
Submitted:
18 March 2024
Posted:
18 March 2024
You are already at the latest version
Abstract
Regulation of the diet in terms of the content of soluble and insoluble fibre is of great importance from the point of view of dietetics, microbiology of the gastrointestinal tract and immunology of the organism. Dietary fibre plays an important role in maintaining the balance between the con-tinuity of gut mucus production (intestinal mucosa), the stability of the intestinal microbiome composition, the regeneration of the intestinal epithelium and the immune status within the gut-associated lymphoid tissue (GALT) are played by pectin and their metabolic products in the form of pectin oligosaccharides. The main purpose of this review was to focus on pectin, their functions, their impact on the human's health and on some of the most promising prebiotics due to their solubility in the gastrointestinal tract in the form of pectin oligosaccharides. They have the properties of pectin, but have also additional features and postbiotics obtained in fermentation process can affect the intestinal microbiome and GALT-related immunity. They can be obtained from plant agro-wastes.
Keywords:
dietary fibre
; pectin
; pectin oligosaccharides
; microbiome
; GALT
1. Introduction
Dietary fibre is one of the components of food products. The foods richest in this dietary ingredient include: fruits, vegetables, cereals, nuts, beans, and algae [1,2,3]. Important source of dietary fibre are also fruits and vegetable by-products [4]. The classic definition of dietary fibre defined it as ‘plant parts that are resistant to the digestive enzymes of humans and animals’, mainly concerned with structural carbohydrates and lignin [5,6] However, in the case of humans and monogastric animals, a more current definition divides dietary fibre (TDF) into soluble (SDF - viscous or fermentable, such as e.g. pectin) undergoing the process of complete or partial fermentation mainly in the large intestine. Insoluble fibre (IDF) can be also subject only partially fermentation process conducted by Lactobacillus spp. or Bacillus spp. strains [7,8,9]. However SDF is easier fermented than IDF, that is why cellulose, lignin or other insoluble elements of dietary fibre mainly functionate as a ballast and having a beneficial effect on intestinal peristalsis, as well as a "diluent of the digestive content" limiting the absorption of nutrients from the gastrointestinal lumen [10,11]. Table 1 presents physical properties of five individual carbohydrates [12,13,14,15,16,17]. Other important factor is particle size of dietary fibre which also can have influence on gut fermentation and SCFA profile [18].
According to the conception of functional fibre it is defined as a nondigestible carbohydrates found in fibre supplements which must show clinical evidence of a health benefit [19]. Dietary fibre can also be supplied as a result of supplementation and have prebiotic properties, act as a ballast or diluent and can also increase the viscosity of the chyme, reducing the absorption of nutrients [20,21]. This type of fibre containing sources for production fructo oligosaccharides (FOS) and pectin oligosaccharides (POS), can be obtained from by-products of the agri-food industry [22,23,24,25]. Pectin plays an important role during the inflammatory process in the gut, but its effect is also important when this process not occurred in terms of keeping overall gut health [26]. Long-term pectin supplementation modulates gut microbiome composition and affect positively on health [27,28]. From the other side pectin is able to modified redox potential which is important factor in case of metabolic and bacterial activity in gut [29]. Additionally last researches emphasize better bioactivities modified (with changes in functional groups) then natural pectin [30]. Nowadays interest in pectin, and POS, has increased significantly in recent years due to their mucus gut layer modulation properties and the prebiotic properties of pectic oligosaccharides utilized selectively mainly by Eubacterium eilgens and Faecalibacterium prausnitzi [31,32]. They consequently affects the maintenance of gut mucus layer continuity and counteracting inflammation of the intestinal epithelium, preventing the uncontrolled increase in the number of pathogens and their adhesion to the epithelial receptors [33]. This process maintaining the intestinal barrier and the efficient action of gut-associated lymphoid tissue (GALT), which affects the immunity of the entire organism human and animals [34,35,36].
2. Dietary Fibre and Its Importance in the Diet
2.1. Dietary Fibre Consumption
Due to the significant degree of urbanization of society and economic growth, many countries are experiencing changes in dietary habits towards the consumption of high-calorie food rich in refined carbohydrates and fats and low in dietary fibre [37]. This type of diet significantly increases the incidence of lifestyle diseases in high-income group of people without awareness from nutritional knowledge and choosing quick prepared meals what in consequence increases the costs of hospitalization in society [38,39].
Dietary fibre consumption varies significantly depending on the degree of industrialization of a given region of the world. Data up to 2015 indicate that the average dietary fibre intake by adults is 12-18 g/day in the USA, 14 g/day in the United Kingdom and 16-29 g/day in Europe [40]. However, in the last decade dietary patterns in development countries changes towards a significant increase in the consumption of processed food due to its lower costs, especially due to the savings in time needed to prepare this type of meal in home [41,42]. This is indicated by greater diversity of the microbiome intestinal in case of increasing the intake of dietary fibre in the diet, but also leading to insulin resistance and antioxidant stress when diet contains high amount of sucrose or refined carbohydrates [43]. The consumption of protein and animal fat in meat increases the number of Bacteroides in its composition, in turn, the high share of dietary fibre in agrarian societies affects the growth of Prevotella sp. [44]. This is especially easy to notice between populations from industrialized and non-industrialized areas [45].
2.2. Properties of Dietary Fibre
The physiological and biochemical properties of dietary fibre in the small and large intestine have important implications for public health in a given population [46]. The most important include: solubility in water, influence on the viscosity of food content, increasing the volume through water absorption, influence on the absorption of nutrients, ability to ferment and binding other chemical compounds [39,47]. As a result of this, the action of dietary fibre in the digestive tract of humans and animals has a number of benefits: reducing cholesterol levels, lowering blood glucose levels, gut transit time, maintaining the health of the digestive tract by influencing peristalsis and mucus, having a positive effect on the bioavailability of calcium, microbial specificity and gut-associated immune function lymphoid tissue (GALT) [46,48].
2.3. Dietary Fibre in Diet and Its Effect on a Health
A diet low in dietary fibre favours the occurrence of inflammatory bowel disease, colon cancer, allergies, autoimmune diseases and obesity, which can be prevented by including an appropriate level of dietary fibre in the diet [49]. This ingredient contained in food affects the proper functioning of the digestive tract, maintaining the continuity of the epithelial structure and the balance of the digestive tract microbiome [45]. As a result, it affects the body's immunity, playing an important role in maintaining the proper functioning of the digestive tract and, therefore, good health of the patient [50]. In addition to the above-mentioned advantages of its use in the diet, dietary fibre also has a number of benefits, reducing the risk of developing ischemic heart disease, stroke, hypertension, diabetes, obesity and digestive problems [51]. Moreover, dietary fibre reduces the concentration of fats in the blood plasma and the level of glucose in the blood, which also has a positive effect on blood pressure [52].
Research by Thompson et al. [53] indicate that the avocado is an important source of dietary fibre in the diet, especially of overweight and obese adults. In addition to the fibre, which in the case of avocados in the diet (175 g in men and 140 g in women) increased the concentration of metabolites of microbial origin in faeces over a 12-week period, it also contains MUFA acids that reduce the pool of bile acids in faeces. As a consequence, regular consumption of avocado in the diet reduces the body weight of overweight and obese people and increases the number of commensal and symbiotic bacteria responsible for the production of postbiotics, including butyric acid, which is important in the process of nourishing the intestinal epithelium and preventing colon disease [53,54]. Walnuts and walnut meals rich in SDF and unsaturated fatty acids have a similar effect on increasing the number of microorganisms producing butyric acid [55,56]. Moreover, like avocados, they lower the state of pro-inflammatory secondary bile acids and LDL cholesterol, reducing the risk of metabolic syndrome. Additionally, Wang et al. [57] in their research, a predictive tool based on the deep learning method based on the metabolic response predictor using Multilayer Perceptrons (McMLP). It allows for more accurate diet selection compared to previously used machine learning methods for a given person, depending on the type of dietary fibre and unsaturated fatty acids content, based on information from the microbiome and metabolome, which will allow in the future to more precisely reduce the risk of obesity and metabolic problems with bound by it.
3. Types of Dietary Fibre and the Specificity of Their Action
3.1. Dietary Fibre Types and Its Action in Gastrointestinal Tract
Dietary fibre is divided into two groups based on solubility. Insoluble fibre consists of cellulose, lignin and part of hemicelluloses. IDF is insoluble in water and slightly fermentable, but mainly is responsible for accelerating the rate of content flow in the digestive tract and increases the frequency of peristaltic movements in the intestines [16]. Dietary fibre and its soluble fraction also influence the body's immune functions by stabilizing the mucus layer in the intestines, increase of flora diversity and reduce intestinal inflammation [58]. Goblet cells of the intestinal epithelium are responsible for the production of mucin, which has the form of a hydrated gel, which is a component of the mucus that prevents most bacteria from lumen to entering through the layer formed by it and contacting the receptors of the intestinal epithelium [59,60]. When the composition of the intestinal microbiome is disturbed and the barrier mechanism of the epithelium in the form of mucus disappears, we talk about "leaky gut” and the associated inflammation [61]. An important role in the process of stabilizing the protective barriers within the GALT is played by substances produced by microorganisms that restore the stabilization of the mucus layer, collectively called postbiotics [62,63].
3.2. Soluble Dietary Fibre and Its Action in Hindgut
Soluble fibre consists of pectin, gums and mucilage. It is characterized by very good solubility in water and is easily fermented in the gastrointestinal tract [64]. The viscosity effect of chyme caused by soluble dietary fibre limits the access of digestive enzymes to food/feed particles, as a result of which, in combination with the increased amount of insoluble fibre in the diet [65]. Consequently by this the phenomenon of limiting the absorption of nutrients is strengthened, which in combination with physical activity allows to counteract the occurrence of overweight or promotes weight reduction when a given person is overweight or obese [66,67].
Looking at this fact from the point of view of chemical compounds, cellulose, as an insoluble dietary fibre, is poorly fermented by the bacteria of the large intestine due to the linear structure of the chain composed of glucose residues of the molecules of this compound, while pectin, especially in highly branched molecules, is well fermentable [68]. Although, on the other hand, soluble fibre, depending on the type of compounds that compose it, may have a different degree of fermentation and then it tends to increase the viscosity of the food content, which will not be a beneficial solution when used in prebiotic preparations [3]. Table 2 presents content of IDF and SDF in food and amount of those types of dietary fibre in percentages [64,69,70,71,72,73].
That makes it an effective remedy in the fight against constipation, and is also a diluent of the diet, which means that, per unit volume, it contains a smaller amount of nutrients available to the body [74,75]. On the other hand, the appropriate level of soluble and insoluble dietary fibre is responsible for slowing down the flow of food and the feeling of satiety.]. That affects intestinal peristalsis and additionally limits the absorption of nutrients, preventing overweight and obesity and reducing the level of cholesterol in the blood serum, especially the LDL fraction [76,77]. Figure 1 presents classification of dietary fibre proposed by EFSA with added kinds of resistant starch from Cione et al. [78,79].
Soluble dietary fibre is characterized by properties that increase the viscosity of the food content, which results in a decrease in the absorption of nutrients from the gastrointestinal tract [80]. Moreover, this type of fibre has hypocholesterolemic properties related to limiting the digestion and absorption of fat and limiting the process of cholesterol synthesis in the liver through the production of propionic acid and other bacterial transformation products by microorganisms and as a result of the increase in the viscosity of the food content and its effect on the level of insulin and other hormones [6].
3.3. SDF and Changes in Microbiome of Hindgut
Additionally microbiome using SDF with a prebiotic effect increases the concentration of SCFA, what resulting in a lower pH and the production of end products in the form of butyrate, which influences the growth of epithelial cells, and soluble fibre is able to influence the thickness of the mucous layer, which makes it a favourable medium for the existence of symbiotic bacteria and facilitates their adhesion to epithelial receptors [81,82].
Synthesis of SCFA by microorganisms affects the stabilization of the microbiome in a given region of the gastrointestinal tract, prevents disruption of the continuity of the epithelial structure and supports the action of GALT [83,84]. A high level of dietary fibre, including soluble fibre in the diet, prevents the development of inflammation bowel disease [85].
Dietary fibre affects the growth of specific groups of microorganisms and serves as a substrate for the production of short-chain fatty acids SCFA, which play an important role in metabolism and immune processes and are included in the definition of postbiotics [50,86]. Dysbiosis in the intestinal microbiome environment may disturb the host's interaction with the microbiome, limit the thickness and quality of the mucous layer and contribute to the development of pathogenic microorganisms, as well as lead to the occurrence of a disease process [87]. Dietary fibre has an impact on maintaining the balance of the microbiome in the gastrointestinal tract of humans and animals and is necessity in rebuilt its status [88,89].
4. Pectin Structure and Their Properties
4.1. Occurrence of Pectin and Its Amount in Various Fruits and By-Products
Water-soluble pectin is an important element of dietary fibre, playing an important role in modulating the immune barrier of the gastrointestinal tract. They are isolated from the cell walls of fruits and vegetables and are found mainly in the peel or shell of: apples, citrus fruits, and potatoes, passion fruit, mango, bergamot, olives and the exception is sugar beets [90,91,92,93,94,95]. However, the share of pectin in individual plants varies and also depends on the plant parts that are taken into account during the analysis and used for the industrial production of pectin or POS (Figure 2) [96,97].
Orange peel contains 30% pectin, apple peel 15%, and onion peel 12% [14,98]. In turn, raw sugar beets contain approximately 18% pectin, 19% cellulose, 28% hemicelluloses and 8% total protein [99]. Mango peel contains 5-11% of pectin [100]. Of course, taking into account the possible prebiotic effect of pectin or oligosaccharides with typically prebiotic properties are isolated from it. They do not have a viscous effect, do not form gels, but are highly fermentable and affect the profile of fatty acids in the colon [101].
Citrus peel is a rich source of pectin and is also a sustainable source of raw material for the production of high-quality pectin and its derivatives [102]. The production of citrus products is associated with the formation of a large amount of by-products in the form of peels and seeds (up to 50% of the fruit weight), which can be used to produce pectic oligosaccharides with prebiotic properties [22]. In turn, during the production of mango products, peel and seeds constitute 35-60% pectin, which can also be used to produce pectic oligosaccharides, especially due to the fact that peel has antioxidant properties and low fat and protein content [103]. Wande et al. [104] after their analysis also confirm the presence of several types of POS in different concentrations in pomelo peel. Gamonpilas et al. [105] after analysing the solubility of dietary fibre found: 86 g TDF/100 g DM, 20 g SDF/100 g DM and 66 g IDF/100 g DM. Valuable source of pectin are also by-products from avocado (Persea americana), from which in biorefineries this product is obtained with efficiency from 75 to 85% [106].
4.2. Pectin in Cell Wall
Pectin is a component of the cell wall and are located between cellulose and hemicellulose microfibrils which support the functioning of the cell wall by combating external stresses, give it elasticity together with hemicelluloses, which allows it to withstand turgor pressure and maintain the cell shape [107,108]. They act as cementing substances in the cell wall, and thanks to their gelling properties, pectin can reduce the rate of gastric emptying and influence the rate of content flow in the small intestine [67]. Like gums and mucilages, pectin are substances that are soluble in water and are easily fermentable in the large intestine [68]. Pectin has the ability to bind metals in the digestive tract, limiting their absorption from light, thus removing heavy metals from the body in urine, reducing the absorption of lead into the blood and lowering the level of strontium in bones and blood, which may be of great importance in people living in industrial or post-industrial areas [109,110].
4.3. Structure of Pectin
Pectin are polymers composed mainly of homogalacturonan, rhamnogalacturonan I, rhamnogalacturonan II and xylogalacturonan [111]. These compounds belong to heteropolysaccharides consisting primarily of galacturonic acid ( GaIA ) residues connected by α-1,4 bonds [112]. Pectin molecules consist of approximately 70% of glucuronic acid residues which is a part of hairy and smooth regions in molecule (Figure 3). The remaining part are branched elements: xylogalacturonan, apiogalacturonan, rhamnogalacturonan I. Those branched structures additionally consisting of galactose, arabinose, rhamnose molecules, and rhamnogalacturonan II. Molecules in the side chains are composed of: rhamnose, apiose, galacturonic acid, glucuronic acid, fructose, 3-deoxy-d-lyxo-2-heptulosaric acid (Dha), 3-deoxy-d-manno-2-octulosonic acid (Kdo), acetyl and methyl esters. Due to the degree of branching, pectin with low and high dry matter content are distinguished.
