, Mullins M, Lampson MA. Regulation of zygotic genome activation and DNA damage checkpoint acquisition at the mid-blastula transition. Cell Cycle. 2014;13(24):3828-38. [CrossRef]
- Farrell JA, Shermoen AW, Yuan K, O’Farrell PH. Embryonic onset of late replication requires Cdc25 down-regulation. Genes Dev. 2012 Apr 1;26(7):714-25. [CrossRef]
- Jockusch, E. L. “An Evolutionary Correlate of Genome Size Change in Plethodontid Salamanders.” Proceedings: Biological Sciences, vol. 264, no. 1381, 1997, pp. 597–604. JSTOR, http://www.jstor.org/stable/50551.
- Rabosky, D., Santini, F., Eastman, J. et al. Rates of speciation and morphological evolution are correlated across the largest vertebrate radiation. Nat Commun 4, 1958 (2013). [CrossRef]
- Arthur, W. The emerging conceptual framework of evolutionary developmental biology. Nature. 2002 Feb 14;415(6873):757-64. [CrossRef]
- Uesaka M, Kuratani S, Irie N. The developmental hourglass model and recapitulation: An attempt to integrate the two models. J Exp Zool B Mol Dev Evol. 2022 Jan; 338(1-2):76-86. [CrossRef]
- 113. Chase D Brownstein, Daniel J MacGuigan, Daemin Kim, Oliver Orr, Liandong Yang, Solomon R David, Brian Kreiser, Thomas J Near, The genomic signatures of evolutionary stasis, Evolution, 2024; qpae028,. [CrossRef]
- Herrick J. Genetic variation and DNA replication timing, or why is there late replicating DNA? Evolution. 2011 Nov;65(11):3031-47. [CrossRef]
- Sclavi, B., Herrick, J. [Submitted on 9 Feb 2013] Slow Evolution of rag1 and pomc Genes in Vertebrates with Large Genomes. https://arxiv.org/ftp/arxiv/papers/1302/1302.2182.pdf.
- Sendell-Price, A.T., Tulenko, F.J., Pettersson, M. et al. Low mutation rate in epaulette sharks is consistent with a slow rate of evolution in sharks. Nat Commun 14, 6628 (2023). [CrossRef]
- 117. Madhav Jagannathan, Yukiko M Yamashita, Defective Satellite DNA Clustering into Chromocenters Underlies Hybrid Incompatibility in Drosophila, Molecular Biology and Evolution, Volume 38, Issue 11, November 2021, Pages 4977–4986, . [CrossRef]
- Ricci, M., Peona, V., Guichard, E. et al. Transposable Elements Activity is Positively Related to Rate of Speciation in Mammals. J Mol Evol 86, 303–310 (2018). [CrossRef]
- Quadrana, L., Etcheverry, M., Gilly, A. et al. Transposition favors the generation of large effect mutations that may facilitate rapid adaption. Nat Commun 10, 3421 (2019). [CrossRef]
- Baduel P, Quadrana L, Hunter B, Bomblies K, Colot V. Relaxed purifying selection in autopolyploids drives transposable element over-accumulation which provides variants for local adaptation. Nat Commun. 2019 Dec 20;10(1):5818. [CrossRef]
- Zeh DW, Zeh JA, Ishida Y. Transposable elements and an epigenetic basis for punctuated equilibria. Bioessays. 2009 Jul;31(7):715-26. [CrossRef]
- Halliday TJD, Dos Reis M, Tamuri AU, Ferguson-Gow H, Yang Z, Goswami A. Rapid morphological evolution in placental mammals post-dates the origin of the crown group. Proc Biol Sci. 2019 Mar 13;286(1898):20182418. [CrossRef]
- Budd GE, Mann RP. The dynamics of stem and crown groups. Sci Adv. 2020 Feb 19;6(8):eaaz1626. [CrossRef]
- Kumar, S. Molecular clocks: four decades of evolution. Nat Rev Genet 6, 654–662 (2005). [CrossRef]
- Martin, C.F., Gordon, R. Differentiation trees, a junk DNA molecular clock, and the evolution of neoteny in salamanders. Volume8, Issue3 May 1995. Pages 339-354.