Pectin is broken down by the action of bacterial pectinase, which is a mixture of endo and exoenzymes:
- polygalactouronases and polymethylgalactouronases hydrolyzing α-1,4-glycosidic bonds of pectin molecules,
- polygalactouronate lyases and polymethylgalactouronase Lyases break down α-1,4-glycosidic bonds by a trans-elimination mechanism
Homogalacturonan is digested with the participation of polygalactouronase , pectin methyl esterase, pectin lyase and pectin acetyl esterase. RG I is degrade by RG lyase, hydrolase and RG acetylesterase. Pectin galactan by β-1,4-galactanase, β-1,6-galactanase and α- arabinosidase. Arabinogalactan is degrade by the action of β-1,3-galactanase, β-1,6-galactanase, α-(1,5) arabinanase and α- L - arabinofuranosidase. Arabinate in the case of α-(1,5) arabinanase and α- L -arabinofuranosidase activity towards to RG II is not precisely defined in terms of families of enzymes that degrade its structures [116].
Pectin consists of linear α-1,4-linked D-galacturonic acid (homogalacturonan) segments and branched rhamnogalacturonan segments [117]. Homogalacturonan pectin are most often isolated from: sunflowers, citrus fruits, apples and rice endosperm cell walls. Homogalacturonan chains in sugar beet pectin are shorter compared to those found in citrus fruits and apples, but beet pulp has a higher share of type I rhamnogalacturonan than apple pomace and citrus peels [118]. The desirable properties of citrus pectin are related to the high degree of branching of their structure, heterogeneous distribution of the galacturonic acid residue and the presence of neutral sugars and acetyl groups in the side chains [102]. Studies in humans and animals have shown that pectin protects intestinal epithelial barrier indirectly by the modulation of microbiome and its activity, and directly blocking Tool-like receptor 2-1 by which preventing doxorubicin-induced ileitis [119,120]. Additionally products of it degradation, especially butyrate can protect and repair intestinal barrier function controlling mechanism synthesis of junctional proteins and molecular cascade of their secretion [36,121].
Pectin is one of the most important hydrocolloids, primarily in the food industry, but it is also used in other sectors of the economy, including pharmaceutical applications for the production of medicines and hydrocolloid dressings [122]. From the point of view of industry and raw materials for further processing, the methoxyl content in such components is important. Citrus and apple pectin contain over 50% of this compound, while sunflower pectin has a low content of this ingredient [123]. Most natural pectin contains pectin with a high methoxyl content with a level of esterification (HM, above 50%), while processed pectin are dominated by pectin with a low methoxyl content (LM, below 50%) due to the fact that it is easier to obtain homogalacturonan in the technological process , as the unbranched part pectin molecules [124,125]. HMs are used as gelling substances in jams, marmalades and jellies, while LMs mainly act as stabilizers in acidified silent drinks, sour dairy products and mixtures of fruit juice and milk [126].
There is also a division according to the degree of branching of pectin into "smooth" and "hairy", corresponding to unbranched and branched forms, respectively [127]. Commercially produced pectin is rich in smooth pectin and is widely used in the food and pharmaceutical industries due to its excellent hydrocolloid properties [128]. It is produced mainly from citrus peels, beet pulp and apple pomace [127]. However, recent studies indicate that the branched form has a beneficial effect on the human microbiome by increasing the number of Bifidobacterium spp., Lactobacillus spp., Faecalibacterium spp. and inhibiting the adhesion of pathogenic bacteria and toxins [129]. This type of form is difficult to obtain in the technological process, but it has greater prebiotic potential compared to pectin [93,130].
5. Prebiotics
5.1. Oligosaccharides
Oligomers are obtained on an industrial scale from dietary fibre and have great potential for use in various industries. The high bioactivity of this type of compounds creates the possibility of their use in the production of foodstuffs and medicines, cosmetology, prebiotics and symbiotics , antioxidants, they can also be used as a source of anticarcinogenic ingredients [99,131]. Enzymatic hydrolysis of pectin using pectin lyase results in obtaining POS with various structures and degrees of polymerization [132,133]. However, in the case of beet pulp, better results by 20% in terms of the amount of POS obtained are obtained in the case of hydrothermal processing [31].
The most commonly used to modulate the body's immunity and microbiome are: FOS, GOS, XOS [134]. In recent years, attention has also been paid to other water-soluble oligosaccharides with probiotic properties: GalOS, COS and the previously mentioned POS [135]. POS is an addition to dietary fibre in the form of functional fibre. It is extracted in an industrial process towards selected beneficial chemical compounds contained in pectin, which affect the health of the organism [136]. The functional fibre includes: vicus fibre - lowering glucose and cholesterol levels and fermentable fibre - affecting colon health and insoluble fibre stimulating peristalsis and reducing the number of constipations [137]. Research by Kang et al. [138] confirms the beneficial effect of pectic oligosaccharides on reducing the level of triglycerides in blood serum and total cholesterol and LDL fraction in the blood.
5.2. Pectin Oligosaccharides
Pectin oligosaccharides, like pectin, have a prebiotic effect in the digestive tract of humans and animals against lactic acid bacteria: Lactobacillus sp., Bifidobacterium sp. [139]. On an industrial scale, they are obtained as a by-product from agri-food waste contained in fruit and vegetable peels: orange peel, onion peel, beet pulp, Actinidia alifolia and fresh artichokes [92,140] . Oligosaccharides obtained from pectin may have similar or additional applications, but they are characterized by a higher degree of fermentation compared to pectin (lower viscosity), hence they may have a greater selective effect on the profile of fatty acids produced in the hindgut [141]. POS also has antioxidant and anti-carcinogenic properties against several types of cancer, causing apoptosis of cancer cells, preventing colon ulcers and inflammation [142]. In addition, like other oligosaccharides not digested in the host organism, they stimulate the growth of beneficial bacteria in the colon and, on the principle of competitive exclusion [143]. Therefore limit the number of pathogens in the mucus layer, preventing their nesting on epithelial receptors and thus preventing the occurrence of inflammation of the large intestine [118].
Pectin oligosaccharides are obtained on an industrial scale as a result of: enzymatic hydrolysis of pectin, acid hydrolysis, hydrothermal treatment and physical decomposition. However, purification of raw POS mixtures occurs through membrane filtration [131]. Oligosaccharides obtained from pectin may include products such as: oligogalactouronides (OGalA), galactooligosaccharides (GalOS), arabinooligosaccharides (AraOS), rhamnogalacturonooligosaccharides (RhaGalAOS) and arabiongalactooligosaccharides (AraGalOS), which may be additionally replaced with a methyl or acetyl group or an esterified ferulic acid [130].
Scholz-Ahrens et al. [144] is available raising the issue of the impact of probiotics, prebiotics and synbiotics on calcium absorption, the mineral composition of bones and their structure, we may assume that the use of oligosaccharides has a beneficial effect on increasing the level of calcium in the body. This effect was confirmed by Chawla and Patil [6] in their review article, pointing to numerous studies using inulin, oligofructose and FOS, in which increased absorption of Ca, Mg and Zn from the gastrointestinal tract was found in the participants.
In addition, POS has a positive effect on lipid metabolism in white adipose tissue in studies on mice fed a high-fat diet. Fan et al. [145] in their experiment using POS derived from hawthorn found a reduction in inflammation of adipose tissue when animals were on a high-fat diet. Moreover, POS had a positive effect on the metabolism in adipose tissue by blocking the formation of triglycerides and fatty acids and facilitating fatty acid oxidation and triglyceride catabolism. In turn, Li et al. [146] in their experiment with POS contained in hawthorn fruits found their very strong antibacterial activity in vitro. POS also have antioxidant and anti-carcinogenic properties against several types of cancer, causing apoptosis of cancer cells, preventing ulcers and inflammation in the colon [131].
Oligosaccharides obtained from pectin are also the subject of the research from the point of view of formulas of milk replacers for infants [147,148]. POS extracted from pectin are acidic, while oligosaccharides found in human milk consist of 75-85% of neutral oligosaccharides and 15-25% of them acidic. However, as research by Fanaro et al. [149] use 0.2g/dL acidic oligosaccharides gave similar results to the standard preparation based on milk proteins (in a 60/40 ratio for whey and casein, respectively), and they also found no difference between the total number of bifidobacteria in feces. However, much better results were achieved by combining pectic oligosaccharides in the amount of 0.2 g/dL with the GOS/FOS mixture in the amount of 0.6 g/dL. Newborns showed greater weight gain, although not statistically significant (p>0.05), whereas a significantly higher number of Bifidobacterium was found in their stools.
Additionally, pectin and POS block the formation of advanced glycation end products (AGE's), which is associated with the occurrence of diabetic complications, the aging process and atherosclerosis [150,151]. Additionally, research by Zhu et al. [147] showed that the content of AGE's in milk replacers in infant formulas based on cow's milk designed to meet the nutritional requirements of infants was up to a thousand times higher than in raw human or cow's milk. For these types of infant formulas, the use of POS at 1.5 mg/ml yielded comparable results compared to the same amount of aminoguanidine. These results were significantly higher compared to the participation of these two ingredients in the amount of 1 and 0.5 mg/ml and the blank sample containing no active substances in relation to AGE's .
6. Pectin and POS - Prebiotic Effect on Intestinal Microbiome
6.1. Prebiotics
The definition of prebiotics used today developed over an 8-year period (from 1995 to 2004) and initially described them as: non-enzymatically digestible dietary components that beneficially affect the host by selectively stimulating the growth and activity of one or a specific number of bacterial strains in the colon that improve the health of the host. Subsequently, it was slightly modified and prebiotics were defined as: selectively fermented ingredients that allow specific changes in both the composition and activity of the gastrointestinal microflora that benefit the host organism [152,153]. From this time definition was subject of gradual modifications and her the most current sound left, proposed by Bindels et al. [154] defines prebiotics as: “A nondigestible compound that, through its metabolism by microorganisms in the gut, modulates the composition and/or activity of the gut microbiota, thus conferring a beneficial physiologic effect on the host.”
Pectin and POS directly influence the intestinal microbiome, similarly to other prebiotics. Comparison between effect on microbiota and SCFA production from pectin source and POS obtain from the same source is presented in a Table 3. Four main types of bacteria dominate the human intestinal microbiome are: Firmicutes, Bacteroidetes, Actinobacteria, Proteobacteria. In western populations, the ratio of Bacteroidetes to Firmicutes is 1:1 [155].
Obesity and irritable bowel syndrome are associated with an increase in Firmicutes and a decrease in Bacteroidetes in the gut microbiome [162,166] Wang et al. [167] highlight three important functions in the modulation of the microbiome through the use of pectin and their derivatives: prebiotic effect by selectively increasing Bifidobacterium and Lactobacillus strains in the colonic microbiome, modifying the ratio of the number of Bacteroidetes and Firmicutes , and reducing the potential of Proteobacteria associated with mucosa, especially the enteric pathogens present in this group of microorganisms.
6.2. Pectin and Its Effect on Microbiome
Pectin in the gastrointestinal tract is responsible for limiting the absorption of glucose from the intestinal lumen, causing hypercholesterolemia and prolonging the gastric emptying time [168]. Additionally, oligosaccharides obtained from pectin are a prospective addition to the diet as new generation prebiotics, characterized by high efficiency and relatively low production costs [90]. By many years there was a conviction that pectin does not have a direct antibacterial effect, it is only a source of substrates for bacteria, which degrade it into oligosaccharides, activating the antibacterial properties of these compounds [169]. However, this is not entirely true because the research by Ishisono et al. [85] and Sahasrabudhe [120] showed that pectin also have a direct effect on antibacterial activity within the intestines by blocking Tool-like signalling receptors in Peyer's patches. Hence, the antibacterial effect is visible on two levels.
The diversity of microorganisms in the intestines plays an important role in the development of inflammatory disease processes, similarly, the occurrence of allergies is related to the disruption of the intestinal colonization process by microorganisms or to the limitation of the diversity of the microbiome in this part of the digestive tract [170]. Additionally, Chung et al. [156], comparing the effect of inulin and pectin on the diversity of the microbiome, found a significantly higher Shanon value diversity index in the case of the microbiome whose substrate was pectin (p = 0.001). Inulin increased the replication of the selected type of bacteria, which is a reason to use functional fibre in the form of both types of probiotics.
Pectin, depending on its structure and the number of branched chains, affects different profiles of fatty acids: acetate, propionate and butyrate. Additionally, the high content of dry matter in pectin was positively correlated with the level of propionate produced by microorganisms. Gram-negative Bacteroidetes, which are part of the native flora of the intestinal microbiome , are mainly responsible for the degradation of pectin [171]. Another interesting species of bacteria that participate in the pectin decomposition process is Eubacterium eligens, belonging to Firmicutes. This bacterium has the ability to reduce the occurrence of inflammation within the intestinal mucus and epithelium by stimulating the production of IL-10 in the intestinal epithelial layer, which affects the stabilization of the microbiome in the colon, but also has a positive impact on the health of patients [172]. Pectin contained in beet pulp contain mainly arabino -oligosaccharides with polymer numbers ranging from 2 to 6. Holck et al. [173] indicate that arabino-oligosaccharides with a polymer number of up to 8 influence the selective stimulation of Bifidobacteria in relation to Clostridia, Lactobacilli and Bacteroides in the case of in vitro fermentation. However, a precise determination of the effect of pectin on the microbiome, and in particular on the entire mechanism of maintaining the balance of the microbiome and developing immunity, requires more research, meta-analyses and systematic reviews.
6.3. Pectin Oligosaccharides - Effect on Microbiome
The increase in the microbiome population is associated with the production of SCFAs in the hindgut, and in addition, the presence of dietary fibre not digested by the host affects its blood lipid profile [174]. Additionally, Wilkowska et al. [32] conducted studies on two age groups of healthy volunteers who used the addition of functional fibre to Western diets in the form of POS obtained from apple fibre. Researchers found that as in the case of the microbiome and animal model studies, also in humans POS work more effectively in the case of younger people leading to a more favourable SCFA profile. However, research of Gu et al. [175] emphasize that influence of different types of pectin on the gut microbiome composition and SCFA profiles of elderly is compared to younger adults. This fact is emphasized by Williams et al. [47], who points out that the criterion for selecting dietary fibre in the diet should be not only its solubility, but also the ability to ferment and thus influence the obtained SCFA profile, what consequently affects the maintenance of the homeostasis of the intestinal microbiome and positively interacts with an increase in immunological resistance in humans and animals. In vitro studies Yang et al. [176] show that pectin are fermented in about 60%, β- glucan , resistant starch in about 50%, inulin in 40%, and guar gum and arabinoxylans in 30%. Of course, obtaining dietary fibre in the form of oligosaccharides from these types significantly increases the efficiency of fermentation in the intestines.
POS influences the composition and diversity of intestinal microbiome [177]. The main bacteria responsible for the degradation of this type of oligosaccharides are Bacteroides and Prevotella, the latter type of bacteria showing an affinity for pectin with a higher methoxyl content, and the profile of SCFA resulting from microbiological transformations contains more propionic acid [139]. Pectin seems to be a better substrate for Bacteroides present in the large intestine than inulin, and 70% of bacteria from this genus use HG and galactan for growth, the breakdown of dietary fibre occurs thanks to enzymes such as: lyases, methylesterases and acetylases [178]. Products resulting from the action of these bacteria increase their number and that of Lactobacilli [179]. Although, on the other hand in studies on: rats, mice and piglets, a reduction in the number of Lactobacillus sp. and Bacteroides spp. was observed [142,180]. Some studies also show a positive correlation between Bacteroidetes and Firmicutes ratios, and the amount of SCFAs [181,182] However, Hu et al. [183] indicate a correlation between the number of Bifidobacterium , Bacteroidetes and Lactobacillus and the level of SCFA, total cholesterol and LDL fraction.
In terms of the overall SCFA profile for pectin, many microorganisms are responsible for the production of acetic acid, but especially those of the Bifidobacteria and Lactobacilli genera. Propionic acid is mainly produced by Bacteroidetes and Firmicutes, while butyric acid is synthesized by Eubacterium rectale, Roseburia intestinalis, Faecalibacterium prausnitzii [184,185]. Butyric acid plays a very important role in the colon related to the nutrition of intestinal epithelial cells, as well as in the prevention and treatment of colon diseases [54,186]. The prebiotic potential of POS has been confirmed in experiments in which they selectively increased the number of beneficial bacteria in the human gastrointestinal tract, such as Bifidobacteria or Eubacterium rectale [141,159]. However, the effect of POS and their probiotic potential depends on the type of plants from which they are obtained and the production process. They can be obtained as a result of physical, chemical and enzymatic processes, which affects their degree of polymerization and thus their properties [15]. Therefore, in the case of large-scale use of POS, it is necessary to standardize the obtained product or obtain a uniform product from POS with different atomic weight on the basis of masterbatches. Of course, in the case of prebiotic properties and the impact on the stabilization of gut mucus layer, intestinal epithelium and immunity, branched forms with a significant share of rhamnogalacturonans I and II, due to high solubility in water and easier access of microorganisms to the structure of the POS molecule.
6.4. Antibacterial and Immunomodulatory Effect of POS
The antibacterial effect of POS was first confirmed by Takenaka et al. [187] using pectin hydrolyzate contained in lemons due to the action of galacturonan oligomers. Foti et al. [188] also demonstrated the antibacterial effect and modulatory effect of POS obtained from citrus pectin on the intestinal microbiome. Wang et al. [58] in their studies demonstrated strong antibacterial activity of POS contained in citrus peels and apple pomace against gram-bacteria. In turn, Gullón et al. [118] showed a very high ability of pectin and POS in modulating the microbiome (especially Faecalibacterium prausnitzii bacteria and Roseburia intestinalis ) compared to other FOS or GOS oligosaccharides. Which is especially important in the case of Faecalibacterium prausnitzii , because as one of the main bacteria from the entire group of microorganisms it is responsible for the health of the organism within the biofilm colonic microbiome [129]. The remaining bacteria include: Bacteroides spp., Coprococcus, Dorea, Ruminococcus, Blautia, Oscillospira, Sutterella, Bifidobacterium, Christensenellaceae.
An important element in the stabilization of the microbiome, as well as the acquisition of immunity, is the combination of the effects of POS, FOS and GOS. Hence this type of research has been carried out in infants and the studies have shown that the combination of ScGOS, lc FOS and pectin results in a higher number in their microbiome: Bifidobacterium sp. and Lactobacilli compared to groups of newborns who received only pectin in the formula or the control group [148,149]. What is important in this case is the number of Lactobacilli and Bifidobacteria, the reduced level of which in the digestive tract of children under 5 years of age resulted in a greater risk of allergies [189]. Stabilization of the microbiome by prebiotics (including pectins and POS) also affects the amount and type of metabolites produced by microorganisms, including SCFA: which is a source of energy for colonocytes, regulators of gene expression, cell differentiation and inflammatory agent [183,190]. This allows for the creation of a cross-feeding network, thanks to which, using metagenomics and metabolomics methods combined with machine learning methods (GutCP) or deep learning, it is possible to modulate the composition of the diet, and thus influence changes in the microbiome and control the health status of a given person. people based on methods of inferring and predicting cross-feeding interactions in the human gut microbiome [57,191]. Similar studies on intestinal microflora and its metabolites collected in the form of a global metabolic interaction network were performed by [190]. Like the others, it allows predicting changes in the composition of the intestinal microbiome, and thus the concentrations of metabolites produced by microorganisms, which allows, based on changes in groups of metabolites, to predict changes that may lead to the occurrence of a disease in order to avoid it.
7. Interaction of Pectin and POS with GALT
7.1. Dietary Fibre and Its Effect on GALT
Dietary fibre is broken down mainly in the human large intestine and its decomposition products are obtained there [192]. However, in the small intestine, it has the ability to directly interact with the cells of the immune barrier in the small intestine, due to the interaction between intestinal epithelial cells and immune cells (Figure 4).
The consequence of these interactions is the strengthening of the mucous layer, the barrier function of epithelial cells and the modulation of the intestinal immune response [193,194]. The direct interaction between dietary fibre and the intestinal immune system may improve health and reduce the occurrence of diseases [195].
Dietary fibre is correlated with reducing the occurrence of diseases, immune disorders and mortality, a special role in this case is played by maintaining the immune barrier of the gastrointestinal tract by stabilizing the microbiome and their metabolites, especially in case use inulin in relation with type 2 inflammation [196]. Additionally, research indicates the immuno-modulatory function of short-chain galactooligosaccharides (scGOS) and long-chain fructooligosaccharides (lcFOS) also supporting the function of human milk oligosaccharides (HMOS) [197]. Additionally, Bernard et al. [198] when used in the diet of mice (pAOS) found stimulation of immune polymerization of Th1 lymphocytes as a result of increasing the release of interferon γ, upregulating the expression of the t-bet gene, decreasing the secretion of interleukin 4 and downregulating the expression of the GATA3 gene.
7.2. Pectin Effect on GALT
Pectin have a large impact on the health of the body by modulating the intestinal immune barrier [199]. The mucosal layer is a protective barrier separating the intestinal epithelium from the microbiome and digestive content [200]. Metabolic products synthetized by microorganisms change the composition and properties of the mucous layer [201]. In the colon mucus is divided on two layers: inner free from bacteria and outer composed from mucus, gut bacteria and dietary material [202].
Citrus pectin exhibit immunomodulatory properties in two ways: neutral RG-1 or RG-2 side chains inhibit the activation of lipopolysaccharide (LPS) induced by pattern recognition receptors tool like receptors 4 (TLR-4), while homogalactouronate (HG) has an immunostimulatory effect through the TLR-4 pathway and the MyD88 protein [183]. Additionally, modified citrus pectin has strong anticancer properties [203].
The immunomodulatory effect of pectin depends mainly on the content of galacturonic acid and methoxyl residues, the percentage of which is most abundant in the parts of pectin composed of RG-I and RG-II that are intact during the POS extraction process [110]. The effect of increasing the thickness of the mucous layer is visible after the use of pectin in the diet by stimulating mucin secretion in the small intestine [204]. Regulation mucus secretion and homeostasis occurs with the participation of Forkhead box protein O1 (Foxo1) [205]. These compounds are produced in goblet cells in response to microorganisms in the intestinal lumen and dietary ingredients as a result of activation of pattern recognition receptors (PRRs), simulation of SCFA production and as a result of cytokine secretion from immune cells [206].
7.3. POS Effect on GALT
Oligosaccharides FOS, POS and GOS are broken down by the intestinal microbiome into SCFAs, the concentration of which affects the functions of leukocytes and the differentiation of T lymphocytes and their functions, allowing SCFAs to be classified as postbiotics [139,207]. POS, like other oligosaccharides, is responsible for reducing the response dependent on Th2 lymphocytes in the course of allergic diseases, limiting the excessive response of the immune system to allergens [208]. The use of POS also affects the regulation of Treg /TH17 levels [167]. Additionally, the results of studies conducted in children and infants indicate a reduction in the frequency of viral and bacterial infections requiring antibiotic treatment [203]. Therefore, in recent years, the administration of prebiotics in the form of oligosaccharides has become a new strategy for preventing acute or chronic infectious diseases , and thus reducing the phenomenon of antibiotic resistance in bacteria. However, Montilla et al. [209] point out that when using pectin and POS, the level of pro-inflammatory biomarkers is reduced and the body's immunity increases. Xie et al [210] noted lack differences in thickness of mucus layer when inulin and POS was used. The mucous layer also contains immunoglobulin A, which counteracts the increase in the number of pathogenic microorganisms, important role in this process plays eosinophils [211].
Oligosaccharides plats also important role in decrease of allergic reaction of organism. Jeurink et al. [212] described the impact of the use of oligosaccharides on immune processes and mechanisms responsible for shaping the body's resistance in connection with exposure to allergens. Cohort studies indicate that one of the factors contributing to the occurrence of food allergies is intestinal dysbiosis [213,214].
Fecalibacterium prausnitzii and Akkermansia muciniphilla play an important role in maintaining the balance of the colon's microbiome and thus improving its health, belong to bacteria that degrade mucin and pectic oligosaccharides, which constitute substrates for the production of butyric acid, which influences the nutrition of intestinal epithelial cells [195]. The first bacterial species reduces inflammation of the epithelium, while the second one strengthens the barrier function of the mucus Wang et al. [132] in their studies confirm the beneficial effect of POS obtained as a result of enzymatic degradation of pectin in restoring the balance of the microbiome and immunological homeostasis in the intestines, both to prevent uncontrolled changes, as well as after ulcerative colitis. Additionally, POS causes an increase in the number of beneficial microorganisms: Bifidobacterium , Lactobacillus and Eubacterium, which ferment carbohydrates without producing toxins, increasing immunity and acting on the principle of competitive exclusion in relation to pathogens. In the study by Guevara-Arauza et al. [215] found an increase in Lactobacilli in response to mucilage oligosachides from Opuntia ficus-indica by 23.8% and Bifidobacteria by 25.0% when POS derived from this plant was used
Pectin and POS have the ability to block the adhesion of pathogens to intestinal epithelial cells [216]. POS reduces the adhesion of pathogenic bacteria: E. coli, Listeria monocytogenes, Salmonella typhimurium, Campylobacter jejuni. However, pectin with lower molecular weight is mainly responsible for this type of effect Moreover, pectin and mainly pectin oligosaccharides enhance the action of commensal and probiotic Lactobacilli strains to the intestinal epithelium, but the final effect in these authors' studies depended on the species and conditions of their performance, which were most often in vitro studies [50]. The mucus layer of the intestinal epithelium is favourable medium for symbiotic bacteria, which promotes their multiplication in this place and also facilitates adhesion to the receptor of the intestinal epithelium, Ouwerkerk et al. [217] defined interaction between those three elements as a glycobiome.
8. Conclusions
The change in eating habits, especially in urbanized societies in the last decade, requires paying more attention to supplying the organism with dietary fibre present in the diet. Insoluble dietary fibre constitute ballast and dietary diluent in both humans and animals GIT, it is also object of partially fermentation. Soluble fibre is divided into fermentable as a source of SCFA) and viscous (limiting the absorption of nutrients). Pectin as an element of SDF contained in agri-food products is an important source of pectin in food industry and pectic oligosaccharides, varied in terms of branching of the side chains of pectin molecules, which affects their properties. Pectin-oligosaccharides have a prebiotic effect in the gastrointestinal tract mainly promoting Lactobacillus sp., Bifidobacterium sp., Bacteroidetes, Firmicutes and Prevotella. Adhesion of pathogens is blocking by: indirect effect as a component, in regeneration of mucus layer in GIT as a barrier for pathogenic bacteria, and synthesis of SCFA by bacteria present in intestines and also in mucus layer. Direct effect on antibacterial activity within the intestines by blocking Tool-like signaling receptors in Peyer's patches. Additionally in organism POS have a positive effect on lipid metabolism (supporting their catabolism), have antioxidant properties by blocking advanced glycolysis products and anti-cancer properties by activating the apoptosis mechanism of cancer cells. Immunomodulation effect is induced by HG, RG-1 and RG-2 mainly by SCFA production from oligomers and stabilization of mucus layer in guts. This type of oligosaccharides is new aspect in science, that is why more research is necessary to find an easier way to obtain POS and to precisely determine their mechanism of action in humans and animals, as well as in synergy with other types of oligosaccharides.
Author Contributions
Conceptualization, T.H, P.W.; software, T.H, A.Z; investigation, all authors; resources all authors; data curation, T.H.; writing—original draft preparation, T.H., P.W.; writing—review and editing, A.S., K.L-S.; visualization, T.H., P.W.; supervision, A.R., A.Z., A.S., K.L-S.; funding acquisition, T.H. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are available from the authors.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- He, Y.; Wang, B.; Wen, L.; Wang, F.; Yu, H.; Chen, D.; Su, X.; Zhang, C. Effects of dietary fiber on human health. Food Sci. Hum. Wellness. 2022, 11, 1–10. [Google Scholar] [CrossRef]
- Maina, N.H.; Rieder, A.; de Bondt, Y.D.; Mäkelä-Salmi, N.; Sahlstrøm, S.; Mattila, O.; Lamothe, L.M.; Nyström, L.; Courtin, C.M.; Katina, K.; Poutanen, K. Process-induced changes in the quality and characteristics of grain dietary fiber. Foods 2021, 10, 2566. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Folkerts, J.; Folkerts, G.; Maurer, M.; Braber, S. Microbiota-dependent and -independent effects of dietary fibre on human health. Br. J. Pharmacol. 2020, 177, 1363–1381. [Google Scholar] [CrossRef] [PubMed]
- Pathania, S.; Kaur, N. Utilization of fruits and vegetable by-products for isolation of dietary fibres and its potential application as a functional ingredients. Bioact. Carbohydr. Diet. Fibre 2022, 27, 100295. [Google Scholar] [CrossRef]
- Buttriss, J.L.; Stokes, C.S. Dietary fibre health: An overwiew. Nutr. Bull. 2008, 33, 186–200. [Google Scholar] [CrossRef]
- Chawla, R.; Patil, G.R. Soluble dietary fiber. Compr. Rev. Food Sci. Food Saf. 2010, 9, 178–196. [Google Scholar] [CrossRef]
- Widaningrum; Flanagan, B.M.; Williams, B.A.; Sonni, F.; Mikkelsen D.; Gidley, M.J. Fruit and vegetable insoluble dietary fibre in vitro fermentation characteristics depend on cell wall type. Bioact. Carbohydr. Diet. Fibre 2020, 23, 100223. [Google Scholar] [CrossRef]
- Liao, A.-M.; Zhang, J.; Yang, Z.L.; Huang, J.-H.; Pan, L.; Hou, Y.C.; Li, X.-X.; Zhao, P.H.; Dong, Y.Q.; Hu, Z. Y.; Hui, M. Structural, physiochemical and functional properties of wheat bran insoluble dietary fiber modified with probiotic fermentation. Front. Nutr. 2022, 9, 803440. [Google Scholar] [CrossRef]
- Ge, Q.; Li, H.-G.; Zheng, Z.-Y.; Yang, K.; Li, P.; Xiao, Z.-G.; Xiao, G.-M.; Mao, J.W. In vitro fecal fermentation characteristics of bamboo insoluble dietary fiber and its impacts on human gut microbiota. Food Res. Int. 2022, 156, 111173. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Bai, Y.; Zhang, G.; Liu, L.; Lai, L. Relationship between dietary fiber fermentation and volatile fatty acids concentration in growing pigs. Animals 2020, 10, 263. [Google Scholar] [CrossRef]
- Slavin, J. Fiber and prebiotics: Mechanism and health benefits. Nutrients, 2013, 5, 1417–1435. [Google Scholar] [CrossRef]
- Lattimer, J.M.; Haub, M.D. Effects of dietary fiber and its components on metabolic health. Nutrients, 2010, 2, 1266–1289. [Google Scholar] [CrossRef]
- Dalzenne, N.M. Oligosaccharides: State of the art. Proc. Nutr. Soc. 2003, 62, 177–182. [Google Scholar] [CrossRef]