- Bock DG, Cai Z, Elphinstone C, González-Segovia E, Hirabayashi K, Huang K, Keais GL, Kim A, Owens GL, Rieseberg LH. Genomics of plant speciation. Plant Commun. 2023 Sep 11;4(5):100599. [CrossRef]
- Hughes SE, Hawley RS. Heterochromatin: a rapidly evolving species barrier. PLoS Biol. 2009 Oct;7(10):e1000233. [CrossRef]
- Wolfsberger, W.W.; Battistuzzi, F.U.; Oleksyk, T.K. Genomics of Adaptation and Speciation. Genes 2022, 13, 1187. [CrossRef]
- C Ryan Campbell, J W Poelstra, Anne D Yoder, What is Speciation Genomics? The roles of ecology, gene flow, and genomic architecture in the formation of species, Biological Journal of the Linnean Society, Volume 124, Issue 4, August 2018, Pages 561–583, . [CrossRef]
Figure 1.
Species Richness and Evenness in Urodela. The black triangles represent the species richness of the 10 salamander families. Species richness varies widely across the respective salamander lineages and their distributions are highly uneven. From [115].
Figure 1.
Species Richness and Evenness in Urodela. The black triangles represent the species richness of the 10 salamander families. Species richness varies widely across the respective salamander lineages and their distributions are highly uneven. From [115].
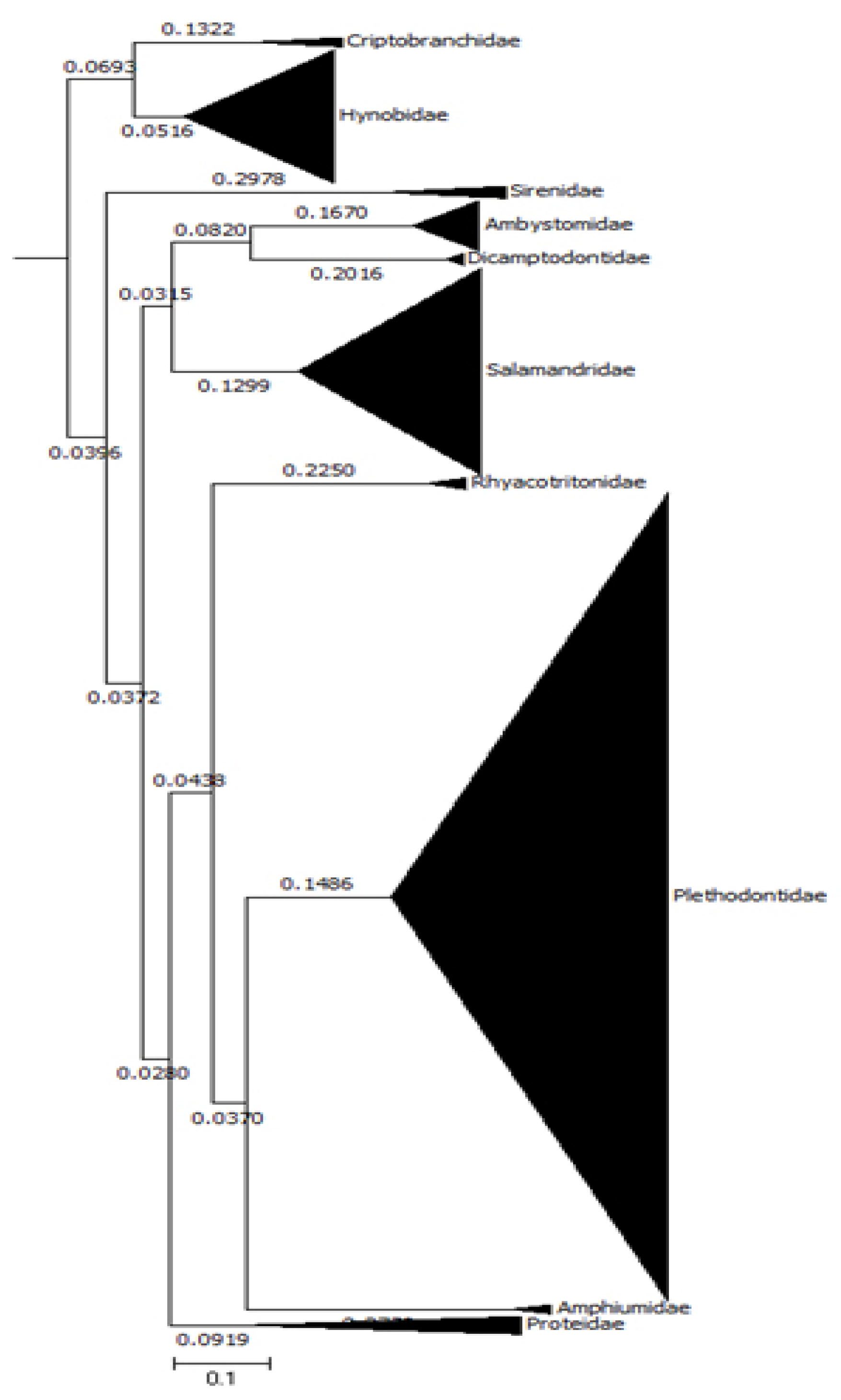
Figure 2.