- Mudgil, D.; Barak, S. Composition properties and health benefits of indigestible carbohydrate polymers as dietary fiber: A review. Int. Biol. Macromol. 2013, 61, 1–6. [Google Scholar] [CrossRef]
- Combo, A.M.M.; Aguedo, M.; Quiévy, N.; Danthine, S.; Goffin, D.; Jacquet, N.; Blecker, C.; Devaux, J.; Paquot, M. Characterization of sugar beet pectic-derived oligosaccharides obtained by enzymatic hydrolysis. Int. J. Biol. Macromol. 2013, 52, 148–156. [Google Scholar] [CrossRef] [PubMed]
- La Torre, D.; Verbeke, K.; Dalile, B. Dietary fibre and gut-brain axis: Microbiota-dependent and independent mechanism of action. Gut Microbiome 2021, 2, 1–26. [Google Scholar] [CrossRef]
- Raninen, K.; Lappi, J.; Mykkänen, H.; Poutanen, K. Dietary fiber type reflects physiological functionality: Comparison of grain fiber, inulin, and polydextrose. Nutr. Rev. 2011, 69, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Yao, H.; Flanagan, B.M.; Williams, B.A.; Mikkelsen, D.; Gidley, M.J. Particle size of dietary fibre has diverse effects on in vitro gut fermentation rate and end-products depending on food source. Food Hydrocoll. 2023, 134, 108096. [Google Scholar] [CrossRef]
- McRorie Jr., J. W.; McKeown, N.M. Understanding the physics of functional fibers in the gastrointestinal tract: An evidence-based approach to resolving enduring misconceptions about insoluble and soluble fiber. J. Acad. Nutr. Diet. 2017, 117, 251–264. [Google Scholar] [CrossRef]
- Gidley, M.J.; Yakubov, G.E. Functional categorization of dietary fibre in foods: Beyond ‘soluble’ vs ‘insoluble’. Trends Sci. Food Technol. 2019, 86, 563–568. [Google Scholar] [CrossRef]
- Bach Knudsen, K.E. The nutritional significance of “dietary fibre” analysis. Anim. Feed Sci. Tech. 2001, 90, 3–20. [Google Scholar] [CrossRef]
- Singh, R.P.; Tingirikari, J.M.R. Agro waste derived pectin poly and oligosaccharides: Synthesis and functional characterization. Biocatal. Agric. Biotechnol. 2021, 31, 101910. [Google Scholar] [CrossRef]
- Pop, C.; Suharoschi, R.; Pop, O.L. Dietary fiber and prebiotic compounds in fruits and vegetables for waste. Sustainability, 2021, 13, 7219. [Google Scholar] [CrossRef]
- Saber, W.I.A.; El-Naggar, N.E. Optimization of fermentation conditions for the biosynthesis of inulinase by the new source; Aspergillus tamari and hydrolysis of some inulin containing agro-wastes. Biotechnology 2009, 8, 425–433. [Google Scholar] [CrossRef]
- Sabater, C.; Calvette-Torre, I.; Villamiel, M.; Javier Moreno, F.; Margolles, A.; Ruiz, L. Vegetable waste and by-products to feed a healthy gut microbiota: Current evidence, machine learning and computational tools to design novel microbiome-targeted foods. Trends Food Sci. Technol. 2021, 118 (Part A), 399–417. [Google Scholar] [CrossRef]
- Elshahed, M.S.; Miron, A.; Aprotosoaie, A.C.; Farag, M.A. Pectin in diet: Interactions with the human microbiome, role in gut homeostasis, and nutrient-drug interactions. Carbohydr. Polym. 2021, 255, 117388. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Liao, C.; Wu,L. ; Tang, J.; Chen, J.; Lei, C.; Zheng, L.; Zhang, C.; Lin, Y.-Y.; Xavier, J.; Dai, L. Ecological dynamics of the gut microbiome in response to dietary fiber. ISME J. 2022, 16, 2040–2055. [Google Scholar] [CrossRef] [PubMed]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Goptenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A.; Biddinger, S.B.; Dutton, R.J.; Turnbaugh, P.J. Diet rapidly and reproducibly alters the human gut microbiome. Nature 2014, 505, 559–563. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.; Feng, N.; Li, Q.; Wang, H.; Su, Y.; Zhu, W. Short-term supplementation of pectin alters substrate dynamics and modulates microbial carbohydrate metabolism in the gut of a pig model. J. Agric. Food Chem. 2023, 71, 10470–10482. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Peng, C.; Mao, A.; Zhong, M.; Hu, Z. An overview of microbial enzymatic approaches for pectin degradation. Int. J. Biol. Macromol. 2024, 254, 127804. [Google Scholar] [CrossRef] [PubMed]
- Lordan, C.; Thapa, D.; Ross, R.P.; Cotter, P.D. Potential for enriching next generation health-promoting gut bacteria through prebiotics and other dietary components. Gut Microbes 2020, 11, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Wilkowska, A.; Nowak, A.; Antczak-Chrobot, A.; Motyl, I.; Czyżowska, A.; Paliwoda, A. Structurally different pectic oligosaccharides produced from apple pomace and their biological activity in vitro. Foods 2019, 8, 365. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.-C.; Chiang, H.-H.; Liu, C.-Y.; Li, Y.-J.; Lu, C.-L.; Lee, Y.-P.; Huang, C.-J.; Lai, C.-L. Intestinal mucosal barrier improvement with prebiotics: Histological evaluation of longish glucomannan hydrolysates-induced innate T-lymphocyte activities in mice. Nutrients 2022, 14, 2220. [Google Scholar] [CrossRef] [PubMed]
- Mao, X.; Ding, X.; Zeng, Q.; Bai, S.; Zhang, K.; Chen, D.; Yu, B.; He, J.; Yu, J.; Luo, J.; Luo, Y.; Wang, J. The effect of dietary pectic oligosaccharide supplementation on intestinal health of broiler breeders with different egg-laying rates. Poultry Science 2021, 100, 100938. [Google Scholar] [CrossRef]
- Chudzik, A.; Orzyłowska, A.; Rola, R.; Stanisz, G.J. Probiotics, prebiotics and postbiotics on mitigation of depression symptoms: Modulation of the brain-gut-microbiome axis. Biomolecules 2021, 11, 1000. [Google Scholar] [CrossRef]
- Chen, H.; Hu, H.; Chen, D.; Tang, J.; Yu, J.; Luo, J.; He, J.; Luo, Y.; Yu, J.; Mao, X. Dietary pectic oligosaccharide administration improves growth performance and immunity in weaned pigs infected by rotavirus. J. Agric. Food. Chem. 2017, 65, 65,2923–2929. [Google Scholar] [CrossRef]
- Barber, T.M.; Kabisch, S.; Pfeiffer, A.F.H.; Weickert, M.O. The health benefits of dietary fibre. Nutrients 2020, 12, 3209. [Google Scholar] [CrossRef]
- O’Keffe, S.J. The association between dietary fibre deficiency and high-income lifestyle- associated diseases: Burkitt’s hypothesis revisited. Lancet Gastroenterol. Hepatol. 2019, 4, 984–996. [Google Scholar] [CrossRef] [PubMed]
- Bach Knudsen, K.E.; Nørskov, N.P.; Bolvig, A.K.; Hedemann, M.S.; Lærke, H.N. Dietary fibers and associated phytochemicals in cereals. Mol. Nutr. Food Res. 2017, 61, 1600518. [Google Scholar] [CrossRef] [PubMed]
- Holscher, H.D. Dietary fiber and prebiotics and the gastrointestinal microbiota. Gut Microbes 2017, 8, 172–184. [Google Scholar] [CrossRef] [PubMed]
- Prado, T.R.; Mazzonetto, A.C.; Botelho, A.M.; Fiates, G.M.R. Home availability of ultraprocessed foods in families who prepare meals at home. Rev. Nutr. 2022, 35, e210249. [Google Scholar] [CrossRef]
- Farmer, N.; Lee, L.J.; Powell-Wiley, T.M.; Wallen, G.R. Cooking frequency and perception of diet among US adults are associated with US healthy and healthy Mediterranean-style dietary related classes: A latent class profile analysis. Nutrients 2020, 12, 3268. [Google Scholar] [CrossRef]
- Kopp, W. How western diet and lifestyle drive the pandemic of obesity and civilization diseases. Diabetes Metab. Syndr. Obes. 2019, 12, 2221–2236. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.D.; Chen, J.; Hoffman, C.; Bittinger, K.; Chen, Y.-Y.; Keilbaugh, S.A.; Bewtra, M.; Knights, D.; Walters, W.A.; Knight, R.; Sinha, R.; Gilroy, E.; Gupta, K.; Baldassano, R.; Nessel, L.; Li, H.; Bushman, F.D.; Lewis, J.D. Linking long-term dietary patterns with gut microbial enterotypes. Science, 2011, 334, 105–108. [Google Scholar] [CrossRef] [PubMed]
- Makki, K.; Deehan, E.C.; Walter, J.; Bäckhed, F. The impact of dietary fiber on gut microbiota in host health and disease. Cell Host Microbe 2018, 23, 705–715. [Google Scholar] [CrossRef]
- Gill, S.K.; Rossi, M.; Bajka, B.; Whelan, K. Dietary fibre in gastrointestinal health and disease. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 101–116. [Google Scholar] [CrossRef] [PubMed]
- Williams, B.A.; Mikkelsen, D.; Flangan, B.M.; Gidley, M.J. “Dietary fibre”: Moving beyond the “soluble/insoluble” classification for monogastric nutrition, with an emphasis on humans and pigs. J. Anim. Sci. Biotechnol. 2019, 10, 45. [Google Scholar] [CrossRef] [PubMed]
- Tungland, B.C.; Meyer, D. Nondigestible oligo- and polysaccharides (dietary fiber): Their physiology and role in human health and food. Comp. Rev. Food Sci. Food Saf., 2002, 3, 90–109. [Google Scholar] [CrossRef]
- Deehan, E.C.; Walter, J. The fiber gap and the disappearing gut microbiome: Implications for human nutrition. Trends Endocrinol. Metab. 2016, 27, P239–P242. [Google Scholar] [CrossRef]
- Beukema, M.; Faas, M.M.; de Vos, P. The effects of different dietary fiber pectin structures on the gastrointestinal immune barrier: Impact via gut microbiota and direct effects on immune cells. Exp. Mol. Med. 2020, 52, 1364–1376. [Google Scholar] [CrossRef]
- Anderson, J.W.; Baird, P.; Davis Jr., R. H.; Ferreri, S,; Knudtson, M.; Koraym, A.; Waters, V.; Williams, C.L. Health benefits of dietary fiber. Nutr. Rev. 2009, 67, 188–205. [Google Scholar] [CrossRef] [PubMed]
- Ötles, S.; Ozgoz, S. Health effects of dietary fiber. Acta Sci. Pol. Technol. Aliment. 2014, 13, 191–202. [Google Scholar] [CrossRef] [PubMed]
- Thompson, S.V.; Bailey, M.A.; Taylor, A.M.; Kaczmarek, J.L.; Mysonhimer, A.R.; Edwards, C.G.; Reeser, G.E.; Burd, N.A.; Khan, N.A.; Holscher, H.D. Avocado consumption alters gastrointestinal bacteria abundance and microbial metabolic concentrations among adults with overweight or obesity: A randomized controlled trial. J. Nutr. 2021, 151, 753–762. [Google Scholar] [CrossRef] [PubMed]
- Bach Knudsen, K.E.; Lærke, H.N.; Hedemann, M.S.; Nielsen, T.S.; Ingerslev, A.K.; Gundelund Nielsen, D.S.; Theil, P.K.; Purup, S.; Hald, S.; Schioldan, A.G.; Marco, M.L.; Gregersen, S.; Hermasen, K. Impact of diet-modulated butyrate production on intestinal barrier function and inflammation. Nutrients, 2018, 10, 1499. [Google Scholar] [CrossRef]
- Wang, G.; Zhong, D.; Lin, H.; Yang, T.; Liang, Q.; Wang, J.; Zhang, R.; Zhang, Y. Water soluble dietary fibre from walnut meal as a prebiotic in preventing metabolic syndrome. J. Funct. Foods, 2021, 78, 104358. [Google Scholar] [CrossRef]
- Holscher, H.D.; Gutterman, H.M.; Swanson, K.S.; An, R.; Matthan, N.R.; Lichtenstein, A.H.; Novotny, J.A.; Baer, D.J. Walnut consumption alters the gastrointestinal microbiota, microbially derived secondary bile acids, and health markers in healthy adults: A randomized controlled trial. J. Nutr. 2018, 148, 861–867. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Holscher, H.D.; Maslov, S.; Hu, F.B.; Weiss, S.T.; Lin, Y.-Y. Predicting metabolic response to dietary intervention using deep learning. bioRxiv 2023. [Google Scholar] [CrossRef]
- Wang, H.; He, C.; Lin, Y.; Zhao, H.; Long, L.; Gai, X.; Zhao, H. Soluble dietary fiber protects intestinal mucosa barrier by improving intestinal flora in a murine model of sepsis. Biomed. Pharmacother. 2020, 129, 110343. [Google Scholar] [CrossRef] [PubMed]
- Nyström, E.E.L.; Martinez-Abad, B.; Arike, L.; Birchenough, G.M.H.; Nonnecke, E.B.; Castillo, P.A.; Svensson, F.; Bevins, C.L.; Hansson, G.C.; Johansson, M.E.V. An intercrypt subpopulation of goblet cells is essential for colonic mucus barrier function. Science 2021, 372, eaabb1590. [Google Scholar] [CrossRef] [PubMed]
- Sauvaitre, T. Etienne-Mesmin, L.; Sivignon, A.; Mosoni, P.; Courtin, C.M.; van de Wiele, T.; Blanquet-Diot, S. Tripartite relationship between gut microbiota, intestinal mucus and dietary fibers: Towards preventive strategies against enteric infections. FEMS Microbiol. Rev. 2020, 45, 1–36. [Google Scholar] [CrossRef]
- Fernández-Tomé, S.; Ortega Moreno, L.; Chaparro, M.; Gisbert, J.P. Gut microbiota and dietary factors as modulators of the mucus layer in inflammatory bowel disease. Int. J. Mol. Sci. 2021, 22, 10224. [Google Scholar] [CrossRef]
- Maguire, M.; Maguire, G. Gut dysbiosis, leaky gut, and intestinal epithelial proliferation in neurological disorders: Towards the development of a new therapeutic using amino acids, prebiotics, probiotics, and postbiotics. Rev. Neurosci., 2019, 30, 179–201. [Google Scholar] [CrossRef] [PubMed]
- Quigley, E.M.M. Microbiota-brain-gut ais and neurodegenerative diseases. Curr. Neurol. Neurosci. Rep. 2017, 17, 94. [Google Scholar] [CrossRef]
- Dhingra, D.; Michael, M.; Rajput, H.; Patil, R.T. Dietary fibre in foods: A review. J. Food Sci. Technol. 2012, 49, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.T.; Bedford, M.R.; Wu, S.-B.; Morgan, N.K. Soluble non-starch polysaccharide modulates broiler gastrointestinal tract environment. Poultry Science 2021, 100, 101183. [Google Scholar] [CrossRef]
- Cameron-Smith, D.; Collier, G.R.; O’Dea, K. Effect of soluble dietary fibre on the viscosity of gastrointestinal contents and the acute glycaemic response in the rat. Br. J. Nutr. 1994, 71, 563–571. [Google Scholar] [CrossRef] [PubMed]
- Tejeda, O.J.; Kim, W.K. Role of dietary fiber in poultry nutrition. Animals 2021, 11, 461. [Google Scholar] [CrossRef] [PubMed]
- Ye, S.; Shah, B.R.; Li, J.; Liang, H.; Zhan, F.; Geng, F.; Li, B. A critical review on interplay between dietary fibres and gut microbiota. Trends Food Sci. Technol. 2022, 124, 237–249. [Google Scholar] [CrossRef]
- Baker, R.A. Reassessment of some fruit and vegetable pectin levels. Food Sci., 1997, 62, 225–229. [Google Scholar] [CrossRef]
- Khanum, F.; Sidallinga Swany, M.; Sudarshana Krishna, K.R.; Santhanam, K.; Viswanathan, K.R. Dietary fiber content at commonly fresh and cooked vegetables consumed in India. Plants Foods Hum. Nutr. 2000, 55, 207–218. [Google Scholar] [CrossRef]
- Schakel, S.F.; Dennis, B.H.; Wold, A.C.; Conway, R.; Zhao, L.; Okuda, N.; Okayama, A.; Moag-Stahlberg, A.; Robertson, C.; van Heel, N.; Buzzard, I.M.; Stamler, J. Enhancing data on nutrient composition of food eaten by participants in the INTERMAP study in China, Japan, the United Kingdom, and the United States. J. Food Compos. Anal. 2003, 16, 395–408. [Google Scholar] [CrossRef]
- Bailoni, L.; Schiavon, S.; Pagnin, G.; Tagliapietra, F.; Bousembiante, M. Quanti-qualitative evaluation of pectins in the dietary fibre of 24 foods. Ital. J. Anim. Sci. 2005, 4, 49–58. [Google Scholar] [CrossRef]
- Choi, H.; Sung, J.Y.; Kim, B.G. Neutral detergent fiber rather than other dietary fiber types as an independent variable increases the accuracy of prediction equation for digestible energy in feeds for growing pigs. Asian-Australas. J. Anim. Sci. 2020, 33, 615–622. [Google Scholar] [CrossRef] [PubMed]
- Bellini, M.; Tonarelli, S.; Barracca, F.; Rettura, F.; Pancetti, A.; Cecarelli, L.; Ricchiuti, A.; Costa, F. , de Bortoli N., Marchi S., Rossi A. Chronic constipation: Is a nutritional approach reasonable. Nutrients, 2021, 13, 3386. [Google Scholar] [CrossRef] [PubMed]
- McRae, M.P. Effectiveness of fiber supplementation for constipation, weight loss and supporting gastrointestinal function: A narrative review of meta-analyses. J. Chiropr. Med. 2020, 19, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Prasadi, V.P.; Joye, I.J. Dietary fibre from whole grains and their benefits on metabolic health. Nutrients, 2020, 12, 3045. [Google Scholar] [CrossRef]
- Chutkan, R.; Fahey, G.; Wright, W.L.; McRorie, J. Viscous versus nonviscous soluble fiber supplements: Mechanism and evidence for fiber – specific health benefits. J. Am. Assoc. Nurse Pract. 2012, 24, 476–487. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Dietetic Products Nutrition, and Allergies (NDA). Scientific opinion on dietary reference values for carbohydrates and dietary fibre . 2010, 8, 1462.
- Cione, E.; Fazio, A.; Curcio, R.; Tucci, P.; Lauria, G.; Capello, A.R.; Dolce, V. Resistant starches and non-communicable diseases: A focus on Mediterranean diet. Foods 2021, 10, 2062. [Google Scholar] [CrossRef]
- Chen, M.; Guo, L.; Nsor-Atindana, J.; Goff, H.D.; Zhang, W.; Zhong, F. The effect of viscous soluble dietary fiber on nutrient digestion and metabolic responses II: In vivo digestion process. Food Hydrocoll. 2020, 107, 105908. [Google Scholar] [CrossRef]
- Larsen, N.; de Souza, C.B.; Krych, L.; Barbosa Cahú, T.; Wiese, M.; Kot, W.; Meyer Hansen, K.; Blennow, A.; Venema, K.; Jespersen, L. Potential of pectins to beneficially modulate the gut microbiota depends on their structural properties. Front. Microbiol. 2019, 10, 223. [Google Scholar] [CrossRef]
- Desai, M.S.; Seekatz, A.M.; Koropatkin, N.M.; Kamada, N.; Hickey, C.A.; Wolter, A.; Pudlo, N.A.; Kitamoto, S.; Terrapon, N.; Muller, A.; Young, V.B.; Henrissat, B.; Wilmes, P.; Stappenbeck, T.S.; Núñez, G.; Martens, E.C. A dietary fiber-deprived gut microbiota degrades the colonic mucus barrier and enhances pathogen susceptibility. Cell, 2016, 167, 1339–1353. [Google Scholar] [CrossRef] [PubMed]
- Ashaolu, T.J. ; Immune boosting functional foods and their mechanism: A critical evaluation of probiotics. Biomed. Pharmacother. 2020, 130, 110625. [Google Scholar] [CrossRef] [PubMed]
- Monteagudo-Mera, A.; Rastall, R.A.; Gibson, G.R.; Charalampopoulos, D.; Chatzifragkou, A. Adhesion mechanism mediated by probiotics and prebiotics and their impact on human health. Appl. Microbiol. Biotechnol. 2019, 103, 6463–6472. [Google Scholar] [CrossRef] [PubMed]
- Ishisono, K.; Mano, T.; Yabe, T.; Kitaguchi, K. Dietary fiber pectin ameliorates experimental colitis in a neutral sugar side chain-dependent manner. Front. Immunol. 2019, 10, 2979. [Google Scholar] [CrossRef]
- Kimura, I.; Inoue, D.; Hirano, K.; Tsujimoto, G. The SCFA receptor GPR43 and energy metabolism. Front. Endocrinol. 2014, 5, 85. [Google Scholar] [CrossRef] [PubMed]
- Lo Conte, M.; Cosovich, I.; Ferrarense, R.; Nobili, A.; Palmieri, V.; Massimino, L.; Lamparelli, L.A.; Liang, W.; Riba, M.; Devecchi, E.; Bolla, A.M.; Pedone; E. ; Scavini, M.; Bosi, E.; Fasano, A.; Ungaro, F.; Diana, J.; Mancini, N.; Falcone, M. Alternations of the intestinal mucus layer correlate with dysbiosis and immune dysregulation in human Type 1 Diabetes. eBioMedicine 2023, 91, 104567. [Google Scholar] [CrossRef] [PubMed]
- Murga-Garrido, S.M.; Hong, Q.; Cross, T.-W. L.; Hutchinson, E.R.; Han, J.; Thomas, S.P.; Vivas, E.I.; Denu, J.; Geschin, D.G.; Tang, Z.-Z.; Rey, F.E. Gut microbiota variation modulates the effects of dietary fiber on host metabolism. Microbiome 2021, 9, 117. [Google Scholar] [CrossRef] [PubMed]
- Wróblewska, P.; Hikawczuk, T.; Wiliczkiewicz, A. Dried sugar beet pulp as a source of soluble dietary fibre in equine nutrition: A review. Anim. Nutr. Feed Technol. 2021, 21, 405–420. [Google Scholar] [CrossRef]
- Long, J.; Li, X.; Xue, L.; Xie, Z.; Jiao, A.; Bai, Y.; Zhou, X.; Chen, L.; Qiu, C.; Xu, X.; Jin, Z. Continuous hydrolysis of mango peel pectin for the production of antibacterial pectic oligosaccharides in packed-bed reactor using immobilized polygalactouronase. Food Biosci. 2022, 50, 102117. [Google Scholar] [CrossRef]
- Vaz Rezende, E.S.V.; Lima, G.C.; Naves, M.M.V. Dietary fibers as beneficial microbiota modulators: A proposed classification by prebiotic categories. Nutrition, 2021, 89, 111217. [Google Scholar] [CrossRef] [PubMed]
- Cano, M.E.; García-Martin, A.; Comendador Morales, P.; Wojtusik, M.; Santos, V.E.; Kovensky, J.; Ladero, M. Production of oligosaccharides from agrofood wastes. Fermentation 2020, 6, 31. [Google Scholar] [CrossRef]
- de Moura, F.A.; Macagnan, F.T.; de Oliveira Petkowicz, C.L. da Silva, L.P. Partially hydrolyzed pectin extracted from passion fruit peel: Molar mass and physicochemical properties. Bioact. Carbohydr. Diet. Fibre 2020, 21, 100206. [Google Scholar] [CrossRef]
- Zhu, R.; Wang, C.; Zhang, L.; Wang, Y.; Chen, G.; Fan, J.; Jia, Y.; Yan, F.; Ning, C. Pectin oligosaccharides from fruit of Actinidia arguta: Structure-activity relationship of prebiotic and antiglycation potentials. Carbohydr. Polym., 2019, 217, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Mandalari, G.; Bennett, R.N.; Kirby, A.R.; Lo Curto, R.B.; Bisignano, G.; Waldron, K.W.; Faulds, G.B. Enzymatic hydrolysis of flavonoids and pectic oligosaccharides from bergamot (Citrus bergamia Risso) peel. J. Agric. Food Chem. 2006, 54, 8307–8313. [Google Scholar] [CrossRef] [PubMed]
- Hutnan, M.; Drtil, M.; Marfkova, L. Anaerobic biodegradation of sugar beet pulp. Biodegradation 2000, 11, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Tariq, A.; Sahar, A.; Usman, M.; Sameen, A.; Azhar, M.; Tahir, R.; Younas, R.; Khan, M.I. Extraction of dietary fiber and polyphenols from mango peel and its therapeutic potential to improve gut health. Food. Biosci. 2023, 53, 102669. [Google Scholar] [CrossRef]
- Gawkowska, D.; Cybulska, J.; Zdunek, A. Structure-related gelling of pectins and linking with other natural components: A review. Polymers, 2018, 10, 762. [Google Scholar] [CrossRef] [PubMed]
- Usmani, Z.; Sharma, M.; Diwan, D.; Tripathi, M.; Whale, E.; Jayakody, L.N.; Moreau, B.; Thankur, V.K.; Tuohy, M.; Gupta, V.K. Valorization of sugar beet pulp to value-added products: A review. Bioresour. Technol. 2022, 346, 126580. [Google Scholar] [CrossRef]
- Wongkaew, M.; Tangjaidee, P.; Leksawasdi, N.; Jantansakulwong, K.; Rachtanapun, P.; Seesuriyachan, P.; Phimolsiripol, Y.; Chaiyaso, T.; Ruksiriwanich, W.; Jantrawut, P.; Sommano, S.R. Mango pectic oligosaccharides: A novel prebiotic for functional food. Front. Nutr. 2022, 9, 798543. [Google Scholar] [CrossRef] [PubMed]
- Verspreet, J.; Damen, B.; Broekaert, W.F.; Verbeke, K.; Delcour, J.A.; Courtin, C.M. A critical look at prebiotics within the dietary fiber concept. Annu. Rev. Food Sci. Technol. 2016, 7, 167–190. [Google Scholar] [CrossRef]
- Pasarin, D.; Ghizdareanu, A.-I.; Teodorescu, F.; Rovinaru, C.; Banu, A. Characterization of pectin oligosaccharides obtained from citrus peel pectin. Fermentation, 2023, 9, 312. [Google Scholar] [CrossRef]
- Kaur, B.; Panesar, P.S.; Anal, A.K. Standardization of ultrasound assisted extraction for the recovery of phenolic compounds from mango peels. J. Food Sci. Tech. 2022, 59, 2813–2820. [Google Scholar] [CrossRef]
- Wandee, Y.; Uttapap, D.; Mischnik, P.; Rungsardthong, V. Production of pectic-oligosaccharides from pomelo peel pectin by oxidative degradation with hydrogen peroxide. Food Chem. 2021, 348, 129078. [Google Scholar] [CrossRef]
- Gamonpilas, C.; Buathongjan, C.; Kirdsawasd, T.; Rattanaprasert, M.; Klomtun, M.; Phonsatta, N.; Methacanon, P. Pomelo pectin and fiber: Some perspectives and applications in food industry. Food Hydrocoll. 2021, 120, 106981. [Google Scholar] [CrossRef]
- Sivamani, S.; Binnal, P.; Roy, C.; Al Khaldi, A.; Al Hamar, F.; Maran, J.P.; Sivarajasekar, N.; Rajeshkumar, G.; Al Dhabi, N.A.; Karuppiah, P. Optimization and characterization of pectin recovered from (Persea americana) peel using statistical and nonstatistical techniques. Biomass Conv. Bioref. 2023, 13, 6501–6514. [Google Scholar] [CrossRef]
- Cosgove, D.J. Diffuse growth of plant cell walls. Plant Physiol. 2018, 176, 16–27. [Google Scholar] [CrossRef] [PubMed]
- Lampugnani, E.R.; Khan, G.A.; Somssich, M.; Persson, S. Building a plant cell wall at a glance. J. Cell. Sci. 2018, 131. [Google Scholar] [CrossRef] [PubMed]
- Eliaz, I.; Hotchkiss, A.T.; Fishman, M.L.; Rode, D. The effect of modified citrus pectin on urinary excretion of toxic elements. Phytother. Res. 2006, 20, 859–864. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Li, S.; Dong, Y.; Zhu, R.; Lin, Y. Antioxidant activity of penta-oligogalacturonide isolated from haw pectin, supressed triglyceride synthesis in mice feed with a high-fat diet. Food Chem. 2014, 145, 335–341. [Google Scholar] [CrossRef] [PubMed]
- Humerez-Flores, J.N.; Kyomugasho, C.; Gutiérrez-Ortiz, A.A.; De Bie, M.; Panozzo, A.; Van Loey, A.M.; Moldenaers, P.; Hendrickx, M.E. Production and molecular characterization of tailored citrus pectin-derived compounds. Food Chem. 2022, 367, 130635. [Google Scholar] [CrossRef] [PubMed]
- Voragen, A.G.J.; Coenen, G.-J.; Verhoef, R.P.; Schols, H.A. Pectin, a versatile polysaccharide present in plant cell walls. Struct. Chem. 2009, 20, 263–275. [Google Scholar] [CrossRef]
- Zdunek, A.; Pieczywek, P.M.; Cybulska, J. The primary, secondary and structures of higher levels of pectin polysaccharides. Compr. Rev. Food Sci. Food Saf. 2021, 20, 1101–1117. [Google Scholar] [CrossRef] [PubMed]
- Babbar, N. Dejonghe, W.; Gatti, M.; Sforza, S.; Elst, K. Pectic oligosaccharides from agricultural by-products: Production, characterization and health benefits. Crit. Rev. Biotechnol. 2016, 36, 594–606. [Google Scholar] [CrossRef] [PubMed]
- Muller, S.; Concha, D.; Vasquez, P.; Rodriguez-Nuñez, K.; Martinez, R.; Bernal, C. Effect of immobilization of pectinase on the molecular weight distribution of pectin oligosaccharides obtained from citrus pectin. Biocatal. Agric. Biotechnol. 2022, 43, 102389. [Google Scholar] [CrossRef]
- Hamaker, B.R.; Tuncil, Y.E. A perspective on the complexity of dietary fiber structures and their potential effect on gut microbiota. J. Mol. Biol. 2014, 426, 3838–3850. [Google Scholar] [CrossRef] [PubMed]
- Zhu, K.; Mao, G.; Wu, D.; Yu, C.; Cheng, H.; Xiao, H.; Ye, X.; Linhardt, R.J.; Orfila, C.; Chen, S. Highly branched RG-I domain enrichment is indispensable for pectin mitigating against high-fat diet-induced obesity. J. Agric. Food Chem. 2020, 68, 8688–8701. [Google Scholar] [CrossRef]
- Gullón, B.; Gómez, B.; Martínez-Sabajanes, M.; Yáñez, R.; Parajó, J.C.; Alonso, J.L. Pectic oligosaccharides: Manufacture and functional properties. Trends Food Sci. Technol. 2013, 30, 153–161. [Google Scholar] [CrossRef]
- Vogt, L.M.; Sahasrabudhe, N.M.; Ramasamy, U.; Meyer, D.; Pullens, G.; Faas, M.M.; Venema, K.; Schols, H.A.; de Vos, P. The impact of lemon pectin characteristics on TLR activation and T84 intestinal epithelial cell barrier function. J. Funct. Foods, 2016, 22, 398–407. [Google Scholar] [CrossRef]
- Sahasrabudhe, N.M.; Beukema, M.; Tian, L.; Troost, B.; Scholte, J.; Bruininx, E.; Bruggeman, G.; van den Berg, M.; Schurink, A.; Schols, H.A.; Faas, M.M.; de Vos, P. Dietary fiber pectin directly blocks tool-like receptor 2-1 and prevents doxorubicin-included ileitis. Front. Immunol. 2018, 9, 383. [Google Scholar] [CrossRef] [PubMed]
- Jiang, T.; Gao, X.; Wu, C.; Tian, F.; Lei, Q.; Bi, J.; Xie, B.; Wang, H.Y.; Chen, S.; Wang, X. Apple-derived pectin modulates gut microbiota, improves gut barrier function, and attenuates metabolic endotoxemia in rats with diet-induced obesity. Nutrients, 2016, 8, 126. [Google Scholar] [CrossRef]
- Naqash, F.; Masoodi, F.A.; Rather, S.A.; Wani, S.M.; Gani, A. Emerging concepts in the nutraceutical and functional properties of pectin – A Review. Carbohydr. Polym. 2017, 168, 227–239. [Google Scholar] [CrossRef] [PubMed]
- Basak, S.; Annapure, U.S. Trends in “green” and novel methods of pectin modification – A review. Carbohydr. Polym. 2022, 278, 118967. [Google Scholar] [CrossRef]
- Sila, D.N.; Van Buggenhout, S.; Duvetter, T.; Fraeye, I.; De Roeck, A.; Van Loey, A.; Hendrickx, M. Pectins in processed fruits and vegetables: Part II-structure-function relationships. Compr. Rev. 2009, 8, 86–104. [Google Scholar] [CrossRef]