Salamanders have relatively slower rates of molecular evolution compared to frogs and fish. The box plot reveals the time in million years to accumulate the same average amount of genetic change (Myr/1% dif). The numbers refer to average C-values (pg) for the respective lineages. From [115].
Figure 2.
Salamanders have relatively slower rates of molecular evolution compared to frogs and fish. The box plot reveals the time in million years to accumulate the same average amount of genetic change (Myr/1% dif). The numbers refer to average C-values (pg) for the respective lineages. From [115].
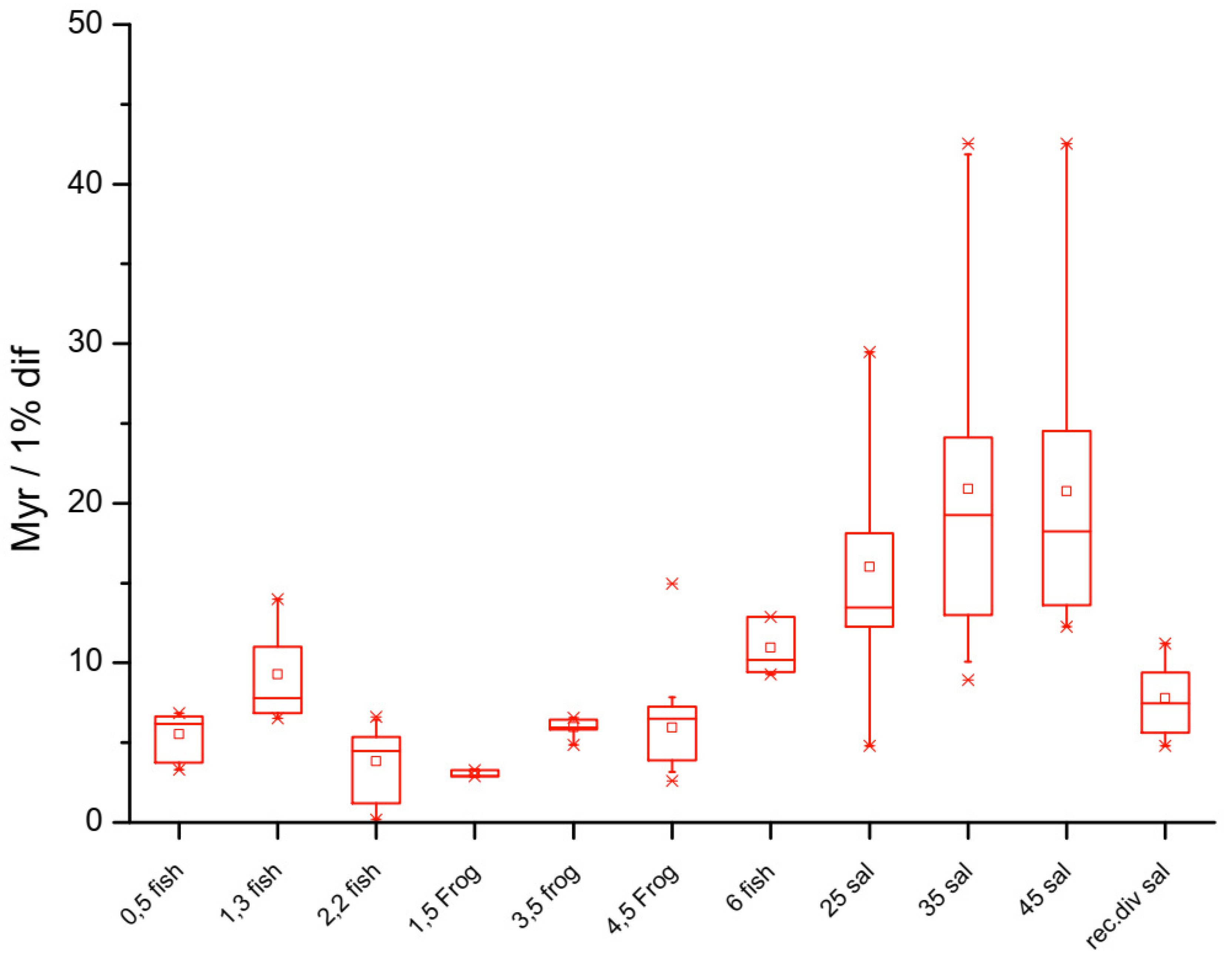
Figure 3.