- Pascale, N.; Gu, F.; Larsen, N.; Jespersen, L.; Respondek, F. The potential of pectins to modulate the human gut microbiota evaluated by in vitro fermentation. Nutrients, 2022, 14, 3629. [Google Scholar] [CrossRef] [PubMed]
- Zykwinska, A.; Boiffard, M.-H.; Kontkanen, H.; Buchert, J.; Thibault, J.-F.; Bonnin, E. Extraction of green labeled pectins and pectic oligosaccharides from plant byproducts. J. Agric. Food Chem. 2008, 56, 8926–8935. [Google Scholar] [CrossRef]
- Arrutia, F.; Adam, M; Calvo-Carrascal, M. Á.; Mao, Y.; Binner, E. Development of a continuous-flow system for microwave-assisted extraction of pectin-derived oligosaccharides from food waste. Chem. Eng. J. 2020, 395, 125056. [Google Scholar] [CrossRef]
- Chandel, V.; Biswas, D.; Roy, S.; Vaidya, D.; Verma, A.; Gupta, A. Current advancements in pectin: Extraction, properties and multifunctional applications. Foods 2022, 11, 2683. [Google Scholar] [CrossRef]
- Mao, G.; Li, S.; Orfila, C.; Shen, X; Zhou, S. ; Linhardt, R.J.; Ye, X.; Chen, S. Depolymerized RG-I-enriched pectin from citrus segment membranes modulates gut microbiota, increases SCFA production, and promotes the growth of Bifidobacterium spp., Lactobacillus spp. and Faecalibacterium spp. Food Funct. 2019, 10, 7828–7843. [Google Scholar] [CrossRef]
- Gómez, B.; Yáñez, R.; Parajó, J.C.; Alonso, J.L. Production of pectin-derived oligosaccharides from lemon peels by extraction, enzymatic hydrolysis and membrane filtration. J. Chem. Technol. Biotechnol. 2016, 91, 234–247. [Google Scholar] [CrossRef]
- Gómez, B.; Gullón, B.; Yáñez, R.; Schols, H.; Alonso, J.L. Prebiotic potential of pectins and pectic oligosaccharides derived from lemon peel wastes and sugar beet pulp: A comparative evaluation. J. Funct. Foods 2016, 20, 108–121. [Google Scholar] [CrossRef]
- Wang, Z.; Xu, B.; Luo, H.; Meng, K.; Wang, Y.; Liu, M.; Bai, Y.; Yao, B.; Tu, T. Production pectin oligosaccharides using Humicola insolens Y1-derived unsual pectate lyase. J. Biosci. Bioeng. 2020, 129, 16–22. [Google Scholar] [CrossRef]
- Zhang, S.; Hu, H.; Wang, L.; Liu, F.; Pan, S. Preparation and prebiotic potential of pectin oligosaccharides obtained from citrus peel pectin. Food Chem. 2018, 232–237. [Google Scholar] [CrossRef]
- Pham, V.T.; Catayud, M.; Rotsaert, C.; Seifert, N.; Richard, N.; van der Abbeele, P.; Marzorati, M.; Steinert, R.E. T.; Catayud, M.; Rotsaert, C.; Seifert, N.; Richard, N.; van der Abbeele, P.; Marzorati, M.; Steinert, R.E. Antioxidant vitamins and prebiotic FOS and XOS differentially shift microbiota composition and function and improve intestinal epithelial barrier in vitro. Nutrients 2021, 13, 1125. [Google Scholar] [CrossRef]
- Conçalves, D.A.; González, A.; Roupar, D.; Teixeira, J.A.; Nobre, C. How prebiotics have been produced from agro-industrial waste: An overview of the enzymatic technologies applied and the models used to validate their health claim. Trends. Food Sci. 2023, 135, 74–92. [Google Scholar] [CrossRef]
- Lunn, J.; Buttriss, J.L. Carbohydrates and dietary fibre. Nutr. Bull. 2007, 32, 21–64. [Google Scholar] [CrossRef]
- Dai, F.-J.; Chau, C.-F. Classification and regulatory perspectives of dietary fiber. J. Food Drug Anal. 2017, 25, 37–42. [Google Scholar] [CrossRef]
- Kang, H.J.; Kwon, J.H.; Ahn, D.U.; Lee, J.W.; Lee, W.K.; Jo, C. Effect of citrus pectin oligosaccharide prepared by irradiation on high cholesterol diet B6.KOR-ApoE mice. Food Sci. Biotechnol. 2009, 18, 884–888. [Google Scholar]
- Zhang, S.; Hu, H.; He, W.; Muhammad, Z.; Wang, L.; Liu, F.; Pan, S. Regulatory roles of pectin oligosaccharides on immunoglobulin production in healthy mice mediated by gut microbiota. Mol. Nutr. Food Res. 2019, 63, 1801363. [Google Scholar] [CrossRef]
- Prandi, B.; Baldassarre, S.; Babbar, N.; Bancalari, E.; Vandezande, P.; Hermans, D.; Bruggeman, G.; Gatti, M.; Elst, K.; Sforza, S. Pectin oligosaccharides from sugar beet pulp: Molecular characterization and potential prebiotic activity. Food Funct. 2018, 9, 1557. [Google Scholar] [CrossRef] [PubMed]
- Olano-Martin, E.; Gibson, G.R.; Rastall, R.A. Comparison of the in vitro bifidogenic properties of pectins and pectic-oligosaccharides. J. Appl. Microbiol. 2002, 93, 505–511. [Google Scholar] [CrossRef]
- Tan, H.; Chen, W.; Liu, Q.; Yang, G.; Li, K. Pectin oligosaccharides ameliorate colon cancer by regulating oxidative stress- and inflammation-activated signaling pathways. Front. Immunol. 2018, 9, 1504. [Google Scholar] [CrossRef]
- Darcy, J.L.; Washburne, A.D.; Robeson, M.S.; Prest, T.; Schmidt, S.K.; Lozupone, C.A. A phylogenic model for the recruitment of species into microbial communities and application to studies of the human microbiome. ISME J. 2020, 14, 1359–1368. [Google Scholar] [CrossRef]
- Scholz-Ahrens, K.E.; Ade, P.; Marten, B.; Weber, P.; Timm, W.; Açil, Y.; Glüer, C.-C.; Schrezenmeir, J. Prebiotics, probiotics, and synbiotics affect mineral absorption, bone mineral content, and bone structure. J. Nutr. 2007, 137, 838S–846S. [Google Scholar] [CrossRef]
- Fan, Z.; Chen, X.; Liu, T.; Yu, Q.; Song, Z.; Wang, F.; Li, T. Pectin oligosaccharides improved lipid metabolism in white adipose tissue of high-fat diet fed mice. Food Sci. Biotechnol. 2022, 31, 1197–1205. [Google Scholar] [CrossRef]
- Li, S.; Li, T.; Jia, Y.; Zhu, R.; Wang, N.; Jin, S.; Guo, M. Fractination and structural characterization of haw pectin oligosaccharides. Eur. Food Res. Technol. 2011, 233, 731–734. [Google Scholar] [CrossRef]
- Zhu, R.; Sun, X.; Zhang, Y.; Yang, T.; Wang, C.; Zhang, J.; Duan, Z.; Shang, F.; Fan, J.; Liu, Y.; Peng, X.; Wang, N.; Chen, G. Effect of pectin oligosaccharides supplementation on infant formulas: The storage stability, formation and intestinal absorption of advanced glycation end products. Food Chem. 2022, 373 (Part B), 131571. [Google Scholar] [CrossRef]
- Kong, C.; Faas, M.M.; de Vos, P.; Akkerman, R. Impact of dietary fibers in infant formulas on gut microbiota on the intestinal immune barrier. Food Funct. 2022, 11, 9445. [Google Scholar] [CrossRef]
- Fanaro, S.; Jelinek, J.; Stahl, B.; Boehm, G.; Kock, R.; Vigi, V. Acidic oligosaccharides from pectin hydrolysate as new component for infant formulae: Effect of intestinal flora, stool characteristics, and pH. J. Pediatr. Gastroenterol. Nutr. 2005, 41, 186–190. [Google Scholar] [CrossRef] [PubMed]
- Poulsen, M.W.; Hedegaard, R.V.; Andersen, J.M.; de Courten, B.; Bügel, S.; Nielsen, J.; Skibsted, L.H.; Dragsted, L.O. Advanced glycation endproducts in food and their effects on health. Food Chem. Toxicol. 2013, 60, 10–37. [Google Scholar] [CrossRef] [PubMed]
- Zhu, R.; Hong, M.; Zhuang, C.; Zhang, L.; Wang, C.; Liu, J.; Duan, Z.; Shang, F.; Hu, F.; Li, T.; Ning, C.; Chen, G. Pectin oligosaccharides from hawtorn (Crataefus pinnatifida Bunge. Var. major) inhibit the formation of advanced glycation end products in infant formula milk powder. Food Funct. 2019, 10, 8081. [Google Scholar] [CrossRef] [PubMed]
- Gibson, G.R.; Roberfroid, M.B. Dietary modulation of the human colonic microbiota: Introducing the concept of prebiotics. J. Nutr. 1995, 125, 1401–1412. [Google Scholar] [CrossRef] [PubMed]
- Gibson, G.R. Fibre and effects on probiotics (the probiotic concept). Clin. Nutr. Suppl. 2004, 1, 25–31. [Google Scholar] [CrossRef]
- Bindels, L.B.; Delzenne, N.M.; Cani, P.D.; Walter, J. Towards a more comprehensive concept for prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Tingirikari, J.M.R. Microbiota-accessible pectic poly- and oligosaccharides in gut health. Food Funct. 2018, 9, 5059. [Google Scholar] [CrossRef]
- Chung, W.S.F.; Walker, A.W.; Louis, P.; Parkhill, J.; Vermeiren, J.; Bosscher, D.; Duncan, S.H.; Flint, H.J. Modulation of the human gut microbiota by dietary fibres occurs at the species level. BMC Biology 2016, 14, 3. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Liang, R.-H.; Lin, W.; Li, T.; Lin, C.M.; Wu, S.-S.; Wang, Z.-J. Pectic-oligosaccharides prepared by dynamic high pressure microfluidization and their in vitro fermentation properties. Carbohydr. Polym. 2013, 91, 175–182. [Google Scholar] [CrossRef]
- Duncan, S.H.; Holtop, G.; Lobley, G.E.; Colder, A.G.; Stewart, C.S.; Flint, H.J. Contribution of acetate to butyrate formation by human faecal bacteria. Brit. J. Nutr. 2004, 91, 915–923. [Google Scholar] [CrossRef]
- Manderson, K.; Pinart, M.; Tuohy, K.M.; Grace, W.E.; Hotchkiss, A.T.; Widmer, W.; Yadhav, M.P.; Gibons, G.R.; Rastall, R.A. In vitro determination of probiotic properties of oligosaccharides derived from an orange juice manufacturing by-product stream. Appl. Environ. Microbiol. 2005, 71. [Google Scholar] [CrossRef]
- Bang, S.-J.; Kim, G.; Lim, M.Y.; Song, E.-J.; Jung, D.-H.; Kum, J.-S.; Nam, Y.-D.; Park, C.-S.; Seo, D.-H. The influence of in vitro pectin fermentation on the human fecal microbiome. AMB Express 2018, 8, 98. [Google Scholar] [CrossRef]
- Gómez, B.; Gullón, B.; Remoroza, B.; Schols, H.A.; Parejó, J.C.; Alonso, J.L. Purification, characterization and prebiotic properties of pectic oligosaccharides from orange peel wastes. J. Agric. Food Chem. 2014, 62, 9769–9782. [Google Scholar] [CrossRef] [PubMed]
- An, R.; Wilms, E.; Smolinska, A.; Hermes, G.D.A; Masclee, A.A.M; de Vos, P.; Schols, H.A; van Schooten, F.J.; Smidt, H.; Jonkers, D.M.A.E.; Zoetendal, E.G.; Troost, F.J. Sugar beet pectin supplementation did not alter profiles of fecal microbiota and exhaled breath in healthy young adults and healthy elderly. Nutrients 2019, 11, 2193. [Google Scholar] [CrossRef]
- Leijdekkers, A.G.M.; Aguirre, M.; Venema, K.; Bosch, G.; Gruppen, H.; Schols, H.A. In vitro fermentability of sugar beet pulp derived oligosaccharides using human and pig fecal inocula. J. Agric. Food Chem. 2014, 62, 1079–1087. [Google Scholar] [CrossRef] [PubMed]
- Gulfi, M.; Arrigoni, E.; Amedó, R. Influence of structure on in vitro fermentability of commercial pectins and partially hydrolysed pectin preparations. Carbohydr. Polym. 2005, 59, 247–255. [Google Scholar] [CrossRef]
- Aguirre, M.; Jonkers, D.M.A.E.; Troost, F.J.; Roeselers, G.; Venema, K. In vitro characterization of the impact of different substrates on metabolite production, energy extraction and composition of gut microbiota from lean to obese subjects. PLoS ONE 2014, 9, e113864. [Google Scholar] [CrossRef] [PubMed]
- Holck, J.; Lorentzen, A.; Vigsnæs, L.K.; Licht, T.R.; Mikkelsen, J.D.; Meyer, A.S. Feruloylated and nonferuloylated arabino-oligosaccharides from sugar beet pectin selectively stimulate the growth of Bifidobacterium spp. in human fecal in vitro fermentations. J. Agric. Food Chem. 2011, 59, 6511–6519. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Liu, N.; Yang, Z.; Zhao, K.; Pang, H.; Shao, K. Zhou, Z.; Li, S.; He, N. Preventive effect of pectic oligosaccharides on acute colitis model mice: Modulating epithelial barrier, gut microbiota and Treg/Th17 balance. Food Funct. 2022, 13, 9999–10012. [Google Scholar] [CrossRef] [PubMed]
- Ho, Y.-Y.; Lin, C.-M.; Wu, M.-C. Evaluation of the prebiotic effects of citrus pectin hydrolysate. J. Food Drug Anal. 2017, 25, 550–558. [Google Scholar] [CrossRef]
- Gao, M.; Wang, X.; Lin, J.; Lin, X.Y.; Qi, D.; Luo, Y.; Aheyeli-kai, Y.; Ma, H. Separation, structural identification and antibacterial activity of pectin oligosaccharides derived from seed melon. Food Biosc. 2023, 53, 102616. [Google Scholar] [CrossRef]
- Amarasekera, M.; Prescott, S.L.; Palmer, D.J. Nutrition in early life, immune-programming and allergies: The role of epigenetics. Asian. Pac. J. Allergy Immunol. 2013, 31, 175–182. [Google Scholar]