Box plots of rates of silent site substitution (dS) in the fish lineage. A clear genome size effect on mutation rates is apparent across the lineages comprising skates and rays, which have the largest fish genomes, and puffer fish, which have the smallest fish genomes. The inset log transformed data indicates a power law relationship between evolutionary rates and genome size across these samples. The exponent is -0.39, suggesting significantly different modes of evolution in fish with small genomes compared to fish with larger genomes; perhaps because of the slower rates of DNA loss in species with larger genomes and a corresponding differential dependence on DNA repair systems between species with large versus small genomes (small C-value: HR > NHEJ; large C-value: HR < NHEJ). See [115].
Figure 3.
Box plots of rates of silent site substitution (dS) in the fish lineage. A clear genome size effect on mutation rates is apparent across the lineages comprising skates and rays, which have the largest fish genomes, and puffer fish, which have the smallest fish genomes. The inset log transformed data indicates a power law relationship between evolutionary rates and genome size across these samples. The exponent is -0.39, suggesting significantly different modes of evolution in fish with small genomes compared to fish with larger genomes; perhaps because of the slower rates of DNA loss in species with larger genomes and a corresponding differential dependence on DNA repair systems between species with large versus small genomes (small C-value: HR > NHEJ; large C-value: HR < NHEJ). See [115].
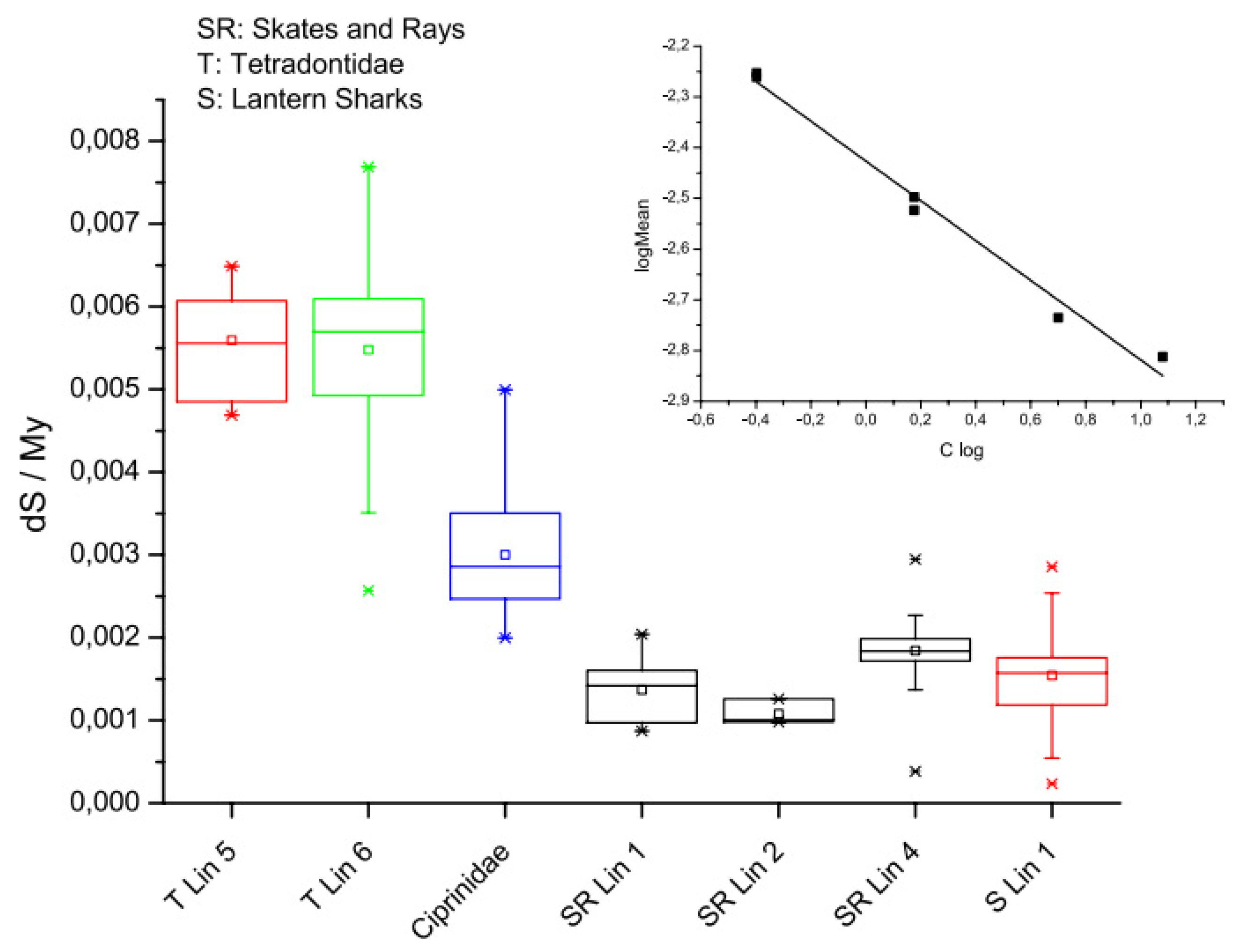
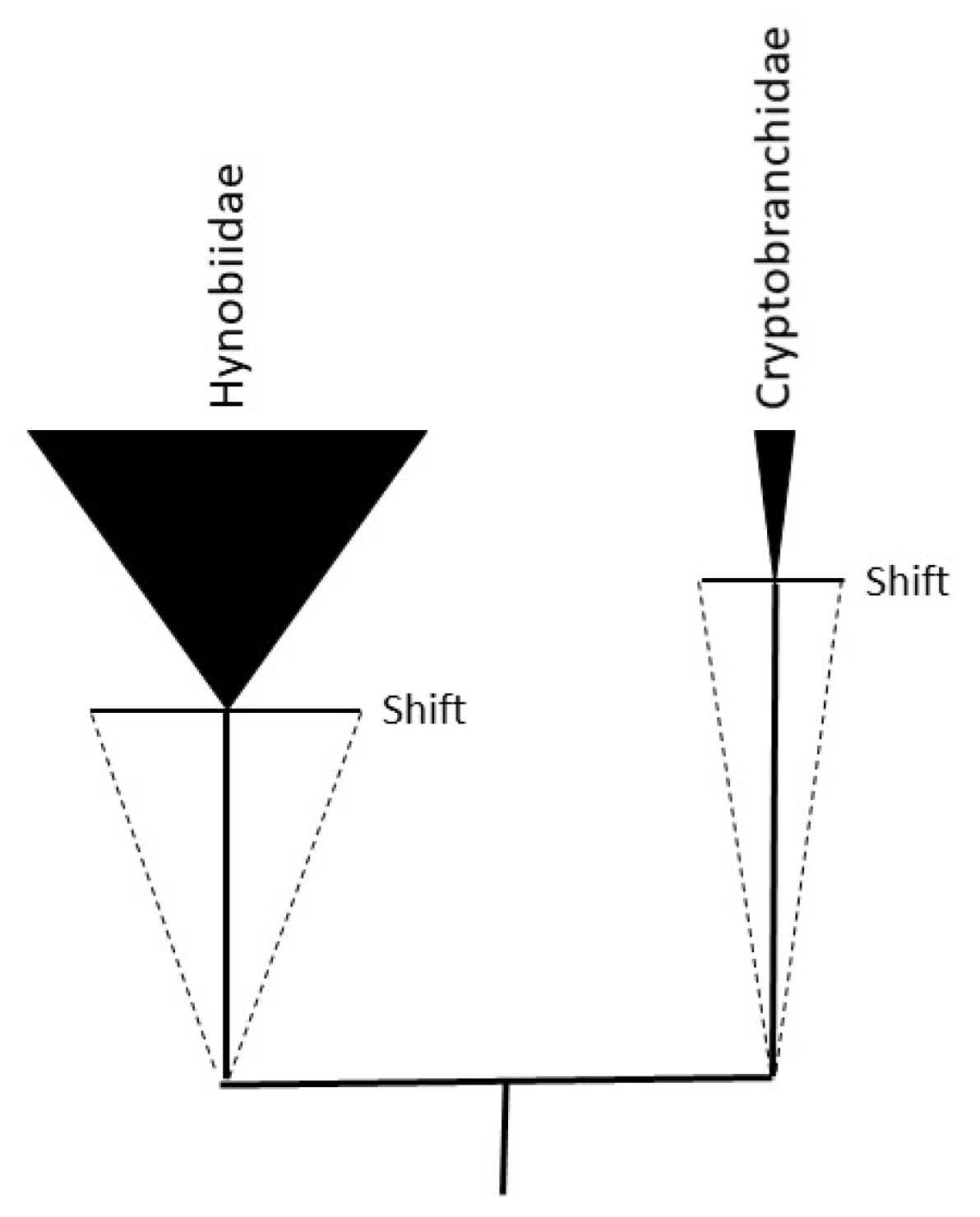
Figure 4.