- Despres, J.; Forano, E.; Lepercq, P.; Comtet-Marre, S.; Jubelin, G.; Chambon, C.; Yeoman, C.J.; Berg Miller, M.E.; Fields, C.J.; Martens, E.; Terrapon, N.; Henrissat, B.; White, B.A.; Mosoni, P. Xylan degradation by the human gut Bacteroides xylanisolvens XB1AT involves two distinct gene clusters that are linked at the transcriptional level. BMC Genom. 2016, 17, 326. [Google Scholar] [CrossRef]
- Chung, W.S.F.; Meijerink, M.; Zeuner, B.; Holck, J.; Louis, P.; Meyer, A.S.; Wells, J.M.; Flint, H.J. Prebiotic potential of pectin and pectic oligosaccharides to promote anti-inflammatory commensal bacteria in the human colon. FEMS Microbiol. Ecol. 2017, 93, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Holck, J.; Hjernø, K.; Lorentzen, A.; Visnæs, K.; Hemmingsen, L.; Licht, T.R.; Mikkelsen, J.D.; Meyer, A.S. Tailored enzymatic production of oligosaccharides from sugar beet pectin and evidence of differential effects of a single DP chain length difference on human faecal microbiota composition after in vitro fermentation. Process Biochem. 2011, 46, 1039–1049. [Google Scholar] [CrossRef]
- Fotschki, B.; Juśkiewicz, J.; Jurgoński, A.; Sójka, M. Fructo-oligosaccharides and pectins enhance beneficial effects of raspberry polyphenols in rats with nonalcoholic fatty liver. Nutrients 2021, 13, 833. [Google Scholar] [CrossRef]
- Gu, F.; Larsen, N.; Pascale, N.; Petersen, S.A.; Khakimov, B.; Respondek, F.; Jespersen, L. Age-related effects of the modulation of gut microbiota by pectins and their derivatives: An in vitro study. Front Microbiol. 2023, 14, 1207837. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Martínez, I.; Walter, J.; Keshavarzian, A.; Rose, D.J. In vitro characterization of the impact of selected dietary fibers on fecal microbiota composition and short chain fatty acid production. Anaerobe 2013, 23, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Wilkowska, A.; Motyl, I.; Antczak-Chrobot, A.; Wojtczak, M.; Nowak, A.; Czyżowska, A.; Motyl, W. Influence of human age on the prebiotic effect of pectin-derived oligosaccharides obtained from apple pomace. Fermentation, 2021, 7, 224. [Google Scholar] [CrossRef]
- Luis, A.S.; Briggs, J.; Zhang, X.; Farnell, B.; Ndeh, D.; Labourel, A.; Baslé, A.; Cartmell, A.; Terrapon, N.; Stoott, K.; Lowe, E.C.; McLean, R.; Shearer, K.; Schükel, J.; Venditto, I.; Ralet, M.-C.; Henrissat, B.; Martens, E.C.; Mosimann, S.C.; Abbott, D.W.; Gilbert, H.J. Dietary pectic glycans are degraded by coordinated enzyme pathways in human colonic Bacteroides. Nat. Microbiol. 2018, 3, 210–219. [Google Scholar] [CrossRef]
- Shinohara, K.; Ohashi, Y.; Kawasumi, K.; Terada, A.; Fujisawa, T. Effect of apple intake on fecal microbiota and meatbolites in humans. Anaerobe 2010, 16, 510–515. [Google Scholar] [CrossRef]
- Paturi, G.; Butts, C.A.; Stoklosinski, H.; Herath, T.D.; Monro, J.A. Short-term feeding of fermentable dietary fibres influences the gut microbiota composition and metabolic activity in rats. Int. J. Food Sci. + Technol. 2017, 52, 2572–2581. [Google Scholar] [CrossRef]
- Koropatkin, N.M.; Cameron, E.A.; Martens, E.C. How glycan metabolism shapes the human gut microbiota. Nat. Rev. Microbiol. 2012, 10, 323–335. [Google Scholar] [CrossRef] [PubMed]
- Trompette, A.; Gollwitzer, E.S.; Yadava, K.; Sichelstiel, A.K.; Sprenger, N. Ngom-Bru, C.; Blanchard, C.; Junt, T.; Nicod, L.P.; Harris, N.L.; Marsland, B.J. Gut microbiota metabolism of dietary fiber influences allergic airway disease and hematopoesis. Nat. Med. 2014, 20, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Zhang, S.; Lin, F. Zhang, P.; Muhammad, Z.; Pan, S. Role of the gut microbiota and their metabolites in modulating the cholesterol-lowering effects of citrus pectin oligosaccharides in C57BL/6 mice. J. Agric. Food Chem. 2019, 67, 11922–11930. [Google Scholar] [CrossRef] [PubMed]
- Reichart, N.; Duncan, S.H.; Young, P.; Brlenguer, A.; McWilliam Leitch, C.; Scott, K.P.; Flint, H.J.; Louis, P. Phytogenetic distribution of three pathways for propionate production within the human gut microbiota. ISME J. 2014, 8, 1323–1335. [Google Scholar] [CrossRef]
- Hirata, S.-I.; Kunisawa, J. Gut microbiome, metabolome and allergic diseases. Allergol. Int. 2017, 66, 523–528. [Google Scholar] [CrossRef] [PubMed]
- Jakobsdottir, G.; Jädert, C.; Holm, L.; Nyman, M.E. Propionic and butyric acids, formed in the caecum of rats fed highly fermentable dietary fibre, are reflected in portal and aortic serum. Br. J. Nutr. 2013, 110, 1565–1572. [Google Scholar] [CrossRef] [PubMed]
- Takenaka, T.; Muto, O.; Yatsunami, K.; Echigo, T. Antibacterial activity of pectin hydrolyzates. Nippon Shokuhin Kogyo Gakkaishi, 1994, 41, 785–792. [Google Scholar] [CrossRef]
- Foti, P.; Ballistreri, G.; Timpanaro, N.; Rapisadra, P.; Romeo, F.V. Prebiotic effects of citrus pectic oligosaccharides. Nat. Prod. Res. 2022, 36, 3173–3176. [Google Scholar] [CrossRef]
- Sjögren, Y.M.; Jenmalmm, M.C.; Böttcher, M.F.; Björkstén, B.; Sverremark-Ekström, E. Altered early infant gut microbiota in children developing allergy up to 5 years of age. Clin. Exp. Allergy 2009, 39, 518–526. [Google Scholar] [CrossRef]
- Sung, J.; Kim, S.; Cabatbat, J.J.T; Jang, S.; Jin, Y.-S.; Jung, G.Y.; Chia, N.; Kim, P.-J. Global metabolic interaction network of the human gut microbiota for context-specific community-scale analysis. Nat. Commun. 2017, 8, 15393. [Google Scholar] [CrossRef]
- Goyal, A.; Wang, T.; Dubinkina, V.; Maslov, S. Ecology-guided prediction of cross-feeding interactions in the human gut microbiome. Nat. Commun. 2021, 12, 1335. [Google Scholar] [CrossRef] [PubMed]
- Guo, K.; Yao, Z.; Tang, T. Intestinal microbiota-mediated dietary fiber bioavailability. Front. Nutr. 2022, 9, 1003571. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Tomé, S.; Ortega-Moreno, L.; Chaparro, M.; Gisbert, J.P. Gut microbiota and dietary factors as modulators of the mucus layer in inflammatory bowel disease. Int. J. Mol. Sci. 2021, 22, 10224. [Google Scholar] [CrossRef] [PubMed]
- Blanco-Pérez, F.; Steióerwald, H.; Schülke, S.; Vieths, S.; Toda, M.; Scheurer, S. The dietary fiber pectin: Health benefits and potential for the treatment of allergies by modulation of gut microbiota. Curr. Allergy Asthma Rep. 2021, 21, 43. [Google Scholar] [CrossRef]
- Cai, J.; Sun, L.; Gonzalez, F.J. Gut microbiota-derived bile acids in intestinal immunity, inflammation and tumorigenesis. Cell Host Microbe 2022, 30, 289–300. [Google Scholar] [CrossRef]
- Cohen, Y.; Elinay, E. Dietary fibers & immunity – more than meets the eye. Cell Res. 2023, 33, 411–412. [Google Scholar] [CrossRef]
- Zuurveld, M.; van Witzenburg, N.P.; Garssen, J.; Folkerts, G.; Sthal, B.; van’t Land, B.; Willemsen, L.E.M. Immunomodulation by human milk oligosaccharides: The potential role in prevention of allergic diseases. Front. Immunol. 2020, 11, 801. [Google Scholar] [CrossRef]
- Bernard, H.; Desseyn, J.-L.; Gottrand, F.; Stahl, B.; Bartke, N.; Husson, M.-O. Pectic-derived acidic oligosaccharides improve the outcome of Pseudomonas aeruginosa lung infection in C57BL/6 Mice. PLoS ONE 2015, 10, e0139686. [Google Scholar] [CrossRef]
- Wikiera, A.; Irla, M.; Mika, M. Health promoting properties of pectin. Postepy Hig. Med. Dosw. 2014, 68, 590–596. [Google Scholar] [CrossRef] [PubMed]
- Mowat, A.M.; Agace, W.W. Regional specialization within the intestinal immune system. Nat. Rev. Immunol. 2014, 14, 667–685. [Google Scholar] [CrossRef] [PubMed]
- Jakobsson, H.E.; Rodriguez-Piñeiro, A.M.; Schütte, A.; Ermund, A.; Boysen, P.; Bemark, M.; Sommer, F.; Bäckhed, F.; Hansson, G.C.; Johansson, M.E.V. The composition of the gut microbiota shapes the colon mucus barrier. EMBO Rep. 2015, 16, 164–177. [Google Scholar] [CrossRef]
- Johansson, M.E.V.; Phillipson, M.; Petersson, J.; Velcich, A.; Holm, L.; Hansson, G.C. The inner of the two Muc2 mucin dependent mucus layers in colon is devoid bacteria. PNAS 2008, 105, 15064–15069. [Google Scholar] [CrossRef]
- Leclere, L.; Van Cutsem, P.; Michiels, C. Anti-cancer activities of pH- or heat – modified pectin. Front. Pharmacal. 2013, 4, 128. [Google Scholar] [CrossRef] [PubMed]
- Hino, S.; Sonoyama, K.; Bito, H.; Kawagishi, H.; Aoe, S.; Morita, T. Low-methoyl pectin stimulates small intestinal mucin secretion irrespective of goblet cell proliferation and is characterized by jejunum Muc2 upregulation in rats. J. Nutr. 2013, 143, 34–40. [Google Scholar] [CrossRef]
- Chen, Z.; Luo, J.; Li, J.; Kim, G.; Chen, E.S. , Xiao, S.; Snapper, S.B.; Bao, B.; An, D.; Blumberg, R.S.; Lin, C.-H.; Wang, S.; Zhong, J.; Lin, K.; Li, Q; Wu, C.; Kuchroo, V.K. Foxo1 controls gut homeostasis and commensalism by regulating mucus secretion. J. Exp. Med. 2021, 218, e20210324. [Google Scholar] [CrossRef]
- Peterson, L.W.; Artis, D. Intestinal epithelial cells: Regulators of barrier function and immune homeostasis. Nat. Rev. Immunol. 2014, 14, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Minello, V.L.; Minello, A.; Ficele, L.; Skublewska-D’Elia, A.; Dargenio, V.N.; Cristofori, F.; Francavilla, R. Gut immunobiosis and biomodulators. Nutrients 2023, 15, 2114. [Google Scholar] [CrossRef]
- Bernard, H.; Desseyn, J.-L.; Bartke, N.; Kleijnjans, L.; Stahl, B.; Belzer, C.; Knol, J.; Gottrand, F.; Husson, M.-O. Dietary pectin-derived acidic oligosaccharides improve the pulmonary bacterial clearance of Pseudomonas aeruginosa lung infection in mice by modulating intestinal microbiota and immunity. J. Infect. Dis. 2015, 211, 156–165. [Google Scholar] [CrossRef]
- Montilla, A.; Muñoz-Alamagro, N.; Villamiel, M. Chapter 6 – A new approach of functional pectin and pectic oligosaccharides: Role as antioxidant and anti-inflammatory compounds. [In] Current Advances for Development of Functional Foods Modulating Inflammation and Oxidative Stress. 2022, 105-120. [CrossRef]
- Xie, J.; Yu, R.; Qi, J.; Zhang, G.; Peng, X.; Luo, J. Pectin and inulin stimulated the mucus formation at a similar level: An omics-based comparative analysis. Food Sci. 2020, 85, 1939–1947. [Google Scholar] [CrossRef]
- Singh, G.; Brass, A.; Knight, C.G; Cruickshank, S.M. Gut eosinophils and their impact on the mucus-resident microbiota. Immunology 2019, 158, 194–205. [Google Scholar] [CrossRef]
- Jeurink, P.V.; van Esch, B.C.; Rijnierse, A.; Garssen, J.; Knippels, L.M. Mechanism underlying immune effects of dietary oligosaccharides. Am. J. Clin. Nutr. 2013, 98, 572S–577S. [Google Scholar] [CrossRef]
- Lee, K.H. , Song, Y.; Wu, W.; Yu, W.; Zhang, G. The gut microbiota, environmental factors and links to the development of food allergy. Clin. Mol. Allergy 2020, 18, 5. [Google Scholar] [CrossRef]
- Nance, C.L.; Deniskin, R.; Diaz, V.C.; Paul, M.; Anvari, S.; Anagnostou, A. The role of the microbiome in food allergy: A Review. Children 2020, 7, 50. [Google Scholar] [CrossRef] [PubMed]
- Guevara-Arauza, J.C.; Ornelas-Paz, J.J.; Pimentel-González, D.J.; Mendoza, S.R.; Guerra, R.E.S.; Paz Maldonado, L.M.T. Prebiotic effect of mucilage and pectic-derived oligosaccharides from Nopal (Opuntia ficus-indica). Food Sci. Biotechnol. 2012, 21, 997–1003. [Google Scholar] [CrossRef]
- Lee, J.H.; Shim, J.S.; Lee, J.S.; Kim, M.-K.; Chung, M.-S.; Kim, K.H. Pectin-like acidic polysaccharide from Panax ginseng with selective antiadhesive activity against pathogenic bacteria. Carbohydr Res. 2006, 341, 1154–1163. [Google Scholar] [CrossRef]
- Ouwerkerk, J.P.; de Vos, W.M.; Belzer, C. Glycobiome: Bacteria and mucus at the epithelial interface. Best. Pract. Res. Clin. Gastroenterol. 2013, 27, 25–38. [Google Scholar] [CrossRef]
Figure 1.
Classification of dietary fibre.
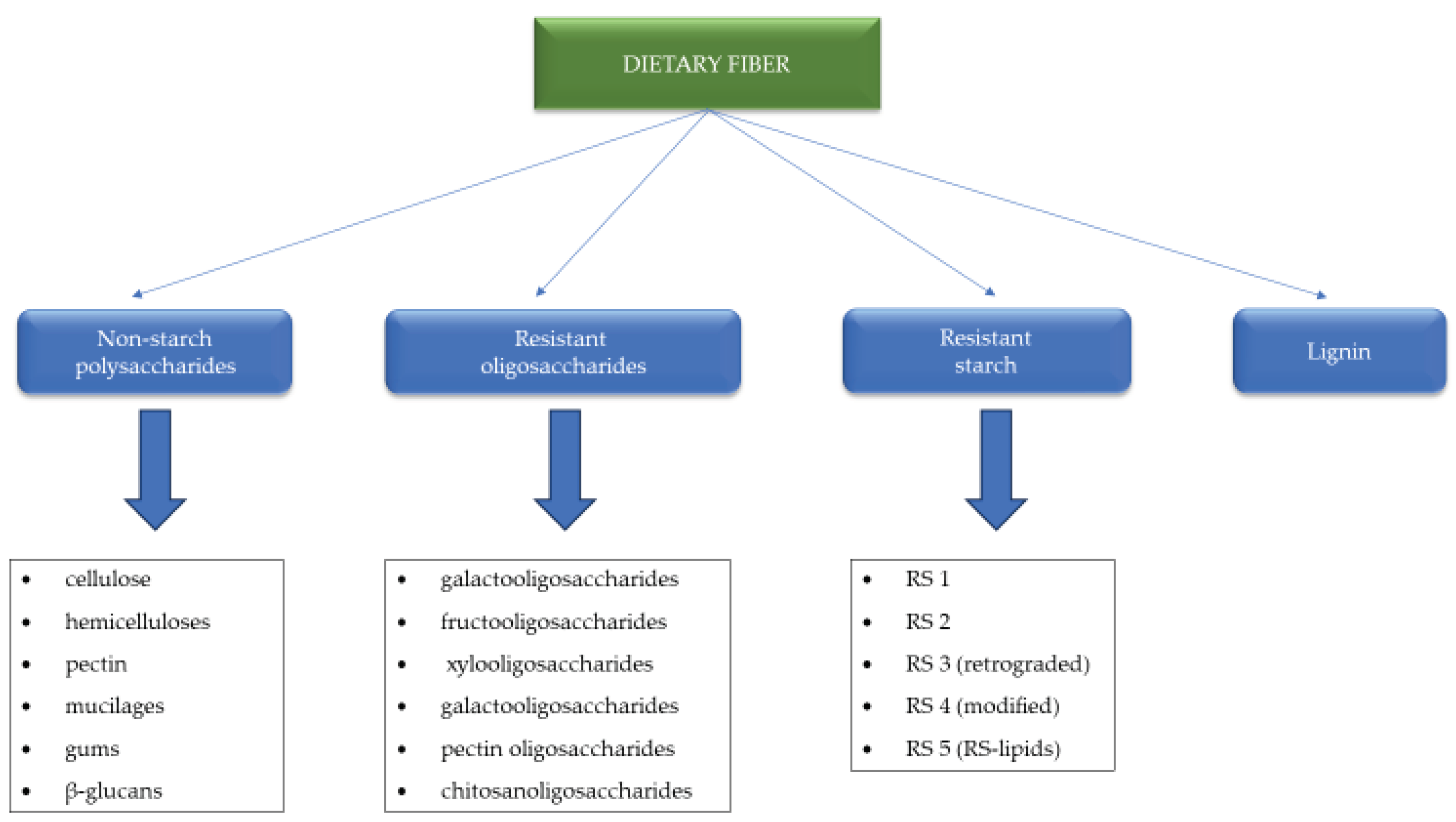
Figure 2.
Pectin content from various agro-waste sources.
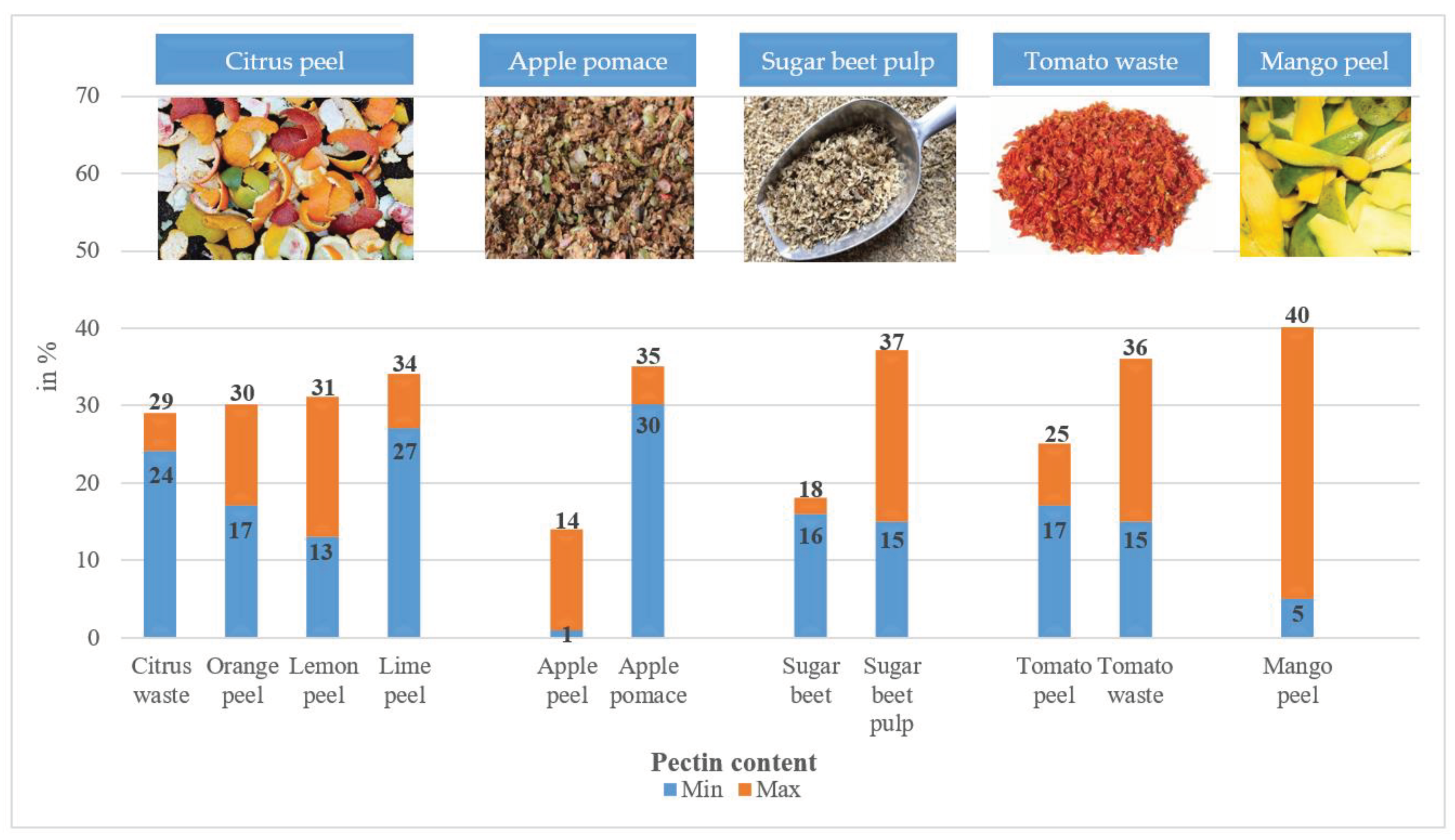
Figure 3.
Structure of pectin with characterization of smooth and hairy region [113].
Figure 3.
Structure of pectin with characterization of smooth and hairy region [113].
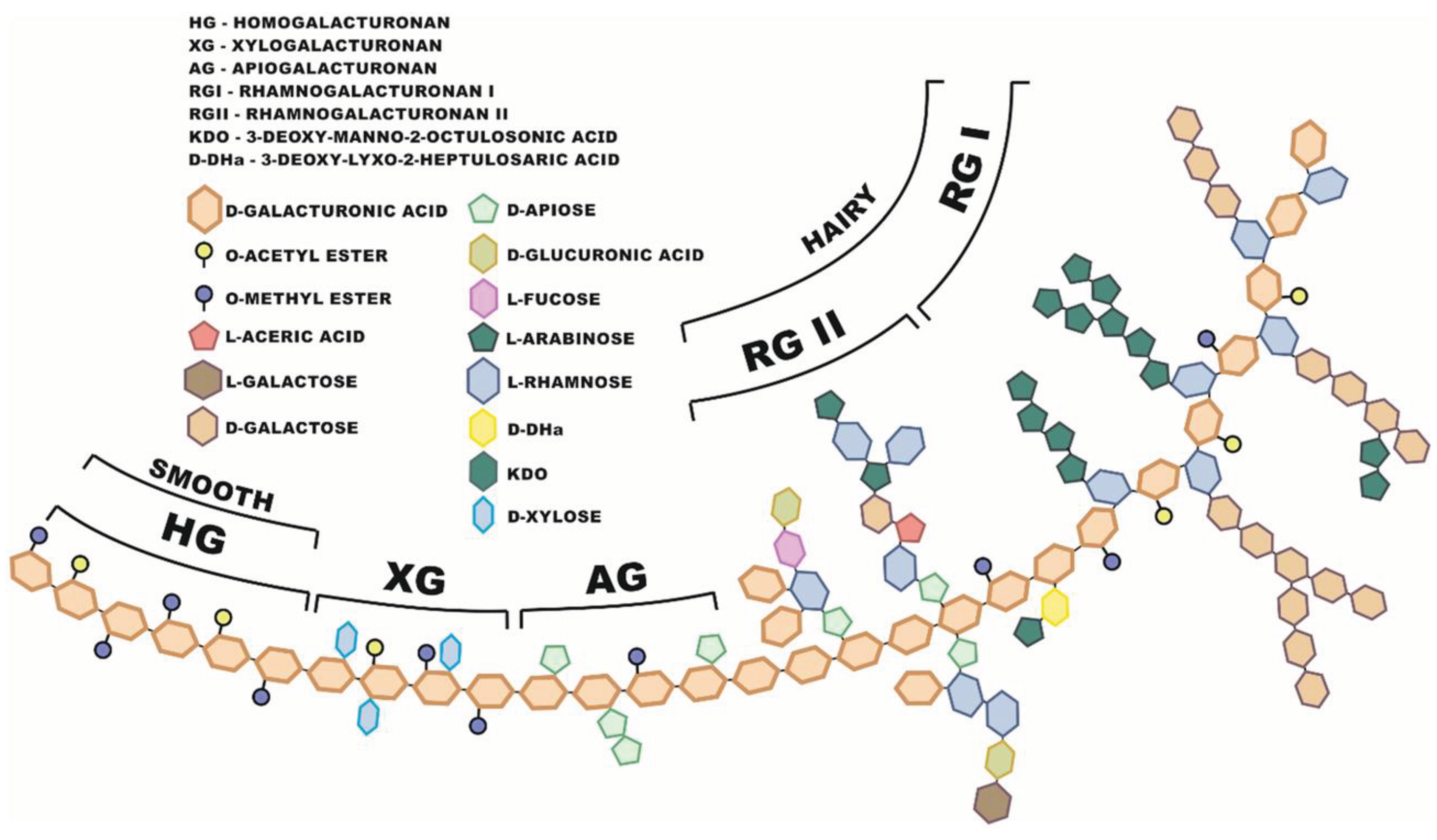
Figure 4.
Pectin oligosaccharides production and its effect for guts and GALT.
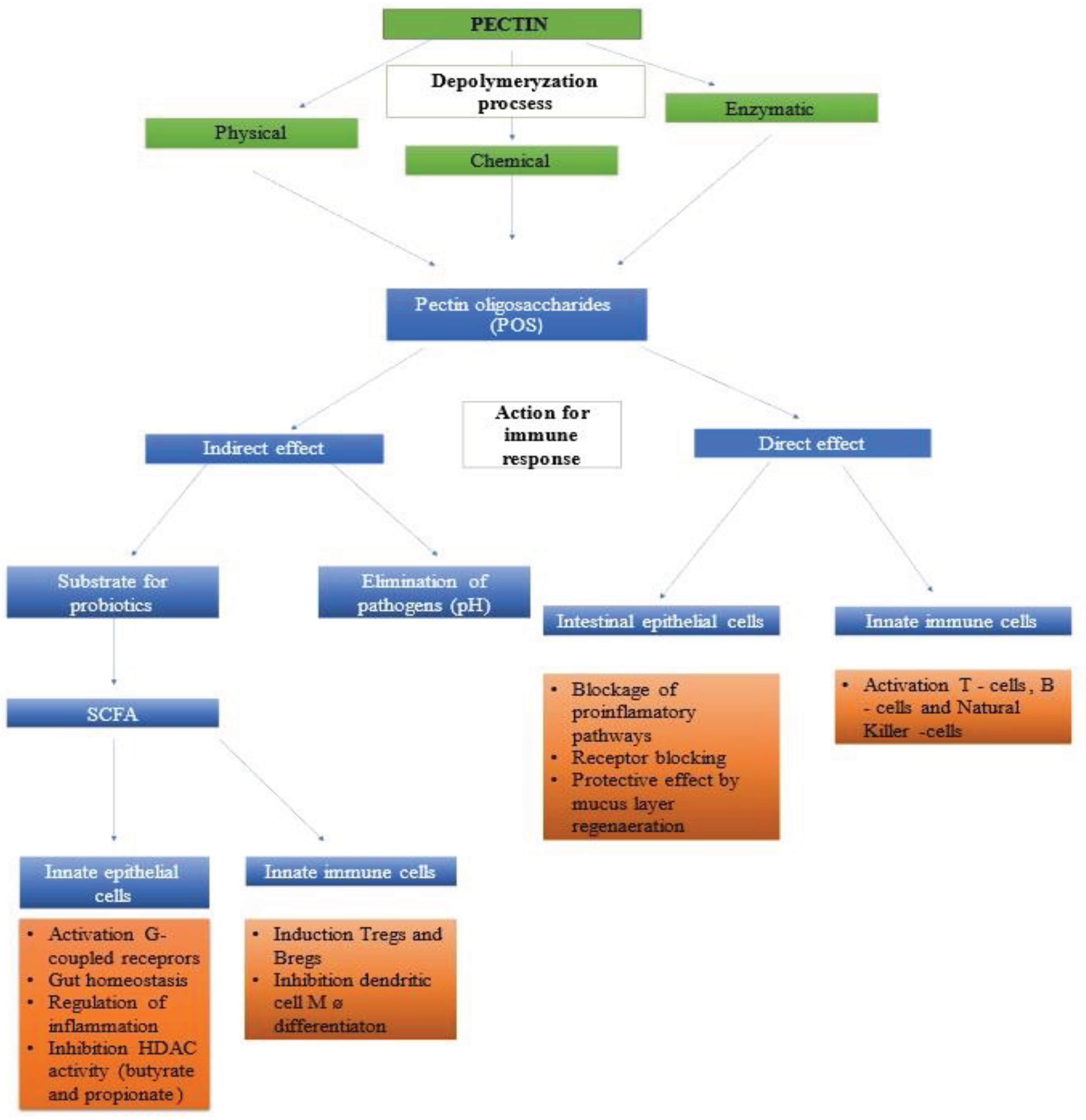

Table 1.
Physical properties of individual carbohydrates in comparison with cellulose.
| Item | Solubility | Fermentability | Viscosity |
|---|---|---|---|
| Cellulose | - | slow | - |
| β-glucan (cereals) | high | rapid | high |
| Inulin | variable | rapid | low |
| FOS | high | rapid | - |
| Pectin | high | rapid | high |
| POS | high | rapid | low |
Table 2.
Dietary fibre content in different products.
| Item | DF (g /100 g edible portion) | Percentage of TDF (%) | For 30 g TDF, person must eat in kg (single component of diet) |
|||
|---|---|---|---|---|---|---|
| Total | IDF | SDF | IDF | SDF | ||
| Flaxseed | 22.3 | 10.1 | 12.2 | 45 | 55 | 0.13 |
| Soybean | 21.5 | 19.4 | 2.1 | 90 | 10 | 0.14 |
| Barley | 19.5 | 15.7 | 3.8 | 81 | 19 | 0.15 |
| Raw white beans | 17.7 | 13.4 | 4.3 | 76 | 24 | 0.17 |
| Wheat grain | 12.6 | 10.2 | 2.3 | 81 | 18 | 0.24 |
| Corn | 11.9 | 10.5 | 1.4 | 88 | 12 | 0.25 |
| Raw lentils | 11.4 | 10.3 | 1.1 | 90 | 10 | 0.26 |
| Almond | 11.2 | 10.1 | 1.1 | 90 | 10 | 0.27 |
| Oats | 10.3 | 6.5 | 3.8 | 63 | 37 | 0.29 |
| Raw coconut | 9.0 | 8.5 | 0.5 | 94 | 6 | 0.33 |
| Dry roasted peanut | 8.0 | 7.5 | 0.5 | 94 | 6 | 0.38 |
| Beetroot | 7.8 | 5.4 | 2.4 | 69 | 31 | 0.38 |
| Sesame seed | 7.8 | 5.9 | 1.9 | 76 | 24 | 0.38 |
| Frozen green peas | 3.5 | 3.2 | 0.3 | 91 | 9 | 0.86 |
| Kiwi | 3.4 | 2.6 | 0.8 | 76 | 24 | 0.88 |
| Pear | 3.0 | 2.0 | 1.0 | 67 | 33 | 1.00 |
| Raw spinach | 2.6 | 2.1 | 0.5 | 81 | 19 | 1.15 |
| Raw carrot | 2.5 | 2.3 | 0.2 | 92 | 8 | 1.20 |
| Strawberry | 2.2 | 1.3 | 0.9 | 59 | 41 | 1.36 |
| Unpeeled apple | 2.0 | 1.8 | 0.2 | 90 | 10 | 1.50 |
| Green beans | 1.9 | 1.4 | 0.5 | 74 | 26 | 1.58 |
| Peach | 1.9 | 1.0 | 0.9 | 53 | 47 | 1.58 |
| Raw cauliflower | 1.8 | 1.1 | 0.7 | 61 | 39 | 1.67 |
| Mango | 1.8 | 1.1 | 0.7 | 61 | 39 | 1.67 |
| Oranges | 1.8 | 0.7 | 1.1 | 39 | 61 | 1.67 |
| Bananas | 1.7 | 1.2 | 0.5 | 71 | 29 | 1.76 |
| Plums | 1.6 | 0.7 | 0.9 | 44 | 56 | 1.88 |
| Raw celery | 1.5 | 1.0 | 0.5 | 67 | 33 | 2.00 |
| Rice dry | 1.3 | 1.0 | 0.3 | 77 | 23 | 2.31 |
| Potato without skin | 1.3 | 1.0 | 0.3 | 77 | 23 | 2.31 |
| Raw tomato | 1.2 | 0.8 | 0.4 | 67 | 33 | 2.50 |
| Pineapple | 1.2 | 1.1 | 0.1 | 92 | 8 | 2.50 |
| Grapes | 1.2 | 0.7 | 0.5 | 58 | 42 | 2.50 |
| Rice cooked | 0.7 | 0.7 | 0.0 | 100 | 0 | 4.29 |
| Peeled cucumber | 0.6 | 0.5 | 0.1 | 83 | 17 | 5.00 |
| Watermelon | 0.5 | 0.3 | 0.2 | 60 | 40 | 6.00 |
Table 3.
Effect of pectin and pectin oligosaccharides from similar sources on specific microorganisms growth and SCFA production.
Table 3.
Effect of pectin and pectin oligosaccharides from similar sources on specific microorganisms growth and SCFA production.
| Research | |||||
|---|---|---|---|---|---|
| with pectin | Microbiota | SCFA | with POS | Microbiota | SCFA |
| Apple pectin [156] |
↑Bacteroidetes | *Acetate | Apple POS [157] |
↑Bifidobacteria | |
| ↑Firmicutes | *Propionate | ↑Lactobacilli | ↑Acetate | ||
| ↑Eubacterium eligens |
*Butyrate |
↓Bacteroides ↓Clostridia |
↑Propionate |
||
| Citrus pectin [111,150,151,152,153,154,155,156,157,158,159,160] |
↑Bifidobacteria | Orange POS [161] |
|||
| ↑Bacteroides | |||||
| ↑Eubacterium rectale | ↑Acetate |
↑Bifidobacteria |
*Acetate | ||
| ↑Lachnospira | ↑Propionate | *Propionate | |||
| ↑Dorea | *Bytyrate | ↑Lactobacilli | *Butyrate | ||
| ↑Clostridium | |||||
| ↑Sutterella | |||||
| Sugar beet Pectin [162] |
↑Prevotella | Sugar beet POS [163] |
↑Bacteroidetes | ||
| ↑Bacteroides | ↑Acetate | ||||
| ↑Bacteroidales | ↑Acetate |
↑Propionate | |||
| ↑Clostridium | ↑Butyrate | ||||
| ↓Lachnospiraceae | ↓Valerate | ||||
| ↓Ruminococcus | |||||
| Apple and Citrus pectin [164] |
↑Acetate | Apple POS and Citrus POS |
|||
| no available data | ↑Propionate | no available data | no available data | ||
| ↑Butyrate | |||||
| Apple and sugar beet pectin [165] | ↑Bifidobacteria |
↑Butyrate ↑Acetate | Apple POS and sugar beet POS | no available data | no available data |
| Citrus and sugar beet pectin [81] | ↑Bacteroidetes | ↑Propionate |
Citrus POS and sugar beet POS | no available data | no available data |
| ↑Enterobacteriaceae | |||||
| with pectin and POS | Microbiota | SCFA | |||
| Citrus pectin, sugar beet pectin and POS from lemon peel [Gomez 2016 JFF] |
↑Bifidobacteria ↑Lactobacilli ↑Faecalibacterium ↑Roseburia |
↑Acetate ↑Propionate ↑Butyrate |
|||
↑Increase number of bacteria or individual SCFA level. ↓ Decrease number of bacteria or individual SCFA level. *Change of individual SCFA level.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