Hypothetical Biphasic Model of Stem and Crown Group dynamics. The black triangles represent extant crown group family species richness (salamanders). The dotted triangles represent extinct stem group species richness. The Cryptobranchidae, for example, are evolving more slowly over evolutionary time (slope of dotted line; stem to crown age) than are the sister clade of Hynobiidae. It is assumed here that speciation occurs in a predominantly neutral niche mode (neutral adaptive radiation) in the ancestral population until an environmental crisis, or shift, drives ecological speciation (adaptive radiation). Both drift and selection, however, are expected to shape simultaneously evolutionary paths. If karyotype diversity evolves neutrally (genetic drift) during an ancestral phase of evolution (dotted triangles), then the rate of karyotype diversification might be greater than or at least equal to the rate of genetic diversification (rate KD >/= rate GD): more than one genotype, for example, specifying a single phenotype. If an environmental shift applies selection pressure on the diversified karyotypes, a transition might take place where selection pressure acts principally, but not exclusively, on genes (rate of GD > rate of KD). The figure depicts one of multiple cycles generating extant species richness (crown group). The ancestral karyotypes surviving the post crisis/Shift will contribute proportionally to the karyotype diversity in the crown group until those ancestors go extinct (Gause’s principle). It should be noted that “living fossils”—the notion that stem group species persist into extant crown group species—is not assumed in this model: rates of molecular evolution over time (molecular clock) will result in crown group species that are descended—and genetically distinct—from extinct stem group species even in the absence of identifiable morphological or phenotypic change. Figure adapted from [123].
Figure 4.
Hypothetical Biphasic Model of Stem and Crown Group dynamics. The black triangles represent extant crown group family species richness (salamanders). The dotted triangles represent extinct stem group species richness. The Cryptobranchidae, for example, are evolving more slowly over evolutionary time (slope of dotted line; stem to crown age) than are the sister clade of Hynobiidae. It is assumed here that speciation occurs in a predominantly neutral niche mode (neutral adaptive radiation) in the ancestral population until an environmental crisis, or shift, drives ecological speciation (adaptive radiation). Both drift and selection, however, are expected to shape simultaneously evolutionary paths. If karyotype diversity evolves neutrally (genetic drift) during an ancestral phase of evolution (dotted triangles), then the rate of karyotype diversification might be greater than or at least equal to the rate of genetic diversification (rate KD >/= rate GD): more than one genotype, for example, specifying a single phenotype. If an environmental shift applies selection pressure on the diversified karyotypes, a transition might take place where selection pressure acts principally, but not exclusively, on genes (rate of GD > rate of KD). The figure depicts one of multiple cycles generating extant species richness (crown group). The ancestral karyotypes surviving the post crisis/Shift will contribute proportionally to the karyotype diversity in the crown group until those ancestors go extinct (Gause’s principle). It should be noted that “living fossils”—the notion that stem group species persist into extant crown group species—is not assumed in this model: rates of molecular evolution over time (molecular clock) will result in crown group species that are descended—and genetically distinct—from extinct stem group species even in the absence of identifiable morphological or phenotypic change. Figure adapted from [123].
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



